Embed Size (px)
Citation preview

Aus dem ZentralNboratorium der Wiener Gebietskrankenkasse.
Stoffwechsel und vegetative Regulation. Z u r F r a g e d e r I n s u l i n w i r k u n g .
Von
Theodor Leipert.
Seit mehr 'als zwei Jahrzehnten hat sich die Ansehauung durch- gesetzt, die F fille aller Auf- und Abbaureaktionen, die wir als Stoff- wechsel bezeiehnen, werde yon vegetativ-nerv6sen Impulsen reguliert. Parallel damit ging d ie gewaltige Entwiekl~ng der Lehre von den Wirkstoffeu :und es wurde der Satz geprfigt: Der Stoffwechsel wird vegetativ reguliert und hormonal gesteuert. Der Regulationsmecha- n~smus allerdings war n~r in grot~en Z/igen ,bekannf, die Einzelheiten blieben unklar.
Man wul]te, dal] das Adrenalin der sgmpathischen Erregung zu- geo.rdnet ist, doch entsprach nicht jeder sy.ml~athis.ctae Reiz einer reinen Adrenalinwirkung am Erfolgs,organ. Frfih wurde daher e in weiterer sympathiseher Wirkstoff gefordert, den man Sgmpathin nam~te und der jfings,t als Noradrenalin idenSfiziert wurde. Als Hormon des Parasgmpathikus gilt das Insulin, das im Sgnergismus mit Aeetglcholin 1 die gleiehen Effekte auslSst, wie eine Reizung parasympathiseher Fasern.
In ihrer Stoffwechselwirkung fiihren die adrenergis,ehen Wirk- stoffe zur Mobilisier'ung des Depotzuekers Glykogen, zur Hyper- glyk~imie, zu gestei,gertem Sauerstoffverbraueh ~nd damit zu einer Erh6hlmg der Oxydafionsintegrale. Die Stoffweehsellage ist aeido- fiseh. Insulin dagegen deponiert im Zusam~.enspiel mit eholinergi- sehen Wirksto~ffen Gly'kogen, senkt ,den Btutzueker, spart Sauers.t~o.ff und erniedrigt den Grun.dumsMz. 'Dabei besteht eine Alkalose. Adrenalin ist demnach der Wirksto[[ der Dissimilationsprozesse, Insulin der Wirkstoff der Assimilation.
An ,dieser allgemeinen Konzepfion braueht aueh heute niehts ge- findert werden. Gremels ~ konnt,e am Herz-Lungenpr/iparat des intak~en, spontan atmenden Tieres zeigen, wie sieh dureh humorale Innervation, d.h. dureh Dauerirffusion kleinster Men gen adrenalin- iihnlieher Stoffe, wie Sympatol ~n.d Adrianol und dureh Infusion
4*

52 Th. Leipert:
ge r ings te r I n s u l i n d o s e n m i t A c e t y l c h o l i n j ede gewi in sch t e S tof f - wechaella, ge einst~l len l~l]t: Diss~.m~latorische Sc h i i be nfi t E r h S h u n g des B lu t zucke r s , Anstie.g der a r t .e r ioven6sen Di f fe renz be i ges te ige r tem S a u e r s t o f f v e r . b r a u e h u n d l an .ganha l t ende a s s imi l a to r i s che E r h o l u n g s - p h a s e n m~t B l ' t a z u e k e r a b f a l l , g e r i n g e r a r t e r i o v e n 6 s e r Di.fferenz u n d Sa 'ue r s to f fe inspa rung .
W i t s te l len bei den f o l g e n d e n U n t e r s u c h u n g e n den K o h l e h y d r a t - s to f fweehse l irt den M i t t e l p u n k t u n s e r e r Betra, e h t u n g aus zwei G,riir~den:
1. E r l i e fe r t die E a e r g i e f i i r j ede Ze l le i s tung; die Zel le sti~rbt, w,enn sie n i e h t Tnehr z u r G l y k o l y s e f fihig ist.
2. Die W e g e des E iwe i l ] s t o f fwechse l s vere in ig~n sich m i t d e m Ab- b a u w e g der K o h l e h y d r a t e . E i w e i g k 6 r p e r u n d Fe t te w e r d e n d a h e r f iber d e n s e l b e n o x y d a t i v e n Z y k l u s w ie K o h l e h y d r a t e zu E n e r g i e - l i e fe ran ten .
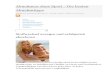
Im intermedidren Stoffwechsel zerfiillt 61ykogen naeh nebenstehendem Schema dureh das Ferment Phosphorylase in Hexose--1-phosphat, das fiber Hexose-6- phosphat in Hexose-1, 6-diphosphat iibergeht und dann in 2 Triosen zerbrieht: Glyeerinaldehydphosphors~iure und Dioxyaeetonphosphors~iure. Die Aldotriose dieses GIeiehgewiehts wird welter zu Phosphorglyeerinsiiure oxydiert, die in Phosphorbrenz- traubensiiure und unter Phosphatvefiust sehlieglieh in Brenztraubens~iure fibergeht. Die Phosphorylierung wird dureh Adenosintriphosphorsfture und Kreafinphosphor- siiure durehgeffihrt.
Mit der Brenztraubensdiure ist ein zentrales Zwisehenprodukt des Kohlehydrat- stoffwechsels erreieht. Sie wird teilweise zu Milchsdure reduziert. Damit ist eine Phase des Kohlehydratstoffweehsels durehlaufen, die als anaerobe Phase gilt und aueh als Glykolyse oder Glykogenolyse bezeiehnet wird. Der Vorgang ist nieht streng anaerob, wie man gew6hnlieh meint, sondern er enthNt eine oxydative Zwisehenstufe:
C6H• - - 4 H = 2 CaK~Oa Glukose Brenztranbens~iure
Nur die Anh~iufung gr6gerer Milehsiiuremengen hinder Wasserstoff und die t/eaktion erseheint als einfaehe Spaltung:
C6Ht~O6 = 2 CaH60~ Glukose Milehs/iure
An die Brenztraubens~iure knfipft die aerobe Phase des Kohlehydratabbaues in eigenartiger Weise an. 1Mol Brenztraubensiiurehydrat wird dureh Brenztrauben- s~iuredehydrase dehydriert und vertiert CO2 unter Bfldung yon Essigs~iureL Ein zweites Mol Brellztraubensiiure nimmt CO.~ auf und geht nach Woo&Werk- man a, a, 5, 6 in Oxalessiits5ure fiber. So entstehen zwei Intermediiirprodukte, ein C~-Khrper, Oxalessigsiiure und ein C2-Khrper, Essigsiiure oder eine aktive Form davon. Beide liefern miteinander eine unges~itfigte Trikarbonsgture mit 6 Kohlen- stoffatomen, die cis Aeonitsdure. Sie tritt durch Was seraufnahme mit Zitronensdiure und IsozitronensOure ins Gleiehgewieht. Isozitronens~mre geht in Ozalbernsteinsdure fiber, diese unter CO2-Verlust in a-Ketoglutarsdiure mit 5 Kohlenstoffatomen, aus der unter weiterem CO2-Vedust eine Ca-Karbonsiiure gebildet wird, die Bernsteinsiiure. Dadureh kommt der Ansehlug an das Ca-Siiuresystem Szent G~16rggis zustande mit der bekannten Umwandlung in Fumarsdiure, dpfelsdiure und Oxatessigsiiure. Es wird

Stoffweehsel und vegetative Regulation. 53
also gleichsam ein C2-KSrper am Kontakt der Oxalessigsiiure verbrannt und die scheinbar unver~inderte Oxalessigsiiure ist nach Ablaut dieses , ,Zitronensdurezyklus" 7 neuerlich zur Aufnahme eines Ce-KSrpers f/ihig. An diesem Kreisprozel3 sind eine Reihe dehydrierender Fermente beteiligt, die man heute unter dem Sammel.begriff Cyklophorasesystem zusammenfal~t.
Glykogen
[
Gh~kose-Iphospl~a~
Trio sephospho~'s~uren
I
Phoslohorbrenztraubens~ure
Phosphoroxal- . essi~sSoure
Alanin ~ ........................... ~- ~ Brenztraubensgure -~--~" Milehs~ure (Serin) v : ~ , (Cystein) ! ~-0xybutters~ure
-. . . . . . . . . . . . . . . . . . . . . . . . ~
�9 ~EssigsS~ure:. ~--~'AeetessigsS~ure ~---~Fett- Asparagins aure ~ .................. ~ iOiaie ssi gs~ u re l s ~iuren
ftlo fels~ure Aeonitsi~ure
Fumars~ure Zitronensi~ure Isozitronens~are
Bernsteins~ure Oxalbernsteins~ure~
Ketogl:ars~ure ~
,r . C02
Glutamins/iure (Prolin, tIistldin)

54 Th. Leipert :
Der h~er skizzierte Weg entspricht der Dissimilationsphase des Kohlehgdratstoffwechsels. Der Angriffspunkt des Adrenalins auf diesem Wege ist un'bekannt. Er wird u~mi~telbar an der Glykogen- spaltung vermutet, vielleicht am Phosphorylasesystem, das Glykogen in Glu.kose-l-phosphat spa.ltet:
Glykogen ~ Phosphat ~-~- Glukose- 1-phosphat.
Die Assimilation yon Kohlehydrat mfil]te den ~mgekehrten Weg gehen. In der Tat sind fast alle Teilreaktionen reversibel. Fiir den Kreispro,zei~ selbst steht die Frage tier Umkehrbarkeit nicht zur Er- 6rteru.ng, da jeder Weg zur Oxaless~,gs.iiure z uriickfiihren muir, d is,si- milator~sch fiber die Siiurea mit 6, 5 und 4 Kohlenstoffatomen oder assimilatorisch ~mter CO2-Bindung, z.B. von der a-Ketoglutarsiiure a u s 8, 9.
Ketoglutars~i~re -~ CO 2 -~ Codehydrase H~ ---- Isozitronensiiure -~ ~- Codehydrase.
Der Zerfall der Zitronens~iure in Oxalessigs~.ure und Essigs~iure steht noch z ur Diskus,sion lo. Dieses Zusammensp~,el von Assimilation un,d Dissimilation im Kreisprozel~ ist durchaus verstiindlich, denn es kann nur etw,as assimiliert werden, was vorher durch Dissimilations- prozesse zum Umbau bereitgestellt wurde.
Die Umkehr der ,,anaeroben" Phase des Kohlehgdratstoffwechsels �9 ist e~g mit der Fiihig~keit des Organismus zur C02-Assimilation ver-
kniipft, die durch Versuche mit isotopem Kohl,enstoff bewiesen wurde. So vermag das isolierte S~iugetierherz dutch isotopen Kohlen- stoff markiertes C02 zu as similierer~ und schweren Kohlenstoff in sein Glykogen einz~ubauen ~. Dabei fo~lgt die COu-Aufn.ahme der Wood- We r kman-R eaktio n :
Brenztrau.bens~iure -~- CO2~ -~ Oxalessigsiiure.
Ge~ebsbrei bildet sowohl mit Brenztrauber~siiure wie mit Oxal- es,sigsiiure die gleichen Substan'zen, Zitronens~iure, Ketoglutars~iure, Bernstei~siiure, Fumarsiiure ~ud )[pfel's~iure und die Umsat'zgr58e is[ yon der C02-Tensio~ abh~ingig 6. Die Sgnthese der Oxalessigsiiure nimmt im AssimiIationsweg der Kohlehydrate eine Schliisselstellung ein, da der riickl~iufige (Jbergan:g von Brenztraubensiiure in Phos- phorbrenztraubensiiure nur unter speziellen Bedingungen, wie An- h~iufung vo~ Kal~umi~onen und Adenosintriphosphors~iure mSglich ist. N0ch Megerhof 1~ hielt ihn ffir unmSglich und vermutete in der irreversiblen Spaltung yon Phosphorbrenztraubens~ure z u Brenz- traubensiiure tin Venti~l, das e~e ,glatte Resynthese ausschliel]t. Dieses Hindernis ist scheinbar. Der Organ~smus ~mg.eht es spielend, indem er die Oxalessigsiiure, vielleicht auch s,chon die Apfel.s~i'ure phospho- ryliert is, ~, 1~ und aus dem Phosphorylierungsproclukt COe ab-

Stoffwechsel und vegetative Regulation. 55
spaltet. Jedenfalls hat Verffitterung von Zitr.onensfiure einen promp- ten Anstieg des Leberglykogens zur Folge 16 ~nd M~s,kelbrei fiihrt Zitronensiiure leicht in Phosphorbrenztraub.en.siiure fiber 17, ein Be- weis, dag der Knotenpunkt yon Anaerobiose und Oxybiose glatt passiert wird. Auch dig Glykogenbildung aus Milehsiiure s,eheint diesen Weg fiber Brenztraubensiiure, Oxalessigsfinre, Pho.sphoroxal- essigsiiure in Phosphorbrenztra~a'bensfiure zu gehen, wie die Ver- teilung des isotopen Kohtens.toffs ergib:t 15, w/ihren,d ein Teil fiber den Krei.sprozefi oxydiert wird (Pasteur-Meyerhofsehe Reaktion).
Es binder also Brenztraubensiiure C02. So bremst gev~isserma~en die bei tier Dissimilation gebi.ldete Kohlensiiure 4en brfisken Abbau und vermittelt die rfiekliiufige Synthese von Kohlehydrat. Sehon Hermann Rein is hatte fiber die M6glie.hkeit einer zentralnerv6sen Regulierung des oxydativen Ges,amtstoffweehsels dutch Kohlensiitlre beriehtet. Er konnte zeig'en, .dal~ Mensch und Tier ihre Saners:toff- aufnahme eins,ehriinken, wenn sie ein Gemisch yon Luft mit 0,5 bis 6% CO2 at.men. Die maximalen UmsatzserLkungen betrugen etwa 20% des Ruheumsatzes und wurden gerade bei ,niedrigen C02-Kon- zentrationen beobaehtet, die durebaus im Bereieh des physiologi- seh,en Gesehehens liegen. Wir kenn,en heute n.och ni'eht den genauen Meehanismus der Wood-Werkman-Reaktion, do,eh wi,ssen wit, dag sie mit Phos,phoryl~erungsprozessen verkniipft ist und dnreh Vitamin B 1 katalysiert wird.
Eine der Bedingungen, dig eine prompte as'similatorische Phase ausl,6st, ist der Sauerstoffmangel. Als ",v.ir den Kohlehydratstoff- weehsel im Unterdruekkammerversu,eh prfiften 19, 20, fanden wit eine alte Beobaehtung yon Evans 21 aufs neue besffitigt. Wenn man hungernde Ratt.en 'mit 0,1% Leberglykogen 24 Stur~d,en bei 0,5 at 5500 m hiilt, dann steigt der Glykogengehalt der Leber auf fiber 1,4%, Gut g.effitterte, glykogenreiehe Tiere zeiger~ im Unterdruek xueh naeh mehrtii:gi.gem Hunger keinen Abfall ihres Lebergly,kogens. Wit konnten in kurzfristigen Unterdruekversu.ehen bereits, naeh drei- stfindi, gem Aufenthalt in 7000 m HShe ein Leberglykogert yon 0,3 his 0,8% naehweisen, Eine solehe Glykogenanhiiufung ist ganz nn- gew6hnlieh; denn :lii~t man Ratt,en bei normalem Druek htmgern, dann si~kt ihr Leberglykogen innerhalb yon 24 Stunden auf nn- geffihr 0,1%, Mei.bt etwa zwei Tage auf fiefem Niveau, um allmiih- liet~ wieder anzusteigen. Im Sauerstoffmangel dagegen tritt diese Glykogenbildung unverzfiglieh ein und man kann ohne beson'deren Eingriff dutch einfaehe Verminderung der Sauerstoffzufuhr eine prompte assimilatorisehe Phase ausl.6sen.
Der neugebildete Zueker stammte si~eher ni.eht aus dem Glykogen- depot der Muskulatur, denn de~en Glykogengehalt .blieb unveriindert.

56 Th. Leii0ert:
Es kam also keine Kohlehydratverschiebu~ag, sondern nur eine Neu- bildung yon Glykogen aus EiweiB oder Fett in Frage. Fiir seine Bildung aus Eiweil~ fand sich in Belastungsversu, chen mit den A~cninosiiuren Gtykokoll 'und Alanin kein AnhaltsFunkt 19. Es schien im Gegenteil, als ob die Glykogenreserve der Leber, abel" auch die der Mus~:ulaCur unter ,dem EinfluB beider Ami.nosii~uren im Sauerstoff- mangel bewegScher wiire, was dadurch verstfindli~zh wiirde, dab ge- rade diese beiden Amino.siiuren die st/irks.re spez,ifis,ch dynamische, also dissimilatorische Wirkung enffalten. Der Weg der Fettsiiuren zum Kohlehydrat ist zweifell.os gangbar, wie Isotopenversuche er- geben 22, 2a. Atmh ist die Theorie dev ~-Oxydation der Fetts~iuren nach Knoop immer wieder bestiitigt worden. Doeh blieb alas Schick- sal der dabei anfallenden C2-Bruchstii.cke ung.ewil~.
Ffir eine lebhafte Fettwanderung im Sauerstoffmangel sprechen alte Bef.unde 24, 2~, 26 Bei Meerschweinchen tritt nach drei Tage langer Hypoxie eine hochgradige Leberverfettung auf mit Ver'meh- rung des Neutralfettes ~nd der Phosphatide bei gleichzeitiger Ab- n.ahme des Depo,tfettes 27, 2s, 29 Kaninchen zeigen unter gleic~en Bedingungen eine Lipiimie verbunden mit einem auffMligen Anstieg des Serumcholesterins 30. Die Erscheinun, gen sind bei Pflanzenfressern stets sehr a~usgeprS:gt, da ihr Fettumsatz weniger intensiv ist, s,o dal~ sich auch alimentfir bedingte Steatosen leicht ausbilden. Sie sind lm- deutlich bei tier omnivoren Ratte. Immerhin fehlen auch h~er nicht die Zeichen einer gesteigerten Fettbewegung.
WiT 'untersn,chien an drei ~aormalen Ratten naeh 24sfiindigem Hunger ,und an zwei Tieren, die aul~erdem no,eh drei Stuuden bei 354 mm I-Ig (6000 m) gehalten wurden, die Leber auf Gesamffett 31, Lipoid-P und -N, Ges.amt- und Es~ereholesteri.n 32 s.o~Me die Jodzahl des Gesamffettes und der Fettsiiuren aa (Ta~b. 1).
Tab. 1. F e t t - u n d G l y k o g e n g e h a l t d e r R a t t e n l e b e r u n t e r n o r - m a l e n V e r h i i l t n i s s e n u n d i n d e r H y p o x i e . ,
I Tier ] ~ des Nr. [ feueht.
Organs
I N0rmal 1, 2, 3 I 4,83
ttypoxie 4, 5 I 4,48
A t h e r e x t r a k t
rag% Cholesterin frei Gesamt
4,69 5,03
4,87 5,00
mg% Lipoid
P N
2,70 2,39
3,13 2,35
P : N Jodzahl
Gesamt Fettsr.
1:1,96 86,7 55,2
1:1.66 86,0 71,8
% Glyko-
0,186
�9 0,348
Eine Leberverfettung tritt bet der Ratte im Sauerstoffmangel ni'cht ein. Sie wiire aueh bet glei.ehzeitigem Anstieg des Leberglyko,gens schwer zu deuten. Sehen wit jedoeh in den Phosphafiden und Phos- phatidsiiuren eine Transporfform der Fettsiiuren, dann zeigt die Ver-
gen

Stoffwechsel und vegetative Regulation. 57
mehrung des Lipoid-P uncl die Senkung der Relation P : N deut~ich chin Vermehrun.g beider Frakt ionen im Sauersr Es wan- dern ,dcmnach in cler Hypoxie Fetlsiiuren zur Leber. An dieser Wan- derung hiram[ ~das Cholesterir~ nicht tell, wie aus den Cholesteri~- werten, insbesondere tuber aus den kaNm vcr~inderien Esierfraktio~nen hervorgeht. Die Joclzahl der Fettsiiuren ist in der Hypoxie erhSht, bet glei,Cbcr Jodzahl des Gesamtfettes, Es ist also eine Desaturierung der Fettsiiurer~ eingetrete~, wie sie bereits frfiher beobachtet und als An- passungsmechanismus an den verminderten SauerstoffaustaNs~ch ge- deutet wurde 34. Gerade di'esc Abnahme des S~ittigungsgrades abcr letter na~ch unserer heuti,gen Anschauung den oxydativer~ AbbaN der Fctts~iuren ein. Uber Wasseranlagertmg und neuerl~che DehydrierNng folgt die Bildung der Ketos~iure, die zur Siiurespaltung fiihig ist und so die Verkiirzung der Fettsiiureker durch ~-OxydaSon ermSglicht unter ,dauerncler Bildung yon C2-Resten.
Wir versuchten nun unter der Annahme, diese Inter m~di~ir- produkte mit zwei Kohlenstoffa tomen wiiren e'mfache Essigs/iure- reste, du rch Verabreichung yon Natr~umacetat aa Ratten eine Zitronens~iurevermehrung z u erzwin,gen, aJso yon einem Intermedidr- produkt des FettsSureabbctues zu einem Zwischenprodukt des oxgda- tiven Kohlehgdratstoffwechsels zN kommen. Die F/ihigkeit der Z}tronens~ture, Glykog,en z u biIden, war bereits erwiesen ~6 nnd konnte yon uns neuerlich best~itigt werden. Wohl liegt die Zitronen- siiure auf einem Seitenwege des oxydativen Kreislaufs, do,ch ist sle [gegeniiber den an de.ren ZwischenkSrpern lei~cht nachzuweisen 35.
Die perorale Belastung unserer Tiere erfolgte, wenn nicht anders angegeben, mit 1 mg de.r zu prfifenden Substanz je 1 qcm KSrperfliiche berecbnet nach der Formel: Oberfl~iche -- 11,36 X Gewicht -"/~ ~0. Die Dosis wurde in der Regel nach 12stiindigem Hunger verabreicht und nach 24stiindigem Hunger wiederholt. Im NormaIversuch blieb das Tier noch weitere 3 Stunden am Leben, im Hypoxieversuch wurde es ebenso lange bet 354mmHg (6060m) gehalten. Als AcetatlSsung diente eine 3,333 molare L5sung yon kristalliertem Nafriumacetat entsprechend 200mg Essigsiiure je 1 ccm. Die erforderlichen Mengen wurden reaktionslos vertragen, ohne Erbrechen und ohne DurchfSlle. Als Oxalessigs~iure stand uns ein selbst dargestelltes Priiparat vom F. P. 1760 zur Verfiigung 37. Unterdruckversuche wie friiher ~0.
Es ge'lang uns eine vermehrte Zitronens~urebildung nur unter zwei Bedingungen na.ch.zuweisen:
1. Das Tier mufite gIykogenarm seth.
2. Es muflte im Satterstoffmangel gehalten werden. Blut, Leber und Muskeln normaler Ratten er/thalten 2 bis 4,5 rag%
Zitronens~iure, Rattennieren bis zu 15 rag% (Tab. 2), }Verte, die mit den Angabert der Literatur iibereinstin~ner~ ss vom Glykogengehait der L eber unabh~ingig sind uncl sich auch im Sauerstoffmangel nicht ~i~ndern.

58 Th. Leipert:
Tab. 2. L e b e r g l y k o g e n u n d Z i t r o n e n s i i u r e g e h a l t v o n R a t t e n - o r g a n e n , n o r m a l u n d i n d e r H y p o x i e .
Tier:Nr. IGeg ieht
9 12 18 20 13 10 23 11
232 225 135 205 258 260 150 135
Leber
2,4 2,4 2,9 2,7
2,1 1,6 2,3
mgO, o Zitronens~iure
Muskel Blur Niere
2,8 2,4 - - 2,7 2,6 14,6 3,3 4,4 6,7 3,7 2,6 5,4 2,9 2,4 - - 4,4 4,2 - - 2,9 3,1 13,1 3,0 3,7 - -
~ Glvkogen
1,629 5,360 0,170 0,150 3,890 0,441 0,267 1,418
Normal - - Glykogenreieh
,, 24 Std. Hunger 24
3 Std. Hypoxie -- (~lykogenreieh 3 . . . . - 24 Std. Hunger 3 . . . . -- 24 . . . . 24 . . . . -- 24 . . . .
Acetatbelastungen be i H u n g e r t i e r e n u ,n ter n o r m a l e n D r u c k -
v e r h [ i l t n i s s e a z e i g e n Z i t r o n e n s ~ i u r e w e r t e a n d e r o b e r e n G r e n z e d e r
N o r m (Tab . 3) , d o e h n i e m a l s w e s e n t l i c h d a r i i b e r . D a s w,ar n i c h t
s o n d e r f i c h i i b e r r a s c h e n d , k a n n m a n do e h s d b s t n a e h I n j e k t i o n y o n
C4-Dicarbons~i ,u ren , also, a u s g e z e i c h n e t e r Z i t r o n e n s f i u r e b i l d n e r u n d '
a n e r k a n n t e r V o r s t u f e n de r Z i t r o n e n s i i u r e d e n Z i t r o . n e n s i i u r e g e h a l t
Tab. 3. L e b e r g l y k o g e n u n d Z i t r o n e n s i i u r e i*n R a t t e n o r g a n e n n a c h A c e t a t b e l a s t u n g u n t e r w e c h s e l n d e n B e d i n g u n g e n .
Tier ~r.
22 36 38
45 14 46 21 19
28
33 37 29
24
25
G-ew. g
185 205 215
125 150 160 220 120
95
230 210 213
215
175
mg% Zitronensiiure
Leber [ Muskel
3,5 4,1 4,1 4,7 3,4 4,3
2,9 3,6 2,3 6.5 3,6 311 5,5 9,7 4,5 11,0
Blur
5,2 4,7 5,6
2,9 6,6 4.8 9;1
10,6
Niere
11,7 16,7
6,8
10,3 58,0 64.2
% Leber- Gly-
kogen
0,111 0,177 0,236
0,090 0,070 0,200 0,150 0,137
2 • Aeetat, 24 Std. Hunger 2X ,, 24 . . . . 2X ,, 24 . . . .
l• Hunger- Hypoxie, 1 8td. 1• ,, ,, ,, 5 , , 2X . . . . . . I ,,
2X . . . . . . 3 ,, 2 • . . . . . . 41/2"
6,0
2,1
4,0 2,5
3,3
4,2
11,3
3,8 4,1 2,5
5,5
5,0
12,6
3,9 7,6 1,7
4,1
4,4
46,8
17.4 1613 9,3
17,t
16,3
0,210
0,150 0,181 0,139
2,880
o,43~
2 • Aeetat- Hunger- Hypoxie, 3 Std. 5 Std. normal
ebenso ebenso
1 X Aeetat- Hunger - Hypoxie, 3 Std. 12 Std. normal
2 x Aeetat + G-iukose 1 rag/1 em s Brotfutter. Hypoxie 3 Std.
ebenso

Stoffwechsel und vegetative Regulation. 59
des Blutes nur wenig erhfhen, w/ihrend gleichzeitig grol~e Men,gen a usgeschieden werden. Man erkliirt das mit der rasehen Aus.seheidung der Zitronensiiure 39 oder m~t ihrer Bildung in der Niere 38
Aul~erdem wird verf/itterte Zitronensiiure reiehlich abgebaut, ohne im Harn zu erscheinen 40, 41. Die Essigsfiure war nicht nur in Ver- su, ch,en mit Gewebs,brei, sondern aueh in vivo im Belastungsversueh kein Z~tron,er~siiurebildner 42, nnd sel,bst doff, wo naeh Acet,at- belastung eine Vermehrun,g tier Zitronensiiurea~asscheidung beob- achtet wurde, war der Alka.lieinflul3 des Acetats auf die Zitronensfiure- bildung yon der Essigsiiurewirkung nicht zu trennen 40.
In der Hgpoxie tritt nach Acetatbelastung ein auffalIender Anstieg der Zitronensi~ure im Muskel, Blut mid in der Niere auf, wiihrend der Zitronensiiuregehalt der Leber unveriindert bleibt. Dieser Anstieg er- folgt la.ngsam. Einst/indige Hypoxie isl unzure}ehend. Der Anst.ieg fib erdauert den Sauerstoffmang~el teilweis,e betriichtlieh u n d e r tritt nur bei mangelndem Leberglykogen ein, d.h. beim Fehten einer leieht mob.ilisierbaren Kohlehydratreserve. Die gleichzeitige Verffitte- rung von Ko.hlehydrat verhindert ihn. Wir ko,mmen auf diese mer.k- wfirdige ~rirkun,g kleiner Zuekermengen noeh zurfiek. GlykcNen wird unter ,unseren Versnehsbedingungen nieht gebildet. Die Werte f/ir Leberglykogen erLtspreehen durehwegs Hungertieren. Es ist be- kannt, dal~ Fettsiiuren Glykogen mobilisieren und eine Hyper- glyk/imie verursa.chen 4a, 44. Sie wirken a~o ,der erwarteten Assimi- lation entgegen.
Man k6nnte den Einwand erheben, die Essigsfiure wirke .in unseren Vers.uehen n'ur indirekt. Sie vermehre den Eiweigabbau ~and die dar- aus hervorgehen.den Ketosiiuren s'eien die eigenfliehen Z~tronensiim'e- bildner. Do.eh wurde ein gesteigerter Eiweil~umsatz na,eh Essigs~iure- zufuhr an der hungernden t/atte aueh in liinger dauern.den Ver- suehen nieht beo,b~aehtet 45.
Die Essigsiiure seheint demnaeh als solehe zur Zitronensfiure- bildung ffihig zu sein und die Armahme ein,er ,,aktiven" Ess'igsiiure oder tier xus zwei Essigsiiu.reresten hervorgehenden Aeetessi~gsfiure als eines reaktionsfiihigeren Zwischenprod'uktes ist durehaus nieht zwin- gend 46, 47, 48
Es war nns aufgef~llen, dal~ der Zitronensiiureanstieg sehr zfgernd erfolgt und au,eh fiber die Hypoxie hinaus noch einige Zeit bestehen bleibt. Es lag daher nahe, diese Sperre im oxgdativen Zgklus als vago- tone Schutzreaktion zu deu~en. Dal~ die H6henkrankheit gemein- s.ame .Symptome mit einer Tonus,erh6hung des Parasympathikus a'ufweist, wurde sehon frfiher behauptet 49 und btieb aueh nieht ganz ohne Widersprueh 50. Mit der Vagotonie ist eine Speieherung von Glykogen verbunden und man hat das Insulin, das diese Speieherung

60 Th. Leipert:
katalysiert, als das Hormon des Parasympath~kus bezeichnet 51. Nach, Gremels 1 arbeitet es synergetisch mJt dem ,,Vagusstoff" Acetylcholin. Nun ist die Hypoxie, eine allerdings unfreiwillige Einsparung yon Sauersteff, in glei~cher Weise mit einem Glykogenzuwaehs in der Leber ,des hungernden Tieres verbunden. ~Vir Suchten daher nach einem Zusammenhang des Insulins mit dem Zitronensdurestof[- wechsel, der nach Mmungsversuchen zweifellos vorhanden sein m n ~ e 52, 53, 54
Wir gaben Ratten nach Acetatbelastung Dauerinfusionen gering- ster Dosen yon Acetylcholin und Insulin wfihrend drei Stunden. Ace~lcholin war in unserer niedrigen D osieruTlg unwi,r.ksam, gr6Bere Dosen dagegen waren lmbrauchbar, da s:ie den Sanerstoffverbra~ch steiger.n. Eine pharmakologische Hemmung der Cholinesterase muBte vermieden werde~, um nicht schwierig zu iiberblickende Verhiilt- nisse zu schaffen; denn Prosfigmin erh6ht den Zilronensfiuregehalt des Btutes 55. Auf Ins~li.n nnd Insulin -]- Acelylcholin spricht der Zitronens~iurespiegel sofort an (Tab. 4), obgleich auch bier u.nter dem Einflug tier Essigs~iure eine Glyko,genzun'ahme in der Leber aus- bleibi.
Tab. 4. D e r Z i t r o n e n s i i u r e g e h a l t y o n R a t t e n o r g a n e n n a c h I n s u I i n u n d A c e t y 1 c h o 1 i n (A. Ch.). Subkulane Dauerinfusion beider Stoffe
in 5 ccm isoton. Na-AcetatlSsung innerhalb yon 3 Stunden.
Tier I Gew. Nr. g
31 ] 205 32 200 34 215
35 I 220
mgO/o Zitronens~ure
Leber [Muskel[ Blur Niece
3,5 5,1 5,8 9,8 3,2 5,4 11,9 51,7 5,1 4,2 9,8 24,1 8,4 6,3 22,7 4 8 8
o/0 Leb er-[ Oly-
kogen
I o,186 [ l • Aeetat + 0,05 7 A. Oh. 0,142 [ 2~ ,, +0,IE Insulin 0,108 2X ,, § ,, 0,178 I 2X ,, +0,1E ,, +0,17
I A. Oh. Kr~mpfe!
Insulin verursacht daher bei Ratten nach Acetatbelastung den gleichen AnsHeg dec Zitronensdure mie eine Verminderung dec Sauer- stoffzufuhr. Wiederum betrifft die Zitronensiiurevermehrung in erster Linie die Niece, dann das Bl,ut, wiihrend ein wesentlieher An- stieg in tier Muskulatur ausMeibt. Dagegen zeigt die Leber im Insulin- krampf, in den Tier 35 unabsichtlich geriel, eine beaehtliehe Ver- mehrung der Zitronensiiure.
~berrasehende Ergebnisse erhielten wir, als wit auch die Oxal- essigsOure (OES.) in unsere Unters~ehungen einbezogen. Wit gaben sie zunfiehst peroral in der iibliehen Dosis yon 1 rag/1 qem, was 1,45 mg Zitronens/iure je Fl~ieheneinheit entsprieht. Sie wurde in wenig Wasser suspendiert und dutch Zuga,be einiger Tropfen kon-

Stoffweehsel uncl vegetative Regulation. 61
zen t r i e r t e r N a t r o n l a u g e bis a u f p H ~ 6 vSll ig in L 6 s u n g g eb rach t . Sp i i te r be i den A c e t a t - O x a l a c e t a t g e m i s c h e n e r h 6 h t e n w i r ,die Men ge a u f 1,38 m g OES. /1 q c m m i t 0,62 m g Ess~gsiiure/1 qcm, g a b e n also die 2 m g Zi t ronens i iu re /1 q c m ~iquivalente Menge. Schl ie f i l ich pa l ] t en w i r u n s u n s e r e r u r sp r f ing l i ch , en Aceta tgRbe y o n 1 m g Es.si,gsfiure je 1 q c m an.
Tab. 5. D e r Z i t T o n e n s i i u r e g e h a l t (ZS.) v o n R a t t e n o r g a n e n n a c h B e l a s t u n g m i t OES. u n d E s s i g s ~ i u r e + O E S .
Tier
48
49
52
56
41
50
53
54 42
43
47
51
55
Gew. g
160
170
120
125
]55
120
140
95 160
200
175
155
115
�9 rag% Zitronens~iure
Leber I .Musk. I Blut Niere
3,9
4,0
5,0
3,8
4,8
4,1
6,1 7,0
5,9
5,6
6,8
7,0]
o/oLeb.- Giy-
kogen
2,4
3;2
4.4
5.3
2,8
4,4
3,5
6,8 8,6
5,7
5,8
6,4
6,4
3,5 10,4
3,4 9,9
6,6 66,6
10,5 46,6
3.7 12,4
8,.~ 43,6
4,9 12,2
12,7 44,3 16~5 69,2
26,7 30,6
- - 56,3
29,8 56,9
34,4 61,3
0,091
0,090
0,180
0,091
0:330
0,084
0,216
0,061 0,124
0~098
0,096
0,184
0,105
mg ~ Blut- zucker
11
38
7O
54
22
O4
73
66
36
77
. . . . 2 • (2 mg ZS/cm 2)
2 X Ace'~at + 1 • OES (2 mg ZS/em0
2 X Aeetat + 2 X 0ES (2 mg ZS/em 2)
2 X Acetat + 1 • 0ES (3,2 mg ZS/cm ~)
. . . . 2 • 0ES, Hypoxie 3 Std. (1,45 mg ZS/cm 2
2 • Aeetat + 1 • 0ES (2 mg ZS/em 2) Hypoxie 3 Std.
2 x Aceta~ + 1 x 0ES (2 mg ZS/em ~) Hypoxie 3 Std.
ebenso 2 • Acetat + 1 • 0ES
(3,2 mg ZS/cm0 ttypoxie - - Tod nach 2 Std.
ebenso Hypoxie - - Tod nach a/4 Std.
2 • Acetat + 2 ~ 0ES (2 mg ZS/cm 2) Hypoxie -- Tod n~ch P/2 Std.
ebenso Hypoxie - - Tod nach 13/4 Std.
2 • Acetat + 1 • 0ES (3,2 mg ZS/cm0 Hypoxie - - Tod naeh ~/4 Std.
W i r f i i g t en z u r e r r e c h n e t e n A c e t a t m e n g e die i iqu iva len te Menge OES . ---- 2,2 i~g OES. /1 qcm. D a d ' u r c h w u r S e n d ie T ie re m i t e inem Gem{sch belas te t , (]as z u r B i l d m l g v o n 3,2 m g Z i t ronens~ure /1 q c m a u.sgereicht h~itte.
Die T ie re e r t r u g e n Belastungen mit OES. seh r g u t u n d o h n e n e n n e n s w e r t e n Ans t i eg der Z i t ronens f iu re , s e w o h l i m N o r m a l v e r s u c h wie in der H y p o x i e (Tab . 5). N a c h Beiga.be de r Essigs~iure k o m m t es a u c h u n t e r n o r m a l e n Verh i i l tn i ssen zu e iner te i lweisen V e r m e h r u ~ g

62 Th. Leipert :
de r Z i t ronens / iu re , b e s o n d e r s n a c h . h o h e n O E S . - D o s e n . Diese Tiere s ind n u n a u l ] e r o r d e n t l i c h e m p f i n d l i c h gegert Sa,uersto,ffmangel . Sie mfiss.ert l a n g s a m dev L u f t v e r d i i n n u n g aus:gesetzt w e r d e n u n d gehen be i h o h e r O E S . - D o s i s h i iu f ig u n t e r Kr~impfer t z u g r u n d e . D a b e i schne l l t alas T i e r p lS t z t i ch in die H6he, ro l l t u m seine L/ings,achse u n d u n t e r k l o n i s c h e n Zuckur~gen tri• der T o d ein. D u r c h E r S f f m l n g der Caro t i s l~il~t s ich n u t wen ig B lu t g e w i n n e n . Die T o t e n s t a r r e m a ~ h t s i ch in weni~e11 Mint~ten b e m e r k b a r . D a s w a r e n auff/i l lSge Z e i c he n eines hyp<rg lyk / im i schen Schocks . Sie w a r e n axm so b e f r e m d e n d e r , als d e n Ketos~iuren g e w S h n l i c h eine s y m p a t h o m i m e t i s c h e , a l so b lu t - z u c k e r s t e i g e r n d e W i r k ' u n g z u g e s c h r i e b e n w~rd ~6. Die B l u t z u c k e r - b e s t i m m u n g ,zeigte in d e r T a t bei diesert T i e r e n gan~z be tHich t l i che H y p o g l y k ~ i m i e n , i m Geg,ensatz z u r H y p e r g l y k ~ i m i e der T ie re u n t e r n o r m a l e n D r u c k v e r h i i l t n i s s e n . So fiig~ s i ch d ie u n g e w S h n l i c h e E m p - f indSchke i t gegen Sauer s to f fman ,ge l s eh r g u t z u m h y p o g l y k ~ m i s c h e n Gesamtb i l d ~ , 5s, 59
Man darf dabei allerdings nieht fibers ehen, dab die Verffitterung der Alkalisalze organiseher S~iuren zu einer Alkalose fiihrt, die eine vorhandene Insulinwirkung verst~irkt. Im Sauerstoffmangel wird darm dureh die anffingliehe Hyperpnoe leicht eine Hypervenfilationstetanie ausgelSst, die mit einer Neigung zur Hypoglyk/imie einhergeht 60, 61. Bemerkenswert ist, dal~ ein tetaniseher Anfall dureh genfigend groBe Zuekergaben verhindert werden kann 6L Nun zeigen die Tiere welehe die Belastung mit Aeetat q- Oxalacetat im Sauerstoffmangel fiberlebten, ebenfalls einen gelegentlichen Tremor, yon dem sie sich aber nach Anpassung an die Luftverdfinnung erholten. Gerade diese Tiere haben jedoeh bei normalem oder erhfhtem Blutzucker nut einen Zitronens/iurespiegel des Blutes yon 4,9--12,7 mg%, wfihrend er bei den in der Hypoglyk/imie verendeten Tieren auf 16,3--34,4 mg% ansteigt, also jene HShe erreicht wie im Insulinschock. Ebensowenig fehlt auch hier die frfiher beobachtete Zitronen- sfiurevermehrung in der Leber (Tier 35, Tab. 4). Wir kSnnen daher dieses mlter- schiedliche Verhalten nur durch eine mehr oder weniger intensive Insulinwirkung erkl/iren, keineswegs durch die aliment/ire Alkalose, die bei beiden Tiergruppen nahezu yon gleicher GrSBenordnung sein muB. Unseren Ergebnissen stehen alte Befunde yon Davies, Haldane und Kennaway 6a sowie von Greenwald 64 sehr nahe, welche die Ursache der Atmungstetanie fin Sauerstoffmangel der Gewebe sehen, be- dingt durch die grSl]ere Best/indigkeit des Oxyh/imoglobins im alkalotischen Organis- mus. Dabei kommt es zur Vermehrung der Milehs/iure und zur Bildung einer un- bekannten nieht flfichligen S/iure, die weder Acetessigs/iure noch ~-Oxybutters~iure ist (Zitronensfiure?).
In der Tat bestehen engste wechselseitige Beziehungen zwischen Alkalose, Sauer- stoffmangel trod lnsulinmirkung einerseits und der Zitronens~iurebildung anderseits.
Insulin verursacht eine Alkalose mit Anstieg der Alkalireserve des Blutes und Zu- nahme der Alkalit/it des Harns. Umgekehrt verst/irkt eine Alkalose die Insulinwirkung worauf Dienst 65 erneut hinwies. Wir selbst fanden bei Blutzuckeruntersuchungen nach Bikarbonatgaben die hypoglykfimische Phase ungewShnlich tief 20. Gerade die Alkalose aber lfif~t auch die Zitronens/iureausscheidung gewaltig ansteigen, ohne dab es zu einer ErhShung des zitronens{iurespiegels im Blute kommt a0, al, 66, 67. Ostberg (zit. bei 66) hielt eine vermehrte Zitronenausscheidung sogar ffir ein Zeichen einer alkalotischen Stoffwechselst6rung, In nahezu allen friiheren Arbeiten fiber Zitronen-

Stoffweehsel und vegetative Regulation. 63
s~iure finden sich znsiitzliche Belastungsversuche mit Bikarbonat, um den ,,Alkalose- fehler" auszuschalten. Setzen wir an Stelle der Alkalose die durch sie verstiirkte Insulinwirktmg, dann erkliirt sich eine Vermehrung der Zitronensiiureausscheidung ebenso zwanglos, wie ihre Verminderung nach Zufuhr von Siiure nnd sauren Salzen 68.
Wir finden einen Anstieg der Zitronens~iure bet Acetat-Tieren im Sauerstoff- mangel. Doch Alkalose bewirkt durch Hemmung der Dissoziation des Oxyhfimo- g]obins ebenfalls eine Hypoxie der Gewebe 63, ~, 6s nnd die sauerstoffsparende Wir- kung des Insulins 1 ist lange bekannt. Im Friihsiadium der Insulinkriimpfe zeigt das arierielle Blut venSsen Charakter miI einer O2-Siittigung yon 80% 89, das venSse Blur aber ist hellrot und maximal mit Sauerstoff ges/itiigt, da die Sauerstoffabgabe in den peripheren Gefiil3gebieten herabgesetzt ist 7o.
W i r schliel~en aus unse ren Vers~achen, daft das Parasympathikus- hormon Insulin am Ubergang oon Anaerobiose zur Oxgbiose eine ent- scheidende Rolle spielt. Das ist d~,e Stelle, w o .die Brertzt.raubensiiure einerseits in Essigsiiure u n d CO 2 zerfiillt, andersei ts aber d u t c h CO2- A u f n a h m e n a c h Wood-Werkman in Oxalessigsiiure i ibergehL Die S u m m e beider I r t te rmedi f i rprodukte is~ Zi t ronensi iure n a e h der ein- ~achen Bi lanz:
2 Brenz t raubens i iu re + O ~--- Zitronensiiure.
Der endogene Schwund oon CO~ k o n n t e y o n u n s a m Abfall des respiratoriscbe~ Quotienten (RQ.) der Ve,rsuehsliere au f Wer t e bi,s 0,68 a u c h bewiesen w e r d e n (Tab. 6).
Tab. 6. R e s p i r a t o r i s e h e Q u o t i e n t e n i m S a u e r s t o f f m a n g e l .
I I ; 0irator sober 0uo ie Nr. Std. Nach hbsetzen des Versuchsbeginn
Futters
7 ] 4 3 ] 0,86 0,74
Satterstoffmangel Std. 1 2 5 12
0,68 0,72 - - - -
0,72 0,72 0,71 0,76
Die Wer te gleichen v611ig den,en, die a m Menschen n a c h [age- lan,gem Verwei len in der U n t e r d r u c k k a . m m e r beobach te t w u r d e n , a u c h im stf i rkeren Abfal l des I/Q., w e n n sein A u s g a n g s w e r t ur- spr / ingl ich h 6 h e r lag 71. Sie beobach~eten W e r t e n n a c h tion von 1938 72
Man hai te diese ex t rem
gleichen a u c h den unmi t t e l ba r a m Berg de m B ericht der Moun t -Everes t -Exped i -
n iederen Wer te bis,her n u r d u r c h ver- mehr t e Sauers tof f re ten t ion an die Fet ts i iureket ten zu deu ten ver- sucht , was den Quotientert CO2/O ~ ern iedr igen mul~te. Sie galtert als Beweis f.fir den ~ b e r g a n g y o n Fet t in Koh lehyd ra t . Der endogene Verl~ast yon CO~ d u t c h B i n d u n g an Brene t raubens i iu re und Bil.dung yon Oxalessigsiiure m u g j edoch in gleicher Weise den RQ. er- niedrigen, ja er f i ihr t d u r e h W e g n a h m e des CO2 aus dem P u f f e n m g s - sys t em N a H C O j C O 2 z u m Uberwiegen des Biearbonats , z u r Alkalose.

64 Th. Leipert:
Dadurch wird die Alkalose der vagotonen Stoffwechsellage ebenso verstiindlich wie alas me rkwfirdige, bis jetzt ungekliirte WechselspM: Insulin macht Alkalose und Alkalose ver'stiirkt ,die Insulinwirkung.
Klar wivd auch tier Synergismus yon Insulin mit Vitamin B1, das als Coferment die beiden Abbauwege der Brer~ztrau.bens~iure kataly- siert und so 4er Insulinwirkung unmittel6ar vorgeschaltet ist. Ein gegenseitiger Einflul~ im synergistischen Sinne is,t schon auf Grund der Massen~wirkung verstiindlich.
Insulin senkt den Brenztraubens~iuregehalt ,des Blutes genau wie Aneurin 73, 74, 75 had f6rdert alIgemein den Umsatz der Brenztraub,en- s~iure v6. Anderseits ffihrt j,ecle stfirkere Beansprtmhung des Insulins, z. B. die Anpassur~g cter Kohlehydratoxyda4ion an die Hypoxie, fiber einen relati.ven Ins~linmangel nicht nur zu einer Abs,ehw~chung der hypoglyk~mischen Phase der Blutzucker-Belastun, gskurven~ son,dern ~uch zu einem Anstieg der Brenztran'b,ens~iure genau wie ein B1- Defizit 19 Aneurin setzt den Nfi.chtern,blutzucker herab, die Be- lastungshyperglykiin~ie ist nach B1-Gabeu geringer als ~m Normalfall und die Insulinhypoglyk~imie t~i~t sich dutch B 1 verstiirken vT-so B}utzuekerkurvert im B1-Mang'el zeigen diabetischen Charakter s~ Selbst ,aloft, wo ein nnmittelbarer Einflul~ des Aneurins auf den Niiehternblutzueker n}cht beobachtet wird, blei~bt seine f6rdernde Wirkung auf die Glykoger~bildung in der Leber na,ch Glakos,ezufuhr eber~so unbestri.tten wie die des Ins'ulins s2 Au~ch zum Zitronensiiure- stoffwechsel wird das Aneurin in Beziehung gebra~cht. W~ihrend des B1-Mangels kommt es zu einer Abnahme der Zitronens~iureaussehei- dung sa, 84 auch nnahhihngig vo.m tIun~ger 85 Inj'iziertes Suecinat geht schlecht in Zitronensiiure fiber ss. Als Ursache wird ,der Ausfall der Goearboxylase angesehen, ,deren Bedeu~ung ffir die Deearbo.xylie- rungen im Zi:tronensiiurekreisprozef~ selbst gestmht wird. Anderseits s~teigert ein Zusatz yon B~ die Zitronensiiuresynthese aus Brenz- trau,bens~inre im Gewebe avitaminotis~eher Ratten s6.
Der Angriffspunkt des Insulins an der Oxalessigs~iure bringt es in nahe Beziehung zum Phosphatumsatz. Von den im Schema auf- gezeigten Wegen, die zu und yon tier Oxales,sigs~iure fiihven, ver- laufen alle unter Mitwirkung phosphorylierender Prozesse: Die De- hydrierung der Brenztraubensiiure, die Wood-Werkman-Reaktion 87, der Weg zur Phosphoroxalessigs~iure, die Reduktion zu Apfelsfmre ss und vielleicht auch der Weg zur Zitronensiiure fiber ein Aeetylphos- phat sg, 90. So kommt es mit jeder Intensivierung der Insulinwirkung zu ausgedehnten Phosphatverschiebungen. Besonders die gesteigerte Insulinaktivitiit in der Hypoxie fiihrt zu einer ungewShnlichen Be- wegu~g der Phosphatreserven des Organismus. Wir sehen bei Kohle- hy, dratassimilation im Sauerstoffmangel einen Sturz des anorgani-

Stoffweehsei und vegetative Regulation. 65
sehen Phosphates im Blute und ein fas:t v611iges Sehwinden der Harnphosphate ~9. Nhnli,ehe Phosphatsehiibe konnten wir weder d~ureh Siiuerung mit Chlorammonium noeh dureh Alkalisieren mit Bicarbonat erzielen 20, sondern nur durch Injektion yon Insulin 9~ Dal~ Insulin selbst nicht phosphoryliert, kann heute wohl ats fest- stehende Tatsaehe gelten 92
Unsere Untersuehungen belassen die Insulinwirkung am fiber- gang oon Glgkolgse zur Atmung, oder naeh der alten Formulierung, Insulin reguliert die Pas.,teur-Meyerhofsehe Reaktion, das Gleieh- gewieht zwisehen Zuekerauf- und -a.bbau. Das steht rail den ne~en Ansehauungen der Sehule Coris 93 ni.eht im Einklang, welehe die Insulinwirkung mit der Hexokinasea,ktivitiit in VerIoind~ng bringen. Die in alien Geweben vorhandene Hexokinase phosphoryliert freie Glukose zu Glukose-l-phos.phat. Naeh Cori hemmt die Hypo,physe diese Reakfio:n und Insulin beseitigt diese Hemmung. So besteehend diese Arbeitshypothese ist, so wird sie doeh dureh neue Arbeiten nut ungenfigend gestiitzt 94, 95, 96, 97. Die Hemmung der Hexo,kinase dureh Hypophysenextrakte ist inkonstant. Hemmungsfaktoren kommen aueh in andeeen Organen vor, beso.nders in der Milz. Nor.malfiere und Alloxantiere ~nterseheiden sieh nieht in der Hexo.kinaseaktivitiit ihrer M, us,kulatur, .die in bei4en Gruppen dutch Ins:ulin nieht ge- steigert wird. Kliniseh konnte die hyperinsuliniire Reaktion bet Insel- zellenadenomeu dutch Hypophysenextrakte nieht zuriiekgedriingt werden 9s.
Trotz vieler Einzelheiten bleibt .der grundlegende Prozefi, an dem das Insulin angreift, aueh wetter noeh im Dunkeln. Die Verkniipfung yon Oxalessigsiiure mit Essigsiiure ,dureh Citrogenase ist es sieher nieht. Viel eher noeh die reeht komplexe Bereitstellung und vor allem die Erhaltung einer bestimmten Oxalessigsdurekonzentration; denn Oxalessigsfiure geht Meht dureh Nebenreaktio,nen aus dem System verloven, so dab dann die Gewebsatmung sinkt. Solehe Neben~ege sind die Aminierung tier Oxalessigsiiure zu Asparaginsiiure, also ihr i)bergang in ei.n.en Eiweigbaustein, oder die Deearboxylierung der Oxalessigsfiure zu Brenztraubens/iure, set es spontan oder dureh ~-Coearboxylas:e. Brenztrau'bensiiure wiederum zerfiillt in Essigsiiure, die mit resflieher Oxalessigsiiure reagiert und fiber den oxydativen KreMauf umgesetzt wird, ein Cireulus vitiosus der Sel'bstverniehtung. DaB ein [3bersehug yon Oxalessigsiiure eine betrfiehtliehe Essigsfiure- vennehrung auslOsen kann, zeigt tier KetonkSrperanstieg im Gewebe unserer Versuehstiere naeh Belastung mit Aeetat-4-Oxalaeetat. Er fibertrifft die dutch Aeetat allein verursaehte Erh6hung um das Dop- pelte his Dreifaehe (Tab. 7 und 8).
Aet~ Neuroveget~iva, Bd. I~ Heft 1--2. 5

66 Th. Leipert:
Tab. 7. D e r K e t o n k 5 r p e r g e h a l t y o n R a t t e n o r g a n e n u n t e r w e c h s e l n d e n B e d i n g u n g e n .
Tier Nr.
18 20 23
24
25 15 22 38 45 44 14 19 21
Leber
rag% Gesamtaceton
5,8 6,2 4,1 2,4 2,7 2,7
5~9 3,1
4,1 2,8 6,9 9,5
Muskel Blur
8,5 2,5 2,0
2,1
1,8 11,0
:Niere
3.3
4,5
],0
Normal - - 24 Std. Hunger 24
3 Std. Hypoxie, 24 8td" Hunger, 61ykogezr 0,267~
3 Std. Hypoxie, 2 • Aeetat + Glukose, Olykogen 2,8800/0
ebenso G]ykogen 0,432o/o 1 • Aeetat -- 24 Std. Hunger
10,8 6,1 8,3
10,8 6,9 8,1 6~9
4,7 4,0 5,2 9,9 5,9 5,9 6,4
8,9 5,3 7,0
12,1 6,4
10,2 8,6
5,5 10,0 10,0 30,5
2 X ,, 24 . . . . 2 x ,, 24 1 • ,, - - Hunger - - Hypoxie 1 Std~ I • . . . . . . 2 ,, 1 X . . . . . . 5 ,,
/ 2 X ,, ,, - 4~ 2- 2 X ,, ,, ,, 3 ,,
Tab. 8. D e r K e t o n k S r p e r g e h a l t y o n R a t t e n o r g a n e n n a c h Be- : l a s t u n g m i t A c e t a t - F O x a l a c e t a t .
Tier Nr.
49 52 56 50
53
54 42
43
47
51
55
Leber
8 ,4
4,5 25,6
5,9
5.3
20,3 6,1
8,5
10,2
12,2
9,7
mg~/o @esamtaeeton
Muskel Blur
5,4 6,7 5,0 4,4
29,5 100,4 6,1 7,8
4,7 22,2
9,8 22,6 7,6 12,7
12,2 15fl
7,5
13,7
12,~
Niere
8,9 �9 5,9 26,9 13,1
59,8
37,5 9,2
19,2
19,8
15,5
53,9
2 • Aeet~t § 1 X 0ES (2 mg ZS/cm 2) 2 • ,, + 2 X O E S (2mgZS/em 2) 2 X . + 1 X 0ES (3,2 mgZS/cm 2) 2 • ,, + 1 X OE8 (2 mgZS/em-') Hypoxie 3 Std. 2 X ,, § 2 X 0ES (2 mg ZS/em~) I{ypoxie 3 Std.
ebenso 2 • hcetat + 1 • 0ES (3,2 nag ZS/em 2) Hypoxie - - Tod nach 2 Std.
ebenso Hylooxie - - Tod naeh 3/4 Std. 2 X Acetat + 2 • 0ES (2 mg ZS/cm s) H y p o x i e - Tod nueh 11/2 Std.
ebenso H y p o x i e - Tod nueh 13/4 Std. 2 • Acetat § 1 • 0ES (3,2 mg ZS/em 2) t I y p o x i e - Tod n'~eh 3/4 Std.
E s is t b e m e r k e n s w e r t , d a B u n t e r d i e s e n U m s t ~ n d e n lnsulinmange[ den Resgntheseweg in keiner Weise blockiert. So erkl~i r t s i eh d ie zu-
n f i ch s t f i b e r r a s e h e n d e T a t s a c h e , daft t i e r v o n Himwich a u f g e s t e l l t e

Stoffwechsel und vegetative Rulegati0n. 67
~Glukose-Milchsgiurezyklus 99 zwischen Muskel und Leber am pan- kreaslosen Tier genau so ungestSrt abliiuft wie am norma:len: Milch- siiure wird vom Muskel freigemacht und vonde r Leber verbraucht, die ihrerseits Glukose ausschfittet zur Verwertung im M'us,kel oder anderen Geweben oder auch zur Ausscheidung. Bei Alloxantieren ist die Glykogenbildung lficht gestSrt lo0. Dam}t 'ist in gleicher Weise tier •bergang yon Eiweifl in Kohlehydrat gesichert: Alanin (Serin, Gystin) - - ~ Brenztraubensiiure, Asparagi~ns~iure--~ Oxales.sigs~iure, GlutamiTls~iure (Hisfdin, Prol in)--~ Ketogl'utarsiiure. Gerade in einer FSrderung der Zuckerbildu~g aus Nichtzuckern sieht eine Reihe von Autoren die Hauptwirk'ung des Insulins. Ix der Tat si~kt der Amino- sfiurespiegel des Blutes nach Insulininjekfion.
Ffir den Fettstoffwechsel ~st der ungestSrte ~ber,gang von Glyko- lyse z'ur Atmung von grundlegender Bedeut~g. Der Fettstoffwechsel is den Organismus mit Essigs~iureresten aus der ~-Oxy- dation tier Fettsiiuren. Fehlt Oxalessigs~iure, der Katalysalor der Acetat~oxydafion, dann treten zwei Essigs~iurereste zu Acetatessig- siiure zusam'n~en, die teilweise reversibel in ~-Oxybutters~iure iiber- geht, te}lweise jedoch in Aceto~a zerfiillt. Die Ketonspaltur~g der Acet- essigsiiure ist nicht un~kehrbar. Man kann darin eme Venfilfunktion vermuten, da alas Aceton nicht nur durch den Ham, sondern auch durch die Atemwege ausgeschieden wird u~d so das (Jbermal~ einer Ketonk5rperanh~iufung wirksam begrenzt, ohne die riickl~iufige Syn- these yon Fettsiiuren aus C2-K5rpern in ihrer Kapazitiit einzu- schr~inken. Ketonk6rper entstehen also synthetisch aus Essi.gsiiure, wie in Iso~openversuchen nachgewiesen wurde, und nicht dutch Hemmung des Fettsiiureabbaues au~f der C4-Stufe. Diese Synthese unterbleibt, solange noch eine geringe Menge verffigbares Kohle- hydrat vorhanden ist, das fiber Brenztraubellsiiure Oxalessi~s~iure [iefert und so den normalen Acetatumsatz gewiihrleistet. Zucker w irkt daher antiketogen.
Noeh aaa ebaer anderen Stelle zeigt sieh die ,,katalyti.sche" Wirkung geringer Zuekermengen. Der Sauerstoff'mangel amterbrieht, wie wir zeigen konnten, den Oxydations.zy~zlus. Die Dehydrierung der ls:o- zitroner~s~iure unterbleibt und Zitronens~iure hiiuft sieh an:
Isozftronel~siiure -[- Codehydrase I I ~ -~ Ketoglutars~iure -[- CO 2 -+- -{- Codehydrase II H e.
Die hier wirksame Dehydrase ist Codehydrase II. Ist sie einmal reduziert, dann kann sie nur unter Mitwikung von Hexosemon0- p'hosphat reakfiviert werden. Beim Mangel einer leieht verffigbaren Kohlehydratreserve ist die Codehydrase II daher bald ersch6pft. Durch Verffitterung yon Kohlehydrat dagegen wird sie wieder auf-
5*

68 Th. Leipert :
geladen und die Blockade des Oxydationszyklus mit ihrer Anhii'uf'ung yon Zitronensiiure aufgehoben.
Es ist klar, dab die Umschaltung auf Assimilation nicht yore Insulin allein gesteuert wird, sondern dab dabei da:s endokrine System in seiner Gesamtheit, Hypophyse, Nebennie~en und Schilddriise, eine wichtige RoBe s pielt. Schon Evans 21 hatte gezeigt, da~ die Glykogen- bildung im Sauerst(~ffmangel n~ach Enffernung der Nebennieren a~s- bleibt. Dadurch fiillt das Erfolgsorgan der adreno,corticotropen Wir- kung der Hypophyse mit seinen gesam~en Stoffwechselfaktoren aus. Zwischen Insulin und diesen Fakt,oren besteht eine enge Wechsel- wirkung. Wenn wir als Test der adrenocorticotropen Wfi:kung die Vitamin-C-Aussch,fittung ~us der Nebenniere ansehen lol, da.nn treibt das Assimilafionshormon Insulin das Vitamin C wieder zuriick ins Gewebe und tier C=Gehalt des Blntes sinkt 102. Seit den Unter- suchungen Ashers weiB man, da{~ Ratten durch Entfernung der S,childdriise au~erordentlich h6henfest werden nnd niederen Saner- stoffdruck ausgezeiehnet ertragen 10a. Dassetbe erreicht man heute durch Th~o:uracil 104. Umgekehrt ma, chen Thyroxininjektionen und die Verffitterung von Schilddriisenpr~iparaten .die Versuchstiere hSchst empfindlieh gegen Sauerst~fffmangel. Wir kommen damit in alas un- gemein interessante Gebiet der h5henklimatischen Steuerung des Stoffwechsels. Sympaihikotrope Hyperthyreosen werden d urch HShenaufenthalt weitgehend gebessert. P~lsfrequenz und Ruhe- umsatz sinken und das K6rpergewi,cht nimmt zu, wie es fiir assiinila, torische Phasen charakteristisch ist. Leider versagr das Experiment bei geringer Abnahme der Sauersto,fftension, doch besteht ke~n Grund, die Gfiltigkeit der g eschilderten Steuerungsmechanistnen a'uch fiir diese Bedingungen anzuzweifeln.
Gewi~ ist die Hypoxie nicht der einzige steuernde Faktor des Stoff- wechsels. Die Nahrung selb.st wirk~ regulierend auf den Umsatz und die altbekann~e spezifisch dyn.amische Reaktion ist der Ausdruck dieser Substratsteuerung (Gremels), die bald pos~tiv dissimilierend, bald negat~v assimilierend auftritt, je nach Art und Menge des zu- gefiihrten Nahrungsstoffes 1. Doch die Nahrung ist wertlos, wen~ kein S~uerstoff zu ihrgm Umsatz bereitsteht. Daher stellten wir die ad~iquate Sauerstoffversorgung als regulierenden Faktor in den Mittelpunkt unserer Un~ersuchungen.
Zusammenfassung.
Ffir die hormonale Steuerm~g des Stoffwechse~s gilt auch weiter- h~n d~e alte Anschauung: Adrenalin ist der sympathischen Erregung zugeordnet und de r Wirks~off der Dissim~lationsprozesse, Insulin da- gegen ist das Parasympathikusho,rmon und der Wirkstoff der AsJsi-

Stoffweehsel und vegetative Regulation. 89
milafion. An der bu~gernden Ratte 15st Sa~erstoffm3ngel eine prompte assilnilatorische Phase arts mi t rascher Neubfldung von Glykogen in der Leber. Eiwei{~ kann als Muttersubsfanz des neu- gehildeten Zuckers ausgeschlossen werden. Dagegen gelingt es, von den Ahbauprodt~kten tier Fetts~i'uren a~sgehend, eine Verbindung mit dem oxydat iven Kohlehydratstoffwechsel herzustellen. Bet glykogen: armen Ratten tritt in der Hypoxie des Unterdruckkammerversuches nach Bel~st~n~g mit Acetat ein betr~ichtlicher Anstieg der Zilro~en- s~iure in B}ut, Mus,keln und Nieren a~f. Damit e r schdnt tier oft er- 5rterte 0bergang tier C2-Bruchstfi,cke ans der ~-Oxydation der Fett- sfiuren hinfiber in de~ Zitronen.s~iurezyklus tier Kohlehydratoxyda- tion fiir die Essigsiiure selbst erwiesen. Essigsii'ure kondensiert sich mit Oxalessigsfi,ure aus dem Kohlehydra tabbau zu Zitronensfiure. Diese Anhiiufung von Zitronensiiure l~i~ sich auch auBe~halb der Unterdruckkan~rmer erzielen, wenn man acetatgefiit terten Ras kleinste Insulindosen in Dauerinfusion zufiihrt. Belastet man jedoch Ratten in der Hypoxie mit eilmm Gemisch von Essigs~iure ~nd O~al- essigs~iure, darm gehen die Tiere im hypoglyk~imischen Scho,ck unter Drel~kr~impfen und Biutzuckerwerten vo.n 36 bis 77 rag% z'ugrunde. Wir s, chlie~en daraus, dal~ Insulin am ~bergang yon Glykolyse z~r Atm~ng eine entscheidende Rolle spielt. D,as ist die Stelle, wo Brenz- trauberlsfiure einerseits in Essigsfiure und CO e zerffillt, anderseits aber durch CO2-Aufnahme in Oxalessigsfiure fibergeht. Die Summe beider Zwischenprodt~kte ist Zitronens~iure. Der endogene Schwund an CO2 konnte von uns am Abfall des respirator'~schen Quotienten der Ver- suchstiere auf Werte bis 0,68 auch bewiesen werden. Man hatte diese extrem nieder.en "VVerte bisher dur,ch vermeh.rte 02-Retentio,n an die Fettsiiueeketten zu deuten versucht, was den Quotienten CO2/Oo er- niedrigen mulate..Sie galten als Beweis fiir den Obergang von Fett in Kohle.hydrat. D.m" endo,gene Verlu.st vo.n CO2 dnreh Bindu~g an Brenztraubensi iure und BBdung vo~ Oxalessigsiiure m'ul] jedo.ch in gleicher Weise den RQ. erni.edrigen, ja er f'iihrf dnrch W e ~ m h l n e des CO2 a'us dem Pufferungs.system NaHCOa/CO 2 z,um ~berwiegen des Bicarbonats zur Alkal.ose. D,ad'ur.eh wird die Alkalose der va,gotonen Stoffweehsellage ebenso verstiindlich wie das W.eehsels.pi.el: Insulin mach t Al.kalose und Alkalose versti~rkt die Ins'ulinwirkung. Klar wird auch die enge Bezi.eh'un,g des Insulins zum Pho.sphatumsatz und der Synergismus ,des Insulins mit Vitamin B1, das als Cofer'ment den Ab- bau der Brenztr.aubensiiure katalysiert un,d so der Insuli~wirkung unmit te lbar vorges.ehaltet ist.
Summary. As to the hormonal regulation of metabolism lhe old view is still valid: Adrem
alin belongs to the sympathetic stimulation and is the agent of the dissimilation pro-

7 0 Th. Leipert:
cess, whereas insulin is the hormone of the parasympathetic nervous system and the agent of assimilation. Hypoxia causes, in the hungry rat, a prompt assimilatory phase with rapid new formation of glycogen in the liver. Albumin can be excluded as mother substance of the newly developed sugar. By starting, however, from the catabolic products of fatty acids it is possible to form a connect ion with the oxydative metabolism of carbohydrates. In rats that are poor in glycogen the citric acid in blood, muscles an, d kidneys i s considerably increased in hypoxia of the underpressure chamber experiment after charging with acetate. Thus the frequently mentioned transit ion of the C~ fragments ~rom the ~ oxydation of ~the fatty acids into the citric acid cycle of the carbohydrate oxyda*tion for the acetic acid itself seems to be proved. Acetic acid condensates with oxalic acetic acid out of the carbohydrate dissimilation into citric acid. This accumulation of citric ~cid can also be achieved outside the underpressure chamber by per- manently infusing minimal doses of insulin rats fed with acelate. If, however, rats in hypoxia are charged with a mixture of acetic acid and oxalic acetate, then the animals die in hypoglycaemic shock with staggers and bloodsugar rates of 36 to 77 mg%. From this we infer that insulin plays a decisive part in the transition from glycolysis to respiration. Here pyro-racemic acid disintegrates on the one hand into acetic acid and CO;, but on the other hand by absorbing CO2 turns into oxalic acetate. The sum of these two intermediate products is citric acid. We were also able to prove 'the endogenous decrease of CO2 in the animals experimented upon by the fall of their respiratory quotient down to levels of 0,68. Up to now one had tried to explain these extremely low figures by increased O,~-retention to the fatty acid chains, which would decrease the quotient C0~/O2. They were considered proof of the transition of fat into carbohydrates. The endogenous loss of CO~ by agglutination to pyro-racemic acid and formation of oxalic acetate must, however, in the same way decrease the respiratory quotient (RQ), and even causes by taking away the bicar- bona te from the ,,buffersystem" NaHCOa/CO2 the preponderance of the bicar- bonate, to alcalosis. Thus the alcalosis of the vagotonic situation of metabolism be- comes as comprehensible as the reciprocal effect: insulin makes alcalosis, and alca- losls increases the action of insulin. This also explains the close relation of insulin ,to phosphate metabol ism and the synergism of insulin with vitamin B1 which is the immediate predecessor of the insulin effect by catalyzing as coferment the dis- similatiol] of pyro-racemic acid.
R6sum6.
E n ce qui concerne la r6gula t ion-hormouale du m6tabolisme, la conception continue d '6tre en vigueur que l',adr6naline est coordonn6e ~ l 'excitation sympa- thique comme substance hormonale des procbs de dissimilation, que l'insuline par contre, l 'hormone parasympathique, est la substance stimulative de l'assimi- la t ion Chez le rat affam6 la r6duction d'oxy@ne provoque promptement une phase assimilatrice avec une r6g6n6ration rapide de glycog~ne darts la foie. L'al- bumine ne peut pas 6tre consid6rge comme substance g6n6ratrice de la glucose r6cemment form6e. On r6ussit d'au~tre part, par tant des produits de dissimilation des acides gras, de trouver une connexit6 avec le m6tabolisme oxydatif des hydro- carbonates. Des rats pauvres en glycog~ne manifestent dans l 'hypoxie produite darts la chambre ~ d@ression, aprbs 6tre charg6s d'ac6tats, une augmentation remarquable de l 'acide citrique clans le sang, les muscles et les re ins . C'est ainsi qne paralt prouv6e pour l'acide ac6tique lui-m~me la transition souvent discut6e des fragments de C~ .de la ~-oxydation des acides gras vers le cycle de l'aclde citrique de l 'oxydation des hydrocarhonates. L'acide ac6tique se condense avee l'acide oxalac6tique de ~a .dissimilation hydrocarbon6e ~ l'ac~de cilrique. C'est aussi en dehors de la chambre ~ .d6pression qu 'on peut obtenir cette accumulation

Stoffweehsel und vegetative Regulation. 71
d'acide e~trique par infusion permanente de doses minimes d'insuline ~ des rats nourris d'ac6tate. En ehargeant eependant .des rats en 6tat d'hypoxie d'une mixture d'acide ae6tique et d'ae6tate oxalique, les animaux sueeombent au choe hypoglye6mique ayant des spasmes rotatoires et des titres glye6miques de 36 h 77mgr%. Nous en eoncluons que l'insuline a u n e importance d6eisive pour la transition de la glyeolyse fi la respiration. C'est l~t le lieu off l'aeide pyrurique se d6eompose ,d'une part en aeide ac6tique et CO_~, se transforme eependant d'autre part en aeide oxalae6fique par combinaison avee CO2. Le produit des deux substances interm6diaires est l'aeide eitrique. C'est ~t l'aide de la chute du quotient respir.atoire des animaux en exp6rienee jusqu'~t des litres de 0,68, que nous avons r6ussi h prouver la diminution en,dog~ne du CO2. On avait essay6 jusqu'5 pr6sent d'expliquer ees titres extr~mement bas par augmel~tation de la r6tention de O~ darts les eha~nes des aeides gras ce qui dezait abaisser le quotient CO~/O2. C'est eux qui 6taient consid6r6s eomme preuve de la transformation des corps gras en hydrocarbonates. La perte endogbne de C Oe eependant par combi- naison avee l'aeide pyrurique en formant d'aeide oxaloae6tique dolt ~ son tour de m6me abaisser le quotient respiratoire, plus encore, par l'61oignemel~t du CO~ du syst~me de r6gulation NaHCOs/CO~, elle mbne ~ la pr6pond6rance du bicar- bonate, ~ l'alealose. C'est ainsi qne nous eomprenons l'alealose de l'6tat vago- tonique du m6tabolisme aussi bien que l'effet, r6ciproque: L'insuline provoque de l'alealose et l'alealose intensifie l'effet de l'insuline. On eomprend ainsi la relation intime de l'insuline ,avee le m6tabolisme des phosphates et le synergisme de Fin- suline avee la vitamine Bt qui ealalyse eomme eoferment la dissimilation .de l'aeide pyrurique rangeant ainsi eomme pr6d6eesseur imm6diat de l 'aetion de l'insuline.
Riassunto.
Circa la regolazione ormonale del ricambio vale aneora il vecchio complesso ehe l'~drenalina esereiti una eeeitazione simpatiea quale sostanza ormonale dei processi di dissimilazione, e l 'insulina ,al contrario, rappresenti la sostanza ormo- nale parasimpa,tiea, dei processi di assimilazione. Nel topo affamato la manc anza di ossigeno provoc a una rapida fase assimilatoria, con raplda formazione di glico- geno nel fegato. L'albumina pub essere eseluso che rappresenti la sostanza di origine degli zuceheri neoformati. D'altra parte b possibile partendo dai prodotti di degradazione degli aeidi grassi, ,trovare un eollegamento con il ricambio ossi- dativo dei carboidrati. In topi privi di glicogeno sperimentando in ambiente privo di ossigeno, in camera a depressione, dopo carico con acetato si osserva nn note- vole aumento dell'acido CilTiCO nel sangue, nei muscoli, nel rene. Quindi sembra prova..ta la transizione spesso dise n s ~ dei frammenti di CO~, provenienti dalla ~-ossidazione degli acidi grassi verso il ciclo dell'acido citrico e della ossidazione dei carboidrati. L'acido acetico si eondensa con l'acido ossalioo proveniente dalla degradazione dei carboidrati ad aeido citrico. Cosi senza l'uso della camera a depressione si pub ottenere un accumulo di acido citrico se si sommilfistrano dosi minime di insulina (s.otto forma di infusione continua) a dei ,topi nutriti con acetato. Se si trat'tano pot dei topi in stato di ipoossia con una mlscela di acldo aeetieo ed aeido ossalieo, gli animali muoiono per sehoch ipoglicemieo con spasmi e con valori della glicemia da 36 e 77 rag%. Da cib not eoneludiamo the l'insulina h a una parte m.olto importante nel passaggio dalla glicolisi alla res,pL razione. Questo ~ il punto in cut l'acido piroraeemieo da una parte si frammen,ta in acido aeetieo e CO2, e d'altra parte si ha il passaggio ad acido ossalico pe~ assnnzione .di CO_,. La somma di ambedue i prodotti intermedi 6 l'acido eitrico. La diminuzione e~dogena di CO~ 6 s,tata dimostrata da not per mezzo della eaduta del quoziente respira}orio degli animali da esperimento a valori fino a 0,68. Finora

79~ Th. Leipert:
questi valori e s t remamente bassi erano spiegati con una aumenta ta r i tenzione di 02 alle ca t ene degli acidi grassi, che avrebbe por ta to ad una diminuzione del quoziente COe/O.~; quesfi valori valgono per il passaggio dai grassi ai carboidrati . La perdita endogena di COs per unione all 'acido piroracemico e successiva for- maz ione di acido ossalico por ta ugualmente ad una dimirmzione dei quoziente respirator io a l lontanando il C02 dal sistema t ampone NaHCO~/CO~. Da ci6 l 'alcalosi deIlo stato ~agotonico del metabol i smo diviene comprensibi le come pure il quadro reciproco: l ' insulina p rovoca l 'aicalosi e l 'alcalosi aumerrta per ef.fetto dell ' insulina. Diviene chiaro ~anche Io stretto rappor to dell 'insulina con il meta- bol ismo dei fosfati e i l sinergismo dell ' insulina con la vitanfirm B1 che lunge da catalizzatore come cofermento alla degrad~azione dell 'acido piroracemico, rap- p resen tando cosi l ' immediato predecessore dell 'insulina.
Literatur.
1. Gremels, H., Arch. exper. Path. (D.), 203 (1944)~ 225. - - 2. Wieland, H. u. Mit- arb., Ann. Chem. 542 (1939), 145. - - 3. Wood, H. G., u. C.H. Werkman, Biochem. J. (Brit.) 30 (1936), 48; 32 (1938), 1262; 35 (1940), 7, 1 2 9 . - 4. Wood, H.G., C. H. Werkman, A. Hemingway u. A. O. Nier, J. biol. Chem. (Am.) 139 (1941), 365, 377; 142, (1941), 31. - - 5. Slade, H.D., H. G. Wood, A. O. Nier, A. Hemingway u. C. H. Werkman, J. biol. Chem. (Am.) 143 (1942), 133. - - 6. Krebs, H. A., u. L. V. Eggle- ston, Biochem. J. (Brit.) 35 (1940), 1383. - - 7. Krebs, H. A., u. W. A. Johnson, Enzymologia (Nd.) 4 (19~7), 148. - - 8. Ochoa, S., J. biol. Chem. (Am.) 159 (1945), 243. - - 9. Ochoa, S., u. E. Weisz Tabori, J. biol. Chem. (Am.) 159 (1945), 245. --- 10. Lipton, M. A., u. E. S. G. Barron, J. biol. Chem. (Am.) 166 (1946), 367. - - 11. Lorber, V., A. Hemingwa!] u. A. O. Nier, J. biol. Chem. 151 (1943), 647. - - 12. Meyerhof, 0., P. Ohlmeyer, W. Gentner u. W. Maier-Leibnitz, Biochem. Z. 298 (1938), 396. - - 13. Lipmann, F., Advanc .Enzymol. 1 (1941), 99. - - 14. Kalckar, H. M., Chem. Rev. 28 (1941), 71. - - 15. Solomon A. K. u. Mitarb., J. biol. Chem. (Am.) 140 (1941), 171. - - 16. MacKay, E. M., H. O. Carne u. A. N. Wick, J. biol. Chem. (Am.) 133 (1940), 59. - - 17. Epstein, F. S., Chem. Zbl. 1942, I, 1525. - - 18 Rein, H., Nachr. Ges. Wissensch., GSttingen, Math. Physik, K. VI, Bd. 2, Nr. 1 4 . - 19. Lei- pert, Th., n. E. Kellersmann, Z. physiol. Chem. 276 (1942), 214, 233. - - 20. Leipert, Th., u. J. Meller, Z. physiol. Chem. 280 (1944), 101. - - 21. Evans, G. T., J. biol: Chem. (Am.) 105, XXVI. (1934). - - 22. Buchanan, J. M., A. B. Hastings u. F. B. Nesbett, J. biol. Chem. (Am.) 150 (1943), 413. - - 23. Choikof[, J. L., S. R. Lerner, C. Entenman u. W. G. Dauben, J. biol. Chem. (Am.) 174 (1948), 1045. - - 24. Schroet- ter, H. v., Verh. dtsch. Path. Ges. 1902. - - 25. Rosin, A., Beitr. path. Anat. 76 (1926), 153. - - 26. Lufl, U. C., Beitr. path. Anat. 98 (1937), 323. - - 27. Loewy, A., Biochem, Z. 185 (1927), 286. - - 28. Loewy, A., u. J. Leibowitz, Biochem. Z. 192 (1927), 67. - - 29. Monasterio, G., Biochem. Z. 218 (1930), 331. - - 30. Griffel, W., Biochem. Z. 222 (1930), 290. - - 31. Reichert, R., Helv. chim acta 27 (1944), 961. - - 32. Schoenheimer, R., u. W. M. Sperry, J. biol. Chem. (Am.) 106 (1934), 745. - - 33. Yasuda, M., J. biol. Chem. 94 (1931/32), 401. - - 34. Rof, Y., u. A. Thurnherr, Z. exper. Med. 88, (1933), 693. - - 35. Pucher, G. W., C. C. Sherman u. H. B. Vickery, J. biol. Chem. (Am.) 113 (1936), 235. - - 36. Carman, G. G., u. H. H. Mitchell, Amer. J. Physiol. 76 (1926), 380. - - 37. Wohl, A., u. P. Claussner, Ber. Chem. Ges. 40 (1907), 2308. - - 38. Orten, J. M., u. A. H. Smith, J. biol. Chem. (Am.) 128 (1939), 101. - - 39. Smith, A. H., u. J. M. Often, J. biol. Chem. (Am.) 124 (1938), 43. - - 40. Ffrth, 0., u. Mitarb., Bio- chem. Z. 269 (1934), 379. - - 41. Sherman, C. C., L. B. Mendel u. A. H. Smith, J. biol. Chem. (Am.) 113 (1936), 247, 265. - - 42. Often, J. M., u. A. H. Smith, J. biol. Chem. (Am.) 117, (1937), 555. - - 43. Markees, S. Klin. Wschr. 1941, 1260. - - 44. Corey, E. L., u. S. W. Britton, Amer. J: Physiol. 131, (1941), 783. - - 45. MacKay,

Stoffweehsel und vegetative Regulat ion. 73
E. M., u. Mitarb., J. biol. Chem. (Am.) 135 (1940), 157. - - 46. Wieland, H., u. Ch. Rosenthal, Ann. Chem. 555 (1943), 241. - - 47. Breusch, F. L., Science 97 (1943), 490. - - 48. Stern, J. R., u. S. Ochoa, J. biol. Chem. (Am.) 179 (1949), 491. - - 49. Altmann, F., Z. klin. Med. 115 (1930), 642. - - 50. Loewg, A., u. R. Helter, Z. exper. Med. 87 (1933), 22. - - 51. Staub, H., Pankreas , Be'thes H a n d b u c h d. n o r m . u. path. Physiol . XVI/1, ( I I /1) , Springer, Berlin, 1930. - - 52. Krebs, H. A., u. L. V. Eggteston , Biochem. J. (Brit.) 32 (1938), 913. - - 53. Rice, L., u. E. A. Evans, Science 97 (1943), 470. - - 54. Stadie, W. C., N. Haugaard u. M. Perlmutter, J. biol. Chem. (Am.) 172 (1948), 567. - - 55. AlwaIl, N., Acta reed. scand. 110 (1942), 476. - - 56. Pollak, L., E r g . inn. Med. 23 (1923), 337. - - 57. Glickman, N., u. E. Gellhorn, Amer. J. Phys io l . 121 (1938), 358. - - 58. Gelthorn, E., R. C. h~graham u. L. Moldavskg, J. Neurophys io l . 1 (1938), 301. - - 59. Dussik, K. Th., Neurochi rurgie 1944/45. - - 60. Gg6rgg, P., u. K. Vollmer, Biochem. Z. 140 (1923), 391. - - 61. Monasterio, G., Z. exper . Med. 81 (1932), 276. - - 62. Mandomsky, C., Z. Klin. Med. 111 (1929), 135. - - 63. Davies, H. W., J. B. S. Haldane u. E. L. Kennaway, Amer. J. Physiol . 54 (1920/21), 32. - - 64. J. Greenwald, J. biol. Chem. (Am.) 54 (1922), 285. - - 65. Dienst, C., Klin. Wschr . 1939, 1614; 1941, 858. - - 66. Fiirth, 0., Sitzg.~Ber. Akad. Wissensch . Wien, Akad. Anzg. Nr. 14 (1933). - - 67. Kugper, A. C., u. H. A. MatiIl, J. biol. Chem. ( A m . ) 1 0 3 (1933), 54. - - 68. Smith, A. H., u. Mitarb. , J. Nutr . 20 (1940), 255. - - 69. Olmsted, J. M. D., u. A. C. Taglor, J. biol. Chem. (Am.) 59, XXX (1923). - - 70. Holzer, H., u. O. Klein, Miinch. Med. Wsch r . 1928, 1284. - - 71. Hasselbaleh, K. A., u. J. Lindhard, Biochem. Z. 74 (1916), 1. - - 72. Warren, C. B. M. J. Physiol . 96 (1939), 34P . - - 73. Euler, H. o., u. B. H6gberg, Naturw. 1940, 29. - - 74. Nylin, G., H. v. Euler u. B. H6gberg, Kiln. Wschr . 1940, 433. - - 75. Mar- kees, S., u. F. W. Meyer, Exper ienf ia 4 (1948), 195. - - 76. Banga, J. S. Ochoa u. R. A. Peters, Biochem. J. (Brit.) 33 (1939), 1109. - - 77. Tislowitz, Klin. Wsehr . 1937, 226. - - 78. Magyar, J., Z. exper. Med. 104 (1938), 495. - - 79. Saka, M. 0., Schweiz. Med. Wschr . 1942, II , 1327. - - 80. Donner~ L., ~asop. ~esk. l~k. 81 (1942), 850. - - 81. Bueding, E., M.H. Stein u. H. Wortis, J. biol. Chem. (Am.) 140 (1041), 697. - - 82. Horn. Z., Klin. Wschr . 1941, 1032. - - 83. Smith, A. H., u. C. E. Meyer, J. biol. Chem. (Am.) 139 (1941), 227 - - 84. Simola, P. E., Aeta physiol , scand. 7 (1944), 115. - - 85. Sober, H.A., M. A. Lipton u. C. A. Elvehjem, J. biol. Chem. (Am.) 135 (1940), 605. - - 86. Guzmann, E. S., Baron u. Milarb. , J. biol. Chem. (Am.) 141 (1941), 957. - - 87. Utter, M. F., u. H. G. Wood, J. biol. Chem. (Am.) 160 (1945), 375. 88. Ruf]o, A., u. C. Cennamo, Boll. soe. ital. biol. sper. 20 (1945), 745. - - 89. Kaplan, N. 0., u. F. Lipmann, J. biol. Chem. (Am.) 174 (1948), 37. - - 90. Soodak, M., u. F. Lip- mann, J. biol. Chem. (Am.) 175 (1948), 999. - - 91. Me,let, J., u. Th. Leipert, Unver- 6ffentlichte Un te r suchungen . - - 92. Levine, R., S. D. Loube u. H. F. Weisberg, Amer. J. Physiol . 159 (1949), 107. - - 93. Colowick, S. P., G. T. Cori u. M. W. Slein, J. biol. Chem. (Am.) 168 (1947), 583. - - 94. Broh-Kalm, R. H., u. J. A. Mirskg, Science 106 (1947), 148. - - 95. Smith, R. H., Biochem. J. (Brit.) 45 Proc. X L I I (1949). - - 96. Bonetti, E., Na tu re 164 (1949), 957. - - 97. Stadie, W. C., u. N. Haugaard, J. biol. Chem. (Am.) 177 (1949), 311. - - 98. Corm u. Louis, J. Clin. Endocr in . 5 (1945), 220. - - 99. Him- wich, H. E., u. Milarb., J. biol. Chem. (Am.) 90 (1931), 417. - - 100. Weber, H., Nature 158 (1946) 627. - - 101. Long, C. N. H. Fed. Proc. 6 (1947), 461. - - t02. Sherrg u. Ralii~ J. Clin Invest ig 27 (M~irz) 1948. - - 103. Asher, L. u. H. Stdmpfli, Bioehem. Z. 185 (1927), 192. - - 104. Gordon, A. S., E. D. Goldsmith u. H. A. Charipper, Endo- crinol. 37 (1945), 223.
5~