Embed Size (px)
Citation preview

RNAi is a widespread eukaryotic regulatory processin which dsRNA triggers the silencing of cognate genes(Hannon 2002). The initiation of RNAi and relatedgene-silencing pathways requires cleavage of inputdsRNA into small RNA fragments, ranging from 21 to27 nucleotides in length, by the enzyme Dicer(Bernstein et al. 2001). The small RNAs are subse-quently bound by members of the Argonaute family ofproteins, which function as the core components of adiverse family of protein–RNA complexes called RISC(Hammond et al. 2001; Song et al. 2004). RISC andrelated complexes use the small RNAs as guides forsequence-specific gene silencing via translationalrepression (Pillai et al. 2005), mRNA degradation(Hammond et al. 2000), and heterochromatin formation(Verdel et al. 2004).
We previously reported the crystal structure of Dicerfrom the parasite G. intestinalis (MacRae et al. 2006).The Giardia Dicer is smaller than Dicer enzymes ofhigher eukaryotes largely because it lacks an amino-terminal helicase and carboxy-terminal dsRNA-bindingdomain (dsRBD). Giardia Dicer is an elongated moleculemeasuring approximately 100 Å long and 30–50 Å widewith a shape resembling a hatchet. The “blade” of themolecule is made up of two RNase III domains thatconnect to the PAZ domain by a long α-helix which spansthe length of the hatchet “handle.”
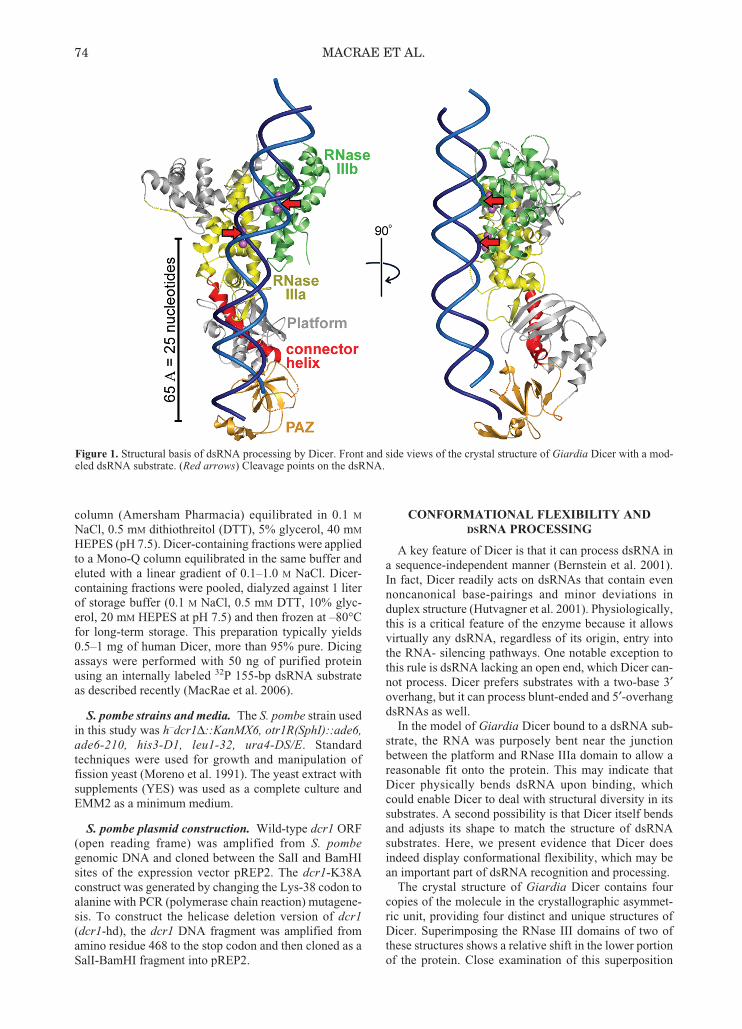
In our model of dsRNA processing, Dicer functions asa molecular ruler to produce dsRNA fragments 25nucleotides in length. Dicer works by recognizing theend of a dsRNA with its PAZ domain and then cuttingthe dsRNA with its RNase III domains (Fig. 1). The dis-tance between the PAZ and RNase III domains deter-mines the length of the small RNAs generated by Dicer.In the case of Giardia Dicer, this distance is 65 Å, whichleads to the production of small RNAs about 25nucleotides in length. On the basis of biochemical data,a similar model for processing of dsRNA was proposedfor human Dicer (Zhang et al. 2004).
METHODS AND MATERIALS
Structural analysis. The crystal structure of GiardiaDicer was taken from the Protein Data Bank (accessionnumber 2FFL), and C-alpha atoms of the A and B chaincopies were superimposed using lsqkab of the CCP4program suite. Normal mode analysis was performedusing the ElNémo interface (http://igs-server.cnrs-mrs.fr/elnemo/index.html).
Human Dicer expression and purification. HumanDicer was expressed in Sf-9 cells using a baculoviralsystem as described previously (Myers et al. 2003).Truncated human Dicer lacking the amino-terminalhelicase domain was generated by expressing residuesM605-S1922 using baculovirus as well. After 72 hours ofbaculoviral infection at 27°C, 1–2 liters of Sf-9 cells (1 x106 cells /ml) were harvested by gentle centrifugation andthen homogenized in 50 ml of lysis buffer containing 0.3 MNaCl, 0.5% Triton X-100, 5% glycerol, 10 mM imid-azole, 0.5 mM TCEP, 50 mM H2PO4 (pH 8.0), and 1 tabletof EDTA-free protease inhibitor cocktail (Roche).Insoluble material was removed by centrifugation, andthe soluble cell extract was then incubated with 5 ml ofNi-NTA resin (QIAGEN) for 30 minutes at 4ºC. Theresin was pelleted by gentle centrifugation and resus-pended in 50 ml of wash buffer (0.3 M NaCl, 5% glycerol,20 mM imidazole, 0.5 mM TCEP, 50 mM H2PO4 at pH8.0). The washed resin was pelleted again, and the washstep was repeated a total of five times. Protein was elutedfrom the Ni-NTA resin by resuspending washed resin in15 ml of elution buffer (0.3 M NaCl, 0.3 M imidazole, 5%glycerol, 0.5 mM TCEP, 50 mM H2PO4 at pH 8.0). Theeluted protein was dialyzed against 1 liter of Ni-NTAcolumn wash buffer at 4°C overnight in the presence ofTEV protease (Lucast et al. 2001), which removes theamino-terminal His6 tag engineered onto Dicer. Thedialyzed protein solution was then passed through a 5-mlNi-NTA column. Unbound material was collected,concentrated to 1 ml, and run over a Superdex 200 16/60
Structure of Dicer and Mechanistic Implications for RNAi
I.J. MACRAE,*‡ F. LI,* K. ZHOU,*‡ W.Z. CANDE,* AND J.A. DOUDNA*†‡§
*Department of Molecular and Cell Biology, †Department of Chemistry, ‡Howard Hughes Medical Institute,University of California, Berkeley, California 94720; §Physical Biosciences Division, Lawrence Berkeley
National Laboratory, Berkeley, California 94720
Dicer is a specialized ribonuclease that processes double-stranded RNA (dsRNA) into small RNA fragments about25 nucleotides in length during the initiation phase of RNA interference (RNAi). We previously determined the crystalstructure of a Dicer enzyme from the diplomonad Giardia intestinalis and proposed a structural model for dsRNA process-ing. Here, we provide evidence that Dicer is composed of three structurally rigid regions connected by flexible hinges andpropose that conformational flexibility facilitates dsRNA binding and processing. We also examine the role of the acces-sory domains found in Dicers of higher eukaryotes but absent in Giardia Dicer. Finally, we combine the structure ofDicer with published biochemical data to propose a model for the architecture of the RNA-induced silencing complex(RISC)-loading complex.
Cold Spring Harbor Symposia on Quantitative Biology, Volume LXXI. © 2006 Cold Spring Harbor Laboratory Press 978-087969817-1 73
073-080_MacRae_Symp71.qxd 2/6/07 3:23 PM Page 73
column (Amersham Pharmacia) equilibrated in 0.1 M
NaCl, 0.5 mM dithiothreitol (DTT), 5% glycerol, 40 mM
HEPES (pH 7.5). Dicer-containing fractions were appliedto a Mono-Q column equilibrated in the same buffer andeluted with a linear gradient of 0.1–1.0 M NaCl. Dicer-containing fractions were pooled, dialyzed against 1 literof storage buffer (0.1 M NaCl, 0.5 mM DTT, 10% glyc-erol, 20 mM HEPES at pH 7.5) and then frozen at –80°Cfor long-term storage. This preparation typically yields0.5–1 mg of human Dicer, more than 95% pure. Dicingassays were performed with 50 ng of purified proteinusing an internally labeled 32P 155-bp dsRNA substrateas described recently (MacRae et al. 2006).
S. pombe strains and media. The S. pombe strain usedin this study was h–dcr1Δ::KanMX6, otr1R(SphI)::ade6,ade6-210, his3-D1, leu1-32, ura4-DS/E. Standardtechniques were used for growth and manipulation offission yeast (Moreno et al. 1991). The yeast extract withsupplements (YES) was used as a complete culture andEMM2 as a minimum medium.
S. pombe plasmid construction. Wild-type dcr1 ORF(open reading frame) was amplified from S. pombegenomic DNA and cloned between the SalI and BamHIsites of the expression vector pREP2. The dcr1-K38Aconstruct was generated by changing the Lys-38 codon toalanine with PCR (polymerase chain reaction) mutagene-sis. To construct the helicase deletion version of dcr1(dcr1-hd), the dcr1 DNA fragment was amplified fromamino residue 468 to the stop codon and then cloned as aSalI-BamHI fragment into pREP2.
CONFORMATIONAL FLEXIBILITY ANDDSRNA PROCESSING
A key feature of Dicer is that it can process dsRNA ina sequence-independent manner (Bernstein et al. 2001).In fact, Dicer readily acts on dsRNAs that contain evennoncanonical base-pairings and minor deviations induplex structure (Hutvagner et al. 2001). Physiologically,this is a critical feature of the enzyme because it allowsvirtually any dsRNA, regardless of its origin, entry intothe RNA- silencing pathways. One notable exception tothis rule is dsRNA lacking an open end, which Dicer can-not process. Dicer prefers substrates with a two-base 3′overhang, but it can process blunt-ended and 5′-overhangdsRNAs as well.
In the model of Giardia Dicer bound to a dsRNA sub-strate, the RNA was purposely bent near the junctionbetween the platform and RNase IIIa domain to allow areasonable fit onto the protein. This may indicate thatDicer physically bends dsRNA upon binding, whichcould enable Dicer to deal with structural diversity in itssubstrates. A second possibility is that Dicer itself bendsand adjusts its shape to match the structure of dsRNAsubstrates. Here, we present evidence that Dicer doesindeed display conformational flexibility, which may bean important part of dsRNA recognition and processing.
The crystal structure of Giardia Dicer contains fourcopies of the molecule in the crystallographic asymmet-ric unit, providing four distinct and unique structures ofDicer. Superimposing the RNase III domains of two ofthese structures shows a relative shift in the lower portionof the protein. Close examination of this superposition
74 MACRAE ET AL.
Figure 1. Structural basis of dsRNA processing by Dicer. Front and side views of the crystal structure of Giardia Dicer with a mod-eled dsRNA substrate. (Red arrows) Cleavage points on the dsRNA.
073-080_MacRae_Symp71.qxd 2/6/07 3:23 PM Page 74

reveals that Dicer is composed of three structurally rigidregions connected by flexible hinges. The “RNase region”is made up of both RNase III domains and the bridgingdomain. The “platform region” is composed of the plat-form domain and the connector helix, and the third regionis the PAZ domain (Fig. 2, left).
The most dramatic difference between the two structuresis the shift in position of the PAZ domain, which movesabout 5 Å. The hinge for this movement (hinge 1) can betraced to a single point in the connector helix where thereis a distinct kink in the helix (Fig. 2, right). The kink ismost likely induced by the presence of Pro-266, which sitsin the apex of the kink. Comparison of Dicer sequencesfrom a variety of organisms shows that proline is strictlyconserved at this position in the primary sequence (MacRaeet al. 2006). Thus, a kinked connector helix is likely astructural feature of all Dicer enzymes. The strict conser-vation of proline at this position argues that the connectorhelix kink and the hinge it forms between the platform andPAZ regions have an important role in Dicer function,most likely facilitating conformational flexibility.
The second hinge in Dicer resides between the plat-form and RNase regions. Unlike the connector helix kink,there is no distinct point in the polypeptide chain thatforms the hinge between the two regions. Instead, thepositional shift is diffused along the platform loop and theloop following the connector helix. The movementbetween the two structures in this region is a 5° rotationof the RNase region around the long axis of the molecule,which approximately coincides with the connector helix.
In addition to comparing static crystal structures ofGiardia Dicer, we also explored possible conformationalflexibility in the protein by normal mode analysis (Suhreand Sanejouand 2004). This analysis predicts similarstructurally rigid regions of Dicer connected by hinges in
approximately the same locations as described above. Thelargest positional shifts in the analysis were between theRNase and platform regions, with the RNase predicted toboth rotate and bend with respect to the platform (Fig. 3).
Structural analysis of Giardia Dicer reveals that theenzyme possesses a substantial degree of conformationalflexibility. We propose that the three structural regions ofDicer flex and rotate relative to each other as the enzymeengages its dsRNA substrates. This is analogous to the“induced fit” classically seen in enzyme-active siteswhen bound to small-molecule substrates, except that inthe case of Dicer, which has a substrate more than 80 Ålong, the induced fit spans the length of the protein.
Flexibility is likely a critical feature of Dicer because itallows the enzyme to adjust its shape to accommodatestructural diversity in its dsRNA substrates. This is particu-larly important in the processing of pre-microRNAs (pre-miRNAs), which are typically riddled with RNA helicalimperfections (Pasquinelli et al. 2000; Lau et al. 2001). Theability of Dicer to cope with deviations from ideal RNAduplex structure allows pre-miRNAs to function as imper-fect duplexes. Evolutionarily, this is an important pointbecause it means that mutations on both sides of miRNAprecursors will be tolerated, allowing nature to create newmiRNAs from old as well as to adjust thermodynamicasymmetry across the pre-miRNA and thereby optimizingthe guide:passenger strand ratio that is incorporated intomiRNPs. Thus, conformational flexibility in Dicer’s struc-ture allows evolutionary flexibility in miRNA function.
ROLES FOR ACCESSORY DOMAINS IN DICER
The structure of Giardia Dicer represents the func-tional core structure that is likely to be common to allDicer enzymes (Zhang et al. 2004). This relatively small
Figure 2. Conformational diversity in Giardia Dicer crystal structures. (Left) Giardia Dicer is composed of three structurally rigidregions termed RNase, Platform, and PAZ. These regions are connected by two flexible hinges in the protein. (Right) Close-up viewof the connector helix. Hinge1 resides at a kink in the connector helix that is likely formed by Pro-266.
STRUCTURE OF DICER AND RNAI 75
073-080_MacRae_Symp71.qxd 2/6/07 3:23 PM Page 75

and simple protein is fully capable of dicing dsRNA invitro and is sufficient to support RNAi in vivo (MacRaeet al. 2006). However, several of the domains commonlyfound in Dicer proteins of higher eukaryotes are not pres-ent in the Giardia enzyme. Most notably, Giardia Dicerlacks the amino-terminal DExD helicase present inhuman, fly, worm, plant, and fission yeast Dicers.Although the presence of the Dicer helicase is broadlyconserved among higher eukaryotes, no function has yetbeen assigned to this domain. Possible activities of theDicer helicase domain include unwinding of productsiRNAs, translocating Dicer along dsRNA substrates,and facilitating the handoff of substrates or products withother components of the RNAi machinery.
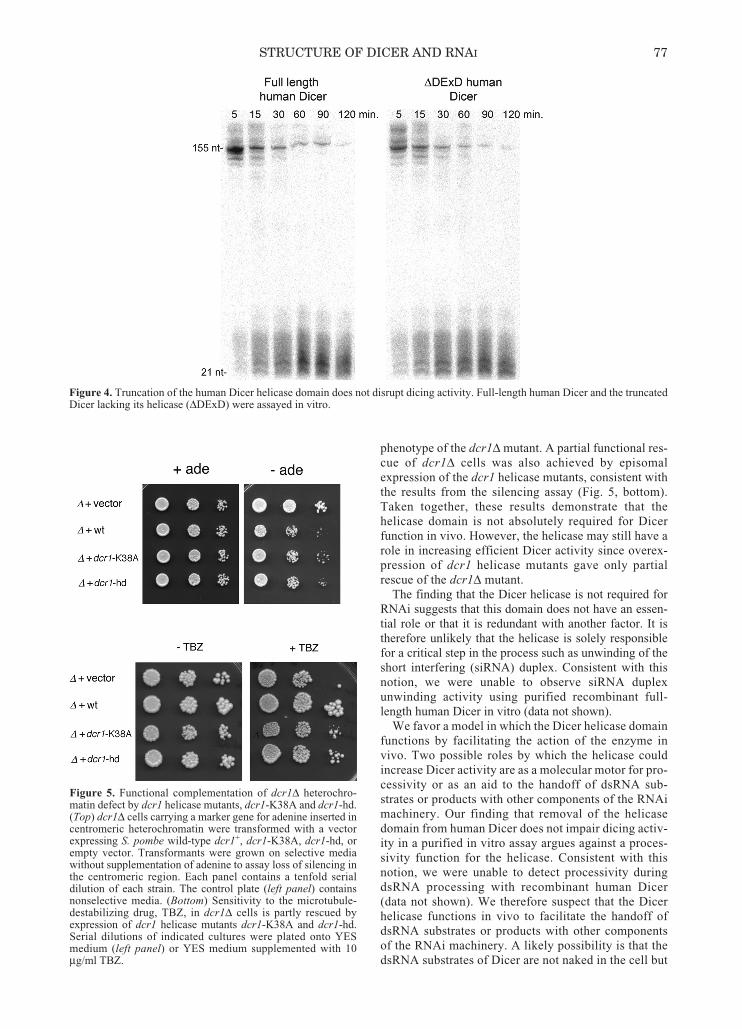
We examined the role of the helicase in dicing by pro-ducing a version of human Dicer that lacks the entirehelicase. The truncated protein was stable and wellbehaved, indicating that the core structure of Dicer wasnot perturbed by the removal of the helicase.Furthermore, the truncated Dicer displayed activity com-parable to that of the wild-type protein in an in vitrodicing assay in both the presence and absence of ATP(Fig. 4). Thus, in the absence of other cellular factors, thehelicase domain does not facilitate processing of dsRNAby human Dicer. This finding is consistent with previousresults which showed that mutation of the ATP-bindingWalker A motif did not impair the activity of purifiedrecombinant human Dicer (Zhang et al. 2002).
To investigate the in vivo function of the Dicer heli-case domain, we produced a mutant version ofSchizosaccharomyces pombe Dicer in which Lys-38, thehelicase Walker A motif lysine, is changed to alanine
(named dcr1-K38A). Mutation of the analogous lysineresidue eliminates ATP binding and hydrolysis in theDExD helicase eIF4AIII (Shibuya et al. 2006). Thismutation should therefore render the Dicer helicasecatalytically inactive while leaving the helicase domainstructurally intact. We also created a version of S. pombeDicer lacking the entire helicase domain (named dcr1-hdfor helicase deletion).
pREP2 expression plasmids carrying either the wild-type S. pombe Dicer or one of the two mutants were intro-duced into a Dicer deletion strain of S. pombe. The S.pombe strain used also contained the marker gene ade6 atthe heterochromatic otr region of the centromere.Transformants were isolated and plated in serial dilutionson Edinburgh minimal medium (EMM) without adenine.Because silencing of the marker gene ade6 was disrupted,the dcr1Δ strain grew well on the plate. As expected, thestrain transformed by the vector with wild-type Dicerdisplayed poor growth, indicating that ectopic expressionof Dicer rescues the dcr1Δ centromeric silencing defect(Fig. 5, top). The two strains carrying the helicasemutants, dcr1-K38A and dcr1-hd, displayed a slowergrowth rate than the dcr1 deletion strain, although not asslow as the wild type, indicating that even the Dicerlacking a helicase domain imparts partial functionalcomplementation to the dcr1Δ mutant.
To further characterize the S. pombe Dicer mutants,sensitivity to the microtubule-destabilizing drug,thiabendazole (TBZ), was investigated. dcr1Δ cellsdisplay a strong sensitivity to TBZ, presumably due toincreased chromosome missegregation. The vector withwild-type dcr1+ fully rescued the TBZ sensitivity
76 MACRAE ET AL.
Figure 3. Comparison of the two lowest-frequency normal modes calculated for Giardia Dicer. In each panel, the crystal structure isshown in blue, and the yellow structure represents the conformational change predicted by the analysis. Structures were aligned intheir Platform regions, and arrows indicate direction of relative movements of RNase and PAZ regions.
073-080_MacRae_Symp71.qxd 2/6/07 3:23 PM Page 76
phenotype of the dcr1Δ mutant. A partial functional res-cue of dcr1Δ cells was also achieved by episomalexpression of the dcr1 helicase mutants, consistent withthe results from the silencing assay (Fig. 5, bottom).Taken together, these results demonstrate that thehelicase domain is not absolutely required for Dicerfunction in vivo. However, the helicase may still have arole in increasing efficient Dicer activity since overex-pression of dcr1 helicase mutants gave only partialrescue of the dcr1Δ mutant.
The finding that the Dicer helicase is not required forRNAi suggests that this domain does not have an essen-tial role or that it is redundant with another factor. It istherefore unlikely that the helicase is solely responsiblefor a critical step in the process such as unwinding of theshort interfering (siRNA) duplex. Consistent with thisnotion, we were unable to observe siRNA duplexunwinding activity using purified recombinant full-length human Dicer in vitro (data not shown).
We favor a model in which the Dicer helicase domainfunctions by facilitating the action of the enzyme invivo. Two possible roles by which the helicase couldincrease Dicer activity are as a molecular motor for pro-cessivity or as an aid to the handoff of dsRNA sub-strates or products with other components of the RNAimachinery. Our finding that removal of the helicasedomain from human Dicer does not impair dicing activ-ity in a purified in vitro assay argues against a proces-sivity function for the helicase. Consistent with thisnotion, we were unable to detect processivity duringdsRNA processing with recombinant human Dicer(data not shown). We therefore suspect that the Dicerhelicase functions in vivo to facilitate the handoff ofdsRNA substrates or products with other componentsof the RNAi machinery. A likely possibility is that thedsRNA substrates of Dicer are not naked in the cell but
STRUCTURE OF DICER AND RNAI 77
Figure 4. Truncation of the human Dicer helicase domain does not disrupt dicing activity. Full-length human Dicer and the truncatedDicer lacking its helicase (ΔDExD) were assayed in vitro.
Figure 5. Functional complementation of dcr1Δ heterochro-matin defect by dcr1 helicase mutants, dcr1-K38A and dcr1-hd.(Top) dcr1Δ cells carrying a marker gene for adenine inserted incentromeric heterochromatin were transformed with a vectorexpressing S. pombe wild-type dcr1+, dcr1-K38A, dcr1-hd, orempty vector. Transformants were grown on selective mediawithout supplementation of adenine to assay loss of silencing inthe centromeric region. Each panel contains a tenfold serialdilution of each strain. The control plate (left panel) containsnonselective media. (Bottom) Sensitivity to the microtubule-destabilizing drug, TBZ, in dcr1Δ cells is partly rescued byexpression of dcr1 helicase mutants dcr1-K38A and dcr1-hd.Serial dilutions of indicated cultures were plated onto YESmedium (left panel) or YES medium supplemented with 10μg/ml TBZ.
073-080_MacRae_Symp71.qxd 2/6/07 3:23 PM Page 77
are bound to a variety of cellular factors. The helicasedomain of human Dicer may help remove or repositionthese factors and allow Dicer to bind and process theRNA. Indeed, the DExD RNA helicases NPH-II andDED1 have been shown to catalyze protein displace-ment from RNA (Jankowsky et al. 2001; Fairman et al.2004). This model is also consistent with the findingthat in HeLa cell extracts, Dicer requires ATP to bindexogenous siRNAs (Pellino et al. 2005), whereas thepurified recombinant human Dicer readily bindssiRNAs in the absence of ATP (Zhang et al. 2002;Pellino et al. 2005). Furthermore, ATP has been shownto stimulate Dicer activity in crude mouse embryonalcarcinoma cell lysates (Billy et al. 2001). On the otherhand, Drosophila Dcr-2 requires ATP to processdsRNA, even when purified to near homogeneity (Liuet al. 2003). Likewise, immunopurified Dcr-1 fromCaenorhabditis elegans shows ATP-dependent dicingactivity (Ketting et al. 2001). It therefore appears thatthe Dicer helicase may serve multiple functions that areemphasized differentially in Dicer enzymes fromdifferent organisms.
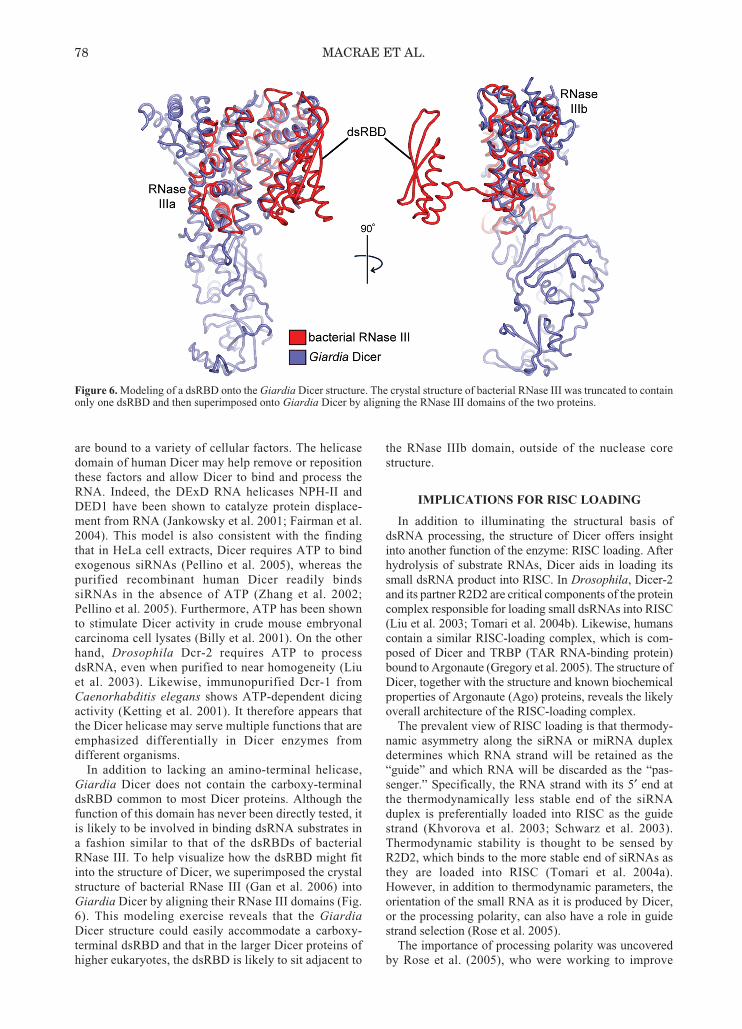
In addition to lacking an amino-terminal helicase,Giardia Dicer does not contain the carboxy-terminaldsRBD common to most Dicer proteins. Although thefunction of this domain has never been directly tested, itis likely to be involved in binding dsRNA substrates ina fashion similar to that of the dsRBDs of bacterialRNase III. To help visualize how the dsRBD might fitinto the structure of Dicer, we superimposed the crystalstructure of bacterial RNase III (Gan et al. 2006) intoGiardia Dicer by aligning their RNase III domains (Fig.6). This modeling exercise reveals that the GiardiaDicer structure could easily accommodate a carboxy-terminal dsRBD and that in the larger Dicer proteins ofhigher eukaryotes, the dsRBD is likely to sit adjacent to
the RNase IIIb domain, outside of the nuclease corestructure.
IMPLICATIONS FOR RISC LOADING
In addition to illuminating the structural basis ofdsRNA processing, the structure of Dicer offers insightinto another function of the enzyme: RISC loading. Afterhydrolysis of substrate RNAs, Dicer aids in loading itssmall dsRNA product into RISC. In Drosophila, Dicer-2and its partner R2D2 are critical components of the proteincomplex responsible for loading small dsRNAs into RISC(Liu et al. 2003; Tomari et al. 2004b). Likewise, humanscontain a similar RISC-loading complex, which is com-posed of Dicer and TRBP (TAR RNA-binding protein)bound to Argonaute (Gregory et al. 2005). The structure ofDicer, together with the structure and known biochemicalproperties of Argonaute (Ago) proteins, reveals the likelyoverall architecture of the RISC-loading complex.
The prevalent view of RISC loading is that thermody-namic asymmetry along the siRNA or miRNA duplexdetermines which RNA strand will be retained as the“guide” and which RNA will be discarded as the “pas-senger.” Specifically, the RNA strand with its 5′ end atthe thermodynamically less stable end of the siRNAduplex is preferentially loaded into RISC as the guidestrand (Khvorova et al. 2003; Schwarz et al. 2003).Thermodynamic stability is thought to be sensed byR2D2, which binds to the more stable end of siRNAs asthey are loaded into RISC (Tomari et al. 2004a).However, in addition to thermodynamic parameters, theorientation of the small RNA as it is produced by Dicer,or the processing polarity, can also have a role in guidestrand selection (Rose et al. 2005).
The importance of processing polarity was uncoveredby Rose et al. (2005), who were working to improve
78 MACRAE ET AL.
Figure 6. Modeling of a dsRBD onto the Giardia Dicer structure. The crystal structure of bacterial RNase III was truncated to containonly one dsRBD and then superimposed onto Giardia Dicer by aligning the RNase III domains of the two proteins.
073-080_MacRae_Symp71.qxd 2/6/07 3:23 PM Page 78

siRNA design for commercial research purposes. It hadpreviously been shown that upon transfection into mam-malian cells, 27-nucleotide dsRNAs are processed byDicer into approximately 21-nucleotide products as theyenter the RNAi pathway (Kim et al. 2005). These Dicer-substrate small RNAs are 100 times more potent inducersof gene silencing than the “prediced” 21-nucleotidesiRNAs traditionally used in gene knockdown experi-ments, presumably because dsRNA cleavage by Dicer iscoupled to RISC loading (Gregory et al. 2005; Maniatakiand Mourelatos 2005). However, a disadvantage of 27-nucleotide dsRNAs is that they can be processed by Dicerfrom either end of the duplex, resulting in a variety ofpossible 21-nucleotide siRNA products and a higherchance for off-target effects. Rose et al. (2005) found thatblunt-ended dsRNAs with DNA bases on the last twonucleotides of the 3′ ends are poorly processed by humanDicer. This finding allowed the design of 27-nucleotidedsRNA substrates that are blocked at one end, forcingDicer cleavage to result in a single desired product.
The motivation for this work was to improve rationaldesign of Dicer-substrate reagents, but the finding alsoallowed inspection of the relationship between dsRNAprocessing polarity and guide strand selection. Rose et al.(2005) prepared pairs of 27-nucleotide dsRNA Dicersubstrates designed to produce the same final 21-nucleotide siRNA duplex, but with opposite processingpolarity. In seven out of nine cases, silencing of a reportergene was more efficient when the 5′ end of the strand anti-sense to the reporter mRNA was positioned at the blockedend of the 27-nucleotide Dicer substrate. Because the twosubstrates result in identical siRNA duplexes upon cleav-age by Dicer, differences in silencing efficiency are inde-pendent of thermodynamic asymmetry and most likelydue entirely to processing polarity. These results are con-sistent with the early studies of Elbashir et al. (2001) inDrosophila extracts, which also correlated dsRNA pro-cessing polarity with selective target strand cleavage.
These results have several interesting implicationswhen considered together with the structure of Dicer.First, the observation that Dicer processing can beblocked by adding two DNA bases to a blunt-endeddsRNA suggests that the human Dicer PAZ domaincannot bind dsRNA modified in this way. Second, andmore importantly, these results reveal that in the absenceof thermodynamic asymmetry, the RNA strand that hasits 3′ end bound to the PAZ domain of Dicer after cleav-age is much more likely to be incorporated as the guidestrand into RISC (Fig. 7, left). Along with the knowledgethat in RISC, the 3′ end of the guide strand is bound to thePAZ domain of Argonaute (Song et al. 2003; Ma et al.2004), this suggests that the siRNA duplex is passed fromthe PAZ domain of Dicer to the PAZ domain ofArgonaute (Fig. 7, right). Thus, a likely model for theRISC-loading complex is one in which the PAZ domainsof Dicer and Argonaute sit adjacent to one another andthe RNase III domains of Dicer sit opposite the PIWIdomain in Argonaute (Fig. 7). TRBP, which has beenshown to interact with both Dicer and Argonaute(Chendrimada et al. 2005; Lee et al. 2006), may residebetween the two where it could function in a mannersimilar to that of its Drosophila ortholog R2D2 in sensingsiRNA thermodynamic asymmetry as the duplex ispassed from Dicer to Argonaute.
CONCLUSIONS
The crystal structure of Dicer has revealed how this fam-ily of specialized ribonucleases measures and producesdsRNAs of specific lengths that function in various gene-silencing pathways. Furthermore, the structure has enableddetailed models for Dicer recognition of imperfectly base-paired substrates and for its interactions with other key pro-teins involved in RNAi. These models can now be testedexperimentally, and provide a framework for investigatingthe activities of different Dicer enzymes in vivo.
STRUCTURE OF DICER AND RNAI 79
Figure 7. A model for the architecture of the RISC-loading complex. (Left) Silencing of a target gene is more effective when the 3′end of the antisense strand in the siRNA duplex is bound to the Dicer PAZ domain. (Right) A model of the RISC-loading complexwith Dicer and Argonaute PAZ domains adjacent to each other could explain the observed relationship between processing polarityand guide strand selection.
073-080_MacRae_Symp71.qxd 2/6/07 3:23 PM Page 79

ACKNOWLEDGMENTS
The authors thank the Life Sciences ResearchFoundation for a postdoctoral fellowship to I.J.M.; thiswork was supported in part by a grant from the NationalInstitutes of Health to J.A.D.
REFERENCES
Bernstein E., Caudy A.A., Hammond S.M., and Hannon G.J.2001. Role for a bidentate ribonuclease in the initiation stepof RNA interference. Nature 409: 363.
Billy E., Brondani V., Zhang H., Muller U., and Filipowicz W.2001. Specific interference with gene expression induced bylong, double-stranded RNA in mouse embryonal teratocarci-noma cell lines. Proc. Natl. Acad. Sci. 98: 14428.
Chendrimada T.P., Gregory R.I., Kumaraswamy E., Norman J.,Cooch N., Nishikura K., and Shiekhattar R. 2005. TRBPrecruits the Dicer complex to Ago2 for microRNA processingand gene silencing. Nature 436: 740.
Elbashir S.M., Lendeckel W., and Tuschl T. 2001. RNA inter-ference is mediated by 21- and 22-nucleotide RNAs. GenesDev. 15: 188.
Fairman M.E., Maroney P.A., Wang W., Bowers H.A., GollnickP., Nilsen T.W., and Jankowsky E. 2004. Protein displace-ment by DExH/D “RNA helicases” without duplex unwind-ing. Science 304: 730.
Gan J., Tropea J.E., Austin B.P., Court D.L., Waugh D.S., andJi X. 2006. Structural insight into the mechanism of double-stranded RNA processing by ribonuclease III. Cell 124: 355.
Gregory R.I., Chendrimada T.P., Cooch N., and Shiekhattar R.2005. Human RISC couples microRNA biogenesis and post-transcriptional gene silencing. Cell 123: 631.
Hammond S.M., Bernstein E., Beach D., and Hannon G.J. 2000.An RNA-directed nuclease mediates post-transcriptionalgene silencing in Drosophila cells. Nature 404: 293.
Hammond S.M., Boettcher S., Caudy A.A., Kobayashi R., andHannon G.J. 2001. Argonaute2, a link between genetic andbiochemical analyses of RNAi. Science 293: 1146.
Hannon G.J. 2002. RNA interference. Nature 418: 244.Hutvagner G., McLachlan J., Pasquinelli A.E., Balint E., Tuschl
T., and Zamore P.D. 2001. A cellular function for the RNA-interference enzyme Dicer in the maturation of the let-7 smalltemporal RNA. Science 293: 834.
Jankowsky E., Gross C.H., Shuman S., and Pyle A.M. 2001.Active disruption of an RNA-protein interaction by aDExH/D RNA helicase. Science 291: 121.
Ketting R.F., Fischer S.E., Bernstein E., Sijen T., Hannon G.J.,and Plasterk R.H. 2001. Dicer functions in RNA interferenceand in synthesis of small RNA involved in developmentaltiming in C. elegans. Genes Dev. 15: 2654.
Khvorova A., Reynolds A., and Jayasena S.D. 2003. FunctionalsiRNAs and miRNAs exhibit strand bias. Cell 115: 209.
Kim D.H., Behlke M.A., Rose S.D., Chang M.S., Choi S., andRossi J.J. 2005. Synthetic dsRNA Dicer substrates enhanceRNAi potency and efficacy. Nat. Biotechnol. 23: 222.
Lau N.C., Lim L.P., Weinstein E.G., and Bartel D.P. 2001. Anabundant class of tiny RNAs with probable regulatory roles inCaenorhabditis elegans. Science 294: 858.
Lee Y., Hur I., Park S.Y., Kim Y.K., Suh M.R., and Kim V.N.2006. The role of PACT in the RNA silencing pathway.EMBO J. 25: 522.
Liu Q., Rand T.A., Kalidas S., Du F., Kim H.E., Smith D.P., andWang X. 2003. R2D2, a bridge between the initiation and effec-tor steps of the Drosophila RNAi pathway. Science 301: 1921.
Lucast L.J., Batey R.T., and Doudna J.A. 2001. Large-scalepurification of a stable form of recombinant tobacco etchvirus protease. Biotechniques 30: 544.
Ma J.B., Ye K., and Patel D.J. 2004. Structural basis foroverhang-specific small interfering RNA recognition by thePAZ domain. Nature 429: 318.
MacRae I.J., Zhou K., Li F., Repic A., Brooks A.N., CandeW.Z., Adams P.D., and Doudna J.A. 2006. Structural basisfor double-stranded RNA processing by Dicer. Science 311:195.
Maniataki E. and Mourelatos Z. 2005. A human, ATP-independent, RISC assembly machine fueled by pre-miRNA.Genes Dev. 19: 2979.
Moreno S., Klar A., and Nurse P. 1991. Molecular geneticanalysis of fission yeast Schizosaccharomyces pombe.Methods Enzymol. 194: 795.
Myers J.W., Jones J.T., Meyer T., and Ferrell J.E., Jr. 2003.Recombinant Dicer efficiently converts large dsRNAs intosiRNAs suitable for gene silencing. Nat. Biotechnol. 21:324.
Pasquinelli A.E., Reinhart B.J., Slack F., Martindale M.Q.,Kuroda M.I., Maller B., Hayward D.C., Ball E.E., Degnan B.,Muller P., et al. 2000. Conservation of the sequence and tem-poral expression of let-7 heterochronic regulatory RNA.Nature 408: 86.
Pellino J.L., Jaskiewicz L., Filipowicz W., and Sontheimer E.J.2005. ATP modulates siRNA interactions with an endoge-nous human Dicer complex. RNA 11: 1719.
Pillai R.S., Bhattacharyya S.N., Artus C.G., Zoller T., CougotN., Basyuk E., Bertrand E., and Filipowicz W. 2005.Inhibition of translational initiation by Let-7 MicroRNA inhuman cells. Science 309: 1573.
Rose S.D., Kim D.H., Amarzguioui M., Heidel J.D.,Collingwood M.A., Davis M.E., Rossi J.J., and Behlke M.A.2005. Functional polarity is introduced by Dicer processingof short substrate RNAs. Nucleic Acids Res. 33: 4140.
Schwarz D.S., Hutvagner G., Du T., Xu Z., Aronin N., andZamore P.D. 2003. Asymmetry in the assembly of the RNAienzyme complex. Cell 115: 199.
Shibuya T., Tange T.O., Stroupe M.E., and Moore M.J. 2006.Mutational analysis of human eIF4AIII identifies regions nec-essary for exon junction complex formation and nonsense-mediated mRNA decay. RNA 12: 360.
Song J.J., Smith S.K., Hannon G.J., and Joshua-Tor L. 2004.Crystal structure of Argonaute and its implications for RISCslicer activity. Science 305: 1434.
Song J.J., Liu J., Tolia N.H., Schneiderman J., Smith S.K.,Martienssen R.A., Hannon G.J., and Joshua-Tor L. 2003. Thecrystal structure of the Argonaute2 PAZ domain reveals anRNA binding motif in RNAi effector complexes. Nat. Struct.Biol. 10: 1026.
Suhre K. and Sanejouand Y.H. 2004. ElNemo: A normal modeweb server for protein movement analysis and the generationof templates for molecular replacement. Nucleic Acids Res.32: W610.
Tomari Y., Matranga C., Haley B., Martinez N., and ZamoreP.D. 2004a. A protein sensor for siRNA asymmetry. Science306: 1377.
Tomari Y., Du T., Haley B., Schwarz D.S., Bennett R., CookH.A., Koppetsch B.S., Theurkauf W.E., and Zamore P.D.2004b. RISC assembly defects in the Drosophila RNAimutant armitage. Cell 116: 831.
Verdel A., Jia S., Gerber S., Sugiyama T., Gygi S., Grewal S.I.,and Moazed D. 2004. RNAi-mediated targeting of het-erochromatin by the RITS complex. Science 303: 672.
Zhang H., Kolb F.A., Brondani V., Billy E., and FilipowiczW. 2002. Human Dicer preferentially cleaves dsRNAs attheir termini without a requirement for ATP. EMBO J. 21:5875.
Zhang H., Kolb F.A., Jaskiewicz L., Westhof E., and FilipowiczW. 2004. Single processing center models for human Dicerand bacterial RNase III. Cell 118: 57.
80 MACRAE ET AL.
073-080_MacRae_Symp71.qxd 2/6/07 3:23 PM Page 80






















![core.ac.uk · 2017-03-09 · double-stranded RNA: dsRNA), 5’-ppp dsRNA, and poly AdT (p-AdT; as DNA:RNA duplex) as indices of ISG56 induction [15]. Having checked several human](https://img.pdfslide.net/doc/110x75/5f07f1427e708231d41f8907/coreacuk-2017-03-09-double-stranded-rna-dsrna-5a-ppp-dsrna-and-poly-adt.jpg)






