Embed Size (px)
Citation preview

ZOOTAXAISSN 1175-5326 (print edition)
ISSN 1175-5334 (online edition)Copyright © 2013 Magnolia Press
Zootaxa 3626 (3): 381–390 www.mapress.com/zootaxa/ Article
http://dx.doi.org/10.11646/zootaxa.3626.3.6http://zoobank.org/urn:lsid:zoobank.org:pub:DD0BD375-D98F-464B-A352-00B04BA8434C
A new species of Gamasiphis Berlese (Acari: Ologamasidae) from North Asia, with a key to the Eurasian species
IRINA I. MARCHENKOInstitute of Systematics and Ecology of Animals, Russian Academy of Sciences, Siberian Branch, Frunze str. 11, Novosibirsk 630091, Russia. E-mail: [email protected]
Abstract
Gamasiphis angaridis sp. n. is described from females and males collected from litter and soil in the North Asian part of Russia–Siberia and the Far East. This is the first species of the large genus Gamasiphis to be described from the northern Palaearctic Region. A key for the separation of females of the 11 recognisable species of Gamasiphis described in Eurasia is provided.
Key words: Mesostigmata, edaphic mites, taxonomy
Introduction
The family Ologamasidae Ryke is a large group of predatory mites encountered in soil, humus and compost (Lindquist et al., 2009). The family is much richer in genera and species in tropical to subantarctic regions of the Southern Hemisphere than in Northern Hemisphere (Lindquist et al., 2009). Gamasiphis is one of the most diverse genera of Ologamasidae, with 72 described species (Castilho et al., 2010a, 2012). Studies by Beaulieu & Walter (2007) and Castilho et al. (2010b) indicate that Gamasiphis species may feed on Nematoda, Collembola and other soft-bodied arthropods.
Ten species of Gamasiphis have been described or recorded from Eurasia. Four species are known from Europe—G. pulchellus (Berlese, 1887) was recorded from North Italy, Belgium, Bulgaria and Spain (Berlese, 1904; Van Daele, 1975; Deltshev et al., 1998; Moraza & Peña, 2005); G. gandensius Van Daele, 1975 from Belgium, G. sextus Vitzthum, 1921 and G. lanceolatus Karg, 1987 from Germany.
Seven species are known from Asia: G. pulchellus from Caucasus, China, Japan (Lee, 1970; Bregetova, 1977; Ma & Ma, 2000, Caucasus specimens in O. Makarova's collections); G. productellus Berlese, 1923, G.novipulchellus Ma & Yin, 1998, G. aduncus Ma, 2004 and G. turgicalcareus Ma, 2009 from China (Berlese, 1923; Ma & Yin, 1998; Ma, 2004; 2009), G. bengalensis Bhattacharyya, 1966 and G. indicus Bhattacharyya, 1978 from India (Bhattacharyya, 1966, 1978). Apart from its occurrence in Europe and Asia, G. pulchellus has also been recorded from Egypt and South Africa (Lee, 1970; Zaher, 1986; Halliday, 2005; El-Banhawya et al., 2006), USA, Mexico and Bermuda (Ewing, 1920; Hennessey & Farrier, 1988; Minor & Norton, 2004), and Java (Castagnoli & Pegazzano, 1985). The objective of this paper is to provide description of a new species of Gamasiphis from the North Asian part of Russia and a key to Eurasian species of this genus.
Material and methods
Samples of soil and litter were collected for many years (1972–2011) by numerous collectors in different regions of the North Asian part of Russia – Altai Mountains, Kemerovo Region, north of Krasnoyarskii Krai, Amur Region, Khabarovskii Krai, Primorskii Krai, Sakhalin and Kuril Islands (Kunashir and Shikotan). Mites were extracted using a Berlese funnel and mounted in Hoyer’s medium. The specimens of Gamasiphis were separated and examined under a microscope Zeiss Axioscop 40, and were found to belong to a new species.
Accepted by B. Halliday: 31 Jan. 2013; published: 14 Mar. 2013 381

Taxonomically relevant structures were illustrated with the use of a Canon Power Shot G11 camera and measured with the use of a graded ocular. Measurements of each structure are given in micrometres (µm) and as a range (or a single value when measurement did not vary) representing the variation among all individuals examined. The dorsal and ventral idiosomal nomenclature of setae is based on Lindquist & Evans (1965) as adapted by Silva et al. (2007) and Castilho et al. (2010a) for the Ologamasidae. The idiosomal adenotaxy and poroidotaxy are based on Johnston & Moraza (1991).
The key provided for the separation of 11 species of Gamasiphis of Eurasia was prepared based on the original descriptions, available redescriptions of the concerned species and on the key to world species of Gamasiphis of Castilho et al. (2012).
Gamasiphis Berlese
Gamasiphis Berlese, 1904: 261. Type species: Gamasus pulchellus Berlese, 1887, by original designation.Gamasiphis.―Berlese, 1906: 101; Berlese, 1914: 137; Bregetova, 1977: 308; Lee, 1970: 42; Karg, 1990: 321; Karg, 1993: 169;
Castilho et al., 2012: 1970.Ologamasellus (Micriphis) Berlese, 1914: 140. Type species: Gamasiphis gamasellus Berlese, 1913, by monotypy. Synonymy
by Lee (1970).Ologamasus (Micriphis).―Baker & Wharton, 1952: 73.Micriphis.―Ryke, 1962: 160.Gamasiphis (Heteroiphis) Trägårdh, 1952: 55. Type species: Gamasiphis (Heteroiphis) arcuatus Trägårdh, 1952, by original
designation. Synonymy by Lee (1970).Heteroiphis.―Ryke, 1962: 160; Bhattacharyya, 1968: 530.Neogamasiphis Trägårdh, 1952: 57. Type species: Neogamasiphis hamifer Trägårdh, 1952, by original designation. Synonymy
by Lee (1970).
Diagnosis. Epistome with three anterior extensions, the anteromedial extension aciculate, spatulate or lanceolate, smooth or serrate, usually longer than the others. Dorsal shield entire, fused anteriorly with peritrematic shields and posteriorly with ventri-anal shield. Pre-sternal area with 0–3 pre-sternal shields. Sternal shield bearing four pairs of setae, totally fused to metasternal and endopodal shields; extending posterolaterally around genital shield. Genital shield with convex anterior margin, truncate posteriorly, bearing one pair of setae. Peritrematic shield fused to metapodal and exopodal shield near coxa IV. Exopodal shields in two pairs – a small subtriangular pair between coxae I–II (not always clearly distinct) and a large elongated pair reaching from midlevel of coxa II to midlevel of coxa IV, the latter shield separated from the peritrematic shield by a strip of unsclerotised cuticle.
Gamasiphis angaridis sp.n.
Diagnosis of adults (female and male). Anteromedial extension of epistome aciculate; all idiosomal setae aciculate; podonotal region of dorsal shield with 23 pairs of setae; opisthonotal region with 12 pairs of setae; seta j2 about twice as long as j1; seta j4 about 0.9 times as long as distance between its base and base of j5; seta z6 about as long as j6; seta s6 about 0.9 times as long as z6; seta j6 about 0.7 times as long as distance between its base and base of J2; four pairs of J setae; two pairs of pre-sternal shields; ventri-anal shields with eight pairs of setae in addition to para-anal setae (JV1-JV5, ZV1-ZV3); seta ZV2 as long as distance between its base and base of ZV3; seta JV5 about 5–6 times as long as para-anal setae; setae JV3 and JV5 situated posterior to unsclerotised line that partially separates dorsal and ventri-anal shields; distance between ends of these lines shorter than or equal to distance between bases of JV3 or these lines fully fused in some specimens.
Female. (Figs 1–7) (35 specimens measured).Gnathosoma: Fixed cheliceral digit 65–70 µm long with seven teeth in addition to apical tooth and a setiform
pilus dentilis (Fig. 1). Movable cheliceral digit 63–65µm long, with four teeth in addition to apical tooth. Dorsal cheliceral seta, lateral (antiaxial) and dorsal lyrifissures distinct. Epistome with anteromedian extension smooth and aciculate and with a pair of short anterolateral spines (Figs 2 A–B). Deutosternal groove of hypostome with
MARCHENKO382 · Zootaxa 3626 (3) © 2013 Magnolia Press
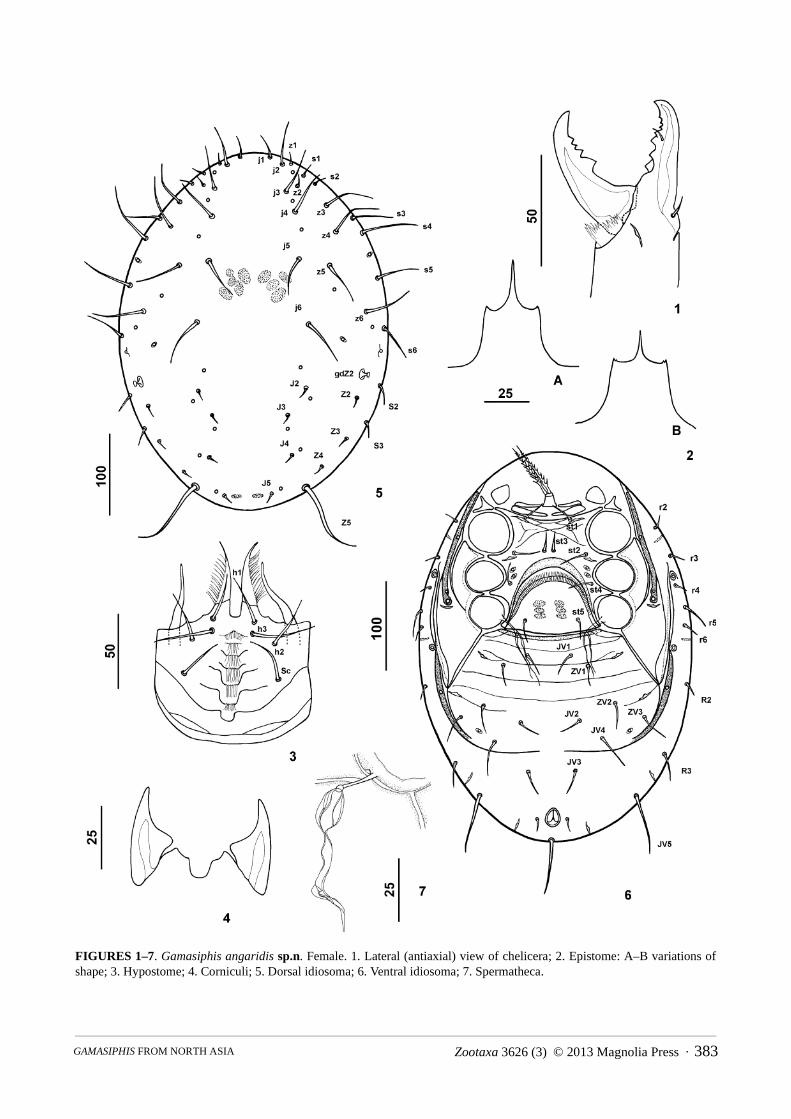
FIGURES 1–7. Gamasiphis angaridis sp.n. Female. 1. Lateral (antiaxial) view of chelicera; 2. Epistome: A–B variations of shape; 3. Hypostome; 4. Corniculi; 5. Dorsal idiosoma; 6. Ventral idiosoma; 7. Spermatheca.
Zootaxa 3626 (3) © 2013 Magnolia Press · 383GAMASIPHIS FROM NORTH ASIA

FIGURES 8–13. Gamasiphis angaridis sp. n. Male. 8. Lateral (antiaxial) view of chelicera: A–B; 9. Corniculi; 10. Palpa; 11. Ventral idiosoma; 12. Leg IV; 13. Leg II.
MARCHENKO384 · Zootaxa 3626 (3) © 2013 Magnolia Press

seven rows each bearing 7–14 denticles, except most basal row smooth (Fig. 2); anterior-most row shaped as an inverted “V”, subsequent rows roughly transverse; margins of deutosternum not distinct. Setae h1, h2 and Sc (h4) equal in length (32–35 µm), seta h3 slightly longer than other (35–37 µm). The salivary styli well developed (Fig. 3). Internal malae fimbriate laterally. Corniculi 38–40 µm long, 17–18 µm wide at the widest point (Fig. 4). Palp chaetotaxy 2-5-6-14-15; palp trochanter provided with one small ventral protuberance; setae al1 and al2 of palp genu slightly stout; palp apotele 3-tined.
Dorsal idiosoma: Dorsal shield entire, ovoid, smooth, totally covering dorsal surface, 425–480 µm long, 320–350 µm wide at level of coxa IV (Fig. 5). Dorsal shield with 35 pairs of acicular setae. Podonotal region with 23 pairs of setae (j1-j6, z1-z6, s1-s6, r2-r6) and with nine pairs of distinguishable lyrifissures and three pairs of pores (posterior and slightly mediad to s6, between r3-r4 and posterior to and mediad of r6); with numerous sigilla posterior to j5. Opisthonotal region with 12 pairs of setae (J2-J5, Z2-Z5, S2-S3, R2-R3) and with six pairs of distinguishable lyrifissures and two pairs of pores, including gdZ2 especially pronounced (anterior and mediad to S2) and one pair of pores on narrow diagonal sclerotised section of dorsal shield. Length of setae: j1 (17–20), j2 (40–45), j3 (40–45), j4 (45–50), j5 (45–50), j6 (45–50), z1 (7–10), z2 (7–10), z3 (45–50), z4 (45–50), z5 (45–50), z6 (45–50), s1 (7–10), s2 (7–10), s3 (40–45), s4 (60–65), s5 (40–45), s6 (40–45), r2 (10–12), r3 (10–12), r4 (10–12), r5 (25–30), r6 (7–10), J2 (7–10), J3 (7–10), J4 (7–10), J5 (7–10), Z2 (12–15), Z3 (12–15), Z4 (12–15), Z5 (85–90), S2 (12–15), S3 (12–15), R2 (12–15), R3 (20–25).
Ventral idiosoma (Fig. 6): Base of tritosternum 18–20 µm long and 20–22 µm wide proximally, laciniae (80–100 µm) totally separated from each other, pilose. Two pairs of pre-sternal shields present, slightly fused laterally. Sternal shield reticulate anteriorly between st1 and st2, smooth posteriorly; 70–75 µm long at mid-line and 135–140 µm wide between coxae II and III; with four pairs of setae (st1, st2, st4 acicular; st3 stout, inserted slightly anterior or slightly posterior or mesad to st2) and four pairs of lyrifissures. Peritreme extending anteriorly to midlevel of coxa I. Peritrematic shield fused with section of exopodal shield near to coxa IV, widest at level of posterior margin of coxa IV, with a lyrifissure posterior to stigma. Length of peritrematic-exopodal shield from stigma to posterior margin 100 µm, width 50 µm at level of posterior margin of coxa IV. Band of dorsal shield extending laterad to the fused peritrematic-exopodal shield ending sharply in posterior margin. Genital shield wider than long, 80–90 µm long, 110–125 µm wide, abutting the sternal shield by hyaline apex; anterior margin convex and posterior margin truncate, with a pair of setae st5 and three pairs of sigilla, distance between st5-st5 65–67 µm. Ventri-anal shield with transverse striations anteriorly to ZV2 and smooth posteriorly; 230 µm long from anterior margin to post-anal seta, 250 µm wide at widest point, with eight pairs of acicular opishogastral setae (JV1-JV5, ZV1-ZV3) in addition to post-anal and para-anal setae, and with four pairs of lyrifissures (anterior to and laterad of ZV1, in transverse line and laterad of ZV2, posterior to and mediad of ZV3 and posterior to and laterad of para-anal seta). Post-anal seta about 5–6 times as long as para-anal setae, the latter usually situated at level of the posterior third or at the midlevel of anal opening. Dorsal and ventri-anal shields partly separated by an unsclerotised line; the distance between the ends of this line is shorter or equal to distance between bases of JV3 or connected into a single fused line in some specimens; region posterior of this line 95 µm long; seta JV3 posterior to this line. Narrow sclerotised diagonal section laterad of ventrianal shield that connects the latter to the dorsal shield with one pair of pores; 7–12 µm wide at the level of pore; ending sharply at anterior part. Length of ventral setae: st1 (25–30), st2 (25–30), st3 (20–25), st4 (20–25), st5 (20–25), JV1 (25–27), JV2 (25–27), JV3 (25–30), JV4 (37–40), JV5 (65–75), ZV1 (25–30), ZV2 (25–32), ZV3 (25–32), para-anal (12–15) and post-anal seta (60–75).
Spermatheca: Opening part of spermathecal apparatus tubular, extending medially from base of coxa IV (Fig.7). Other parts of spermatheca not clearly visible.
Legs: Lengths: I: 380–400, II: 237–288, III: 258–270, IV: 340–365 µm. Chaetotaxy of legs I–IV: coxa 2, 2, 2, 1; trochanter 6, 5, 5, 5; femur (2 3/2 2/2 2), (2 3/1 2/2 1), (1 2/1 2/0 0), (0 2/2 2/0 0); genu (2 3/2 3/1 2), (2 3/1 2/1 2), (2 2/1 2/0 1), (2 2/1 3/0 0); tibia (2 3/2 3/2 2), (2 2/1 2/1 2), (2 1/1 2/1 1), (2 1/1 3/1 1). All leg setae acicular, except one antero-lateral pilose seta on trochanter II. All legs with pretarsus, elongate ambulacral stalk, a pair of strongly sclerotised claws and rounded pulvillus.
Male. (Figs 8–13) (24 specimens measured). Gnathosoma: Fixed cheliceral digit 45 µm long, with five teeth in addition to apical tooth, with transverse line
across the digit and a setiform pilus dentilis (Fig. 8 A–B). Movable cheliceral digit 50 µm long, with one tooth in addition to apical tooth. Spermatodactyl 50–55 µm long, curved, widest at proximal part and gradually narrowing apically; with an internal canal along its entire length. Dorsal cheliceral seta, lateral (antiaxial) and dorsal
Zootaxa 3626 (3) © 2013 Magnolia Press · 385GAMASIPHIS FROM NORTH ASIA

lyrifissures distinct. Corniculi 37–38 µm long, 10 µm wide; with extension at inner side (Fig. 9). Epistome and hypostome as in female. Setae h1, h2 and Sc equal length (30–33 µm), h3 slightly longer than other (35–37 µm). Palps of male as in female (Fig. 10).
Dorsal idiosoma: Dorsal shield 415–460 µm long, 275–330 µm wide, similar to that of female. Measurements of setae: j1 (17–20), j2 (35–40), j3 (35–40), j4 (35–40), j5 (35–40), j6 (35–40), z1 (7–10), z2 (7–10), z3 (35–40), z4 (35–45), z5 (35–45), z6 (35–45), s1 (7–10), s2 (7–10), s3 (30–35), s4 (50–55), s5 (40–45), s6 (35–40), r2 (10–12), r3 (10–12), r4 (10–12), r5 (25–30), r6 (7–12), J2 (7–10), J3 (7–10), J4 (7–10), J5 (7–10), Z2 (10–12), Z3 (10–15), Z4 (10–12), Z5 (55–70), S2 (12–15), S3 (12–15), R2 (12–15), R3 (20–25).
Ventral idiosoma: Base of tritosternum 12–15 µm long and 20–22 µm wide proximally, with pilose lacinae (70–75 µm) (Fig. 11). Except for the fusion of sternal and genital shields (sternitigenital shield), shape, pattern and fusions of ventral shields as in female. Sternitigenital shield reticulate between st1 and st2, smooth posteriorly; 117–120 µm long, 137–145 µm wide at widest point between coxae II and III; with five acicular setae (st1–st5), distance st1–st1 55 µm, st2–st2 75 µm, st3–st3 90 µm, st4–st4 90 µm and st5–st5 90 µm; with four pairs of lyrifissures; genital opening on anterior margin of shield, flanked by two rounded flaps, with elongate pointed extensions. Ventri-anal shield 225–235 µm long from anterior margin to post-anal seta and 237–250 µm wide at widest point; with eight pairs of setae (JV1–JV5, ZV1–ZV3) in addition to post-anal and para-anal setae; and with four pairs of lyrifissures (anterior to and laterad of ZV1, anterior to and laterad of ZV2, posterior to and mediad of ZV3 and posterior to and laterad of para-anal seta); post-anal seta about 4–5 times as long as para-anal seta. Length of ventral setae: st1 (25–30), st2 (25–30), st3 (20–25), st4 (20–25), st5 (20–25), JV1 (25–27), JV2 (25–27), JV3 (25–30), JV4 (27–37), JV5 (50–65), ZV1 (25–30), ZV2 (35–37), ZV3 (20–25), para-anal (12–15) and post-anal seta (50–65).
Legs: Lengths: I: 400–437, II: 287–295, III: 263–285, IV: 360–368 µm. Chaetotaxy of legs similar to that of female. All setae of legs I, III, IV acicular (Fig. 12). Legs II with one antero-lateral pilose seta on trochanter (similar to female); femur with two ventral spur-like setae: one large spur 40 µm length, in shape of twisted “long ear” and one small spur on elevated base; genu with two small ventral spur-like setae; tibia with one small ventral spur; tarsus with one small anterolateral spur near distal margin of basitarsus and with one acicular ventral seta on raised base (Fig. 13). All legs with pretarsus, elongate ambulacral stalk, a pair of strongly sclerotised claws and rounded pulvillus similar to those of female.
Material examined. Holotype—female, Russia, South Siberia, North-Eastern Altai, Turochak district, environs of Kebezen village (51°55' N, 87°06' E), Pinus sylvestris—Abies sibirica—Betula pubescens forest, in litter, 22 June 2007, coll. I. Marchenko. Paratypes—6 females on the same slide as holotype; 38 females, Altai, environs of Kebezen village, Pinus sylvestris forest, 11 August 2006, coll. I. Marchenko; 11 females, 2 males, Altai, environs of Obogo village (51°30' N, 87°18' E), Abies sibirica—Pinus sibirica forest, 6 August 2007, coll. I. Marchenko; 5 females, 2 males, Altai, environs of Saidyp village (52°34' N, 86°34' E), Betula pubescens—Populus tremula forest, 22 August 2006, coll. I. Marchenko; 2 females, 1 male; 2 females, 1 male, Middle Siberia, Krasnoyarskii Krai, environs of Turukhansk (65°47' N, 87°58' E), Picea sibirica—Pinus sibirica forest, 26 July 2003, coll. A. Babenko; 2 females, 1 male, Far East of Russia, Khabarovskii Krai, Grand Khekhtsir ridge (48°12' N, 134°52' E, 800 m a.s.l.), dark coniferous forest, 7 June 2011, coll. V. Dubatolov; 1 females, 6 males, Khabarovskii Krai, Nikolaevsk-na-Amure District, estuary of Amur River, environs of Arkhangelskoe village (53°05' N, 140°59' E), mixed forest with Quercus mongolica, 22 September 2009, coll. V. Dubatolov; 2 females, 3 males, Khabarovskii Krai, environs of Chegdomyn village (51°07' N, 132°58' E), Larix gmelinii forest, 18 July 2005, coll. I. Lyubechansky; 9 females, 2 males, Primorskii Krai, Sikhote-Alin Ridge (46°59' N, 136°53' E), June 2009, coll. D. Berman.
Holotype is deposited in Zoological Museum of Institute of Systematics and Ecology of Animals, Novosibirsk, Russia; 20 paratypes (17 females and 3 males) are kept in arthropods collection of Manchester Museum, Manchester, United Kingdom; other paratypes are deposited in Zoological Museum of Institute of Systematics and Ecology of Animals, Novosibirsk, Russia.
Other examined material: Russia, South Siberia: 240 females, 8 males, Altai, different districts (51°13'– 52°34’ N, 85°33' – 87°48' E); 5 females, Kemerovo Region, Mountain Shoria (53°20' N, 87°11' E); specimens were collected in litter and soil of South Siberian forests with Larix sibirica, Pinus sibirica, P. sylvestris, Abies sibirica, Picea abovata, Betula pubescens, Populus tremula, in the nests of Formica polyctena; Far East of Russia: 4 females, Amur Region (55°19' N, 123°09' E); 9 females, Khabarovskii Krai, Komsomolsk-na-Amure District
MARCHENKO386 · Zootaxa 3626 (3) © 2013 Magnolia Press

(50°29' N, 137°04' E); 6 females, 4 males, Nikolaevsk-na-Amure District, estuary of Amur River, environs of Arkhangelskoe village (53°05' N, 140°59' E); 28 females, 6 males, Grand Khekhtsir Ridge; 2 females, Primorskii Krai, Vladivostok city (43°10' N, 131°55' E); 26 females, Sikhote-Alin Ridge (46°59' N, 136°53' E); 76 females, 15 males, Ussuri State Reserve (43°48' N, 131°56' E); on mainland Far East of Russia specimens were collected in litter and soil of mixed forests with prevalence of Querqus mongolica and other mixed broad-leaved forests; 71 females, 15 males, Sakhalin Island (46°59'–51°49' N, 142°51'–143°09' E); Kuril Islands: 6 females, Kunashir (43°50’ N, 145°30' E); 2 males, Shikotan (43°48' N, 146°51' E); specimens were collected in litter and soil of island type forests with Betula ermanii, Quercus mongolica and bamboo Sasa spp.
Etymology. The name angaridis refers to the name of the paleo-continent Angarida which existed in the Paleozoic time and became the Siberian Platform of Eurasia in modern time.
Remarks. Females of G. angaridis are similar to those of G. novipulchellus Ma & Yin, 1998. However, the latter has 17 pairs of setae in the podonotal region and 10 pairs in the opisthonotal region of dorsal shield; seta s6 is about 0.3 times as long as z6; the narrow dorsal sclerotised diagonal section latered of ventri-anal shield is not indicated; exopodal shields between I and II coxae are not distinguishable; separating line at ventri-anal shield scarcely reaching bases of JV4 setae; and the distance between bases of JV4 and JV3 setae is about 0.5 times as long as JV4 seta. Males of G. angaridis are very similar to those of G. novipulchellus, but the latter has whip-shaped spermatodactyl, about twice as long as the movable digit of the chelicerae; and tarsus II lacks spur-setae.
Females of G. angaridis are similar also to those of G. aduncus Ma, 2004. However the latter has 18 pairs of setae in the podonotal region and 10 pairs in the opisthonotal region of the dorsal shield; exopodal shields between coxae I and II are fused to the sternal shield; the separating line at ventri-anal shield scarcely reaches the bases of JV3 setae; seta ZV2 is about half as long as distance between its base and the base of ZV3; seta JV2 is about 0.3 times as long as distance between its base and base of JV3. Males of G. angaridis are very similar to those G.aduncus, but the latter has a whip-shaped spermatodactyl about twice as long as the movable digit of the chelicerae; the basal part of spermatodactyl is narrow; femur II has a hook-shaped spur-seta; and tarsus II lacks a spur-seta.
Key to Gamasiphis species of Eurasia (Females)
Eleven species of the genus Gamasiphis are known in Eurasia.
1. With one pair of pre-sternal shields. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Gamasiphis gandensius Van Daele, 1975; Belgium- With two pairs of of pre-sternal shields . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 22. Distance between transverse level of posteriormost pair of ventrianal setae (excluding para-anal setae) and anterior margin of
anal orifice corresponding to about the length of the anal orifice. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 3- Insertion of posteriormost pair of ventrianal setae (excluding para-anal setae) varying from slightly anterior to posterior to anal
orifice . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 53. Split band of dorsal shield adjacent to fused peritrematic-exopodal shield with a broad blunt posteriorly end. . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Gamasiphis productellus Berlese, 1923; China- Split band of dorsal shield adjacent to united peritrematic-exopodal shield with a sharply pointed posterior end . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Gamasiphis bengalensis Bhattacharyya, 1966; India5. At least Z5 and post-anal setae blunt or distally expanded . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .6- All idiosomal setae with sharp tips . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 76. With three pairs of J setae. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Gamasiphis pulchellus (Berlese 1887);
Europe: Italy, Belgium, Canary Islands, Bulgaria; Asia: Caucasus, China, Japan; Africa: Egypt, South Africa; North and South America; Bermudas; Java
- With four pairs of J setae. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .Gamasiphis lanceolatus Karg, 1987; Germany7. Seta j6 at least 1.2 times as long as distance between their bases . . . . . . . . . . . . . Gamasiphis sextus Vitzthum, 1921; Germany- Seta j6 at most 0.7 times as long as distance between their bases . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 88. Ventri-anal shield with seven pairs of setae in addition to para-anal setae (JV5 absent) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Gamasiphis turgicalcareus Ma, 2009; China- Ventri-anal shield with eight pairs of setae in addition to para-anal setae (JV5 present) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .99. Seta st5 at most 0.6 times as long as JV1; seta JV2 about as long as distance between its base and base of JV4 . . . . . . . . . . . . .
. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Gamasiphis indicus Bhattacharyya, 1978; India - Seta st5 about as long as JV1; seta JV2 at most 0.7 times as long as distance between its base and base of JV4 . . . . . . . . . . . 1010. Seta ZV2 about half as long as the distance between its base and the base of ZV3; seta JV2 about 0.3 times as long as the dis-
tance between its base and the base of JV3 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Gamasiphis aduncus Ma, 2004; China
Zootaxa 3626 (3) © 2013 Magnolia Press · 387GAMASIPHIS FROM NORTH ASIA

- Seta ZV2 about as long as the distance between its base and the base of ZV3; seta JV2 about half as long as the distance between its base and the base of JV3 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
11. Distance between the ends of separating line at ventri-anal shields as long as distance between bases of JV4 setae; distance between bases of JV4 and JV3 setae about 0.5 times as long as JV4 seta. . . . . . . . . . .G. novipulchellus Ma & Yin, 1998; China
- Distance between the ends of separating line at ventri-anal shields is shorter or equal to distance between bases of JV3 or con-nected into a complete fusion line in some specimens; distance between bases of JV4 and JV3 setae as long as JV4 seta. . . . . .. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . Gamasiphis angaridis; Russia: Altai Mountains, Middle Siberia, Far East
Discussion
Species of Gamasiphis had not been found higher than 50 degrees of latitude north or south before this study (Lee, 1970; Karg & Schorlemmer, 2009). The most northerly recorded Gamasiphis species are G. novipulchellus in Asia, China, Heilongjiang Province (47°10' N) and two undescribed species in Vancouver Island, Canada (48°46' N) (Emberson, 1968). The new record of the northernmost distribution of the genus Gamasiphis in the world is due to the current studies such huge and poorly studied region as North Asia.
Gamasiphis angaridis is the only species of Gamasiphis that is widely distributed in North Asia, from South Siberia, Altai Mountains (51°21'N) to Middle Siberia, Turukhansk (65°47'N) and to east from Altai Mountains (85°33'E) to South Kuril Islands (146°51'E). It is common in litter and soil at many different localities: birch-aspen, coniferous forests in Altai and Middle Siberia as well as broad-leaved and mixed forests in the Far East of Russia (Marchenko, 2010, 2012).
The distribution of G. angaridis is not limited by specific climatic conditions. It is common in both the temperate zone (Far East of Russia) or in the area with a severe continental climate of Middle Siberia and Altai Mountains, where the range of annual air temperature reaches 80° (between – 40° C in winter and + 40° C in summer). The distribution of soil gamasid mites in Siberia is not determinated only by recent climatic conditions (latitudinal aspect), but also historical (mainly longitudinal) ranges caused by paleoclimatic events that occurred in the Pleistocene. The Altai Mountains and Middle Siberia (old Siberian Platform) avoided marine transgressions in Pleistocene time (Arkhipov & Volkova, 1994). It is likely that soil as the most protected natural medium has enabled some representatives of thermophilous mite genera to survive the Pleistocene Ice Ages and protects them from harsh Siberian climatic conditions in a recent time.
Acknowledgements
I am grateful to my colleagues A.B. Babenko, O.G. Berezina, D.I. Berman, A.M. Dolgikh, V.V. Dubatolov, S.V. Kalabin, A.A. Lyashchev, I.I. Lyubechansky, N.A. Riabinin and E.V. Sleptsova for the collected material. Special thanks to Raphael Castilho, Bruce Halliday and Olga Makarova for access to information and their help in preparation of this paper.
References
Arkhipov, S.A. & Volkova, V.S. (1994) Geological history, landscapes and climate in Pleistocene time of Western Siberia. United Institute of Geology, Geophysics and Mineralogy, Novosibirsk, 106 pp.
Baker, E.W. & Wharton, G.W. (1952) An Introduction to Acarology. New York, Macmillan, 465 pp.Beaulieu, F. & Walter, D.E. (2007) Predation in suspended and forest floor soils: observation on Australian Mesostigmatic
mites. Acarologia, 57, 43–54.Berlese, A. (1887) Acari, Myriapoda et Scorpiones hucusque in Italia reperta. Portici et Padova, 39 (4), 4.Berlese, A. (1904) Acari nuovi. Manipulus II. Redia, 1, 258–280.Berlese, A. (1906) Monographia dei genere Gamasus Latr. Redia, 3, 65–304.Berlese, A. (1913) Acari nuovi. Manipulus VII–VIII. Redia, 9, 77–111.Berlese, A. (1914) Acari nuovi. Manipulus IX. Redia, 10, 113–150.Berlese, A. (1923) Centuria sesta di Acari nuovi. Redia, 15, 237–262.Bhattacharyya, S.K. (1966) Studies on Indian mites (Acarina: Mesostigmata), Part 3. Zoologischer Anzeiger, 177, 151–157.
http://dx.doi.org/10.1080/00305316.1971.10434033Bhattacharyya, S.K. (1968) Studies on Indian mites (Acarina: Mesostigmata). 6. Six records and descriptions of nine new
MARCHENKO388 · Zootaxa 3626 (3) © 2013 Magnolia Press

species. Acarologia, 10, 52–549.Bhattacharyya, S.K. (1978) Five new species and a new record of Mesostigmata (Acari) from West Bengal, India. Indian
Journal of Acarology, 2, 78–87.Bregetova, N.G. (1977) Family Ologamasidae Ryke. In: Ghilyarov, M.S. & Bregetova, N.G. (Eds). Key to the Soil-Inhabiting
Mites Mesostigmata. Nauka, Leningrad, pp. 483–554.Castagnoli, M. & Pegazzano, F. (1985) Catalogue of the Berlese Acaroteca. Istituto Sperimentale per la Zoologia Agraria,
Florence, Italy, 490 pp.Castilho, R.C., Moraes, G.J. de & Narita, J.P.Z. (2010a) A new species of Gamasiphis (Acari: Ologamasidae) from Brazil, with
a key to species from the Neotropical Region. Zootaxa, 2452, 31–43.Castilho, R.C., Iwanicki, N., Narita, J.P.Z. & Moraes G.J. de (2010b) Predation potential of Gamasiphis paulista Castilho,
Moraes & Narita (Acari: Ologamasidae). In: XIII International Congress of Acarology, August 23–27, 2010, Recife-PE,Brasil, 47.
Castilho, R.C., Narita, J.P.Z. & Moraes, G.J. de (2012) Three new species of Gamasiphis (Acari: Mesostigmata: Ologamasidae) from Brazil, with complementary information about Gamasiphis plenosetosus Karg and a key to the world species of the genus. Journal of Natural History, 46, 1969–1998.
Deltshev, C., Andreev, S., Blagoev, G., Golemansky, V., Milojkova, G., Peneva, V., Dobrev D., Todorov, M. & Hubenov, Z. (1998) Invertebrates (Non-Insecta) in Bulgaria. In: Bulgaria’s Biological Diversity. Available from: http://www.worldwildlife.org/bsp/publications/europe/bulgaria/bulgaria5.html (date of access 25 October 2010).
El-Banhawy, E.M., Nasr, A.K. & Afia, S.I. (2006) Survey of predaceous soil mites (Acari: Mesostigmata) in citrus orchards of the Nile Delta and Middle Egypt with notes on the abundance of the citrus parasitic nematode Tylenchulus semipenetrans(Tylenchida: Tylenchulidae). International Journal of Tropical Insect Science, 26, 64–69. http://dx.doi.org/10.1079/IJT200692
Emberson, R.M. (1968) The Mesostigmata of certain coniferous forest soils in Western Quebec, with a preliminary account of the North American Rhodacaridae (Acarina). Ph.D. dissertation, Department of Entomology, McGill University, Montreal, Canada, pp. 349.
Ewing, H.E. (1920) New predaceous and parasitic mites of the superfamily Gamasoidea (Acar.). Entomological news, 31, 286–293.
Halliday, R.B. (2005) Predatory mites from crops and pastures in South Africa: potential natural enemies of redlegged earth mite Halotydeus destructor (Acari: Pentaleidae). Zootaxa, 1079, 11–64.
Hennessey, M.K. & Farrier, M.H. (1988) Systematic revision of thirty species of free-living, soil-inhabiting Gamasine mites (Acari: Mesostigmata) of North America. Technical Bulletin, North Carolina Agricultural Research Service, 285, 1–121.
Johnston, D.E. & Moraza M.L. (1991) The idiosomal adenotaxy and poroidotaxy of Zerconidae (Mesostigmata: Zerconina). InF. Dusbábek & V. Bukva (eds), Modern Acarology, SPB Academic Publishing, The Hague, Netherlands, Volume 2, 349–356.
Karg, W. (1987) Zur Kenntnis der Raubmilbenarten der Gattung Gamasiphis Berlese, 1904 (Acarina, Parasitiformes). Mitteilungen aus dem Zoologischen Museum in Berlin, 63, 301–308. http://dx.doi.org/10.1002/mmnz.19870630205
Karg, W. (1990) Neue Raubmilbenarten der Gattung Gamasiphis Berlese, 1904 (Acarina, Parasitiformes). Acarologia, 31, 321–355. http://dx.doi.org/10.1002/mmnz.19870630205
Karg, W. (1993) Zur Kenntnis der Raubmilbengattung Gamasiphis Berlese, 1904 (Acarina, Parasitiformes). Zoologische Jahrbücher, Abteilung für Systematic, Oecologie und Geographie der Tiere, 120, 169–188. http://dx.doi.org/10.1002/mmnz.19870630205
Karg, W. & Schorlemmer, A. (2009) New insights into predatory mites (Acarina, Gamasina) from tropical rain forests with special reference to distribution and taxonomy. Zoosystematics and Evolution, 85, 57–91. http://dx.doi.org/10.1002/zoos.200800016
Lee, D.C. (1970) Rhodacaridae (Acari: Mesostigmata): classification, external morphology and distribution of genera. Records of the South Australian Museum, 16 (3), 1–219.
Lindquist, E.E. & Evans, G.O. (1965) Taxonomic concepts in the Ascidae, with a modified setal nomenclature for the idiosoma of the Gamasina (Acarina: Mesostigmata). Memoirs of the Entomological Society of Canada. 47, 1–64. http://dx.doi.org/10.4039/entm9747fv
Lindquist, E.E., Krantz, G.W. & Walter, D.E. (2009) Order Mesostigmata. In: Krantz, G.W. & Walter, D.E. (eds). A Manual of Acarology, Third Edition, Texas Tech University Press, Lubbock, Texas, pp. 124–232. http://dx.doi.org/10.1653/024.092.0323
Ma, L.-M. (2004) Description of Gamasiphis aduncus sp. nov. and supplemental characters of Gamasiphis novipulchellus(Acari: Gamasina: Ologamasidae). Acta Arachnologica Sinica, 13 (1), 23–27.
Ma, L.-M. (2009) Description of Gamasiphis turgicalcareus sp. nov. (Acari: Mesostigmata: Ologamasidae). Acta Arachnologica Sinica, 18 (2), 78–79.
Ma, L.-M. & Ma, D.-M. (2000) Collections of gamasid mites in Dandong City (II) with descriptions of male and deutonymph of Parasitus mammilatus (Acari). Entomological Journal of East China, 912, 117–119.
Ma, L.-M. & Yin, X.-Q. (1998) A new species of the genus Gamasiphis (Acari: Ologamasidae). Acta Entomologica Sinica, 41,
Zootaxa 3626 (3) © 2013 Magnolia Press · 389GAMASIPHIS FROM NORTH ASIA

319–322.Marchenko, I.I. (2010) Soil-inhabiting gamasid mires (Acari, Mesostigmata) on North-East Altai: transformation of taxonomic,
geographical and population structure of communities along an altitudinal gradient. Euroasian Entomological Journal, 9, 741–756.
Marchenko, I.I. (2012) Soil gamasid mites (Acari, Mesostigmata) of North Siberia. Euroasian Entomological Journal, 6, 517–528.
Minor, M.A. & Norton, R.A. (2004) Effects of soil amendments on assemblages of soil mites (Acari: Oribatida, Mesostigmata) in short-rotation willow plantings in Central New York. Canadian Journal of Forest Research, 34, 1417–1425. http://dx.doi.org/10.1139/x04-035
Moraza, M.L. & Peña, M.A. (2005) Ácaros Mesostigmata (Acari, Mesostigmata) de hàbitats seleccionados de la Gomera (Islas Canarias, España). Graellsia, 61 (1), 1–6. http://dx.doi.org/10.3989/graellsia.2005.v61.i1.9
Ryke, P.A.J. (1962) The subfamily Rhodacaridae with notes on a new subfamily Ologamasinae (Acarina: Rhodacaridae). Entomologische Berichten, 22 (1), 155–162.
Silva, E.S., Moraes, G.J. de, Krantz, G.W. (2007) A new species of Ologamasus (Acari: Ologamasidae) from Brazil. Zootaxa, 1462, 61–68.
Trägårdh, I. (1952) Acarina, collected by the Mangarevan expedition to South Polynesia in 1934 by the Bernice P. Bishop Museum, Honolulu, Hawaii, Mesostigmata. Arkiv for Zoologi, 4, 45–90.
Van Daele, E. (1975) A new Gamasiphis species from azalea culture. Biologisch Jaarboek Dodonaea, 43, 266–270.Vitzthum, H.G. (1921) Acarologische Beobachtungen. 5 Reihe. Archiv für Naturgeschichte, 87 (4), 1–77.Zaher, M.A. (1986) Predaceous and nonphytophagous mites in Egypt (Nile Valley and Delta). Cairo, Cairo University, 567 pp.
MARCHENKO390 · Zootaxa 3626 (3) © 2013 Magnolia Press





























