Embed Size (px)
Citation preview

SUPLEMENTASI ASAM AMINO PADA PAKAN AYAM
BROILER TINGGI LEMAK UNTUK MENGURANGI
DEPOSIT LEMAK ABDOMEN
MOHAMMAD MAGHFURI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2014


PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Suplementasi Asam
Amino Pada Pakan Ayam Broiler Tinggi Lemak untuk Mengurangi Deposit Lemak Abdomen adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Maret 2014
Mohammad Maghfuri
NIM D152110031


RINGKASAN
MOHAMMAD MAGHFURI. Suplementasi Asam Amino pada Pakan
Ayam Broiler Tinggi Lemak untuk Mengurangi Deposit Lemak Abdomen. Dibimbing oleh ASEP SUDARMAN dan RITA MUTIA.
Lemak atau minyak pada nutrisi unggas berfungsi sebagai sumber energi,
sumber asam lemak esensial, dan sebagai pelarut vitamin larut lemak. Minyak yang paling banyak digunakan pada industri pakan Indonesia adalah minyak sawit yang memiliki energi metabolis 8000 kkal kg-1. Kandungan asam lemak yang utama pada minyak sawit adalah asam palmitat 39.23% dan oleat 38.21%. Penggunaan minyak sawit sampai 6% tidak berpengaruh terhadap deposit lemak abdomen. Lemak abdomen menjadi masalah pada industri ayam broiler modern. Lemak abdomen tidak hanya menurunkan persentase hasil karkas dan efisiensi pakan. Lemak abdomen dapat diturunkan dengan meningkatkan kecernaan dan metabolisme lemak pakan melalui suplementasi asam amino. Glisina dan taurina merupakan asam amino yang berperan dalam biosintesis garam empedu yang mengemulsi lemak. Lisina dan metionina merupakan asam amino prekursor biosintesis L-karnitin yang digunakan sebagai transportasi asam lemak dalam proses �-oksidasi di mitokondria. Penelitian ini bertujuan untuk mengkaji jenis asam amino yang tepat disuplementasi pada pakan ayam broiler tinggi lemak untuk meningkatkan penyerapan dan metabolisme lemak sehingga mengurangi deposit lemak abdomen.
Penelitian ini menggunakan 240 ekor anak ayam broiler (DOC) strain Cobb. Ayam broiler ditempatkan pada 24 unit kandang secara acak dengan densitas 10 ekor per m2. Ayam broiler diberi pakan dan minum ad libitum sampai umur 42 hari. Pada umur 21 dan 42 hari, setiap perlakuan diambil empat ekor ayam broiler dari masing-masing ulangan untuk diambil serum darahnya dan disembelih. Perlakuan pada penelitian ini terdiri dari R0 (ransum basal mengandung 5% minyak sawit), R1 (R0 + 0.25% glisina), R2 (R0 + 0.25% taurina), R3 (R0 + 15% total lisina dan metionina ransum basal), R4 (R0 + 0.25% glisina + 0.25% taurina), R5 (R0 + 0.25% glisina + 0.25% taurina + 15% total lisina dan metionina ransum basal. Penelitian ini menggunakan rancangan acak lengkap (RAL) dengan 6 perlakuan dan 4 ulangan. Data yang diperoleh dianalisis sidik ragam kecuali mortalitas, kemudian bila rataan berbeda nyata dilanjutkan dengan Uji Jarak Duncan. Variabel yang diukur meliputi performa, profil metabolit serum, karakteristik empedu, koefisien kecernaan semu nutrien, kandungan nutrien daging, dan berat relatif karkas dan organ ayam broiler.
Hasil penelitian menunjukan bahwa semua suplementasi asam amino nyata menurunkan konsumsi pakan, bobot badan dan pertambahan bobot badan ayam broiler umur 1-42 hari. Konversi pakan ayam broiler umur 1-42 hari yang diberi pakan perlakuan R2 dan R3 nyata meningkat, R1 dan R4 tidak berbeda nyata, dan R5 nyata menurun dibandingkan kontrol. Mortalitas ayam broiler yang diberi pakan R3 paling tinggi dibandingkan perlakuan lainnya. Semua suplementasi asam amino nyata meningkatkan asam urat dan trigliserida serum, tetapi nyata menurunkan kolesterol serum ayam broiler umur 21 dan 42 hari. Berat relatif kantung empedu ayam broiler umur 21 hari perlakuan R1 dan R4 nyata meningkat, tetapi perlakuan R2 yang nyata meningkat pada umur 42 hari. Semua

suplementasi asam amino nyata meningkatkan berat kantung empedu ayam broiler umur 21 hari, kecuali R3. Selanjutnya, perlakuan R3 nyata menurunkan berat kantung empedu ayam broiler umur 42 hari. Berat dan volume cairan empedu ayam broiler umur 21 hari nyata meningkat dengan perlakuan R1, R2 dan R4, tetapi tidak berbeda nyata pada umur 42 hari. Semua suplementasi asam amino nyata menurunkan berat jenis cairan empedu ayam broiler umur 21 hari kecuali R1, tetapi tidak berbeda nyata pada umur 42 hari.
Koefisien kecernaan semu bahan kering ayam broiler umur 21 hari perlakuan R1 nyata meningkat, sedangkan perlakuan R1, R2 dan R4 yang nyata meningkat pada umur 42 hari. Selanjutya, koefisien kecernaan semu lemak kasar ayam broiler umur 21 hari perlakuan R1 dan R4 nyata meningkat, tetapi perlakuan R3 nyata menurun pada umur 42 hari. Koefisien kecernaan semu protein kasar ayam broiler umur 21 hari nyata menurun dengan perlakuan R3 dan R4, tetapi nyata meningkat dengan perlakuan R1 pada umur 42 hari. Koefisien kecernaan semu energi ayam broiler umur 21 hari perlakuan R3 nyata menurun. Selanjutnya, koefisien kecernaan semu energi ayam broiler umur 42 hari perlakuan R3 nyata menurun, R2 dan R5 tidak berbeda nyata, serta perlakuan R1 dan R4 nyata meningkat.
Bahan kering daging ayam broiler umur 21 hari perlakuan R4 dan R5 nyata meningkat, tetapi perlakuan R2 dan R4 yang meningkat pada umur 42 hari. Protein kasar daging ayam broiler umur 21 hari perlakuan R2 nyata meningkat, tetapi suplementasi asam amino lainnya tidak berpengaruh nyata terhadap protein kasar daging ayam broiler umur 21 dan 42 hari. Lemak kasar daging ayam broiler umur 21 hari meningkat dengan perlakuan R1, R3, dan R5. Selanjutnya, semua suplementasi asam amino nyata meningkatkan lemak kasar daging ayam broiler umur 42 hari. Ayam broiler umur 21 dan 42 hari yang diberi pakan R3 cenderung menurunkan L-karnitin daging, tetapi cenderung meningkat dengan perlakuan R5. Semua suplementasi asam amino tidak berpengaruh nyata terhadap berat relatif karkas, hati dan pankreas ayam broiler umur 21 dan 42 hari. Berat relatif lemak abdomen ayam broiler umur 21 hari perlakuan R2 nyata meningkat, R1, R4 dan R5 tidak berbeda nyata, serta R3 nyata menurun. Akan tetapi, semua suplementasi asam amino tidak berpengaruh nyata terhadap berat relatif lemak abdomen ayam broiler umur 42 hari.
Kesimpulannya adalah kombinasi suplementasi lisina dan metionina (R3) dapat menurunkan lemak abdomen ayam broiler umur 21 hari dari 1.30 menjadi 0.76%. Kombinasi suplementasi glisina, taurina, lisina, dan metionina (R5) cenderung meningkatkan L-karnitin daging ayam broiler umur 21 dan 42 hari dengan konsentrasi masing-masing 15.28 dan 12.70 mg 100 g-1. Suplementasi glisina, taurina, maupun kombinasin keduanya dapat meningkatkan volume cairan empedu dan kecernaan lemak, khususnya pada umur 21 hari. Peningkatan volume cairan empedu mencapai 97.10%, sedangkan koefisien kecernaan semu lemak kasar mencapai 2.24%.
Kata kunci: ayam broiler, garam empedu, lemak abdomen, L-karnitin,
suplementasi asam amino

SUMMARY
MOHAMMAD MAGHFURI. Amino Acid Supplementation into High Fat Broiler Diet for Reducing Abdominal Fat Deposition. Supervised by ASEP SUDARMAN and RITA MUTIA.
Fats or oils are used in poultry nutrition as source of energy, essential fatty
acids and carrier of fat soluble vitamins. Most oil that used in Indonesia feed industry is palm oil that has metabolizable energy 8000 kcal kg-1. Fatty acids contained in palm oil are greatly palmitic acid of 39.23% and oleic acid of 38.21%. Palm oil utilization up to 6% do not affect abdominal fat deposition. Abdominal fat become a problem in modern broiler chicken industry. Abdominal fat do not only reduce carcass yield percentage and feed efficiency, but is also rejected by costumer. Abdominal fat might be reduced with the increasing digestibility and metabolisibility fat in the diet by amino acids supplementation. Glycine and taurine were amino acids that have role to synthesize bile salt for fat emulsification. Lysine and methionine were precursor to synthesize L-carnitine that has function to transport fatty acid for �-oxidation process in mitochondria. The aim of this study was to elaborate appropriate amino acids supplementation into high fat broiler diet to improve fat absorption and metabolism in order to reduce abdominal fat deposition.
This study used 240 day old broilers Cobb strain. They were placed randomly into 24 pens with density of 10 birds/m2. Broiler chickens were given feed and water ad libitum for 42 days. At 21st and 42nd day, each treatments were taken four broiler chicken come from each replication. They were slaughtered and their blood serum were taken. The treatments diet were R0 (basal diet contained 5% palm oil), R1 (R0 + 0,25% glycine), R2 (R0 + 0,25% taurine), R3 (R0 + 15% total lysine and methionine of basal diet), R4 (R0 + 0,25% glycine + 0,25% taurine), and R5 (R0 + 0,25% glycine + 0,25% taurine + 15% total lysine and methionine of basal diet). Completely randomized design (CRD) with 6 treatments and 4 replicates was used in this study. The variables measured were performances, serum metabolites, gall bladder characteristics, apparent digestion coefficient of nutrients, nutrients content of meat, and relative weight of carcass and organs broiler chicken. Data were subjected to analysis of variance and if any means significant differences were continued by Duncan multiple range test, except mortality.
The results showed that all amino acids supplementation significantly reduced feed consumption, body weight and weight gain at 1-42 day old broiler chickens. Feed conversion broiler chicken for 1-42 day old fed R2 and R3 significantly increased, R1 and R4 were not significant, and R5 significantly reduced compared control. Chicken fed R3 had highest mortality than those of other groups. All supplementation of amino acids significantly increased serum uric acid and triglyceride, but significantly reduced serum cholesterol broiler chicken at 21 and 42 days old. Gall blader relative weight for 21 days fed R1 and R4 significantly increased, but fed R2 significantly increased at 42 days. All amino acid supplementations increased gall blader weight broiler chicken at 21 days, except fed R3. Whereas, broiler chicken fed R3 significantly reduced gall blader weight at 42 days. Weight and volume of bile liquid broiler chicken fed R1,

R2 and R4 at significantly increased 21 days, but there were no differences at 42 days. All amino acid supplementations significantly reduced bile liquid density broiler chicken at 21 except R1, but there were no differences at 42 days.
Apparent digestibility coefficient of dry matter broiler chicken for 21 days fed R1 significantly increased, nevertheless fed R1, R2 and R4 were significantly increased at 42 days. Furthermore, apparent digestibility coefficient of crude fat broiler chicken for 21 days fed R1 and R4 significantly increased, but fed R3 sigificantly reduced at 42 days. Apparent digestibility coefficient of crude protein broiler chicken for 21 days significantly increased with fed R3 and R4, but increased with fed R1 at 42 days. Apparent digestibility coefficient of energy broiler chicken for 21 days fed R3 significantly reduced. Moreover, apparent digestibility coefficient of energy fed R3 significantly reduced, R2 and R5 were not significantly different, then R1 and R4 significantly increased at 42 days.
Meat dry matter broiler chicken at 21 days fed R4 and R5 significantly increased, but fed R2 and R4 significantly increased at 42 days. Meat crude protein broiler chicken at 21 days fed R2 significantly increased, but other amino acid supplementations did not effected meat crude protein broiler chicken at 21 and 42 days. Meat crude fat broiler chicken at 21 days fed R1, R3 and R5 significantly increased. Furthermore, all amino acid supplementations significantly increased meat crude fat broiler chicken at 42 days. All amino acids supplementation did not affect to relative weight of carcass, liver, and pancreas broiler chicken at 21 and 42 days. Broiler chicken given feed R3 had tendency reduced meat L-carnitine, but they fed R5 increased at 21 and 42 days old. Abdominal fat relative weight broiler chicken at 21 days fed R2 significantly increased, R1, R4 and R5 did not significant, and R3 significantly reduced. Nevertheless, all amino acid supplementations did not effect to abdominal fat relative weight at 42 days.
It is concluded that mixed of supplementations of lysine and methionine (R3) could reduce abdominal fat at 21 days old from 1.30 be 0.76%. Mixed supplementation of glycine, taurine, lysine and methionine (R5) tendency improve L-carnitine meat content of broiler chicken at 21 and 42 days with each concentration 15.28 and 12.70 mg 100 g-1. Supplementations of glycine, taurine, or both mixed could improve bile liquid volume and fat apparent digestibly coefficient, especially at 21 days old. Bile liquid volume increased up to 97.10%, then fat apparent digestibly coefficient increased 2.24%. Key words: Abdominal fat, amino acid supplementation, broiler chicken, bile salt,
L-carnitine

© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan
IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis ini
dalam bentuk apa pun tanpa izin IPB


Tesis sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Studi Ilmu Nutrisi dan Pakan
SUPLEMENTASI ASAM AMINO PADA PAKAN AYAM
BROILER TINGGI LEMAK UNTUK MENGURANGI
DEPOSIT LEMAK ABDOMEN
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
2014
MOHAMMAD MAGHFURI

Penguji Luar Komisi pada Ujian Tesis : Dr Ir Sumiati, MSc

Judul Tesis : Suplementasi Asam Amino pada Pakan Ayam Broiler Tinggi Lemak untuk Mengurangi Deposit Lemak Abdomen.
Nama : Mohammad Maghfuri NIM : D152110031
Disetujui oleh
Komisi Pembimbing
Dr Ir Asep Sudarman, MRurSc Ketua
Dr Ir Rita Mutia, MAgrSc Anggota
Diketahui oleh Ketua Program Studi Ilmu Nutrisi dan Pakan
Dr Ir Dwierra Evvyernie A, MS MSc
Dekan Sekolah Pascasarjana Dr Ir Dahrul Syah, MScAgr
Tanggal Ujian: 5 Maret 2014
Tanggal Lulus:

T esis : Suplementasi Asam Amino pada Pakan Ayam Broiler Tinggi Lemak untuk Mengurangi Deposit Lemak Abdomen.
: Mohammad Maghfuri : D152110031
Disetujui oleh
Komisi Pembimbing
Dr Ir Asep Sudarman, MRurSc Dr Ir Rita Mutia, MAgrSc Ketua Anggota
Diketahui oleh
Ketua Program Studi Ilmu Nutrisi dan Pakan
.;,--, . ,
Dr IT Dwierra Evvyernie A, MS MSc
Tanggal Ujian: 5 Maret 2014 Tanggal Lulus: 08 APR 20 14
Sekolah Pascasatjana


PRAKATA
Innalhamdalillah, puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala rahmat dan hidayah-Nya sehingga karya ilmiah ini berhasil diselesaikan. Judul penelitian dalam karya ilmiah ini adalah Suplementasi Asam Amino pada Pakan Ayam Broiler Tinggi Lemak untuk Mengurangi Deposit Lemak Abdomen yang telah dilaksanakan dengan baik pada bulan Oktober 2012 sampai Pebruari 2013.
Penulis menyampaikan terima kasih kepada Bapak Dr Ir Asep Sudarman MRurSc dan Ibu Dr Ir Rita Mutia MAgrSc, selaku komisi pembimbing yang telah berkenan meluangkan waktu, memberikan bimbingan dan arahan pada penulis dalam menyelesaikan karya ilmiah ini. Ungkapan terima kasih juga penulis kepada Dr Ir Sumiati MSc atas saran dan masukan selama proses penelitian dan penyempurnaan karya ilmiah ini. Penulis juga menyampaikan terima kasih kepada Bapak Ir Nugroho Adi H, MSi dan PT. Cheil Jedang Feed Indonesia yang telah memberikan bantuan materi selama proses penelitian. Terima kasih juga penulis ucapkan kepada Kementerian Pendidikan dan Kebudayaan atas Beasiswa Kementerian Pendidikan Nasional yang penulis terima mulai tahun 2011.
Terima kasih penulis ucapkan kepada Ayahanda Kasmuri Santoso dan Ibunda Nurjanah, saudara-saudaraku tercinta (Muflikhatun, Aji Wijaya dan Zami), serta istri (Herwanti) dan anakku (Zafira) tercinta atas doa, kasih sayang dan dorongan baik moral, materi maupun spiritual. Kepada Ibu Dr Ir Ning Iriyanti MP dan Prof Agus Iriyanto PhD, penulis ucapkan terima kasih telah menjadi pembimbing, motivator dan orang tua kedua bagi penulis. Penulis mengucapkan terima kasih kepada teman-teman UNSOED di Wisma Soedirman, teman-teman INP 2011, khususnya Mas Arif, Mas Sichi, dan Mas Alam yang telah memberikan bantuan dan motivasi selama penelitian. Penulis juga menyampaikan terimakasih kepada Ibu Lanjarsih, Bapak Supri, Ibu Ade, Sofian, serta semua pihak yang tidak bisa ditulis satu persatu yang telah membantu jalannya penelitian.
Semoga karya ilmiah ini bermanfaat bagi semua pembaca sekalian.
Bogor, Maret 2014
Mohammad Maghfuri


DAFTAR ISI
DAFTAR TABEL ix�
DAFTAR GAMBAR ix�
DAFTAR LAMPIRAN x�
PENDAHULUAN 1�Latar Belakang 1�Tujuan Penelitian 2�Hipotesis Penelitian 2�Kerangka Pemikiran 3�
TINJAUAN PUSTAKA 4�Lemak pada Nutrisi Unggas 4�Peran Glisina dan Taurina dalam Biosintesis Garam Empedu 4�Peran Garam Empedu pada Proses Pencernaan dan Penyerapan Lemak 6�Peran Lisina dan Metionina dalam Biosintesis L-karnitin 6�Peran L-Karnitin dalam Metabolisme Lemak 8�Lemak Abdomen pada Ayam Broiler 9�Biosintesis Lemak Abdomen 9�Metode Menurunkan Deposit Lemak Abdomen 11�
METODE 13�Bahan 13�Alat 13�Waktu dan Lokasi Percobaan 13�Rancangan Percobaan 13�Metode Percobaan 15�Prosedur Pengukuran Parameter 16�Analisis Data 18�
HASIL DAN PEMBAHASAN 19�Pengaruh Suplementasi Asam Amino terhadap Performa 19�Pengaruh Suplementasi Asam Amino terhadap Metabolit Serum 29�Pengaruh Suplementasi Asam Amino terhadap Karakteristik Empedu 33�Pengaruh Suplementasi Asam Amino terhadap Koefisien Kecernaan Semu Nutrien 38�Pengaruh Suplementasi Asam Amino terhadap Kandungan Nutrien Daging Dada 43�Pengaruh Suplementasi Asam Amino terhadap Berat Relatif Karkas dan Organ 49�
SIMPULAN DAN SARAN 53�Simpulan 53�Saran 53�

DAFTAR PUSTAKA 54�
LAMPIRAN 63�
RIWAYAT HIDUP 82�

DAFTAR TABEL
1 Susunan ransum basal untuk penelitian 14�2 Pengaruh suplementasi asam amino terhadap performa ayam broiler 19�3 Pengaruh suplementasi asam amino terhadap metabolit serum ayam
broiler (mg dl-1) 29�4 Pengaruh suplementasi asam amino terhadap karakteristik empedu
ayam broiler 34�5 Korelasi volume cairan empedu dengan konsumsi glisina, taurina
dan glisina+taurina 37�6 Pengaruh suplementasi asam amino terhadap koefisen kecernaan
semu nutrien ayam broiler (%) 39�7 Pengaruh suplementasi asam amino terhadap nutrien daging paha
ayam broiler (%) 43�8 Pengaruh suplementasi asam amino terhadap berat relatif karkas dan
organ ayam broiler (% BB) 49�
DAFTAR GAMBAR
1 Kerangka Pemikiran Penelitian 3�2 Proses konjugasi asam empedu dengan glisina dan taurina (Sumber :
Märt 2013) 5�3 Peran garam empedu dalam proses pencernaan dan penyerapan lemak
(Sumber : Liz 2013) 6�4 Peran lisina dan metionina dalam proses biosintesis L-karnitin
(Sumber : Scholte 2003) 7�5 Peran L-karnitin pada proses metabolisme asam lemak (Sumber :
Mehta 2013) 8�6 Proses biosintesis lemak abdomen 10�7 Mekanisme suplementasi asam amino menurunkan performa ayam
broiler 20�8 Proses metabolisme kelebihan asam amino pada serum 30�9 Ringkasan pengaruh suplementasi glisina dan taurina terhadap
karakteristik empedu 38�10 Hubungan antara volume cairan empedu dan koefisien kecernaan
semu nutrien 41�11 Kandungan L-karnitin daging ayam broiler umur 21 dan 42 hari 46�12 Korelasi konsumsi lisina dan metionina dengan kandungan L-karnitin
daging ayam broiler umur 21 (kiri) dan 42 (kanan) hari 48�13 Mekanisme rendahnya korelasi L-karnitin daging dengan konsumsi
lisina dan metionina 48�

DAFTAR LAMPIRAN
1 Prosedur analisis L-karnitin dengan Metode HPLC 63�2 Analisis sidik ragam dan uji lanjut Duncan performa 64�3 Analisis sidik ragam dan uji lanjut Duncan profil metabolit serum 69�4 Analisis sidik ragam dan uji lanjut Duncan karakteristik empedu 72�5 Analisis sidik ragam dan uji lanjut Duncan koefisien kecernaan semu
nutrien 75�6 Analisis sidik ragam dan uji lanjut Duncan nutrien daging paha 78�7 Analisis sidik ragam dan uji lanjut Duncan berat relatif karkas dan
organ 81�

PENDAHULUAN
Latar Belakang
Lemak atau minyak pada nutrisi unggas berfungsi sebagai sumber energi, sumber asam lemak esensial dan sebagai pelarut vitamin larut lemak. Kandungan energi lemak sebesar 9.4 kkal g-1 atau 2.25 kali energi karbohidrat. Proses pencernaan dan metabolisme lemak membutuhkan banyak faktor fisiologis seperti asam empedu, lipase pankreas, protein pengikat asam lemak dan L-karnitin untuk proses � oksidasi asam lemak (Larbier dan Leclerq 1994). Efisiensi penggunaan lemak dalam pakan sangat tergantung pada komposisi asam lemaknya. Lemak yang kaya akan asam lemak jenuh lebih sulit dicerna dibandingkan dengan asam lemak tidak jenuh (Crespo dan Esteve-Garcia 2001; Baião dan Lara 2005; Shahriar et al. 2007). Penambahan lemak pada pakan dapat meningkatkan pertumbuhan dan efisiensi pakan ayam broiler (Leeson dan Summer 2001; Firman et al. 2008). Akan tetapi, penggunaan lemak pada ayam muda kurang efektif, khususnya ketika diberikan sumber asam lemak jenuh. Hal ini disebabkan karena rendahnya sintesis garam empedu yang berfungsi sebagai pengemulsi pada proses pencernaan lemak (Green dan Kellogg 1987; Al-Marzooqi 1998; Baião dan Lara 2005). Asam lemak jenuh pada pakan juga meningkatkan deposit lemak abdomen, karena lemak lebih banyak dideposit dari pada dikonversi menjadi energi (Firman et al. 2008). Hal ini dapat disebabkan karena kurangnya biosintesis L-karnitin yang berfungsi membantu oksidasi asam lemak menjadi energi.
Deposit lemak abdomen menjadi masalah pada industri ayam broiler modern karena menyebabkan penurunan hasil karkas dan efisiensi pakan (Kessler et al. 2000). Secara ekonomi, jika rata-rata lemak abdomen 2% dari bobot ayam broiler dan pada tahun 2012 Indonesia memproduksi 2.4 miliar kg ayam broiler hidup (Utoyo 2012), maka sekitar 480 juta kg lemak abdomen dihasilkan pada tahun 2012. Peningkatan deposit lemak abdomen sangat dipengaruhi oleh genetik dan pakan (T�mová dan Teimouri 2010). Seleksi genetik tidak hanya berkontribusi meningkatkan laju pertumbuhan, tetapi juga meningkatkan lemak tubuh, khususnya lemak abdomen (Tavernari et al. 2009). Gaya et al. (2005) mengestimasikan bahwa terjadi peningkatan berat lemak abdomen ayam broiler sekitar 0.39 g tahun-1. Selanjutnya, kandungan nutrien pakan seperti lemak kasar dan asam lemak pakan juga berpengaruh terhadap deposit lemak abdomen. Wongsuthavas (2007) melaporkan bahwa pakan yang mengandung lemak kasar 9% meningkatkan lemak abdomen. Akan tetapi, penggunaan minyak biji bunga matahari yang kaya asam lemak linoleat dapat menurunkan lemak abdomen (Crespo dan Esteve-Garcia 2001; Poorghasemi et al. (2013).
Penelitian ini menggunakan 5% minyak sawit pada campuran pakan untuk meningkatkan kandungan lemak kasar, sehingga disebut “pakan tinggi lemak”. Minyak sawit dipilih karena merupakan jenis lemak yang paling banyak digunakan pada industri pakan Indonesia. Kandungan asam lemak yang utama pada minyak sawit adalah asam palmitat 39.23% dan oleat 38.21% (Pesti et al. 2002). Penelitian sebelumnya menunjukan bahwa penggunaan minyak sawit sampai 6% tidak berpengaruh terhadap berat lemak abdomen dibandingkan yang menggunakan minyak kedelai (Pesti et al. 2002; Baião dan Lara 2005; Firman et

2
al. 2008). Industri pakan biasanya menggunakan minyak sawit dalam formula pakan ayam broiler sekitar 3%, sedangkan penelitian ini menggunakannya sampai 5%. Oleh karena itu, salah satu manfaat yang diperoleh dari penelitian ini adalah dapat meningkatkan penggunaan minyak sawit pada formula pakan ayam broiler.
Lemak abdomen dapat diturunkan dengan meningkatkan kecernaan dan metabolisme lemak pakan melalui suplementasi asam amino. Industri pakan unggas biasa menggunakan asam amino sebagai feed supplement untuk mengoptimalkan performa ayam broiler. Asam amino yang disuplementasi pada penelitian ini meliputi glisina dan taurina yang berperan dalam biosintesis garam empedu (Gropper et al. 2009; Cheeke dan Dierenfeld 2010), serta lisina dan metionina yang berperan dalam biosintesis L-karnitin (Arslan 2006). Suplementasi asam amino dilaporkan dapat meningkatkan proses pencernaan dan metabolisme karena dapat meningkatkan sintesis garam empedu dan L-karnitin (Lee et al. 2004; Alzawqari et al. 2010; dan Bouyeh dan Gevorgyan 2011). Garam empedu memiliki peran penting mengemulsikan lemak dengan memecah butiran lemak menjadi lebih kecil sehingga dapat diserap oleh usus (Gropper et al. 2009; Cheeke dan Dierenfeld 2010). Selanjutnya, L karnitin digunakan sebagai sarana transportasi asam lemak rantai panjang untuk masuk ke membran dalam mitokondria pada proses �-oksidasi asam lemak menjadi ATP (Adenosina Trifosfat) (Harmeyer 2002; Arslan 2006).
Suplementasi asam amino glisina dan taurina diharapkan dapat meningkatkan produksi garam empedu untuk meningkatkan kecernaan lemak. Suplementasi asam amino lisina dan metionina juga diharapkan dapat meningkatkan biosintesis L-karnitin yang berperan dalam �-oksidasi asam lemak menjadi energi. Oleh karena itu, kombinasi suplementasi asam amino tersebut diharapkan mampu memaksimalkan proses pencernaan dan metabolisme lemak ayam broiler, sehingga dapat menurunkan deposit lemak abdomen.
Tujuan Penelitian
Penelitian ini bertujuan menentukan jenis asam amino yang tepat disuplementasikan pada pakan ayam broiler tinggi lemak untuk menurunkan deposit lemak abdomen. Penelitian ini juga bertujuan untuk mengkaji proses biosintesis garam empedu dan L-karnitin ayam broiler dengan suplementasi asam amino pada pakan ayam broiler tinggi lemak.
Hipotesis Penelitian
Hipotesis dari penelitian ini adalah suplementasi asam amino dengan kombinasi yang tepat dapat menurunkan deposit lemak abdomen ayam broiler serta meningkatkan biosintesis garam empedu dan L-karnitin ayam broiler.

3
Kerangka Pemikiran
Kerangka pemikiran penelitian ini berdasarkan permasalahan pada latar belakang diilustrasikan pada Gambar 1.
Gambar 1 Kerangka Pemikiran Penelitian
Asam Amino Lisina dan Metionina :
Meningkatkan Biosintesis L-karnitin�
Meningkatkan Metabolisme Lemak
Pemanfaatan Lemak pada Pakan Ayam Broiler
Asam Amino Glisina dan Taurina :
Meningkatkan Biosintesis Garam Empedu
Sumber Energi :
9.4 kkal g-1 atau 2.25 Energi Karbohidrat Sumber Asam Lemak Esensial :
Oleat, linoleat, dan linolenat
Masalah :
1. Pemanfaatan Lemak Terbatas 2. Deposit Lemak Abdomen Meningkat
Meningkatkan Absorbsi Lemak
Output :
1. Menurunkan Deposit Lemak Abdomen 2. Meningkatkan Penggunaan Minyak Sawit
Upaya Penyelesaian Masalah :
Suplemantasi Asam Amino
Kerugian :
480 ton Limbah Lemak Abdomen
Kerugian :
Efisiensi Pakan Menurun

4
TINJAUAN PUSTAKA
Lemak pada Nutrisi Unggas
Lemak merupakan senyawa yang tersusun dari asam lemak dan gliserol (Baião dan Lara 2005). Penggunaan lemak pada pakan unggas dapat menurunkan heat increment dan meningkatkan kecernaan nutrien (Kessler et al. 2009). Oleh karena itu, penggunaan lemak dapat meningkatkan efisiensi energi dan performa, serta dapat memberikan pengaruh “extra caloric”. Keuntungan lain penggunaan lemak pada pakan adalah meningkatkan palatabilitas, mengurangi debu dan kehilangan pakan, serta sebagai pelumas untuk alat di pabrik pakan. Sumber lemak yang sering digunakan pada pakan unggas adalah lemak sapi, lemak babi, minyak bunga matahari, minyak kedelai, dan minyak sawit (Firman et al. 2008). Unggas yang baru menetas memiliki keterbatasan fisiologis untuk mencerna dan menyerap lemak. Hal ini disebabkan karena sekresi garam empedu dan sintesis L-karnitin yang rendah (Krogdahl 1985; Arslan 2006). Oleh karena itu, kecernaan lemak pada ayam muda 6% lebih rendah dibandingkan ayam jantan dewasa (Freitas et al. 2005). Keterbatasan unggas muda dalam mencerna lemak menyebabkan kerugian ekonomi, karena penggunaan lemak pakan menjadi kurang efisien (Kessler et al. 2009).
Minyak sawit merupakan minyak yang paling banyak digunakan dalam industri pakan Indonesia. Minyak sawit diproduksi dari daging buah yang lunak dan biji buah sawit. Minyak sawit banyak mengandung asam lemak jenuh, jadi penggunaannya terbatas pada unggas muda (Leeson dan Summer 2005). Profil asam lemak minyak sawit terdiri dari 3.80% asam laurat (12:0), 2.12% asam miristat (14:0), 39.23% asam palmitat (16:0), 5.07% asam stearat (18:0), 38.21% asam oleat (18:1), dan 9.86% asam linoleat (18:2) (Pesti et al. 2002). Selanjutnya, kandungan energi metabolis minyak sawit ayam broiler muda sampai berumur tiga minggu adalah 7200 kkal kg-1, sedangkan pada umur diatas tiga minggu 8000 kkal kg-1. Nilai energi metabolis minyak sawit untuk ayam muda lebih rendah dibandingkan yang lebih tua. Hal ini disebabkan karena kecernaan lemak ayam broiler muda lebih rendah dibandingkan yang lebih tua (Baião dan Lara 2005). Sebagai tambahan, Leeson dan Summer (2005) menyatakan bahwa energi metabolis lemak pada ayam broiler umur 1-7 hari hanya 88% dari energi metabolis pada umur 28 hari.
Peran Glisina dan Taurina dalam Biosintesis Garam Empedu
Asam amino merupakan nutrien penting pada nutrisi unggas dan cukup berpengaruh terhadap biaya formulasi pakan (Nascimento 2004). Oleh karena itu, penentuan kebutuhan asam amino yang akurat dapat menurunkan biaya pakan (Suida 2001). Gisina tergolong sebagai asam amino non esensial pada unggas, tetapi sintesis glisina tidak mencukupi untuk pertumbuhan yang optimal (Corzo et
al. 2004; Dean et al. 2006; Powell et al. 2009). Glisina terlibat dalam berbagai fungsi metabolik yang biasanya berperan dalam sintesis protein, seperti glutation dan keratin. Glisina juga dapat dikonversi menjadi serina, sehingga kebutuhan glisina dipengaruhi oleh kandungan serina pakan (Dean et al. 2006; Corzo 2012).

Selanjutnya, tnon karena taurin tidak berikatan dengan senyawa protein oleh ikatan peptida (Suarez 2006).sulfat dibkurang Taurinataurina homeostasi kalsium (Lambert 2004). dan empedu
menjadi asam tersebut merupakan bentuk utama asam empedu (80%), sedangkan sisanya berasal dari produk sekunder yang dikenodeoksikolatdeoksikolatakan berkonjugasi dengan gProses konjugasi asam empedu menghasilkan ionisasi yang lebih baik, sehingga meningkatkan kemampuan membentuk kenodeoksikolatmembentuk asam glikoberkonjugasi dengan asam amino dan asam disajikan
Selanjutnya, tnon esensial yang karena taurin tidak berikatan dengan senyawa protein oleh ikatan peptida (Suarez 2006). Martin sulfat dibandkurang untuk mendukung pertumbuhan yang maksimum pada ayamTaurina tidak dapat digunakan untuk sintesis protein atau sumber energi, tetapi taurina berpengaruh dalam stabilitas membran, modulasi phomeostasi kalsium (Lambert 2004). dan taurina berperan empedu (Corzo
Asam empedu disintesis dari kolesterol melalui serangkaian proses oksidasi menjadi asam tersebut merupakan bentuk utama asam empedu (80%), sedangkan sisanya berasal dari produk sekunder yang dikenodeoksikolatdeoksikolat akan berkonjugasi dengan gProses konjugasi asam empedu menghasilkan ionisasi yang lebih baik, sehingga meningkatkan kemampuan membentuk kenodeoksikolatmembentuk asam glikoberkonjugasi dengan asam amino dan asam taurodisajikan pada Gambar 2
Gambar 2
Selanjutnya, taurina (2-
esensial yang banyak ditemukan pada jakarena taurin tidak berikatan dengan senyawa protein oleh ikatan peptida (Suarez
Martin et al. (1966) mengemukakan bahwa taurina banyak disintesis dari andingkan dari
untuk mendukung pertumbuhan yang maksimum pada ayamtidak dapat digunakan untuk sintesis protein atau sumber energi, tetapi berpengaruh dalam stabilitas membran, modulasi p
homeostasi kalsium (Lambert 2004). berperan penting
(Corzo et al. 2004; Powell Asam empedu disintesis dari kolesterol melalui serangkaian proses oksidasi
menjadi asam kenodeoksikolattersebut merupakan bentuk utama asam empedu (80%), sedangkan sisanya berasal dari produk sekunder yang dikenodeoksikolat dirubah
(Gropper et al.
akan berkonjugasi dengan gProses konjugasi asam empedu menghasilkan ionisasi yang lebih baik, sehingga meningkatkan kemampuan membentuk misel (Gropper kenodeoksikolat dan asam membentuk asam glikoberkonjugasi dengan asam amino
taurokolat. Proses pada Gambar 2
Proses konjugasi asam empedu dengan glisina dan taurina (Sumber : Märt 2013)
-aminoethanesulfonic acid
banyak ditemukan pada jakarena taurin tidak berikatan dengan senyawa protein oleh ikatan peptida (Suarez
(1966) mengemukakan bahwa taurina banyak disintesis dari dari sisteina,
untuk mendukung pertumbuhan yang maksimum pada ayamtidak dapat digunakan untuk sintesis protein atau sumber energi, tetapi berpengaruh dalam stabilitas membran, modulasi p
homeostasi kalsium (Lambert 2004). penting dalam 2004; Powell et al.
Asam empedu disintesis dari kolesterol melalui serangkaian proses oksidasi kenodeoksikolat dan asam
tersebut merupakan bentuk utama asam empedu (80%), sedangkan sisanya berasal dari produk sekunder yang diproduksi dari hasil aktifitas bakteri
dirubah bakteri et al. 2009; Cheeke
akan berkonjugasi dengan glisina dan Proses konjugasi asam empedu dengan glisina dan taurinamenghasilkan ionisasi yang lebih baik, sehingga meningkatkan kemampuan
(Gropper et al. 2009; sam kolat berkonjugasi dengan asam amino g
membentuk asam glikokenodeoksikolatberkonjugasi dengan asam amino taurina
. Proses konjugasi asam empedu pada Gambar 2.
konjugasi asam empedu dengan glisina dan taurina (Sumber :
aminoethanesulfonic acid
banyak ditemukan pada jaringan sebagai asam amino bebas, karena taurin tidak berikatan dengan senyawa protein oleh ikatan peptida (Suarez
(1966) mengemukakan bahwa taurina banyak disintesis dari isteina, serta taurina
untuk mendukung pertumbuhan yang maksimum pada ayamtidak dapat digunakan untuk sintesis protein atau sumber energi, tetapi berpengaruh dalam stabilitas membran, modulasi p
homeostasi kalsium (Lambert 2004). Kedua asam amino tersebut, yaitu gdalam proses konjugasi asam empedu menjadi garam
et al. 2009; Gropper Asam empedu disintesis dari kolesterol melalui serangkaian proses oksidasi
dan asam kolat tersebut merupakan bentuk utama asam empedu (80%), sedangkan sisanya berasal
produksi dari hasil aktifitas bakteribakteri menjadi litokolat dan asam kolat menjadi 2009; Cheeke dan
dan taurina membentuk asam empdengan glisina dan taurina
menghasilkan ionisasi yang lebih baik, sehingga meningkatkan kemampuan 2009; Stamp
berkonjugasi dengan asam amino gkenodeoksikolat dan asam
taurina membentuk asam taurokonjugasi asam empedu
konjugasi asam empedu dengan glisina dan taurina (Sumber :
aminoethanesulfonic acid) juga merupakan asam amino ringan sebagai asam amino bebas,
karena taurin tidak berikatan dengan senyawa protein oleh ikatan peptida (Suarez (1966) mengemukakan bahwa taurina banyak disintesis dari
taurina diproduksi untuk mendukung pertumbuhan yang maksimum pada ayamtidak dapat digunakan untuk sintesis protein atau sumber energi, tetapi berpengaruh dalam stabilitas membran, modulasi p
Kedua asam amino tersebut, yaitu gproses konjugasi asam empedu menjadi garam
2009; Gropper et al.
Asam empedu disintesis dari kolesterol melalui serangkaian proses oksidasi di dalam hepatosit
tersebut merupakan bentuk utama asam empedu (80%), sedangkan sisanya berasal produksi dari hasil aktifitas bakteri
litokolat dan asam kolat menjadi Dierendfeld 2010). Asam empedu
membentuk asam empdengan glisina dan taurina
menghasilkan ionisasi yang lebih baik, sehingga meningkatkan kemampuan dan Jenkins 2008).
berkonjugasi dengan asam amino gdan asam glikokolatmembentuk asam tauro
konjugasi asam empedu dengan glisina dan taurina
konjugasi asam empedu dengan glisina dan taurina (Sumber :
juga merupakan asam amino ringan sebagai asam amino bebas,
karena taurin tidak berikatan dengan senyawa protein oleh ikatan peptida (Suarez (1966) mengemukakan bahwa taurina banyak disintesis dari
diproduksi dalam jumlah yang untuk mendukung pertumbuhan yang maksimum pada ayamtidak dapat digunakan untuk sintesis protein atau sumber energi, tetapi berpengaruh dalam stabilitas membran, modulasi pertumbuhan dan
Kedua asam amino tersebut, yaitu gproses konjugasi asam empedu menjadi garam
et al. 2009). Asam empedu disintesis dari kolesterol melalui serangkaian proses oksidasi
di dalam hepatosit. Asam tersebut merupakan bentuk utama asam empedu (80%), sedangkan sisanya berasal
produksi dari hasil aktifitas bakterilitokolat dan asam kolat menjadi
Dierendfeld 2010). Asam empedu membentuk asam empedu konjugasi.
dengan glisina dan taurina bertujuan untuk menghasilkan ionisasi yang lebih baik, sehingga meningkatkan kemampuan
Jenkins 2008). Pada manusia, berkonjugasi dengan asam amino glisina
glikokolat. Sisanya (25%) membentuk asam taurokenodeoksikolat
dengan glisina dan taurina
konjugasi asam empedu dengan glisina dan taurina (Sumber :
5
juga merupakan asam amino ringan sebagai asam amino bebas,
karena taurin tidak berikatan dengan senyawa protein oleh ikatan peptida (Suarez (1966) mengemukakan bahwa taurina banyak disintesis dari
dalam jumlah yang untuk mendukung pertumbuhan yang maksimum pada ayam broiler. tidak dapat digunakan untuk sintesis protein atau sumber energi, tetapi
ertumbuhan dan Kedua asam amino tersebut, yaitu glisina
proses konjugasi asam empedu menjadi garam
Asam empedu disintesis dari kolesterol melalui serangkaian proses oksidasi . Asam empedu
tersebut merupakan bentuk utama asam empedu (80%), sedangkan sisanya berasal produksi dari hasil aktifitas bakteri. Asam
litokolat dan asam kolat menjadi Dierendfeld 2010). Asam empedu
edu konjugasi. bertujuan untuk
menghasilkan ionisasi yang lebih baik, sehingga meningkatkan kemampuan Pada manusia,
lisina (75%) isanya (25%)
kenodeoksikolat dengan glisina dan taurina
konjugasi asam empedu dengan glisina dan taurina (Sumber :
edu konjugasi.

6
Peran Garam Empedu pada Proses Pencernaan dan Penyerapan Lemak
Kantung empedu merupakan tempat pemekatan dan penyimpanan empedu yang dibuat oleh hati sampai empedu dibutuhkan untuk mencerna lemak di usus halus. Empedu adalah cairan berwarna hijau kekuningan yang susunan utamanya adalah asam empedu dan garam empedu (Gropper et al. 2009). Volume sekresi empedu dipengaruhi oleh laju sirkulasi darah, status nutrisi unggas, tipe pakan yang diberikan dan efisiensi sirkulasi enterohepatik garam empedu (Marzooqi dan Leeson 1998). Hormon kolesistokinin bekerja menstimulasi kontraksi kantung empedu dan melepaskan empedu ke duodenum. Sebaliknya, somatostatin bekerja menghambat kontraksi kantung empedu (Gropper et al. 2009). Asam empedu dan garam empedu berfungsi untuk mengemulsi lemak di saluran pencernaan (Gropper et al. 2009; Cheeke dan Dierendfeld 2010).
Salah satu aspek fundamental dalam proses pencernaan lemak adalah proses melarutkan lemak pada media cair saluran pencernaan yang biasa disebut emulsifikasi. Proses pengemulsian lemak terjadi di dalam usus halus dengan bantuan garam empedu dan phospolipid sebagai pengemulsi. Garam empedu dapat mengemulsikan lemak karena memiliki kemampuan amphipathik, yang berarti bahwa garam empedu dapat larut pada media air dan lemak karena adanya variasi polaritas di dalam molekulnya (Cheeke dan Dierenfeld 2010). Pengemulsian lemak berfungsi memfasilitasi kerja dari lipase pankreas, untuk pembentukan misel. Misel merupakan senyawa yang tersusun atas asam lemak dan kolesterol pada bagian tengahnya, sedangkan bagian luarnya dikelilingi oleh garam empedu (Larbier dan Leclerq 1994). Peran garam empedu dalam proses pencernaan dan penyerapan lemak disajikan pada Gambar 3.
Peran Lisina dan Metionina dalam Biosintesis L-karnitin
Lisina dan metionina merupakan dua asam amino yang paling esensial pada pakan yang berbasis jagung dan bungkil kedelai. Oleh karena itu, suplementasi
Gambar 3 Peran garam empedu dalam proses pencernaan dan penyerapan lemak
(Sumber : Liz 2013)

7
lisina dan metionina sintetik pada pakan ayam broiler biasa dilakukan industri pakan unggas untuk memenuhi kebutuhan asam amino tersebut dan menurunkan biaya pakan (Si et al. 2001). Lisina merupakan asam amino yang sangat penting untuk pertumbuhan unggas. Defisiensi lisina menyebabkan pertumbuhan terganggu, depigmenasi pada unggas dan penurunan hemoglobin dan hematokrit pada ayam broiler (Osti dan Pandey 2004). Selanjutnya, metionina juga merupakan asam amino bersulfur yang berperan dalam biosintesis protein dan anti bodi. Metionina merupakan pendonor gugus metil utama di dalam tubuh yang dibutuhkan untuk biosintesis senyawa penting seperti keratin, poliamin, epineprin, melatonin, kolin dan karnitin (Baker et al. 1996). Dua asam amino tersebut, yaitu lisina dan metionina merupakan prekursor L-karnitin yang berperan penting dalam metabolisme lemak dan energi (Arslan 2006). Oleh karena itu, penelitian pada manusia menunjukan bahwa konsumsi lisina dan metionina berkorelasi positif dengan kadar karnitin dalam darah (Kraj�ovi�ova-Kudlá�ková et al. 2000).
Karnitin merupakan senyawa amina larut air (�-hydroxy �-trimethylamino-
butyrate) yang ditemukan pada jaringan manusia, binatang, tumbuhan, dan beberapa mikroorganisme dengan konsentrasi yang berbeda-beda. Karnitin berfungsi seperti vitamin dalam proses katabolisme lemak dan penyediaan energi (Walter 2000). Pada manusia dengan pola makan normal, sekitar � kebutuhan karnitin berasal dari makanan, sedangkan sisanya berasal dari biosintesis di dalam tubuh (Krähenbuhl 1996). Proses biosintesis karnitin melibatkan empat rantai karbon dari lisina, dan gugus metil dari metionina. Biosintesis karnitin diawali dengan proses metilasi asam amino lisina oleh S-adenosylmetionina (SAM). Metionina, magnesium, asam askorbat, besi, dan pyridoxal-5’-phospat (P5P) dan niasin berperan sebagai kofaktor untuk memperbaharui SAM dari homosisteina yang dibutuhkan dalam sintesis karnitin (Gregory dan Kelly 1998). Peran lisina dan metionina pada proses biosintesis L-karnitin disajikan pada Gambar 4.
Gambar 4 Peran lisina dan metionina dalam proses biosintesis L-karnitin
(Sumber : Scholte 2003)

8
Peran L-Karnitin dalam Metabolisme Lemak
L-karnitin memiliki banyak fungsi di dalam tubuh, tetapi ada dua fungsi utama L-karnitin. Fungsi yang pertama adalah sebagai katalisator pembakaran asam lemak di mitokondria dan fungsi yang kedua adalah sebagai buffer terhadap kelebihan residu asil (Arslan 2006). L-karnitin pada proses �-oksidasi digunakan sebagai sarana transportasi asam lemak rantai panjang untuk masuk ke membran dalam mitokondria. Proses �-oksidasi diawali dengan masuknya asam lemak yang berasal dari makanan atau jaringan adiposa ke dalam sel melalui sirkulasi darah. Asam lemak kemudian dibawa ke sitosol dan ditransfer ke mitokondria. Transportasi asam lemak dari sitosol ke mitokondria diawali dengan aktivasi asam lemak pada membran luar mitokondria dengan bantuan enzim Acyl-CoA-
Synthetase menjadi senyawa asil-KoA-lemak. Selanjutnya, asil-KoA-lemak akan mengikat L-karnitin menjadi asil-karnitin-lemak untuk ditransportasikan ke matrik mitokondria. Asil-karnitin-lemak kemudian dikonversi menjadi asil-KoA lemak dan melepaskan karnitin bebas. Asil-KoA lemak di matrik mitondria dirubah menjadi energi melalui B-oksidasi (Gropper et al. 2009).
L-karnitin juga berfungsi membantu menjaga rasio asetil-KoA/KoA bebas tetap optimal. Pembentukan energi dalam mitokondria dapat berlangsung normal jika KoA bebas di matrik mitokondria tersedia dalam jumlah cukup. Hal ini disebabkan karena KoA bebas dibutuhkan sebagai kofaktor pada banyak reaksi seluler. L-karnitin berfungsi menyimpan kelebihan residu asetil dengan membawa kelebihan asetil dari asetil-KoA, sehingga KoA bebas dapat digunakan pada reaksi selular yang lainnya. Berdasarkan fakta ini, maka L-karnitin berfungsi sebagai buffer asetil atau deposit intermediet dari kelompok asetil (Stevens 1996; Gibson dan Harris 2002; Gropper et al. 2009).
Gambar 5 Peran L-karnitin pada proses metabolisme asam lemak (Sumber :
Mehta 2013)

9
Lemak Abdomen pada Ayam Broiler
Lemak di dalam tubuh ayam broiler dapat berasal dari luar tubuh (makanan), dalam tubuh (sintesis di hati) dan tulang. Deposit lemak biasanya terjadi ketika keseimbangan energi positif atau kandungan nutrien pakan yang tidak seimbang (Haro 2005). Lemak biasanya disimpan pada lemak abdomen (termasuk lemak di sekitar gizard, proventrikulus, bursa fabrisius, kloaka, dan perbatasan dengan daging), lemak sartorial (lemak subkutikular di daerah kranial paha), lemak di daerah leher (lemak subkutikular daerah leher ventro-kaudal) dan lemak mesentrium (lemak yang menempel pada mesentrium saluran pencernaan). Lemak abdomen merupakan tempat utama untuk deposit lemak yang berhubungan erat dengan lemak karkas ayam broiler. Hal ini berdasarkan kenyataan dimana lemak tubuh terdiri dari 20% lemak abdomen, 18% lemak subkutan, 15% lemak tulang, serta 2.5% hati dan bulu (T�mová dan Teimouri 2010). Berat relatif lemak abdomen biasanya sekitar 2 sampai 3% dari bobot badan ayam broiler (Leeson dan Summer 2005). Produksi lemak abdomen dapat diprediksi dengan menggunakan metode ultrasound dengan persamaan LA = -48.566 + 4.5265 (PLA) + 0.04240 BB. Dimana LA = lemak abdomen (g) ; PLA = permukaan lemak abdomen (mm2) dan BB = bobot badan ayam broiler (g). Persamaan tersebut memiliki nilai R2 = 0.74 dan r = 0.64 (Melo et al. 2003).
Deposit lemak abdomen dipengaruhi oleh umur, jenis kelamin, iklim pada suatu daerah dan terutama oleh faktor genetik dan pakan (Jones dan Farrell 1987). Secara genetik, diestimasikan peningkatan performa ayam broiler secara bersamaam meningkatkan berat lemak abdomen sekitar 0.39 g per tahun (Gaya et
al. 2005). Deposit lemak abdomen juga sangat dipengaruhi pakan, khususnya jumlah lemak dan profil asam lemak pakan. Pakan yang mengandung lemak tinggi akan meningkatkan berat dan persentase lemak abdomen (Jones dan Farrell 1989; Pesti et al. 2002). Selanjutnya, penggunaan minyak tumbuhan yang kaya asam lemak tidak jenuh seperti minyak biji bunga matahari dilaporkan dapat menurunkan deposit lemak abdomen ayam broiler (Sanz et al. 2000; Crespo dan Esteve-Garcia 2001).
Biosintesis Lemak Abdomen
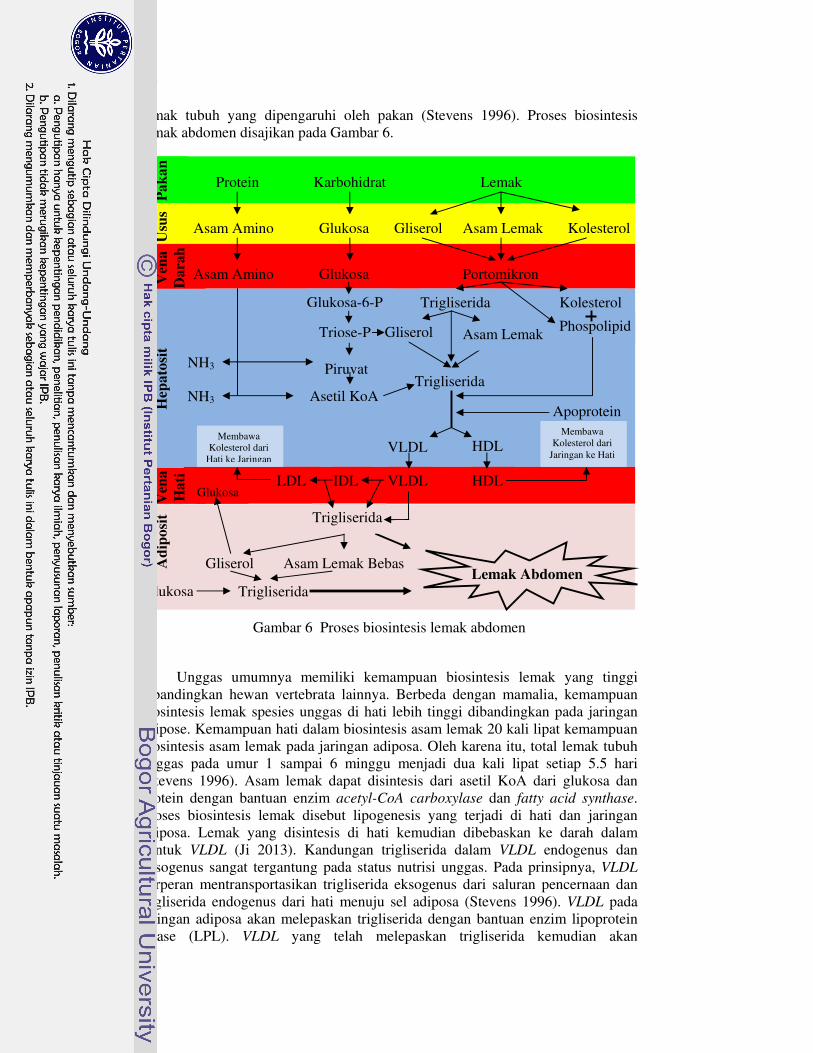
Lemak abdomen merupakan bagian dari jaringan adiposa yang berfungsi menyimpan kelebihan energi. Di dalam tubuh, energi disimpan dalam bentuk trigliserida yang terdiri dari tiga senyawa asam lemak terestersifikasi dengan gliserol (Ji 2013). Asam lemak penyusun trigliserida dapat berasal dari luar tubuh (pakan) dan dari dalam tubuh (biosintesis), sedangkan gliserol diproduksi dari glukosa (Gibson dan Harris 2002; Gropper et al. 2009). Asam lemak hasil pencernaan lemak pakan diserap oleh enterosit dengan dibungkus kolesterol, lipoprotein, dan lemak lainnya menjadi protomikron (sama dengan kilomikron pada mamalia) kemudian ditransportasikan pada darah. Protomikron di darah kemudian dikirim ke hati untuk dikonversi menjadi very low density lipoprotein
(VLDL) dan high density lipoprotein (HDL), sebagian lainnya dikirim ke jaringan adiposa untuk dideposit atau dioksidasi menjadi energi. Pada unggas yang diberi pakan butiran, kontribusi portomikron dari pakan sangat rendah karena hanya 5%
10
lemak tubuh yang dipengaruhi oleh pakan (Stevens 1996). Proses biosintesis lemak abdomen disajikan pada Gambar 6.
Gambar 6 Proses biosintesis lemak abdomen
Unggas umumnya memiliki kemampuan biosintesis lemak yang tinggi
dibandingkan hewan vertebrata lainnya. Berbeda dengan mamalia, kemampuan biosintesis lemak spesies unggas di hati lebih tinggi dibandingkan pada jaringan adipose. Kemampuan hati dalam biosintesis asam lemak 20 kali lipat kemampuan biosintesis asam lemak pada jaringan adiposa. Oleh karena itu, total lemak tubuh unggas pada umur 1 sampai 6 minggu menjadi dua kali lipat setiap 5.5 hari (Stevens 1996). Asam lemak dapat disintesis dari asetil KoA dari glukosa dan protein dengan bantuan enzim acetyl-CoA carboxylase dan fatty acid synthase. Proses biosintesis lemak disebut lipogenesis yang terjadi di hati dan jaringan adiposa. Lemak yang disintesis di hati kemudian dibebaskan ke darah dalam bentuk VLDL (Ji 2013). Kandungan trigliserida dalam VLDL endogenus dan eksogenus sangat tergantung pada status nutrisi unggas. Pada prinsipnya, VLDL berperan mentransportasikan trigliserida eksogenus dari saluran pencernaan dan trigliserida endogenus dari hati menuju sel adiposa (Stevens 1996). VLDL pada jaringan adiposa akan melepaskan trigliserida dengan bantuan enzim lipoprotein lipase (LPL). VLDL yang telah melepaskan trigliserida kemudian akan
Ad
iposi
t V
ena
Hati
VLDL HDL IDL LDL
Asam Lemak Bebas Gliserol Lemak Abdomen
Trigliserida Glukosa
Trigliserida
Glukosa
Usu
s
Protein Lemak Karbohidrat P
ak
an
Asam Amino Asam Lemak Glukosa Gliserol Kolesterol
Ven
a
Da
rah
Asam Amino Glukosa Portomikron
Gliserol
Glukosa-6-P
Piruvat
VLDL
Hep
ato
sit
Triose-P
Asetil KoA
Trigliserida Kolesterol
Phospolipid Asam Lemak
Apoprotein
Trigliserida
HDL
NH3
Membawa Kolesterol dari
Jaringan ke Hati
Membawa Kolesterol dari
Hati ke Jaringan
NH3

11
membentuk low density lipoprotein (LDL) yang bersama HDL berperan dalam proses metabolisme kolesterol (lihat Gambar 6). Trigliserida pada jaringan adiposa kemudian akan di deposit menjadi lemak tubuh seperti lemak abdomen (Gropper et al. 2009).
Unggas yang diberi pakan berbasis butiran yang rendah lemak, komposisi asam lemak yang disimpan dalam tubuh kebanyakan terdiri dari asam lemak jenuh (saturated), seperti asam palmitat (16:0) dan stearat (18:0) (Gurr 1992). Hal ini disebabkan karena asam lemak banyak disintesis dari karbohidrat, ditambah kemampuan unggas yang rendah sintesis asam lemak tidak jenuh (lihat Gambar 6). Selanjutnya, profil asam lemak pada pakan berhubungan dengan profil asam lemak pada lemak abdomen. Misalnya, ayam broiler yang diberi pakan mengandung minyak sawit akan meningkatkan konsentrasi asam lemak palmitat dan stearat pada lemak abdomen dibandingkan pakan yang mengandung biji dan minyak bunga matahari (Ortiz et al. 2006). Penambahan minyak pada pakan akan meningkatkan konsumsi lemak, sehingga menyebabkan penurunan laju sintesis de
novo asam lemak. Hal ini disebabkan karena menurunnya aktifitas enzim ATP
citrate lyase dan malate di hati untuk mengkatalisis produksi asetil KoA yang dibutuhkan dalam biosintesis asam lemak (Stevens 1996). Oleh karena itu, penambahan minyak pada pakan akan meningkatkan deposit lemak dari pakan dan menurunkan deposit lemak dari hasil biosintesis lemak di dalam tubuh.
Metode Menurunkan Deposit Lemak Abdomen
Ada tiga metode utama dalam menurunkan deposit lemak abdomen ayam broiler, yaitu melalui rekayasa genetik, manajemen pemeliharaan dan nutrisi dalam pakan. Secara genetik, ayam broiler modern memiliki pertumbuhan yang cepat dan konversi pakan yang rendah. Akan tetapi, peningkatan pertumbuhan menyebabkan peningkatan deposit lemak abdomen, kematian, dan gangguan metabolis. Oleh karena itu, industri pembibitan berusaha untuk menciptakan ayam broiler dengan karakteristik pertumbuhan cepat, tetapi rendah deposit lemak abdomen. T�mová dan Teimouri (2010) melaporkan bahwa urutan strain ayam broiler dengan lemak abdomen terendah adalah Cobb 500 < Hubbard < Hybro=Ross 308 < Arbor Arces. Kesimpulannya adalah strain dengan deposit lemak abdomen terendah adalah Cobb 500 dan tertinggi adalah Arbor Arces.
Manajemen pemeliharaan ayam broiler juga berpengaruh terhadap deposit lemak abdomen khususnya sistem pemberian pakan. Penelitian sebelumnya menemukan bahwa pembatasan pakan (Hakimi 1992; Jones dan Farrel 1987) dan skip day feeding (Nosrati et al. 2006) nyata menurunkan lemak abdomen ayam broiler. Hasil penelitian Hakimi (1992) menunjukan bahwa pembatasan pakan 90% dari konsumsi ad libitum secara nyata menurunkan lemak abdomen. Selanjutnya, penelitian Nosrati et al. (2006) menemukan bahwa skip feeding pada umur 18-28 hari dapat menurunkan deposit lemak abdomen. Faktor manajemen lainnya seperti cuaca, tipe kandang, tipe lantai, dan pencahayaan gagal untuk menurunkan lemak abdomen (Hakimi 1992).
Deposit lemak abdomen secara nyata dapat diturunkan dengan manipulasi nutrisi pada pakan. Manipulasi pakan dapat dilakukan dengan mengatur keseimbangan energi dengan protein, manipulasi profil asam lemak dan

12
suplementasi herbal. Kandungan energi dan protein pada pakan berpengaruh besar terhadap komposisi karkas, karena keduanya dibutuhkan untuk pertumbuhan (T�mová dan Teimouri 2010). Oleh karena itu, keseimbangan nutrien pakan harus diperhatikan dengan menghitung rasio energi dan protein. Rasio energi dan protein yang tinggi akan menyebabkan energi banyak dideposit sebagai lemak, sedangkan apabila rasionya rendah akan menyebabkan kelebihan protein. Selanjutnya, Leeson dan Summer (2005) merekomendasikan rasio energi dan protein yang optimal untuk ayam broiler umur 35 sampai 49 hari adalah 178.4.
Jumlah lemak dan profil asam lemak dalam pakan berpengaruh terhadap deposit lemak abdomen. Wongsuthavas (2007) melaporkan bahwa pakan yang mengandung lemak 9% nyata meningkatkan lemak abdomen dibandingkan pakan yang mengandung lemak 3 dan 6%. Selanjutnya, Crespo dan Esteve-Garcia (2001) dan Poorghasemi et al. (2013) melaporkan bahwa penggunaan minyak biji bunga matahari dapat menurunkan lemak abdomen dibandingkan menggunakan lemak sapi dan minyak canola. Minyak biji bunga matahari kaya akan asam lemak tidak jenuh, sehingga penggunaan minyak biji bunga matahari akan meningkatkan asam lemak tidak jenuh pada pakan. Ayam broiler yang diberi pakan mengandung asam lemak tidak jenuh tinggi dapat mengurunkan deposit lemak abdomen. Hal ini disebabkan karena ayam yang diberi pakan mengandung asam lemak tidak jenuh dapat menurunkan sintesis lemak di dalam tubuh dan meningkatkan oksidasi lemak menjadi energi (Sanz et al. 2000; Wongsuthavas 2007). Selanjutnya, Nosrati et al. (2006) dan Wongsuthavas (2007) merekomendasikan rasio asam lemak jenuh dengan asam lemak tidak jenuh yang direkomendasikan untuk menurunkan lemak abdomen adalah 1:5.
Banyak penelitian telah dilakukan untuk menurunkan deposit lemak abdomen ayam broiler dengan suplementasi tanaman herbal. Wardah et al. (2012) melaporkan bahwa suplementasi 5% tepung daun Phyllanthus buxifolius nyata menurunkan lemak abdomen, lemak intramuskuler, dan lemak daging. Hal ini disebabkan karena suplementasi Phyllanthus buxifolius menyebabkan peningkatan kandungan serat dalam pakan. Serat berfungsi menghambat penyerapan lemak di dalam saluran pencernaan dan mengurangi lipogenesis di dalam tubuh. Suplementasi bawang putih 4% juga dilaporkan dapat menurunkan lemak abdomen dengan cara menekan biosintesis trigliserida di hati (Shahriari et al. 2009). Ma�lank dan Pisarski (2009) menambahkan suplementasi 2% Viola
tricolor L., Mentha piperita L. dan Urtica dioica L. dapat menurunkan lemak abdomen dengan cara meningkatkan rasio asam lemak tidak jenuh dengan asam lemak jenuh, sehingga meningkatkan oksidasi lemak.

13
METODE
Bahan
Penelitian ini menggunakan 240 ekor ayam broiler strain Cobb dan 24 petak kandang. Bahan-bahan yang digunakan menyusun ransum basal penelitian seperti jagung, dedak padi, bungkil kedelai, minyak sawit, asam amino sintetik dan premix vitamin dan mineral. Bahan-bahan yang digunakan untuk analisis bahan kering, protein kasar dan lemak kasar (AOAC 1995). Complete Test Kit dan akuades untuk analisis asam urat, trigliserida dan kolesterol serum. Bahan-bahan yang digunakan untuk analisis L-karnitin daging seperti larutan standar L-karnitin, larutan Ba(OH)2 dan NaH2PO4.H2O.
Alat
Alat-alat yang digunakan selama pemeliharaan ayam broiler seperti tempat pakan, tempat minum dan alat timbang. Alat-alat yang digunakan untuk analisis bahan kering, protein kasar dan lemak kasar (AOAC 1995). Alat-alat yang digunakan untuk analisis asam urat, trigliserida dan kolesterol serum seperti spektrofotometer, mikropipet dan sentrifus. Alat-alat yang digunakan analisis koefisien kecernaan semu nutrien dan energi seperti kandang metabolis, oven, mortal dan blender. Kalorimeter tipe Parr 6200 Calorimeter untuk analisis kandungan energi bruto pakan perlakuan dan ekskreta. Alat-alat yang digunakan untuk analisis L-karnitin daging seperti High Performance Liquid Cromatography (HPLC) tipe Waters Alliance E2695, detektor UV-VIS 2489, labu ukur dan pipet. Alat-alat yang digunakan untuk analisis berat relatif karkas dan organ seperti pisau, gunting bedah dan nampan.
Waktu dan Lokasi Percobaan
Penelitian telah dilaksanakan pada bulan Oktober 2012 sampai Pebruari 2013. Pemeliharaan ayam broiler dilakukan di Laboratorium Lapang Departemen Ilmu Nutrisi dan Teknologi Pakan Institut Pertanian Bogor (IPB). Analisis profil darah dilakukan di Laboratorium Ilmu Nutrisi Ternak Daging dan Kerja IPB. Analisis proksimat kandungan nutrien pakan, ekskreta, dan daging dilakukan di Laboratorium Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB. Analisis energi bruto dilakukan di Laboratorium Ilmu dan Teknologi Pakan IPB. Analisis L-karnitin daging dilakukan di Laboratorium Saraswanti Bogor. Analisis koefisien kecernaan semu nutrien dan energi, serta berat relatif karkas dan organ dilakukan di Laboratorium Nutrisi Unggas IPB.
Rancangan Percobaan
Penelitian ini menggunakan rancangan acak lengkap (RAL) dengan enam perlakuan dan empat ulangan, setiap ulangan terdiri dari sepuluh ekor ayam
14
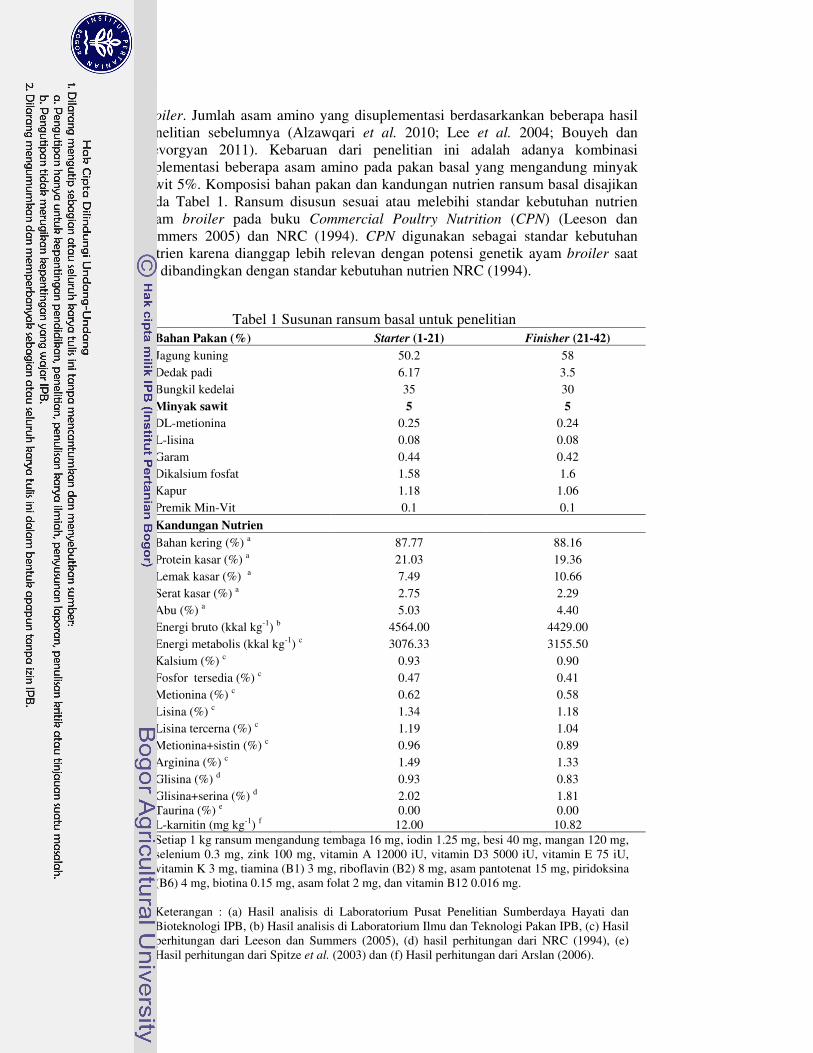
broiler. Jumlah asam amino yang disuplementasi berdasarkankan beberapa hasil penelitian sebelumnya (Alzawqari et al. 2010; Lee et al. 2004; Bouyeh dan Gevorgyan 2011). Kebaruan dari penelitian ini adalah adanya kombinasi suplementasi beberapa asam amino pada pakan basal yang mengandung minyak sawit 5%. Komposisi bahan pakan dan kandungan nutrien ransum basal disajikan pada Tabel 1. Ransum disusun sesuai atau melebihi standar kebutuhan nutrien ayam broiler pada buku Commercial Poultry Nutrition (CPN) (Leeson dan Summers 2005) dan NRC (1994). CPN digunakan sebagai standar kebutuhan nutrien karena dianggap lebih relevan dengan potensi genetik ayam broiler saat ini dibandingkan dengan standar kebutuhan nutrien NRC (1994).
Tabel 1 Susunan ransum basal untuk penelitian Bahan Pakan (%) Starter (1-21) Finisher (21-42)
Jagung kuning 50.2 58 Dedak padi 6.17 3.5 Bungkil kedelai 35 30 Minyak sawit 5 5
DL-metionina 0.25 0.24 L-lisina 0.08 0.08 Garam 0.44 0.42 Dikalsium fosfat 1.58 1.6 Kapur 1.18 1.06 Premik Min-Vit 0.1 0.1
Kandungan Nutrien
Bahan kering (%) a 87.77 88.16 Protein kasar (%) a 21.03 19.36 Lemak kasar (%) a 7.49 10.66 Serat kasar (%) a 2.75 2.29 Abu (%) a 5.03 4.40 Energi bruto (kkal kg-1) b 4564.00 4429.00 Energi metabolis (kkal kg-1) c 3076.33 3155.50 Kalsium (%) c 0.93 0.90 Fosfor tersedia (%) c 0.47 0.41 Metionina (%) c 0.62 0.58 Lisina (%) c 1.34 1.18 Lisina tercerna (%) c 1.19 1.04 Metionina+sistin (%) c 0.96 0.89 Arginina (%) c 1.49 1.33 Glisina (%) d 0.93 0.83 Glisina+serina (%) d 2.02 1.81 Taurina (%) e 0.00 0.00 L-karnitin (mg kg-1) f 12.00 10.82 Setiap 1 kg ransum mengandung tembaga 16 mg, iodin 1.25 mg, besi 40 mg, mangan 120 mg, selenium 0.3 mg, zink 100 mg, vitamin A 12000 iU, vitamin D3 5000 iU, vitamin E 75 iU, vitamin K 3 mg, tiamina (B1) 3 mg, riboflavin (B2) 8 mg, asam pantotenat 15 mg, piridoksina (B6) 4 mg, biotina 0.15 mg, asam folat 2 mg, dan vitamin B12 0.016 mg. Keterangan : (a) Hasil analisis di Laboratorium Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB, (b) Hasil analisis di Laboratorium Ilmu dan Teknologi Pakan IPB, (c) Hasil perhitungan dari Leeson dan Summers (2005), (d) hasil perhitungan dari NRC (1994), (e) Hasil perhitungan dari Spitze et al. (2003) dan (f) Hasil perhitungan dari Arslan (2006).

15
Perlakuan Penelitian
Perlakuan penelitian ini terdiri dari : R0 = Ransum basal (mengandung minyak sawit 5%) R1 = Ransum basal + 0.25% glisina R2 = Ransum basal + 0.25% taurina R3 = Ransum basal + 15% total lisina dan metionina ransum basal R4 = Ransum basal + 0.25% glisina + 0.25% taurina R5 = Ransum basal + 0.25% glisina + 0.25% taurina + 15% total lisina dan
metionina ransum basal Peubah yang Diamati
Peubah yang diamati pada penelitian ini meliputi : 1. Performa ayam broiler : konsumsi pakan, bobot badan (BB), pertambahan
bobot badan (PBB), konversi pakan dan mortalitas. 2. Profil metabolit serum : asam urat, trigliserida dan kolesterol. 3. Karakteristik empedu : berat relatif empedu, berat organ empedu, serta berat,
volume dan berat jenis cairan empedu. 4. Koefisien kecernaan semu : bahan kering, lemak kasar, protein kasar dan
energi. 5. Komposisi daging : bahan kering, protein kasar, lemak kasar dan L-karnitin. 6. Karakteristik karkas dan organ pencernaan : berat relatif karkas, lemak
abdomen, hati dan pankreas.
Metode Percobaan
Tahap Persiapan Pemeliharaan
Kandang dan peralatannya disucihamakan menggunakan disinfektan. 240 ekor DOC ayam broiler ditempatkan pada 24 unit kandang berukuran 1 m2 secara acak. Tiap kandang berisi satu tempat pakan dan minum, serta lantai kandang dilapisi dengan sekam padi. Pada saat brooding, DOC diberi lampu bulb 60 watt yang berfungsi sebagai penerangan dan penghangat.
Tahap Pelaksanaan Pemeliharaan
Pemberian pakan dan air minum diberikan ad libitum sampai umur 42 hari. Jumlah pakan yang diberikan dan sisa pakan ditimbang setiap minggu untuk menentukan konsumsi pakan. Penimbangan bobot badan dilakukan setiap minggu untuk mengetahui bobot badan dan menghitung pertambahan bobot badan.
Pengambilan Sampel Serum
Pengambilan sampel darah dilakukan pada saat ayam broiler berumur 21 dan 42 hari pada pembuluh darah vena jugularis. Sampel serum perlakuan diambil dari empat ekor ayam broiler yang berasal dari masing-masing ulangan (empat ulangan). Sampel darah diambil sebanyak 1 ml menggunakan disposable syringes. Luka bekas pengambilan darah kemudian dioleskan alkohol 96% untuk menghindari terjadinya infeksi. Sampel darah kemudian disentrifus dengan kecepatan 1.500 rpm selama 15 menit untuk diambil sampel serum darahnya. Serum darah kemudian dianailisis kadar asam urat, trigliserida dan kolesterol.

16
Pengambilan Sampel Ekskreta.
Sampel ekskreta digunakan untuk mengukur koefisien kecernaan semu nutrien dan energi ayam broiler. Sampel ekskreta diambil dari empat ekor ayam broiler setiap perlakuan (satu ekor setiap ulangan). Pengukuran koefisien kecernaan nutrien dilakukan ketika ayam broiler berumur 21 dan 42 hari. Pengukuran koefisien kecernaan nutrien dilakukan dengan menempatkan ayam broiler pada kandang metabolis. Ayam broiler kemudian dipuasakan makan selama 24 jam untuk menghilangkan sisa pakan di saluran pencernaan, kemudian diberi pakan selama 2 jam (08.00-10.00 WIB). Pengumpulan ekskreta dilakukan selama 24 jam setelah pemberian pakan. Setiap empat jam sekali ekskreta disemprot menggunakan larutan H2SO4 0.01% dan formalin 10%. Ekskreta hasil koleksi total selama 24 jam kemudian ditimbang dan dikeringkan dalam oven 50 oC sampai kering. Ekskreta yang telah kering kemudian ditimbang kembali dan dihaluskan menggunakan blender dan mortal. Ekskreta kemudian dianalisis kandungan bahan kering, lemak kasar, protein kasar, dan energi bruto.
Pengukuran Berat Relatif Karkas dan Organ
Sampel berat relatif karkas dan organ diambil dari empat ekor ayam broiler dari setiap perlakuan. Pengukuran berat relatif karkas dilakukan pada saat ayam broiler berumur 21 dan 42 hari. Satu ekor ayam broiler dari masing-masing ulangan diambil secara acak kemudian disembelih menggunakan metode penyembelihan Kosher style. Ayam yang telah disembelih dicelupkan pada air dengan suhu 65 oC sampai 80 oC selama 5 sampai 30 detik, kemudian bulu ayam dicabut sampai bersih dan jeroan dikeluarkan. Berat relatif karkas dan organ kemudian dianalisis.
Pengambilan Sampel Daging
Sampel daging diambil dari empat ekor ayam broiler setiap perlakuan pada umur 21 dan 42 hari. Sampel daging yang digunakan berasal dari daging paha bagian atas. Sampel daging dihaluskan menggunakan meat blender sampai halus dan homogen. Sampel daging kemudian dianalisis bahan kering, lemak kasar, protein kasar dan L-karnitin. Sampel daging untuk analisis L-karnitin merupakan hasil komposit daging paha semua ulangan.
Pengambilan Sampel Empedu
Sampel empedu diambil dari empat ekor ayam broiler setiap perlakuan pada umur 21 dan 42 hari. Sampel empedu diambil dari jeroan ayam broiler yang telah dihitung berat relatif karkas dan organ. Empedu yang diperoleh kemudian diukur berat organ empedu, serta berat dan volume cairan empedu.
Prosedur Pengukuran Parameter
Data Performa
Pengukuran performa ayam dilakukan untuk mengkaji pengaruh suplementasi asam amino terhadap performa ayam broiler, khususnya konversi pakan. Hasil penelitian sebelumnya menunjukan bahwa suplementasi asam amino dapat menurunkan konversi pakan dan meningkatkan bobot badan. Konsumsi
pakan (g) dihitung dari jumlah pakan yang diberikan dikurangi dengan sisa pakan.

17
Bobot badan (g) diperoleh dari hasil penimbangan dilakukan setiap minggu. Pertambahan bobot badan harian (g) diperoleh dari hasil pengurangan berat badan akhir dikurangi dengan bobot badan awal. Konversi pakan diperoleh dari hasil pembagian konsumsi pakan dengan pertambahan bobot badan selama periode penelitian. Mortalitas (ekor perlakuan
-1) yaitu jumlah ayam yang mati
selama periode penelitian.
Data Karakteristik Empedu
Data karakteristik empedu diukur untuk mengetahui apakah suplementasi asam amino dapat meningkatkan produksi garam empedu yang berfungsi mengemulsi lemak pada pakan. Berat relatif empedu (% BB) diperoleh dari hasil pembagian berat organ empedu dengan bobot hidup ayam broiler dikalikan 100%. Berat organ empedu (g) diperoleh dari hasil penimbangan berat organ empedu. Volume (ml) dan berat (g) cairan empedu diperoleh dari pengukuran volume dan berat cairan empedu menggunakan disposable syringes. Berat jenis
cairan empedu merupakan hasil pembagian antara berat dan volume cairan empedu.
Data Koefisien Kecernaan Semu Nutrien
Koefisien kecernaan semu nutrien merupakan istilah yang digunakan untuk menghitung nutrien yang dicerna dan diserap oleh ayam broiler (pembahasan lebih dalam lihat halaman 38). Koefisien kecernaan semu nutrien dihitung untuk mengetahui apakah suplementasi glisina dan taurina dapat meningkatkan kecernaan lemak serta lisina dan metionina dapat meningkatkan metabolisme lemak. Pengukuran koefisien kecernaan semu nutrien dilakukan dengan mengukur kandungan bahan kering, lemak kasar dan protein kasar, serta energi bruto pada pakan dan ekskreta. Prosedur pengukuran kadar bahan kering, lemak kasar, dan protein kasar sesuai dengan prosedur AOAC (1995).
Kandungan energi dianalisis menggunakan bomb kalorimeter tipe Parr
6200 Calorimeter yang merupakan kalorimeter balistik. Analisis sampel energi diawali dengan menyiapkan cruicable yang telah dibersihkan dan diisi sampel seberat 0.5 g. Cruicable yang berisi sampel diletakkan pada bagian kepala bomb. Benang kalori yang telah terhubung dengan katoda dan anoda ditempelkan pada sampel. Kepala Bomb kemudian digabungkan dengan bomb bagian bawah sampai terpasang rapat. Bomb diberikan gas oksigen dengan menghubungkan pipa oksigen dengan bomb, tombol Fill pada panel kemudian ditekan dan ditunggu selama satu menit. Bomb yang telah diisi oksigen kemudian dimasukkan ditengah bucket kalorimeter. Anoda dan katoda dihubungkan dengan kepala bomb, kemudian jacket ditutup rapat. Tombol “Burn” ditekan dan ditunggu sekitar 7 menit sampai bomb dapat mengukur kandungan energi bruto sampel. Koefisien kecernaan semu nutrien (KKSN) dan Koefisien kecernaan semu energi (KKSE) dihitung menggunakan rumus :
�������� �����������������������������������������������
�����������������������������
������� ����������������������� ������������������������ �
���������������������� ���������

18
Data Profil Metabolit Serum
Pengukuran profil metabolit serum digunakan untuk mengetahui adanya hubungan antara penambahan asam amino terhadap profil metabolit serum. Profil metabolit serum berfungsi menguji apakah suplementasi asam amino dapat efisien digunakan oleh ayam broiler dengan mengukur asam urat serum. Pengukuran metabolit serum juga bertujuan untuk mengatahui pengaruh suplementasi asam amino terhadap kadar trigliserida dan kolestesterol serum ayam broiler yang diberi pakan mengandung minyak sawit 5%. Asam urat serum dianalisis sesuai prosedur analisis AKL 10101804028 Complete Test Kit menggunakan Metode Uricase-PAP. Trigliserida serum dianalisis sesuai prosedur analisis AKL 10101804027 Complete Test Kit menggunakan Metode GPO-PAP. Kolesterol serum dianalisis sesuai prosedur analisis INF 1001701 GB Complete Test Kit menggunakan Metode CHOD-PAP.
Data Berat Relatif Karkas dan Organ
Sebagian peneliti melaporkan bahwa suplementasi asam amino dapat meningkatkan berat relatif karkas dan jantung, dan menurunkan lemak abdomen. Berat relatif yang diamati pada penelitian ini meliputi berat relatif karkas, lemak abdomen, hati, pankreas dan empedu. Karkas yang dimaksud pada penelitian ini adalah adalah tubuh ayam tanpa bulu, kepala, leher, kaki dan jeroan. Rumus perhitungan menentukan karakteristik karkas adalah sebagai berikut :
�������� ���!������� ���������� ��������������� �����������
��"���"�#������������
Data Kandungan Nutrien dan L-karnitin Daging
Pengukuran kandungan nutrien daging dilakukan untuk mengetahui pengaruh suplementasi asam amino terhadap kandungan bahan kering, protein kasar, lemak kasar dan L-karnitin daging. Suplementasi asam amino diharapkan dapat menurunkan kadar lemak dan meningkatkan L-karnitin daging. Kandungan bahan kering, protein kasar dan lemak kasar dianalisis sesuai prosedur AOAC (1995). Kandungan L-karnitin daging dianalisis menggunakan metode HPLC, prosedur analisis dapat dilihat pada Lampiran 1.
Analisis Data
Data yang diperoleh dianalisis sidik ragam kecuali mortalitas. Apabila dari hasil analisis sidik ragam terdapat perbedaan yang nyata dilanjutkan dengan Uji Jarak Duncan (Steel dan Torrie 1994). Model matematis yang digunakan adalah :
Yij = � + �i + � ij Keterangan : Yij = Respon suplementasi asam amino ke-i dan ulangan ke-j. µ = Nilai tengah respon τI = Pengaruh suplementasi asam amino ke-i. εij = Galat percobaan dari suplementasi asam amino ke-i dan ulangan ke-j i = Banyaknya perlakuan (6 perlakuan). j = Banyaknya ulangan per perlakuan (4 ulangan).
19
HASIL DAN PEMBAHASAN
Pengaruh Suplementasi Asam Amino terhadap Performa
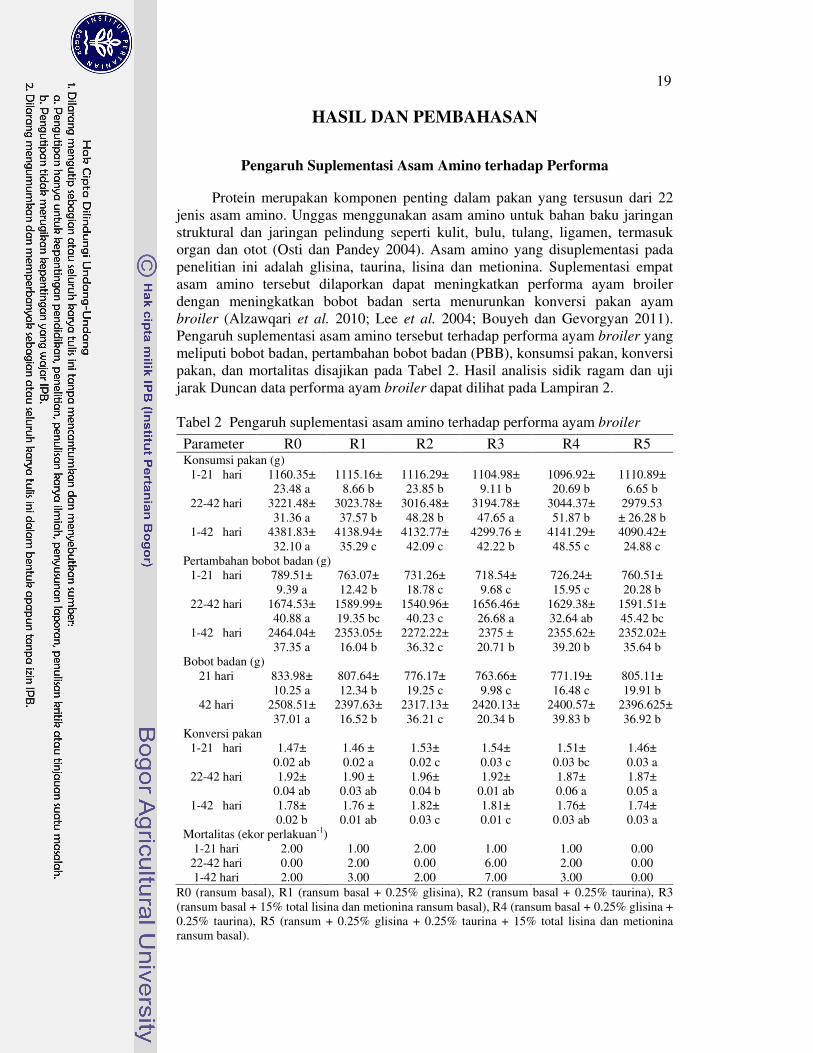
Protein merupakan komponen penting dalam pakan yang tersusun dari 22 jenis asam amino. Unggas menggunakan asam amino untuk bahan baku jaringan struktural dan jaringan pelindung seperti kulit, bulu, tulang, ligamen, termasuk organ dan otot (Osti dan Pandey 2004). Asam amino yang disuplementasi pada penelitian ini adalah glisina, taurina, lisina dan metionina. Suplementasi empat asam amino tersebut dilaporkan dapat meningkatkan performa ayam broiler dengan meningkatkan bobot badan serta menurunkan konversi pakan ayam broiler (Alzawqari et al. 2010; Lee et al. 2004; Bouyeh dan Gevorgyan 2011). Pengaruh suplementasi asam amino tersebut terhadap performa ayam broiler yang meliputi bobot badan, pertambahan bobot badan (PBB), konsumsi pakan, konversi pakan, dan mortalitas disajikan pada Tabel 2. Hasil analisis sidik ragam dan uji jarak Duncan data performa ayam broiler dapat dilihat pada Lampiran 2. Tabel 2 Pengaruh suplementasi asam amino terhadap performa ayam broiler
Parameter R0 R1 R2 R3 R4 R5 Konsumsi pakan (g)
1-21 hari 1160.35± 23.48 a
1115.16± 8.66 b
1116.29± 23.85 b
1104.98± 9.11 b
1096.92± 20.69 b
1110.89± 6.65 b
22-42 hari 3221.48± 31.36 a
3023.78± 37.57 b
3016.48± 48.28 b
3194.78± 47.65 a
3044.37± 51.87 b
2979.53 ± 26.28 b
1-42 hari 4381.83± 32.10 a
4138.94± 35.29 c
4132.77± 42.09 c
4299.76 ± 42.22 b
4141.29± 48.55 c
4090.42± 24.88 c
Pertambahan bobot badan (g) 1-21 hari 789.51±
9.39 a 763.07± 12.42 b
731.26± 18.78 c
718.54± 9.68 c
726.24± 15.95 c
760.51± 20.28 b
22-42 hari 1674.53± 40.88 a
1589.99± 19.35 bc
1540.96± 40.23 c
1656.46± 26.68 a
1629.38± 32.64 ab
1591.51± 45.42 bc
1-42 hari 2464.04± 37.35 a
2353.05± 16.04 b
2272.22± 36.32 c
2375 ± 20.71 b
2355.62± 39.20 b
2352.02± 35.64 b
Bobot badan (g) 21 hari 833.98±
10.25 a 807.64± 12.34 b
776.17± 19.25 c
763.66± 9.98 c
771.19± 16.48 c
805.11± 19.91 b
42 hari 2508.51± 37.01 a
2397.63± 16.52 b
2317.13± 36.21 c
2420.13± 20.34 b
2400.57± 39.83 b
2396.625± 36.92 b
Konversi pakan 1-21 hari 1.47±
0.02 ab 1.46 ± 0.02 a
1.53± 0.02 c
1.54± 0.03 c
1.51± 0.03 bc
1.46± 0.03 a
22-42 hari 1.92± 0.04 ab
1.90 ± 0.03 ab
1.96± 0.04 b
1.92± 0.01 ab
1.87± 0.06 a
1.87± 0.05 a
1-42 hari 1.78± 0.02 b
1.76 ± 0.01 ab
1.82± 0.03 c
1.81± 0.01 c
1.76± 0.03 ab
1.74± 0.03 a
Mortalitas (ekor perlakuan-1) 1-21 hari 2.00 1.00 2.00 1.00 1.00 0.00
22-42 hari 0.00 2.00 0.00 6.00 2.00 0.00 1-42 hari 2.00 3.00 2.00 7.00 3.00 0.00
R0 (ransum basal), R1 (ransum basal + 0.25% glisina), R2 (ransum basal + 0.25% taurina), R3 (ransum basal + 15% total lisina dan metionina ransum basal), R4 (ransum basal + 0.25% glisina + 0.25% taurina), R5 (ransum + 0.25% glisina + 0.25% taurina + 15% total lisina dan metionina ransum basal).

20
Asam Amino Esensial Asam Amino Esensial
Asam Urat Serum Meningkat Asam Urat Serum Meningkat
Konsumsi Pakan
Tabel 1 menunjukan bahwa konsumsi pakan ayam broiler umur 1-21 hari berkisar antara 1096.92 sampai 1160.35 g, kemudian berkisar dari 2979.53 sampai 3221.48 g pada umur 22-42. Secara keseluruhan, konsumsi pakan ayam broiler umur 1-42 hari yang tertinggi adalah R0 yaitu 4381.83 g dan terendah adalah R5 yaitu 4090.42 g. Suplementasi asam amino berpengaruh sangat nyata (P<0.01) terhadap konsumsi pakan ayam broiler pada semua periode pemeliharaan. Semua suplementasi asam amino nyata menurunkan konsumsi pakan ayam broiler pada periode starter dan finisher. Penurunan konsumsi pakan dapat disebabkan karena suplementasi asam amino menyebabkan kandungan asam amino pada pakan menjadi berlebihan dan tidak seimbang. Penurunan konsumsi pakan juga disebabkan karena suplementasi asam amino menyebabkan ayam broiler mengalami stres metabolis karena harus memetabolis kelebihan asam amino untuk diekskresikan dalam bentuk asam urat. Mekanisme suplementasi asam amino menyebabkan penurunan konsumsi pakan dan performa ayam broiler diilustrasikan pada Gambar 7. Pesti et al. (2005) juga menyatakan bahwa asam amino yang berlebih dan tidak seimbang akan menurunkan konsumsi pakan dan pertumbuhan. Selanjutnya, konsumsi pakan berkorelasi positif dengan bobot badan dan PBB. Oleh karena itu, penurunan konsumsi pakan menyebabkan penurunan bobot badan dan PBB ayam broiler.
Gambar 7 Mekanisme suplementasi asam amino menurunkan performa ayam broiler
Suplementasi glisina (R1) nyata menurunkan konsumsi pakan ayam broiler umur 1-21, 22-42, dan 1-42 hari. Hasil penelitian sebelumnya juga menunjukan bahwa suplementasi dan peningkatan level glisina pada pakan nyata menurunkan konsumsi pakan (Han et al. 1999; Sohail et al. 2003; Dahiya et al. 2005). Penurunan konsumsi pakan disebabkan karena peningkatan kandungan glisina menyebabkan asam amino non esensial pada pakan menjadi tidak seimbang, serta menyebabkan terjadinya enteritis nekrotik secara subklinis (Dahiya et al. 2005). Suplementasi glisina pada pakan basal cukup protein juga dianggap kurang efektif.
Asam Amino Nonesensial
Pakan Basal dengan Asam Amino Cukup dan Seimbang
Asam Amino Pakan Berlebih dan Tidak Seimbang
Suplementasi Asam Amino
Asam Amino Dideaminasi atau Transaminasi
Stres Metabolis Mortalitas Meningkat
Diekskresi dalam Bentuk Asam Urat
Suplementasi Asam Amino Tidak Efisien Digunakan
Eksresi Nitrogen Meningkat Asam Urat Serum Meningkat
Konsumsi Pakan Menurun PBB dan bobot Badan Menurun Konversi Pakan Meningkat

21
Hal ini disebabkan karena kebutuhan glisina lebih rendah pada pakan cukup protein dibandingkan pakan rendah protein (Dean et al. 2006).
Suplementasi taurina (R2) nyata menurunkan konsumsi pakan ayam broiler
pada semua periode pemeliharaan. Konsumsi pakan ayam broiler yang diberi pakan R2 paling rendah pada umur 1-42. Hasil penelitian ini sesuai dengan hasil penelitian sebelumnya yang menunjukan suplementasi taurina sedikit menurunkan konsumsi pakan (Lee et al. 2004; Zeweil et al. 2011). Suplementasi taurina menurunkan konsumsi pakan dengan menekan pelepasan GABA (�-aminobutyrate) bergantung Ca pada korteks otak tikus, sehingga menekan stimulasi pada area hipotalamus untuk makan. GABA diketahui berperan mengatur nafsu makan dengan cara menghambat kerja neurotransmitter yang mengirim sinyal kenyang ke hipotalamus (Burdakov et al. 2003).
Suplementasi lisina dan metionina (R3) nyata menurunkan konsumsi pakan ayam broiler umur 1-21 dan 1-42 hari, tetapi tidak nyata pada umur 22-42 hari. Hasil penelitian ini sesuai dengan hasil penelitian sebelumnya yang menunjukan suplementasi lisina dan metionina pada pakan menurunkan konsumsi pakan (Ojano-Dirain dan Waldroup 2002; Ukpabi et al. 2008 dan Ahmed dan Abbas 2011). Konsumsi pakan yang menurun disebabkan karena suplementasi lisina dan metionina menyebabkan asam amino pada pakan berlebih dan tidak seimbang. Kelebihan asam amino terjadi karena kebutuhan lisina pada suhu tinggi seperti pada penelitian ini (29-32 oC) lebih rendah dibandingkan ketika dipelihara di zona nyaman (Ojano-Dirain dan Waldroup 2002). Oleh karena itu, salah satu cara untuk meningkatkan konsumsi pakan pada suhu tinggi adalah dengan menurunkan rasio lisina:energi metabolis. Selanjutnya, asam amino lisina merupakan asam amino yang digunakan sebagai standar dalam konsep asam amino ideal. Suplementasi lisina tanpa mempertimbangkan konsep asam amino ideal akan merusak keseimbangan asam amino pada pakan. Onu et al. (2010) menambahkan bahwa asam amino esensial yang tidak seimbang pada pakan dapat menurunkan nilai biologi dari pakan dan menurunkan konsumsi pakan. Asam amino yang melebihi kebutuhan dan tidak seimbang akan dideaminasi dan diekskresi dalam bentuk asam urat dan amonia melalui ekskreta. Hal ini menyebabkan terjadinya polusi lingkungan dan biaya produksi yang tinggi (Osti dan Pandey 2004).
Ayam broiler yang diberi pakan dengan kombinasi suplementasi glisina dan taurina (R4) pada umur 1-42 hari sedikit lebih tinggi dibandingkan suplementasi tunggal glisina (R1) atau taurina (R2). Hal ini sesuai dengan hasil penelitian Powell et al. (2009) yang menunjukan suplementasi glisina pada pakan yang kelebihan asam amino bersulfur sedikit meningkatkan konsumsi pakan. Kombinasi suplementasi glisina dan taurina juga menimbulkan pengaruh sinergis, sehingga dapat meningkatkan konsumsi pakan. Pengaruh sinergis terjadi karena glisina berfungsi membantu ekskresi kelebihan taurina, sedangkan taurina berfungsi meningkatkan kebutuhan glisina untuk pembentukan asam urat. Suplementasi glisina, taurina, lisina dan metionina (R5) nyata menurunkan konsumsi pakan ayam broiler. Selama periode pemeliharaan, konsumsi pakan R5 paling rendah dibandingkan perlakuan yang lain. Hal ini sesuai dengan penelitian Sohail et al. (2003) yang menemukan suplementasi lisina dan metionina yang ditambah glisina 1% pada pakan defisien asam amino, konsumsi pakannya paling rendah dibandingkan perlakuan lain. Sebagai tambahan, ayam broiler umur 7-21 hari yang diberi pakan dengan kombinasi suplementasi sistina, treonina dan

22
glisina juga paling rendah bila dibandingkan yang disuplementasi salah satu asam amino tersebut (McGill et al. 2012).
Pertambahan Bobot Badan (PBB) dan Bobot Badan
Suplementasi asam amino juga berpengaruh sangat nyata (P<0.01) terhadap PBB ayam broiler umur 1-21, 22-42, dan 1-42 hari. Suplementasi asam amino menyebabkan penurunan PBB ayam broiler umur 1-21 hari dari 3.32% sampai 8.96%, tetapi pada umur 22-42 menurun berkisar 1.08 sampai 7.98%. Penurunan PBBH pada periode finisher akibat suplementasi asam amino, tidak drastis seperti pada peride starter. Hal ini menunjukan bahwa pada periode finisher, ayam broiler lebih toleran terhadap kelebihan dan ketidak seimbangan asam amino pada pakan. Pesti et al. (2005) juga menyatakan unggas memiliki kemampuan untuk beradaptasi terhadap asam amino berlebih dan tidak seimbang pada pakan. Unggas beradaptasi dengan cara mempercepat pemecahan kelebihan asam amino menjadi asam urat. Oleh karena itu, pada periode finisher ayam broiler dapat mengurangi pengaruh negatif asam amino pada pakan yang berlebih dan tidak seimbangan. Secara keseluruhan, suplementasi asam amino pada pakan ayam broiler pada umur 1-42 hari nyata menurunkan PBB sampai 7.81%. Penurunan PBB ayam broiler disebabkan karena menurunnya konsumsi pakan (lihat Gambar 7). PBB berkorelasi kuat dengan konsumsi pakan, korelasi konsumsi pakan dengan PBB pada penelitian ini adalah 0.82. Oleh karena itu, semakin tinggi konsumsi pakan maka semakin tinggi pula PBB ayam broiler. Namroud et al. (2010) juga menyatakan bahwa laju pertumbuhan sangat dipengaruhi oleh konsumsi dan komposisi pakan, serta sedikit dipengaruhi oleh aktifitas hormon tiroid. Penurunan PBB juga disebabkan karena suplementasi asam amino menyebabkan penurunan koefisien kecernaan protein kasar, khususnya ketika ayam broiler umur 21 hari (lihat Tabel 6).
Data bobot badan ayam broiler menunjukan pola yang sama dengan data PBB, dimana suplementasi asam amino menyebabkan penurunan bobot badan ayam broiler. Rataan bobot badan ayam broiler umur 21 hari berkisar 763.66 sampai 833.98 g, kemudian berkisar 2317.13 sampai 2508.51 g pada umur 42 hari. Suplementasi asam amino sangat nyata (P<0.01) berpengaruh terhadap bobot badan ayam broiler umur 21 dan 42 hari. Hasil penelitian menunjukan bahwa semua ransum yang disuplementasi asam amino nyata menurunkan bobot badan ayam broiler umur 21 dan 42 hari. Perlakuan bobot badan tertinggi pada umur 21 dan 42 hari adalah R0, sedangkan bobot badan terendah pada umur 21 dan 42 hari masing-masing adalah R3 dan R2. Penurunan bobot badan disebabkan karena menurunnya konsumsi pakan karena asam amino pakan yang berlebihan dan tidak seimbang. Beberapa literatur juga menyebutkan bahwa asam amino yang berlebih dan tidak seimbang pada pakan menyebabkan penurunan bobot badan (Baker 2003; Dean et al. 2006; Powell et al. 2009).
Suplementasi glisina nyata menurunkan PBB dan bobot badan ayam broiler umur 21 dan 42 hari. Hasil penelitian sebelumnya juga menunjukan suplementasi dan peningkatan level glisina pada pakan menurunkan PBB dan bobot badan ayam broiler (Bae et al. 1999; Sohail et al. 2003; Dahiya et al. 2005; Alzawqari et
al. 2010). Penurunan PBB dan bobot badan ayam broiler dapat disebabkan karena suplementasi glisina menyebabkan asam amino pada pakan menjadi tidak seimbang dan berlebihan. Suplementasi glisina menyebabkan perubahan rasio

23
glisina+serina : lisina, sehingga keseimbangan asam amino menjadi terganggu. Hasil penelitian terbaru merekomendasikan imbangan glisina+serina : lisina tercerna adalah 161% untuk pertumbuhan maksimal (Corzo 2012). Pakan basal starter dan finisher secara berurutan memiliki imbangan glisina+serina : lisina tercerna 170 dan 174%, sedangkan pada R1 191 dan 198%. Imbangan glisina+serina : lisina tercerna pada pakan basal lebih mendekati imbangan yang direkomendasikan Corzo (2012), sehingga PBB dan bobot badan R0 lebih tinggi dibandingkan R1. Suplementasi glisina juga menyebabkan asam amino pada pakan berlebihan, hasil penelitian Corzo et al. (2004) menemukan bahwa kebutuhan glisina pada pakan stater untuk bobot badan yang maksimal adalah 0.98%. Pakan basal starter penelitian mengandung glisina 0.93%, maka suplementasi glisina 0.25% kandungan glisina pada pakan menjadi 1.18%. Oleh karena itu, suplementasi glisina menyebabkan asam amino pada pakan menjadi tidak seimbang dan berlebihan.
Pentingnya glisina pada ayam broiler tumbuh masih menjadi perdebatan (Bae et al. 1999). Akan tetapi, beberapa hasil penelitian terbaru menunjukan bahwa sintesis glisina dalam tubuh tidak cukup untuk mendukung performa maksimum ayam broiler (Corzo et al. 2004; Dean et al. 2006; Powell et al. 2009). Glisina termasuk asam amino non esensial karena dapat disintesis dari serina dengan melepas atom �-karbon, sehingga serina dapat menggantikan glisina pada pakan. Akan tetapi, Dean dan Scot (1965) menemukan bahwa kebutuhan glisina pada pakan bebas serina adalah 1.6%. Terdapat perbedaan mengenai estimasi kebutuhan glisina pada ayam broiler. Hal ini disebabkan karena adanya perbedaan sumber protein dan konsentrasi asam amino lain yang terlibat dalam metabolisme glisina seperti serina dan treonina (Dean et al. 2006). Oleh karena itu, suplementasi glisina pada pakan yang cukup mengandung serina sedikit direspon pada unggas. Sebagai tambahan, respon suplementasi glisina juga dipengaruhi oleh kandungan protein kasar, fortifikasi asam amino, bahan baku pakan, serta umur dan jenis unggas (Dean et al. 2006; McGill et al. 2012).
PBB dan bobot badan ayam broiler umur 21 dan 42 hari nyata menurun dengan suplementasi taurina 0.25%. Bahkan, bobot badan ayam broiler umur 42 hari yang diberi pakan dengan suplementasi taurina paling rendah dibandingkan perlakuan lain. Hasil yang sama dilaporkan oleh Lee et al. (2004) dimana suplementasi taurina berjenjang dari 0.1 sampai 0.4% menurunkan bobot badan, meskipun tidak berbeda nyata. Hasil penelitian Tufft dan Jensen (1992) juga menunjukan bahwa suplementasi taurina berpengaruh kecil terhadap performa ayam broiler dan kalkun. Lee et al. (2004) menyatakan bahwa respon suplementasi taurina terhadap PBB dan bobot badan dipengaruhi oleh kandungan asam amino bersulfur, khususnya metionina. Suplementasi taurina memberikan respon positif pada performa pertumbuhan ketika diberikan pada pakan dengan level asam amino bersulfur yang rendah. Kandungan asam amino bersulfur pada pakan basal periode starter dan finisher sudah sesuai dengan rekomendasi Leeson
dan Summer (2005) yaitu 0.96 dan 0.89%. Selanjutnya, suplementasi taurina juga dapat menghemat kebutuhan asam amino bersulfur pada ayam broiler, sehingga suplementasi taurina menyebabkan kelebihan asam amino bersulfur pada pakan.
Penurunan PBB dan bobot badan dengan suplementasi taurina juga dapat disebabkan karena adanya penurunan konsumsi pakan yang nyata. Hasil penelitian pada tikus juga menunjukan terjadi penurunan PBB dan bobot badan

24
dengan suplementasi taurina yang disebabkan karena konsumsi pakan yang menurun (Yamamoto et al. 2000; Park et al. 2007). Penurunan PBB dan bobot badan juga menunjukan taurina yang disuplementasikan tidak dapat di metabolisme dengan baik. Hal ini ditunjukan dengan meningkatnya kadar asam urat pada serum ayam broiler umur 21 dan 42 hari (lihat Tabel 3). Hasil penelitian Powell et al. (2009) juga menunjukan bahwa terjadi peningkatan asam urat pada serum ayam broiler yang kelebihan asam amino bersulfur. Powell et al. (2009) juga merekomendasikan rasio total asam amino bersulfur (TSAA) : lisina tidak melebihi 0.72. Rasio TSAA : lisina pada pakan basal penelitian adalah 0.69, sedangkan pada R2 dengan asumsi taurina dihitung sebagai TSAA, maka rasionya menjadi 0.88. Rasio TSAA : lisina yang tidak ideal menyebabkan terjadinya penurunan PBB dan bobot badan ayam broiler.
Suplementasi taurina sintetik banyak digunakan pada pakan hewan karnivora seperti kucing, anjing dan ikan karnivora. Pada unggas, tidak ada penelitian yang menunjukan bahwa suplementasi taurina memberikan pengaruh menguntungkan terhadap performa (EFSA 2012). Kebutuhan taurina pada ternak sebagian dipenuhi dari makanan dan sebagian yang lain berasal dari biosintesis dari metionina dan sisteina (Tsuboyama et al. 1996). Taurina disintesis dari metionina dan sisteina melalui serangkaian reaksi enzimatis. Enzim sisteina sulfinat dekarboksilase merupakan enzim yang berperan dalam biosintesis taurina. Akan tetapi, jumlah enzim tersebut terbatas, sehingga menghambat proses biosintesis taurina pada unggas dan mamalia (Jacobsen dan Smith 1968). Sturman
dan Hayes (1980) juga melaporkan aktivitas enzim sisteina sulfinate karboksilase lebih rendah pada otak dan hati fetus manusia, monyet, kelinci, tikus dan kucing dibandingkan pada saat dewasa. Akan tetapi, taurina memiliki peran penting dalam pertumbuhan, khususnya pada periode starter (Zeweil et al. 2011). Hasil penelitian Zeng et al. (2009) menunjukan bahwa peningkatan taurina 0.1% pada pakan dapat meningkatkan rataan PBB puyuh pada periode starter, tetapi tidak berbeda nyata pada periode finisher.
Suplementasi lisina dan metionina nyata menurunkan PBB ayam broiler umur 1-21 dan 1-42 hari, tetapi tidak berbeda nyata pada umur 22-42 hari. Hasil penelitian ini menunjukan pola yang sama dengan hasil penelitian Bouyeh dan
Gevorgyan (2011) yang menunjukan bahwa suplementasi lisina dan metionina 30% dari standar NRC menurunkan PBB ayam broiler umur 21, tetapi meningkatkan PBB pada umur 42 hari. Fakta ini membuktikan bahwa ayam broiler dapat mengkompensasi pertubuhannya pada umur 22 sampai 42 hari. Kidd et al. (1998) juga menemukan bahwa kandungan lisina pada pakan periode starter berhubungan dengan kandungan lisina periode finisher. Ayam broiler yang defisien lisina pada periode starter dapat mengkompensasi sebagian pertumbuhannya pada periode grower-finisher ketika diberi pakan mengandung lisina 125% dari standar NRC (1994). Onu et al. (2010) menambahkan bahwa suplementasi lisina dan metionina sintetik secara individu maupun kombinasinya dapat meningkatkan performa ayam broiler ketika diberikan pada pakan yang kekurangan lisina dan metionina.
Suplementasi lisina dan metionina juga nyata menurunkan bobot badan ayam broiler umur 21 dan 42 hari. Banyak penelitian sebelumnya melaporkan bahwa peningkatan level lisina dan metionina dari standar NRC (1994) dapat meningkatkan bobot badan ayam broiler (Café dan Waldroup 2006; Onu et al.

25
2010; Bouyeh dan Gevorgyan 2011). Hasil penelitian ini bertentangan dengan hasil penelitian sebelumnya, hal ini disebabkan karena pada penelitian ini tidak memperhitungkan rasio lisina : TSAA dan tidak menambahkan 0.1% NaHCO3 yang digunakan sebagai buffer dari kelebihan klorida dari penambahan L-lisina HCl seperti yang dilakukan oleh Bouyeh dan Gevorgyan (2011). Perbedaan performa dengan hasil penelitian sebelumnya juga dapat disebabkan karena perbedaan strain broiler, pakan basal, dan lingkungan selama periode penelitian. Akan tetapi, hasil penelitian ini sesuai dengan penelitian yang dilakukan oleh Abudabos dan Aljumaah (2010) yang menunjukan bahwa peningkatan level lisina 30% dari rekomendasi NRC (1994) memberikan pengaruh negatif terhadap bobot badan ayam broiler.
Penurunan PBB dan bobot badan dapat disebabkan asam amino pada pakan menjadi tidak seimbang dengan suplementasi lisina dan metionina. Asam amino menjadi tidak seimbang karena hanya mensuplementasi lisina dan metionina, tanpa mempertimbangkan keseimbangan asam amino yang lain. Saat ini dikenal adanya konsep asam amino ideal yang merupakan salah satu konsep yang digunakan untuk memenuhi kebutuhan asam amino ayam broiler yang kompleks (Baker 2003). Konsep ideal asam amino mengasumsikan bahwa meskipun kebutuhan asam amino berubah pada berbagai situasi, rasio antar asam amino akan tetap stabil. Lisina biasanya digunakan sebagai standar dalam asam amino ideal, sedangkan level optimal asam amino lainnya diperoleh menggunakan rasio optimal masing-masing asam amino (Vieira et al. 2004). Hasil penelitian sebelumnya menunjukan bahwa aplikasi konsep asam amino ideal berpengaruh besar terhadap performa ayam broiler (Vieira et al. 2004; Siqueira et al. 2013).
Suplementasi lisina dan metionina juga menyebabkan kelebihan lisina dan metionina. Banyak penelitian telah dilakukan untuk mengetahui kebutuhan lisina dan metionina, tetapi hasilnya bervariasi karena adanya perbedaan genetik, nutrisi dan manajemen ayam broiler. Bernal et al. (2012) merekomendasikan kebutuhan lisina tercerna untuk memaksimalkan performa dari ayam broiler COBB 500 pada periode starter untuk jantan 1.22% dan betina 1.24%, serta 1.16% untuk jantan dan betina pada periode finisher. Suplementasi lisina dan metionina pada pakan mengakibatkan kandungan lisinanya berlebih karena mengandung lisina tercerna 1.37% pada periode starter dan 1.19% pada finisher. Leclercq (1998) menambahkan bahwa kebutuhan lisina untuk memaksimalkan pertambahan bobot badan lebih rendah bila dibandingkan dengan kebutuhan lisina untuk meningkatkan daging dada, serta menurunkan konversi pakan dan lemak abdomen. Selanjutnya, Goulart et al. (2011) merekomendasikan metionina+sisteina tercerna pada pakan ayam periode stater dan finisher adalah 0.75 dan 0.61%. Kandungan metionina+siteina tercerna R0 pada periode starter dan finisher adalah 0.79 dan 0.70%, sedangkan pakan R3 0.88 dan 0.78%. Hal ini menunjukan suplementasi lisina dan metionina juga menyebabkan kelebihan metionina. Hasil penelitian Adeyemo (2012) juga menunjukan suplementasi metionina pada pakan tinggi protein menyebabkan penurunan bobot badan. Hal ini disebabkan karena kelebihan metionina dapat menekan pertumbuhan ayam broiler.
Kombinasi suplementasi glisina dan taurina nyata menurunkan PBB dan bobot badan ayam broiler umur 21 dan 42 hari dibandingkan kontrol. PBB dan bobot badan ayam broiler umur 1-42 hari yang diberi pakan R4 nyata meningkat dibandingkan R2, tetapi tidak nyata berbeda dengan R1. Kombinasi suplementasi

26
glisina dan taurina dapat sedikit meningkatkan PBB dan bobot badan ayam broiler umur 42 hari dibandingkan suplementasi tunggal glisina atau taurina. Hasil penelitian ini sesuai dengan Powell et al. (2009) yang menunjukan suplementasi glisina dapat meningkatkan PBB ayam broiler yang kelebihan asam amino bersulfur. Kombinasi suplementasi glisina, taurina, lisina dan metionina (R5) juga nyata menurunkan PBB dan bobot badan ayam broiler umur 21 dan 42 hari dibandingkan kontrol. PBB dan bobot badan ayam broiler umur 42 hari yang diberi pakan R5 tidak berbeda nyata dengan R1, R3, dan R4, tetapi nyata lebih tinggi dibandingkan R2. Hasil penelitian ini mendukung penelitian Sohail et al. (2003) yang menunjukan penambahan glisina dengan metionina dan lisina meningkatkan pertumbuhan. Hasil penelitian kedua McGill et al. (2012) juga menunjukan bahwa kombinasi suplementasi glisina, sistina, dan treonina sedikit meningkatkan PBB bila dibandingkan dengan suplementasi tunggal sistina, treonina dan glisina.
Konversi Pakan
Hasil penelitian menunjukan bahwa suplementasi asam amino pada ayam broiler umur 1-21 hari sangat nyata (P<0.01) berpengaruh terhadap konversi pakan. Konversi pakan pada perlakuan R2, R3, dan R4 nyata lebih tinggi dibandingkan kontrol. Pada periode finisher, suplementasi asam amino hanya berpengaruh nyata (P<0.05) dan hanya perlakuan R2 yang nyata meningkatkan konversi pakan. Konversi pakan yang membaik pada periode finisher dapat disebabkan karena unggas telah mampu mengurangi pengaruh negatif dari kelebihan asam amino pada pakan. Suplementasi asam amino pada pakan ayam broiler umur 1-42 sangat nyata (P<0.01) berpengaruh terhadap konversi pakan. Konversi pakan ayam broiler umur 1-42 hari perlakuan R5 lebih rendah, R1 dan R4 sama, serta R2 dan R3 lebih tinggi dari pada kontrol. Meningkatnya konversi pakan dapat disebabkan karena suplementasi asam amino pada pakan cukup protein menyebabkan penggunaan protein dan asam amino menjadi tidak efisien (lihah Gambar 7). Hal ini ditandai dengan meningkatnya kadar asam urat pada serum (lihat Tabel 3). Hasil penelitian Siqueira et al. (2013) juga menunjukan bahwa peningkatan konsentrasi asam amino menyebabkan efisiensi penggunaannya menurun. Hal ini disebabkan asam amino lebih banyak digunakan untuk proses metabolis alternatif yang tidak berhubungan dengan deposit protein. Penggunaan protein pakan lebih efisien pada pakan rendah protein dan seimbang dibandingkan dengan pakan tinggi protein tetapi tidak seimbang (Huyghebaert dan Park 1996). Selanjutnya, hasil penelitian Vieira et al. (2004) menunjukan unggas yang diberi pakan dengan kandungan protein 26.0% membutuhkan 0.45 kg protein per kg PBB, tetapi unggas yang diberi pakan dengan kandungan protein 20.5% hanya membutuhkan 0.35 kg protein per kg PBB.
Suplementasi glisina tidak berpengaruh nyata terhadap konversi pakan pada semua periode starter dan finisher. Hasil penelitian Alzawqari et al. (2010) juga menunjukan bahwa suplementasi glisina berpengaruh tidak nyata terhadap konversi pakan. Bae et al. (1999) dan Corzo et al. (2004) juga menemukan bahwa peningkatan level glisina pada pakan tidak berpengaruh terhadap konversi pakan ayam broiler umur 21 hari. Suplementasi glisina tidak berpengaruh nyata terhadap konversi pakan disebabkan karena kandungan protein kasar pada pakan basal pada fase stater dan finisher sudah mencukupi yaitu 22% dan 20%. Suplementasi

27
glisina secara nyata dapat menurunkan konversi pakan jika diberikan pada pakan dengan kandungan protein rendah (Dean et al. 2006). Selanjutnya, Corzo (2012) merekomendasikan bahwa rasio (glisina+serina : lisina tercerna) yang direkomendasikan untuk memaksimalkan konversi pakan adalah 184.6%. Penelitian pada manusia menunjukan bahwa glisina berperan penting dalam pelepasan hormon petumbuhan yang berperan meningkatkan efisiensi sintesis protein (Eklund et al. 2005; Dean et al. 2006). Glisina juga berperan sebagai prekursor untuk sintesis sistina yang dapat meningkatkan penggunaan asam amino lainnya untuk sintesis protein daging (Powell et al. 2009).
Suplementasi taurina pada penelitian ini nyata meningkatkan konversi pakan ayam broiler selama periode pemeliharaan (1-42 hari). Konversi pakan dengan suplementasi taurina paling tinggi dibandingkan dengan perlakuan lainnya. Hasil penelitian ini bertentangan dengan hasil penelitian Lee et al. (2004) dan Zeweil et al. (2011) yang menunjukan bahwa suplementasi taurina dapat menurunkan konversi pakan. Akan tetapi, hasil penelitian ini sesuai dengan penelitian Ruiz-Feria et al. (1999) yang menemukan suplementasi taurina 5% pada tikus nyata menurunkan efisiensi pakan dari 0.37 menjadi hanya 0.31. Suplementasi taurina tidak konsisten dapat meningkatkan performa ayam broiler. Hal ini mungkin disebakan karena respon suplementasi taurin berhubungan dengan kandungan metionina pada pakan. Taurina tidak efisien disuplementasikan pada pakan tinggi metionina (Lee et al. 2004).
Suplementasi lisina dan metionina (R3) nyata meningkatkan konversi pakan ayam broiler umur 1-21 dan 1-42 hari. Konversi pakan ayam broiler umur 22-42 hari yang disuplementasi lisina dan metionina memiliki nilai yang sama dengan kontrol. Hasil penelitian Khalifeh-Gholi dan Jahanian (2012) juga menunjukan bahwa peningkatan level metionina dari 100 menjadi 110% standar NRC pada pakan yang mengandung cukup arginina meningkatkan konversi pakan dari 1.72 menjadi 1.75. Konversi pakan yang meningkat disebabkan karena terjadi kelebihan asam amino. Aftab et al. (2007) menemukan bahwa kebutuhan lisina pada daerah tropis lebih rendah dibandingkan dengan kebutuhan lisina pada zona thermonetral. Kebutuhan kandungan lisina total pada pakan untuk konversi pakan maksimal di daerah tropis Pakistan adalah 0.97% (setara 0.84% lisina tercerna). Pada kondisi yang berlebihan, asam amino tidak digunakan untuk pertumbuhan tetapi akan didegradasi dan diekskresi sebagai nitrogen atau ditransformasi menjadi asam amino non esensial (Vieira et al. 2004). Penurunan konversi pakan juga dapat disebabkan karena teknik formulasi suplementasi kurang efektif dilakukan. Siqueira et al. (2013) menemukan bahwa unggas yang diberi pakan dengan teknik formulasi “dilution” lebih efisien menggunakan lisina (75.57%) dibandingkan pakan dengan teknik formulasi “supplementation” (73.22%). Formulasi pakan dengan teknik “supplementation” menyebabkan rasio asam amino pada pakan menjadi bervariasi, sedangkan teknik “dilution” menjaga rasio asam amino tetap konstan.
Ayam broiler umur 1-42 hari yang diberi pakan R4 memiliki konversi pakan sedikit lebih rendah dibandingkan R0, walaupun secara statistik tidak nyata. Selanjutnya, ayam broiler umur 1-42 hari yang diberi pakan R4 memiliki konversi pakan lebih rendah dibandingkan R1 dan R2. Hal ini menunjukan bahwa ayam broiler yang diberi pakan R4, dapat menggunakan pakan lebih efisien dibandingkan R1 dan R2. Peningkatan efisiensi pakan dapat disebabkan karena

28
meningkatnya efisiensi penggunaan asam amino yang ditunjukan dengan penurunan kadar asam urat serum R4 dibandingkan R1 dan R2. Hasil penelitian Powell et al. (2009) juga menunjukan bahwa pakan dengan lisina 1.35% dan rasio TSAA:lisina 0.76 yang disuplementasi glisina memiliki efisiensi pakan yang lebih baik dibandingkan tanpa suplementasi glisina. Kombinasi suplementasi glisina, taurina, lisina dan metionina nyata menurunkan konversi pakan pada ayam broiler umur 1-42 hari dan memiliki konversi pakan paling rendah dibandingkan perlakuan lain. Hasil penelitian Sohail et al. (2003) juga menunjukan bahwa kombinasi suplementasi lisina, metionina, dan glisina 0.05% pada pakan rendah protein konversi pakannya paling rendah dibandingkan perlakuan lainnya ketika ayam broiler berumur enam minggu. Selanjutnya, kombinasi suplementasi sistina, treonina, dan glisina pada pakan rendah protein yang dilakukan pada penelitian kedua McGill et al. (2012) juga sedikit menurunkan konversi pakan dari 1.34 menjadi 1.32. Menurunnya konversi pakan dengan kombinasi suplementasi glisina, taurina, lisina dan metionina dapat disebabkan karena meningkatnya keseimbangan asam amino esensial dan non esensial.
Mortalitas
Mortalitas ayam broiler umur 1-42 hari yang diberi pakan dengan suplementasi glisina berjumlah 3 ekor. Hasil penelitian sebelumnya menunjukan bahwa suplementasi glisina atau treonina tidak berpengaruh nyata terhadap mortalitas (Waldroup et al. 2005). Akan tetapi, penelitian Dahiya et al. (2005) menemukan mortalitas ayam broiler yang disuplementasi glisina mencapai 8.33%. Peningkatan mortalitas disebabkan karena ayam broiler terkena enteritis nekrotik dan stres panas. Selanjutnya, mortalitas ayam broiler umur 1-42 hari yang diberi pakan dengan suplementasi taurina berjumlah 2 ekor. Hasil penelitian sebelumnya menunjukan suplementasi taurina 0.25% tidak berpengaruh pada mortalitas (Lee et al. 2004). Taurina banyak diteliti berkaitan dengan upaya untuk menurunkan kematian karena sudden death syndrome (SDS), karena taurina berperan dalam mengatur kerja jantung (Ruiz-Feria et al. 1999). Akan tetapi, suplementasi taurina hanya sedikit menurunkan kejadian SDS (Jacob 1989).
Hasil penelitian menunjukan bahwa suplementasi lisina dan metionina tanpa mengikuti konsep asam amino ideal dapat meningkatkan mortalias ayam broiler. Mortalitas ayam broiler perlakuan R3 berjumlah 7 ekor sampai akhir pemeliharaan. Mortalitas R3 paling tinggi dibandingkan perlakuan lain, karena terjadi peningkatan mortalitas pada ayam broiler umur 22-42 hari yang mencapai 6 ekor. Hasil penelitian yang dilakukan Café dan Waldroup (2006) dan Bouyeh
dan Gevorgyan (2011) menunjukan bahwa level lisina dan metionina tidak berpengaruh terhadap mortalitas. Akan tetapi, hasil penelitian Kidd et al. (1998) dan Corzo (2012) menunjukan bahwa peningkatan level lisina menyebabkan peningkatan mortalitas ayam broiler pada periode starter dan grower. Peningkatan mortalitas pada starter disebabkan karena penyakit pada kaki, sedangkan pada finisher disebabkan karena asites dan suddent death syndrome.
Mortalitas ayam broiler sampai akhir penelitian yang diberi pakan dengan kombinasi suplementasi glisina dan taurina berjumlah 3 ekor. Mortalitas ayam broiler yang diberi pakan R4 mirip dengan ayam yang diberi pakan R1 dan R2. Hal ini menunjukan bahwa kelebihan asam amino glisina dan taurina tidak berpengaruh terhadap mortalitas. Kelebihan glisina dan taurina hanya berdampak
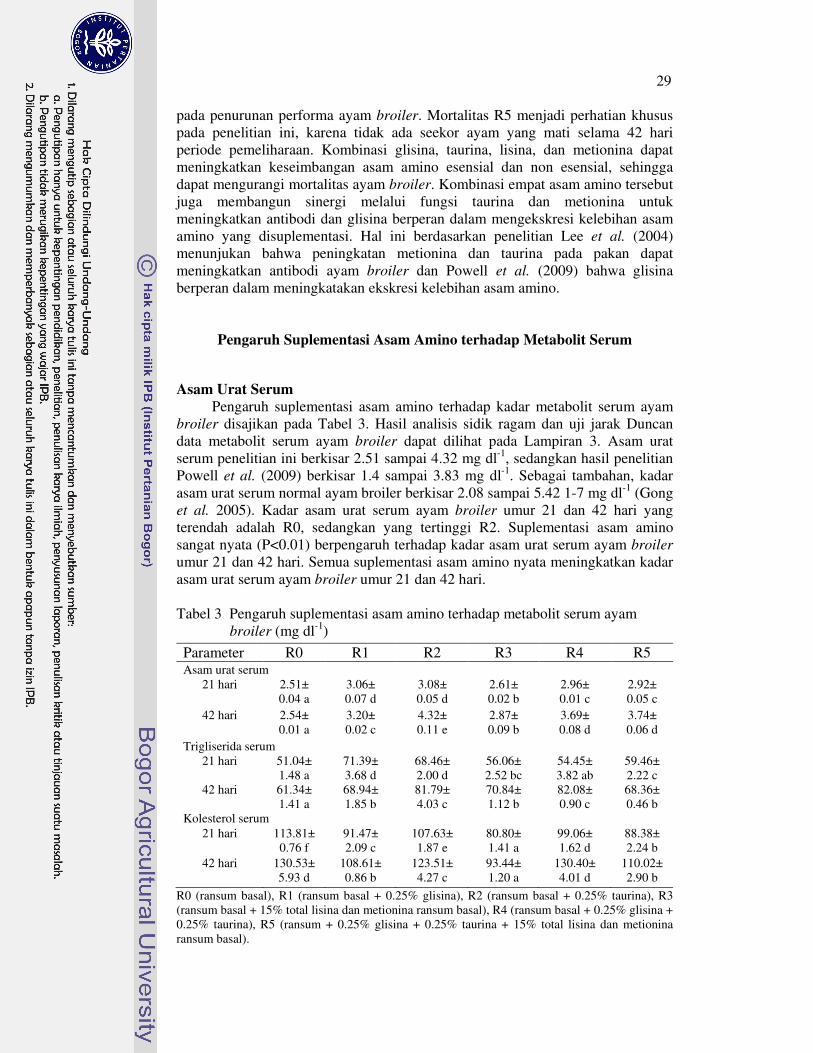
29
pada penurunan performa ayam broiler. Mortalitas R5 menjadi perhatian khusus pada penelitian ini, karena tidak ada seekor ayam yang mati selama 42 hari periode pemeliharaan. Kombinasi glisina, taurina, lisina, dan metionina dapat meningkatkan keseimbangan asam amino esensial dan non esensial, sehingga dapat mengurangi mortalitas ayam broiler. Kombinasi empat asam amino tersebut juga membangun sinergi melalui fungsi taurina dan metionina untuk meningkatkan antibodi dan glisina berperan dalam mengekskresi kelebihan asam amino yang disuplementasi. Hal ini berdasarkan penelitian Lee et al. (2004) menunjukan bahwa peningkatan metionina dan taurina pada pakan dapat meningkatkan antibodi ayam broiler dan Powell et al. (2009) bahwa glisina berperan dalam meningkatakan ekskresi kelebihan asam amino.
Pengaruh Suplementasi Asam Amino terhadap Metabolit Serum
Asam Urat Serum
Pengaruh suplementasi asam amino terhadap kadar metabolit serum ayam broiler disajikan pada Tabel 3. Hasil analisis sidik ragam dan uji jarak Duncan data metabolit serum ayam broiler dapat dilihat pada Lampiran 3. Asam urat serum penelitian ini berkisar 2.51 sampai 4.32 mg dl-1, sedangkan hasil penelitian Powell et al. (2009) berkisar 1.4 sampai 3.83 mg dl-1. Sebagai tambahan, kadar asam urat serum normal ayam broiler berkisar 2.08 sampai 5.42 1-7 mg dl-1 (Gong et al. 2005). Kadar asam urat serum ayam broiler umur 21 dan 42 hari yang terendah adalah R0, sedangkan yang tertinggi R2. Suplementasi asam amino sangat nyata (P<0.01) berpengaruh terhadap kadar asam urat serum ayam broiler umur 21 dan 42 hari. Semua suplementasi asam amino nyata meningkatkan kadar asam urat serum ayam broiler umur 21 dan 42 hari.
Tabel 3 Pengaruh suplementasi asam amino terhadap metabolit serum ayam
broiler (mg dl-1)
Parameter R0 R1 R2 R3 R4 R5 Asam urat serum
21 hari 2.51± 0.04 a
3.06± 0.07 d
3.08± 0.05 d
2.61± 0.02 b
2.96± 0.01 c
2.92± 0.05 c
42 hari 2.54± 0.01 a
3.20± 0.02 c
4.32± 0.11 e
2.87± 0.09 b
3.69± 0.08 d
3.74± 0.06 d
Trigliserida serum 21 hari 51.04±
1.48 a 71.39± 3.68 d
68.46± 2.00 d
56.06± 2.52 bc
54.45± 3.82 ab
59.46± 2.22 c
42 hari 61.34± 1.41 a
68.94± 1.85 b
81.79± 4.03 c
70.84± 1.12 b
82.08± 0.90 c
68.36± 0.46 b
Kolesterol serum 21 hari 113.81±
0.76 f 91.47± 2.09 c
107.63± 1.87 e
80.80± 1.41 a
99.06± 1.62 d
88.38± 2.24 b
42 hari 130.53± 5.93 d
108.61± 0.86 b
123.51± 4.27 c
93.44± 1.20 a
130.40± 4.01 d
110.02± 2.90 b
R0 (ransum basal), R1 (ransum basal + 0.25% glisina), R2 (ransum basal + 0.25% taurina), R3 (ransum basal + 15% total lisina dan metionina ransum basal), R4 (ransum basal + 0.25% glisina + 0.25% taurina), R5 (ransum + 0.25% glisina + 0.25% taurina + 15% total lisina dan metionina ransum basal).

30
Asam urat serum menggambarkan kemampuan unggas dalam menggunakan protein dan asam amino pada pakan. Kadar asam urat akan meningkat apabila protein atau asam amino pada pakan berlebihan atau tidak seimbang. Peningkatan kadar asam urat bertujuan untuk mengekskresikan kelebihan protein melalui ekskreta. Kelebihan asam amino akan didegradasi dan membebaskan amonia yang kemudian disintesis menjadi asam urat (Bouyeh dan Gevorgyan 2011). Sintesis asam urat menggunakan jalur metabolis yang sama dengan sintesis adenin dan guanin. Atom nitrogen yang digunakan dalam biosintesis berasal dari glisin, glutamine dan aspatat, sedangkan atom karbonnya berasal dari CO2 dan tetrahidrofolat (Larbier dan Leclerq 1994; Klasing 2006). Proses metabolisme kelebihan asam amino pada serum disajikan pada Gambar 8. Asam urat hasil biosintesis kemudian dikirim ke ginjal untuk diekskresikan (Gong et al. 2005). Asam urat diekskresikan sekitar 60 sampai 80% nitrogen urin pada unggas (Larbier dan Leclerq 1994).
Gambar 8 Proses metabolisme kelebihan asam amino pada serum Kombinasi glisina dan taurina dapat menurunkan asam urat serum
dibandingkan yang diberikan taurina secara individu. Hal ini menunjukan kombinasi suplementasi glisina dan taurina lebih menguntungkan dibandingkan suplementasi glisina dan taurina secara individu. Selanjutnya, kadar asam urat serum yang diberi pakan R5 secara umum lebih tinggi dibandingkan yang diberi pakan R1, R3, dan R4, tetapi lebih rendah dibandingkan R2. Penurunan kadar asam urat serum dengan kombinasi glisina dapat disebabkan karena glisina berfungsi membantu metabolisme kelebihan TSAA dengan meningkatkan penggunaan metionine untuk membentuk sistina atau dengan meningkatkan eksresi kelebihan TSAA sebagai asam urat. Hasil penelitian Powell et al. (2009) menunjukan bahwa peningkatan rasio TSAA : lisina pada ayam broiler umur 21 hari meningkatkan kadar asam urat serum, tetapi suplementasi glisina dapat menurunkan kadar asam urat serum. Hal ini berkaitan dengan peran glisina sebagai komponen asam urat yang penting untuk mengekskresikan kelebihan protein pada unggas (Corzo et al. 2004; Powell et al. 2009).
Asam Amino Tidak Seimbang dan Berlebih
Deaminasi
Kerangka Karbon Amonia (NH3) Asam Amino Nonesensial
NH3
Glikogen
Asam Urat Urea Glukogenik Ketogenik
Asetil KoA Asam Urat Serum
Meningkat
Piruvat Asetoasetat
Kolesterol
Trigliserida Glukosa
Glukosa darah
Meningkat
Kolesterol
Serum Menurun
Trigliserida Serum
Meningkat
Transaminasi

31
Trigliserida Serum
Suplementasi asam amino sangat nyata (P<0.01) berpengaruh terhadap kadar trigliserida serum ayam broiler umur 21 dan 42 hari. Semua suplementasi asam amino nyata meningkatkan kadar trigliserida serum ayam broiler umur 21 dan 42 hari. Kadar trigliserida serum ayam broiler umur 21 berkisar 51.04 sampai 71.39 mg dl-1, sedangkan umur 42 hari berkisar dari 61.34 sampai 82.08 mg dl-1. Kadar trigliserida serum yang tertinggi pada umur 21 dan 42 hari masing-masing adalah R1 dan R4. Rataan kadar trigliserida serum ayam broiler umur 21 hari yaitu 60.14 mg/dl, lebih rendah dibandingkan pada umur 42 (72.23 mg dl-1). Hasil penelitian ini sesuai dengan penelitian Gong et al. (2005) yang menunjukan kadar trigliserida serum ayam broiler yang diberi pakan dengan rasio treonina:lisina yang berbeda pada umur 42 hari lebih tinggi dari umur 21 hari. Hasil penelitian Gong et al. (2005) juga menunjukan bahwa kadar trigliserida serum normal pada ayam broiler berkisar 19.33 sampai 26.50 mg dl-1. Kadar trigliserida serum penelitian ini lebih tinggi dibandingkan trigliserida serum normal dikarenakan terjadi kelebihan asam amino.
Suplementasi glisina dan taurina nyata meningkatkan kadar trigliserida serum ayam broiler umur 21 dan 42 hari. Hasil penelitian Alzawqari et al. (2010) juga menunjukan bahwa suplementasi 0.25% glisina meningkatkan kadar trigliserida serum ayam broiler umur 21 dan 42 hari, walaupun tidak nyata. Hasil penelitian pada tikus juga menunjukan suplementasi glisina meningkatkan kadar trigliserida darah dan hati, meskipun secara statistik tidak berbeda nyata. Peningkatan trigliserida serum dapat disebabkan karena penurunan enzim oksidasi asam lemak dan peningkatan enzim lipogenik pada darah sehingga meningkatkan jumlah asam lemak bebas (Senthilkumar et al. 2003). Berikutnya, hasil penelitian Shim et al. (2009) menemukan bahwa injeksi taurina nyata meningkarkan kadar trigliserida serum, tetapi nyata menurunkan trigliserida hati. Hal ini menunjukan bahwa taurina menstimulasi sekresi trigliserida dari hati ke darah pada ayam broiler dan tikus. Hasil penelitian Lee et al. (2004) juga menemukan bahwa ada kecenderungan suplementasi taurina pada pakan cukup metionina menyebabkan peningkatan trigliserida. Oleh karena itu, kadar trigliserida tidak hanya dipengaruhi oleh suplementasi taurina, tetapi juga level metionina pada pakan.
Suplementasi lisina dan metionina nyata meningkatkan kadar trigliserida serum ayam broiler umur 21 dan 42 hari. Hasil penelitian ini didukung hasil penelitian Khalifeh-Gholi dan Jahanian (2012) yang menunjukan bahwa peningkatan level metionina dari 90% menjadi 110% dari standar NRC (1999) sedikit meningkatkan kadar trigliserida serum pada ayam broiler umur 21 dan 42 hari. Hasil penelitian Sachan et al. (1984) juga menunjukan suplementasi prekursor karnitin (lisina dan metionina) gagal menurunkan trigliserida serum, tetapi suplementasi karnitin nyata menurunkan kadar trigliserida plasma. Hal ini dapat disebabkan karena asam amino yang berlebih dapat menurunkan produksi karnitin dalam tubuh dan mengganggu pengaruh hipolipidemik dan lipotropik dari karnitin. Sachan et al. (1984) menemukan bahwa suplementasi lisina dan metionina pada pakan yang mengandung etanol nyata menurunkan L-karnitin total dan L-karnitin bebas plasma darah tikus. Peningkatan kadar trigliserida serum juga dapat disebabkan karena kelebihan asam amino lisina dan metionina akan dioksidasi. Proses tersebut menghasilkan asetil KoA yang kemudian disintesis menjadi trigliserida. Oleh karena itu, trigliserida serum berkorelasi positif dengan

32
asam urat serum. Peningkatan konsentrasi asam urat berarti akan menyediakan kerangka karbon yang lebih banyak untuk sintesis trigliserida. Koefisien korelasi kadar trigliserida dengan asam urat serum ayam broiler umur 21 dan 42 hari adalah 0.79. Ros (2000) menjelaskan bahwa proses biosintesis trigliserida dimulai dengan pengaktifan asam lemak menjadi asetil KoA, kemudian asetil KoA diestersifikasi untuk dikonversi menjadi trigliserida (lihat Gambar 8).
Kombinasi suplementasi glisina dan taurina nyata meningkatkan trigliserida serum ayam broiler umur 21 dan 42 hari dibandingkan kontrol. Kombinasi suplementasi glisina dan taurina menurunkan trigliserida serum umur 21 hari dibandingkan yang diberikan secara individu, tetapi trigliserida serum meningkat pada umur 42 hari. Hal ini menunjukan umur ayam broiler berpengaruh terhadap respon suplementasi asam amino terhadap kadar trigliserida serum. Hasil penelitian Khalifeh-Gholi dan Jahanian (2012) juga menunjukan bahwa peningkatan level metionina menjadi 110% dari standar NRC (1994) menurunkan kadar trigliserida serum ayam broiler umur 21 hari, tetapi cenderung meningkat pada umur 42 hari. Selanjutnya, hasil penelitian Tappia et al. (2012) menunjukan bahwa suplementasi taurina secara individu nyata meningkatkan trigliserida plasma darah tikus, tetapi kombinasi taurina dengan sisteina cenderung menurunkan trigliserida plasma.
Kolesterol Serum
Suplementasi asam amino sangat nyata (P<0.01) berpengaruh terhadap kadar kolesterol darah. Semua suplementasi asam amino nyata menurunkan kadar kolesterol darah ayam broiler umur 21 dan 42 hari. Kadar kolesterol serum ayam broiler umur 42 hari lebih tinggi dibandingkan umur 21 hari. Rataan kadar kolesterol pada umur 42 hari adalah 116.08 mg dl-1, sedangkan pada umur 21 hari hanya 96.83 mg dl-1. Hasil penelitian ini sesuai dengan Khalifeh-Gholi dan
Jahanian (2012) yang menunjukan bahwa kolesterol ayam broiler umur 42 hari yang diberi pakan dengan metionina berjenjang lebih tinggi dibandingkan umur 21 hari. Hasil penelitiannya menunjukan bahwa rataaan kolesterol darah pada umur 42 hari adalah 120.38 mg dl-1, sedangkan pada umur 21 hari 110.88 mg dl-1. Kadar kolesterol darah ayam broiler umur 21 hari turun 29.00%, sedangkan pada umur 42 hari kolesterol darah turun 28.41%.
Suplementasi glisina dan taurina nyata menurunkan kadar kolesterol serum ayam broiler umur 21 dan 42 hari. Hasil penelitian sebelumnya juga menunjukan suplementasi glisina pada pakan yang ditambahkan kolesterol (Sugiyama et al 1989) dan histidina 3.5% (Ohmura et al. 1988) nyata menurunkan kolesterol darah. Suplementasi glisina dapat menurunkan kolesterol serum dikarenakan kolesterol pada serum digunakan dalam biosintesis asam empedu. Ohmura et al. (1988) melaporkan bahwa suplementasi glisina meningkatkan asam empedu yang berkonjugasi dengan glisina, serta menstimulasi ekskresi asam empedu. Oleh karena itu, suplementasi glisina menurunkan level kolesterol serum (lihat Gambar 9). Cheeke dan Dierenfeld (2010) menjelaskan bahwa sebagian besar kolesterol diekskresikan dalam bentuk asam empedu. Selanjutnya, hasil penelitian Lee et al. (2004) dan Zeweil et al. (2011) menunjukan bahwa suplementasi taurina nyata menurunkan kolesterol serum ayam broiler. Suplementasi taurina pada tikus yang diberikan pakan tinggi kolesterol juga menunjukan penurunan kadar kolesterol serum (Park et al. 2007; Shim et al. 2009). Kadar kolesterol menurun disebabkan

33
karena taurina bersifat hypocholesterolemic (Park et al. 2007). Aksi hypocholesterolemic taurina dengan membatasi aktivitas cholesterol 7 �-
hydroxylase, menekan HMG-CoA reduktase, reabsorbsi asam empedu pada ilium dan mungkin sebagai reseptor LDL. Suplementasi glisina dan taurina juga akan meningkatkan konjugasi asam kolik yang secara tidak langsung mengurangi ketersediaan asam kolik bebas untuk aktifitas kolesterol esterase yang berperan membantu penyerapan kolesterol. Oleh karena itu suplementasi glisin, taurin dan kombinasi keduanya dapat menurunkan kadar kolesterol serum.
Suplementasi lisina dan metionina nyata menurunkan kadar kolesterol serum ayam broiler umur 21 dan 42 hari. Hasil penelitian Fillios et al. (1954) menunjukan bahwa suplementasi sisteina dan metionina pada tikus yang diberi pakan tinggi kolesterol nyata menurunkan hypercholesteremia. Lee et al. (2004) menambahkan bahwa suplementasi metionina akan meningkatkan kadar asam amino bebas, khususnya metionina dan taurina pada serum. Peningkatan taurina pada serum bersifat hypocholesterolemic sehingga menurunkan kolesterol serum pada tikus (Park et al. 2007). Suplementasi glisina dan taurina (R4) nyata menurunkan kadar kolesterol serum ayam broiler umur 21 hari, tetapi tidak berbeda nyata pada umur 42 hari. Kadar kolesterol serum ayam broiler umur 21 dan 42 yang diberi pakan R5 nyata lebih rendah dibandingkan R0. Selanjutnya, Bouyeh dan Gevorgyan (2011) menjelaskan bahwa kolesterol pada darah sangat dinamis dibandingkan kolesterol pada jaringan seperti daging dada dan paha.
Penurunan kadar kolesterol juga dapat disebabkan karena terbatasnya suplai asetil KoA sebagai bahan baku proses biosintesis kolesterol. Kolesterol dapat disintesis dari konversi 2 atom karbon (C2) bagian asetil KoA menjadi menjadi sebuah prekursor isoprenoid asam mevalonat (C6). Enam asam mevalonat kemudian dikonversi menjadi skualan (C30), selanjutnya skualan ditransformasi menjadi kolesterol (C27) (Mathews 2000). Hasil biosintesis kolesterol didistribusikan keseluruh tubuh untuk pembentukan membran sel, ikut serta dalam proses transportasi asam lemak darah, penyusun garam empedu dan sebagai prekursor vitamin D dan hormon steroid (Prosser 1991). Terbatasnya suplai asetil KoA untuk biosintesis kolesterol disebabkan karena asetil KoA juga digunakan dalam proses biosintesis asam lemak dan glukosa. Oleh karena itu, terjadi kompetisi penggunaan asetil KoA untuk pembentukan asam lemak, glukosa dan kolesterol. Hasil penelitian Bouyeh dan Gevorgyan (2011) menunjukan bahwa asetil KoA hasil pemecahan kelebihan asam amino lisina dan metionina mengalami proses glukoneogenesis di sitosol, sehingga nyata meningkatkan kadar glukosa dan menurunkan kolesterol plasma (lihat Gambar 8).
Pengaruh Suplementasi Asam Amino terhadap Karakteristik Empedu
Berat Relatif Kantung Empedu
Pengaruh suplementasi asam amino terhadap karakteristik empedu disajikan pada Tabel 4. Hasil analisis sidik ragam dan uji jarak Duncan data karakteristik empedu ayam broiler dapat dilihat pada Lampiran 4. Berat relatif kantung empedu ayam broiler umur 21 hari berkisar 0.13 sampai 0.23%, sedangkan pada umur 42 hari berkisar 0.14 sampai 0.19%. Berat relatif kantung empedu hasil penelitian ini,
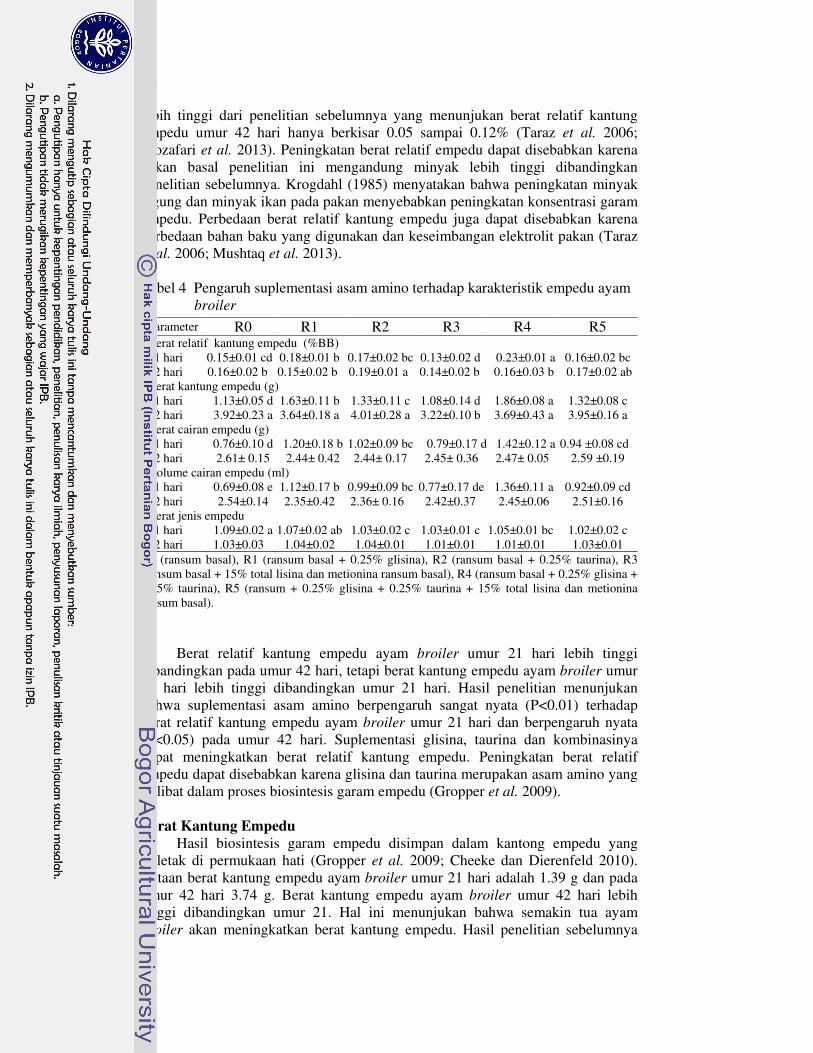
34
lebih tinggi dari penelitian sebelumnya yang menunjukan berat relatif kantung empedu umur 42 hari hanya berkisar 0.05 sampai 0.12% (Taraz et al. 2006; Mozafari et al. 2013). Peningkatan berat relatif empedu dapat disebabkan karena pakan basal penelitian ini mengandung minyak lebih tinggi dibandingkan penelitian sebelumnya. Krogdahl (1985) menyatakan bahwa peningkatan minyak jagung dan minyak ikan pada pakan menyebabkan peningkatan konsentrasi garam empedu. Perbedaan berat relatif kantung empedu juga dapat disebabkan karena perbedaan bahan baku yang digunakan dan keseimbangan elektrolit pakan (Taraz et al. 2006; Mushtaq et al. 2013).
Tabel 4 Pengaruh suplementasi asam amino terhadap karakteristik empedu ayam
broiler Parameter R0 R1 R2 R3 R4 R5 Berat relatif kantung empedu (%BB) 21 hari 0.15±0.01 cd 0.18±0.01 b 0.17±0.02 bc 0.13±0.02 d 0.23±0.01 a 0.16±0.02 bc 42 hari 0.16±0.02 b 0.15±0.02 b 0.19±0.01 a 0.14±0.02 b 0.16±0.03 b 0.17±0.02 ab Berat kantung empedu (g) 21 hari 1.13±0.05 d 1.63±0.11 b 1.33±0.11 c 1.08±0.14 d 1.86±0.08 a 1.32±0.08 c 42 hari 3.92±0.23 a 3.64±0.18 a 4.01±0.28 a 3.22±0.10 b 3.69±0.43 a 3.95±0.16 a Berat cairan empedu (g) 21 hari 0.76±0.10 d 1.20±0.18 b 1.02±0.09 bc 0.79±0.17 d 1.42±0.12 a 0.94 ±0.08 cd 42 hari 2.61± 0.15 2.44± 0.42 2.44± 0.17 2.45± 0.36 2.47± 0.05 2.59 ±0.19 Volume cairan empedu (ml) 21 hari 0.69±0.08 e 1.12±0.17 b 0.99±0.09 bc 0.77±0.17 de 1.36±0.11 a 0.92±0.09 cd 42 hari 2.54±0.14 2.35±0.42 2.36± 0.16 2.42±0.37 2.45±0.06 2.51±0.16 Berat jenis empedu 21 hari 1.09±0.02 a 1.07±0.02 ab 1.03±0.02 c 1.03±0.01 c 1.05±0.01 bc 1.02±0.02 c 42 hari 1.03±0.03 1.04±0.02 1.04±0.01 1.01±0.01 1.01±0.01 1.03±0.01
R0 (ransum basal), R1 (ransum basal + 0.25% glisina), R2 (ransum basal + 0.25% taurina), R3 (ransum basal + 15% total lisina dan metionina ransum basal), R4 (ransum basal + 0.25% glisina + 0.25% taurina), R5 (ransum + 0.25% glisina + 0.25% taurina + 15% total lisina dan metionina ransum basal).
Berat relatif kantung empedu ayam broiler umur 21 hari lebih tinggi
dibandingkan pada umur 42 hari, tetapi berat kantung empedu ayam broiler umur 42 hari lebih tinggi dibandingkan umur 21 hari. Hasil penelitian menunjukan bahwa suplementasi asam amino berpengaruh sangat nyata (P<0.01) terhadap berat relatif kantung empedu ayam broiler umur 21 hari dan berpengaruh nyata (P<0.05) pada umur 42 hari. Suplementasi glisina, taurina dan kombinasinya dapat meningkatkan berat relatif kantung empedu. Peningkatan berat relatif empedu dapat disebabkan karena glisina dan taurina merupakan asam amino yang terlibat dalam proses biosintesis garam empedu (Gropper et al. 2009).
Berat Kantung Empedu
Hasil biosintesis garam empedu disimpan dalam kantong empedu yang terletak di permukaan hati (Gropper et al. 2009; Cheeke dan Dierenfeld 2010). Rataan berat kantung empedu ayam broiler umur 21 hari adalah 1.39 g dan pada umur 42 hari 3.74 g. Berat kantung empedu ayam broiler umur 42 hari lebih tinggi dibandingkan umur 21. Hal ini menunjukan bahwa semakin tua ayam broiler akan meningkatkan berat kantung empedu. Hasil penelitian sebelumnya

35
menunjukan bahwa berat kantung empedu berkisar 1.24 sampai 2.22 g (Taraz et
al. 2006). Suplementasi asam amino sangat nyata (P<0.01) meningkatkan berat kantung empedu ayam broiler umur 21 dan 42 hari. Hasil penelitian ini sesuai dengan pernyataan Krogdahl (1985) bahwa beberapa komponen nonlemak seperti asam amino pada pakan berpengaruh terhadap sintesis dan sekresi garam empedu. Komponen tersebut banyak mendapat perhatian, karena peningkatan sintesis dan ekskresi garam empedu nyata menurunkan kadar kolesterol darah.
Hasil penelitian sebelumnya menunjukan bahwa perbedaan sumber protein pakan dapat menyebabkan perbedaan berat kantung empedu. Penggantian bungkil kedelai dengan bungkil rapeseed nyata meningkatkan berat kantung empedu. Hal ini disebabkan sphincter oddi mengalami hipothiroidism yang menyebabkan lambatnya pengosongan kantung empedu (Taraz et al. 2006). Perubahan komposisi pakan juga akan merubah mikroflora saluran pencernaan, sehingga menyebabkan terjadinya konversi garam empedu primer menjadi garam empedu sekunder (Krogdahl 1985). Asam empedu sekunder terbentuk dari proses metabolisme mikroba pada saluran pencernaan bagian belakang. Pada nutrisi manusia, asam empedu sekuder menjadi perhatian karena bersifat karsinogen, dan dapat memicu berkembangnya kanker kolon. Sebagian besar asam empedu yang disekresikan kantong empedu akan direabsorbsi melalui saluran pencernaan bagian bawah dan dihilangkan dari darah oleh kerja hati. Hal ini dikenal sebagai siklus entero-hepatik empedu. Siklus ini berkontribusi untuk menjaga metabolisme kolesterol tetap normal. Kolesterol serum dapat turun oleh senyawa pakan yang dapat mengikat dan mengekskresikan kolesterol. Hal ini termasuk obat penurun kolesterol (misalnya cholestyramine) dan senyawa alami (misalnya saponin) (Gropper et al. 2009). Berikutnya, hasil penelitian Kussaibati dan
Leclercq (1984) menunjukan bahwa suplementasi 0.5 dan 1.0% sorbitol dan 0.5% garam empedu dapat meningkatkan berat kantung empedu. Hal ini disebabkan karena sorbitol berperan dalam pemekatan dan sekresi garam empedu, sedangkan suplementasi garam empedu dapat meningkatkan akumulasi garam empedu pada kantung empedu.
Berat dan Volume Cairan Empedu
Cairan empedu berwarna kehijauan, sedikit asam (pH 6), mengandung garam empedu dan lemak (phospolipid dan kolesterol) (Larbier dan Leclerq 1994). Pada manusia, kantung empedu mampu menyimpan sekitar 40-50 ml cairan empedu (Gropper et al. 2009). Hasil penelitian menunjukan bahwa rataan berat cairan empedu ayam broiler umur 21 hari adalah 1.02 g, sedangkan pada umur 42 hari 2.50 g. Berat cairan empedu berhubungan erat dengan volume cairan empedu, sehingga berat dan volume cairan empedu menunjukan pola yang sama. Volume cairan empedu ayam broiler umur 21 hari berkisar 0.69 sampai 1.36 ml, sedangkan pada umur 42 hari berkisar 2.25 sampai 2.54 ml. Volume cairan empedu umur 21 hari lebih rendah dibandingkan umur 42 hari, tetapi volume cairan empedu umur 21 hari lebih sensitif terhadap suplementasi asam amino dibandingkan umur 42 hari. Hal ini menunjukan bahwa produksi cairan empedu ayam broiler umur 21 hari masih terbatas, maka suplementasi asam amino dapat menstimulasi sintesis cairan empedu. AKan tetapi, pada umur 42 ayam broiler telah mampu memproduksi cairan empedu dalam jumlah yang cukup. Oleh karena itu, ayam broiler umur 21 hari lebih sulit memanfaatkan lemak pakan

36
dibandingkan pada umur 42 hari. Hasil penelitian ini sesuai dengan hasil penelitian oleh Pullen dan Polin (1984) yang menunjukan bahwa suplementasi tepung empedu ayam dan asam kolat pada pakan ayam broiler jantan umur delapan minggu tidak berpengaruh nyata pada sekresi cairan empedu.
Suplementasi asam amino berpengaruh sangat nyata (P<0.01) terhadap volume cairan empedu ayam broiler umur 21 hari, tetapi tidak berpengaruh nyata (P>0.05) pada umur 42 hari. Hasil penelitian ini sulit dibandingkan dengan hasil penelitian sebelumnya, karena sangat sedikit peneliti yang melaporkan data berat dan volume cairan empedu pada kantung empedu. Akan tetapi, hasil penelitian Pullen dan Polin (1984) dengan memasang cannulate pada pembuluh empedu menunjukan bahwa rataan harian volume empedu yang disekresikan ayam broiler jantan umur 8 minggu yang diberi pakan 0.08% tepung empedu adalah 21 ml dengan kandungan bahan kering 20%. Sebagai tambahan, Marzooqi dan Leeson (1998) menyatakan bahwa volume sekresi empedu tergantung pada laju sirkulasi darah, status nutrisi unggas, tipe pakan yang diberikan dan efisiensi sirkulasi enterohepatik garam empedu. Laju aliran empedu juga dipengaruhi oleh konsentrasi protein dan asam amino, level lemak, kualitas lemak, sterol dan level lesitin pada pakan (Krogdahl 1985).
Berat Jenis (BJ)
Kantong empedu merupakan tempat pemekatan dan penyimpanan empedu yang disintesis oleh hati sampai empedu dibutuhkan untuk mencerna lemak di usus halus. Selama periode pencernaan, empedu dikirim dari hati ke kantung empedu untuk dikonsentrasikan dan disimpan. Cairan empedu yang telah dikonsentrasikan mengandung air lebih dari 90%, sedangkan sisanya merupakan elektrolit yang direabsorbsi oleh mukosa kantung empedu. Sodium merupakan kation utama pada empedu, akan tetapi potassium dan kalsium juga ditemukan pada cairan empedu. Proses pengkonsentrasian empedu memungkinkan kantung empedu untuk menyimpan empedu yang diproduksi oleh hati sampai dibutuhkan dalam proses pencernaan makanan (Gropper et al. 2009; Cheeke dan Dierenfeld 2010). Salah satu indikator untuk dapat menilai proses pemekatan garam empedu adalah BJ cairan empedu.
Rataan berat jenis empedu ayam broiler umur 21 adalah 1.05 kemudian pada umur 42 hari 1.03. BJ cairan empedu umur 21 hari lebih tinggi dibandingkan umur 42 hari, hal ini disebabkan karena pada umur 21 hari asam empedu lebih banyak berkonjugasi dengan glisina. Berat jenis glisina lebih tinggi dibandingkan taurina (1.595 vs 1.494), sehingga BJ cairan empedu umur 21 lebih tinggi dari umur 42 hari. Suplementasi asam amino berpengaruh sangat nyata (P<0.01) terhadap BJ empedu ayam broiler umur 21 hari, tetapi tidak berpengaruh nyata (P>0.05) pada umur 42 hari. Suplementasi asam amino nyata menurunkan BJ empedu ayam broiler umur 21 hari. Hal ini dapat disebabkan karena suplementasi asam amino meningkatkan volume cairan empedu, sehingga penurunan konsentrasi cairan empedu. Akan tetapi, berat jenis empedu tidak berpengaruh terhadap efektifitas garam empedu dalam mencerna lemak. Hal ini ditunjukan dengan meningkatnya koefisien kecernaan semu lemak kasar bersamaan dengan meningkatnya volume cairan empedu, meskipun berat jenis cairan empedu menurun. Oleh karena itu, proses pencernaan lemak lebih dipengaruhi oleh volume cairan empedu dibandingkan berat jenis cairan empedu.
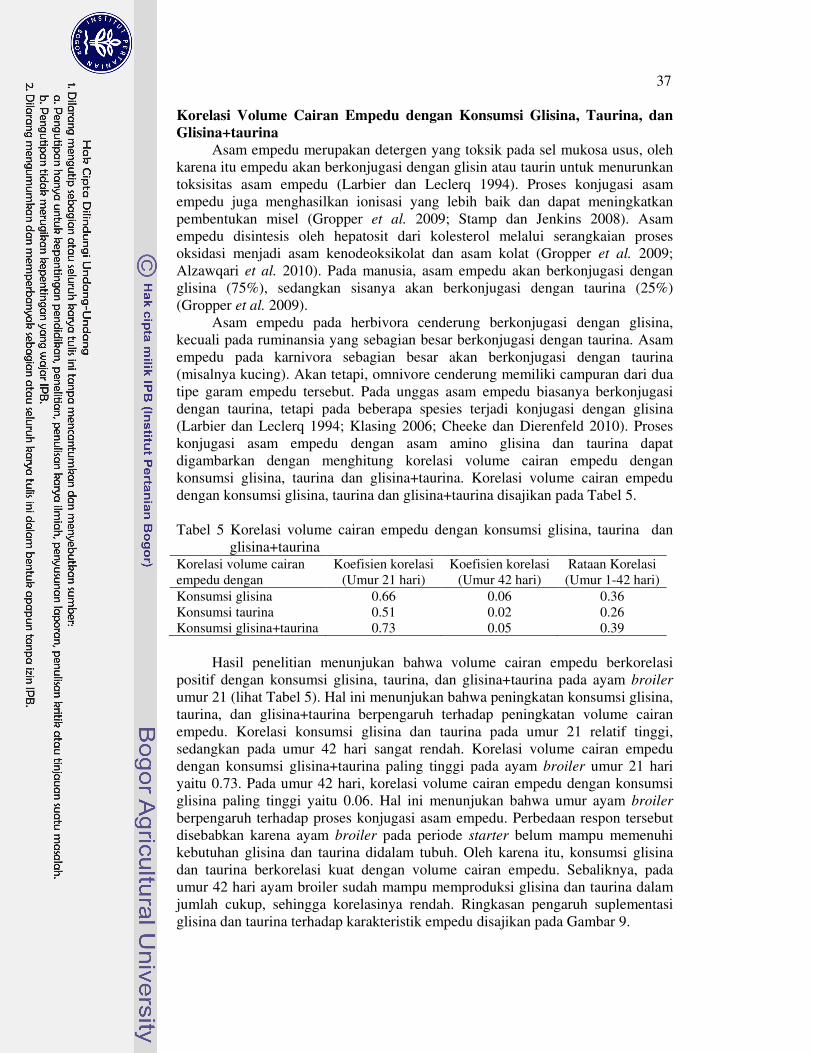
37
Korelasi Volume Cairan Empedu dengan Konsumsi Glisina, Taurina, dan
Glisina+taurina
Asam empedu merupakan detergen yang toksik pada sel mukosa usus, oleh karena itu empedu akan berkonjugasi dengan glisin atau taurin untuk menurunkan toksisitas asam empedu (Larbier dan Leclerq 1994). Proses konjugasi asam empedu juga menghasilkan ionisasi yang lebih baik dan dapat meningkatkan pembentukan misel (Gropper et al. 2009; Stamp dan Jenkins 2008). Asam empedu disintesis oleh hepatosit dari kolesterol melalui serangkaian proses oksidasi menjadi asam kenodeoksikolat dan asam kolat (Gropper et al. 2009; Alzawqari et al. 2010). Pada manusia, asam empedu akan berkonjugasi dengan glisina (75%), sedangkan sisanya akan berkonjugasi dengan taurina (25%) (Gropper et al. 2009).
Asam empedu pada herbivora cenderung berkonjugasi dengan glisina, kecuali pada ruminansia yang sebagian besar berkonjugasi dengan taurina. Asam empedu pada karnivora sebagian besar akan berkonjugasi dengan taurina (misalnya kucing). Akan tetapi, omnivore cenderung memiliki campuran dari dua tipe garam empedu tersebut. Pada unggas asam empedu biasanya berkonjugasi dengan taurina, tetapi pada beberapa spesies terjadi konjugasi dengan glisina (Larbier dan Leclerq 1994; Klasing 2006; Cheeke dan Dierenfeld 2010). Proses konjugasi asam empedu dengan asam amino glisina dan taurina dapat digambarkan dengan menghitung korelasi volume cairan empedu dengan konsumsi glisina, taurina dan glisina+taurina. Korelasi volume cairan empedu dengan konsumsi glisina, taurina dan glisina+taurina disajikan pada Tabel 5.
Tabel 5 Korelasi volume cairan empedu dengan konsumsi glisina, taurina dan
glisina+taurina Korelasi volume cairan empedu dengan
Koefisien korelasi (Umur 21 hari)
Koefisien korelasi (Umur 42 hari)
Rataan Korelasi (Umur 1-42 hari)
Konsumsi glisina 0.66 0.06 0.36 Konsumsi taurina 0.51 0.02 0.26 Konsumsi glisina+taurina 0.73 0.05 0.39
Hasil penelitian menunjukan bahwa volume cairan empedu berkorelasi
positif dengan konsumsi glisina, taurina, dan glisina+taurina pada ayam broiler umur 21 (lihat Tabel 5). Hal ini menunjukan bahwa peningkatan konsumsi glisina, taurina, dan glisina+taurina berpengaruh terhadap peningkatan volume cairan empedu. Korelasi konsumsi glisina dan taurina pada umur 21 relatif tinggi, sedangkan pada umur 42 hari sangat rendah. Korelasi volume cairan empedu dengan konsumsi glisina+taurina paling tinggi pada ayam broiler umur 21 hari yaitu 0.73. Pada umur 42 hari, korelasi volume cairan empedu dengan konsumsi glisina paling tinggi yaitu 0.06. Hal ini menunjukan bahwa umur ayam broiler berpengaruh terhadap proses konjugasi asam empedu. Perbedaan respon tersebut disebabkan karena ayam broiler pada periode starter belum mampu memenuhi kebutuhan glisina dan taurina didalam tubuh. Oleh karena itu, konsumsi glisina dan taurina berkorelasi kuat dengan volume cairan empedu. Sebaliknya, pada umur 42 hari ayam broiler sudah mampu memproduksi glisina dan taurina dalam jumlah cukup, sehingga korelasinya rendah. Ringkasan pengaruh suplementasi glisina dan taurina terhadap karakteristik empedu disajikan pada Gambar 9.

38
Gambar 9 Ringkasan pengaruh suplementasi glisina dan taurina terhadap karakteristik empedu
Asam empedu lebih banyak berkonjugasi dengan taurina pada kondisi asam,
karena asam empedu terkonjugasi dengan taurina lebih mudah larut dibandingkan yang terkonjugasi dengan glisina (Cheeke dan Dierenfeld 2010). Akan tetapi, proses konjugasi juga dipengaruhi oleh ketersediaan glisina dan taurina pada pakan. Hewan herbivora lebih banyak berkonjugasi dengan glisina karena pada tumbuhan banyak ditemukan glisina. Selanjutnya, hewan karnivora banyak berkonjugasi dengan taurina karena taurina banyak ditemukan pada daging. Oleh karena itu, proses konjugasi asam empedu dengan glisina atau taurina juga dipengaruhi oleh ketersediaan glisina dan taurina pada pakan. Hasil penelitian menunjukan bahwa asam empedu lebih berkorelasi positif dengan glisina, hal ini disebabkan karena pakan lebih banyak mengandung glisina dibandingkan taurina.
Pengaruh Suplementasi Asam Amino terhadap Koefisien Kecernaan Semu
Pakan yang dikonsumsi ayam broiler akan dicerna di saluran pencernaan menjadi nutrien yang siap diserap seperti asam amino, asam lemak dan glukosa. Istilah “kecernaan” biasa digunakan menyatakan jumlah nutrien yang dicerna dan diserap di saluran pencernaan (Kussaibati dan Lerlercq 1984; Smits et al. 1998; Leeson dan Summer 2001; Adrizal et al. 2002; Gong et al. 2005). Akan tetapi, sebagian peneliti menggunakan istilah “absorbsi” (Leeson dan Summer 2001; Matin et al. 2013), serta sebagian lainnya menggunakan istilah “retensi” (Pullen dan Polin 1984; Adeyemo et al. 2013). Penelitian ini menggunakan istilah “kecernaan” karena lebih banyak peneliti yang menggunakan istilah ini dibandingkan istilah lainnya. Hal ini menunjang penulis untuk dapat mencari litaratur menjadi lebih komperhensif. Kecernaan dapat diketahui dengan menghitung konsumsi nutrien dan nutrien pada ekskreta. Data konsumsi nutrien dan nutrien ekskreta digunakan untuk menghitung kecernaan (lihat halaman 17). Kecernaan nutrien dapat dihitung menggunakan metode penciri inert dan koleksi total. Metode pengukuran kecernaan yang digunakan pada penelitian ini menggunakan metode koleksi total dengan cara mengukur konsumsi nutrien dan jumlah nutrien pada ekskreta.
Istilah “kecernaan” dianggap kurang relevan digunakan karena pada unggas sangat sulit memisahkan antara feses dan urin karena keduanya dieksresikan dalam ekskreta melalui kloaka. Oleh karena itu, penulis menambahkan istilah “semu” karena ada sebagian nutrien, khususnya protein yang berasal dari urin.
Volume Cairan empedu Meningkat
Kolesterol Serum Menurun
Kolesterol
Asam Empedu
Garam Empedu Terkonjugasi Taurina
Garam Empedu Terkonjugasi Glisina
Korelasi 0.36
Korelasi 0.26
Glisina Taurina
Sekresi Empedu Meningkat
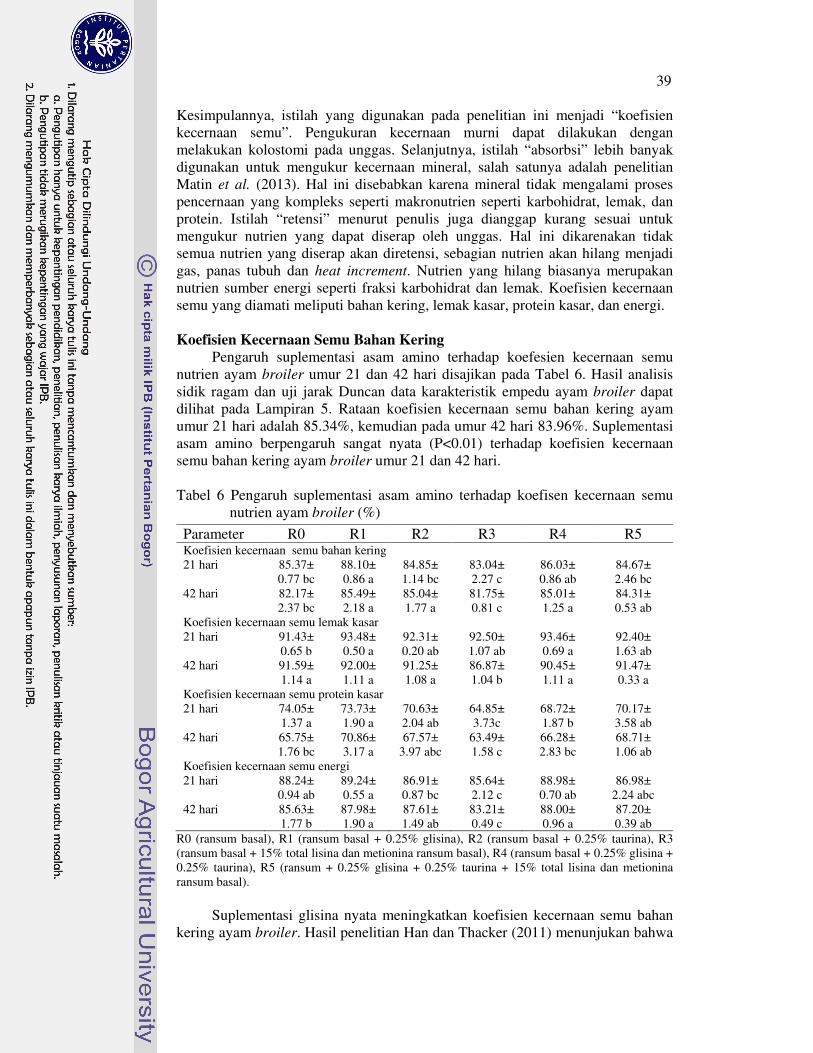
39
Kesimpulannya, istilah yang digunakan pada penelitian ini menjadi “koefisien kecernaan semu”. Pengukuran kecernaan murni dapat dilakukan dengan melakukan kolostomi pada unggas. Selanjutnya, istilah “absorbsi” lebih banyak digunakan untuk mengukur kecernaan mineral, salah satunya adalah penelitian Matin et al. (2013). Hal ini disebabkan karena mineral tidak mengalami proses pencernaan yang kompleks seperti makronutrien seperti karbohidrat, lemak, dan protein. Istilah “retensi” menurut penulis juga dianggap kurang sesuai untuk mengukur nutrien yang dapat diserap oleh unggas. Hal ini dikarenakan tidak semua nutrien yang diserap akan diretensi, sebagian nutrien akan hilang menjadi gas, panas tubuh dan heat increment. Nutrien yang hilang biasanya merupakan nutrien sumber energi seperti fraksi karbohidrat dan lemak. Koefisien kecernaan semu yang diamati meliputi bahan kering, lemak kasar, protein kasar, dan energi.
Koefisien Kecernaan Semu Bahan Kering
Pengaruh suplementasi asam amino terhadap koefesien kecernaan semu nutrien ayam broiler umur 21 dan 42 hari disajikan pada Tabel 6. Hasil analisis sidik ragam dan uji jarak Duncan data karakteristik empedu ayam broiler dapat dilihat pada Lampiran 5. Rataan koefisien kecernaan semu bahan kering ayam umur 21 hari adalah 85.34%, kemudian pada umur 42 hari 83.96%. Suplementasi asam amino berpengaruh sangat nyata (P<0.01) terhadap koefisien kecernaan semu bahan kering ayam broiler umur 21 dan 42 hari.
Tabel 6 Pengaruh suplementasi asam amino terhadap koefisen kecernaan semu
nutrien ayam broiler (%)
Parameter R0 R1 R2 R3 R4 R5 Koefisien kecernaan semu bahan kering 21 hari 85.37±
0.77 bc 88.10± 0.86 a
84.85± 1.14 bc
83.04± 2.27 c
86.03± 0.86 ab
84.67± 2.46 bc
42 hari 82.17± 2.37 bc
85.49± 2.18 a
85.04± 1.77 a
81.75± 0.81 c
85.01± 1.25 a
84.31± 0.53 ab
Koefisien kecernaan semu lemak kasar 21 hari 91.43±
0.65 b 93.48± 0.50 a
92.31± 0.20 ab
92.50± 1.07 ab
93.46± 0.69 a
92.40± 1.63 ab
42 hari 91.59± 1.14 a
92.00± 1.11 a
91.25± 1.08 a
86.87± 1.04 b
90.45± 1.11 a
91.47± 0.33 a
Koefisien kecernaan semu protein kasar 21 hari 74.05±
1.37 a 73.73± 1.90 a
70.63± 2.04 ab
64.85± 3.73c
68.72± 1.87 b
70.17± 3.58 ab
42 hari 65.75± 1.76 bc
70.86± 3.17 a
67.57± 3.97 abc
63.49± 1.58 c
66.28± 2.83 bc
68.71± 1.06 ab
Koefisien kecernaan semu energi 21 hari 88.24±
0.94 ab 89.24± 0.55 a
86.91± 0.87 bc
85.64± 2.12 c
88.98± 0.70 ab
86.98± 2.24 abc
42 hari 85.63± 1.77 b
87.98± 1.90 a
87.61± 1.49 ab
83.21± 0.49 c
88.00± 0.96 a
87.20± 0.39 ab
R0 (ransum basal), R1 (ransum basal + 0.25% glisina), R2 (ransum basal + 0.25% taurina), R3 (ransum basal + 15% total lisina dan metionina ransum basal), R4 (ransum basal + 0.25% glisina + 0.25% taurina), R5 (ransum + 0.25% glisina + 0.25% taurina + 15% total lisina dan metionina ransum basal).
Suplementasi glisina nyata meningkatkan koefisien kecernaan semu bahan
kering ayam broiler. Hasil penelitian Han dan Thacker (2011) menunjukan bahwa

40
suplementasi glisina 0.1% pada pakan ayam petelur meningkatkan kecernaan ileum bahan kering, walaupun tidak nyata. Hal ini dapat disebabkan karena glisina dapat berpengaruh langsung maupun tidak langsung terhadap peningkatan fungsi dari mukosa usus (Ospina-Rojas et al. 2013). Suplementasi taurina tidak berpengaruh nyata terhadap koefisien kecernaan semu bahan kering ayam broiler umur 21, tetapi nyata meningkat pada ayam broiler umur 42 hari. Hasil penelitian ini sesuai dengan hasil penelitian Zeweil et al. (2011) yang menunujukan bahwa suplementasi taurina dapat meningkatkan koefisien kecernaan bahan kering ayam broiler umur 42 hari. Peningkatan kecernaan nutrien disebabkan karena taurina memiliki peran penting dalam metabolisme lemak dan meningkatkan fungsi pankreas (Bouckenooghe et al. 2006). Meningkatnya koefisien kecernaan semu bahan kering juga dapat disebabkan karena suplementasi glisina dan taurina dapat meningkatkan kecernaan asam lemak jenuh.
Suplementasi lisina dan metionina tidak berpengaruh nyata terhadap koefisien kecernaan semu bahan kering. Hasil penelitian ini sesuai dengan hasil penelitian Ukpabi et al. (2008) yang menunjukan bahwa suplementasi lisina dan metionina pada pakan yang mengandung kacang tanah Bambarra tidak berpengaruh pada koefisien kecernaan bahan kering. Suplementasi lisina dan metionina tidak berpengaruh terhadap koefisien kecernaan semu bahan kering karena lisina dan metionina tidak terlibat langsung dalam proses biosintesis garam empedu, seperti halnya glisina dan taurina. Oleh karena itu, suplementasi lisina dan metionina tidak dapat meningkatkan berat kantung empedu, tetapi suplementasi glisina dan taurina nyata meningkatkan berat kantung empedu umur 21 hari. Peningkatan produksi empedu akan meningkatkan proses emulsifikasi lemak, sehingga akan mengurangi nutrien non lemak yang dibatasi oleh droplet lemak. Oleh karena itu, lebih banyak nutrien yang tersedia untuk dicerna oleh enzim pencernaan dan diabsorbsi oleh usus halus.
Koefisien Kecernaan Semu Lemak Kasar
Koefisien kecernaan semu lemak kasar ayam broiler yang diberi pakan mengandung minyak sawit 5% dengan suplementasi asam amino berkisar 86.87 sampai 93.46%. Suplementasi asam amino berpengaruh nyata (P<0.05) terhadap koefisien kecernaan lemak kasar ayam broiler umur 21 hari, dan berpengaruh sangat nyata (P<0.01) pada umur 42 hari. Suplementasi asam amino nyata meningkatkan koefisien kecernaan lemak kasar ayam broiler umur 21 hari. Akan tetapi, suplementasi asam amino tidak berpengaruh nyata terhadap koefisien kecernaan semu lemak kasar ayam broiler umur 42 hari. Hal ini dapat disebabkan karena ayam broiler umur 42 hari telah mampu memproduksi garam empedu dan enzim lipase dalam jumlah cukup untuk dapat mencerna lemak pakan.
Suplementasi glisina dan taurina nyata meningkatkan koefisien kecernaan semu lemak kasar. Akan tetapi, glisina lebih nyata meningkatkan koefisien kecernaan semu lemak kasar dibandingkan. Hasil penelitian ini sesuai dengan hasil penelitian sebelumnya yang menunjukan bahwa suplementasi glisina (Alzawqari et al. 2010) dan taurina (Zeweil et al. 2011) meningkatkan koefisien kecernaan semu lemak kasar. Peningkatan koefisien kecernaan semu lemak kasar ayam broiler umur 21 hari dapat disebabkan karena suplementasi glisina dan taurina mampu meningkatkan biosintesis empedu. Asam empedu yang disintesis dari kolesterol akan berkonjugasi dengan glisina atau taurina untuk membentuk

41
garam empedu memiliki peran yang penting dalam dalam pencernaan dan penyerapan lemak pada pakan dan nutrien lain yang larut dalam lemak (Zeweil et
al. 2011). Suplementasi lisina dan metionina tidak berpengaruh nyata terhadap
koefisien kecernaan semu lemak ayam broiler umur 21 hari, tetapi sigifikan menurunkan koefisien kecernaan lemak pada ayam broiler umur 42 hari. Hasil penelitian Ukpabi et al. (2008) juga menunjukan suplementasi lisina dan methinoni pada pakan yang mengandung kacang tanah Bambarra tidak nyata mempengaruhi koefisien kecernaan lemak kasar. Penurunan koefisien kecernaan semu lemak kasar pada ayam broiler umur 42 hari dapat disebabkan karena rendahnya produksi cairan empedu. Empedu dapat berfungsi sebagai antimikroba, maka produksi garam empedu yang rendah akan meningkatkan mikroba pada saluran pencernaan dan meningkatkan katabolisme empedu di saluran pencernaan (Alzawqari et al. 2010). Katabolisme garam empedu oleh mikroba di saluran pencernaan akan meningkatkan ekskresi garam empedu. Hal ini disebabkan karena aktivitas mikroba disaluran pencernaan akan mengganggu proses sirkulasi enterohepatik garam empedu (Smits et al. 1998).
Kombinasi suplementasi lisina dan metionina dengan glisina dan taurina juga menurunkan kemampuan glisina untuk meningkatkan koefisien kecernaan semu lemak kasar. Hasil penelitian Kussaibati dan Leclercq (1984) menunjukan bahwa berat empedu berkorelasi positif dengan koefisien kecernaan semu lemak kasar. Sebagai tambahan, suplementasi garam empedu dapat meningkatkan koefisien kecernaan semu lemak kasar ayam broiler yang diberi pakan mengandung asam lemak jenuh (Krogdahl 1985; Adrizal et al. 2002; Alzawqari et
al. 2010). Korelasi berat kantung empedu dengan koefisien kecernaan semu lemak kasar pada ayam broiler umur 21 dan 42 hari masing-masing adalah 0.84 dan 0.83. Hubungan antara volume cairan empedu dan koefisien kecernaan semu nutrien dapat diilustrasikan pada Gambar 10.
Gambar 10 Hubungan antara volume cairan empedu dan koefisien kecernaan semu nutrien
Koefisien Kecernaan Semu Protein Kasar
Koefisien kecernaan semu protein kasar ayam broiler berkisar 64.85 sampai 74.05%. Hasil penelitian ini sesuai dengan penelitian Adrizal et al. (2002) yang menunjukan bahwa koefisen kecernaan semu protein ekskreta berkisar 62.2 sampai 70.6%. Suplementasi asam amino berpengaruh sangat nyata (P<0.01) terhadap koefisien kecernaan semu protein kasar ayam broiler umur 21 hari, tetapi berpengaruh nyata (P<0.05) pada umur 42 hari. Hasil penelitian sebelumnya menunjukan bahwa suplementasi glisina pada pakan ayam petelur tidak
Biosintesis Asam Empedu
Volume Cairan Empedu Meningkat
Meningkatkan Koefisien Kecernaan BK, LK dan Energi
Meningkatkan Kerja Enzim untuk Mencerna Nutrien
Membantu Memecah Droplet Lemak

42
berpengaruh terhadap kecernaan ileum protein kasar dan asam amino (Han dan
Thacker 2011). Suplementasi taurina meningkatkan koefisien kecernaan protein umur 42 hari secara linier (Zeweil et al. 2011). Selanjutnya, peningkatan lisina dari 1.00 menjadi 1.18% pada pakan sedikit meningkatkan kecernaan semu ileum asam amino esensial dan non esensial. Perbedaan respon penelitian ini dengan penelitian sebelumnya dapat disebabkan karena perbedaan kandungan protein dan asam amino pada pakan basal.
Semua suplementasi asam amino menurunkan koefisien kecernaan semu protein kasar ayam broiler umur 21 hari. Akan tetapi, suplementasi asam amino meningkatkan koefisien kecernaan semu protein kasar ayam umur 42 hari, kecuali R3. Perbedaan respon suplementasi tersebut menunjukan bahwa koefisien kecernaan semu protein kasar dipengaruhi oleh umur ayam broiler. Hasil penelitian sebelumnya juga menunjukan bahwa kecernaan protein dipengaruhi oleh umur ayam broiler (Adrizal et al. 2002). Perbedaan respon suplementasi ini dapat disebabkan karena koefisien kecernaan semu protein kasar berkorelasi negatif dengan dengan konversi pakan. Oleh karena itu, peningkatan koefisien kecernaan semu protein kasar ayam broiler akan diiringi dengan penurunan konversi pakan. Korelasi koefisien kecernaan semu protein kasar dengan konversi pakan pada ayam broiler umur 21 dan 42 hari masing-masing adalah (-0.77) dan (-0.54). Hasil penelitian Gong et al. (2005) dan Selle et al. (2007) juga menunjukan bahwa koefisien kecernaan semu protein kasar berkorelasi negatif dengan konversi pakan.
Koefisien Kecernaan Semu Energi
Koefisien kecernaan semu energi ayam broiler berkisar 83.21 sampai 89.24%. Rataan koefisien kecernaan semu energi ayam broiler umur 21 adalah 87.66% sedangkan pada umur 42 hari 86.61%. Hasil penelitian ini menunjukan bahwa koefisien kecernaan semu energi ayam broiler umur 21 hari lebih tinggi dibandingkan umur 42 hari. Meningkatnya koefisien kecernaan semu energi dapat disebabkan meningkatnya koefisien kecernaan semu bahan kering ayam broiler umur 21 hari. Suplementasi asam amino berpengaruh nyata (P<0.05) terhadap koefisien kecernaan semu energi pada ayam broiler umur 21 hari dan berpengaruh sangat nyata (P<0.01) pada umur 42 hari. Suplementasi asam amino cenderung meningkatkan koefisien kecernaan semu energi, kecuali perlakuan R3. Koefisien kecernaan semu energi ayam yang diberi pakan dengan perlakuan R3 nyata menurun. Suplementasi glisina berpengaruh nyata terhadap koefisien kecernaan semu energi. Hal ini sesuai dengan hasil penelitian Han dan Thacker (2011) yang menunjukan bahwa suplementasi glisina sampai 0.1% pada pakan ayam petelur secara linier meningkatkan kecernaan ileum energi sampai 1.99%. Glisina berperan meningkatkan fungsi dari mukosa usus, sehingga suplementasi glisin dapat meningkatkan efisiensi penggunaan energi pada pakan (Ospina-Rojas et al. 2013). Glisina dan taurina juga merupakan prekursor dalam biosintesis garam empedu yang berperan penting dalam proses pencernaan dan penyerapan lemak, nutrien larut lemak, serta nutrien non lemak dalam pakan (lihat Gambar 10) (Stamp dan Jenkins 2008; Alzawqari et al. 2010).
Suplementasi lisina dan metionina nyata menurunkan koefisien kecernaan semu energi. Hasil penelitian sebelumnya menunjukan bahwa peningkatan kandungan lisina pakan dari 1.00 menjadi 1.18% menurunkan energi metabolis
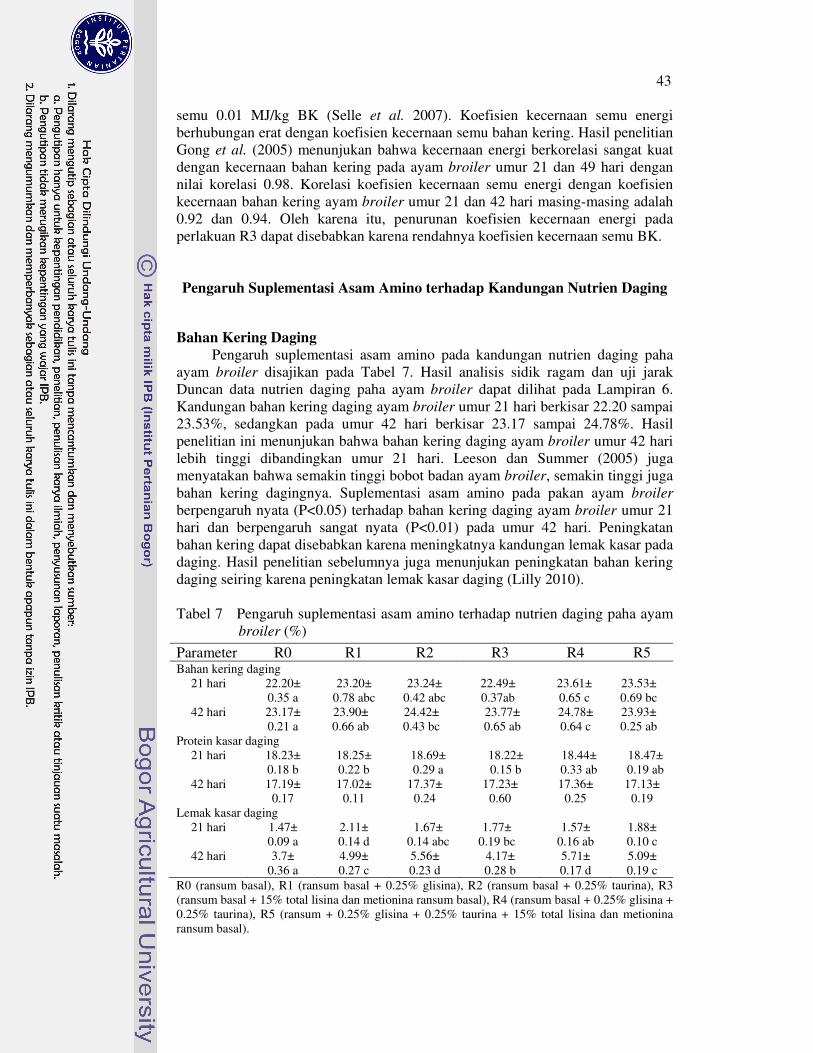
43
semu 0.01 MJ/kg BK (Selle et al. 2007). Koefisien kecernaan semu energi berhubungan erat dengan koefisien kecernaan semu bahan kering. Hasil penelitian Gong et al. (2005) menunjukan bahwa kecernaan energi berkorelasi sangat kuat dengan kecernaan bahan kering pada ayam broiler umur 21 dan 49 hari dengan nilai korelasi 0.98. Korelasi koefisien kecernaan semu energi dengan koefisien kecernaan bahan kering ayam broiler umur 21 dan 42 hari masing-masing adalah 0.92 dan 0.94. Oleh karena itu, penurunan koefisien kecernaan energi pada perlakuan R3 dapat disebabkan karena rendahnya koefisien kecernaan semu BK.
Pengaruh Suplementasi Asam Amino terhadap Kandungan Nutrien Daging
Bahan Kering Daging
Pengaruh suplementasi asam amino pada kandungan nutrien daging paha ayam broiler disajikan pada Tabel 7. Hasil analisis sidik ragam dan uji jarak Duncan data nutrien daging paha ayam broiler dapat dilihat pada Lampiran 6. Kandungan bahan kering daging ayam broiler umur 21 hari berkisar 22.20 sampai 23.53%, sedangkan pada umur 42 hari berkisar 23.17 sampai 24.78%. Hasil penelitian ini menunjukan bahwa bahan kering daging ayam broiler umur 42 hari lebih tinggi dibandingkan umur 21 hari. Leeson dan Summer (2005) juga menyatakan bahwa semakin tinggi bobot badan ayam broiler, semakin tinggi juga bahan kering dagingnya. Suplementasi asam amino pada pakan ayam broiler berpengaruh nyata (P<0.05) terhadap bahan kering daging ayam broiler umur 21 hari dan berpengaruh sangat nyata (P<0.01) pada umur 42 hari. Peningkatan bahan kering dapat disebabkan karena meningkatnya kandungan lemak kasar pada daging. Hasil penelitian sebelumnya juga menunjukan peningkatan bahan kering daging seiring karena peningkatan lemak kasar daging (Lilly 2010).
Tabel 7 Pengaruh suplementasi asam amino terhadap nutrien daging paha ayam
broiler (%)
Parameter R0 R1 R2 R3 R4 R5 Bahan kering daging
21 hari 22.20± 0.35 a
23.20± 0.78 abc
23.24± 0.42 abc
22.49± 0.37ab
23.61± 0.65 c
23.53± 0.69 bc
42 hari 23.17± 0.21 a
23.90± 0.66 ab
24.42± 0.43 bc
23.77± 0.65 ab
24.78± 0.64 c
23.93± 0.25 ab
Protein kasar daging 21 hari 18.23±
0.18 b 18.25± 0.22 b
18.69± 0.29 a
18.22± 0.15 b
18.44± 0.33 ab
18.47± 0.19 ab
42 hari 17.19± 0.17
17.02± 0.11
17.37± 0.24
17.23± 0.60
17.36± 0.25
17.13± 0.19
Lemak kasar daging 21 hari 1.47±
0.09 a 2.11± 0.14 d
1.67± 0.14 abc
1.77± 0.19 bc
1.57± 0.16 ab
1.88± 0.10 c
42 hari 3.7± 0.36 a
4.99± 0.27 c
5.56± 0.23 d
4.17± 0.28 b
5.71± 0.17 d
5.09± 0.19 c
R0 (ransum basal), R1 (ransum basal + 0.25% glisina), R2 (ransum basal + 0.25% taurina), R3 (ransum basal + 15% total lisina dan metionina ransum basal), R4 (ransum basal + 0.25% glisina + 0.25% taurina), R5 (ransum + 0.25% glisina + 0.25% taurina + 15% total lisina dan metionina ransum basal).

44
Suplementasi asam amino menyebabkan asam amino pada pakan menjadi berlebihan. Kelebihan asam amino akan dideaminasi dan atau ditransaminasi, proses ini menghasilkan rangka karbon yang kemudian digunakan untuk sintesis lemak (lihat Gambar 8). Oleh karena itu, suplementasi asam amino meningkatkan kandungan lemak kasar daging. Hasil penelitian ini dan penelitian sebelumnya menunjukan suplementasi asam amino pada pakan cukup asam amino hanya sedikit berpengaruh terhadap peningkatan protein kasar daging (Namroud et al. 2010; Lilly 2010). Akan tetapi, hasil penelitian Zeweil et al. (2011) menunjukan suplementasi taurina dapat meningkatkan bahan kering daging dada yang disebabkan meningkatnya kandungan protein. Kandungan bahan kering daging R4 paling tinggi dibandingkan perlakuan lainnya, hal ini disebabkan karena terjadi peningkatan kandungan lemak dan protein secara bersamaan.
Protein Kasar Daging
Protein daging ayam broiler umur 21 berkisar 18.22 sampai 18.69%, sedangkan pada umur 42 hari berkisar 17.02 sampai 17.37%. Suplementasi asam amino berpengaruh nyata (P<0.05) terhadap kandungan protein daging ayam broiler umur 21, tidak berpengaruh nyata (P>0.05) pada umur 42 hari. Leeson dan
Summer (2005) menyatakan bahwa protein pada karkas sedikit dipengaruhi oleh nutrisi, dengan asumsi tidak ada asam amino yang defisiensi dan hasil daging sesuai dengan potensi genetiknya. Beberapa hasil penelitian juga menunjukan bahwa ayam broiler yang diberi pakan dengan kandungan protein dan level energi yang berbeda tidak berpengaruh nyata terhadap protein kasar daging dada (Barteczko dan Lasek 2008; Horniakova dan Abas 2009; Namroud et al. 2010). Suplementasi asam amino dilaporkan dapat meningkatkan kandungan protein daging ketika disuplementasi pada pakan dengan sumber protein nabati atau pakan yang defisien asam amino. Zeweil et al. (2011) menyatakan bahwa suplementasi taurina pada pakan nabati nyata meningkatkan kandungan protein kasar daging. Hasil penelitian lainnya menunjukan peningkatan densitas asam amino pada pakan defisiensi asam amino nyata meningkatkan kandungan protein kasar daging (Lilly 2010). Asam amino yang defisien dan tidak seimbang akan mengganggu sintesis protein, sehingga menyebabkan penurunan protein daging (Kidd et al. 1998). Suplementasi taurina nyata meningkatkan kandungan protein daging ayam broiler umur 21 hari. Hal ini dapat disebabkan karena meningkatnya konsentrasi taurina bebas pada daging paha. Hasil penelitian Lee et al. (2004) menunjukan suplementasi taurina sintetik 0.5% meningkatkan kandungan taurina daging paha 246% dibandingkan kontrol. Sebagai tambahan, kandungan taurina pada otot dada adalah 778 µg/g.
Lemak Kasar Daging
Kandungan lemak kasar daging dada ayam broiler umur 21 hari berkisar 1.47 sampai 2.11%, sedangkan pada umur 42 berkisar 3.70 sampai 5.71%. Suplementasi asam amino berpengaruh sangat nyata (P<0.01) terhadap kandungan lemak kasar daging paha ayam broiler umur 21 dan 42 hari. Hasil penelitian juga menunjukan rataan kandungan lemak kasar daging paha umur 42 hari lebih tinggi dibandingkan umur 21 hari. Hasil penelitian ini sesuai dengan pernyataaan Leeson
dan Summer (2005) yang menyatakan bahwa kandungan lemak pada karkas sangat dipengaruhi nutrisi dan umur ayam broiler. Semakin besar energi yang

45
dikonsumsi dan semakin tua ayam broiler, semakin besar pula potensi lemak dideposit. Hasil penelitian juga menunjukan bahwa suplementasi asam amino pada ayam broiler umur 21 hari terlihat belum optimum berpengaruh terhadap pembentukan lemak daging. Akan tetapi, pengaruh pakan terhadap lemak daging lebih jelas terlihat pada umur 42 hari, terutama pengaruh glisina dan taurina. Selanjutnya, kandungan asam lemak pada jaringan adiposa tergantung pada jenis lemak yang dikonsumsi (Crespo et al. 2001). Kandungan asam lemak daging yang diberi pakan mengandung sawit 6% sebagian besar terdiri dari oleat (18:1), palmitat (16:0) dan stearat (18:0) (Panja et al. 1995).
Peningkatan kandungan lemak juga dapat disebabkan karena suplementasi asam amino menyebabkan asam amino pada pakan tidak seimbang dan berlebihan sehingga tidak dapat digunakan untuk sintesis protein. Pada saat kelebihan protein bersamaan dengan intake karbohidrat yang cukup, maka rangka karbon dari asam amino akan digunakan untuk sintesis glukosa dan asam lemak (Gropper et al. 2009). Oleh karena itu, suplementasi asam amino menyebabkan peningkatan kandungan trigliserida serum dan lemak kasar daging. Hasil penelitian sebelumnya menunjukan kandungan lemak daging berkorelasi positif terhadap kadar trigliserida darah (Lilly 2010; Zeweil et al. 2011; Bouyeh dan Gevorgyan 2011). Asam amino yang tidak seimbang dan defisien juga menyebabkan penurunan sekresi hormon tiroksin, sehingga meningkatkan deposit lemak (Namroud et al. 2010). Kandungan lemak kasar daging ayam broiler yang paling tinggi adalah perlakuan R1 untuk umur 21 hari dan R4 untuk umur 42 hari. Tingginya kandungan lemak kasar daging disebabkan karena ayam dengan perlakuan tersebut paling tinggi trigliserida serumnya. Kandungan trigliserida serum ayam yang diberi perlakuan R1 pada umur 21 hari adalah 71.39 mg dl-1, sedangkan ayam perlakuan R4 pada umur 42 hari 82.08 mg dl-1.
L-Karnitin Daging
Kandungan L-karnitin pada daging ayam broiler berkisar 7.49 sampai 15.28 mg 100 g-1. Akan tetapi, hasil penelitian Xu et al. (2003) menunjukan bahwa kandungan karnitin bebas pada daging dada berkisar 0.96 sampai 1.15 µmol g-1, atau setara dengan 15.36 sampai 18.40 mg 100 g-1. Perbedaan konsentrasi karnitin tersebut dapat disebabkan karena perbedaan sampel yang digunakan dan bentuk karnitin yang diamati. Karnitin yang diamati pada penelitian ini adalah L-karnitin pada daging paha, sedangkan penelitian Xu et al. (2003) mengamati karnitin bebas pada daging dada.
Konsentrasi L-karnitin pada daging ayam broiler umur 21 dan 42 hari disajikan pada Gambar 11. Kandungan L-karnitin daging ayam broiler umur 21 lebih tinggi dibandingkan umur 42 hari, kecuali pada R0. Kandungan L-karnitin daging ayam broiler umur 21 hari berkisar 8.83 sampai 15.28 mg 100 g-1, sedangkan pada umur 42 hari berkisar 7.49 sampai 12.70 mg 100 g-1. Konsentrasi L-karnitin sangat bergantung pada spesies, tipe jaringan, status nutrisi, serta bentuk senyawanya (Harmeyer 2002; Arslan 2006). Asam amino lisina dan metionina merupakan prekursor dalam proses biosintesis L-karnitin. Ketersediaan lisina dan metionina untuk biosintesis L-karnitin di dalam tubuh dipengaruhi oleh koefisien kecernaan protein kasar. Oleh karena itu, kandungan L-karnitin daging berkorelasi positif dengan koefisien kecernaan semu protein kasar. Koefisien kecernaan semu protein kasar ayam broiler umur 21 lebih tinggi dibandingkan
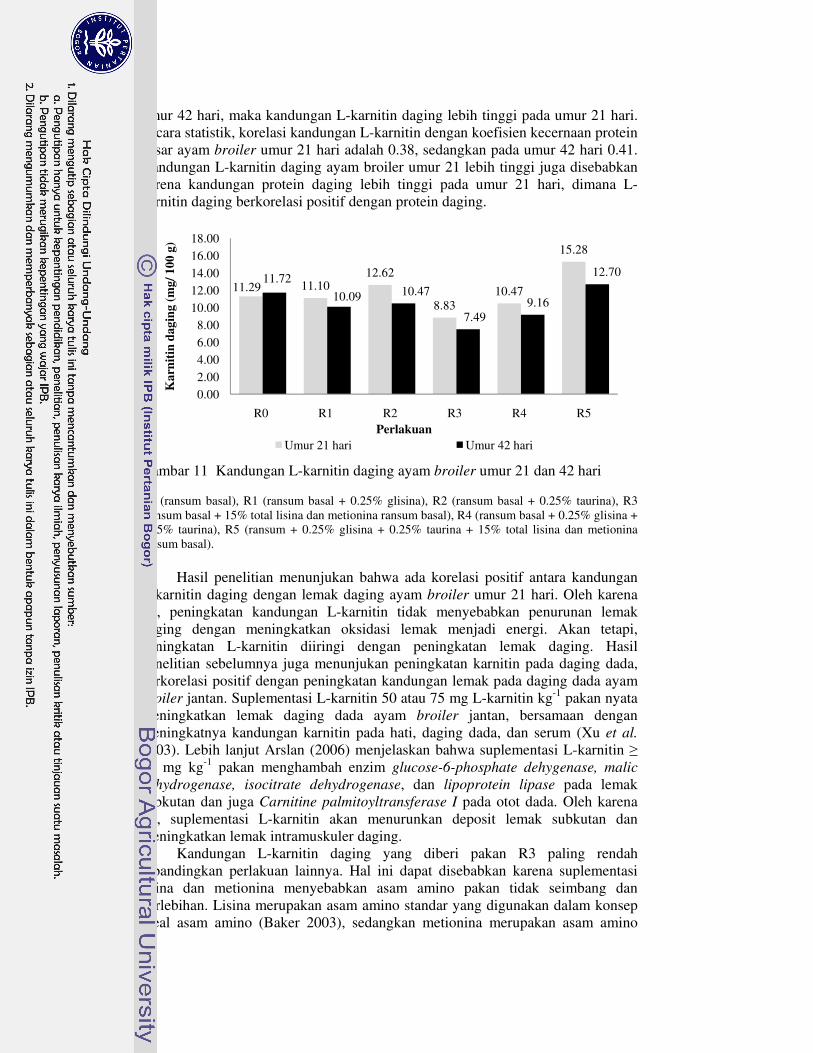
46
umur 42 hari, maka kandungan L-karnitin daging lebih tinggi pada umur 21 hari. Secara statistik, korelasi kandungan L-karnitin dengan koefisien kecernaan protein kasar ayam broiler umur 21 hari adalah 0.38, sedangkan pada umur 42 hari 0.41. Kandungan L-karnitin daging ayam broiler umur 21 lebih tinggi juga disebabkan karena kandungan protein daging lebih tinggi pada umur 21 hari, dimana L-karnitin daging berkorelasi positif dengan protein daging.
Gambar 11 Kandungan L-karnitin daging ayam broiler umur 21 dan 42 hari R0 (ransum basal), R1 (ransum basal + 0.25% glisina), R2 (ransum basal + 0.25% taurina), R3 (ransum basal + 15% total lisina dan metionina ransum basal), R4 (ransum basal + 0.25% glisina + 0.25% taurina), R5 (ransum + 0.25% glisina + 0.25% taurina + 15% total lisina dan metionina ransum basal).
Hasil penelitian menunjukan bahwa ada korelasi positif antara kandungan
L-karnitin daging dengan lemak daging ayam broiler umur 21 hari. Oleh karena itu, peningkatan kandungan L-karnitin tidak menyebabkan penurunan lemak daging dengan meningkatkan oksidasi lemak menjadi energi. Akan tetapi, peningkatan L-karnitin diiringi dengan peningkatan lemak daging. Hasil penelitian sebelumnya juga menunjukan peningkatan karnitin pada daging dada, berkorelasi positif dengan peningkatan kandungan lemak pada daging dada ayam broiler jantan. Suplementasi L-karnitin 50 atau 75 mg L-karnitin kg-1 pakan nyata meningkatkan lemak daging dada ayam broiler jantan, bersamaan dengan meningkatnya kandungan karnitin pada hati, daging dada, dan serum (Xu et al. 2003). Lebih lanjut Arslan (2006) menjelaskan bahwa suplementasi L-karnitin $ 50 mg kg-1 pakan menghambah enzim glucose-6-phosphate dehygenase, malic
dehydrogenase, isocitrate dehydrogenase, dan lipoprotein lipase pada lemak subkutan dan juga Carnitine palmitoyltransferase I pada otot dada. Oleh karena itu, suplementasi L-karnitin akan menurunkan deposit lemak subkutan dan meningkatkan lemak intramuskuler daging.
Kandungan L-karnitin daging yang diberi pakan R3 paling rendah dibandingkan perlakuan lainnya. Hal ini dapat disebabkan karena suplementasi lisina dan metionina menyebabkan asam amino pakan tidak seimbang dan berlebihan. Lisina merupakan asam amino standar yang digunakan dalam konsep ideal asam amino (Baker 2003), sedangkan metionina merupakan asam amino
11.29 11.1012.62
8.8310.47
15.28
11.72
10.09 10.47
7.499.16
12.70
0.00
2.00
4.00
6.00
8.00
10.00
12.00
14.00
16.00
18.00
R0 R1 R2 R3 R4 R5
Ka
rnit
in d
ag
ing
(m
g/
10
0 g
)
Perlakuan
Umur 21 hari Umur 42 hari

47
yang paling toksik dibandingkan asam amino lain (Adeyemo et al. 2010). Oleh karena itu, suplementasi lisina dan metionina menyebabkan stres metabolis pada ternak, sehingga penurunan L-karnitin daging. Hal ini berdasarkan pernyataan Arslan (2006) yang menyatakan bahwa hewan dalam kondisi stres dapat menyebabkan proses biosintesis L-karnitin tidak cukup. Hasil penelitian Sachan et
al. (1984) juga menunjukan bahwa tikus yang stres metabolis karena diberi pakan mengandung etanol nyata menurunkan karnitin plasma. Sebagai tambahan, suplementasi lisina dan metionina pada kondisi tersebut tidak mampu meningkatkan kandungan karnitin plasma. Kondisi stres metabolis salah satunya ditunjukan dengan meningkatnya berat hati. Hasil penelitian sebelumnya menunjukan penambahan etanol (Sachan et al. 1984) serta suplementasi lisina dan metionina (Bouyeh dan Gevorgyan 2011) meningkatkan berat hati. Hasil penelitian ini juga menunjukan bahwa suplementasi lisina dan metionina cenderung meningkatkan berat hati ayam broiler umur 21 dan 42 hari, walaupun tidak nyata (lihat Tabel 8).
Kandungan L-karnitin daging ayam broiler yang diberi pakan R5 pada umur 21 dan 42 hari cenderung meningkat. Hal ini menunjukan bahwa kombinasi suplementasi lisina, metionina, glisina dan taurina dapat meningkatkan keseimbangan asam amino sehingga dapat meningkatkan kandungan L-karnitin pada daging ayam broiler. Akan tetapi, suplementasi sintetik L-karnitin lebih efisien untuk meningkatkan kandungan karnitin pada jaringan, dibandingkan suplementasi prekursor dan kofaktor L-karnitin. Hasil penelitian Xu et al. (2003) menunjukan bahwa suplementasi L-karnitin nyata meningkatkan kandungan karnitin pada hati, daging dada, dan serum ayam broiler jantan. Sebaliknya, suplementasi lisina dan metionina gagal meningkatkan kandungan karnitin plasma pada tikus yang diberi pakan mengandung etanol (Sachan et al. 1984). Arslan (2006) juga menyatakan bahwa suplementasi lisina dan metionina pada pakan tidak berpengaruh terhadap level karnitin pada jaringan.
Respon suplementasi L-karnitin dan prekursornya pada unggas dipengaruhi oleh kandungan L-karnitin dan prekursornya dalam pakan, lama periode suplementasi, suplai energi metabolis, jenis kelamin, status genotip dan fisiologi ternak, dan kondisi lingkungan (Arslan 2006). Berbeda dengan kandungan karnitin pada jaringan ruminansia yang diatur oleh kontrol metabolis dan tidak dipengaruhi oleh pakan (Greenwood et al. 2001). Biosintesis L-karnitin dalam tubuh dapat memenuhi kebutuhan L-karnitin normal pada semua mamalia dan spesies burung, ketika prekursor dan kofaktor tersedia dalam jumlah cukup dalam pakan. Akan tetapi, pada hewan yang baru lahir (proses biosintesis belum cukup berkembang), kondisi stres, ternak dengan performa tinggi, kelelahan fisik dan ketika diberi pakan kaya lemak (Arslan 2006).
Korelasi Konsumsi Lisina+metionina dengan Kandungan L-karnitin Daging
Korelasi konsumsi lisina dan metionina dengan kandungan karnitin daging ayam broiler umur 21 dan 42 hari disajikan pada Gambar 12. Korelasi konsumsi lisina dan metionina dengan kandungan L-karnitin daging ayam broiler umur 21 hari adalah 0.386 sedangkan pada umur 42 hari hanya 0.122. Hal ini menunjukan bahwa konsumsi lisina dan metionina memiliki hubungan yang rendah dengan kandungan L-karnitin pada daging paha. Sama halnya seperti hubungan konsumsi lisina dan metionina terhadap kadar karnitin bebas pada manusia dengan pola
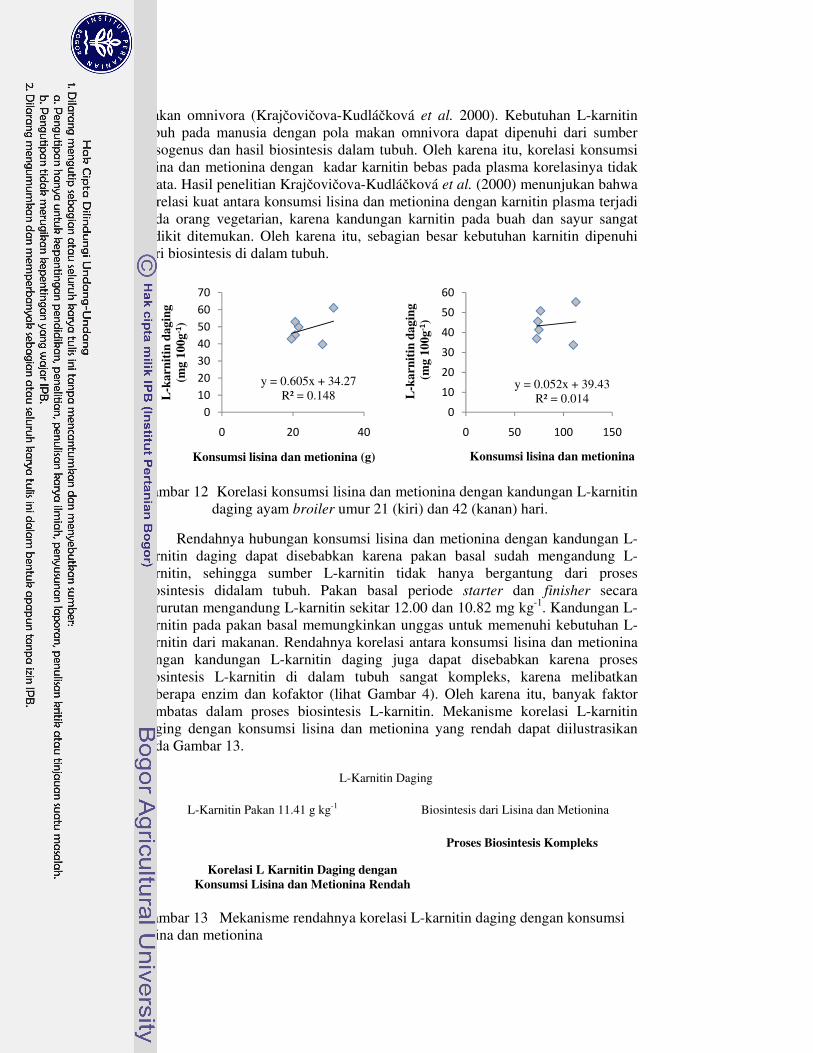
48
makan omnivora (Kraj�ovi�ova-Kudlá�ková et al. 2000). Kebutuhan L-karnitin tubuh pada manusia dengan pola makan omnivora dapat dipenuhi dari sumber eksogenus dan hasil biosintesis dalam tubuh. Oleh karena itu, korelasi konsumsi lisina dan metionina dengan kadar karnitin bebas pada plasma korelasinya tidak nyata. Hasil penelitian Kraj�ovi�ova-Kudlá�ková et al. (2000) menunjukan bahwa korelasi kuat antara konsumsi lisina dan metionina dengan karnitin plasma terjadi pada orang vegetarian, karena kandungan karnitin pada buah dan sayur sangat sedikit ditemukan. Oleh karena itu, sebagian besar kebutuhan karnitin dipenuhi dari biosintesis di dalam tubuh.
Gambar 12 Korelasi konsumsi lisina dan metionina dengan kandungan L-karnitin
daging ayam broiler umur 21 (kiri) dan 42 (kanan) hari.
Rendahnya hubungan konsumsi lisina dan metionina dengan kandungan L-karnitin daging dapat disebabkan karena pakan basal sudah mengandung L-karnitin, sehingga sumber L-karnitin tidak hanya bergantung dari proses biosintesis didalam tubuh. Pakan basal periode starter dan finisher secara berurutan mengandung L-karnitin sekitar 12.00 dan 10.82 mg kg-1. Kandungan L-karnitin pada pakan basal memungkinkan unggas untuk memenuhi kebutuhan L-karnitin dari makanan. Rendahnya korelasi antara konsumsi lisina dan metionina dengan kandungan L-karnitin daging juga dapat disebabkan karena proses biosintesis L-karnitin di dalam tubuh sangat kompleks, karena melibatkan beberapa enzim dan kofaktor (lihat Gambar 4). Oleh karena itu, banyak faktor pembatas dalam proses biosintesis L-karnitin. Mekanisme korelasi L-karnitin daging dengan konsumsi lisina dan metionina yang rendah dapat diilustrasikan pada Gambar 13.
Gambar 13 Mekanisme rendahnya korelasi L-karnitin daging dengan konsumsi lisina dan metionina
Biosintesis dari Lisina dan Metionina
L-Karnitin Daging
L-Karnitin Pakan 11.41 g kg-1
Korelasi L Karnitin Daging dengan
Konsumsi Lisina dan Metionina Rendah
Proses Biosintesis Kompleks
y = 0.605x + 34.27R² = 0.148
�
��
��
��
��
��
��
�
� �� ��
L-k
arn
itin
da
gin
g
(mg
10
0g��
)
Konsumsi lisina dan metionina (g)
y = 0.052x + 39.43R² = 0.014
�
��
��
��
��
��
��
� �� ��� ���
L-k
arn
itin
da
gin
g
(mg
10
0g��
)
Konsumsi lisina dan metionina (g)
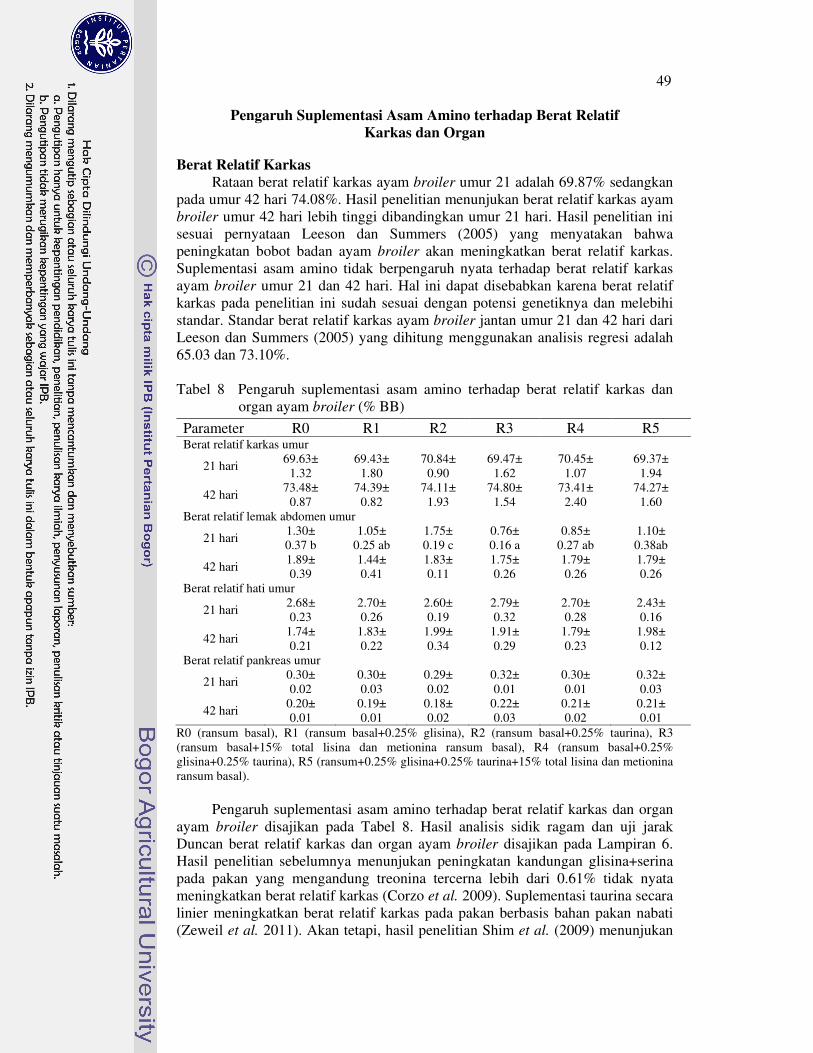
49
Pengaruh Suplementasi Asam Amino terhadap Berat Relatif
Karkas dan Organ
Berat Relatif Karkas
Rataan berat relatif karkas ayam broiler umur 21 adalah 69.87% sedangkan pada umur 42 hari 74.08%. Hasil penelitian menunjukan berat relatif karkas ayam broiler umur 42 hari lebih tinggi dibandingkan umur 21 hari. Hasil penelitian ini sesuai pernyataan Leeson dan Summers (2005) yang menyatakan bahwa peningkatan bobot badan ayam broiler akan meningkatkan berat relatif karkas. Suplementasi asam amino tidak berpengaruh nyata terhadap berat relatif karkas ayam broiler umur 21 dan 42 hari. Hal ini dapat disebabkan karena berat relatif karkas pada penelitian ini sudah sesuai dengan potensi genetiknya dan melebihi standar. Standar berat relatif karkas ayam broiler jantan umur 21 dan 42 hari dari Leeson dan Summers (2005) yang dihitung menggunakan analisis regresi adalah 65.03 dan 73.10%.
Tabel 8 Pengaruh suplementasi asam amino terhadap berat relatif karkas dan
organ ayam broiler (% BB)
Parameter R0 R1 R2 R3 R4 R5 Berat relatif karkas umur
21 hari 69.63±
1.32 69.43±
1.80 70.84±
0.90 69.47±
1.62 70.45±
1.07 69.37±
1.94
42 hari 73.48±
0.87 74.39±
0.82 74.11±
1.93 74.80±
1.54 73.41±
2.40 74.27±
1.60 Berat relatif lemak abdomen umur
21 hari 1.30± 0.37 b
1.05± 0.25 ab
1.75± 0.19 c
0.76± 0.16 a
0.85± 0.27 ab
1.10± 0.38ab
42 hari 1.89± 0.39
1.44± 0.41
1.83± 0.11
1.75± 0.26
1.79± 0.26
1.79± 0.26
Berat relatif hati umur
21 hari 2.68± 0.23
2.70± 0.26
2.60± 0.19
2.79± 0.32
2.70± 0.28
2.43± 0.16
42 hari 1.74± 0.21
1.83± 0.22
1.99± 0.34
1.91± 0.29
1.79± 0.23
1.98± 0.12
Berat relatif pankreas umur
21 hari 0.30± 0.02
0.30± 0.03
0.29± 0.02
0.32± 0.01
0.30± 0.01
0.32± 0.03
42 hari 0.20± 0.01
0.19± 0.01
0.18± 0.02
0.22± 0.03
0.21± 0.02
0.21± 0.01
R0 (ransum basal), R1 (ransum basal+0.25% glisina), R2 (ransum basal+0.25% taurina), R3 (ransum basal+15% total lisina dan metionina ransum basal), R4 (ransum basal+0.25% glisina+0.25% taurina), R5 (ransum+0.25% glisina+0.25% taurina+15% total lisina dan metionina ransum basal).
Pengaruh suplementasi asam amino terhadap berat relatif karkas dan organ
ayam broiler disajikan pada Tabel 8. Hasil analisis sidik ragam dan uji jarak Duncan berat relatif karkas dan organ ayam broiler disajikan pada Lampiran 6. Hasil penelitian sebelumnya menunjukan peningkatan kandungan glisina+serina pada pakan yang mengandung treonina tercerna lebih dari 0.61% tidak nyata meningkatkan berat relatif karkas (Corzo et al. 2009). Suplementasi taurina secara linier meningkatkan berat relatif karkas pada pakan berbasis bahan pakan nabati (Zeweil et al. 2011). Akan tetapi, hasil penelitian Shim et al. (2009) menunjukan

50
bahwa injeksi taurina menurunkan sintesis in vitro protein 54-kDa, 37-kDa dan 24-kDa. Selanjutnya, suplementasi lisina dan metionina tidak berpengaruh terhadap berat relatif karkas ayam broiler (Cengiz et al. 2008; Lilly 2010; Ahmed
dan Abbas 2011). Peningkatan level lisina pada pakan yang cukup lisina juga tidak berpengaruh terhadap retensi protein ayam broiler (Tavernari et al. 2009). Akan tetapi, penelitian lain menunjukan bahwa suplementasi lisina dan metionina pada pakan nyata meningkatkan berat relatif karkas (Bouyeh dan Gevorgyan 2011; Malomo et al. 2013). Perbedaan respon berat relatif karkas dengan suplementasi asam amino dapat disebabkan karena perbedaan komposisi pakan, khususnya kandungan lisina dan protein pakan basal. Suplementasi dan peningkatan level lisina pakan yang sesuai dengan rekomendasi NRC (1994) tidak berpengaruh nyata terhadap berat relatif karkas. Suplementasi lisina pada pakan nyata meningkatkan berat relatif karkas ketika diberikan pada pakan yang defisien protein atau asam amino (Kidd et al. 1998; Kerr et al. 1999; Vieira et al. 2004; Corzo et al. 2009; Ahmed dan Abbas 2011).
Berat Relatif Lemak Abdomen
Rataan berat relatif lemak abdomen ayam broiler umur 21 hari adalah 1.14% kemudian pada umur 42 hari 1.82%. Hal ini menunjukan bahwa berat relatif lemak abdomen lebih tinggi pada ayam broiler umur 42 hari dibandingkan umur 21 hari. Leeson dan Summers (2005) juga menyatakan bahwa bertambahnya umur ayam broiler akan meningkatkan lemak dalam tubuh, khususnya lemak abdomen. Ayam broiler mengandung lemak abdomen sekitar 2 sampai 3%. faktor utama yang berpengaruh pada perlemakan abdomen ayam broiler adalah jenis kelamin, umur, efisiensi pakan, genetik, dan komposisi pakan. Jaringan lemak pada betina lebih besar pada ayam broiler betina dan dewasa dibandingkan pada ayam broiler jantan dan muda. Selanjutnya, ayam broiler dengan konversi pakan yang baik memiliki lemak abdomen yang rendah (Tavernari et al. 2009).
Suplementasi asam amino berpengaruh sangat nyata (P<0.01) terhadap berat relatif ayam lemak abdomen ayam broiler umur 21 hari, tetapi tidak nyata (P>0.05) pada umur 42 hari. Suplementasi asam amino cenderung menurunkan berat relatif lemak abdomen. kecuali R2. Suplementasi glisina sedikit menurunkan berat relatif lemak abdomen, walaupun tidak nyata. Hasil penelitian sebelumnya, menunjukan bahwa suplementasi glisina dan glutamat pada pakan rendah protein menurunkan berat relatif lemak abdomen (Namroud et al. 2010). Hal ini disebabkan karena glisina dapat mengaktifkan fungsi kelenjar tiroid dengan meningkatkan sekresi hormon triiodotironin dan tiroksin. Hormon triiodotironin dapat menurunkan aktifitas enzim malic yang memiliki aktifitas lipogenesis. Selanjutnya, hormon tiroksin berkorelasi negatif dengan berat relatif lemak abdomen. Oleh karena itu, hormon tiroksin dapat digunakan untuk memprediksi lemak karkas ayam pullet yang diberi pakan ad libitum (Sun et al. 2006).
Hasil penelitian Zeweil et al. (2011) juga menemukan bahwa suplementasi taurina pada pakan nabati nyata menurunkan lemak abdomen. Akan tetapi, suplementasi taurina pada penelitian ini nyata meningkatkan berat relatif lemak abdomen. Meningkatnya lemak abdomen dapat disebabkan karena ayam broiler yang diberi pakan dengan suplementasi taurina memiliki konsumsi pakan yang rendah dan konversi pakan yang tinggi. Lemak abdomen berkorelasi positif

51
dengan konversi pakan dan berkorelasi negatif dengan konsumsi pakan. Namroud et al. (2010) menjelaskan bahwa lemak abdomen berkorelasi positif dengan konsumsi pakan, hal ini disebakan karena konsumsi pakan juga berkorelasi positif dengan fungsi kelenjar tiroid. Selanjutnya, Kiraz dan %engül (2005) menemukan bahwa berat relatif lemak abdomen berkorelasi positif dengan konversi pakan. Hasil penelitian ini juga menunjukan bahwa berat relatif lemak abdomen ayam broiler umur 21 hari berkorelasi positif dengan konversi pakan (0.44) dan berkorelasi negatif dengan konsumsi pakan (-0.02). Meningkatnya konversi pakan disebabkan karena energi pakan lebih banyak digunakan untuk pembentukan lemak tubuh dibandingkan pembentukan daging (Kidd et al. 1998).
Suplementasi lisina dan metionina nyata menurunkan berat relatif lemak abdomen ayam broiler umur 21 hari, tetapi tidak berpengaruh nyata pada umur 42 hari. Hasil penelitian sebelumnya menunjukan peningkatan level metionina (Ahmed dan Abbas 2011) dan lisina (Grisoni et al. 1991) nyata menurunkan berat relatif lemak abdomen ayam broiler. Hasil penelitian pertama Kerr et al. (1999) juga menunjukan bahwa peningkatan lisina pada pakan nyata menurunkan lemak abdomen, namun tidak nyata pada penelitian kedua. Selanjutnya, Kerr et al. (1999) menjelaskan bahwa level lemak abdomen sangat bervariasi yang tidak hanya dipengaruhi oleh lisina pada pakan, tetapi juga oleh genetik dan umur pemotongan. Suplementasi asam amino tunggal dan peningkatan protein pada pakan dapat menurunkan berat relatif lemak abdomen. Penurunan berat relatif lemak abdomen disebabkan karena menurunnya aktivitas enzim malic yang berperan dalam proses biosintesis asam lemak (Grisoni et al. 1991; Taraz et al. 2006; Namroud et al. 2010).
Berat Relatif Hati
Rataan berat relatif hati ayam broiler umur 21 adalah 2.65%, sedangkan umur 42 hari 1.87%. Hati merupakan organ aksesoris yang penting dalam sistem pencernaan. Hati berfungsi memetabolisme nutrien yang diabsorbsi dan memproduksi asam dan garam empedu. Suplementasi asam amino tidak berpengaruh nyata (P>0.05) terhadap berat relatif hati ayam broiler umur 21 dan 42 hari. Hasil penelitian sebelumnya menunjukan bahwa suplementasi glisina hingga 0.1% tidak berpengaruh terhadap berat relatif hati ayam petelur (Han dan
Thacker 2011). Suplementasi taurina 0.05% juga tidak berpengaruh nyata terhadap berat giblet (hati, jantung, gizzard) ayam broiler umur 42 hari (Zeweil et
al. 2011). Selanjutnya, hasil penelitian Cengiz et al. (2008) menunjukan suplementasi lisina dan metionina tidak berpengaruh terhadap berat relatif hati ayam broiler (Cengiz et al. 2008). Hasil penelitian lainnya juga menunjukan bahwa level energi dan lisina (Ullah et al. 2012), serta metionina (Ahmed dan
Abbas 2011) tidak berpengaruh nyata terhadap berat relatif hati ayam broiler. Akan tetapi, hasil penelitian Bouyeh dan Gevorgyan (2011) menunjukan
bahwa suplementasi lisina dan metionina nyata meningkatkan berat relatif hati pada ayam broiler umur 42 hari. Hal ini dapat disebabkan karena suplementasi lisina dan metionina meningkatkan laju metabolisme untuk sintesis L-karnitin, glukosa, kolesterol, protein dan juga asam urat hasil degradasi lisina dan metionina yang berlebih (Bouyeh dan Gevorgyan, 2011). Suplementasi lisina dan metionina pada penelitian ini juga cenderung meningkatkan berat relatif hati, meskipun tidak nyata. Suplementasi asam amino tidak berpengaruh nyata

52
terhadap berat relatif hati dapat disebabkan karena kelebihan asam amino masih dapat ditoleransi oleh ayam broiler. Hasil penelitian ini juga menunjukan bahwa proses biosintesis empedu tidak berhubungan dengan berat relatif relatif hati. Oleh karena itu, peningkatan berat relatif empedu tidak diiringi dengan meningkatnya berat relatif hati.
Berat Relatif Pankreas
Rataan berat relatif pankreas ayam broiler umur 21 hari adalah 0.30% sedangkan pada umur 42 hari 0.20%. Pankreas terdiri dari tiga buah kelenjar pencernaan yang terletak pada bagian dalam duodenum unggas (Pesti et al. 2005). Pankreas memproduksi getah pankreas yang berisi enzim pencernaan seperti amilase, lipase, tripsin, kimotripsin, karboksipeptidase, ribonuklease, dan elastase. Pankreas juga menghasilkan bikarbonat yang berfungsi sebagai buffer pH saluran pencernaan (Klansing 2006). Sekresi getah pankreas distimulasi oleh kolesistokinin dan dihambat oleh somatotropin dan glukagon (Larbier dan Leclerq 1994). Suplementasi asam amino tidak berpengaruh nyata (P>0.05) terhadap berat relatif pankreas ayam broiler umur 21 dan 42 hari. Hasil penelitian sebelumnya juga menunjukan suplementasi glisina tidak berpengaruh terhadap berat relatif pankreas ayam petelur (Han dan Thacker 2011). Selanjutnya, hasil penelitian Zeweil et al. (2011) menunjukan bahwa suplementasi taurina 0.025% tidak berpengaruh nyata terhadap berat relatif pankreas ayam broiler. Asam amino glisina, taurina, lisina dan metionina tidak terlibat dalam sintesis getah pankreas. Oleh karena itu, suplementasi asam amino tidak berpengaruh terhadap berat relatif pankreas. Hasil penelitian ini juga menunjukan bahwa peningkatan koefisien kecernaan lemak semu ayam broiler umur 21 hari dengan suplementasi asam amino tidak dipengaruhi oleh peningkatan berat relatif pankreas.

53
SIMPULAN DAN SARAN
Simpulan
Suplementasi glisina serta kombinasi glisina dan taurina mampu meningkatkan koefisien kecernaan semu lemak kasar ayam broiler umur 21 hari. Akan tetapi, kombinasi suplementasi lisina dan metionina yang mampu menurunkan deposit lemak abdomen. Proses biosintesis garam empedu lebih dipengaruhi oleh glisina dibandingkan taurina. Suplementasi glisina, taurina dan kombinasi keduanya mampu meningkatkan berat kantung empedu ayam broiler umur 21 hari. Selanjutnya, kandungan L-karnitin daging sedikit dipengaruhi oleh konsumsi lisina dan metionina. Kombinasi suplementasi lisina dan metionina cenderung menurunkan kandungan L-karnitin daging ayam broiler umur 21 dan 42 hari. Akan tetapi, kombinasi suplementasi glisina, taurina, lisina dan metionina cenderung meningkatkan kandungan L-karnitin daging ayam broiler umur 21 dan 42 hari. Umur ayam broiler berpengaruh terhadap respon suplementasi asam amino pada pakan ayam broiler tinggi lemak.
Saran
Suplementasi asam amino pada pakan ayam broiler tinggi lemak untuk meningkatkan produksi garam empedu, koefisien kecernaan semu dan L-karnitin daging disarankan pada periode starter. Akan tetapi, suplementasi asam amino dapat menyebabkan asam amino pakan berlebihan dan tidak seimbang yang memberikan pengaruh tidak baik pada performa ayam broiler. Oleh karena itu, suplementasi asam amino pada pakan juga disarankan memperhatikan ketersediaan dan keseimbangan asam amino.

54
DAFTAR PUSTAKA
Abudabos A, Aljumaah R. 2010. Evaluation of digestible lysine needs for male broiler. Int J Poult Science. 9(12):1146-1151.
Adabi SG, Moghaddam G, Taghizadeh A, Namatollahi A, Farahvash T. 2006. Effect of L-carnitine and vegetable fat on broiler breeder fertility, hatchability, egg yolk and serum cholesterol and triglyceride. Int J Poult Science. 5(10): 970-974.
Adeyemo GO, Ologhobo AD, Adebiyi OA. 2010. Effect of graded levels of dietary methionine on the haematology and serum biochemistry of broilers. Int
J Poult Science. 9(2):158-161. Adeyemo GO. 2012. Performance broiler starters fed varying levels of dietary
methionine. Int J AgriScience. 2(2):143-148. Adeyemo IA, Sani A, Aderibigbe TA. 2013. Growth performance and nutrient
retention of broiler chickens fed aspergillus niger hydrolysed cassava peel based diet. Am J Res Communication. 1(7):294-306.
Adrizal, Ohtani S, Yayota M. 2002. Dietary energy source and supplements in broiler diets containing defatted rice bran. J Appl Poult Res.11:410–417.
Aftab U, Ashraf M, Mumtaz AS, Jiang Z. 2007. Lysine requirement of broiler chickens fed low-density diets under tropical conditions. Int J Poult Science. 20(6):939-953.
Ahmed ME, Abbas TE. 2011. Effects of dietary levels of methionine on broiler performance and carcass characteristics. Int J Poult Science. 10(2):145-151.
Al-Marzooqi WS. 1998. Use of Supplemental lipase enzyme and detergent to improve fat digestion in poultry [thesis]. Guelph (Canada). The University of Guelph.
Alzawqari M, Kermanshahi H, Moghaddam HN. 2010. The effect of glycine and desiccated ox bile supplementation on performance, fat digestibility, blood chemistry and ileal digesta viscosity of broiler chickens. Glob Vet. 5(3):187-194.
Arslan C. 2006. L-Carnitine and its use as a feed additive in poultry feeding a review. Revue Méd Vét. 157(3):134-142.
[AOAC] Association of Official Analytical Chemists. 1995. Official Methods of
Analysis. 16th
ed. Virginia (US). Arlington. Bae SH, Kim JH, Shin IS, Han IK. 1999. Partition of amino acid requirements of
broilers between maintenance and growth. IV. Threonine and glycine. Asian-
Aust J Anim Science. 12(3): 381-387. Baião NC, Lara LJC. 2005. Oil and fat in broiler nutrition [review]. Braz J Poult
Science. 7(3):129–141. Baker DH. 2003. Ideal amino acid patterns for broiler chicks. Pages 223–235 in
Amino Acids in Animal Nutrition. 2nd
ed. Edited by D’Mello JPF. Wallingford (GB). CAB Int Publ.
Barteczko J, Lasek O. 2008. Effect of varied protein an energy contents in mixture on meat quality of broiler chicken. Slovak J Anim Science. 41(4):173-178.
Bernal LEP, Tavernari FDC, Lelis GR, Rostagno HS, Albino LFT. 2012. Nutritiona requirement of digestible lysine for Cobb 500 broiler chickens [supplement]. W Poult Conv, 5-9 August, Salvador. Brazil.

55
Bouckenooghe T, Remacle C, Reusens B. 2006. "Is taurin a functional nutrien?" Curr Opin Clin Nutr. 9 : 728–733.
Bouyeh M, Gevorgyan OK. 2011. Influence of excess lysine and methionine on cholesterol, fat and performance of broiler chicks. J Anim Vet Adv.
10(12):1546-1550. Braake FHT. Schierbeek K. Groof AD, Vermes M, Longini G, Buonocore,
Goudoever JBV. 2008. Glutathione synthesis rates after amino acid administration directly after birth in preterm infants. Am J Clin Nutr. 88:333-339.
Burdakov D, Liss B, Ashcroft FM. 2003. Orexin excites GABA-ergic neuron of the arcuate nucleus by activating the sodium-calcium exchanger. J Neurosci 25:4951-4957.
Café MB, Waldroup PW. 2006. Interactions between levels of methionine and lysine in broiler diets changed at typical industry intervals. Int J Poult Science. 5(11):1008-1015
Cengiz Ö, Önol AG, Sevim Ö, Öztürk M, Sari M, Daskiran M. 2008. Influence of excessive lysine and/or methionine supplementation on growth performance and carcass traits in broiler chicks. Rev Méd Vét. 159(4):230-236.
Cheeke PR, Dierenfeld E. 2010. Comparative Animal Nutrition and Metabolism. Oxfordshire (GB). CAB Int.
Corzo A, Kidd MT, Burnham DJ, Kerr BJ. 2004. Dietary glycine needs of broiler chicks. Poult Science. 83:1382-1384.
Corzo A, Kidd MT, Dozier WA III, Kerr BJ. 2009. Dietary glycine and threonine interactive effects in broilers. J Appl Poult Res. 18:79-84. doi:10.3382/japr. 2008-00078.
Corzo A. 2012. Determination of the arginine, tryptophan, and glycine ideal-protein ratios in haigh-yield broiler chicks. Appl Poult Res. 21:79-87.doi:10.3382/japr.2011-00362.
Crespo N, Esteve-Garcia E. 2001. Dietary fatty acid profile modifies abdominal fat deposition in broiler chickens. Poult Science. 80: 71–78.
Dahiya JP, Hoehler D, Wilkie DC, Kessel AGV, Drew MD. 2005. Dietary glycine concentration affects intestinal Clostidium perfingens and Lactobacilli populations in broiler chickens. Poult Science. 84:1875-1885.
Dean WF, Scott HM. 1965. The development of an amino acid reference diet for the early growth of chicks. Poult Science. 44:803.
Dean DW, Bidner TD, Southern LL. 2006. Glycine supplementation to low crude protein, amino acid-supplemented diets supports optimal performance of broiler chicks. Poult Science. 85:288-296.
[EFSA] European Food Safety Authority. 2012. Scientific opinion on the safety and efficacy of taurine as a feed additive for animal species. EFSA Journ.
10(6):2736. Eklund M, Bauer E, Wamatu J, Mosenthin R. 2005. Potential nutritional and
physiological functions of betaine in livestock. Nutr Res Rev. 18:31-48. Fillios LC, Mann GV. 1954. Influence of sulfur amino acid deficiency on
cholesterol metabolism. Metabolism. 3:16. Firman JD, Kamyab A, Leigh H. 2008. Comparison of fat sources in rations of
broilers from hatch to market . Int J Poult Science. 7 (12): 1152-1155.

56
Freitas ER, Sakomura NK, Neme R, Santos ALD. 2002. Valores de energia metabolizável do óleo ácido de soja para aves. In: Anais da 39º Reunião Anual da Sociedade Brasileira de Zootecnia.
Gaya LG, Mourão GB, Rezende FM, Mattos EC, Filho TM, Figueiredo LGG, Ferraz JBS, Eler JP. 2005. Genetic trends of abdominal fat content in a male broiler chicken line. Genet Mol Res. 4(4):760-765.
Gibson DM, Harris RA. 2002. Metabolic Regulation in Mammals. London (GB). Taylor & Francis.
Goulart CDC, Costa FGP, Silva JHVD, Souza JGD, Rodrigues VP, Oliveira CFSD. 2011. Requirements of digestible methionine + cystine for broiler chickens at 1 to 42 days of age. R Bras Zootec. 40(4):797-803.
Gong LM, Lai CH, Qiao SY, Li D, Ma YX, Liu YL. 2005. Growth performance, carcass characteristics, nutrient digestibility and serum biochemical parameters of broiler fed low-protein diets supplemented with various ratios of threonine to lysine. Asian-Aust J Anim Science. 18(8):1164-1170.
Gregory S. Kelly ND. 1998. L-carnitine: Therapeutic applications of a conditionally-essential amino acid. Alter Med Rev. 3(5):345-360.
Green J. Kellogg TF. 1987. Bile acid concentrations in serum, bile, jejuna contens and excreta of male broiler chicks during the first six weeks post hatched. Poult
Science. 66:535-540. Greenwood RH, Titgemeyer EC, Stokka GL, Droillard JS, Löest CA. 2001.
Effects of L-carnitine on nitrogen retention and blood metabolites of growing steers and performance of finishing steers. J Anim Science. 79:254-260.
Grisoni ML, Uzu G, Larbier M, Geraert PA. 1991. Effect of dietary lysine level on lipogenesis in broilers. Reprod Nutr Dev. 31:683-690.
Gropper SS, Smith JL, Groff JL. 2009. Advanced Nutrition and Human
Metabolism. 5th ed. Wadsworth (US). Cengage Learning.
Gurr MI. 1992. Role of Fat in Food and Nutrition. 2nd
ed. New York (US). Elsevier Science Publishers.
Han YK, Thacker PA. 2011. Influence of energy level and glycine supplementation on performance, nutrient digestibility and egg quality in laying hens. Asian-Aust J Anim Science. 24(10):1447-1455.
Hakimi AY. 1992. Studies of manajerial variables on broiler performance and abdominal fat levels [disertation]. Oregon State University.
Harmeyer J. 2002. The physiological role of L-carnitine. Lohmann Inf. 27:1-8. Haro C V. 2005. Interaction between dietary polyunsaturated fatty acids and
vitamin E in body lipid composition and &-tocopherol content of broiler chickens. [disertation]. Barcelona (ES). Universitat Autonoma de Barcelona.
Horniakova E, Abas KA. 2009. Influence of low levels of protein and sex on carcass traits and nutrient content in broiler meats. Slovak J Anim Science. 42(2): 75-78.
Huyghebaert G, Park M. 1996. Effects of dietary protein content addition of nonessential aminoacids and dietary methionine to cysteine balance on responses to dietary sulphur-containing amino acids in broilers. Br Poult
Science. 37:623–639. Jacob JP. 1989. The influence of diet composition and tissue taurine content on
the incidene of suddent death syndrome in male broiler chickens [thesis]. Vancouver (Canada). The University of British Columbia.

57
Jacobsen JG, Smith LH. 1968. Biochemistry and physiology of taurine and taurine derivatives. Physiol Rev. 48:424-511.
Ji B. 2013. Pathway profiling identifies mechanisms of adipose deposition in domestic chickens [disertation]. Knoxville. The University of Tennessee.
Jones GPD, Farrell DJ. 1987. Reducing broiler fat in broiler : preliminary result [Internet]. [diunduh 2014 Maret 17]. Tersedia pada : http://www. Livestocklibrary.com.au/handle/1234/19524.
Kerr BJ, Kidd MT, Halpin KM, McWard GW, Quarles CL. 1999. Lysine level increases live performance and breast yield in male broilers. J Appl Poult Res. 8:381-390.
Kessler AM, Lubisco DS, Vieira MM, Ribeiro AML, Penz AM Jr. 2009. Fatty-acid composition of free-choice starter broiler diets. Braz J Poult Science.
11(1):31–38. Khalifeh-Gholi M, Jahanian R. 2012. Effect of dietary arginine by methionine
interaction on performance and some blood biochemical parameters in broiler chicks [supplement]. W Poult Conv, 5-9 August, Salvador. Bazil. Poster presentation.
Kidd MT, Kerr BJ, Halpin KM, McWard GW, Quarles CL. 1998. Lysine levels in starter and grower-finisher diets affect broiler performance and carcass traits. J
Appl Poult Res. 7:351-358. Kiraz S, %engül T. 2005. Relationship between abdominal fat and methionine
deficiency in broilers. Czech J Anim Science. 50(8):362–368. Klansing KC. 2006. Comparative Avian Nutrition. Oxfordshire (GB). CAB Int. Kraj�ovi�ova-Kudlá�ková M, Šimon�i� M, Béderová A, Babinská K, Béder I.
2000. Correlation of methionin level to methionine and lysine intake. Physiol
Res. 49:399-402. Krähenbuhl S. 1996. Carnitine metabolism in chronic liver disease. Life Science
59: 1579-1599. Krogdahl A. 1985. Digestion and absorption of lipids in poultry. J Nutrition.
115:675-685. Kussaibati R, Leclercq B. 1984. Effect of sorbitol and added bile salts on food
utilization and morphological changes in the liver, gall-blader and caeca of young chicks. Ann Zootech. 33(1):51-58.
Lambert IH. 2004. Regulation of the cellular content of the organic osmolyte taurine in mammalian cells. Neurochem Res. 29:27–63.
Larbier M, Leclerq B. 1994. Nutrition and Feeding of Poultry. Edited and translated by Wiseman J. Nottingham (GB). Nottingham Univ Pr.
Leclercq B. 1998. Spesific effects of lysine on broiler production: comparison with threonine and valine. Poult Science. 77:118-123.
Lee DN, Cheng YH, Chuang YS, Shive JL, Lian YM, Wei HW, Weng CF. 2004. Effects of dietary taurine supplementation on growth performance, serum constituents and antibody production of broilers. Asian-Aust J Anim Science.
17(1):109-115. Lee SM, Lim HS, Kim WY, Paik IK. 2004. The effects of dietary
supplementation of feather meal digests on the performances and muscular taurine contents in broiler chickens. J Anim Science Technol. 46(5):753-752.
Leeson S, Summers JD. 2001. Nutrition of The Chicken. 4th
ed. Guelph (Canada). Univ Books.

58
Leeson S, Summers JD. 2005. Commercial Poultry Nutrition. 3rd
ed. Nottingham (GB). Nottingham Univ Pr.
Lilly RA. 2010. The effects of dietary amino acid density in broiler feed on carcass characteristics and meat quality [thesis]. Mississippi. Mississippi State Univ.
Liz K. 2013. Digestion [Internet]. [diunduh 2014 Pebruari 14]. Tersedia pada : http://www.studyblue.com/notes/note/n/digestion/deck/6199485.
Malomo GA, Bolu SA, Olutade SG, Suleiman ZG. 2013. Effects of feeding low protein diets with methionine and lysine supplementation on the performance and nitrogen economy of broilers. Res Opin Anim Vet Science. 3(9):330-334.
Märt. 2013. Emulsifiers [Internet]. [diunduh pada 2014 Pebruari 18]. Tersedia pada : http://sciencevaria.blogspot.com/2013_02_01_archive.html.
Martin WG, Miraglia RJ, Spaeth DG, Patrick H. 1966. Synthesis of taurin from sulfate by the chick. J Biol Med 122:841-844.
Ma�lanko W, Pisarski RK. 2009. The effect of herbs on the share of abdominal fat and its fatty acid profile in broiler chickens. Sectio EE. 27(3):28-34. Doi: 10.2478/v10083-009-0013-y.
Mathews CK, Holde KEV, Kevin GA. 2000. Biochemistry. 3rd
Ed. San Francisco (US). Benjamin Cummings.
Matin HRH, Dashtbin F, Salari J. 2013. Absorption and macromineral interactions in broiler production: an overview. Glob Veterinaria. 11(1):49-54.
McGill E, Kamyab A, Firman JD. 2012. Low crude protein corn and soybean meal diet with amino acid supplementation for broilers in the starter period 1. Effects of feeding 15% crude protein. Int J Poult Science. 11(3):161-165.
Mehta S. 2013. Activation and transportation of fatty acids to the mitochondria via the carnitine shuttle with animation [Internet]. [diunduh 2014 Pebruari 18]. Tersedia pada : http://pharmaxchange.info/press/2013/10/activation-and-transportation-of-fatty-acids-to-the-mitochondria-via-the-carnitine-shuttle-with-animation.
Melo JE, Motter MM, Morao LR, Huguet MJ, Canet Z, Miquel MC. 2003. Use of in-vivo measurements to estimate breast and abdominal fat content of a free-range broiler strain. Anim Science. 77: 23-31.
Mozafari O, Ghazi S, Moeini MM. 2013. The effects of different levels of edible potato (Solanum tubresum) replacing maize on performance, serum metabolite and immune system of broiler chicks. Iran J Appl Anim Science. 3(3): 583-588
Mushtaq MMH, Pasha TN, Saima, Akram M, Mushtaq T, Parvin R, Farooq U, Mehmood S, Iqbal KJ, Hwangbo J. 2013. Growth performance, carcass traits and serum mineral chemistry as affected by dietary sodium and sodium salts fed to broiler chickens reared under phase feeding system. Asian-Aust J Anim
Science. 26(12):1742-1752.doi:10.5713/ ajas.2013.13266. Namroud NF, Shivazad M, Zaghari M, Shahneh AZ. 2010. Effect of glycine and
glutamic acid to low protein diets on performance, thyroid function and fat deposition in chickens. South Afr J Anim Science. 40 (3):238-244.
Nascimento AH. 2004. Deposição de proteína na carcaça de frangos de corte. Av
world. 2:2-3. [NRC] National Research Council. 1994. Nutrien Requirements of Poultry. 9th
Ed. Washington DC (US). National Ac Pr.

59
Nosrati M, Qutbi A, Navidshad B, Mirhoseini Z, Sayadi AJ, Royan M. 2006. Effects skip day feeding and dietary fat type on abdominal fat pad and blood lipids in broiler chickens. Pakist J Biol Science. 9(15):2748:2752.
Ohmura E, Ishikawa T, Takagi M, Aoyama Y, Yoshida A. 1998. Effects of dietary methionine, taurine, cysteine, and glycine on liver and serum lipids in rats fed with a histidine-excess diet. Agric Biol Chem. 52(4):1027-1032.
Ojano-Dirain CP, Waldroup PW. 2002. Evaluation of lysine, methionine and threonine needs of broilers three to six week of age under moderate temperature stress. Int J Poult Science. 1(1):16-21.
Onu PN, Ayo-Enwerm MC, Ahaotu EO. 2010. Evaluation of carcass characteristics and cost effectiveness of broiler chicks fed synthetic lysine and methionine supplemented soybean-based diets. Int J Science Nat. 1(1):22-26.
Ortiz LT, Alzueta C, Rebolé A, Rodriíguez ML, Arija I, Brenes A. 2006. Effect of dietary high-oleic acid and conventional sunflower seeds and their refined oils on fatty acid composition of adipose tissue and meat in broiler chickens. J
Anim Feed Science. 15:83–95. Ospina-Rojas IC, Murakami AE, Guerra AFQG. 2013. Effect of glycine
supplementation on performance, development of mucosa and nutrient utilization of broiler chickens fed low-protein grower diets with different levels of threonine [abstract]. Poult Science. 92(12):112.
Osti NP, Pandey SB. 2004. Methionine and lysine supplementation in low quality feed ingredient based diets on performance of broiler chicken. Fourth national conference on science and technology; 2014 March 23–26; Kathmandu, Nepal. Royal Nepal Academy of Science and Technology (RONAST). 1-8 p.
Panja P, Kassim H, Jalaludin S. 1995. Effects of palm oil and soybean oil as fat sources in isonitrogenous and isocaloric diets on the performance of broilers. Asian-Aust J Anim Science. 8(3):223-229.
Park IS, Kang YH, Kang JS. 2007. Effects of taurine on plasma and liver lipids, erythrocyte ouabain sensitive Na efflux and platelet aggregation in Sparague Dawley rats. Nut Res Pract. 1(3):200-205.
Pacific Biomarkers. 2013. Bile acids: beyond cholesterol metabolism & fat absorption [Internet]. [diunduh pada 2014 Pebruari 2]. Tersedia pada : http:// www.pacbio.com/publications/bile-acid-whitepaper.php
Pesti GM, Bakalli RI, Qiao M, Sterling KG. 2002. A comparison of eight grades of fat as broiler feed ingredients. Poult Science. 81:382–390.
Pesti GM, Bakalli RI, Driver JP, Atencio A, Foster EH. 2005. Poultry Nutrition
and Feeding. Victoria (Canada). Trafford Pub. Poorghasemi M, Seidavi A, Qotbi AAA, Laudadio V, Tufarelli V. 2013. Influence
of dietary fat source on growth performance responses and carcass traits of broiler chicks. Asian-Aust J Anim Science. 26(5):705-710.
Powell S, Bidner TD, Southern LL. 2009. The interactive effects of glycine, total sulfur amino acids, and lysine supplementation to corn-soybean meal diets on growth performance and serum uric acid and urea concentrations in broilers. Poult Sc. 88:1407-1412. doi:10.3382/ps.2008-00433.
Prosser CL. 1991. Environmental and Metabolic Animal Physiology Comparative
Animal Physiology. Illinois. John Wiley.

60
Ros E. 2000. Intestinal absorbtion of triglyceride and cholesterol. Dietary and pharmacological inhibition to reduce cardiovascular risk [review]. Atherosclerosis. 151:357–379.
Ruiz-Feria CA, Berrs KW, Kidd MT, Wideman RF Jr. 1999. Plasma taurine levels in broilers with pulmonary hypertension syndrome induced by unilateral pulmonary artery occlusion. Poult Science. 78:1627-1633.
Sachan DS, Rhew TH, Ruark RA. 1984. Ameliorating effects of carnitine and its precursors on alcohol-induced fatty acid. Am J Clin Nutr. 39:738-744.
Sanz M, Lopez-Bote CJ, Menoyo D, Bautista JM. 2000. Abdominal fat Deposition and Fatty Acid Synthesis Are Lower and b-Oxidation Is Higher in Broiler Chickens Fed Diets Containing Unsaturated Rather than Saturated Fat. J Nutr. 130:3034–3037.
Scholte HR. 2003. Carnitine [Internet]. [diunduh 2014 Pebruari 2014]. Tersedia pada : http://www.chm.bris.ac.uk/motm/carnitine/Carnitine.htm.
Selle PH, Ravindran V, Ravindran G, Bryden WL. 2007. Effects of dietary lysine and microbial phytase on growth performance and nutrient utilisation of broiler chickens. Asian-Aust J Anim Science. 20(7):1100-1107.
Senthilkumar R, Viswanathan P, Nalini N. 2003. Glycine modulates hepatic lipid accumulation in alcohol-induced liver injury. Pol J Pharmacol. 55:603-611.
Shahriar HA, Rezaei A, Lak A, Ahmadzadeh A. 2007. Effect of dietary fat sources on blood and tissue biochemical factors of broiler. J Anim Vet Adv. 6:1304-1307.
Shahriari A, Tabatabaie RF, Jafari RA, Ghorbanzadeh B. 2009. Modulation of serum and liver triglyceride and abdominal fat pad weight by dietary garlic in male broilers. Int J Vet Res. 3(2) :101-105.
Shim KS, Jung HJ, Na CS, Yoon C, Park GH. 2009. Effects of taurine on lipid metabolism and protein synthesis in poultry and mice. Asian-Aust J Anim
Science. 22(6):865-870. Si J, Fritts CAD, Burnham J, Waldroup PW. 2001. Relationship of Dietary Lisin
level to the concentration of all essential amino acids in broiler diets. Poult
Science. 80: 1472-1479. Siqueira JC, Sakomura NK, Dourado LRB, Ezequiel JMB, Barbosa NAA,
Fernandes JBK. 2013. Diet formulation techniques and lysine requirements of 1-to 22-day-old broilers. Braz J Poult Science. 15(2):123-134.
Smits CHM, Veldman A, Verkade HJ, Beynen AC. 1998. The inhibitory effect of carboxymethylcellulose with high viscosity on lipid absorption in broiler chickens coincides with reduced bile salt concentration and raised microbial numbers in the small intestine. Poult Science. 77:1534–1539.
Sohail SS, Bryant M, Roland DA Sr. 2003. The effect of glycine supplementation on performance of broilers fed sub-marginal protein with adequate synthetic methionine and lysine. Int J Poult Science. 2(6):394-397.
Spitze AR, Wong DL, Rogers QR, Fascetti AJ. 2003. Taurine concentrations in animal feed ingredients; cooking influences taurine content. J Anim Physiol
Anim Nutr. 87:251–262. Stamp D, Jenkins G. 2008. An overview of bile acid synthesis, chemistry and
function. Pages 1-13 In: Bile acids: Toxicology and Bioactivity (Ed. G. Jenkins and L. Hardie). London (GB). Royal Society of Chemistry.

61
Steel RGD, Torrie JH. 1994. Principles and Procedures of Statistics. London (GB). Mc Graw-Hill Int Book Com.
Stevens L. 1996. Avian Biochemistry and Molecular Biology. New York (US). Cambridge University Press.
Sturman JA, Hayes KC. 1980. The biology of taurine in nutrition and development. Adv Nutr Res. 3:231–299.
Suarez EC, Marie DR, Salazar MT, Lopez MGN, David AVC, Ortega AC. 2006. Taurin: An Amino Acid Rich in Fish Meal. Advances en nutricion acuicola VIII. VIII Simposium Inernacional de Nutricion Acuicola. 15-17 Noviembre. Universidad Autonoma de Nuevo Leeon, Monterrey, Nuevo Leon, Mexico.
Sugiyama K, Ohishi, A, Ohnuma Y, Muramatsu K. 1989. Comparison between the plasma cholesterol lowering effects of glycine and taurine in rats fed on high cholesterol diets. Agr Bio Chem. 53(6):1647-1652.
Suida D. 2001. Formulação por proteína ideal e conseqüências técnicas, econômicas e ambientais. In: Simpósio Internacional De Nutrição Animal: Proteína Ideal, Energia Liquida E Modelagem. 27-43, Anais. Santa Maria: Embrapa.
Sun JM, Richards MP, Rosebrough RW, Ashwell CM, McMurtry JP, Coon CN. 2006. The relationship of body composition, feed intake, and metabolic hormones for broiler breeder females. Poult Science. 85:1173-1184.
Tappia PS, Xu YJ, Leyva DR, Aroutiounova N, Dhalla NS. 2012. Cardioprotective effects of cysteine alone or combination with taurine in diabetes. Physiolog Res, forthcoming. Di dalam : Proceedings of the 15th international SHR Symposium/the 48th Scientific Meeting of the Japanese Society for Hypertension-Related Disease Model Research; 2012 September 27- 28, , Melbourne, Australia. Melbourne (AU). Pages 171-178.
Taraz Z, Jalali SMA, Rafeie F. 2006. Effects of replacement of soybean meal with rapeseed meal on organs weight, some blood biochemical parameters and performance of broiler chicks. Int J Poult Science. 5(12):1110-1115.
Tavernari FDC, Buteri CB, Rostagno HS, Albino LFT. 2009. Effects of dietary digestible lysine levels on protein and fat deposition in the carcass of broilers. Braz J Poult Science.11(2):99-107.
Tsuboyama N, Hosokawa YM, Totani J, Oka A, Matsumoto T, Kodama H. 1996. Structural organization and tissue-specific expression of the gene encoding rat cysteine dioxygenase. Gene 181:161–165.
T�mová E, Teimouri A. 2010. Fat deposition in the broiler chicken: a review. Cient Agricult Bohemica. 41(2):121-128.
Ukpabi UH, Amaefule KU, Amaefule OM. 2008. Performance of broilers fed raw bambarra groundnut Vigna subterranean (L.) offal diets supplemented with lysine and or methionine. Int J Poult Science. 7(12):1177-1181.
Ullah MS, Pasha TN, Ali Z, Saima, Khattak FM, Hayat Z. 2012. Effects of different pre-starter diets on broiler performance, gastro intestinal tract morphometry and carcass yield. J Anim Plant Science. 22(3): 570-575.
Vieira SL, Lemme A, Goldenberg DB, Brugalli I. 2004. Responses of growing broilers to diets with increased sulfur amino acids to lysine ratios at two dietary protein levels. Poult Science. 83:1307:1313.

62
Waldroup PW, Jiang Q, Fritts CA. 2005. Effect of glycine and threonine supplementation on performance of broiler chicks fed diets low in crude protein. Int J Poult Science. 4(5):250-257.
Walter P. 2000. L-karnitin-a vitamin-like substance for functional food. Ann Nutr
Metab. 44:75-96. Wardah, Sopandi T, Askono H EB, Kusriningrum. 2012. Reduction of
intracellular lipid accumulation, serum leptin, and cholesterol levels in broiler fed diet supplemented with powder leaves of Phyllanthus buxilofilus. Asian J
Agricult Res. 1-12. Doi:10.3923/ajar.2012. Wongsuthavas S. 2007. Dietary fat type, body composition and fatty acid
metabolism in broiler chickens [disertation]. Utrecht (ND). Utrecht University. Xu ZR, Wang MQ, Mao HX, Zhan XA, Hu CH. 2003. Effects of L-karnitin on
growth performance, carcass composition and metabolism of lipids in male broilers. Poult Science. 82: 408-413.
Yamamoto K, Yoshitama A, Sakono M, Nasu T, Murakami S, Fukuda N. 2000. Dietary taurine decreases hepatic secretion of cholesterol ester in rats fed a high-cholesterol diet. Pharmacology 60:27-33.
Zeng D, Gao Z, Zhao J, Huang XL, Duo L, Tian Y. 2009. Effects of taurine on growth performance, immune organ development and antioxidative ability of broilers. Chin J Vet Science. 29 : 774-778.
Zeweil HS, Abdallah AA, Basyony M. 2011. Effect of taurin supplementation on productive performance, carcass quality, immune organs development and blood constituents of broiler chicks. Egypt Poult Science. 31(4):681-694.

63
LAMPIRAN
Lampiran 1 Prosedur analisis L-karnitin dengan Metode HPLC
Prosedur analisis L-karnitin adalah sebagai berikut :
Preparasi standar Standar L-karnitin ditimbang 25 mg dalam labu ukur 25 ml (standar 1000
mg l-1). H2O ditambahkan sampai volume larutan 25 ml dan homogen. Larutan standar dipipet 1 ml kedalam labu ukur 10 ml, kemudian ditambahkan H2O sampai volumenya 10 ml dan homogen (standar 100 mg l-1). Larutan standar 1000 mg l-1 dipipet 100 dan 500 µl dan larutan standar 100 mg l-1 dipipet 50, 100, dan 500 µl kedalam labu ukur 10 ml. H2O ditambahkan sampai volume 10 ml dan homogen (standar 50, 10, 5, 1 dan 0.5 mg l-1).
Dialisis sampel
20 ml sampel dimasukkan dalam membran, kemudian membran ditutup. Membran didialisis dengan 500 ml Larutan Krebs-Ringer fosfat selama 36 jam pada suhu 4 oC. Larutan buffer diganti sebanyak tiga kali dengan interval 8 sampai 10 jam. Sampel hasil dialilisis siap digunakan.
Pengendapan plasma protein dan prosedur ekstraksi sampel dan standar
Larutan internal standar (20 µg ml-1) dipipet kedalam tabung, kemudian ditambahkan 300 ul sampel dan standar yang mengandung L-karnitin. Larutan kemudian ditambahkan 0.3 M Ba(OH)2 dan ZnSO4 10% sebanyak 300 µl. Larutan divorteks dengan kecepatan tinggi selama satu menit, kemudian larutan disentrifus dan diambil supernatannya. Supernatan divorteks kembali selama 2 menit, kemudian disentrifus dengan kecepatan 10000 rpm selama 10 menit. Supernatan dipisahkan dari endapan dan digunakan untuk derivatisasi sampel.
Derivatisasi sampel dan standar
Supernatan dipipet 500 µl kemudian ditambahkan 0.01 M NaH2PO4.H2O (pH 3.5) sebanyak 500 µl. Larutan kemudian ditambahkan 0.3 M Ba(OH)2 sebanyak 300 µl, ditambahkan HCl 1 M sebanyak 20 µl, dan ditambahkan larutan laminoantracene dan EDAC masing-masing sebanyak 100 µl. Larutan kemudian divorteks dan diinkubasi selama 20 menit pada suhu 25 oC. Kelebihan reagen dicuci dengan menambahkan 5 ml dietilether. Aliquat (fase air) dipipet 300 µl kedalam tabung kemudian ditambahkan 0.01 M NaH2PO4.H2O sampai pH 9.1. Pipet kembali aliquat sebanyak 500 µl kedalam tabung, kemudian diencerkan dengan 0.01 M NaH2PO4.H2O (pH 3.5) sebanyak 500 µl. Larutan kemudian divorteks dan disaring dengan milipore 0.45 µm. Larutan diinjeksikan ke HPLC.
Kondisi HPLC
HPLC yang digunakan merupakan HPLC tipe Waters Alliance E2695 dan detektor UV-VIS 2489. Tipe kolom yang digunakan adalah C18, 150 x 4.6 dengan ukuran partikel 5 µm. Detektor yang digunakan FLD dengan panjang gelombang eksitasi 248 nm dan panjang gelombang emisi 418 nm. Fase Gerak yang digunakan 700 ml 0.1 M Ammonium acetat dalam pH 3.5 ditambah 300 ml
64
acetonitril dengan sistem gradien isokratik. Laju alir 1.0 ml menit-1 dengan volume injeksi 20 µl.
Perhitungan kadar L karnitin adalah sebagai berikut :
Kadar L-karnitin (mg/kg) = '������������(�
� �(�����!������(�����)�������*� ���� �"�
"�"������(� ������
Lampiran 2 Analisis sidik ragam dan uji lanjut Duncan performa
Parameter Sumber variasi
Jumlah kuadrat
DB Kuadrat tengah
F hitung Sig
Konsumsi pakan umur 1-21 hari (g)
Perlakuan 9868.81 5 1973.763 6.766 .001
Galat 5251.03 18 291.724
Total 15119.84 23
Konsumsi pakan umur 22-42 hari (g)
Perlakuan 207000.39 5 41400.079 23.938 .000
Galat 31131.06 18 1729.504
Total 238131.46 23
Konsumsi pakan umur 1-42 hari (g)
Perlakuan 266710.87 5 53342.175 36.341 .000
Galat 26420.94 18 1467.830
Total 293131.82 23
Bobot badan umur 21 hari (g)
Perlakuan 14641.62 5 2928.324 12.589 .000
Galat 4186.86 18 232.603
Total 18828.48 23
Bobot badan umur 42 hari (g)
Perlakuan 75158.19 5 15031.637 14.277 .000
Galat 18951.01 18 1052.834
Total 94109.20 23
Pertambahan bobot badan umur 1-21 hari (g)
Perlakuan 14912.21 5 2982.443 13.209 .000
Galat 4064.34 18 225.797
Total 18976.56 23
Pertambahan bobot badan umur 22-42 hari (g)
Perlakuan 48482.45 5 9696.490 7.752 .000
Galat 22514.56 18 1250.809
Total 70997.01 23
Pertambahan bobot badan umur 1-42 hari (g)
Perlakuan 75445.44 5 15089.088 14.582 .000
Galat 18626.16 18 1034.786
Total 94071.60 23
Konversi pakan umur 1-21 hari
Perlakuan .025 5 .005 6.095 .002
Galat .015 18 .001
Total .039 23
Konversi pakan umur 22-42 hari
Perlakuan .025 5 .005 3.024 .037
Galat .030 18 .002
Total .055 23
Konversi pakan umur 1-42 hari
Perlakuan .020 5 .004 8.662 .000
Galat .008 18 .000
Total .029 23
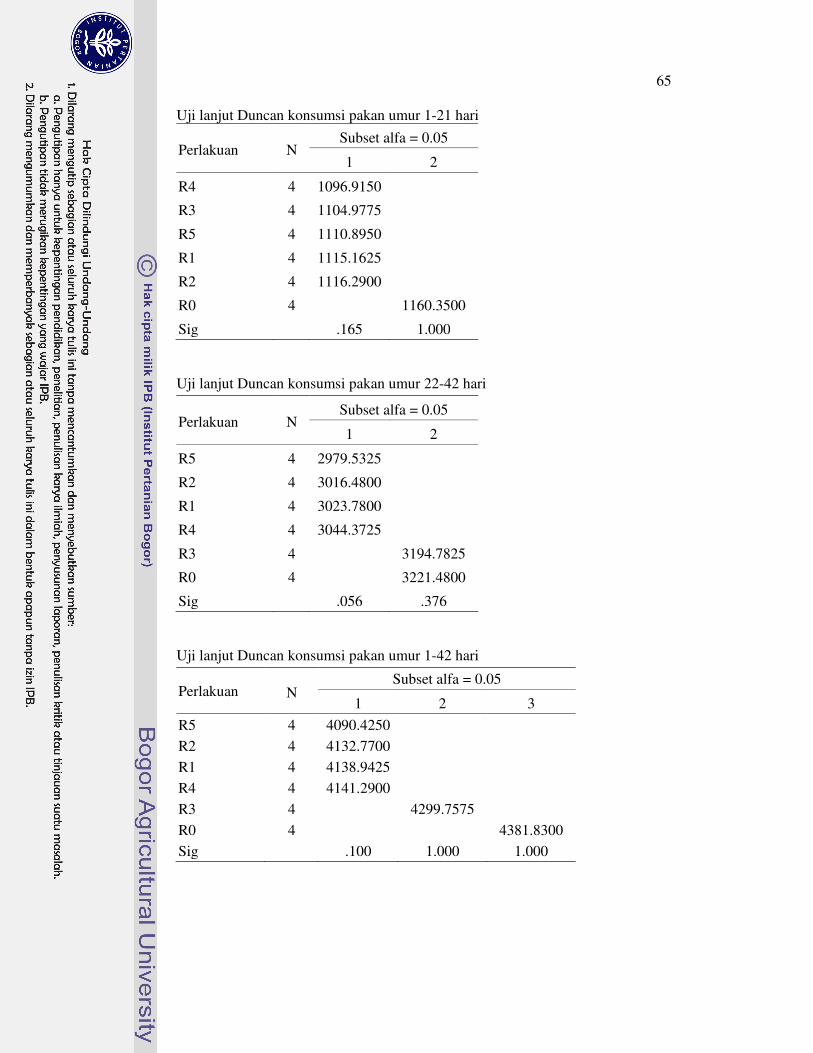
65
Uji lanjut Duncan konsumsi pakan umur 1-21 hari
Perlakuan N Subset alfa = 0.05
1 2
R4 4 1096.9150
R3 4 1104.9775
R5 4 1110.8950
R1 4 1115.1625
R2 4 1116.2900
R0 4 1160.3500
Sig .165 1.000
Uji lanjut Duncan konsumsi pakan umur 22-42 hari
Perlakuan N Subset alfa = 0.05
1 2
R5 4 2979.5325
R2 4 3016.4800
R1 4 3023.7800
R4 4 3044.3725
R3 4 3194.7825
R0 4 3221.4800
Sig .056 .376
Uji lanjut Duncan konsumsi pakan umur 1-42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R5 4 4090.4250
R2 4 4132.7700
R1 4 4138.9425
R4 4 4141.2900
R3 4 4299.7575
R0 4 4381.8300
Sig .100 1.000 1.000
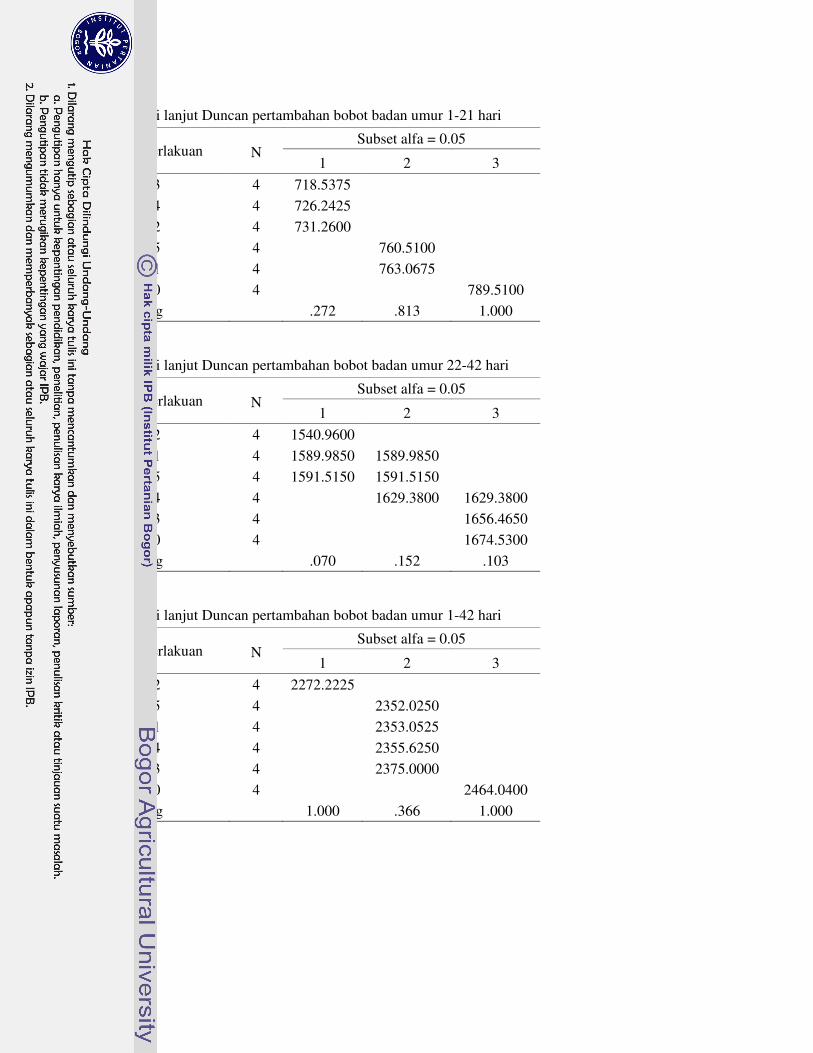
66
Uji lanjut Duncan pertambahan bobot badan umur 1-21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 718.5375
R4 4 726.2425
R2 4 731.2600
R5 4 760.5100
R1 4 763.0675
R0 4 789.5100
Sig .272 .813 1.000 Uji lanjut Duncan pertambahan bobot badan umur 22-42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R2 4 1540.9600
R1 4 1589.9850 1589.9850
R5 4 1591.5150 1591.5150
R4 4 1629.3800 1629.3800
R3 4 1656.4650
R0 4 1674.5300
Sig .070 .152 .103
Uji lanjut Duncan pertambahan bobot badan umur 1-42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R2 4 2272.2225
R5 4 2352.0250
R1 4 2353.0525
R4 4 2355.6250
R3 4 2375.0000
R0 4 2464.0400
Sig 1.000 .366 1.000
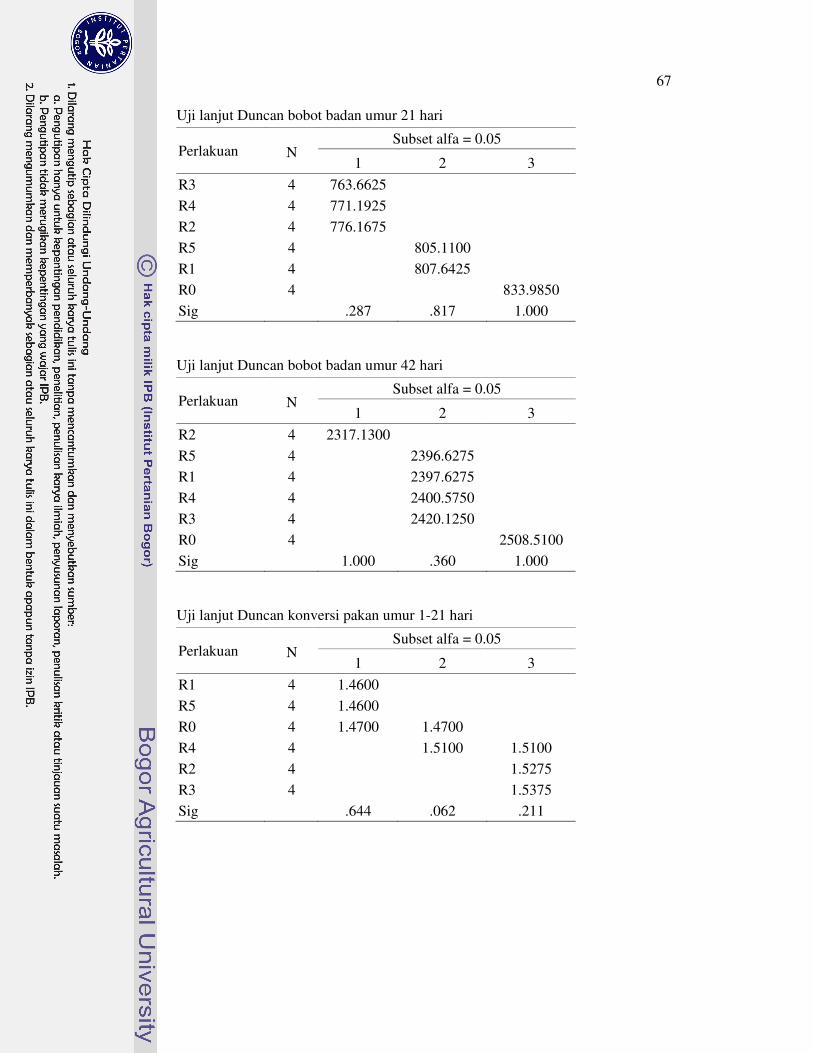
67
Uji lanjut Duncan bobot badan umur 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 763.6625
R4 4 771.1925
R2 4 776.1675
R5 4 805.1100
R1 4 807.6425
R0 4 833.9850
Sig .287 .817 1.000 Uji lanjut Duncan bobot badan umur 42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R2 4 2317.1300
R5 4 2396.6275
R1 4 2397.6275
R4 4 2400.5750
R3 4 2420.1250
R0 4 2508.5100
Sig 1.000 .360 1.000 Uji lanjut Duncan konversi pakan umur 1-21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R1 4 1.4600
R5 4 1.4600
R0 4 1.4700 1.4700
R4 4 1.5100 1.5100
R2 4 1.5275
R3 4 1.5375
Sig .644 .062 .211
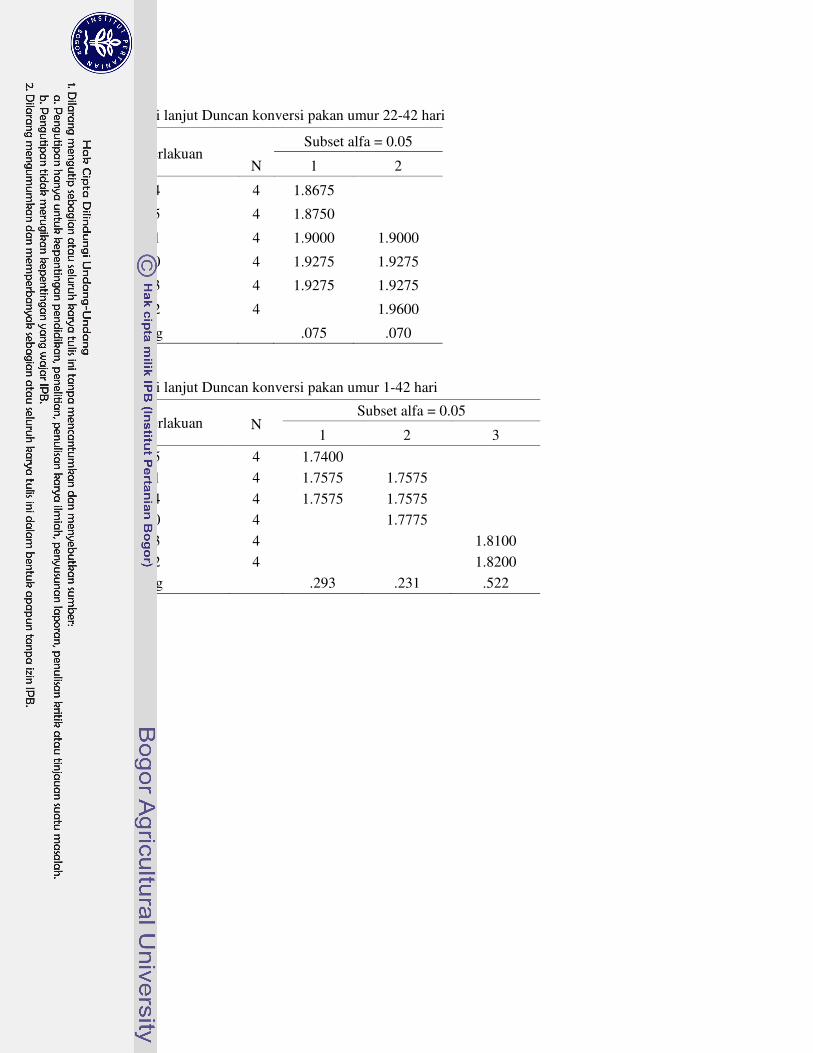
68
Uji lanjut Duncan konversi pakan umur 22-42 hari
Perlakuan Subset alfa = 0.05
N 1 2
R4 4 1.8675
R5 4 1.8750
R1 4 1.9000 1.9000
R0 4 1.9275 1.9275
R3 4 1.9275 1.9275
R2 4 1.9600
Sig .075 .070 Uji lanjut Duncan konversi pakan umur 1-42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R5 4 1.7400
R1 4 1.7575 1.7575
R4 4 1.7575 1.7575
R0 4 1.7775
R3 4 1.8100
R2 4 1.8200
Sig .293 .231 .522
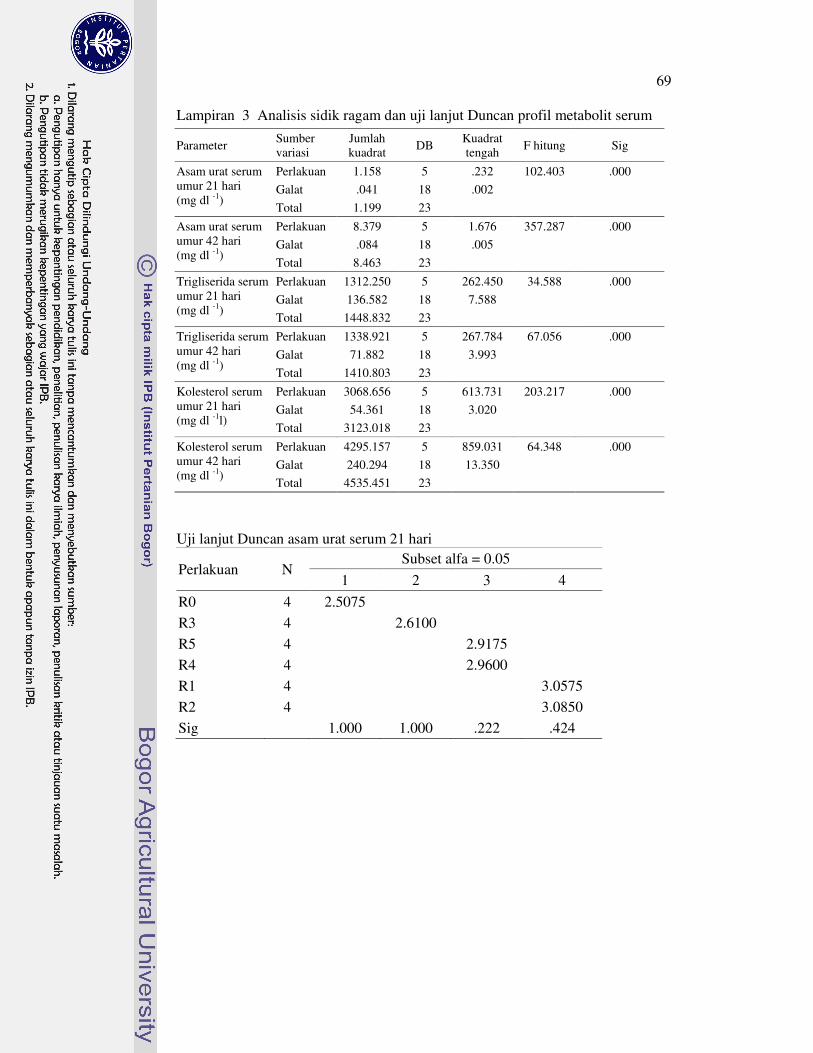
69
Lampiran 3 Analisis sidik ragam dan uji lanjut Duncan profil metabolit serum
Uji lanjut Duncan asam urat serum 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4
R0 4 2.5075
R3 4 2.6100
R5 4 2.9175
R4 4 2.9600
R1 4 3.0575
R2 4 3.0850
Sig 1.000 1.000 .222 .424
Parameter Sumber variasi
Jumlah kuadrat
DB Kuadrat tengah
F hitung Sig
Asam urat serum umur 21 hari (mg dl -1)
Perlakuan 1.158 5 .232 102.403 .000
Galat .041 18 .002
Total 1.199 23
Asam urat serum umur 42 hari (mg dl -1)
Perlakuan 8.379 5 1.676 357.287 .000
Galat .084 18 .005
Total 8.463 23
Trigliserida serum umur 21 hari (mg dl -1)
Perlakuan 1312.250 5 262.450 34.588 .000
Galat 136.582 18 7.588
Total 1448.832 23
Trigliserida serum umur 42 hari (mg dl -1)
Perlakuan 1338.921 5 267.784 67.056 .000
Galat 71.882 18 3.993
Total 1410.803 23
Kolesterol serum umur 21 hari (mg dl -1l)
Perlakuan 3068.656 5 613.731 203.217 .000
Galat 54.361 18 3.020
Total 3123.018 23
Kolesterol serum umur 42 hari (mg dl -1)
Perlakuan 4295.157 5 859.031 64.348 .000
Galat 240.294 18 13.350
Total 4535.451 23
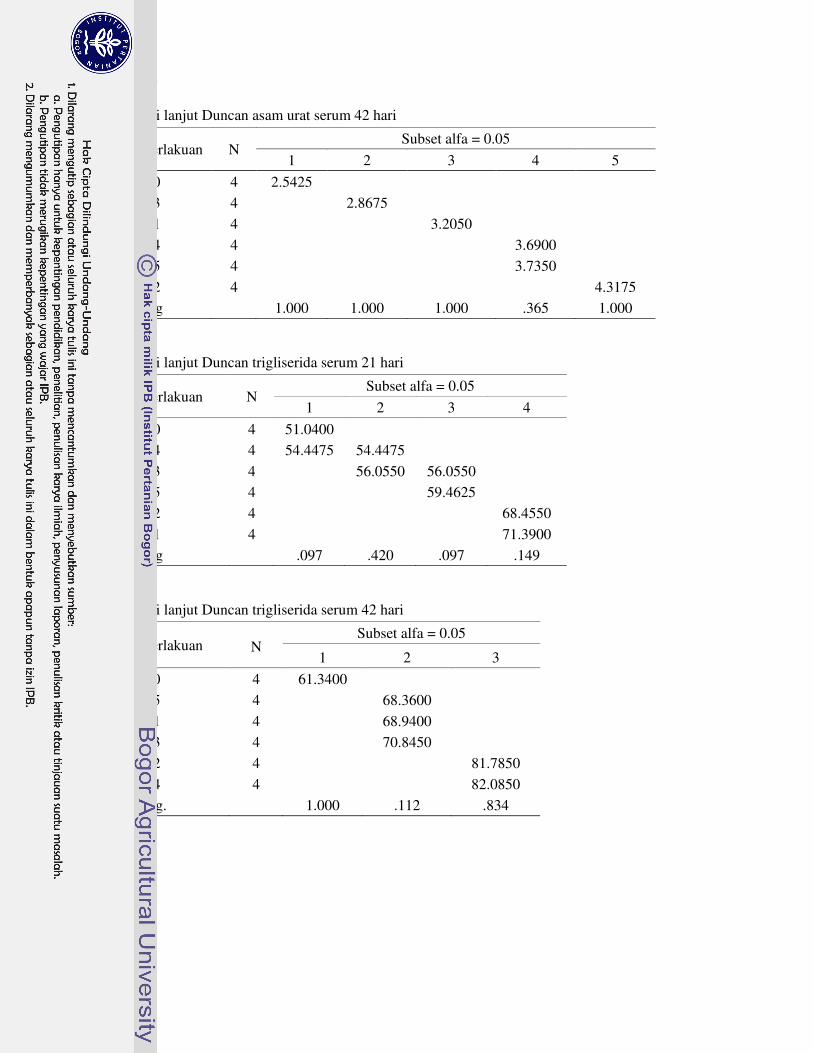
70
Uji lanjut Duncan asam urat serum 42 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4 5
R0 4 2.5425
R3 4 2.8675
R1 4 3.2050
R4 4 3.6900
R5 4 3.7350
R2 4 4.3175
Sig 1.000 1.000 1.000 .365 1.000 Uji lanjut Duncan trigliserida serum 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4
R0 4 51.0400
R4 4 54.4475 54.4475
R3 4 56.0550 56.0550
R5 4 59.4625
R2 4 68.4550
R1 4 71.3900
Sig .097 .420 .097 .149 Uji lanjut Duncan trigliserida serum 42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R0 4 61.3400
R5 4 68.3600
R1 4 68.9400
R3 4 70.8450
R2 4 81.7850
R4 4 82.0850
Sig. 1.000 .112 .834

71
Uji lanjut Duncan kolesterol serum 21 hari
Perlakuan N
Subset alfa = 0.05
1 2 3 4 5 6
R3 4 80.7975
R5 4 88.3825
R1 4 91.4750
R4 4 99.0575
R2 4 107.6300
R0 4 113.8150
Sig 1.000 1.000 1.000 1.000 1.000 1.000 Uji lanjut Duncan kolesterol serum 42 hari
Perlakuan Subset alfa = 0.05
N 1 2 3 4
R3 4 93.4425
R1 4 108.6175
R5 4 110.0175
R2 4 123.5075
R4 4 130.3925
R0 4 130.5325
Sig 1.000 .595 1.000 .957
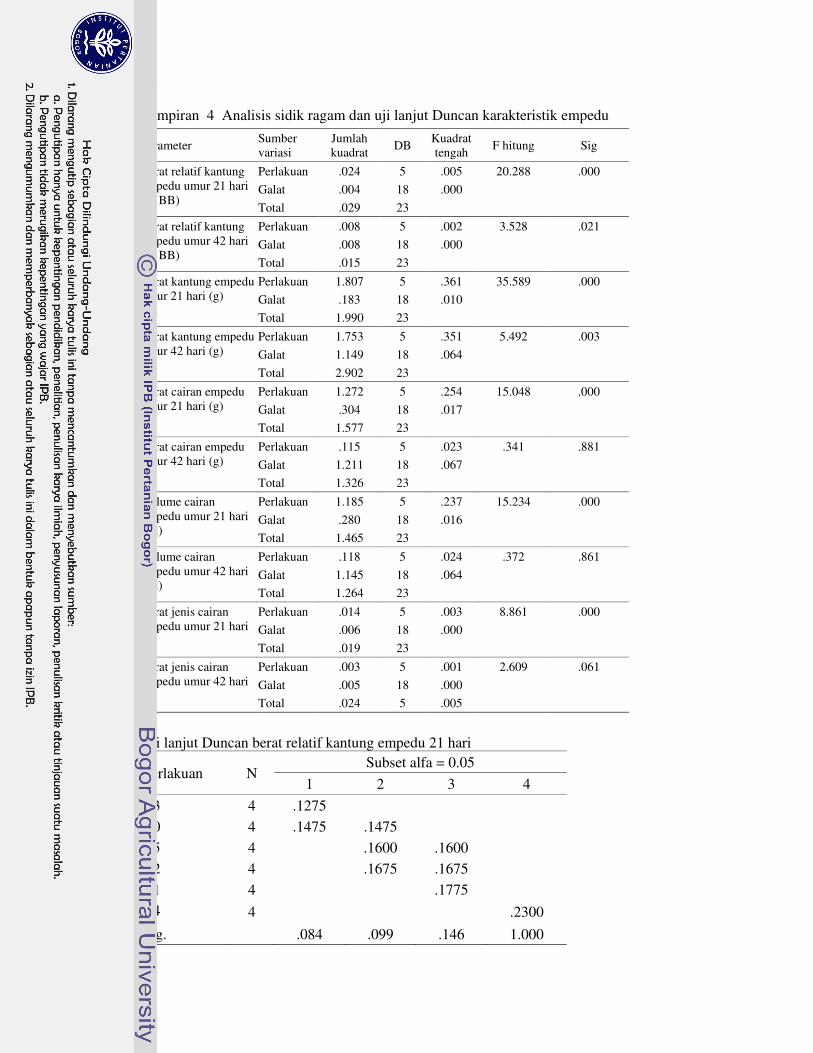
72
Lampiran 4 Analisis sidik ragam dan uji lanjut Duncan karakteristik empedu
Uji lanjut Duncan berat relatif kantung empedu 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4
R3 4 .1275
R0 4 .1475 .1475
R5 4 .1600 .1600
R2 4 .1675 .1675
R1 4 .1775
R4 4 .2300
Sig. .084 .099 .146 1.000
Parameter Sumber variasi
Jumlah kuadrat
DB Kuadrat tengah
F hitung Sig
Berat relatif kantung empedu umur 21 hari (% BB)
Perlakuan .024 5 .005 20.288 .000
Galat .004 18 .000
Total .029 23
Berat relatif kantung empedu umur 42 hari (% BB)
Perlakuan .008 5 .002 3.528 .021
Galat .008 18 .000
Total .015 23
Berat kantung empedu umur 21 hari (g)
Perlakuan 1.807 5 .361 35.589 .000
Galat .183 18 .010
Total 1.990 23
Berat kantung empedu umur 42 hari (g)
Perlakuan 1.753 5 .351 5.492 .003
Galat 1.149 18 .064
Total 2.902 23
Berat cairan empedu umur 21 hari (g)
Perlakuan 1.272 5 .254 15.048 .000
Galat .304 18 .017
Total 1.577 23
Berat cairan empedu umur 42 hari (g)
Perlakuan .115 5 .023 .341 .881
Galat 1.211 18 .067
Total 1.326 23
Volume cairan empedu umur 21 hari (ml)
Perlakuan 1.185 5 .237 15.234 .000
Galat .280 18 .016
Total 1.465 23
Volume cairan empedu umur 42 hari (ml)
Perlakuan .118 5 .024 .372 .861
Galat 1.145 18 .064
Total 1.264 23
Berat jenis cairan empedu umur 21 hari
Perlakuan .014 5 .003 8.861 .000
Galat .006 18 .000
Total .019 23
Berat jenis cairan empedu umur 42 hari
Perlakuan .003 5 .001 2.609 .061
Galat .005 18 .000
Total .024 5 .005
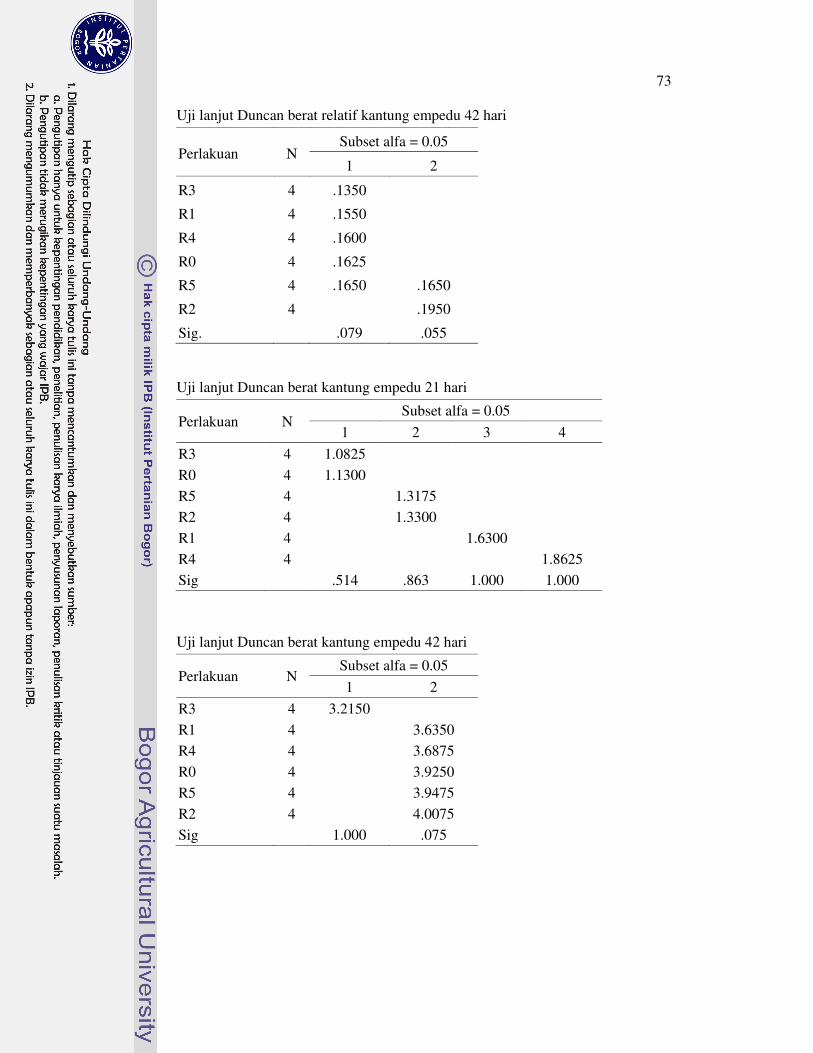
73
Uji lanjut Duncan berat relatif kantung empedu 42 hari
Perlakuan N Subset alfa = 0.05
1 2
R3 4 .1350
R1 4 .1550
R4 4 .1600
R0 4 .1625
R5 4 .1650 .1650
R2 4 .1950
Sig. .079 .055 Uji lanjut Duncan berat kantung empedu 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4
R3 4 1.0825
R0 4 1.1300
R5 4 1.3175
R2 4 1.3300
R1 4 1.6300
R4 4 1.8625
Sig .514 .863 1.000 1.000
Uji lanjut Duncan berat kantung empedu 42 hari
Perlakuan N Subset alfa = 0.05
1 2
R3 4 3.2150
R1 4 3.6350
R4 4 3.6875
R0 4 3.9250
R5 4 3.9475
R2 4 4.0075
Sig 1.000 .075
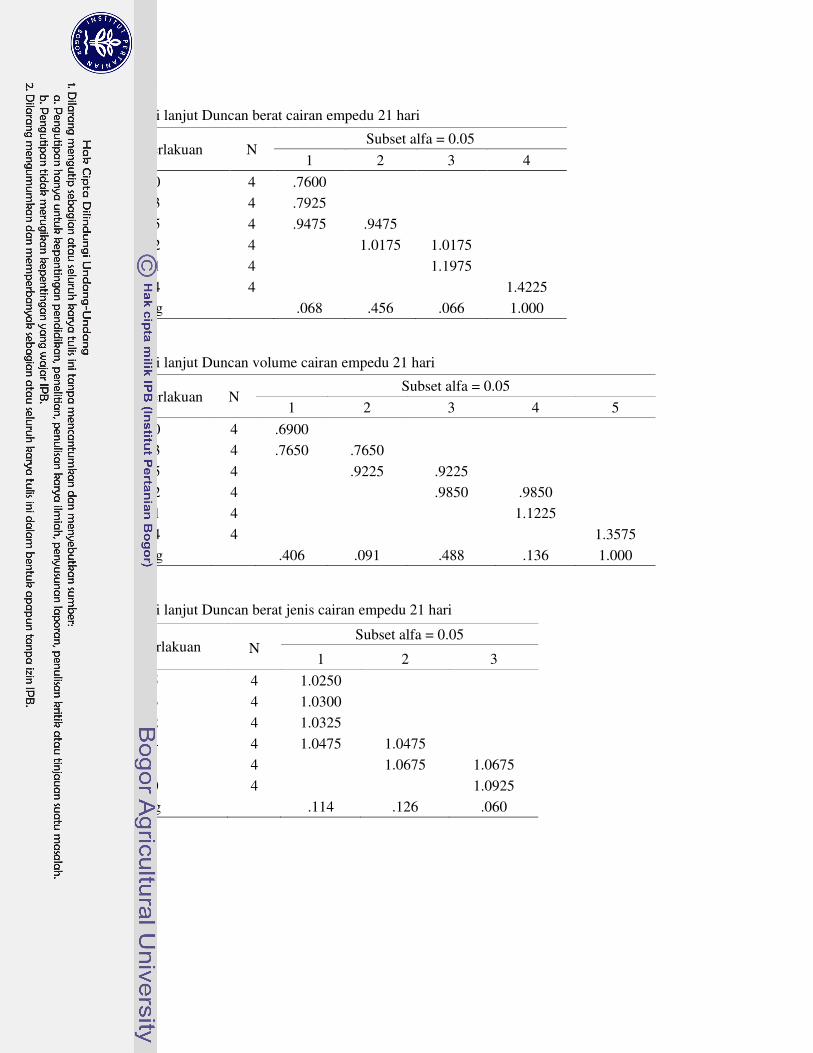
74
Uji lanjut Duncan berat cairan empedu 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4
R0 4 .7600
R3 4 .7925
R5 4 .9475 .9475
R2 4 1.0175 1.0175
R1 4 1.1975
R4 4 1.4225
Sig .068 .456 .066 1.000 Uji lanjut Duncan volume cairan empedu 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4 5
R0 4 .6900
R3 4 .7650 .7650
R5 4 .9225 .9225
R2 4 .9850 .9850
R1 4 1.1225
R4 4 1.3575
Sig .406 .091 .488 .136 1.000 Uji lanjut Duncan berat jenis cairan empedu 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R5 4 1.0250
R3 4 1.0300
R2 4 1.0325
R4 4 1.0475 1.0475
R1 4 1.0675 1.0675
R0 4 1.0925
Sig .114 .126 .060
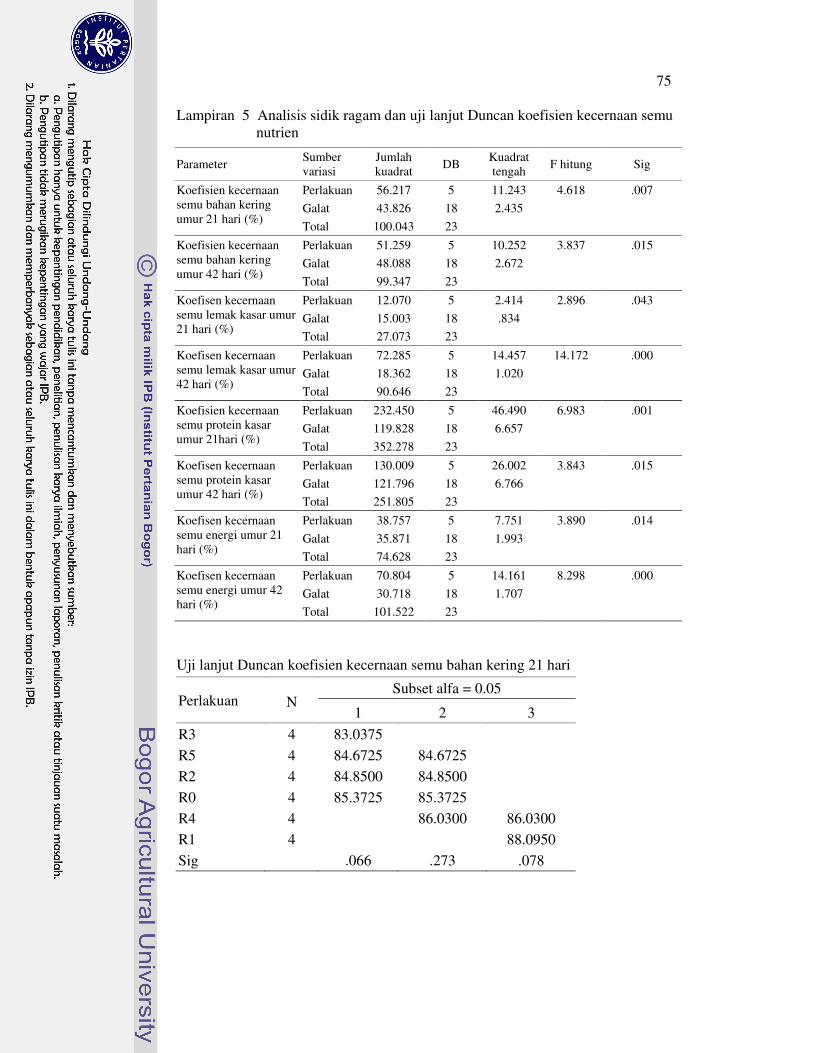
75
Lampiran 5 Analisis sidik ragam dan uji lanjut Duncan koefisien kecernaan semu nutrien
Uji lanjut Duncan koefisien kecernaan semu bahan kering 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 83.0375
R5 4 84.6725 84.6725
R2 4 84.8500 84.8500
R0 4 85.3725 85.3725
R4 4 86.0300 86.0300
R1 4 88.0950
Sig .066 .273 .078
Parameter Sumber variasi
Jumlah kuadrat
DB Kuadrat tengah
F hitung Sig
Koefisien kecernaan semu bahan kering umur 21 hari (%)
Perlakuan 56.217 5 11.243 4.618 .007
Galat 43.826 18 2.435
Total 100.043 23
Koefisien kecernaan semu bahan kering umur 42 hari (%)
Perlakuan 51.259 5 10.252 3.837 .015
Galat 48.088 18 2.672
Total 99.347 23
Koefisen kecernaan semu lemak kasar umur 21 hari (%)
Perlakuan 12.070 5 2.414 2.896 .043
Galat 15.003 18 .834
Total 27.073 23
Koefisen kecernaan semu lemak kasar umur 42 hari (%)
Perlakuan 72.285 5 14.457 14.172 .000
Galat 18.362 18 1.020
Total 90.646 23
Koefisien kecernaan semu protein kasar umur 21hari (%)
Perlakuan 232.450 5 46.490 6.983 .001
Galat 119.828 18 6.657
Total 352.278 23
Koefisen kecernaan semu protein kasar umur 42 hari (%)
Perlakuan 130.009 5 26.002 3.843 .015
Galat 121.796 18 6.766
Total 251.805 23
Koefisen kecernaan semu energi umur 21 hari (%)
Perlakuan 38.757 5 7.751 3.890 .014
Galat 35.871 18 1.993
Total 74.628 23
Koefisen kecernaan semu energi umur 42 hari (%)
Perlakuan 70.804 5 14.161 8.298 .000
Galat 30.718 18 1.707
Total 101.522 23
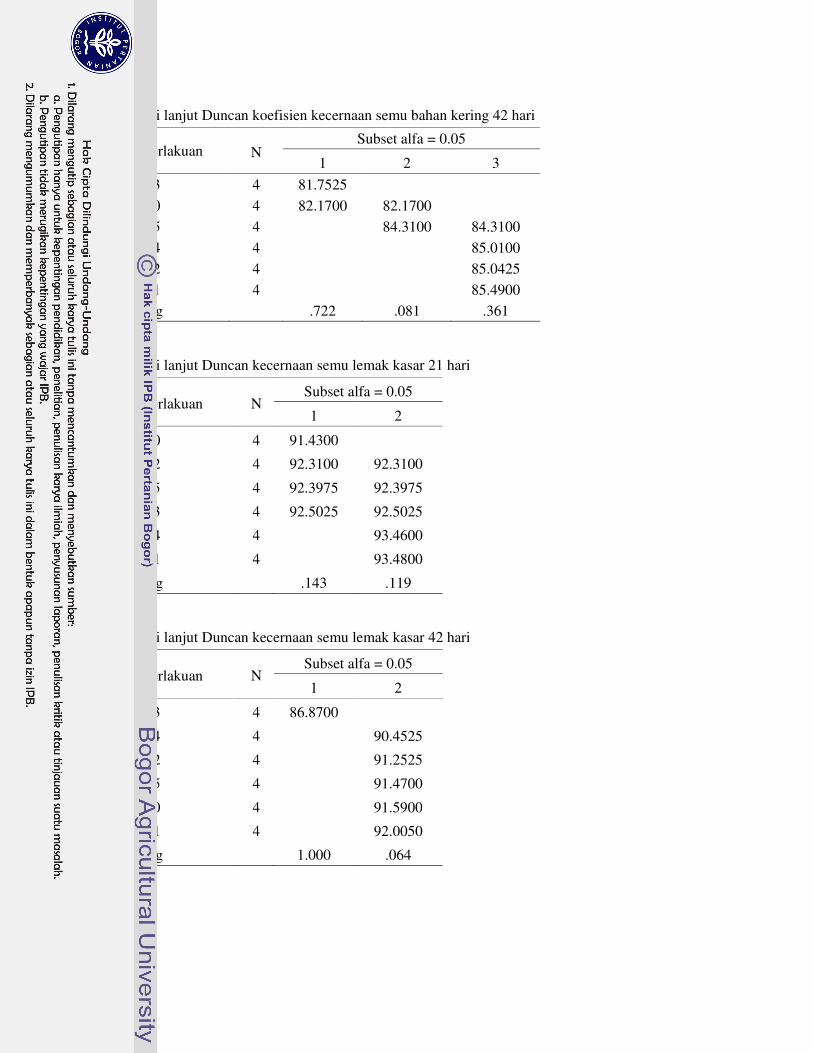
76
Uji lanjut Duncan koefisien kecernaan semu bahan kering 42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 81.7525
R0 4 82.1700 82.1700
R5 4 84.3100 84.3100
R4 4 85.0100
R2 4 85.0425
R1 4 85.4900
Sig .722 .081 .361 Uji lanjut Duncan kecernaan semu lemak kasar 21 hari
Perlakuan N Subset alfa = 0.05
1 2
R0 4 91.4300
R2 4 92.3100 92.3100
R5 4 92.3975 92.3975
R3 4 92.5025 92.5025
R4 4 93.4600
R1 4 93.4800
Sig .143 .119 Uji lanjut Duncan kecernaan semu lemak kasar 42 hari
Perlakuan N Subset alfa = 0.05
1 2
R3 4 86.8700
R4 4 90.4525
R2 4 91.2525
R5 4 91.4700
R0 4 91.5900
R1 4 92.0050
Sig 1.000 .064
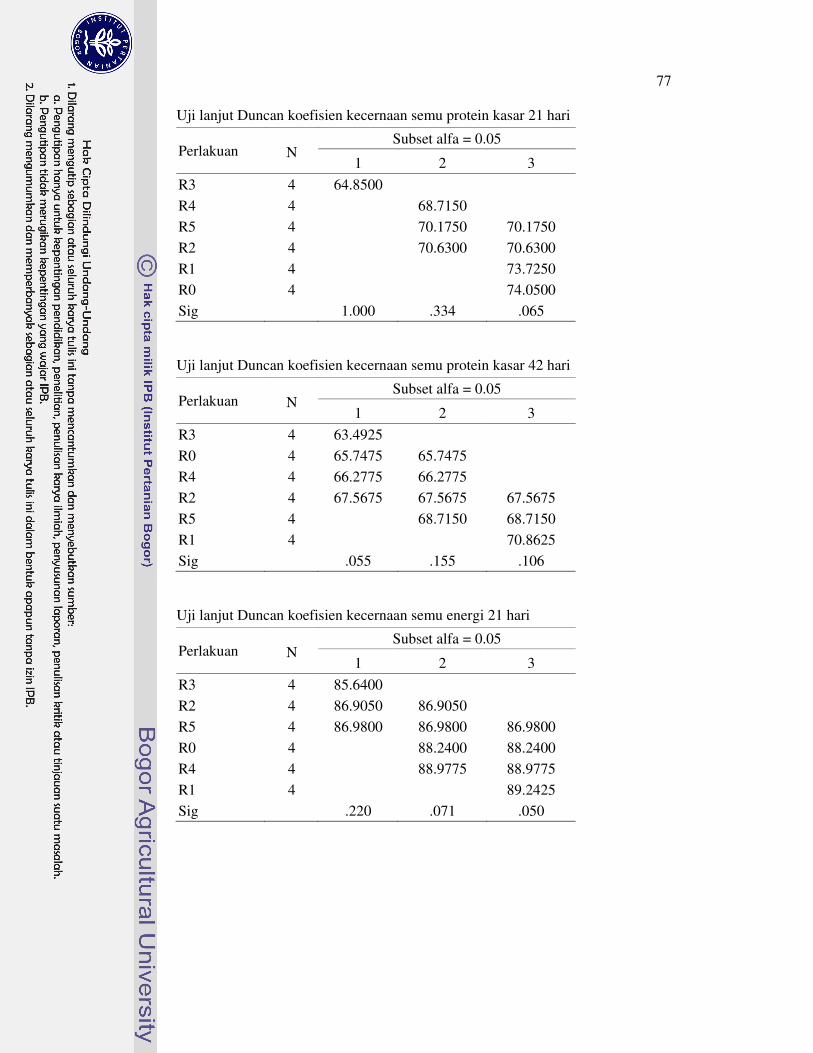
77
Uji lanjut Duncan koefisien kecernaan semu protein kasar 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 64.8500
R4 4 68.7150
R5 4 70.1750 70.1750
R2 4 70.6300 70.6300
R1 4 73.7250
R0 4 74.0500
Sig 1.000 .334 .065 Uji lanjut Duncan koefisien kecernaan semu protein kasar 42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 63.4925
R0 4 65.7475 65.7475
R4 4 66.2775 66.2775
R2 4 67.5675 67.5675 67.5675
R5 4 68.7150 68.7150
R1 4 70.8625
Sig .055 .155 .106 Uji lanjut Duncan koefisien kecernaan semu energi 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 85.6400
R2 4 86.9050 86.9050
R5 4 86.9800 86.9800 86.9800
R0 4 88.2400 88.2400
R4 4 88.9775 88.9775
R1 4 89.2425
Sig .220 .071 .050
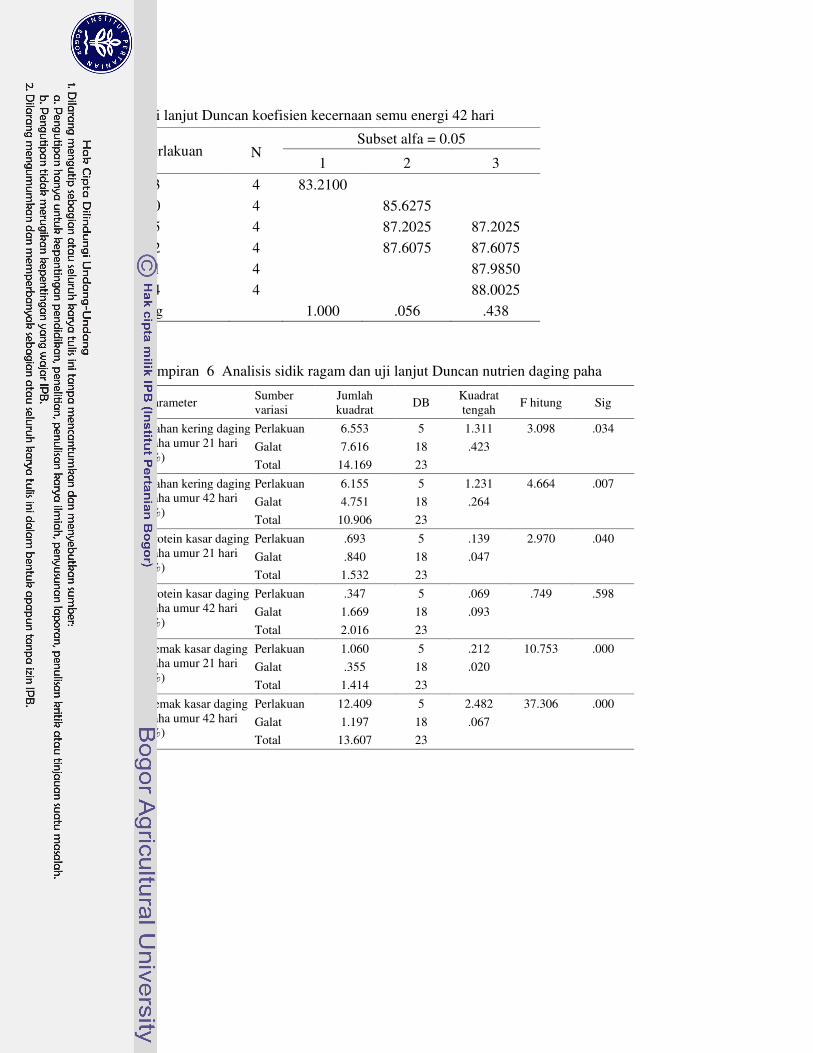
78
Uji lanjut Duncan koefisien kecernaan semu energi 42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 83.2100
R0 4 85.6275
R5 4 87.2025 87.2025
R2 4 87.6075 87.6075
R1 4 87.9850
R4 4 88.0025
Sig 1.000 .056 .438
Lampiran 6 Analisis sidik ragam dan uji lanjut Duncan nutrien daging paha
Parameter Sumber variasi
Jumlah kuadrat
DB Kuadrat tengah
F hitung Sig
Bahan kering daging paha umur 21 hari (%)
Perlakuan 6.553 5 1.311 3.098 .034
Galat 7.616 18 .423
Total 14.169 23
Bahan kering daging paha umur 42 hari (%)
Perlakuan 6.155 5 1.231 4.664 .007
Galat 4.751 18 .264
Total 10.906 23
Protein kasar daging paha umur 21 hari (%)
Perlakuan .693 5 .139 2.970 .040
Galat .840 18 .047
Total 1.532 23
Protein kasar daging paha umur 42 hari (%)
Perlakuan .347 5 .069 .749 .598
Galat 1.669 18 .093
Total 2.016 23
Lemak kasar daging paha umur 21 hari (%)
Perlakuan 1.060 5 .212 10.753 .000
Galat .355 18 .020
Total 1.414 23
Lemak kasar daging paha umur 42 hari (%)
Perlakuan 12.409 5 2.482 37.306 .000
Galat 1.197 18 .067
Total 13.607 23
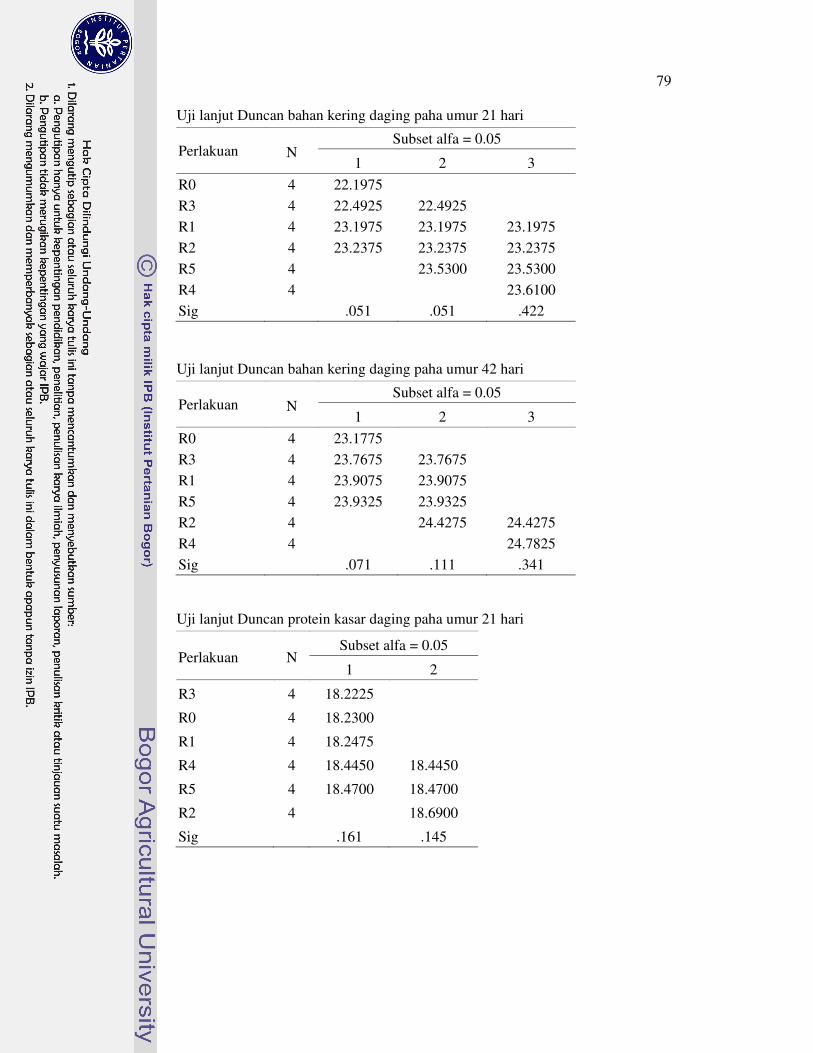
79
Uji lanjut Duncan bahan kering daging paha umur 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R0 4 22.1975
R3 4 22.4925 22.4925
R1 4 23.1975 23.1975 23.1975
R2 4 23.2375 23.2375 23.2375
R5 4 23.5300 23.5300
R4 4 23.6100
Sig .051 .051 .422
Uji lanjut Duncan bahan kering daging paha umur 42 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R0 4 23.1775
R3 4 23.7675 23.7675
R1 4 23.9075 23.9075
R5 4 23.9325 23.9325
R2 4 24.4275 24.4275
R4 4 24.7825
Sig .071 .111 .341 Uji lanjut Duncan protein kasar daging paha umur 21 hari
Perlakuan N Subset alfa = 0.05
1 2
R3 4 18.2225
R0 4 18.2300
R1 4 18.2475
R4 4 18.4450 18.4450
R5 4 18.4700 18.4700
R2 4 18.6900
Sig .161 .145
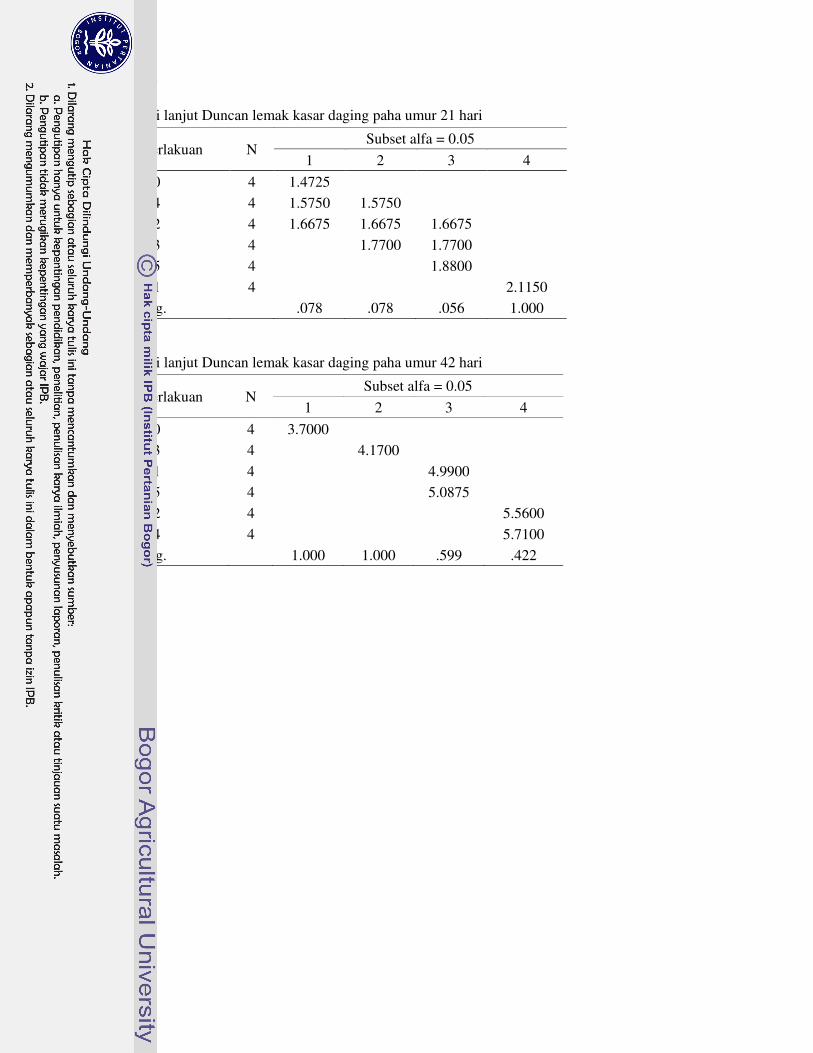
80
Uji lanjut Duncan lemak kasar daging paha umur 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4
R0 4 1.4725
R4 4 1.5750 1.5750
R2 4 1.6675 1.6675 1.6675
R3 4 1.7700 1.7700
R5 4 1.8800
R1 4 2.1150
Sig. .078 .078 .056 1.000 Uji lanjut Duncan lemak kasar daging paha umur 42 hari
Perlakuan N Subset alfa = 0.05
1 2 3 4
R0 4 3.7000
R3 4 4.1700
R1 4 4.9900
R5 4 5.0875
R2 4 5.5600
R4 4 5.7100
Sig. 1.000 1.000 .599 .422
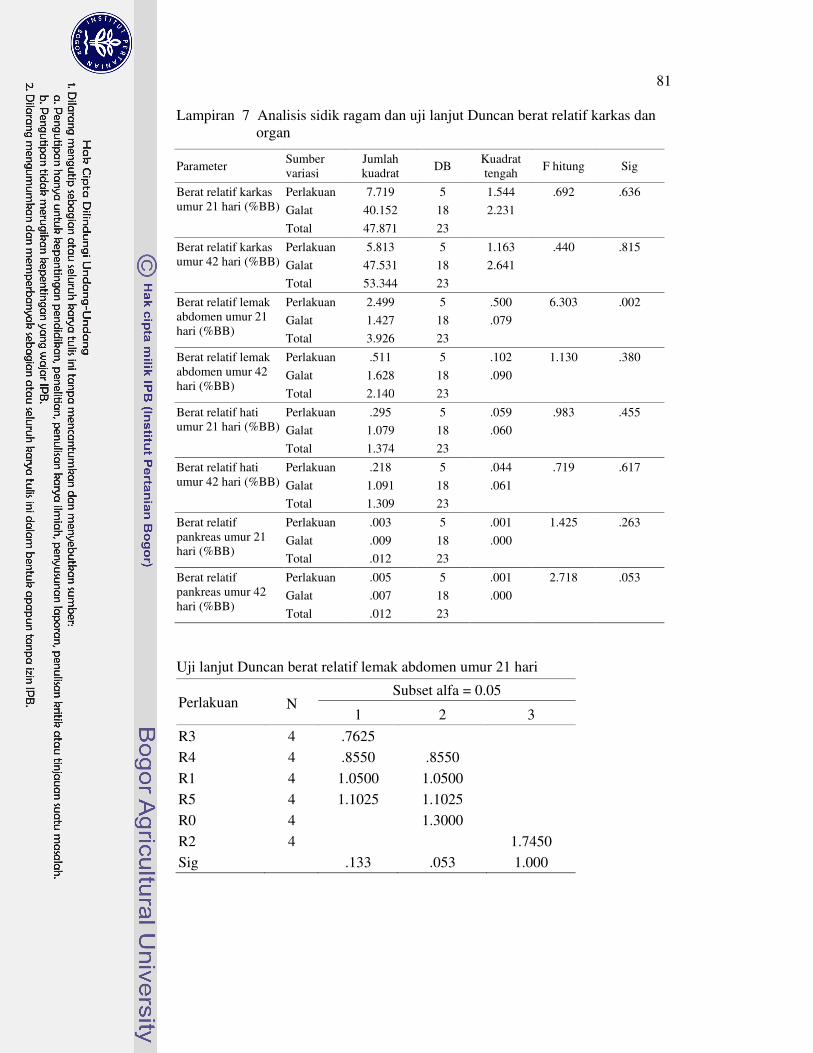
81
Lampiran 7 Analisis sidik ragam dan uji lanjut Duncan berat relatif karkas dan organ
Uji lanjut Duncan berat relatif lemak abdomen umur 21 hari
Perlakuan N Subset alfa = 0.05
1 2 3
R3 4 .7625
R4 4 .8550 .8550
R1 4 1.0500 1.0500
R5 4 1.1025 1.1025
R0 4 1.3000
R2 4 1.7450
Sig .133 .053 1.000
Parameter Sumber variasi
Jumlah kuadrat
DB Kuadrat tengah
F hitung Sig
Berat relatif karkas umur 21 hari (%BB)
Perlakuan 7.719 5 1.544 .692 .636
Galat 40.152 18 2.231
Total 47.871 23
Berat relatif karkas umur 42 hari (%BB)
Perlakuan 5.813 5 1.163 .440 .815
Galat 47.531 18 2.641
Total 53.344 23
Berat relatif lemak abdomen umur 21 hari (%BB)
Perlakuan 2.499 5 .500 6.303 .002
Galat 1.427 18 .079
Total 3.926 23
Berat relatif lemak abdomen umur 42 hari (%BB)
Perlakuan .511 5 .102 1.130 .380
Galat 1.628 18 .090
Total 2.140 23
Berat relatif hati umur 21 hari (%BB)
Perlakuan .295 5 .059 .983 .455
Galat 1.079 18 .060
Total 1.374 23
Berat relatif hati umur 42 hari (%BB)
Perlakuan .218 5 .044 .719 .617
Galat 1.091 18 .061
Total 1.309 23
Berat relatif pankreas umur 21 hari (%BB)
Perlakuan .003 5 .001 1.425 .263
Galat .009 18 .000
Total .012 23
Berat relatif pankreas umur 42 hari (%BB)
Perlakuan .005 5 .001 2.718 .053
Galat .007 18 .000
Total .012 23

82
RIWAYAT HIDUP
Penulis dilahirkan di Brebes pada tanggal 18 April 1989. Penulis merupakan anak ketiga dari empat bersaudara dari pasangan Bapak H Kasmuri Santoso dan Ibu Hj Nurjanah. Penulis merupakan suami dari Herwanti AMd Kes dan ayah dari Zafira Almira. Pendidikan sarjana ditempuh di Pogram Studi Ilmu Nutrisi dan Makanan Ternak, Fakultas Peternakan UNSOED. Penulis menyelesaikan pendidikan sarjana pada tahun 2011 dengan predikat “Dengan Pujian” dan “Lulusan Terbaik” Fakultas Peternakan pada periode wisuda Mei 2011. Pada tahun yang sama, penulis diterima di Program Studi Ilmu Nutrisi dan Pakan pada Program Pascasarjana IPB dan menamatkannya pada tahun 2014. Penulis memperoleh Beasiswa Unggulan Kementerian Pendidikan Nasional (KEMDIKNAS) pada tahun 2011.
Selama mengikuti program magister, penulis menjadi wakil ketua Himpunan Mahasiswa Pascasarjana (HIWACANA) di Program Studi Ilmu Nutrisi dan Pakan. Penulis juga menerima penghargaan INTP award sebagai mahasiswa berprestasi pada tahun 2012 dan 2013. Karya ilmiah berjudul Effects of Amino
Acid Supplementation on Nutritional Values and L-Carnitine of Broiler Meat akan diterbitkan pada jurnal Animal Production. Karya ilmiah tersebut merupakan bagian dari hasil penelitian program S-2 penulis.





























