Embed Size (px)
Citation preview

The University of Chicago
Evolution of Suites of Traits in Response to Environmental StressAuthor(s): F. Stuart Chapin III, Kellar Autumn and Francisco PugnaireReviewed work(s):Source: The American Naturalist, Vol. 142, Supplement: Evolutionary Responses toEnvironmental Stress (Jul., 1993), pp. S78-S92Published by: The University of Chicago Press for The American Society of NaturalistsStable URL: http://www.jstor.org/stable/2462710 .Accessed: 27/09/2012 03:30
Your use of the JSTOR archive indicates your acceptance of the Terms & Conditions of Use, available at .http://www.jstor.org/page/info/about/policies/terms.jsp
.JSTOR is a not-for-profit service that helps scholars, researchers, and students discover, use, and build upon a wide range ofcontent in a trusted digital archive. We use information technology and tools to increase productivity and facilitate new formsof scholarship. For more information about JSTOR, please contact [email protected].
.
The University of Chicago Press, The American Society of Naturalists, The University of Chicago arecollaborating with JSTOR to digitize, preserve and extend access to The American Naturalist.
http://www.jstor.org

Vol. 142, Supplement The American Naturalist July 1993
EVOLUTION OF SUITES OF TRAITS IN RESPONSE TO ENVIRONMENTAL STRESS
F. STUART CHAPIN III,* KELLAR AUTUMN,t AND FRANCISCO PUGNTAIREt
*Department of Integrative Biology, University of California, Berkeley, California 94720; tDepartment of Integrative Biology, Museum of Vertebrate Zoology, University of California, Berkeley, California 94720; tDepartamento de Ecologia, Universidad Complutense, E-28040
Madrid, Spain
Abstract.-Traits that enable plants to exploit low-resource environments (e.g., slow tissue turnover, low transpiration rate, high root: shoot ratio, and high concentrations of plant defenses against pathogens and herbivores) are physiologically linked to key growth-related traits (low rates of photosynthesis, nutrient uptake, and growth). Similar suites of traits occur as both phenotypically plastic and evolutionary responses to stress. We suggest that a genetic change in a switch or underlying trait that turns on this stress resistance syndrome (SRS), which causes it to be expressed over a wider range of environmental circumstances, would effectively convert a high-resource genotype into one that is more stress-tolerant. Because of physiological linkages between growth-related traits and the SRS, any heritable change in a key growth-related trait will pleiotropically affect the SRS. Therefore, heritable changes in these key growth-related traits could be accompanied by evolution of the entire SRS. Evidence for this hypothesis comes from single-gene mutants that differ in many stress-related traits, rapid evolution of metal- tolerant populations that are broadly stress-resistant, and consistent patterns of traits in species along gradients in resource availability. Similar evolutionary patterns occur in many animal taxa, which suggests that it is a general evolutionary phenomenon. We suggest that rapid evolution in response to changing environmental stress may allow many short-lived species to respond to human-induced environmental change and provide opportunities to develop stress-resistant crops. However, the time lag between generations of long-lived species that dominate most natural vegetation may not allow mature individuals of these species to keep pace with rapid global change.
Most plants characteristic of low-resource environments share a common suite of traits, including low rates of growth, photosynthesis, and nutrient absorption, high root: shoot ratios, low rates of tissue turnover, and high concentrations of secondary metabolites (Grime 1977; Chapin 1980; Bryant et al. 1983; Tilman 1988) (table 1) . On the basis of these general patterns, Grime (1977) suggested that this suite of traits, which we term a stress resistance syndrome (SRS), represents an "adaptive strategy" enabling plants to survive in stressful environments. In this article we suggest that this SRS is a consequence of physiological and develop- mental linkages among traits. Therefore, change in a key growth-related character can result in a cascade of effects that modify the entire SRS.
There is long-standing debate about whether large evolutionary changes occur by accumulation of many small mutational events or by a few mutations with large pleiotropic effects (Eldridge and Gould 1972; Lande 1983; Macnair 1991).
Am. Nat. 1993. Vol. 142, pp. S78-S92. ? 1993 by The University of Chicago. 0003-0147/93/4207-0006$02.00. All rights reserved.
EVOLUTION OF SUITES OF TRAITS IN PLANTS S79
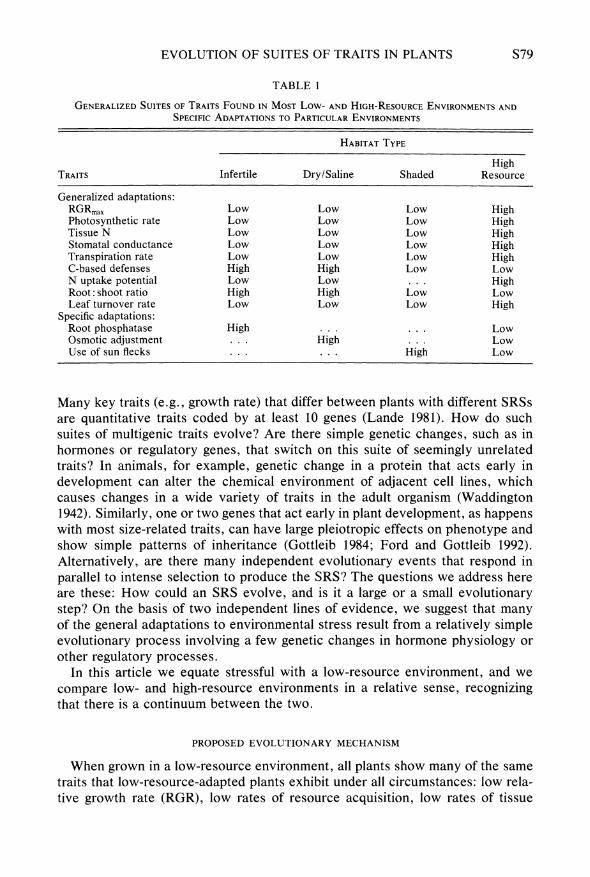
TABLE 1
GENERALIZED SUITES OF TRAITS FOUND IN MOST Low- AND HIGH-RESOURCE ENVIRONMENTS AND SPECIFIC ADAPTATIONS TO PARTICULAR ENVIRONMENTS
HABITAT TYPE
High TRAITS Infertile Dry/Saline Shaded Resource
Generalized adaptations: RGRmax Low Low Low High Photosynthetic rate Low Low Low High Tissue N Low Low Low High Stomatal conductance Low Low Low High Transpiration rate Low Low Low High C-based defenses High High Low Low N uptake potential Low Low ... High Root: shoot ratio High High Low Low Leaf turnover rate Low Low Low High
Specific adaptations: Root phosphatase High ... Low Osmotic adjustment ... High ... Low Use of sun flecks ... High Low
Many key traits (e.g., growth rate) that differ between plants with different SRSs are quantitative traits coded by at least 10 genes (Lande 1981). How do such suites of multigenic traits evolve? Are there simple genetic changes, such as in hormones or regulatory genes, that switch on this suite of seemingly unrelated traits? In animals, for example, genetic change in a protein that acts early in development can alter the chemical environment of adjacent cell lines, which causes changes in a wide variety of traits in the adult organism (Waddington 1942). Similarly, one or two genes that act early in plant development, as happens with most size-related traits, can have large pleiotropic effects on phenotype and show simple patterns of inheritance (Gottleib 1984; Ford and Gottleib 1992). Alternatively, are there many independent evolutionary events that respond in parallel to intense selection to produce the SRS? The questions we address here are these: How could an SRS evolve, and is it a large or a small evolutionary step? On the basis of two independent lines of evidence, we suggest that many of the general adaptations to environmental stress result from a relatively simple evolutionary process involving a few genetic changes in hormone physiology or other regulatory processes.
In this article we equate stressful with a low-resource environment, and we compare low- and high-resource environments in a relative sense, recognizing that there is a continuum between the two.
PROPOSED EVOLUTIONARY MECHANISM
When grown in a low-resource environment, all plants show many of the same traits that low-resource-adapted plants exhibit under all circumstances: low rela- tive growth rate (RGR), low rates of resource acquisition, low rates of tissue
S80 THE AMERICAN NATURALIST
Hgh-resource o / ~~~~~~species 0
O/
/ ~~~~~species
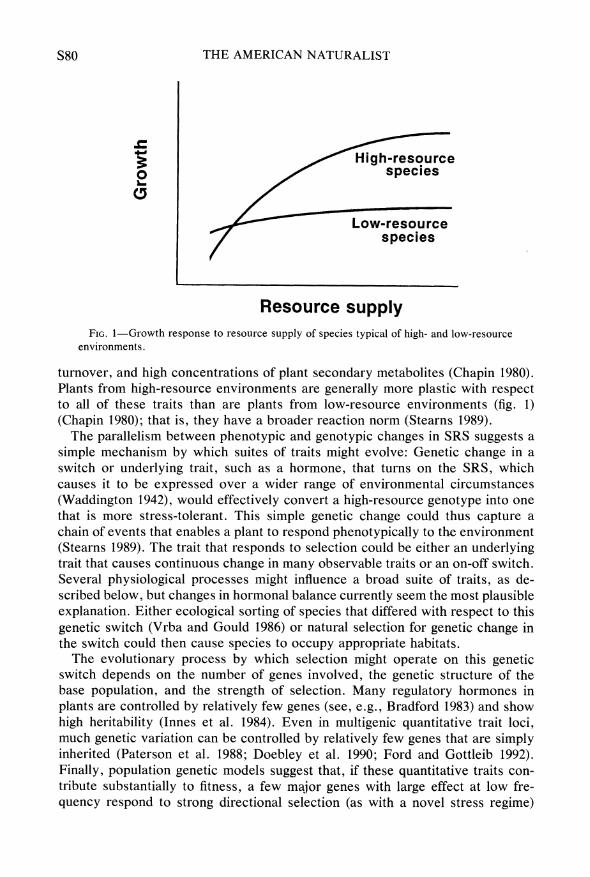
Resource supply FIG. 1-Growth response to resource supply of species typical of high- and low-resource
environments.
turnover, and high concentrations of plant secondary metabolites (Chapin 1980). Plants from high-resource environments are generally more plastic with respect to all of these traits than are plants from low-resource environments (fig. 1) (Chapin 1980); that is, they have a broader reaction norm (Stearns 1989).
The parallelism between phenotypic and genotypic changes in SRS suggests a simple mechanism by which suites of traits might evolve: Genetic change in a switch or underlying trait, such as a hormone, that turns on the SRS, which causes it to be expressed over a wider range of environmental circumstances (Waddington 1942), would effectively convert a high-resource genotype into one that is more stress-tolerant. This simple genetic change could thus capture a chain of events that enables a plant to respond phenotypically to the environment (Stearns 1989). The trait that responds to selection could be either an underlying trait that causes continuous change in many observable traits or an on-off switch. Several physiological processes might influence a broad suite of traits, as de- scribed below, but changes in hormonal balance currently seem the most plausible explanation. Either ecological sorting of species that differed with respect to this genetic switch (Vrba and Gould 1986) or natural selection for genetic change in the switch could then cause species to occupy appropriate habitats.
The evolutionary process by which selection might operate on this genetic switch depends on the number of genes involved, the genetic structure of the base population, and the strength of selection. Many regulatory hormones in plants are controlled by relatively few genes (see, e.g., Bradford 1983) and show high heritability (Innes et al. 1984). Even in multigenic quantitative trait loci, much genetic variation can be controlled by relatively few genes that are simply inherited (Paterson et al. 1988; Doebley et al. 1990; Ford and Gottleib 1992). Finally, population genetic models suggest that, if these quantitative traits con- tribute substantially to fitness, a few major genes with large effect at low fre- quency respond to strong directional selection (as with a novel stress regime)

EVOLUTION OF SUITES OF TRAITS IN PLANTS S81
more than would many genes of small effect at intermediate gene frequency (Parsons 1987; Macnair 1991). In cases in which the new stress regime does not differ strongly from the original environment, polygenic adaptation involving changes in many genes, each with small effect, may better explain evolutionary changes in quantitative traits (Lande 1983; Macnair 1991). Thus, there are several plausible evolutionary mechanisms by which a few genes that exert strong control over many traits might respond to selection, giving rise to an SRS.
In animals there is also considerable evidence for developmental switches and simply inherited underlying traits with widespread morphological and physiologi- cal consequences (see, e.g., Wright 1927; Kauffman 1971; Eldridge and Gould 1972; Sternberg and Horvitz 1981; Raff and Kaufman 1983; Arnold 1987). There is also evidence from insects that acclimation and genetic responses to stress can involve the same physiological mechanism (Hoffmann 1990). Thus, it would be surprising if the phenomenon did not occur in plants.
PHYSIOLOGICAL LINK BETWEEN GROWTH AND STRESS RESISTANCE
There are several direct and indirect ways in which slow rates of resource acquisition and growth confer stress resistance. Slow growth causes slow turn- over of tissues produced previously, particularly in plants that produce multiple cohorts of leaves (e.g., grasses and evergreen species: Langer 1966; Chapin 1980). The reduction in tissue turnover conserves carbon and nutrients because most of the carbon and half of the nutrients in a leaf (Chapin and Kedrowski 1983; Chapin 1991) and most of the carbon and nutrients in a root (Nambiar and Fife 1991) are lost at senescence. Slow growth also minimizes the growth respiration associated with production of new tissues (Penning de Vries et al. 1974). By reducing re- source loss, slow growth reduces dependence on the environment for new re- sources. The low rate of photosynthesis associated with slow growth also reduces water loss because of the tight relationship of photosynthesis to leaf area and stomatal conductance (Farquhar and Sharkey 1983; Collatz et al. 1991), which thereby reduces water requirements and confers stress resistance in dry or saline habitats.
Slow growth indirectly confers stress resistance by reducing carbon demands for growth, which thereby allows greater allocation to other processes that di- rectly contribute to stress resistance such as storage (Chapin et al. 1990), chemical defense against pathogens and herbivores (Mooney and Gulmon 1982; Bryant et al. 1983; Coley et al. 1985), or active detoxification of heavy metals or byproducts of anaerobic metabolism found in flooded soils.
Conversely, any process that increases growth rate also increases tissue turn- over and loss of carbon, nutrients, and water and decreases allocation to storage and plant defense, which renders plants less stress resistant.
PHYSIOLOGICAL BASIS OF THE SRS
Tight integration among physiological processes provides several mechanisms by which changes in one or a few key traits have widespread physiological rami-
S82 THE AMERICAN NATURALIST
Low soil resource water nutrient oxygen
Low temperature
I Root signal
Hydraulic t ABA conductance ; Cytokinins
Others ? ; Water content
Turgor Cell wall Stomatal Plant N \extensibility conductance status
;Leaf growth
; Carbon demand *p Photosynthesis
G; RGR
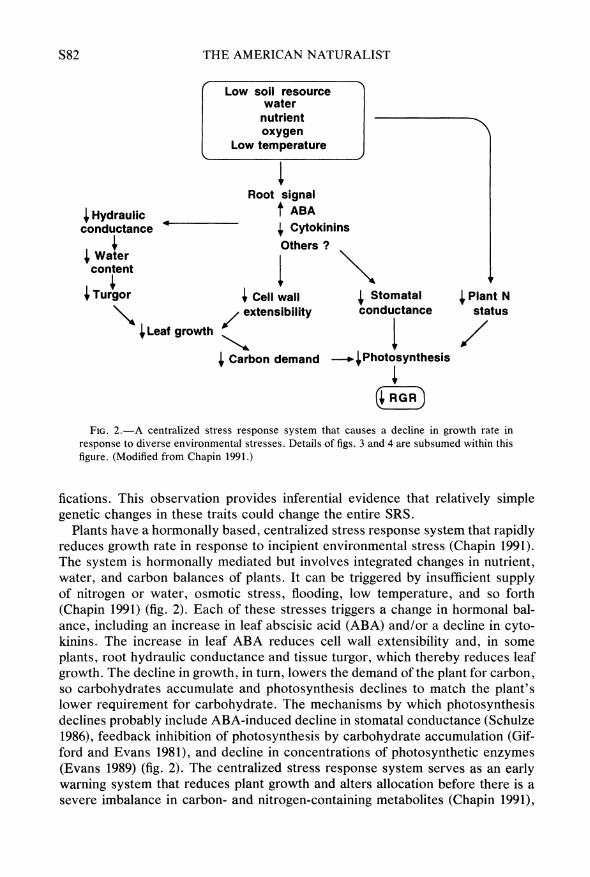
FIG. 2.-A centralized stress response system that causes a decline in growth rate in response to diverse environmental stresses. Details of figs. 3 and 4 are subsumed within this figure. (Modified from Chapin 1991.)
fications. This observation provides inferential evidence that relatively simple genetic changes in these traits could change the entire SRS.
Plants have a hormonally based, centralized stress response system that rapidly reduces growth rate in response to incipient environmental stress (Chapin 1991). The system is hormonally mediated but involves integrated changes in nutrient, water, and carbon balances of plants. It can be triggered by insufficient supply of nitrogen or water, osmotic stress, flooding, low temperature, and so forth (Chapin 1991) (fig. 2). Each of these stresses triggers a change in hormonal bal- ance, including an increase in leaf abscisic acid (ABA) and/or a decline in cyto- kinins. The increase in leaf ABA reduces cell wall extensibility and, in some plants, root hydraulic conductance and tissue turgor, which thereby reduces leaf growth. The decline in growth, in turn, lowers the demand of the plant for carbon, so carbohydrates accumulate and photosynthesis declines to match the plant's lower requirement for carbohydrate. The mechanisms by which photosynthesis declines probably include ABA-induced decline in stomatal conductance (Schulze 1986), feedback inhibition of photosynthesis by carbohydrate accumulation (Gif- ford and Evans 1981), and decline in concentrations of photosynthetic enzymes (Evans 1989) (fig. 2). The centralized stress response system serves as an early warning system that reduces plant growth and alters allocation before there is a severe imbalance in carbon- and nitrogen-containing metabolites (Chapin 1991),
EVOLUTION OF SUITES OF TRAITS IN PLANTS S83
(Large seed
Large leaf I
Large source-sink
ratio
Greater Low photosynthetic Low leaf self-shading potential allocation
Low photosynthetic High maintenance rate respiration \ /
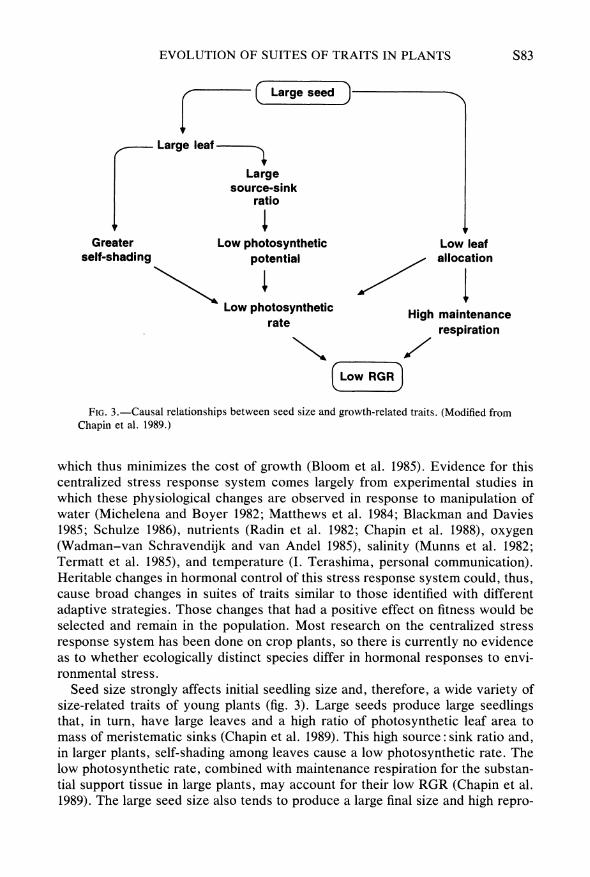
FIG. 3.-Causal relationships between seed size and growth-related traits. (Modified from Chapin et al. 1989.)
which thus minimizes the cost of growth (Bloom et al. 1985). Evidence for this centralized stress response system comes largely from experimental studies in which these physiological changes are observed in response to manipulation of water (Michelena and Boyer 1982; Matthews et al. 1984; Blackman and Davies 1985; Schulze 1986), nutrients (Radin et al. 1982; Chapin et al. 1988), oxygen (Wadman-van Schravendijk and van Andel 1985), salinity (Munns et al. 1982; Termatt et al. 1985), and temperature (I. Terashima, personal communication). Heritable changes in hormonal control of this stress response system could, thus, cause broad changes in suites of traits similar to those identified with different adaptive strategies. Those changes that had a positive effect on fitness would be selected and remain in the population. Most research on the centralized stress response system has been done on crop plants, so there is currently no evidence as to whether ecologically distinct species differ in hormonal responses to envi- ronmental stress.
Seed size strongly affects initial seedling size and, therefore, a wide variety of size-related traits of young plants (fig. 3). Large seeds produce large seedlings that, in turn, have large leaves and a high ratio of photosynthetic leaf area to mass of meristematic sinks (Chapin et al. 1989). This high source: sink ratio and, in larger plants, self-shading among leaves cause a low photosynthetic rate. The low photosynthetic rate, combined with maintenance respiration for the substan- tial support tissue in large plants, may account for their low RGR (Chapin et al. 1989). The large seed size also tends to produce a large final size and high repro-

S84 THE AMERICAN NATURALIST
ductive effort. Evidence for this chain of events comes largely from correlations among traits in related species with different seed sizes: Seed size correlates positively with seedling size; seedling size correlates positively with adult size and reproductive effort (Harper 1977; Fenner 1985); leaf size correlates inversely with photosynthetic rates (Gifford and Evans 1981); seedling size correlates in- versely with RGR (Fenner 1983; Chapin et al. 1989). There are, of course, many other factors that also influence growth rate and reproductive effort, so these correlations are generally seen only among closely related species or genotypes.
Manipulative experiments demonstrate a causal basis for these correlations. When seed size is experimentally reduced by restricting resource supply to the parent plant (maternal effect) (Harper 1977) or by excising part of the storage endosperm (Chapin and Bieleski 1982), plant size declines, RGR increases, and reproductive effort decreases. Similarly, if the source strength of leaves is re- duced by removing part of the leaves, whole-plant RGR and photosynthetic rate of remaining leaves increase, even under optimal water and nutrient supply (Mc- Naughton 1983; Nowak and Caldwell 1984), which presumably reflects changes in the source: sink ratio. Conversely, if sink strength is reduced by removing developing grains, photosynthetic rate declines (Chapin and Wardlaw 1988). Thus, change in traits affecting seed size could have widespread physiological ramifications for photosynthesis, growth, and reproduction (i.e., traits that are key components of distinct adaptive strategies). Plants that germinate predictably in dry or shaded habitats tend to have large seeds (Baker 1972), which suggests a relationship between seed size and stress resistance. Seed size is one of the least variable traits in plants, typically varying less than 10% in response to environment (as compared to 10 orders of magnitude variation among species) (Harper 1977; but see Winn 1988). This indicates that evolution has substantially extended the narrow range of seed size made possible by phenotypic plasticity (Fenner 1985), but it is difficult to evaluate the relative importance of mobility and environmental stress in selecting for or sorting these patterns of seed size.
There are well-documented physiological links among tissue nitrogen (N) con- centration, photosynthesis, stomatal conductance, and transpiration (Field 1991) (fig. 4). Plants growing in low-N soils have low tissue N concentrations, low concentrations of photosynthetic enzymes, and, therefore, low photosynthetic potential (Evans 1983; Field and Mooney 1986). Because plants appear to regulate internal carbon dioxide concentration by matching stomatal conductance to photosynthetic potential (Farquhar and Sharkey 1983), plants with low photo- synthetic rate have low stomatal conductance and, therefore, low transpiration rate. Low water availability can independently initiate the same chain of events through changes in stomatal conductance, leaf area (Nobel 1983), and/or soil N availability (Chapin 1991). These traits remain physiologically linked when comparing a given genotype grown at different N concentrations (Evans 1983), different genotypes of a species, or different species or growth forms (Field and Mooney 1986). Therefore, any simple genetic changes that alter tissue N concen- tration or stomatal conductance would alter this entire suite of traits, in a fashion that parallels phenotypic response to environmental stress. The divergence in these traits among species (Field and Mooney 1986) is much greater than the
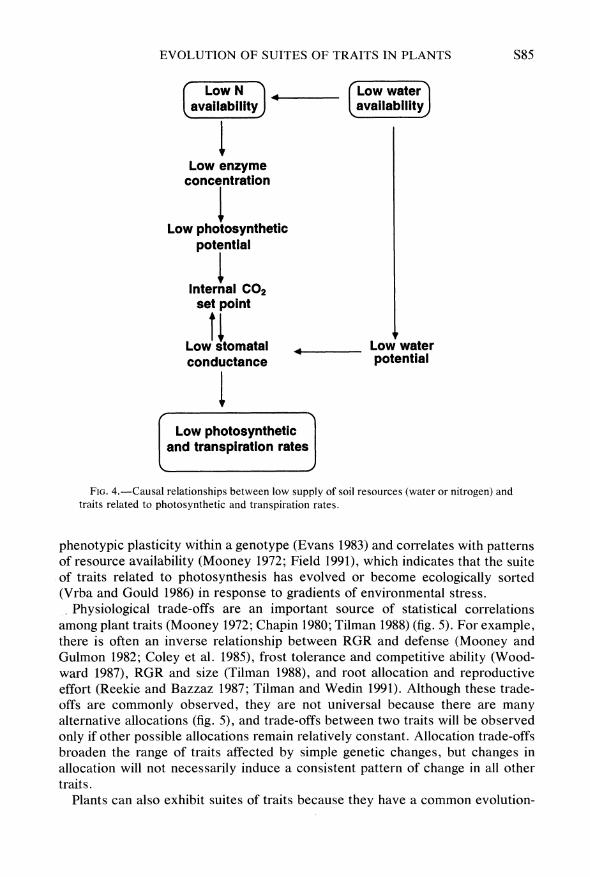
EVOLUTION OF SUITES OF TRAITS IN PLANTS S85
Low N ) Low water availability availability
Low enzyme concentration
Low photosynthetic potential
Internal CO2 set point
Low stomatal * Low water conductance potential
Low photosynthetic and transpiration rates
FIG. 4.-Causal relationships between low supply of soil resources (water or nitrogen) and traits related to photosynthetic and transpiration rates.
phenotypic plasticity within a genotype (Evans 1983) and correlates with patterns of resource availability (Mooney 1972; Field 1991), which indicates that the suite of traits related to photosynthesis has evolved or become ecologically sorted (Vrba and Gould 1986) in response to gradients of environmental stress.
Physiological trade-offs are an important source of statistical correlations among plant traits (Mooney 1972; Chapin 1980; Tilman 1988) (fig. 5). For example, there is often an inverse relationship between RGR and defense (Mooney and Gulmon 1982; Coley et al. 1985), frost tolerance and competitive ability (Wood- ward 1987), RGR and size (Tilman 1988), and root allocation and reproductive effort (Reekie and Bazzaz 1987; Tilman and Wedin 1991). Although these trade- offs are commonly observed, they are not universal because there are many alternative allocations (fig. 5), and trade-offs between two traits will be observed only if other possible allocations remain relatively constant. Allocation trade-offs broaden the range of traits affected by simple genetic changes, but changes in allocation will not necessarily induce a consistent pattern of change in all other traits.
Plants can also exhibit suites of traits because they have a common evolution-

S86 THE AMERICAN NATURALIST
r RGR, Wood
Soil ~~~~~~~~Defense, ( i3 0 Carbon frost hardeness resource gain root allocation etc.
Reproduction
Seed Seed size number
Disperser attraction
FIG. 5.-Allocation trade-offs responsible for negative correlations among plant traits in response to variations in resource supply. (Modified from Hobbie et al. 1993.)
ary history rather than because these traits are functionally related (Lechowicz 1984; Herrera 1992). If traits co-occur for historical reasons, there is no reason to expect a genetic change in one trait to alter other traits in any predictable fashion.
EVOLUTIONARY EVIDENCE
Genetic and evolutionary studies provide the second line of evidence for simple heritability of the SRS.
Single-gene mutants that do not produce or respond to ABA differ from the wild type in many stress-related traits. For example, the flacca mutant of tomato and droopy mutant of potato produce less ABA, have lower hydraulic conduc- tance, and have lower rates of photosynthesis, nutrient absorption, and growth than the wild type (Quarrie 1982; Bradford 1983; Neill and Horgan 1985; Chapin 1990). Theflacca mutant is more sensitive to water stress because it cannot close its stomata, which indicates that a single-gene change in hormonal balance can trigger a change in the centralized stress response system and consequently in a broad suite of seemingly unrelated physiological traits.
Another example of a single-gene change with widespread ramifications for stress resistance involves a point mutation that alters a single amino acid in a chloroplast protein. This change affects not only resistance to the herbicide tri- azine but also photosynthesis, growth, flowering time, and sensitivity to environ- mental stress (Warwick 1991). Dwarfing genes, which operate by altering sensitiv- ity to gibberellins, are also simply inherited and cause widespread changes in growth, allocation, photosynthesis, and yield (Evans 1980; Gottleib 1984).
Many populations have substantial variability in stress-related traits on which natural selection could operate (Bradshaw 1984). When selection is imposed for

EVOLUTION OF SUITES OF TRAITS IN PLANTS S87
a single stress-related character, other stress-related characters are often also altered. For example, there is a threefold variation in ABA concentration among wheat genotypes (Quarrie 1981). Selection for high-ABA genotypes results in substantial changes in drought resistance, yield, and other associated traits (Innes et al. 1984). This study demonstrates that hormones can have a central role in the evolution of suites of traits. Change in a hormone, which orchestrates a broad array of physiological processes, may have less deleterious effects on growth than would change in other gene products with comparably widespread pleiotro- pic effects.
When novel environmental stresses are imposed, there is often rapid evolution of stress tolerance in a population (Wu et al. 1975; Bradshaw 1984). For example, beneath zinc-coated fences, on mine tailings, and near roadsides, populations often develop within years to decades that are metal-tolerant and possess most associated adaptations to environmental stress such as slow growth and low rates of photosynthesis, nutrient uptake, and transpiration (Jowett 1959). Although metal tolerance is a multigenic trait, a few genes often exert major effects (Mac- nair 1989, 1991), which leads to relatively simple patterns of inheritance. Simi- larly, in contrasting nutritional environments, populations have diverged in many stress-related traits (Snaydon and Bradshaw 1962; Gates 1987). In most cases, evolution probably resulted from natural selection for stress-resistant genotypes present in the parent populations at low frequency (see, e.g., Gartside and McNeilly 1974; Bradshaw 1984; Macnair 1991). Birch (Betula nana), which spread to Iceland, where mammalian herbivores were absent until recently, lost its ability to produce papyriferic acid, an energetically costly chemical defense, and it shows a correspondingly higher growth rate in absence of herbivores than do populations that produce the defense (Bryant et al. 1989). These population studies indicate that suites of traits related to environmental stress can be gained or lost by some species, often within decades.
A final line of evolutionary evidence comes from the similarity among divergent taxa in the distribution of suites of traits along environmental gradients (Grime 1977; Chapin 1980). For example, Chionochloa has speciated in response to mountain building in New Zealand. Those species that occur on the most infertile soils have low rates of nutrient uptake and growth and are relatively insensitive to variation in nutrient supply (Chapin et al. 1982). Similar divergence in suites of stress-related traits has occurred with speciation in many groups (Clarkson 1967; Beadle 1968; Veerkamp et al. 1980). If the same suite of traits consistently recurs in diverse evolutionary lineages, it seems likely that it evolved as a coher- ent unit rather than by parallel selection of each individual trait.
Although we have argued that suites of stress-related traits can evolve through relatively simple genetic changes, we recognize that there are many additional traits (e.g., capacity for osmotic regulation, capacity to immobilize heavy metals) that are essential to occupy specific extreme environments (table 1). Many of these traits entail energetic costs that might further reduce allocation to growth. We suggest that effective evolution in response to stress cannot occur without a change in the broad suite of growth-related traits, which may be a relatively

S88 THE AMERICAN NATURALIST
simple evolutionary process, but that occupation of extremely stressful environ- ments also requires certain habitat-specific traits. For this reason relatively few taxa can occupy any particular extreme habitat.
IMPLICATIONS FOR GLOBAL CHANGE
As a consequence of exponential human population growth, there have been rapid changes in local, regional, and global patterns of environmental stress. Local and regional changes in nutrient availability result from either erosional losses associated with overgrazing and deforestation or from enrichment associ- ated with fertilizer inputs, nitrogen deposition from acid rain, and eutrophication from pollution. Similarly, water use for industry, recreation, and agriculture alters water availability locally and perhaps regionally (Shukla et al. 1990). Human activities often cause novel stresses or combinations of stresses such as the com- bination of acidity and increased nitrogen availability associated with acid rain, ozone and acidity in urban pollutants, and salinity and high fertility associated with agricultural salinization.
The rapidity with which stress-tolerant populations of some species evolve suggests that natural populations of some species may accommodate evolution- arily to changes in environmental stress, as well as through phenotypic plasticity of individuals and changes in species composition of communities. Detectable changes in stress resistance required five generations of intensive selection for high ABA (Innes et al. 1984). Populations of metal-tolerant grasses appear within a few decades (Antonovics et al. 1971; Bradshaw 1984), approximately the same time scale as global climate change. Thus, short-lived species may show substan- tial turnover of individuals and adjust evolutionarily to global changes in environ- ment. The unanswered question is which species will evolve in response to which stresses and which range of stresses can each species accommodate evolution- arily. By contrast, long-lived perennial species, such as trees, shrubs, and peren- nial grasses, which dominate most of the world's natural landscapes, turn over more slowly. These long-lived species may have greater difficulty adjusting to rapid environmental change than short-lived species because of longer intervals between successful seedling cohorts and large changes in environmental stress within the lifetime of a single individual. The capacity of these long-lived, large- statured species to respond to environmental change is critical because these species have the greatest impact on ecosystem and global processes (Chapin 1993).
Rapid evolution of stress tolerance provides a complement to the green revolu- tion in providing food in developing nations. Because most of the fertile lands of the world are already employed in agricultural production, expansion of agricul- tural potential must come largely from farming in environmentally marginal habi- tats (Chapin 1980; Vose 1984). Where economics preclude massive additions of water and fertilizer, it seems feasible to select for stress-tolerant genotypes that would grow more slowly and have lower maximum yield but would dependably produce a crop under a wide range of environmental conditions. Development of varieties that are genetically variable with respect to stress tolerance could hedge

EVOLUTION OF SUITES OF TRAITS IN PLANTS S89
bets against stochastic variation in weather and localized patchiness in resource supply.
ACKNOWLEDGMENTS
We thank B. Hungate, T. Jackman, M. Lynch, K. Schwaegerle, M. Slatkin, D. Wake, M. Wake, and two anonymous reviewers for insightful comments and critical feedback. These ideas grew out of research funded by the National Sci- ence Foundation (BSR87-05323 and BSR90-20135); F.P. was supported by the Spanish Ministry of Education.
LITERATURE CITED
Antonovics, J., A. D. Bradshaw, and R. G. Turner. 1971. Heavy metal tolerance in plants. Advances in Ecological Research 7:1-85.
Arnold, S. J. 1987. Genetic correlation and the evolution of physiology. Pages 189-212 in M. E. Feder, E. F. Bennett, W. W. Burggren, and R. B. Huey, eds. New directions in ecological physiology. Cambridge University Press, Cambridge.
Baker, H. G. 1972. Seed weight in relation to environmental conditions in California. Ecology 53:997-1010.
Beadle, N. C. W. 1968. Some aspects of the ecology and physiology of Australian xeromorphic plants. Australian Journal of Science 30:348-355.
Blackman, P. G., and W. J. Davies. 1985. Root to shoot communication in maize plants of the effects of soil drying. Journal of Experimental Botany 36:39-48.
Bloom, A. J., F. S. Chapin, III, and H. A. Mooney. 1985. Resource limitation in plants-an economic analogy. Annual Review of Ecology and Systematics 16:363-392.
Bradford, K. J. 1983. Water relations and growth of the flacca tomato mutant in relation to abscisic acid. Plant Physiology 72:251-255.
Bradshaw, A. D. 1984. The importance of evolutionary ideas in ecology-and vice versa. Pages 1-26 in B. Shorrocks, ed. Evolutionary ecology. Blackwell, Oxford.
Bryant, J. P., F. S. Chapin, III, and D. R. Klein. 1983. Carbon/nutrient balance of boreal plants in relation to vertebrate herbivory. Oikos 40:357-368.
Bryant, J. P., J. Tahvanainen, M. Sulkinoja, R. Julkunen-Tiitto, P. Reichardt, and T. Green. 1989. Biogeographic evidence for the evolution of chemical defense by boreal birch and willow against mammalian browsing. American Naturalist 134:20-34.
Chapin, F. S., III. 1980. The mineral nutrition of wild plants. Annual Review of Ecology and Systemat- ics 11:233-260. 1990. Effects of nutrient deficiency on plant growth: evidence for a centralized stress-response system. British Society for Plant Growth Regulation 21:135-148. 1991. Integrated responses of plants to stress. BioScience 41:29-36. 1993. Functional role of growth forms in ecosystem and global processes. Pages 287-312 in J. R. Ehleringer and C. B. Field, eds. Scaling physiological processes: leaf to globe. Aca- demic Press, San Diego.
Chapin, F. S., III, and R. L. Bieleski. 1982. Mild phosphorus stress in barley and a related low- phosphorus-adapted barley grass: phosphorus fractions and phosphate absorption in relation to growth. Physiologia Plantarum 54:309-317.
Chapin, F. S., III, and R. A. Kedrowski. 1983. Seasonal changes in nitrogen and phosphorus fractions and autumn retranslocation in evergreen and deciduous taiga trees. Ecology 64:376-391.
Chapin, F. S., III, and I. F. Wardlaw. 1988. Effect of phosphorus deficiency on source-sink interac- tions between the flag leaf and developing grain in barley. Journal of Experimental Botany 39:165-177.
Chapin, F. S., III, J. Follett, and K. F. O'Connor. 1982. Growth, phosphate absorption, and phospho- rus chemical fractions in two Chionochloa species. Journal of Ecology 70:305-321.

S90 THE AMERICAN NATURALIST
Chapin, F. S., III, C. H. S. Walter, and D. T. Clarkson. 1988. Growth response of barley and tomato to nitrogen stress and its control by abscisic acid, water relations, and photosynthesis. Planta 173:352-366.
Chapin, F. S., III, R. H. Groves, and L. T. Evans. 1989. Response of growth, photosynthesis, and phosphate absorption to phosphorus stress in wild, weedy, and cultivated Hordeum species. Oecologia (Berlin) 79:96-105.
Chapin, F. S., III, E.-D. Schulze, and H. A. Mooney. 1990. The ecology and economics of storage in plants. Annual Review of Ecology and Systematics 21:423-447.
Clarkson, D. T. 1967. Phosphorus supply and growth rate in species of Agrostis L. Journal of Ecology 55:111-118.
Coley, P. D., J. P. Bryant, and F. S. Chapin, III. 1985. Resource availability and plant antiherbivore defense. Science (Washington, D.C.) 230:895-899.
Collatz, G. J., J. T. Ball, C. Grivet, and J. A. Berry. 1991. Physiological and environmental regulation of stomatal conductance, photosynthesis and transpiration: a model that includes a laminar boundary layer. Agricultural and Forest Meteorology 54:107-136.
Doebley, J., A. Stec, J. Wendel, and M. Edwards. 1990. Genetic and morphological analysis of a maize-teosinte F2 population: implications for the origin of maize. Proceedings of the Na- tional Academy of Sciences of the USA 87:9888-9892.
Eldridge, N., and S. J. Gould. 1972. Punctuated equilibria: an alternative to phyletic gradualism. Pages 82-115 in T. J. M. Schopf, ed. Models in paleobiology. W. H. Freeman, San Francisco.
Evans, J. R. 1983. Nitrogen and photosynthesis in the flag leaf of wheat (Triticum aestivum L.). Plant Physiology 72:297-302. 1989. Photosynthesis and nitrogen relationships in leaves of C3 plants. Oecologia (Berlin) 78:9-19.
Evans, L. T. 1980. The natural history of crop yield. American Scientist 68:388-397. Farquhar, G. D., and T. D. Sharkey. 1982. Stomatal conductance and photosynthesis. Annual Review
of Plant Physiology 33:317-345. Fenner, M. 1983. Relationships between seed weight, ash content and seedling growth in twenty-four
species of Compositae. New Phytologist 95:697-706. 1985. Seed ecology. Chapman & Hall, London.
Field, C. 1991. Ecological scaling of carbon gain to stress and resource availability. Pages 35-65 in H. A. Mooney, W. E. Winner, and E. J. Pell, eds. Integrated responses of plants to stress. Academic Press, New York.
Field, C., and H. A. Mooney. 1986. The photosynthesis-nitrogen relationship in wild plants. Pages 25-55 in T. J. Givnish, ed. On the economy of plant form and function. Cambridge University Press, Cambridge.
Ford, V. S., and L. D. Gottleib. 1992. Bicalyx is a natural homeotic floral variant. Nature (London) 358:671-673.
Gartside, D. W., and T. McNeilly. 1974. The potential for evolution of heavy metal tolerance in plants. II. Copper tolerance in normal populations of different plant species. Heredity 32:335-348.
Gates, K. 1987. Response of Poa annua populations to competition along a nutrient gradient. M. S. thesis. West Virginia University, Morgantown.
Gifford, R. M., and L. T. Evans. 1981. Photosynthesis, carbon partitioning and yield. Annual Review of Plant Physiology 32:485-509.
Gottleib, L. D. 1984. Genetics and morphological evolution in plants. American Naturalist 123: 681-709.
Grime, J. P. 1977. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. American Naturalist 111:1169-1194.
Harper, J. L. 1977. Population biology of plants. Academic Press, London. Herrera, C. M. 1992. Historical effects and sorting processes as explanations for contemporary ecolog-
ical patterns: character syndromes in Mediterranean woody plants. American Naturalist 140:421-446.
Hobbie, S. E., D. B. Jensen, and F. S. Chapin, III. 1993. Resource supply and disturbance as controls over present and future plant diversity. Pages 385-408 in E.-D. Schulze and H. A. Mooney, eds. Ecosystem function of biodiversity. Springer, Berlin.

EVOLUTION OF SUITES OF TRAITS IN PLANTS S91
Hoffmann, A. A. 1990. Acclimation for desiccation resistance in Drosophila melanogaster and the association between acclimation responses and genetic variation. Journal of Insect Physiol- ogy 36:885-891.
Innes, P., R. D. Blackwell, and S. A. Quarrie. 1984. Some effects of genetic variation in drought- induced abscisic acid accumulation on the yield and water use of spring wheat. Journal of Agricultural Science (Cambridge) 102:341-351.
Jowett, D. 1959. Adaptation of a lead-tolerant population of Agrostis tenuis to low soil fertility. Nature (London) 184:43.
Kauffman, S. 1971. Gene regulation networks: a theory for their global structure and behaviors. Current Topics in Developmental Biology 6:145-182.
Lande, R. 1981. The minimum number of genes contributing to quantitative variation between and within populations. Genetics 99:541-553. 1983. The response to selection on major and minor mutations affecting a metrical trait. Heredity 50:47-65.
Langer, R. H. M. 1966. Mineral nutrition of grasses and cereals. Pages 213-226 in F. L. Milthorpe and J. D. Ivins. The growth of cereals and grasses. Butterworths, London.
Lechowicz, M. J. 1984. Why do temperate deciduous trees leaf out at different times? adaptation and ecology of forest communities. American Naturalist 124:821-842.
Macnair, M. R. 1989. The genetics of metal tolerance in natural populations. Pages 235-254 in J. Shaw, ed. Heavy metal tolerance in plants: evolutionary aspects. CRC, Boca Raton, Fla. 1991. Why the evolution of resistance to anthropogenic toxins normally involves major gene changes: the limits to natural selection. Genetica 84:213-219.
Matthews, M. A., E. Van Volkenburgh, and J. S. Boyer. 1984. Acclimation of leaf growth to low water potentials in sunflower. Plant Cell and Environment 7:199-206.
McNaughton, S. J. 1983. Compensatory plant growth as a response to herbivory. Oikos 40:329-336. Michelena, V. A., and J. S. Boyer. 1982. Complete turgor maintenance at low water potentials in the
elongating region of maize leaves. Plant Physiology 69:1145-1149. Mooney, H. A. 1972. The carbon balance of plants. Annual Review of Ecology and Systematics
3:315-346. Mooney, H. A., and S. L. Gulmon. 1982. Constraints on leaf structure and function in reference to
herbivory. BioScience 32:198-206. Munns, R., H. Greenway, R. Delane, and J. Gibbs. 1982. Ion concentration and carbohydrate status
of the elongating leaf tissue of Hordelum ivulgare growing at high external NaCI. Journal of Experimental Botany 33:574-583.
Nambiar, E. K. S., and D. N. Fife. 1991. Nutrient retranslocation in temperate conifers. Tree Physiol- ogy 9:185-207.
Neill, S. J., and R. Horgan. 1985. Abscisic acid production and water relations in wilty tomato mutants subjected to water deficiency. Journal of Experimental Botany 36:1222-1231.
Nobel, P. S. 1983. Biophysical plant ecology. W. H. Freeman, San Francisco. Nowak, R. S.. and M. M. Caldwell. 1984. A test of compensatory photosynthesis in the field: implica-
tions for herbivory tolerance. Oecologia (Berlin) 61:311-318. Parsons, P. A. 1987. Evolutionary rates under environmental stress. Evolutionary Biology 21:
311-347. Paterson, A. H., E. S. Lander, J. D. Hewitt, S. Peterson, S. E. Lincoln, and S. D. Tanksley. 1988.
Resolution of quantitative traits into Mendelian factors by using a complete linkage map of restriction fragment length polymorphisms. Nature (London) 335:721-726.
Penning de Vries, F. W. T., A. H. M. Brunsting, and H. H. van Laar. 1974. Products, requirements and efficiency of biosynthesis: a quantitative approach. Journal of Theoretical Biology 45:339-377.
Quarrie, S. A. 1981. Genetic variability and heritability of drought-induced abscisic acid accumulation in spring wheat. Plant Cell and Environment 4:147-151. 1982. Droopy: a wilty mutant of potato deficient in abscisic acid. Plant Cell and Environment 5:23-26.
Radin, J. W., L. L. Parker, and G. Guinn. 1982. Water relations of cotton plants under nitrogen

S92 THE AMERICAN NATURALIST
deficiency. V. Environmental control of abscisic acid accumulation and stomatal sensitivity to abscisic acid. Plant Physiology 70:1066-1070.
Raff, R. A., and T. C. Kaufman. 1983. Embryos, genes and evolution. Macmillan, New York. Reekie, E. G., and F. A. Bazzaz. 1987. Reproductive effort in plants. III. Effect of reproduction on
vegetative activity. American Naturalist 129:907-919. Schulze, E.-D. 1986. Carbon dioxide and water vapor exchange in response to drought in the atmo-
sphere and in the soil. Annual Review of Plant Physiology 37:247-274. Shukla, J., C. Nobre, and P. Sellers. 1990. Amazon deforestation and climate change. Science (Wash-
ington, D.C.) 247:1322-1325. Snaydon, R. W., and A. D. Bradshaw. 1962. Differences between natural populations of Trifolium
repens L. in response to mineral nutrients. I. Phosphate. Journal of Experimental Botany 13:422-434.
Stearns, S. C. 1989. The evolutionary significance of phenotypic plasticity. BioScience 39:436-445. Sternberg, P. W., and H. R. Horvitz. 1981. Gonadal cell lineages of the nematode Panagrellus redivivus
and implications for evolution by modification of cell lineage. Developmental Biology 88:147-166.
Termatt, A., J. B. Passioura, and R. Munns. 1985. Shoot turgor does not limit shoot growth of NaCl-affected wheat and barley. Plant Physiology 77:869-872.
Tilman, D. 1988. Plant strategies and the dynamics and structure of plant communities. Princeton University Press, Princeton, N.J.
Tilman, D., and D. Wedin. 1991. Dynamics of nitrogen competition between successional grasses. Ecology 72: 1038-1049.
Veerkamp, M. T., W. J. Corre, B. J. Atwell, and P. J. C. Kuiper. 1980. Growth rate and phosphate utilization of some Carex species from a range of oligotrophic to eutrophic swamp habitats. Physiologia Plantarum 50:237-240.
Vose, P. B. 1984. Effects of genetic factors on nutritional requirements of plants. Pages 67-114 in P. B. Vose and S. G. Blixt, eds. Crop breeding, a contemporary basis. Pergamon, Oxford.
Vrba, E. S., and S. J. Gould. 1986. The hierarchical expansion of sorting and selection: sorting and selection cannot be equated. Paleobiology 12:217-228.
Waddington, C. H. 1942. Canalization of development and the inheritance of acquired characters. Nature (London) 150:563-565.
Wadman-van Schravendijk, H., and 0. M. van Andel. 1985. Interdependence of growth, water relations and abscisic acid level in Phaseolus vulgaris during water-logging. Physiologia Plantarum 63:215-220.
Warwick, S. I. 1991. Herbicide resistance in weedy plants: physiology and population biology. Annual Review of Ecology and Systematics 22:95-114.
Winn, A. A. 1988. Ecological and evolutionary consequences of seed size in Prunella vulgaris. Ecol- ogy 69:1537-1544.
Woodward, F. I. 1987. Climate and plant distribution. Cambridge University Press, Cambridge. Wright, S. 1927. The effects in combination of the major color-factors of the guinea pig. Genetics
12:530-569. Wu, L., A. D. Bradshaw, and D. A. Thurman. 1975. The potential for evolution of heavy metal
tolerance in Agrostis stolonifera. Heredity 34:165-187.





























