Embed Size (px)
Citation preview

JOURNAL OF CELLULAR PHYSIOLOGY 134:137-142 (1988)
Temporal Relationship of the Prolactin-Dependent LH-Induced LH Receptor to the LH Stimulus
MOHAMED 0. HUSSEIN AND WILLIAM B. ZIPF* Departments of Pediatrics and Physiology, Children’s Hospital, Ohio State University,
Columbus, Ohio 43205
The time course for LH induction of luteinizing hormone (LH) receptors as reflected in binding of 1251-labeled hCG was investigated in hypophysecto- mized adult male rats. A low dose of oLH (10 p g ) was administered to hy- pophysectomized adult male rats following pretreatments with prolactin, follicle-stimulating hormone (FSH), growth hormone (CH), or saline. Testicular binding of hCG was determined at different times following the LH injection using Leydig cell membrane preparations from a testicular homogenate. Seven days after hypophysectomy, hCG binding was at a nadir of 19 & 7% (mean SD) of control values. Pretreatment with prolactin (100 pg/day) for 7 days was associated with a nonsignificantly different hCG binding that was 30 f 5% of control values. Prolactin pretreatment plus a single 10 p g LH i.p. injection increased lZ5l hCG binding up to 56 k 10% of control values within 30 minutes of the LH injection. Luteinizing hormone-induced hCG binding per- sisted at a high level (51 k 4% of control values) for 2 hours but returned to hypophysectomized control levels 6 hours after the i.p. LH injection.
Seven days pretreatment with FSH or GH at 100 &day plus IO-pg LH injections was also tested. Neither FSH nor GH had a statistically significant effect on hCG binding nor could they mimic the ability of prolactin to allow for LH induction of hCG binding in the hypophysectomized adult male rats.
These studies suggest that the induction or ”up-regulation” of Leydig cell hCG binding by ovine LH is rapid and specifically dependent upon pre- exposure to prolactin.
The regulation of LH receptors has been shown to be dependent upon luteinizing hormone (LH), prolactin (PRL), follicle-stimulating hormone (FSH), and growth hormone (GH) (Bohnet et al., 1976; Chan et al., 1981; Chen et al., 1977; Erickson et al., 1982; Huhtaniemi et al., 1981; Morris and Saxena 1980; Zipf et al., 1978a; Zipf and Berntson, 1981). However, the effects are vari- able depending upon the animal’s age, presence or ab- sence of the pituitary gland or pituitary hormonal replacements. A preponderance of evidence has accu- mulated suggesting that one of the most important hor- mones regulating the number of LH receptors is its homologous hormone, LH. At high doses, LH has a pro- nounced ability to “down regulate” the LH receptor in both the normal and hypophysectomized rat as well as in cultured Leydig cell or granulosa cell systems (Nozu et al., 1981; Jia and Hsueh, 1984). More detailed studies have shown an ability of low and possibility more phys- iologic doses of LH to “up-regulate” or induce its own receptor. Chan et al. (1981) and Huhtaniemi et al. (1981) have shown that low-dose LH injections (10 pglday) in- crease the LH receptor level within 40 to 120 minutes followed by a prolonged period of down-regulation.
Additional studies have suggested a supportive role for other pituitary hormones in the control of the LH receptor. Prolactin may have isolated effects or work in
0 1988 ALAN R. LISS, INC
concert with LH in control of the LH receptor. Prolactin pretreatment mitigates the ability of LH to down-regu- late LH receptors and appears to unmask the ability of low doses of LH to induce new LH receptors in the hypophysectomized male rat (Zipf et al., 1978a; Zipf and Berntson, 1981; Zipf and Wukie, 1983). FSH has also been reported to have an effect on the LH receptor in both the male and female, although in the male this effect seems to be limited to the immature animal. FSH was reported to increase the number of LH receptors and steroid response to LH in immature intact and im- mature hypophysectomized male rats (Chen et al., 1977) and in cultured granulosa cells from female rats (Erick- son et al., 1982), but other studies in the adult male rat have been unable to demonstrate a n effect of FSH on LH/hCG binding (Zipf et al., 1978a) suggesting that FSH plays an important role in Leydig cell differentiation but not in LH receptor maintenance once an animal has achieved sexual maturity.
Received April 28,1987; accepted September 14,1987.
*To whom reprint requests/correspondence should be addressed, at Pediatric Endocrinology, Children’s Hospital, 700 Children’s Drive, Columbus, Ohio 43205.
HUSSEIN AND ZIPF 138
I
Z 30 m
20
10
0 i i ; , L.H. Hours After Last LH Injection
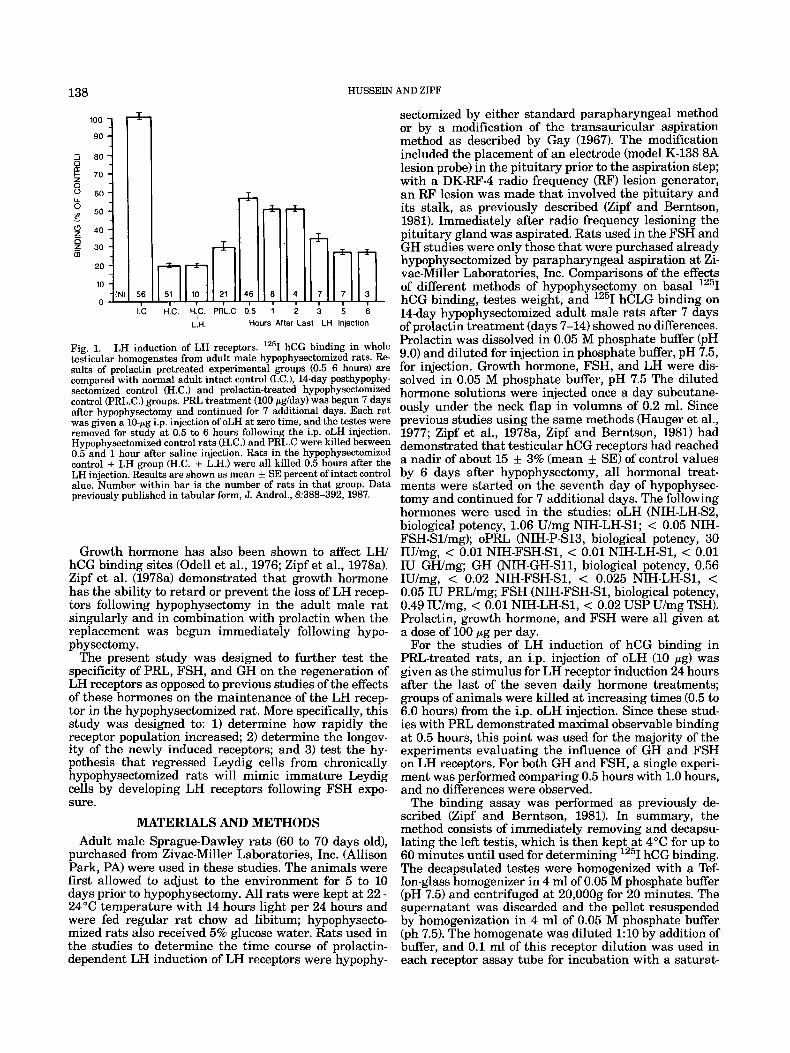
Fig. 1. LH induction of LH receptors. lZ5I hCG binding in whole testicular homogenates from adult male hypophysectomized rats. Re- sults of prolactin pretreated experimental groups (0.5-6 hours) are compared with normal adult intact control (I.C.), 14-day posthypophy- sectomized control (H.C.) and prolactin-treated hypophysectomized control (PRL.C.) groups. PRL treatment (100 pg/day) was begun 7 days after hypophysectomy and continued for 7 additional days. Each rat was given a 10-pg i.p. injection of oLH at zero time, and the testes were removed for study at 0.5 to 6 hours following the i.p. oLH injection. Hypophysectomized control rats (H.C.) and PRL.C were killed between 0.5 and 1 hour after saline injection. Rats in the hypophysectomized control + LH group (H.C. + L.H.) were all killed 0.5 hours after the LH injection. Results are shown as mean k SE percent of intact control alue. Number within bar is the number of rats in that group. Data previously published in tabular form, J. Androl., 8:388-392, 1987.
Growth hormone has also been shown to affect LW hCG binding sites (Ode11 et al., 1976; Zipf et al., 1978a). Zipf et al. (1978a) demonstrated that growth hormone has the ability to retard or prevent the loss of LH recep- tors following hypophysectomy in the adult male rat singularly and in combination with prolactin when the replacement was begun immediately following hypo- phy sectomy.
The present study was designed to further test the specificity of PRL, FSH, and GH on the regeneration of LH receptors as opposed to previous studies of the effects of these hormones on the maintenance of the LH recep- tor in the hypophysectomized rat. More specifically, this study was designed to: 1) determine how rapidly the receptor population increased; 2) determine the longev- ity of the newly induced receptors; and 3) test the hy- pothesis that regressed Leydig cells from chronically hypophysectomized rats will mimic immature Leydig cells by developing LH receptors following FSH expo- sure.
MATERIALS AND METHODS Adult male Sprague-Dawley rats (60 to 70 days old),
purchased from Zivac-Miller Laboratories, Inc. (Allison Park, PA) were used in these studies. The animals were first allowed to adjust to the environment for 5 to 10 days prior to hypophysectomy. All rats were kept at 22- 24°C temperature with 14 hours light per 24 hours and were fed regular rat chow ad libitum; hypophysecto- mized rats also received 5% glucose water. Rats used in the studies to determine the time course of prolactin- dependent LH induction of LH receptors were hypophy-
sectomized by either standard parapharyngeal method or by a modification of the transauricular aspiration method as described by Gay (1967). The modification included the placement of an electrode (model K-138 8A lesion probe) in the pituitary prior to the aspiration step; with a DK-RF-4 radio frequency (RF) lesion generator, an RF lesion was made that involved the pituitary and its stalk, as previously described (Zipf and Berntson, 1981). Immediately after radio frequency lesioning the pituitary gland was aspirated. Rats used in the FSH and GH studies were only those that were purchased already hypophysectomized by parapharyngeal aspiration at Zi- vac-Miller Laboratories, Inc. Comparisons of the effects of different methods of hypophysectomy on basal lZ5I hCG binding, testes weight, and lz5I hCLG binding on 14-day hypophysectomized adult male rats after 7 days of prolactin treatment (days 7-14) showed no differences. Prolactin was dissolved in 0.05 M phosphate buffer (pH 9.0) and diluted for injection in phosphate buffer, pH 7.5, for injection. Growth hormone, FSH, and LH were dis- solved in 0.05 M phosphate buffer, pH 7.5 The diluted hormone solutions were injected once a day subcutane- ously under the neck flap in volumns of 0.2 ml. Since previous studies using the same methods (Hauger et al., 1977; Zipf et al., 1978a, Zipf and Berntson, 1981) had demonstrated that testicular hCG receptors had reached a nadir of about 15 3% (mean f SE) of control values by 6 days after hypophysectomy, all hormonal treat- ments were started on the seventh day of hypophysec- tomy and continued for 7 additional days. The following hormones were used in the studies: oLH (NIH-LH-S2, biological potency, 1.06 U/mg NIH-LH-S1; < 0.05 NIH- FSH-Sl/mg); oPRL (NIH-P-S13, biological potency, 30 IU/mg, < 0.01 NIH-FSH-S1, < 0.01 NIH-LH-S1, < 0.01 IU GWmg; GH (NIH-GH-S11, biological potency, 0.56 IU/mg, < 0.02 NIH-FSH-S1, < 0.025 NIH-LH-S1, < 0.05 IU PRL/mg; FSH (NIH-FSH-S1, biological potency, 0.49 IU/mg, < 0.01 NIH-LH-S1, < 0.02 USP U/mg TSH). Prolactin, growth hormone, and FSH were all given at a dose of 100 pg per day.
For the studies of LH induction of hCG binding in PRL-treated rats, an i.p. injection of oLH (10 pg) was given as the stimulus for LH receptor induction 24 hours after the last of the seven daily hormone treatments; groups of animals were killed at increasing times (0.5 to 6.0 hours) from the i.p. oLH injection. Since these stud- ies with PRL demonstrated maximal observable binding at 0.5 hours, this point was used for the majority of the experiments evaluating the influence of GH and FSH on LH receptors. For both GH and FSH, a single experi- ment was performed comparing 0.5 hours with 1.0 hours, and no differences were observed.
The binding assay was performed as previously de- scribed (Zipf and Berntson, 1981). In summary, the method consists of immediately removing and decapsu- lating the left testis, which is then kept a t 4°C for up to 60 minutes until used for determining 1251 hCG binding. The decapsulated testes were homogenized with a Tef- lon-glass homogenizer in 4 ml of 0.05 M phosphate buffer (pH 7.5) and centrifuged at 20,OOOg for 20 minutes. The supernatant was discarded and the pellet resuspended by homogenization in 4 ml of 0.05 M phosphate buffer (ph 7.5). The homogenate was diluted 1 : l O by addition of buffer, and 0.1 ml of this receptor dilution was used in each receptor assay tube for incubation with a saturat-
PRL-DEPENDENT LH INDUCTION OF LH RECEPTORS 139 100-
90-
80-
70 - 60-
50-
40 - 30-
20 - 10 - 0
+
+ - - . . - - +
-(N) 56 51 , 10 4 4 4 4 I I I I , I - J-5 12 24
+ LH Hours After Last FSH Injection
*Assayed 112 hour after LH injection
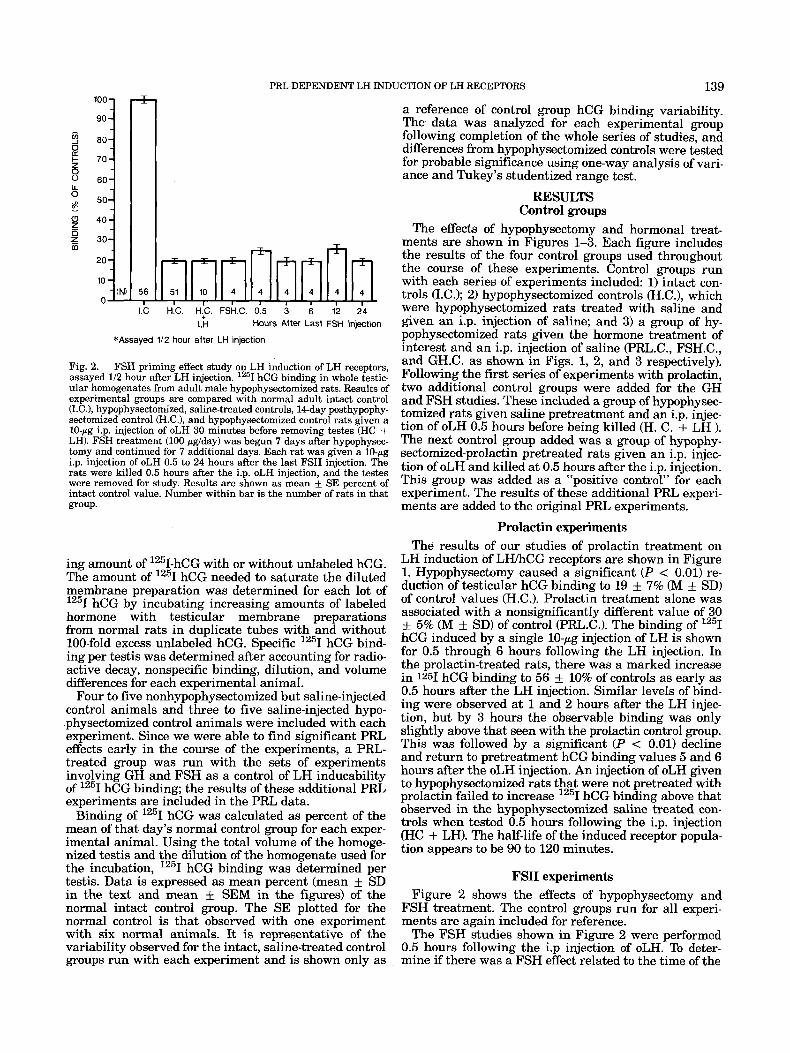
Fig. 2. FSH priming effect study on LH induction of LH receptors, assayed 1/2 hour after LH injection. 1251 hCG binding in whole testic- ular homogenates from adult male hypophysectomized rats. Results of experimental groups are compared with normal adult intact control U.C.), hypophysectomized, saline-treated controls, 14-day posthypophy- sectomized control cH.C.), and hypophysectomized control rats given a 10-pg i.p. injection of oLH 30 minutes before removing testes (HC + LH). FSH treatment (100 pglday) was begun 7 days after hypophysec- tomy and continued for 7 additional days. Each rat was given a 10-pg i.p. injection of oLH 0.5 to 24 hours after the last FSH injection. The rats were killed 0.5 hours after the i.p. oLH injection, and the testes were removed for study. Results are shown as mean f SE percent of intact control value. Number within bar is the number of rats in that group.
ing amount of 1251-hCG with or without unlabeled hCG. The amount of 1251 hCG needed to saturate the diluted membrane preparation was determined for each lot of 1251 hCG by incubating increasing amounts of labeled hormone with testicular membrane preparations from normal rats in duplicate tubes with and without 100-fold excess unlabeled hCG. Specific 1251 hCG bind- ing per testis was determined after accounting for radio- active decay, nonspecific binding, dilution, and volume differences for each experimental animal.
Four to five nonhypophysectomized but saline-injected control animals and three to five saline-injected hypo- physectomized control animals were included with each experiment. Since we were able to find significant PRL effects early in the course of the experiments, a PRL- treated group was run with the sets of experiments involving GH and FSH as a control of LH inducability of 1251 hCG binding; the results of these additional PRL experiments are included in the PRL data.
Binding of 1251 hCG was calculated as percent of the mean of that day’s normal control group for each exper- imental animal. Using the total volume of the homoge- nized testis and the dilution of the homogenate used for the incubation, 1251 hCG binding was determined per testis. Data is expressed as mean percent (mean + SD in the text and mean + SEM in the figures) of the normal intact control group. The SE plotted for the normal control is that observed with one experiment with six normal animals. It is representative of the variability observed for the intact, saline-treated control groups run with each experiment and is shown only as
a reference of control group hCG binding variability. The data was analyzed for each experimental group following completion of the whole series of studies, and differences from hypophysedomized controls were tested for probable significance using one-way analysis of vari- ance and Tukey’s studentized range test.
RESULTS Control groups
The effects of hypophysectomy and hormonal treat- ments are shown in Figures 1-3. Each figure includes the results of the four control groups used throughout the course of these experiments. Control groups run with each series of experiments included: 1) intact con- trols (I.C.1; 2) hypophysectomized controls (H.C.), which were hypophysectomized rats treated with saline and given an i.p. injection of saline; and 3) a group of hy- pophysectomized rats given the hormone treatment of interest and an i.p. injection of saline (PRL.C., FSH.C., and GH.C. as shown in Figs. 1, 2, and 3 respectively). Following the first series of experiments with prolactin, two additional control groups were added for the GH and FSH studies. These included a group of hypophysec- tomized rats given saline pretreatment and an i.p. injec- tion of oLH 0.5 hours before being killed (H. C. + LH ). The next control group added was a group of hypophy- sectomized-prolactin pretreated rats given an i.p. injec- tion of oLH and killed at 0.5 hours after the i.p. injection. This group was added as a “positive control” for each experiment. The results of these additional PRL experi- ments are added to the original PRL experiments.
Prolactin experiments The results of our studies of prolactin treatment on
LH induction of LWhCG receptors are shown in Figure 1. Hypophysectomy caused a significant (P < 0.01) re- duction of testicular hCG binding to 19 & 7% (M f SD) of control values (H.C.). Prolactin treatment alone was associated with a nonsignificantly different value of 30 k 5% (M + SD) of control (PRL.C.1. The binding of 1251 hCG induced by a single 10-pg injection of LH is shown for 0.5 through 6 hours following the LH injection. In the prolactin-treated rats, there was a marked increase in 1251 hCG binding to 56 k 10% of controls as early as 0.5 hours after the LH injection. Similar levels of bind- ing were observed at 1 and 2 hours after the LH injec- tion, but by 3 hours the observable binding was only slightly above that seen with the prolactin control group. This was followed by a significant (P < 0.01) decline and return to pretreatment hCG binding values 5 and 6 hours after the oLH injection. An injection of oLH given to hypophysectomized rats that were not pretreated with prolactin failed to increase 1251 hCG binding above that observed in the hypophysectomized saline treated con- trols when tested 0.5 hours following the i.p. injection (HC + LH). The half-life of the induced receptor popula- tion appears to be 90 to 120 minutes.
FSH experiments Figure 2 shows the effects of hypophysectomy and
FSH treatment. The control groups run for all experi- ments are again included for reference.
The FSH studies shown in Figure 2 were performed 0.5 hours following the i.p injection of oLH. To deter- mine if there was a FSH effect related to the time of the
HUSSEIN AND ZIPF 140
100
90
c 40
m 20
10 56 - 11 I] 1.k H k . H:C. GH.C 0:5
3 3 2 6 Jl 12 24
Lh. Hours After Last GH Injection
*Assayed 112 hour after LH injection
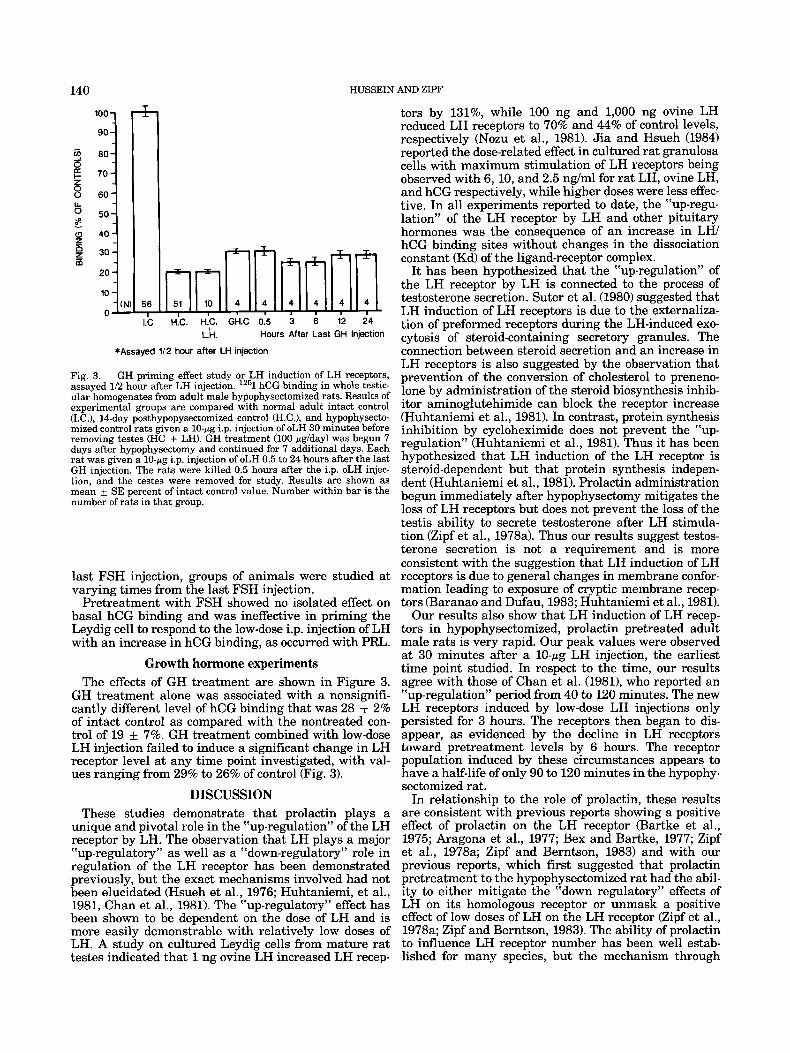
Fig. 3. GH priming effect study or LH induction of LH receptors, assayed 1/2 hour after LH injection. lZ5I hCG binding in whole testic- ular homogenates from adult male hypophysectomized rats. Results of experimental groups are compared with normal adult intact control (I.C.), 14-day posthypopysectomized control (H.C.), and hypophysecto- mized control rats given a 10-pg i.p. injection of oLH 30 minutes before removing testes (HC + LH). GH treatment (100 pg/day) was begun 7 days after hypophysectomy and continued for 7 additional days. Each rat was given a 10-pg i.p. injection of oLH 0.5 to 24 hours after the last GH injection. The rats were killed 0.5 hours after the i.p. oLH injec- tion, and the testes were removed for study. Results are shown as mean k SE percent of intact control value. Number within bar is the number of rats in that group.
last FSH injection, groups of animals were studied at varying times from the last FSH injection.
Pretreatment with FSH showed no isolated effect on basal hCG binding and was ineffective in priming the Leydig cell to respond to the low-dose i.p. injection of LH with an increase in hCG binding, as occurred with PRL.
Growth hormone experiments The effects of GH treatment are shown in Figure 3.
GH treatment alone was associated with a nonsignifi- cantly different level of hCG binding that was 28 f 2% of intact control as compared with the nontreated con- trol of 19 k 7%. GH treatment combined with low-dose LH injection failed to induce a significant change in LH receptor level at any time point investigated, with val- ues ranging from 29% to 26% of control (Fig. 3).
DISCUSSION These studies demonstrate that prolactin plays a
unique and pivotal role in the “up-regulation” of the LH receptor by LH. The observation that LH plays a major “up-regulatory” as well as a “down-regulatory” role in regulation of the LH receptor has been demonstrated previously, but the exact mechanisms involved had not been elucidated (Hsueh et al., 1976; Huhtaniemi, et al., 1981, Chan et al., 1981). The “up-regulatory” effect has been shown to be dependent on the dose of LH and is more easily demonstrable with relatively low doses of LH. A studv on cultured Levdig cells from mature rat
tors by 131%, while 100 ng and 1,000 ng ovine LH reduced LH receptors to 70% and 44% of control levels, respectively (Nozu et al., 1981). Jia and Hsueh (1984) reported the dose-related effect in cultured rat granulosa cells with maximum stimulation of LH receptors being observed with 6,10, and 2.5 ng/ml for rat LH, ovine LH, and hCG respectively, while higher doses were less effec- tive. In all experiments reported to date, the “up-regu- lation” of the LH receptor by LH and other pituitary hormones was the consequence of an increase in LW hCG binding sites without changes in the dissociation constant (Kd) of the ligand-receptor complex.
It has been hypothesized that the “up-regulation” of the LH receptor by LH is connected to the process of testosterone secretion. Suter et al. (1980) suggested that LH induction of LH receptors is due to the externaliza- tion of preformed receptors during the LH-induced exo- cytosis of steroid-containing secretory granules. The connection between steroid secretion and an increase in LH receptors is also suggested by the observation that prevention of the conversion of cholesterol to preneno- lone by administration of the steroid biosynthesis inhib- itor aminoglutehimide can block the receptor increase (Huhtaniemi et al., 1981). In contrast, protein synthesis inhibition by cycloheximide does not prevent the Yip- regulation” (Huhtaniemi et al., 1981). Thus it has been hypothesized that LH induction of the LH receptor is steroid-dependent but that protein synthesis indepen- dent (Huhtaniemi et al., 1981). Prolactin administration begun immediately after hypophysectomy mitigates the loss of LH receptors but does not prevent the loss of the testis ability to secrete testosterone after LH stimula- tion (Zipf et al., 1978a). Thus our results suggest testos- terone secretion is not a requirement and is more consistent with the suggestion that LH induction of LH receptors is due to general changes in membrane confor- mation leading to exposure of cryptic membrane recep- tors (Baranao and Dufau, 1983; Huhtaniemi et al., 1981).
Our results also show that LH induction of LH recep- tors in hypophysectomized, prolactin pretreated adult male rats is very rapid. Our peak values were observed at 30 minutes after a 10-pg LH injection, the earliest time point studied. In respect to the time, our results agree with those of Chan et al. (1981), who reported an “up-regulation” period from 40 to 120 minutes. The new LH receptors induced by low-dose LH injections only persisted for 3 hours. The receptors then began to dis- appear, as evidenced by the decline in LH receptors toward pretreatment levels by 6 hours. The receptor population induced by these circumstances appears to have a half-life of only 90 to 120 minutes in the hypophy- sectomized rat.
In relationship to the role of prolactin, these results are consistent with previous reports showing a positive effect of prolactin on the LH receptor (Bartke et al., 1975; Aragona et al., 1977; Bex and Bartke, 1977; Zipf et al., 1978a; Zipf and Berntson, 1983) and with our previous reports, which first suggested that prolactin pretreatment to the hypophysectomized rat had the abil- ity to either mitigate the “down regulatory” effects of LH on its homologous receptor or unmask a positive effect of low doses of LH on the LH receptor (Zipf et al., 1978a; Zipf and Berntson, 1983). The ability of prolactin to influence LH receptor number has been well estab-
testes indicked that 1 ng ovi”ne LH increased LH recep- lished for many species, but the mechanism through

PRL-DEPENDENT LH INDUCTION OF LH RECEPTORS 141
which it functions has not yet been defined. Prolactin alone has been shown to allow for a persistence of LH receptors in the acutely hypophysectomized rat (Zipf et al., 1978b), suggesting prolactin plays a maintenance role. Other studies using a more chronic hypophysec- tomized rat model have demonstrated the ability of pro- lactin to increase or induce hCG binding independent of any other hormonal preparations (Zipf and Berntson, 1981). Our current series of studies also shows a higher level of hCG binding in some animals treated with pro- lactin alone as opposed to saline, and the percent of control values is almost identical to our previous reports (Zipf and Berntson, 1981; Zipf and Wukie, 1983). How- ever, using more conservative statistical analysis and comparing the results with hypophysectomized rats that show slightly greater residual binding (19% vs. 15%), the prolactin-treated group as a whole in this series of studies was not different from the hypophysectomized saline treated group. In addition, the variability in the treated group makes it difficult to demonstrate a pure prolactin effect. However, the emerging role for very low concentrations of LH to regulate the LH receptor sug- gested to us that the variability might be a consequence of residual but variable LH secretion occuring in some of the hypophysectomized animals. Gross and Page (1979) have shown that in the chronic hypophysecto- mized animal, the pars tuberalis is not always removed, and this residual pituitary stalk tissues has functioning gonadotrophs. To test this possibility, we evaluated for a difference in prolactin-dependent hCG binding in rats hypophysectomized with the standard posterior pharyn- geal method and in rats in which the pars tuberalis was initially destroyed with RF lesioning using stereotaxic techniques before aspiration of the pituitary gland. De- spite this addition to the hypophysectomy method, we did not find a difference between the animals in which the hypophysectomy had been performed using standard methods and those in which the hypothalamus, stalk, and pituitary were first RF-lesioned. Another possibility for consideration is that there is suffkient LH in the prolactin preparations used to allow for an LH “up- regulatory” effect to be observed in the supposed “non- LH” treated rats. In this instance, however, we could normally expect to see the same effect in all animals. Although the question of an independent prolactin effect in the chronic hypophysectomized rat still remains to be answered, the evidence still suggests that the “up-regu- latory” effect of LH is wholly dependent upon prior and recent exposure of the Leydig cell to prolactin.
The mechanism of control of LH receptors by FSH was investigated by Erickson et al. (19821, using granulosa cells from immature hypophysectomized estrogen- primed rats. Freshly collected granulosa cells show a low specific binding to labeled hCG (0.18 fmole/million cells). A marked increase in hCG binding was observed when the cells were cultured in serum-free media with 10 ng/ml FSH, with maximal binding attained after 3 days of incubation (19.2 fmole/million cells). Cholera toxin, prostaglandin (PGEz), and cyclic nucleotide ana- logues mimicked, in part, the FSH effect with maximum induction of hCG binding sites closely parallel with their maximum stimulating effects on cyclic AMP formation. A study by Chen et al. (1977) indicated that at 40-80 pg FSH per day in intact 15-day-old male rats significantly increased the number of LH receptors but did not alter testicular responsiveness to LH.
Although these previous studies indicate a positive effect of FSH on the LH receptor in immature rats, our results showed no significant effect of FSH on LH recep- tors in the mature hypophysectomized rat. The lack of an observable FSH effect suggests that even after chronic hypophysectomy, the Leydig cell or residual Leydig cell precursors do not revert to a primordial or fetal condition that is once again responsive to FSH.
Growth hormone has also been shown to have effects on the LH receptor in the mature rat. Our previous studies have demonstrated a substantial effect of growth hormone under different experimental condittions (Zipf et al., 1978b; Zipf and Berntson, 1981; Zipf and Wukie, 1983). Growth hormone injections (100 pg/day) started on the first day after hypophysectomy in mature male rats maintained LH receptors at 50% of control levels 7 days after hypophysectomy, independent of any other pituitary hormone replacements. This effect was addi- tive to those of prolactin, suggesting these two hormones played different roles. The results of this series of stud- ies support the conclusion that PRL and GH effect LH receptors in different ways. We found that after hypo- physectomy induced regression of the Leydig cells and loss of LH receptors, GH did not exert a significant effect on hCG binding. In these studies we clearly did not see the development of a LH receptor population to the 50% level, as is seen when GH is started immediately after hypophysectomy and continued for 1 week. However, as with prolactin, there was a suggestion of some effect, and it is possible that the statistical method used and the higher baseline binding of the hypophysectomized controls may have obscured a modest positive effect. Vari- ability in the ability of GH to affect LH receptors has been noted previously (Bohnet and Friesen, 1976). Whether or not there is a subtle growth hormone effect on the basal levels of Leydig cell LH receptors of the regressed testis from the chronic hypophysectomized rat is yet to be clarified; however, our studies clearly dem- onstrate an absence of any LH and GH synergism, an effect clearly seen with prolactin.
In summary, these studies demonstrate that the Ley- dig cell “up-regulation” of LWhCG binding response induced by LH is rapid, short-lived, and specifically de- pendent upon pre-exposure to prolactin.
These observations are important to both the design and interpretation of experiments on this hormone-de- pendent event; they help establish the uniqueness of prolactin in the control of receptor membrane density, and they suggest that the primary effect of LH, in re- spect to receptor control, is to induce or maintain the LH receptors on the Leydig cell membrane, as opposed to its role “down-regulating” the receptor.
ACKNOWLEDGMENTS This work was supported through CHRF Grant 74-
220.
LITERATURE CITED Baranao, J.L.S., and Dufau, M.L. (1983) Gonadotropin-induced changes
in the luteinizing hormone receptors of cultured Leydig cells. J. Biol. Chem., 258:7322-7330.
Bohnet, H.G., and Friesen, H.G. (1976) Effect of prolactin and growth hormone on prolactin and LH receptors in the dwarf mouse. J. Re- prod. Fertil., 48:307-311.
Chan. V., Katikineni, M., Davies, T.F., and Catt. K.J. (1981) Hormonal regulation of testicular luteinizing hormone and prolactin receptors. Endocrinology, 108:1607-1612.

142 HUSSEIN AND ZIPF
Chen, Y.I., Shaw, M.J., and Payne, A.H. (1977) Steroid and FSH action on LH receptors and LH-sensitive testicular responsiveness during sexual maturation of the rat. Mol. Cell. Endocrinol., 8:291-299.
Erickson, G.G., Wang C., Casper, R., Mattson, G., and Hofeditz, C. (1982) Studies on the mechanisms of LH receptor control by FSH. Mol. Cell. Endocrinol. 27:17-30.
Gay, V.L. (1967) A stereotaxic approach to transauricular hypophysec- tomy in the rat. Endocrinology, 81:1177-1179.
Hauger, R.L., Chen, Y.D.I., Kelch, R.P. and Payne, A.H. (1977) Pitu- itary regulation of Leydig cell function in the adult male rat. J . Endocrinol. 74:57-66.
Hsueh, A.J.W., Dufau, M.L., and Catt, K.J. (1976) Regulation of lutein- izing hormone receptors in testicular intesitial cells by gonadotropin. Biochem. Biophys. Res. 72:1145-1152.
Huhtaniemi, I., Katikineni, M., and Catt, K.J. (1981) Regulation of luteinizing hormone receptors and steroidogenesis in neonatal rat testis. Endocrinology, 109~588-595.
Jia, X., and Hsueh, A.J.W (1984) Homologous regulation of hormone receptors: Luteinizing hormone increases its own receptors in cul- tured rat granulosa cells. Endocrinology, 1152433-2439.
Leinonen, P., Bolton, N., and Vihko, R. (1982) Human testicular LH receptors: Correlation with circulating gonadotropins and testicular steroid secretion. Int. J. Androl., 5:145-157.
Morris, P.L., and Saxena, B.B. (1980) Dose and age dependent effects of PRL on LH and PRL binding sites in rat Leydig cell homogenates.
Endocrinology, 107~1639-1645. Nozu. D.. Dehejia, A,, Zawistowich, L., Catt, K.J., and Dufau, M.L.
(1981) Gonadotropin-induced receptor regulation and steroidogenic lesions in cultured Leydig cells. J. Biol. Chem., 256~12875-12882.
Suter, D.E., Fletcher, P.W., Sluss, P.M., Reichert, L.E., and Niswender, G.D. (1980) Alterations in the number of ovine luteal receptors for LH and progesterone secretion induced by homologous hormone. Biol. Reprod., 22:205-209.
Warren, D.W., Dufau, M.L., and Catt, K.L. (1982) Hormonal realation of gonadotropin receptors and steroidogenesis in cultured Fetal rat testes. Science, 218:375-377.
Zipf, W.B., and Berntson, G.C., (1981) Hormone dependent subpopula- tion of rat testicular luteinizing hormone (LH) receptors. Biol. Re- prod., 24:306-310.
Zipf. W.B.. and Wukie, J.J. (1983) Role of Drolactin and mowth hor- mone in'the maintenance of normal Leydig cell function. In: Male Reproduction and Fertility. A. Negro-Vilar, ed. Raven Press, New York, pp. 65-75.
Zipf, W.B., Payne, A.H., and Kelch, R.P. (1978a) Prolactin, growth hormone and luteinizing hormone in the maintenance of testicular luteinizing hormone receptors. Endocrinology, 103:595-600.
Zipf, W.B., Payne, A.H., and Kelch, R.P. (1978b) Dissociation of lutro- pin-induced loss of testicular lutropin receptors and lutropin-induced desensitization of testosterone synthesis. Biochem. Biophys. Acta, 540~330-336.





![Update in Anaesthesia [14] · Increased hepatic glucose output. Figure 3. The widespread effects of the pituitary gland FSH LH Anterior pituitary TSH Prolactin ACTH Growth hormone](https://img.pdfslide.net/doc/110x75/5f9b863faf8dea73ca4dcae7/update-in-anaesthesia-14-increased-hepatic-glucose-output-figure-3-the-widespread.jpg)























