Embed Size (px)
Citation preview

Research Article
TFAP2A Regulates Nasopharyngeal Carcinoma Growth andSurvival by Targeting HIF-1a Signaling Pathway
Dingbo Shi1, Fangyun Xie1, Yun Zhang1, Yun Tian1, Wangbing Chen1, Lingyi Fu1, Jingshu Wang1,Wei Guo3, Tiebang Kang1, Wenlin Huang1,2, and Wuguo Deng1,2,3
AbstractTFAP2A is a transcription factor that orchestrates a variety of cell processes, including cell growth
and tissue differentiation. However, the regulation of TFAP2A in human nasopharyngeal carcinoma
tumorigenesis and its precise mechanism of action remain largely unknown. In this study, we
investigated the biologic role and clinical significance of TFAP2A in nasopharyngeal carcinoma growth
and progression and identified the underlying molecular mechanisms. We found that TFAP2A
was highly expressed in various nasopharyngeal carcinoma cell lines and tumor tissue specimens
and was significantly correlated with hypoxia-inducible factor-1a (HIF-1a) expression. A positive
correlation of TFAP2A overexpression with advanced tumor stage, local invasion, clinical progression,
and poor prognosis of patients with nasopharyngeal carcinomas were also observed. Moreover, we
found that knockdown of TFAP2A expression by siRNA significantly inhibited tumor cell growth in
nasopharyngeal carcinoma cell lines and in a subcutaneous xenograft mouse model by targeting
the HIF-1a–mediated VEGF/pigment epithelium–derived factor (PEDF) signaling pathway. Treatment
of nasopharyngeal carcinoma cells with TFAP2A siRNA dramatically inhibited the expression and
the release of VEGF protein but did not change the level of PEDF protein, resulting in a significant
reduction of the ratio of VEGF/PEDF. Pretreatment with a HIF-1a siRNA did not significantly change
the TFAP2A siRNA-mediated inhibition in cell viability. Our results indicate that TFAP2A regulates
nasopharyngeal carcinoma growth and survival through the modulation of the HIF-1a–mediated
VEGF/PEDF signaling pathway, and suggest that TFAP2A could be a potential prognostic biomarker
and therapeutic target for nasopharyngeal carcinoma treatment. Cancer Prev Res; 7(2); 266–77. �2013
AACR.
IntroductionNasopharyngeal carcinoma is a squamous epithelial can-
cer arising from the lateral wall surface of nasopharynx (1).Unlike other head and neck cancers, nasopharyngeal car-cinoma shows a clear regional and racial prevalence. Theincidence rate of nasopharyngeal carcinoma is very high inChina, Japan, and other Southeast Asian countries. But thesouthern Chinese have one of the highest incidences of
nasopharyngeal carcinoma in the world (2). Etiology ofnasopharyngeal carcinoma is considered to be related witha complex interaction of environmental and genetic factorsas well as Epstein–Barr virus infection (3). At present,treatment of nasopharyngeal carcinoma is usually via radio-therapy. Nasopharyngeal carcinoma is more sensitive toionizing radiation than other cancers (4). However, thetreatment success mostly depends on the tumor, node, andmetastasis (TNM) stage classification (5), which tend to bein the advanced stages at the point of diagnosis because theprimary anatomical site of cancer growth is located in thesilent painless area (6). Moreover, nasopharyngeal carcino-ma has a poor prognosis because of late presentation oflesions, poor understanding of the molecular mechanisms,no suitable markers for early detection, and poor responseto available therapies (7, 8). Therefore, there is an urgentneed for further understanding the molecular mechanismsin nasopharyngeal carcinoma tumorigenesis and for iden-tifying effective prognostic and diagnostic biomarkersand new therapeutic targets to improve the prognosis ofpatients.
TFAP2A, also called AP-2a, is a member of the AP-2transcription factor family proteins, which includes 5
Authors' Affiliations: 1Sun Yat-sen University Cancer Center, State KeyLaboratory of Oncology in South China, Collaborative Innovation Center ofCancer Medicine; 2State Key Laboratory of Targeted Drug for Tumors ofGuangdong Province, Guangzhou Double Bioproduct Inc., Guangzhou;and 3Institute of Cancer Stem Cell, Dalian Medical University CancerCenter, Dalian, China
Note:Supplementary data for this article are available atCancer PreventionResearch Online (http://cancerprevres.aacrjournals.org/).
Corresponding Authors: Wuguo Deng, Sun Yat-sen University CancerCenter, 651 Dongfeng East Road, Guangzhou 510060, China. Phone: 86-20-87343170; Fax: 86-20-87343170; E-mail: [email protected]; andWenlin Huang, [email protected]
doi: 10.1158/1940-6207.CAPR-13-0271
�2013 American Association for Cancer Research.
CancerPreventionResearch
Cancer Prev Res; 7(2) February 2014266
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271

different yet closely related 50 kDa proteins known as: AP-2a, AP-2b, AP-2g , AP-2d, and AP-2e (9–12). AP-2 factorsorchestrate a variety of cell processes, including cell growth,apoptosis, and tissue differentiation during embryogenesis(13). The AP-2 family is a DNA-binding protein (14)that presents a conserved helix–span–helix dimerizationdomain preceded by a DNA binding and a transactivationdomain (15). AP-2 homodimers and heterodimers canactivate transcription via GC-rich DNA sequences (16,17). AP-2 has been shown to bind the palindromic con-sensus sequence 50-GCCN3GGC-30, found in various cel-lular and viral enhancers. AP-2 family is known to exhibitboth activating and repressing effects on target genes (18). Anumber of genes involved in cell growth, cell shape, cellmovement, cell fate, and cell communication are regulatedbyAP-2 family (19–22). Reduced, or overexpressed TFAP2Aexpression is often detected in several types of cancers, suchas melanoma, prostate, breast, ovary, neuroglioma, gastric,colon, and bladder cancers (23–32), indicating that loss ofTFAP2A function, or TFAP2A overexpression may contrib-ute to tumorigenesis and development of tumor malignan-cy. However, the biological role, prognostic value, andclinical significance of TFAP2, as well as its mechanism ofaction in nasopharyngeal carcinoma growth and progres-sion remain unclear.Hypoxia-inducible factor-1a (HIF-1a) is a transcription
factor that activates the expression of a number of genesinvolved in diverse aspects of cellular and physiologicprocesses (33, 34). Under hypoxic conditions, HIF-1aforms a heterodimer with HIF-1b, and binds to the hypox-ia-responsive elements of the promoters to activatedownstream hypoxia-responsive genes, including VEGF, toincrease angiogenesis and tumor metastasis or to promotecancer cell proliferation andmigration (35–37). VEGF has acrucial role in pathologic angiogenesis associated withtumors, intraocular neovascular disorders, and other con-ditions (36, 37). The potent angiogenic inhibitor pigmentepithelium–derived factor (PEDF) counterbalances theeffect of VEGF (38). The activity of HIF-1a is upregulatedby a variety of nonhypoxic signals, including the activationby several oncogenic pathways such as Src, HER-2, Ha-Ras,and mitogen-activated protein kinase signaling pathways(39). HIF-1a is overexpressed in many human cancers, andseveral lines of evidence have indicated its essential role intumorigenesis (40, 41). However, the detailed mechanismof action of HIF-1a and its relationship with TFAP2A innasopharyngeal carcinoma tumorigenesis is still largelyunknown.In this study, we investigated the expression of TFAP2A
in nasopharyngeal carcinoma cell lines and tumor tis-sues. The relationship between TFAP2A and HIF-1aexpression and the clinicopathologic factors, as well asthe functions and mechanisms by which TFAP2A regu-lated nasopharyngeal carcinoma growth and survival,were also evaluated. The regulation of TFAP2A in naso-pharyngeal carcinoma tumor growth was further con-firmed in a nasopharyngeal carcinoma xenograft mousemodel in vivo. Our findings provide new insights into
understanding the regulation of TFAP2A in nasopharyn-geal carcinoma tumorigenesis and exploring the poten-tial therapeutic targets for nasopharyngeal carcinomatherapy.
Materials and MethodsCell lines and cell culture
The human nasopharyngeal carcinoma cell lines (CNE1,CNE2, HONE-1, and SUNE-1) and the immortalized nor-mal nasopharyngeal cell line (NP69) were purchased fromthe American Type Culture Collection. Two CNE2 sub-clones (S18 and S26) and 2 SUNE-1 subclones (5-8F and6-10B) were generously provided by Dr. M. Zeng (Sun Yat-sen University Cancer Center, Guangzhou, China). These 4nasopharyngeal carcinoma cell lines and 4 subclones ofnasopharyngeal carcinoma cells were cultured in Dulbec-co’s Modified Eagle Medium supplemented with 10% FBS,100 U/mL penicillin, and 100 mg/mL streptomycin. TheNP69 cells were cultured in Keratinocyte-SFM (Invitrogen).Each cell line was authenticated in March 2013 and main-tained at 37�C in a humidified atmosphere and 5% CO2
according the recommendations of the providers.
Induction of hypoxiaFor hypoxic induction, the nasopharyngeal carcinoma
cells were cultured in a modulator incubator chamber(autoflow gas mixing instruments incubator; Chang JinInc.) with 94% N2, 1% O2, and 5% CO2 at 37�C at 37�Cfor 24 hours.
Patients and tissue specimensThis study was conducted on a total of 179 paraffin-
embedded nasopharyngeal carcinoma samples, whichwere histologically and clinically diagnosed from theCancer Center (Sun Yat-sen University) between 1999and 2008. For the use of these clinical materials forresearch purposes, prior patient’s consent and approvalfrom the Institute Research Ethics Committee wereobtained. The disease stages of all the patients wereclassified or reclassified according to the 1992 nasopha-ryngeal carcinoma staging system of China. Clinical infor-mation of the samples was described in detail in Table 1.Patients included 139 males and 40 females, of agesranging from 20 to 78 years (mean, 46.1 years). Thefigures on metastasis pertain to its presence at any timein follow-up. The median follow-up time for overallsurvival was 54.6 months for patients still alive at thetime of analysis, and ranged from 12 to 128 months. Atotal of 44 (24.6%) patients died during follow up and 27(15.1%) patients experienced metastasis.
RNA extraction and reverse transcription-PCRTotal RNA from cells was extracted using Trizol reagent
(Life Technologies). The RNA was pretreated with DNaseand used for cDNA synthesis with random hexamers. Thefull-length open reading frame of TFAP2A, VEGF, PEDF,and HIF-1awere PCR amplified from cDNA samples of thenasopharyngeal carcinoma cell lines. The primerswere used
TFAP2A Regulates Nasopharyngeal Carcinoma Growth and Survival
www.aacrjournals.org Cancer Prev Res; 7(2) February 2014 267
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271
for amplification of TFAP2A: sense primer, 5-GCGGCATG-AATCTATTGGAC-3, antisense primer, 5-CCCAGGAAGCC-GTCTTTATT-3; VEGF: sense primer, 5-AGGAGGGCAGAAT-CATCACG-3, antisense primer 5-CAAGGCCCACAGGGAT-TTTCT-3; PEDF: sense primer, 5-TCCAATGCAGAGGAG-TAGCA-3, antisense primer, 5-TGTGCAGGCTTAGAGGG-ACT-3; HIF-1a: sense primer, 5-GCGGCATGAATCTATTG-GAC-3, antisense primer, 5-CCCAGGAAGCCGTCTTTATT-3; GAPDH: sense primer, 5-CAAATTCCATGGCACCGTCA-3, antisense primer, 5-GGAGTGGGTGTCGCTGTTGA-3.The PCR products was confirmed by 1.2% agarose gelelectrophoresis.
Western blot analysisEqual amounts of whole cell lysates were resolved by
SDS-PAGE and electrotransferred on a polyvinylidenedifluoride (PVDF) membrane. The tissues were then incu-bated with primary rabbit monoclonal antibodies againstb-actin, TFAP2A (Cell Signaling). VEGF (Merck Millipore),HIF-1a, and PEDF (Santa Cruz Biotechnology). The proteinbands were detected by enhanced chemiluminescence.
Immunocytochemical stainingTumor cells grown on glass cover slips were washed with
PBS in triplicate and fixed with 4% formaldehyde. Endog-enous peroxidase activity from the cells was blocked with3%hydrogen peroxide. The cover slips were then incubatedwith 1% bovine serum albumin (BSA) in PBS for 1 hour atroom temperature and with primary antibody overnight at4�C, followed by incubation with a secondary antibody for30 minutes. The cover slips were then counter stained withhematoxylin solution.
Immunohistochemical stainingThe 4-mm sections were cut from formalin-fixed paraffin-
embedded tissue blocks and then deparaffinized in xyleneand rehydrated in successive washes of ethanol. The sec-tions were then heated in a microwave oven at mediumpower for 2 minutes in citrate buffer (pH 6.0) for heat-induced epitope retrieval. The sections were subjected toblockade of endogenous peroxidase activity and nonspe-cific binding of the primary antibody, and then targetprotein localization with the first antibody and visualiza-tion with the secondary antibody and color reaction asdescribed above. Subsequently, the slides were incubatedwith rabbit monoclonal antibody anti-TFAP2A (Cell Sig-naling Technology; 1:100 dilution), anti-VEGF (Cell Sig-naling Technology; 1:50 dilution), anti-PEDF (Cell Signal-ing Technology; 1:50 dilution), anti-HIF-1a (Cell SignalingTechnology; 1:25 dilution), and anti-CD34 (Life Technol-ogy; 1:100) for overnight at 4�C in a moist chamber. Theslides were sequentially incubated with a secondary anti-body for 1 hour at room temperature, and stained with 3,3-diaminobenzidine. Finally, the sections were counter-stained with Mayer’s hematoxylin, dehydrated, andmounted. A negative control was obtained by replacing theprimary antibody with PBS. Known immunostaining pos-itive slides were used as positive controls.
Stained tumor cells and paraffin sections were reviewedand scored using a light microscope by a pathologistblinded to the treatment group. Positivity of the stainedtumor cells on cover slips and paraffin sectionswere definedby staining intensity and % of tumor cells: the stainingintensity of protein expression was classified semiquanti-tatively into negative and weak, moderate, and strongly
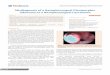
Table 1. Clinicopathologic characteristics and TFAP2A expression of patients with nasopharyngealcarcinoma
Characteristics N % Characteristics N %
Gender N classificationMale 139 77.7 0 39 21.8Female 40 22.3 1 80 44.7
Age 2 43 24�45 88 49.2 3 17 9.6>45 91 50.8
Clinical stage TFAP2A expressionI 7 3.9 Negative 10 5.6II 52 29.1 Positive 169 94.4III 69 38.5 Low 82 45.8IV 51 28.5 High 97 54.2
T classification HIF-1a expression1 23 12.9 Negative 33 18.42 55 30.7 Positive 146 81.63 65 36.3 Low 87 48.64 36 20.1 High 92 51.4
Distant metastasis RecurrenceYes 27 15.1 Yes 11 6.1No 152 84.9 No 168 93.9
Shi et al.
Cancer Prev Res; 7(2) February 2014 Cancer Prevention Research268
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271

positive (0, þ, þþ, and þþþ, respectively). In our study,gene expression was considered positive if the stainingintensity was moderate or strong and the percentage ofpositive stained cells were >25%. However, the density ofmicrovessels staining positive for CD34 was defined aspositive at 400 power of a microscope field.
Cell viability assayCell viability was determined by a MTT assay (Roche
Diagnosis). Briefly, cells plated in 96-well plates (2,000cells/well) were treated with siRNA of TFAP2A and/orHIF-1a at the indicated doses. At 48 hours after treatment,cell viability was determined.
Determination of VEGF and PEDF production by ELISAThe nasopharyngeal carcinoma cells were seeded in 96-
well plates and treated with siRNAs of TFTFAP2A and HIF-1a at 100nmol/L for 48 hours. VEGF andPEDF levels in cellculture media were quantified using a VEGF ImmunoassayKit (968962; R&D Systems) and a Chemikine PEDF ELISAKit (CYT420; Chemikine) according to the manufacturer’sprotocols.
Immunofluorescence and confocal microscopyCells were fixed with 4% paraformaldehyde (w/v) for 20
minutes, quenched for 20 minutes with 50 mmol/L NH4Clin PBS and permeabilized with 0.2% (w/v) saponin in PBSfor 20 minutes. The saturation step was performed for 20minutes in PBS containing 1% BSA and 0.2% saponin(w/v).Cellswere then incubated overnightwith the primaryTFAP2A and HIF-1a antibodies diluted in PBS containing1% BSA and 0.2% saponin. After PBS washings, cells wereincubated for 2 hours with secondary fluorescein isothio-cyanate or tetra-methyl rhodamine isothiocyanate–conju-gated antibodies. TFAP2A and HIF-1a protein localizationwas assessed using a Leica confocal microscopy (ModelTCS-NT). Files of microphotographs were processed withthe Adobe Photoshop 5.0 software.
Co-immunoprecipitation assaysThe nuclei protein extracts were prepared and incubated
with the antibodies for TFAP2A and HIF-1a for 24 hours at4�C. Then, the sepharose-conjugated protein-A/G beads(Santa Cruz Biotech) were added and the mixture wasincubated at 4�C for another 12 hours. After extensivewashing with radioimmunoprecipitation assay buffer, thebeads were boiled. The precipitated proteins were separatedby SDS-PAGE and transferred to PVDF membranes forWestern blot analysis.
Preparation of DOTAP-cholesterol nanoparticles andencapsulation of siRNAsDOTAP-cholesterol was purchased from Avanti Polar-
lipids Inc. High-performance liquid chromatography(HPLC)-grade chloroformwas obtained fromSigmaChem-ical Co. The DOTAP-cholesterol nanoparticles were pre-pared by an EmulsiFlex-B3 high-pressure homogenizer(HPH; Avestin Inc.). In brief, the nanoparticles were pre-
pared from amixture of 2 lipids: cholesterol andDOTAP, atthe molar ratio of 1:1. The lipids were dissolved in 15 mLHPLC-grade chloroform in a round bottom flask and thendried under nitrogen gas and overnight vacuum. The result-ing films of the lipids were hydrated in deionized water togive a final concentration of 10 mmol/L. The lipid disper-sions were warmed and mixed at 50�C for 45 minutes byrotation, followed bywarming again at 35�C for another 10minutes. The resultant dispersion was stored at room tem-perature for 3 hours before it was transferred into a scintil-lation vial and warmed again at 50�C for 10 minutes. Thefinal lipid dispersion was homogenized using HPH at20,000 psi for 5 cycles. Each time, 2.5mL of lipid dispersionwas subjected to homogenization and the resultantDOTAP-cholesterol nanoparticles were collected in anotherscintillation vial. The DOTAP-cholesterol nanoparticleswere kept at room temperature for 1 hour before overnightstorage at 4�C. TFAP2A or HIF-1a siRNAs duplexes werepurchased from Santa Cruz Biotechnology and encapsulat-ed using the DOTAP-cholesterol nanoparticles.
Tumor growth inhibition by TFAP2A siRNA in axenograft mouse model
To determine the effect of TFAP2A siRNA on nasopha-ryngeal carcinoma cell growth in a xenograft model, CNE2cells (2 � 106) were inoculated subcutaneously into theflank of the nude mice. Once palpable tumors wereobserved, tumor volume measurements were taken every3 days using calipers. The tumor volume was calculatedusing the following formula:V¼ (width2� length)/2. Bodyweights were also recorded. Two weeks after injection, themice were randomized into 2 groups (7 mice/group).Group 1 received injection with In Vivo Ready nonspecificsiRNA, and group 2 with In Vivo Ready TFAP2A siRNA.DOTAP-cholesterol nanoparticles encapsulated siRNAduplexes were injected into the tumors using insulin syrin-ges at a concentration of 10 mg of siRNA/50 mm3 of tumorvolume. All 2 groups were treated twice a week for 4 weeks.Upon termination, tumors were harvested and weighted.Animal experiments were approved by the Animal ResearchCommittee of Sun Yat-sen University Cancer Center andwere performed in accordance with established guidelines.
Evaluation of TFAP2A knockdown and angiogenesisfactors in xenograft tumor tissues
Tumor tissues from the above treated animals were col-lected andplaced in 10% formalin and embedded inparaffinfor below analysis. The sections were stained with hematox-ylin and eosin according to standard immunohistochemicalprocedures. Knockdown of TFAP2A by siRNA was deter-mined by immunohistochemical staining with anti-TFAP2Aantibody. To assess the effect of TFAP2A siRNA in vivo onangiogenesis factor VEGF, PEDF, and CD34, the embeddedtissues were stained using anti-PEDF, VEGF, or CD34 anti-body to determine the expression of angiogenic factors. Anegative control was obtained by replacing the primaryantibody with a normal rabbit or mouse immunoglobulinG. The immunoreactivity positive cells from each of the
TFAP2A Regulates Nasopharyngeal Carcinoma Growth and Survival
www.aacrjournals.org Cancer Prev Res; 7(2) February 2014 269
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271

differently treated tumor tissue sections were measured at�200magnificationusing a lightmicroscope. The amountofproteins was analyzed by integral optical density using IPP(ImagePlusPro6.0,MediaCybernetics, Inc., Rockville,MD).
Statistical analysisStatistical analysis was performed using the SPSS statis-
tical software package (standard version 16.0; SPSS). StrongTFAP2A immunoreactivity was assessed for the associationwith clinicopathologic variables such as gender, age, andpathologic TNM stage by using the Pearson x2 test. Survivalcurveswere calculated from the date of surgery to the timeofdeath related to nasopharyngeal carcinoma or to the lastfollow-up observation. Kaplan–Meier curves were calculat-ed for each relevant variable and for TFAP2A expression;differences in survival times among patient subgroups wereanalyzed by the log-rank test. Univariate and multivariateanalyses were done with the Cox proportional hazardregression model to determine the associations betweenclinicopathologic variables and cancer-related mortality.First, we analyzed the associations between death andpossible prognostic factors, including TFAP2A expression,gender, clinical stage,metastasis, T stage, andN stage, taking
into consideration one factor at a time. Second,multivariateanalysis was applied on forward (stepwise) procedures.
ResultsTFAP2A is highly expressed in nasopharyngealcarcinoma cell lines and tumor tissues
Wefirst examined the expressionof TFAP2AatmRNAandprotein levels in 8 human nasopharyngeal carcinoma celllines and an immortalized normal human nasopharyngealepithelial cell line (NP69)by reverse transcription-PCR (RT-PCR) and Western blot analysis, respectively. The TFAP2AmRNA levels were upregulated in 7 nasopharyngeal carci-noma tumor cell lines (CNE1,CNE2, S18,HONE1, SUNE1,5-8F, 6-10B) by comparison with the expression of TFAP2AmRNA in normal cell line NP69 (Fig. 1A). Similarly,TFAP2A proteins were also highly expressed in 6 nasopha-ryngeal carcinoma cell lines (CNE1, CNE2, S18, S26,SUNE1, 5-8F). By contrast, TFAP2A protein was notdetected in NP69 cells (Fig. 1B).
We also tested the expression of TFAP2A protein and itscellular localization in nasopharyngeal carcinoma cells byimmunocytochemical staining. Consistent with the resultsfrom RT-PCR and Western blot analysis, the high levels of
Di ii
viii
A
TFAP2A
GAPDH
B
SU
NE
1
S26
HO
NE
1
TFAP2A
β-Actin
NP
69
CN
E1
CN
E2
S18
5-8
F
6-1
0B
C
HONE1
5-8F 6-10B
NP69 Positive CNE2
CNE1
SUNE1
CNE2
S18 S26
NP
69
CN
E1
CN
E2
S18
S26
HO
NE
1
SU
NE
1
5-8
F
6-1
0B
Figure 1. TFAP2A is highlyexpressed in nasopharyngealcarcinoma cells and tumor tissues.The expression of TFAP2A inhuman normal NP69 cell andvarious nasopharyngealcarcinoma cell lines wasanalyzed by RT-PCR (A),Western blot analysis (B), andimmunocytochemical analysis (C).The expression of TFAP2A innasopharyngeal carcinoma tumortissues and the matched adjacentnasopharyngeal tissues wasdetected by immunohistochemicalstaining (D). i, low expression ofTFAP2A; ii, high expression ofTFAP2A; iii, negative expression ofTFAP2A; iv, low expression ofTFAP2A in the matched adjacentnasopharyngeal tissues.Magnification, �200.
Shi et al.
Cancer Prev Res; 7(2) February 2014 Cancer Prevention Research270
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271

TFAP2A proteins were detected in 6 nasopharyngeal carci-noma cell lines (CNE1, CNE2, SUNE1, S18, S26, 5-8F). TheHONE1 and 6-10B cells had weak staining of TFAP2A.Moreover, the results showed that most of the staining ofTFAP2A proteins localized in the cell nuclei. By contrast, thenormal nasopharyngeal cell line (NP69) had a relativelylow expression of endogenous TFAP2A (Fig. 1C).To further confirm the high expression of TFAP2 in
nasopharyngeal carcinoma cells and to investigate the clin-icopathologic significance of TFAP2A expression, we alsoanalyzed the expression of TFAP2A in tumor tissues frompatients with nasopharyngeal carcinoma by immunohisto-chemical staining assay. TFAP2A-positive staining wasobserved in the nucleus of nasopharyngeal carcinoma cellsin tumor tissues, but stainingwas negative in any of the cellsin adjacent nontumor tissue samples surrounding tumors(Fig. 1D). These results confirm the in vitro findings aboveand suggest that high levels TFAP2A may be a potentialbiomarker for nasopharyngeal carcinomas.
TFAP2Aexpression is associatedwith clinicopathologiccharacteristicsTo gain further insight into the prognostic value of
TFAP2A expression in patients with nasopharyngeal carci-noma, paraffin-embedded tissue sections (n ¼ 179) withhistopathologically confirmed nasopharyngeal carcinomawere examined using immunohistochemistry. The levels ofTFAP2A immunoreactivity varied between the tumor tissuesamples and the adjacent nontumor tissue samples. Highpositive TFAP2A expressionwas localized to the nuclei in 97(54.2%) of the resected tumor tissue samples, whereas theremaining 82 cases (45.8%) displayed low levels nucleiTFAP2A expression (Table 1). Immunohistochemical deter-mination of TFAP2A levels was also statistically analyzed toidentify its association with the clinicopathologic featuresof nasopharyngeal carcinoma. TFAP2A expression was sig-nificantly correlatedwith gender (P<0.001), T classification(P¼ 0.002), N stage (P¼ 0.001), clinical stage (P < 0.001),and distant metastasis (P < 0.001). However, there was nosignificant correlation between TFAP2A expression andage and recurrence (P ¼ 0.925 and P ¼ 0.516) (Supple-mentary Table S1). Spearman correlation analysis alsorevealed that TFAP2A expression levels were positivelycorrelated with clinical stage (r ¼ 0.286; P < 0.001), Tclassification (r ¼ 0.277; P < 0.001), N classification (r ¼0.248; P < 0.001), HIF-1a (r ¼ 0.512; P < 0.001), andmetastasis (r ¼ 0.294; P < 0.001; Supplementary TableS2). Our data indicate that high TFAP2A expressionsignificantly correlated with advanced tumor stage, localinvasion, and clinical progression.
TFAP2A overexpression is associated with poorprognosis for patientswith nasopharyngeal carcinomaTo investigate the biologic and clinicopathologic signif-
icance of TFAP2A in nasopharyngeal carcinoma carcino-genesis, we carried out immunohistochemical staininganalysis in tumor tissues from the patients with nasopha-ryngeal carcinoma. The median survival time of the 179
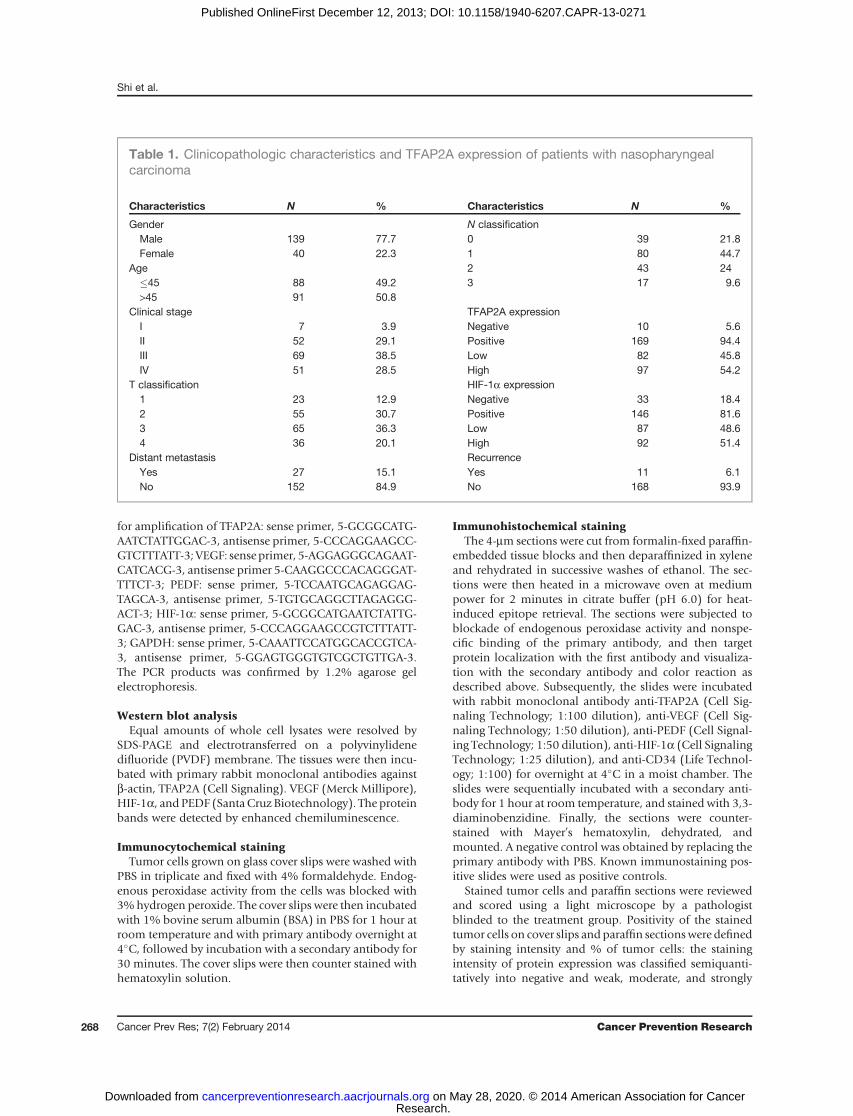
patients with nasopharyngeal carcinoma was 54.6 months(range 12–128months). The overall survival rate in the lowTFAP2A expression group was significantly improved com-pared with the group with high TFAP2A expression (P <0.001, Fig. 2A). In addition, the high HIF-1a expressiongroup had significantly low rate of overall survival than thegroup with low HIF-1a expression (P ¼ 0.04, Fig. 2B).Moreover, the group with low expression of both TFAP2Aand HIF-1a had high overall survival rate than the groupwith high expression of both TFAP2A and HIF-1a (P ¼0.008; Fig. 2C).
We further evaluated the potential correlation betweenexpression of TFAP2A and HIF-1a in our nasopharyngealcarcinoma cohort. The results from immunohistochemicalanalysis showed a positive correlation between expressionof TFAP2A and HIF-1a in nasopharyngeal carcinoma (Fig.2D and E and Table S2). For the 97 nasopharyngeal carci-noma cases with high expression of TFAP2A, an average of69.7% of the carcinoma cells stained positive with HIF-1aprotein; the percentage was significantly higher than that(30.3%) in the remaining 82 nasopharyngeal carcinomaswith low expression of TFAP2A (P < 0.001; Fig. 2D).
TFAP2A knockdown inhibits nasopharyngealcarcinoma cell growth by targeting HIF-1a–mediatedVEGF/PEDF signaling
To determine the role TFAP2A in regulating the growthof nasopharyngeal carcinoma cells, we examined theeffect of TFAP2A knockdown by DOTAP-cholesterol–based TFAP2A siRNA nanoparticles (si-TFAP2A) on cellviability in various kinds of human nasopharyngeal car-cinoma cell lines, including high differentiated cell lineCNE1, low differentiated cell line CNE2, and its clonesS18 and S26, by a MTT analysis. The results showed thatknockdown of TFAP2A expression by TFAP2A siRNA (si-TFAP2A, 100 nmol/L) for 72 hours significantly inhibitedcell viability as compared with the transfection with thenonspecific scramble siRNA (si-NS; 100 nmol/L) andmock control groups, resulting in a 17% to 55% inhibi-tions in cell viability in the 4 kinds of nasopharyngealcarcinoma cell lines (Fig. 3A).
Because TFAP2A expression positively correlates withHIF-1a levels in nasopharyngeal carcinomas and the upre-gulatedHIF-1a canpromoted the expressionofVEGFgenes,we next determined the effect of TFAP2A knockdownon theexpression of VEGF and PEDF at mRNA (Fig. 3B) andprotein levels (Fig. 3C) in nasopharyngeal carcinoma cellsby RT-PCR and Western blot analysis, and on the release ofVEGF and PEDF proteins in cell culture media by ELISA inCNE2 cells (Fig. 3D). Transfectionwith TFAP2A siRNA (100nmol/L) dramatically inhibited the expression of VEGFproteins, but did not significantly decrease the levels ofPEDF protein expression (Fig. 3C). Similarly, treatment ofnasopharyngeal carcinoma cells with TFAP2A siRNA (100nmol/L) also significantly inhibited the release of VEGFprotein, but did not affect the levels of PEDF in cell culturemedia, resulting in a significant reduction of the ratio ofVEGF/PEDF (Fig. 3D). Knockdown of TFAP2A expression
TFAP2A Regulates Nasopharyngeal Carcinoma Growth and Survival
www.aacrjournals.org Cancer Prev Res; 7(2) February 2014 271
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271
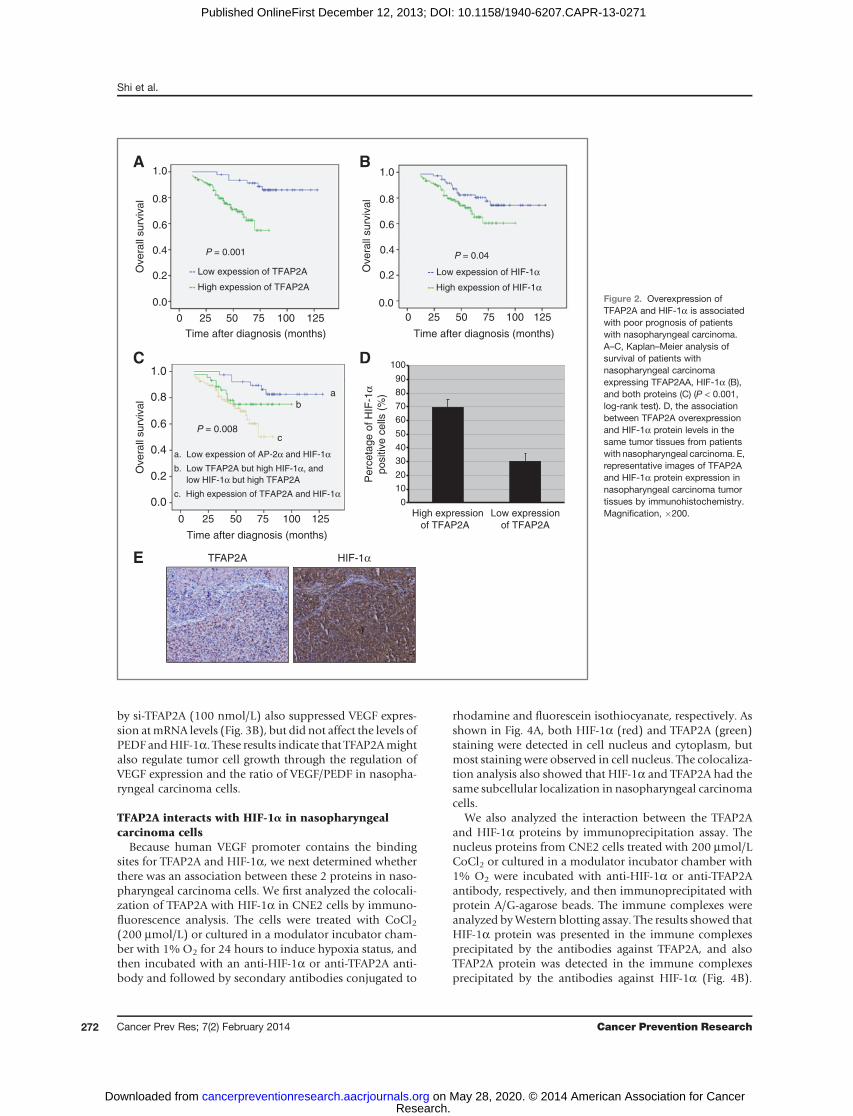
by si-TFAP2A (100 nmol/L) also suppressed VEGF expres-sion atmRNA levels (Fig. 3B), but did not affect the levels ofPEDF andHIF-1a. These results indicate that TFAP2Amightalso regulate tumor cell growth through the regulation ofVEGF expression and the ratio of VEGF/PEDF in nasopha-ryngeal carcinoma cells.
TFAP2A interacts with HIF-1a in nasopharyngealcarcinoma cells
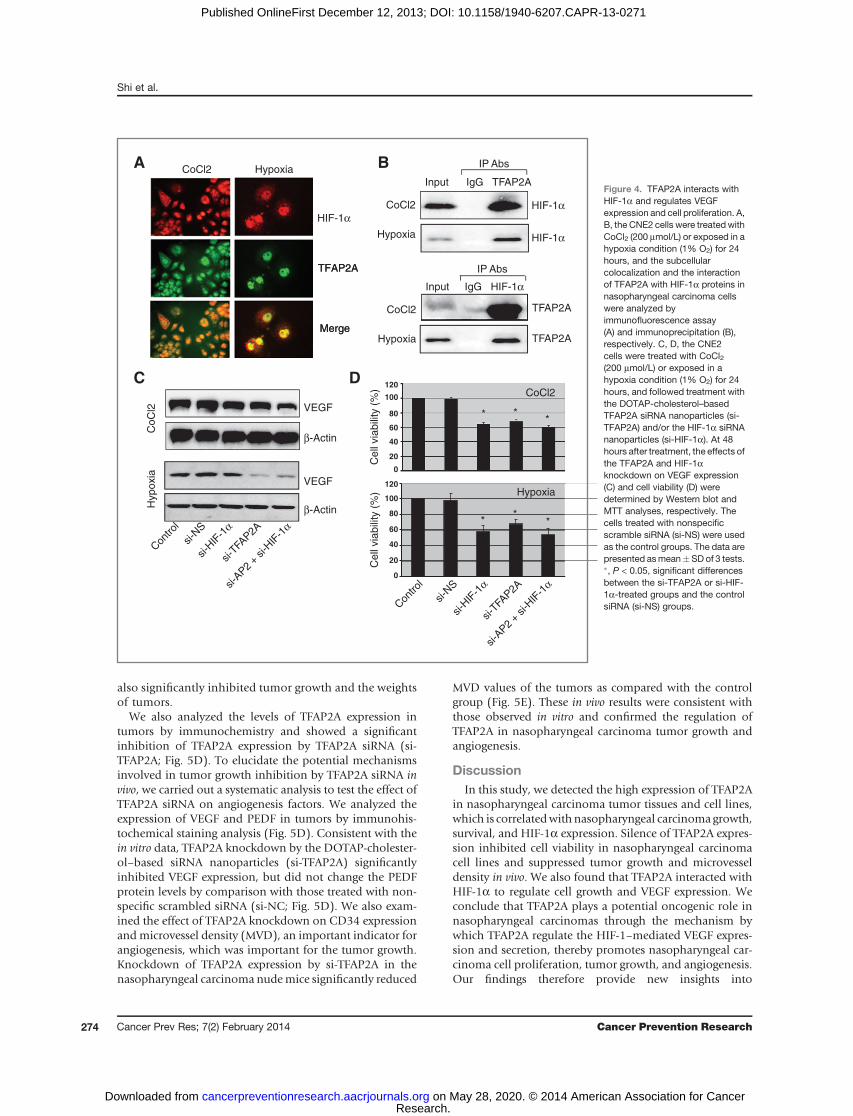
Because human VEGF promoter contains the bindingsites for TFAP2A and HIF-1a, we next determined whetherthere was an association between these 2 proteins in naso-pharyngeal carcinoma cells. We first analyzed the colocali-zation of TFAP2A with HIF-1a in CNE2 cells by immuno-fluorescence analysis. The cells were treated with CoCl2(200 mmol/L) or cultured in a modulator incubator cham-ber with 1% O2 for 24 hours to induce hypoxia status, andthen incubated with an anti-HIF-1a or anti-TFAP2A anti-body and followed by secondary antibodies conjugated to
rhodamine and fluorescein isothiocyanate, respectively. Asshown in Fig. 4A, both HIF-1a (red) and TFAP2A (green)staining were detected in cell nucleus and cytoplasm, butmost staining were observed in cell nucleus. The colocaliza-tion analysis also showed that HIF-1a and TFAP2A had thesame subcellular localization in nasopharyngeal carcinomacells.
We also analyzed the interaction between the TFAP2Aand HIF-1a proteins by immunoprecipitation assay. Thenucleus proteins from CNE2 cells treated with 200 mmol/LCoCl2 or cultured in a modulator incubator chamber with1% O2 were incubated with anti-HIF-1a or anti-TFAP2Aantibody, respectively, and then immunoprecipitated withprotein A/G-agarose beads. The immune complexes wereanalyzed byWestern blotting assay. The results showed thatHIF-1a protein was presented in the immune complexesprecipitated by the antibodies against TFAP2A, and alsoTFAP2A protein was detected in the immune complexesprecipitated by the antibodies against HIF-1a (Fig. 4B).
C D
High expression
of TFAP2A
Perc
eta
ge o
f H
IF-1
αpositiv
e c
ells
(%
)
0
10
20
30
40
50
60
70
80
90
100
Low expression
of TFAP2A
A B
Time after diagnosis (months)
Overa
ll surv
ival
0 25 50 75 100 125
1.0
0.8
0.6
0.4
0.2
0.0
a. Low expession of AP-2α and HIF-1α
c. High expession of TFAP2A and HIF-1α
P = 0.008
Time after diagnosis (months)
Ove
rall
su
rviv
al
0 25 50 75 100 125
1.0
0.8
0.6
0.4
0.2
0.0
-- Low expession of HIF-1α
-- High expession of HIF-1α
Time after diagnosis (months)
Overa
ll surv
ival
0 25 50 75 100 125
1.0
0.8
0.6
0.4
0.2
0.0
-- Low expession of TFAP2A
-- High expession of TFAP2A
TFAP2A HIF-1αE
b
c
b. Low TFAP2A but high HIF-1α, and
low HIF-1α but high TFAP2A
P = 0.001
a
P = 0.04
Figure 2. Overexpression ofTFAP2A and HIF-1a is associatedwith poor prognosis of patientswith nasopharyngeal carcinoma.A–C, Kaplan–Meier analysis ofsurvival of patients withnasopharyngeal carcinomaexpressing TFAP2AA, HIF-1a (B),and both proteins (C) (P < 0.001,log-rank test). D, the associationbetween TFAP2A overexpressionand HIF-1a protein levels in thesame tumor tissues from patientswith nasopharyngeal carcinoma. E,representative images of TFAP2Aand HIF-1a protein expression innasopharyngeal carcinoma tumortissues by immunohistochemistry.Magnification, �200.
Shi et al.
Cancer Prev Res; 7(2) February 2014 Cancer Prevention Research272
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271
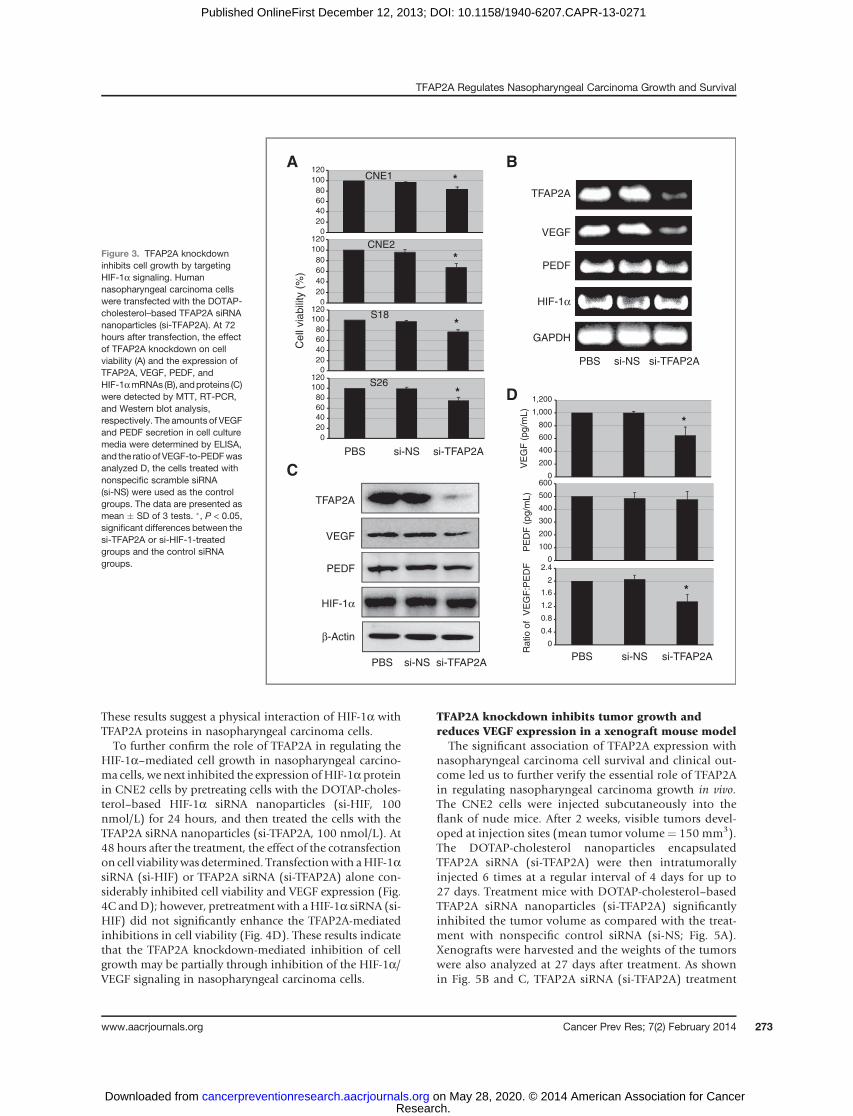
These results suggest a physical interaction of HIF-1a withTFAP2A proteins in nasopharyngeal carcinoma cells.To further confirm the role of TFAP2A in regulating the
HIF-1a–mediated cell growth in nasopharyngeal carcino-ma cells, we next inhibited the expression ofHIF-1a proteinin CNE2 cells by pretreating cells with the DOTAP-choles-terol–based HIF-1a siRNA nanoparticles (si-HIF, 100nmol/L) for 24 hours, and then treated the cells with theTFAP2A siRNA nanoparticles (si-TFAP2A, 100 nmol/L). At48 hours after the treatment, the effect of the cotransfectionon cell viabilitywas determined. Transfectionwith aHIF-1asiRNA (si-HIF) or TFAP2A siRNA (si-TFAP2A) alone con-siderably inhibited cell viability and VEGF expression (Fig.4C andD); however, pretreatment with a HIF-1a siRNA (si-HIF) did not significantly enhance the TFAP2A-mediatedinhibitions in cell viability (Fig. 4D). These results indicatethat the TFAP2A knockdown-mediated inhibition of cellgrowth may be partially through inhibition of the HIF-1a/VEGF signaling in nasopharyngeal carcinoma cells.
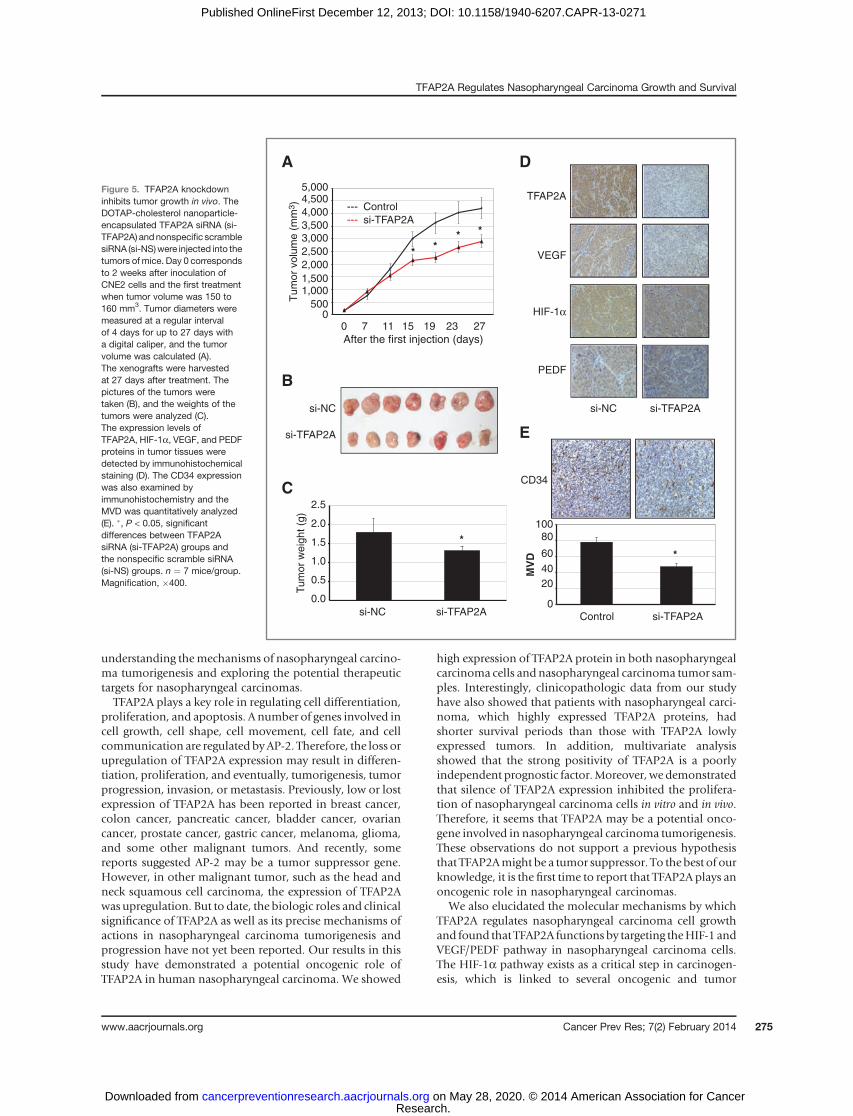
TFAP2A knockdown inhibits tumor growth andreduces VEGF expression in a xenograft mouse model
The significant association of TFAP2A expression withnasopharyngeal carcinoma cell survival and clinical out-come led us to further verify the essential role of TFAP2Ain regulating nasopharyngeal carcinoma growth in vivo.The CNE2 cells were injected subcutaneously into theflank of nude mice. After 2 weeks, visible tumors devel-oped at injection sites (mean tumor volume¼ 150 mm3).The DOTAP-cholesterol nanoparticles encapsulatedTFAP2A siRNA (si-TFAP2A) were then intratumorallyinjected 6 times at a regular interval of 4 days for up to27 days. Treatment mice with DOTAP-cholesterol–basedTFAP2A siRNA nanoparticles (si-TFAP2A) significantlyinhibited the tumor volume as compared with the treat-ment with nonspecific control siRNA (si-NS; Fig. 5A).Xenografts were harvested and the weights of the tumorswere also analyzed at 27 days after treatment. As shownin Fig. 5B and C, TFAP2A siRNA (si-TFAP2A) treatment
A
0
20
40
60
80
100
120
0
20
40
60
80
100
120
0
20
40
60
80
100
120
0
20
40
60
80
100
120
CNE1
CNE2
S18
S26
Ce
ll via
bili
ty (
%)
*
*
*
*
si-TFAP2Asi-NSPBS
B
VEGF
PEDF
HIF-1α
GAPDH
TFAP2A
si-TFAP2Asi-NSPBS
0
200
400
600
800
1,000
1,200
0
100
200
300
400
500
600
0
0.4
0.8
1.2
1.6
2
2.4
VE
GF
(p
g/m
L)
PE
DF
(pg/m
L)
Ratio o
f V
EG
F:P
ED
F
si-TFAP2Asi-NSPBS
*
*
TFAP2A
VEGF
PEDF
HIF-1α
β-Actin
si-TFAP2Asi-NSPBS
C
D
Figure 3. TFAP2A knockdowninhibits cell growth by targetingHIF-1a signaling. Humannasopharyngeal carcinoma cellswere transfected with the DOTAP-cholesterol–based TFAP2A siRNAnanoparticles (si-TFAP2A). At 72hours after transfection, the effectof TFAP2A knockdown on cellviability (A) and the expression ofTFAP2A, VEGF, PEDF, andHIF-1amRNAs (B), andproteins (C)were detected by MTT, RT-PCR,and Western blot analysis,respectively. The amounts of VEGFand PEDF secretion in cell culturemedia were determined by ELISA,and the ratio of VEGF-to-PEDFwasanalyzed D, the cells treated withnonspecific scramble siRNA(si-NS) were used as the controlgroups. The data are presented asmean � SD of 3 tests. �, P < 0.05,significant differences between thesi-TFAP2A or si-HIF-1-treatedgroups and the control siRNAgroups.
TFAP2A Regulates Nasopharyngeal Carcinoma Growth and Survival
www.aacrjournals.org Cancer Prev Res; 7(2) February 2014 273
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271
also significantly inhibited tumor growth and the weightsof tumors.
We also analyzed the levels of TFAP2A expression intumors by immunochemistry and showed a significantinhibition of TFAP2A expression by TFAP2A siRNA (si-TFAP2A; Fig. 5D). To elucidate the potential mechanismsinvolved in tumor growth inhibition by TFAP2A siRNA invivo, we carried out a systematic analysis to test the effect ofTFAP2A siRNA on angiogenesis factors. We analyzed theexpression of VEGF and PEDF in tumors by immunohis-tochemical staining analysis (Fig. 5D). Consistent with thein vitro data, TFAP2A knockdown by the DOTAP-cholester-ol–based siRNA nanoparticles (si-TFAP2A) significantlyinhibited VEGF expression, but did not change the PEDFprotein levels by comparison with those treated with non-specific scrambled siRNA (si-NC; Fig. 5D). We also exam-ined the effect of TFAP2A knockdown on CD34 expressionandmicrovessel density (MVD), an important indicator forangiogenesis, which was important for the tumor growth.Knockdown of TFAP2A expression by si-TFAP2A in thenasopharyngeal carcinoma nudemice significantly reduced
MVD values of the tumors as compared with the controlgroup (Fig. 5E). These in vivo results were consistent withthose observed in vitro and confirmed the regulation ofTFAP2A in nasopharyngeal carcinoma tumor growth andangiogenesis.
DiscussionIn this study, we detected the high expression of TFAP2A
in nasopharyngeal carcinoma tumor tissues and cell lines,which is correlatedwithnasopharyngeal carcinomagrowth,survival, and HIF-1a expression. Silence of TFAP2A expres-sion inhibited cell viability in nasopharyngeal carcinomacell lines and suppressed tumor growth and microvesseldensity in vivo. We also found that TFAP2A interacted withHIF-1a to regulate cell growth and VEGF expression. Weconclude that TFAP2A plays a potential oncogenic role innasopharyngeal carcinomas through the mechanism bywhich TFAP2A regulate the HIF-1–mediated VEGF expres-sion and secretion, thereby promotes nasopharyngeal car-cinoma cell proliferation, tumor growth, and angiogenesis.Our findings therefore provide new insights into
DC
A B
HIF-1α
Input IgG TFAP2A
IP Abs
HIF-1α
CoCl2
TFAP2A
TFAP2A
Input IgG HIF-1α
IP AbsTFAP2A
Merge
Hypoxia
β-Actin
VEGF
β-Actin
VEGF
Hypoxia
CoC
l2
Cell
via
bili
ty (
%)
** *
100
80
60
40
20
0
120
***
100
80
60
40
20
0
120
Cell
via
bili
ty (
%)
CoCl2
HIF-1α
Merge
CoCl2
TFAP2A
Hypoxia
CoCl2
Hypoxia
Hypoxia
Con
trol
si-N
S
si-T
FAP2A
si-H
IF-1
α
si-A
P2 +
si-H
IF-1
α
Con
trol
si-N
S
si-T
FAP2A
si-H
IF-1
α
si-A
P2 +
si-H
IF-1
α
Figure 4. TFAP2A interacts withHIF-1a and regulates VEGFexpression and cell proliferation. A,B, the CNE2 cells were treated withCoCl2 (200 mmol/L) or exposed in ahypoxia condition (1% O2) for 24hours, and the subcellularcolocalization and the interactionof TFAP2A with HIF-1a proteins innasopharyngeal carcinoma cellswere analyzed byimmunofluorescence assay(A) and immunoprecipitation (B),respectively. C, D, the CNE2cells were treated with CoCl2(200 mmol/L) or exposed in ahypoxia condition (1% O2) for 24hours, and followed treatment withthe DOTAP-cholesterol–basedTFAP2A siRNA nanoparticles (si-TFAP2A) and/or the HIF-1a siRNAnanoparticles (si-HIF-1a). At 48hours after treatment, the effects ofthe TFAP2A and HIF-1aknockdown on VEGF expression(C) and cell viability (D) weredetermined by Western blot andMTT analyses, respectively. Thecells treated with nonspecificscramble siRNA (si-NS) were usedas the control groups. The data arepresented asmean�SDof 3 tests.�, P < 0.05, significant differencesbetween the si-TFAP2A or si-HIF-1a-treated groups and the controlsiRNA (si-NS) groups.
Shi et al.
Cancer Prev Res; 7(2) February 2014 Cancer Prevention Research274
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271
understanding themechanisms of nasopharyngeal carcino-ma tumorigenesis and exploring the potential therapeutictargets for nasopharyngeal carcinomas.TFAP2A plays a key role in regulating cell differentiation,
proliferation, and apoptosis. A number of genes involved incell growth, cell shape, cell movement, cell fate, and cellcommunication are regulated byAP-2. Therefore, the loss orupregulation of TFAP2A expression may result in differen-tiation, proliferation, and eventually, tumorigenesis, tumorprogression, invasion, or metastasis. Previously, low or lostexpression of TFAP2A has been reported in breast cancer,colon cancer, pancreatic cancer, bladder cancer, ovariancancer, prostate cancer, gastric cancer, melanoma, glioma,and some other malignant tumors. And recently, somereports suggested AP-2 may be a tumor suppressor gene.However, in other malignant tumor, such as the head andneck squamous cell carcinoma, the expression of TFAP2Awas upregulation. But to date, the biologic roles and clinicalsignificance of TFAP2A as well as its precise mechanisms ofactions in nasopharyngeal carcinoma tumorigenesis andprogression have not yet been reported. Our results in thisstudy have demonstrated a potential oncogenic role ofTFAP2A in human nasopharyngeal carcinoma. We showed
high expression of TFAP2A protein in both nasopharyngealcarcinoma cells and nasopharyngeal carcinoma tumor sam-ples. Interestingly, clinicopathologic data from our studyhave also showed that patients with nasopharyngeal carci-noma, which highly expressed TFAP2A proteins, hadshorter survival periods than those with TFAP2A lowlyexpressed tumors. In addition, multivariate analysisshowed that the strong positivity of TFAP2A is a poorlyindependent prognostic factor.Moreover, we demonstratedthat silence of TFAP2A expression inhibited the prolifera-tion of nasopharyngeal carcinoma cells in vitro and in vivo.Therefore, it seems that TFAP2A may be a potential onco-gene involved in nasopharyngeal carcinoma tumorigenesis.These observations do not support a previous hypothesisthat TFAP2Amight be a tumor suppressor. To the best of ourknowledge, it is the first time to report that TFAP2A plays anoncogenic role in nasopharyngeal carcinomas.
We also elucidated the molecular mechanisms by whichTFAP2A regulates nasopharyngeal carcinoma cell growthand found that TFAP2A functions by targeting theHIF-1 andVEGF/PEDF pathway in nasopharyngeal carcinoma cells.The HIF-1a pathway exists as a critical step in carcinogen-esis, which is linked to several oncogenic and tumor
si-NC
si-TFAP2A
B
si-NC si-TFAP2A
Tum
or
we
igh
t (g
)
0.5
1.0
1.5
2.0
2.5
0.0
C
A
Tum
or
volu
me (
mm
3)
0 7 11 15 19 23 27
After the first injection (days)
5000
1,500
2,500
3,500
4,500
1,000
2,000
3,000
4,000
5,000
D
E
CD34
MV
D
PEDF
HIF-1α
VEGF
TFAP2A
si-NC si-TFAP2A
Control---
--- si-TFAP2A
Control si-TFAP2A
**
** *
*
20
0
40
60
80100
Figure 5. TFAP2A knockdowninhibits tumor growth in vivo. TheDOTAP-cholesterol nanoparticle-encapsulated TFAP2A siRNA (si-TFAP2A) andnonspecific scramblesiRNA (si-NS)were injected into thetumors of mice. Day 0 correspondsto 2 weeks after inoculation ofCNE2 cells and the first treatmentwhen tumor volume was 150 to160 mm3. Tumor diameters weremeasured at a regular intervalof 4 days for up to 27 days witha digital caliper, and the tumorvolume was calculated (A).The xenografts were harvestedat 27 days after treatment. Thepictures of the tumors weretaken (B), and the weights of thetumors were analyzed (C).The expression levels ofTFAP2A, HIF-1a, VEGF, and PEDFproteins in tumor tissues weredetected by immunohistochemicalstaining (D). The CD34 expressionwas also examined byimmunohistochemistry and theMVD was quantitatively analyzed(E). �, P < 0.05, significantdifferences between TFAP2AsiRNA (si-TFAP2A) groups andthe nonspecific scramble siRNA(si-NS) groups. n ¼ 7 mice/group.Magnification, �400.
TFAP2A Regulates Nasopharyngeal Carcinoma Growth and Survival
www.aacrjournals.org Cancer Prev Res; 7(2) February 2014 275
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271

suppressor gene pathways in cancers. As a transcriptionfactor, HIF-1a heterodimerizes with the constitutivelyexpressed HIF-1b subunit, and they activate the expressionof a number of genes, including VEGF, to take part in tumorangiogenesis and tumor cell proliferation and invasion. Bybinding to the hypoxia-responsive elements on VEGF pro-moter, HIF-1 leads to the transcriptional activation of theVEGF gene (42). VEGF plays an important role in tumorangiogenesis, tumor cell growth, and survival. PEDF coun-terbalances the effect of VEGF. PEDF and VEGF have aninverse relationship with each other, especially in cancer inwhich low levels of PEDF are associated with the increasedincidence of metastasis and poor malignancy prognosis(43). PEDF suppressed proliferation and induced apoptosisof the tumor cell (44). The increased ratio of VEGF/PEDF isrequired for angiogenesis and tumor growth. In this study,we found that HIF-1a interacted with TFAP2A to regulateVEGF expression in nasopharyngeal carcinoma cells, andknockdown of TFAP2A by siRNA inhibited the ratio of theVEGF/PEDF. Thus, our results show that HIF-1a signalingcontributes, at least in part, to TFAP2A-mediated nasopha-ryngeal carcinoma carcinogenesis.
Moreover, we demonstrated that knockdown of TFAP2AbyDOTAP-cholesterol–based TFAP2A siRNA nanoparticlesmarkedly inhibited tumor growth in a nasopharyngealcarcinoma xenograftmousemodel at least partially throughthemodulation of VEGF signaling. Immunohistochemistryanalysis for the xenograft tumors showed that TFAP2Aknockdown inhibited the angiogenesis-related protein fac-tor VEGF. Thus, these in vivo studies confirmed the tumorinhibition effects by TFAP2A knockdown in vitro and pro-vide a rationale for pharmacologic investigation of TFAP2Aas a novel therapeutic target in nasopharyngeal carcinoma.The detailedmechanisms by which TFAP2A regulates VEGFremain to be elucidated.
In conclusion, our study demonstrate that high TFAP2Aexpression independently predicts worse overall survival inpatients with nasopharyngeal carcinoma and TFAP2A playsa critical role in regulating human nasopharyngeal carci-noma carcinogenesis by targeting HIF-1a and VEGF/PEDF-dependent pathways. These results indicate that TFAP2Aoverexpression in patients with nasopharyngeal carcinomais associated with a poor prognosis and could be a potentialprognostic biomarker and therapeutic target for nasopha-ryngeal carcinoma treatment.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: D. Shi, W. DengDevelopment of methodology: D. Shi, Y. Zhang, L. FuAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): D. Shi, F. Xie, Y. Tian, W. Chen, L. FuAnalysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis): D. Shi, L. Fu, W. DengWriting, review, and/or revision of the manuscript: D. Shi, F. Xie,W. DengAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): D. Shi, F. Xie, J. Wang, W. GuoStudy supervision: D. Shi, T. Kang, W. Huang, W. Deng
Grant SupportThis work was supported by the funds from the National Natural Science
Foundation of China (81272195, 81071687, 81372133,W. Deng), the State"863 Program" of China (SS2012AA020403, W. Deng), the State "973Program" of China (2014CB542005, X. He), the Doctoral Programs Foun-dation ofMinistry of Education of China (20110171110077, W. Deng), andthe State Key Laboratory of Oncology in South China (W. Deng).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received July 24, 2013; revisedNovember 1, 2013; acceptedNovember 21,2013; published OnlineFirst December 12, 2013.
References1. Brennan B. Nasopharyngeal carcinoma. Orphanet J Rare Dis 2006;
26:23.2. Sen S. Aneuploidy and cancer. Curr Opin Oncol 2000;12:82–8.3. Desalvo MN. Radiation necrosis of the pons after radiotherapy for
nasopharyngeal carcinoma: diagnosis and treatment. J Radiol CaseRep 2012;6:9–16.
4. Zhao Z, Lan Y, Bai S, Shen J, Xiao S, Lv R, et al. Late-onset radiationinduced optic neuropathy after radiotherapy for nasopharyngeal car-cinoma. J Clin Neurosci 2013;20:702–6.
5. Wan XB, Zhao Y, Fan XJ, Cai HM, Zhang Y, Chen MY, et al. Molecularprognostic prediction for locally advanced nasopharyngeal carcinomaby support vector machine integrated approach. PLoS One 2012;7:e31989.
6. Motley AM, Berg N, Taylor MJ, Sahlender DA, Hirst J, Owen DJ, et al.Functional analysis of AP-2a andmu2 subunits. Mol Biol Cell 2006;17:5298–308.
7. Pellikainen J, Kataja V, Ropponen K, Kellokoski J, Pietil€ainen T, B€ohmJ, et al. Reduced nuclear expression of transcription factor AP-2associates with aggressive breast cancer. Clin Cancer Res 2002;8:3487–95.
8. Ropponen KM, Kellokoski JK, Pirinen RT, Moisio KI, Eskelinen MJ,Alhava EM, et al. Expression of transcription factor AP-2 in colorectal
adenomas and adenocarcinomas comparison of immunohistochem-istry and in situ hybridisation. J Clin Pathol 2001;54:533–8.
9. Eckert D, Buhl S, Weber S, J€ager R, Schorle H. The AP-2 family oftranscription factors. Genome Biol 2005;6:246.
10. Cheng C, Ying K, Xu M, Zhao W, Zhou Z, Huang Y, et al. Cloning andcharacterization of a novel human transcription factor AP-2b like gene(TFAP2BL1). Int J Biochem Cell Biol 2002;34:78–86.
11. Zhao F, Satoda M, Licht JD, Hayashizaki Y, Gelb BD. Cloning andcharacterization of a novel mouse AP-2 transcription factor, AP-2d,with unique DNA binding and transactivation properties.J Biol Chem2001;276:40755–60.
12. Feng W, Williams T. Cloning and characterization of the mouse AP-2egene: a novel family member expressed in the developing olfactorybulb. Mol Cell Neurosci 2003;24:460–75.
13. Wang HV, Vaupel K, Buettner R, Bosserhoff AK, Moser M. Identifica-tion and embryonic expression of a new AP-2 transcription factor, AP-2e. Dev Dyn 2004;231:128–35.
14. Hilger-Eversheim K, Moser M, Schorle H, Buettner R. Regulatory rolesof AP-2 transcription factors in vertebrate development, apoptosis andcell-cycle control. Gene 2000;260:1–12.
15. Werling U, Schorle H. Transcription factor gene AP-2g essential forearly murine development. Mol Cell Biol 2002;22:3149–56
Shi et al.
Cancer Prev Res; 7(2) February 2014 Cancer Prevention Research276
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271

16. Wu F, Lee AS. YY1 as a regulator of replication-dependent hamsterhistone H3.2 promoter and an interactive partner of AP-2. J Biol Chem2001;276:28–34.
17. McPherson LA, Loktev AV, Weigel RJ. Tumor suppressor activity ofAP-2a mediated through a direct interaction with p53. J Biol Chem2002;277:45028–33.
18. Batsch�eE,MuchardtC,Behrens J,HurstHC,Cr�emisiC.RBandc-Mycactivate expression of the E-cadherin gene in epithelial cells throughinteraction with transcription factor AP-2. Mol Cell Biol 1998;18:3647–58.
19. Aqeilan RI, Palamarchuk A, Weigel RJ, Herrero JJ, Pekarsky Y, CroceCM. Physical and functional interactions between the Wwox tumorsuppressor protein and the AP-2g transcription factor. Cancer Res2004;64:8256–61.
20. Braganca J, Eloranta JJ, Bamforth SD, Ibbitt JC, Hurst HC, Bhatta-charya S. Physical and functional interactions among AP-2 transcrip-tion factors, p300/CREB-binding protein, and CITED2. J Biol Chem2003;278:16021–9.
21. Karjalainen JM, Kellokoski JK, Eskelinen MJ, Alhava EM, Kosma VM.Down-regulation of transcription factor AP-2 predicts poor survivalin stage I cutaneous malignant melanoma. J Clin Oncol 1998;16:3584–91.
22. Bar-Eli M. Role of AP-2 in tumor growth and metastasis of humanmelanoma. Cancer Metastasis Rev 1999;18:377–85.
23. Ruiz M, Pettaway C, Song R, Stoeltzing O, Ellis L, Bar-Eli M. Activatorprotein 2a inhibits tumorigenicity and represses vascular endothelialgrowth factor transcription in prostate cancer cells. Cancer Res 2004;64:631–8.
24. Douglas DB, Akiyama Y, Carraway H, Belinsky SA, Esteller M, Gab-rielson E, et al. Hyper-methylation of a small CpG uanine-rich regioncorrelates with loss of activator protein-2a expression during progres-sion of breast cancer. Cancer Res 2004;64:1611–20.
25. Anttila MA, Kellokoski JK, Moisio KI, Mitchell PJ, Saarikoski S,Syrj€anen K, et al. Expression of transcription factor AP-2a predictssurvival in epithelial ovarian cancer. Br J Cancer 2000;82:1974–83.
26. Heimberger AB, McGary EC, Suki D, Ruiz M,WangH, et al. Loss of theAP-2a transcription factor is associated with the grade of humangliomas. Clin Cancer Res 2005;11:267–72.
27. Wang W, Lv L, Pan K, Zhang Y, Zhao JJ, Chen JG, et al. Reducedexpression of transcription factor AP-2a is associated with gastricadenocarcinoma prognosis. PLoS One 2011;6:e24897.
28. Schwartz B,Melnikova VO, Tellez C,Mourad-Zeidan A, BlehmK, ZhaoYJ, et al. Lossof AP-2a results in deregulation of E-cadherin andMMP-9 and an increase in tumorigenicity of colon cancer cells in vivo.Oncogene 2007;26:4049–58.
29. Nordentoft I, Dyrskjøt L, Bødker JS,Wild PJ, HartmannA, Bertz S, et al.Increased expression of transcription factor TFAP2a correlates withchemo-sensitivity in advanced bladder cancer. BMC Cancer 2011;11:135.
30. Sumigama S, Ito T, Kajiyama H, Shibata K, Tamakoshi K, Kikkawa F,et al. Suppression of invasion andperitoneal carcinomatosis of ovariancancer cells by over-expression of AP-2a. Oncogene 2004;23:5496–504.
31. Kaluz S, Kaluzov�a M, Liao SY, Lerman M, Stanbridge EJ. Transcrip-tional control of the tumor- and hypoxia-marker carbonic anhydrase 9:a one transcription factor (HIF-1) show? Biochim Biophys Acta2009;1795:162–72.
32. L�opez-L�azaro M. Role of oxygen in cancer: looking beyond hypoxia.Anticancer Agents Med Chem 2009;9:517–25.
33. Forsythe JA, Jiang BH, Iyer NV, Agani F, Leung SW, Koos RD, et al.Activation of vascular endothelial growth factor gene transcription byhypoxia-inducible factor 1. Mol Cell Biol 1996;16:4604–13.
34. Cheeseman MT, Tyrer HE, Williams D, Hough TA, Pathak P, RomeroMR, et al. HIF-VEGF pathways are critical for chronic otitis media inJunbo and Jeff mouse mutants. PLoS Genet 2011;7:e1002336.
35. Rathinavelu A, Narasimhan M, Muthumani P. A novel regulation ofVEGF expression by HIF-1a and STAT3 in HDM2 transfected prostatecancer cells. J Cell Mol Med 2012;16:1750–7.
36. Falk T, Congrove NR, Zhang S, McCourt AD, Sherman SJ, McKay BS.PEDF and VEGF-A output from human retinal pigment epithelial cellsgrown on novel micro-carriers. J Biomed Biotechnol 2012;2012:278932.
37. Lombaert N, Castrucci E, Decordier I, Van Hummelen P, Kirsch-Volders M, Cundari E, et al. Hard-metal (WC-Co) particles trigger asignaling cascade involving p38 MAPK, HIF-1a, HMOX1, and p53activation in human PBMC. Arch Toxicol 2013;87:259–68.
38. TungKH, LinCW, KuoCC, Li LT, KuoYH, LinCW, et al. CHCpromotestumor growth and angiogenesis through regulation of HIF-1a andVEGF signaling. Cancer Lett 2013;331:58–67.
39. Phillips RJ, Mestas J, Gharaee-Kermani M, Burdick MD, Sica A,Belperio JA, et al. Epidermal growth factor and hypoxia-inducedexpressionofCXCchemokine receptor 4onnon-small cell lung cancercells is regulated by the phosphatidylinositol 3-kinase/PTEN/AKT/mammalian target of rapamycin signaling pathway and activation ofhypoxia inducible factor-1a. J Biol Chem 2005;280:22473–81.
40. Rapisarda A,Melillo G. Role of the VEGF/VEGFR axis in cancer biologyand therapy. Adv Cancer Res 2012;114:237–67.
41. Ahluwalia A, Tarnawski AS. Critical role of hypoxia sensor—HIF-1a inVEGF gene activation. Implications for angiogenesis and tissue injuryhealing. Curr Med Chem 2012;19:90–7.
42. ThomasAL,MorganB,Drevs J,UngerC,WiedenmannB, VanhoeferU,et al. Vascular endothelial growth factor receptor tyrosine kinaseinhibitors: PTK787/ZK 222584. Semin Oncol 2003;30:32–8.
43. Lai LJ, Ho TC. Pigment epithelial-derived factor inhibits c-FLIP expres-sion and assists ciglitazone induced apoptosis in hepatocellular car-cinoma. Anticancer Res 2011;31:1173–80.
44. Hoshina D, Abe R, Yamagishi SI, Shimizu H. The role of PEDF in tumorgrowth and metastasis. Curr Mol Med 2010;10:292–5.
TFAP2A Regulates Nasopharyngeal Carcinoma Growth and Survival
www.aacrjournals.org Cancer Prev Res; 7(2) February 2014 277
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271

2014;7:266-277. Published OnlineFirst December 12, 2013.Cancer Prev Res Dingbo Shi, Fangyun Xie, Yun Zhang, et al.
Signaling PathwayαSurvival by Targeting HIF-1TFAP2A Regulates Nasopharyngeal Carcinoma Growth and
Updated version
10.1158/1940-6207.CAPR-13-0271doi:
Access the most recent version of this article at:
Material
Supplementary
1
http://cancerpreventionresearch.aacrjournals.org/content/suppl/2013/12/12/1940-6207.CAPR-13-0271.DCAccess the most recent supplemental material at:
Cited articles
http://cancerpreventionresearch.aacrjournals.org/content/7/2/266.full#ref-list-1
This article cites 44 articles, 17 of which you can access for free at:
Citing articles
http://cancerpreventionresearch.aacrjournals.org/content/7/2/266.full#related-urls
This article has been cited by 2 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerpreventionresearch.aacrjournals.org/content/7/2/266To request permission to re-use all or part of this article, use this link
Research. on May 28, 2020. © 2014 American Association for Cancercancerpreventionresearch.aacrjournals.org Downloaded from
Published OnlineFirst December 12, 2013; DOI: 10.1158/1940-6207.CAPR-13-0271