Embed Size (px)
Citation preview

Mutagenesis vol. 26 no. 6 pp. 771–781, 2011 doi:10.1093/mutage/ger045Advance Access Publication 19 July 2011
The ability of the mouse lymphoma TK assay to detect aneugens
Michael D. Fellows*, Ann T. Doherty, CatherineC. Priestley, Victoria Howarth and Michael R. O’Donovan
AstraZeneca, R&D Alderley Park, Macclesfield, Cheshire SK10 4TG, UK.
*To whom correspondence should be addressed. Tel: þ44 1625 230401; Faxþ44 1625 231281; Email: [email protected]
Received on March 31, 2011; revised on May 23, 2011;accepted on May 31, 2011
There is some evidence that the mouse lymphoma TKassay (MLA) can detect aneugens, and this is accepted inthe current International Conference on Harmonisationguidance for testing pharmaceuticals. However, whetheror not it can be used as a reliable screen for aneugenicityhas been the subject of debate. Consequently, aneugenswith diverse mechanisms of action were tested in theMLA using 24-h exposure. No evidence of increasedmutant frequency was seen with noscapine, diazepam orcolchicine and increases were seen with taxol, carben-dazim, econazole and chloral hydrate only at high levelsof toxicity (for all but one taxol concentration survivalreduced to £10% of control). None of these agents wouldbe unequivocally classified as positive using currentlyaccepted criteria. The largest increases in mutantnumber were seen with taxol and carbendazim; there-fore, trifluorothymidine (TFT)-resistant clones resultingfrom treatment with them were cultured and analysedfor chromosome 11 copy number using fluorescent in situhybridisation (FISH) and loss of heterozygosity (LOH).High concentrations of these aneugens induced LOH atall loci examined indicating only one chromosome 11 waspresent but, perhaps surprisingly, all were found to havetwo copies of chromosome 11 using FISH. This would beconsistent with loss of the tk1 chromosome 11b withconcomitant duplication of chromosome 11a, which hasbeen proposed as a likely mechanism for induction ofTFT-resistant clones. However, it was also surprisingthat analysis of centromere size showed that almost allthe clones had both small and large centromeres, i.e.suggesting the presence of both chromosomes 11a and11b. In conclusion, it appears that the TFT-resistantmutants resulting from treatment with toxic concen-trations of some aneugens such as taxol and carbenda-zim have undergone complex genetic changes. However,these data show that the MLA cannot be used asa routine screen to detect aneugens.
Introduction
Aneuploidy has been linked with several conditions delete-rious to human health, including 35% of spontaneousabortions, 0.3% of live births with increased chance of birth
defects, carcinogenesis and the progression of cancer (1,2).Hence, it would be valuable to identify chemicals witha potential aneugenic mechanism in an in vitro screeningassay. The mouse lymphoma TK assay (MLA) has beenshown to be able to detect a range of genetic events, includingpoint mutations and chromosomal rearrangements (3).Although the International Conference on HarmonisationS2B guidance states that the MLA is able to detectcompounds that induce numerical chromosomal damage andthat the detection of aneuploidy inducers is enhanced if the24-h treatment regimen is used with the microtitre method (4),over the last decade, there has been some debate aboutwhether, or not, the MLA responds to aneugens as a class.Theoretically, if an aneugen induces loss of the copy ofchromosome 11b containing the functional tkþ allele, thisshould be expressed as a trifluorothymidine (TFT)-resistantmutant. Several studies have reported positive results withaneugens including colchicine (5 and Dr Raymond Proudlock,personal communication, Charles Rivers Laboratories), vin-blastine (5) carbendazim (6) and taxol (7). However, with theexception of carbendazim, these positive results have not beeneasy to reproduce in different laboratories (6). Furthermore,although a strain of L5178Y cells with only one intact copy ofchromosome 11 is known (8), it has been suggested thatL5178Y TKþ/� clone 3.7.2c cells (the cell line used in theMLA) that are monosomic for chromosome 11 are not viable(9,10). This would be a pre-requisite if TFT-resistant mutantswere to be induced by simple chromosome loss. If aneugensdo induce TFT-resistant mutant cells, it is possible that theyare the result of more complex mechanisms, for examplechromosome loss and subsequent recombination and dupli-cation (11,12) or simply the induction of chromosomedamage since it is known that most aneugens can also inducestructural changes to some extent.
‘Aneugen’ is an umbrella term for compounds that caninteract with various components of the cell division apparatussuch as microtubules, centrosomes and centromere-associatedproteins to give changes in chromosome number. In order toinvestigate the response of the MLA to aneugens, sevencompounds with differing modes of action were tested (Figure1). To elucidate the mechanism of the responses to carbenda-zim and taxol, loss of heterozygosity (LOH) at four micro-satellite markers on chromosome 11 and chromosome copynumber changes was investigated using fluorescent in situhybridisation (FISH) with chromosome 11 probes. Further-more, it has previously been observed that the two copies ofchromosome 11 have different size centromeres and the largercentromere is the TK competent 11b (24,25); the initialobservations were made using G-banding and subsequentlyconfirmed using FISH (7). Accordingly, centromere sizeanalysis was performed to determine which copies ofchromosome 11 were present in TFT-resistant clones followinganeugen treatment.
� The Author 2011. Published by Oxford University Press on behalf of the UK Environmental Mutagen Society.
All rights reserved. For permissions, please e-mail: [email protected]. 771
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from
Materials and methods
All chemicals and reagents were purchased from Sigma Aldrich unlessotherwise stated.
Test agents
Taxol, carbendazim, noscapine, econazole, chloral hydrate, diazepam,colchicine and 4-nitroquinoline-N-oxide (4-NQO) were dissolved in dimethylsulphoxide immediately before use.
Cell line and culture conditions
Mouse lymphoma L5178Y TKþ/� cells, clone 3.7.2c, were obtained fromDr J. Cole (MRC Cell Mutation Unit, Brighton, UK) to whom they had beensupplied by Dr Don Clive (then at Burroughs Welcome, Research Triangle
Park, USA) in 1978. The provenance of the cells is such that it can bedemonstrated that the cryopreserved master stock cultures in this laboratoryhave spent no more than 3 weeks in continuous culture since they weresupplied by Dr Don Clive and working cell cultures were cleansed of pre-existing TFT-resistant mutants by growth in medium containing methotrexateon only one occasion (Figure 2). All cells were cultured in RPMI 1640 medium(Invitrogen, Paisley, UK) containing 10% heat-inactivated donor horse serum(DHS), 2 mmol/l L-glutamine, 2 mmol/l sodium pyruvate, 1% Pluronic F68,200 IU/ml penicillin, 200 lg/ml streptomycin and maintained at 37�C ina humidified atmosphere of 5% CO2 in air. The average doubling time of theL5178Y cells in stationary culture was �9 to 10 h.
The karyotype of the master cultures including modal chromosome numberand the copy number of chromosome 11 were determined using the methodsdescribed below.
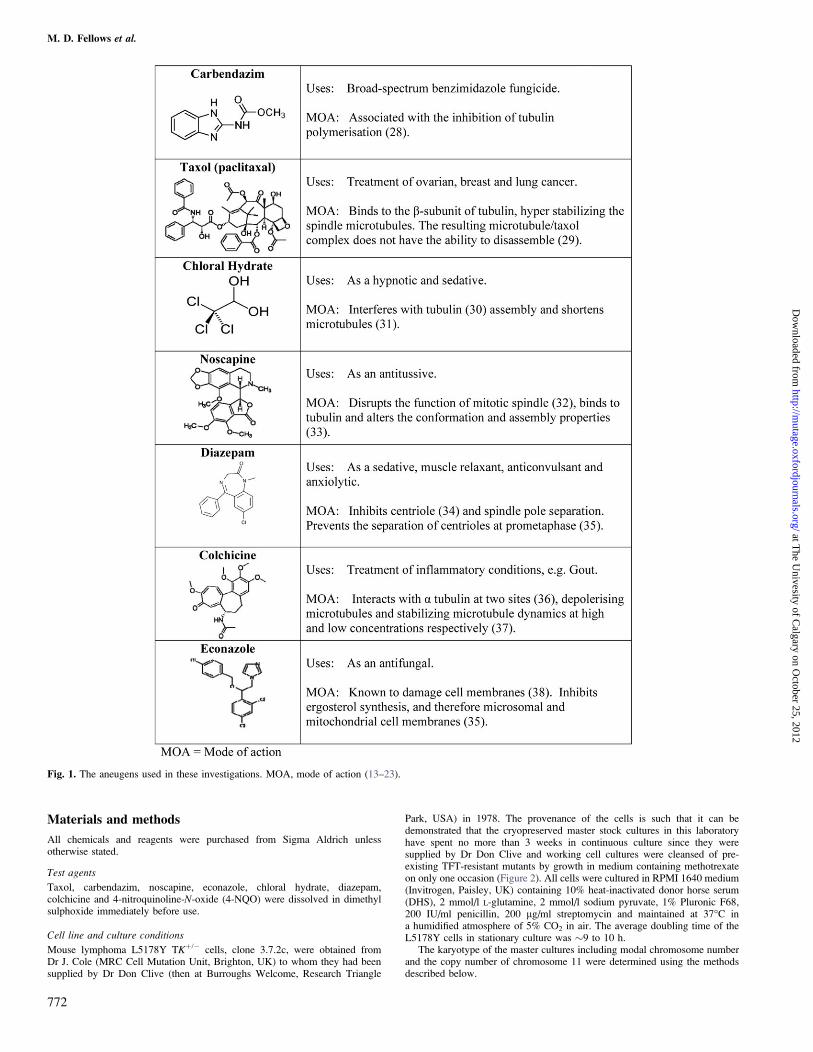
Fig. 1. The aneugens used in these investigations. MOA, mode of action (13–23).
M. D. Fellows et al.
772
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from
Microtitre method to determine mutant frequency at the tk locus (MLA)
The MLA was performed essentially as described by Clements (28) withtreatment for 24 h in the absence of an exogenous metabolising system. Thisexposure condition is considered to be the most suitable to identify division-dependent mutagens and aneugens (29). For each treatment, 4 � 106 cellswere suspended in 20 ml RPMI containing 10% DHS and the test agent,solvent and positive control solutions were added at 1% v/v. Cultures wereprepared for controls and each test agent concentration as identified in theTables II–IV. 4-NQO was used as the positive control. Following treatment,the cells were centrifuged, washed once and resuspended in 20 ml RPMImedium containing 10% DHS, counted and adjusted to 2 � 105 cells/ml.Cultures were maintained at between �1.5 � 105 and 1 � 106 cells/ml for 2days and then selected to determine cell viability and resistance to TFT. Todetermine viability, cultures were plated into two 96-well plates at �1.6 cellsper well in RPMI medium containing 20% DHS. To determine TFTresistance, cultures were plated into two 96-well plates at �2000 cells perwell in RPMI medium containing 20% DHS and 3 lg/ml TFT. Plates wereincubated for up to 14 days, prior to scoring; for TFT-resistant colonies,
mutant clones were designated as large (covering approximately one quarteror more of the area of the well) or small (covering less than one quarter of thearea of the well). Viability and mutant frequency were calculated usingstandard methods (28).
A positive result was determined using the global evaluation factor, i.e. forany concentration, an increase in mutant frequency above concurrent solventcontrol of �126 � 10�6 was considered significant. Toxicity was measuredby reduction in relative total growth (RTG), which is the product ofsuspension growth and Day 2 cloning efficiency. This is the recommendedmethod for toxicity assessment in the MLA (28). Increases in mutantfrequency seen at ,10% RTG were not considered to be biologicallysignificant (30).
In order to investigate the possible genetic changes resulting in TFT-resistant clones, clones were isolated after treatment with two of the aneugensthat gave the largest increases in mutant frequency. For carbendazim (Test 3)and taxol (Test 4), an additional four TFT plates for each control and testcompound concentration were prepared. Where possible, �7 and 14 days afterplating at least 10 large and 10 small mutant clones, respectively, for eachconcentration were picked and grown in RPMI medium containing 10% DHSfor 7–14 days for subsequent LOH and FISH analysis of chromosome 11.
Analysis for LOH
Genomic DNA was extracted from TFT-resistant clones using the QiaAmpDNA mini kit according to the manufactures protocol (Qiagen, Crawley, UK).Primer sequences for the polymerase chain reaction (PCR) analysis of LOHon chromosome 11 at microsatellites D11Agl1 (Tk), D11Mit29, D11Mit42and D11Mit74 (Table I) were taken from Singh et al. (31) and confirmed forMus musculus using the National Centre for Biotechnology Information(NCBI) primer Basic Local Alignment Search Tool (BLAST) database (32).These markers were at a location close to the centromere and both proximaland distal to the tk gene. Each amplification reaction contained 10 pmol ofeach forward and reverse primer (Eurogentech, Southampton, UK), 50 ngDNA and 45 ll of 1.1� Platinum� Blue PCR supermix (Invitrogen) in a finalreaction volume of 50 ll. For D11Agl1 and D11Mit74 loci, amplificationreactions were carried out in multiplex reactions. Reactions were performed ina DNA Engine Dyad� Peltier Thermal Cycler (MJ Research). Cyclingconditions consisted of an initial incubation at 95�C for 2 min, followed bynine cycles of 90�C for 45 sec and 60�C for 45 sec, with a decrease intemperature by 1�C per cycle, followed by 24 cycles of 90�C denaturation for45 sec, 50�C annealing for 45 sec and 72�C extension for 45 sec, with a finalextension at 72�C for 10 min. Reaction products (20 ll in almost all cases)were resolved by electrophoresis on a 2% agarose gel, supplemented with 0.6mg/ml ethidium bromide (Thermo Scientific). Control genomic DNA wasused as a positive control for each run using the same conditions. Initialreactions were performed to confirm LOH at D11Agl1(tk) and D11Mit74 loci(the tk locus and the extremes of the investigated span of chromosome 11),followed by further reactions at internal chromosome 11 loci, D11Mit29 andD11Mit42.
Cytogenetic analysis
Analysis of karyotype. Metaphases were prepared by adding colcemid (10 lg/ml) for 30 mins to cultures containing 5 � 105 cells/ml. The cells wereharvested by suspending in 0.56% potassium chloride at 37�C for 12 min andthen resuspending in 3:1 methanol/acetic acid three times for 5 min and eachthen dropped onto clean slides.
Whole chromosome paints (Cambio) were applied to slides and hybrid-isiation was performed using the HYBrite� in situ hybridisation machine at72�C for 5 min followed by 37�C for 16–20 h. Following hybridisation, slideswere washed in 0.4� saline–sodium citrate (SSC) containing 0.1% triton X at72�C and then removed into 2� SSC wash solution at room temperature.
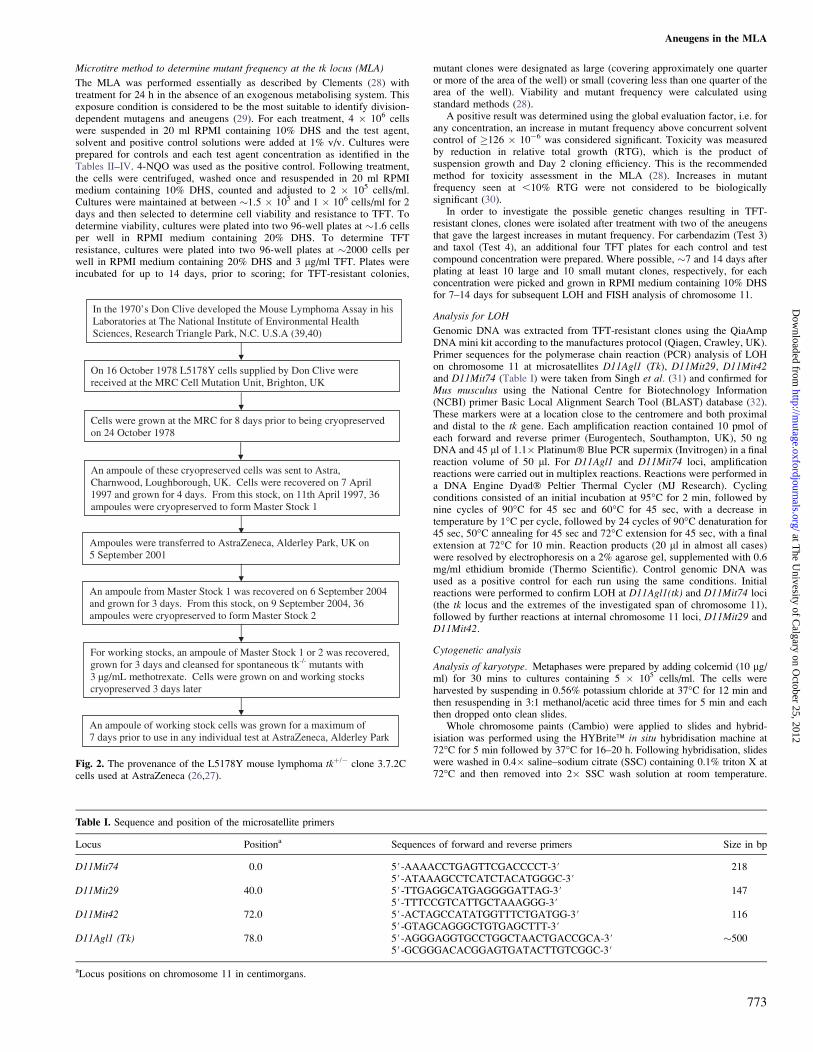
In the 1970’s Don Clive developed the Mouse Lymphoma Assay in his Laboratories at The National Institute of Environmental Health Sciences, Research Triangle Park, N.C. U.S.A (39,40)
On 16 October 1978 L5178Y cells supplied by Don Clive were received at the MRC Cell Mutation Unit, Brighton, UK
Cells were grown at the MRC for 8 days prior to being cryopreserved on 24 October 1978
An ampoule of these cryopreserved cells was sent to Astra, Charnwood, Loughborough, UK. Cells were recovered on 7 April 1997 and grown for 4 days. From this stock, on 11th April 1997, 36 ampoules were cryopreserved to form Master Stock 1
Ampoules were transferred to AstraZeneca, Alderley Park, UK on 5 September 2001
An ampoule from Master Stock 1 was recovered on 6 September 2004 and grown for 3 days. From this stock, on 9 September 2004, 36 ampoules were cryopreserved to form Master Stock 2
For working stocks, an ampoule of Master Stock 1 or 2 was recovered, grown for 3 days and cleansed for spontaneous tk-/- mutants with 3 µg/mL methotrexate. Cells were grown on and working stocks cryopreserved 3 days later
An ampoule of working stock cells was grown for a maximum of 7 days prior to use in any individual test at AstraZeneca, Alderley Park
Fig. 2. The provenance of the L5178Y mouse lymphoma tkþ/� clone 3.7.2Ccells used at AstraZeneca (26,27).
Table I. Sequence and position of the microsatellite primers
Locus Positiona Sequences of forward and reverse primers Size in bp
D11Mit74 0.0 5#-AAAACCTGAGTTCGACCCCT-3# 2185#-ATAAAGCCTCATCTACATGGGC-3#
D11Mit29 40.0 5#-TTGAGGCATGAGGGGATTAG-3# 1475#-TTTCCGTCATTGCTAAAGGG-3#
D11Mit42 72.0 5#-ACTAGCCATATGGTTTCTGATGG-3# 1165#-GTAGCAGGGCTGTGAGCTTT-3#
D11Agl1 (Tk) 78.0 5#-AGGGAGGTGCCTGGCTAACTGACCGCA-3# �5005#-GCGGGACACGGAGTGATACTTGTCGGC-3#
aLocus positions on chromosome 11 in centimorgans.
Aneugens in the MLA
773
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from
Following addition of VectaShield anti-fade mountant containing 4#,6-diamidine-2#-phenylindole dihyrdrochloride (DAPI) (Vector Laboratories),cells were visualised using a fluorescent microscope. Where possible, 100metaphase cells were analysed per slide.
Analysis of L5178Y TFT-resistant mutant clones
Method to determine chromosome 11 copy number. Slides were prepared bycytocentrifugation of 200 ll containing 5 � 105 cells for 8 min at 800revolutions/min using a Shandon Cytospin�. Slides were air-dried and fixed in100% methanol; three replicate slides were prepared for each mutant clone.Whole chromosome paint for chromosome 11 fluorescein isothiocyanate(FITC)-labelled (Cambio UK) and a Cy3�-labelled pancentromeric probe(Cambio UK) were added to each slide. Slides were aged for at least 48 hbefore 10 ll of the probe mix was applied. Cohybridisation of the probes wasperformed using the HYBrite� in situ hybridisation machine at 69�C for 5 minfollowed by 42�C for 16 h. Following hybridisation, slides were washed in0.4� SSC containing 0.1% triton X at 72�C and then removed into 2� SSCwash solution at room temperature. Following addition of VectaShield anti-fade mountant containing DAPI, cells were visualised using a fluorescentmicroscope. Where possible, chromosome 11 copy number was scored until�50 cells with the same copy number were recorded per slide.
Identification of chromosomes 11a and 11b
Nine clones (five large and four small) showing LOH at all four markers wereharvested for metaphase preparations in order to determine the origin of thecopies of chromosome 11. FISH was applied, as above, to the centromeres ofall chromosomes with a Cy3� or FITC-labelled pancentromeric probe and the
individual copies of chromosome 11 identified by a FITC or Cy3�-labelledwhole chromosome paint. In each of the nine clones, 20 metaphase preparationswere scored and all images captured for peer review.
Results
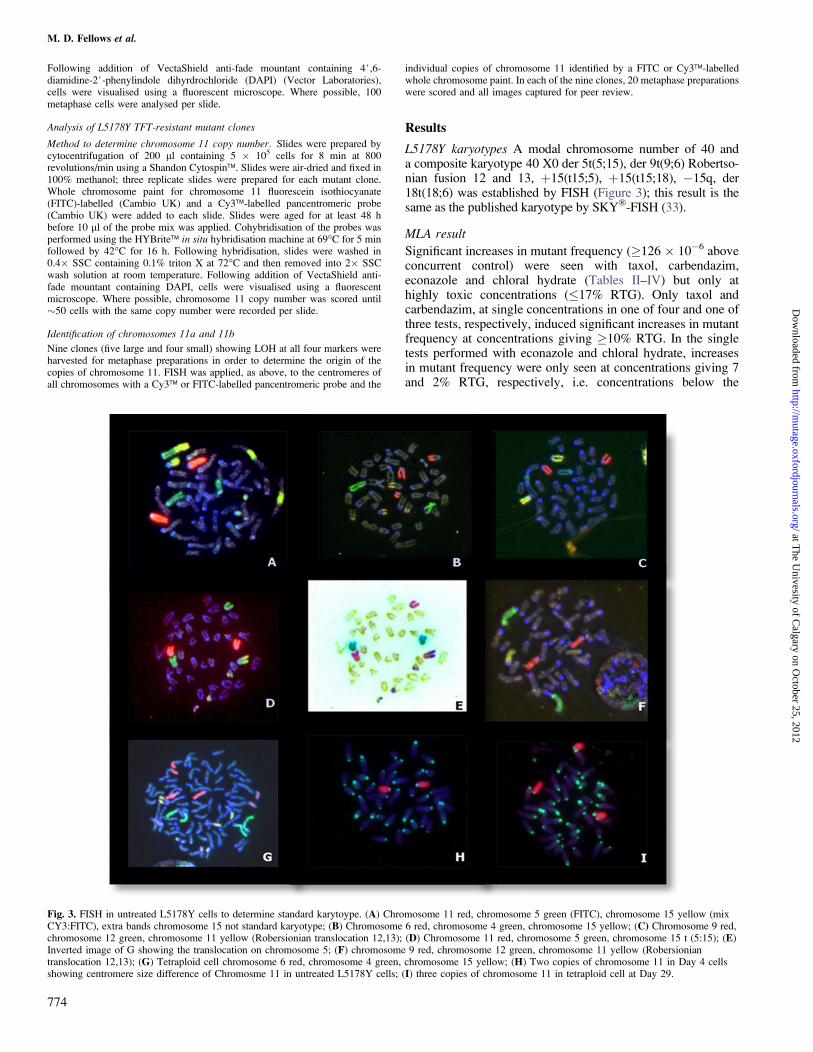
L5178Y karyotypes A modal chromosome number of 40 anda composite karyotype 40 X0 der 5t(5;15), der 9t(9;6) Robertso-nian fusion 12 and 13, þ15(t15;5), þ15(t15;18), �15q, der18t(18;6) was established by FISH (Figure 3); this result is thesame as the published karyotype by SKY�-FISH (33).
MLA result
Significant increases in mutant frequency (�126 � 10�6 aboveconcurrent control) were seen with taxol, carbendazim,econazole and chloral hydrate (Tables II–IV) but only athighly toxic concentrations (�17% RTG). Only taxol andcarbendazim, at single concentrations in one of four and one ofthree tests, respectively, induced significant increases in mutantfrequency at concentrations giving �10% RTG. In the singletests performed with econazole and chloral hydrate, increasesin mutant frequency were only seen at concentrations giving 7and 2% RTG, respectively, i.e. concentrations below the
Fig. 3. FISH in untreated L5178Y cells to determine standard karytoype. (A) Chromosome 11 red, chromosome 5 green (FITC), chromosome 15 yellow (mixCY3:FITC), extra bands chromosome 15 not standard karyotype; (B) Chromosome 6 red, chromosome 4 green, chromosome 15 yellow; (C) Chromosome 9 red,chromosome 12 green, chromosome 11 yellow (Robersionian translocation 12,13); (D) Chromosome 11 red, chromosome 5 green, chromosome 15 t (5:15); (E)Inverted image of G showing the translocation on chromosome 5; (F) chromosome 9 red, chromosome 12 green, chromosome 11 yellow (Robersioniantranslocation 12,13); (G) Tetraploid cell chromosome 6 red, chromosome 4 green, chromosome 15 yellow; (H) Two copies of chromosome 11 in Day 4 cellsshowing centromere size difference of Chromosme 11 in untreated L5178Y cells; (I) three copies of chromosome 11 in tetraploid cell at Day 29.
M. D. Fellows et al.
774
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from
acceptable level of toxicity for analysis in the MLA (30). Also,where there were significant increases in mutant frequency,there was no clear evidence of a preferential increase in eitherthe number of small or the large mutant clones.
None of these results would be considered to be unequivocallypositive using the accepted evaluation criteria for the MLA.However, given the small but reproducible concentration-relatedincreases in mutant frequency, taxol and carbendazim wereconsidered to be weakly positive. Consequently, a further experi-ment was performed with each in order to investigate chromosome11 copy number, using FISH, and LOH to examine the possiblemechanisms responsible for the small increases in mutant frequency.
No significant increase in mutant frequency was seenfollowing exposure to noscapine, diazepam or colchicine, whentested up to concentrations determined by toxicity (Table II).
In all tests where it was used, the positive control 4-NQOinduced a highly significant increase in mutant frequency.
LOH analysis
The mutation assays with taxol and carbendazim and the resultsof PCR analysis for LOH of chromosome 11 of clones fromthem are shown in Tables III and IV and examples of the PCRproducts from carbendazim-treated clones are shown in Figure4. As would be expected from an assay investigating mutationin the tk gene, LOH was often observed at the D11Agl1 (tk)microsatellite marker, independent of LOH at D11Mit74.However, for all clones that demonstrated LOH at tk andD11Mit74, LOH was also seen at D11Mit29 and D11Mit42,regardless of treatment. The vast majority of small clones (95%in taxol Test 4 and 91% in carbendazim Test 3) but fewer large
Table II. Econazole, chloral hydrate, noscapine, diazepam, colchicine, carbendazim and taxol mouse lymphoma assay data
Econazole Chloral hydrate
Concentration (lmol/l) RTG MF % Small clones Concentration (lmol/l) RTG MF % Small clones
0 100 66 43 0 100 88 4014 45 75 53 1700 40 92 4417 29 107 31 2100 27 127 5921 14 154 52 2700 14 116 7627 7 242* 63 3400 2 249* 844-NQO 0.33 39 734* 43 4-NQO 0.33 29 877* 40
Noscapine Diazepam
Concentration (lmol/l) RTG MF % Small clones Concentration (lmol/l) RTG MF % Small clones
0 100 59 45 0 100 52 5234 62 95 21 330 61 98 6342 44 43 50 410 54 67 5852 20 88 29 510 52 74 3666 7 103 36 640 0 130 1004-NQO 0.33 29 636* 44 4-NQO 0.33 40 540* 34
Colchicine Test 1 Colchicine Test 2
Concentration (lmol/l) RTG MF % Small clones Concentration (lmol/l) RTG MF % Small clones
0 100 107 50 0 100 55 330.0125 64 137 42 0.04 78 46 460.025 60 118 47 0.06 52 49 510.031 32 115 44 0.07 20 54 490.038 13 128 35 0.08 5 38 21
Carbendazim Test 1 CarbendazimTest 2
Concentration (lmol/l) RTG MF % Small clones Concentration (lmol/l) RTG MF % Small clones
0 100 65 44 0 100 120 485 91 70 59 5 83 83 658 32 107 39 8 53 139 4310 15 137 27 10 17 231 3013 1 430* 43 13 8 318* 37
16 2 568* 414-NQO 0.33 11 1594* 38 4-NQO 0.33 42 909* 42
Taxol Test 1 Taxol Test 2 Taxol Test 3
Concentration (lmol/l) RTG MF % Small clones Concentration (lmol/l) RTG MF % Small clones Concentration (lmol/l) RTG MF % Small clones
0 100 66 49 0 100 88 34 0 100 97 680.017 62 80 53 0.028 39 104 58 0.018 63 75 410.021 35 92 55 0.036 25 90 16 0.023 31 102 320.026 14 178 34 0.045 7 310* 109 0.028 12 174 370.033 2 521* 64 0.056 2 293* 126 0.036 2 652* 494-NQO 0.33 39 734* 43 4-NQO 0.33 29 877 40 4-NQO 0.33 50 762* 40
MF, mutant frequency � 10�6; RTG, relative total growth.*Significantly different from the control (induced mutant frequency �126 � 10�6).
Aneugens in the MLA
775
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from
clones (77% taxol test and 57% carbendazim test) demon-strated LOH at tk. This is in agreement with previousobservations (9,34,35), where LOH at marker tk was reportedto be more common in small than large mutant clones,indicating a probable intergenic deletion on chromosome 11.
A concentration-dependent increase in LOH at all markerswas seen for both taxol and carbendazim clones, with the highestconcentration of both aneugens giving 95% of small clones and100% of large clones with LOH at all markers. This increase inLOH was not always related to a significant increase in mutantfrequency. In contrast, the incidence of LOH at all markers in theconcurrent control cultures was 22 or 50% for small clones and47 or 25% for large clones in taxol and carbendazim tests,respectively. Furthermore, with 4-NQO, LOH at all markers wasonly seen in �10% to 30% of all clones from both tests.
FISH analysis
The results of the FISH analysis of taxol and carbendazim clonesare shown in Tables III and IV, data and images ofrepresentative cells are shown in Figures 5 and 6. Nearly, allclones, 5 of 5 controls and 27 of 31 aneugen-treated, containeda majority of cells with two copies of chromosome 11 and theremaining 4 clones contained cells with a majority of three or
more copies. The trisomic clones (all induced by carbendazim)were large and also had multiple translocations of chromosome11 material. However, the increase in copy number was notconcentration related. No clone was seen with a majority singlecopy of chromosome 11. The majority of diploid clones didcontain a small proportion (,25%) of cells with one or � threecopies of chromosome 11. No more than 5% of monosomic cellswere seen in any clone and there was no indication ofa relationship between this incidence and aneugen concentration.
Nine clones (five large and four small) isolated after taxoltreatment showing LOH at all four markers were examined todetermine the size of the chromosome 11 centromeres (Table Vwith representative images in Figure 7). In all clones examined,the majority of cells contained two copies of chromosome 11with different sized centromeres. This was consistent with thepublished karyotype for L5178Y cells containing chromo-somes 11a and 11b (Figure 3H) but is not what would beexpected for clones showing apparent complete LOH.
Discussion
The original objective of this study was to determine whetheror not the MLA is able to detect aneugens, although it is
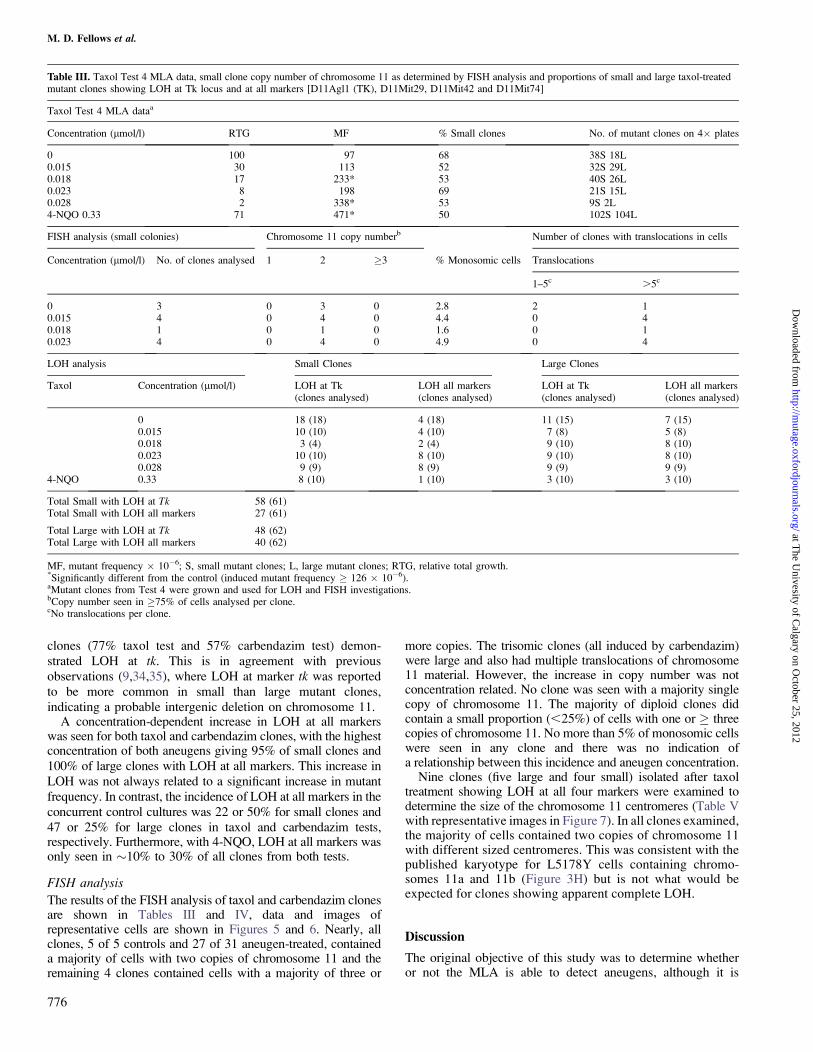
Table III. Taxol Test 4 MLA data, small clone copy number of chromosome 11 as determined by FISH analysis and proportions of small and large taxol-treatedmutant clones showing LOH at Tk locus and at all markers [D11Agl1 (TK), D11Mit29, D11Mit42 and D11Mit74]
Taxol Test 4 MLA dataa
Concentration (lmol/l) RTG MF % Small clones No. of mutant clones on 4� plates
0 100 97 68 38S 18L0.015 30 113 52 32S 29L0.018 17 233* 53 40S 26L0.023 8 198 69 21S 15L0.028 2 338* 53 9S 2L4-NQO 0.33 71 471* 50 102S 104L
FISH analysis (small colonies) Chromosome 11 copy numberb Number of clones with translocations in cells
Concentration (lmol/l) No. of clones analysed 1 2 �3 % Monosomic cells Translocations
1–5c .5c
0 3 0 3 0 2.8 2 10.015 4 0 4 0 4.4 0 40.018 1 0 1 0 1.6 0 10.023 4 0 4 0 4.9 0 4
LOH analysis Small Clones Large Clones
Taxol Concentration (lmol/l) LOH at Tk(clones analysed)
LOH all markers(clones analysed)
LOH at Tk(clones analysed)
LOH all markers(clones analysed)
0 18 (18) 4 (18) 11 (15) 7 (15)0.015 10 (10) 4 (10) 7 (8) 5 (8)0.018 3 (4) 2 (4) 9 (10) 8 (10)0.023 10 (10) 8 (10) 9 (10) 8 (10)0.028 9 (9) 8 (9) 9 (9) 9 (9)
4-NQO 0.33 8 (10) 1 (10) 3 (10) 3 (10)
Total Small with LOH at Tk 58 (61)Total Small with LOH all markers 27 (61)
Total Large with LOH at Tk 48 (62)Total Large with LOH all markers 40 (62)
MF, mutant frequency � 10�6; S, small mutant clones; L, large mutant clones; RTG, relative total growth.*Significantly different from the control (induced mutant frequency � 126 � 10�6).aMutant clones from Test 4 were grown and used for LOH and FISH investigations.bCopy number seen in �75% of cells analysed per clone.cNo translocations per clone.
M. D. Fellows et al.
776
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from
recognised that these are not a single class but rather a diverseset of chemicals that can interact with various components ofthe cell division apparatus. It seems clear that the MLA doesnot respond to all aneugens since no evidence of increasedmutant frequency was seen following exposure to noscapine,diazepam or colchicine, even when tested up to concentrationsreducing survival as measured by RTG to ,10%. Althoughsome evidence of mutagenicity was seen with taxol, carbenda-zim, econazole and chloral hydrate, the increases weregenerally at levels of survival ,10% and it is likely that, iftested as unknown compounds, the majority of these resultswould not be considered positive using currently acceptedmouse lymphoma assay evaluation criteria.
It should be noted that the results obtained with taxol,colchicine and chloral hydrate do not agree with the positiveresponses previously reported for these compounds (5,7,36). Inthe current study, toxicity above and below 10% survival wasseen at the same concentrations of taxol in different experi-ments and this may account for the apparent disagreementbetween laboratories, i.e. results were called positive ornegative depending on whether the increases were seen justabove or below 10% survival. Furthermore, in the laboratorywhere taxol has been reported to be positive, consistent resultswere not obtained between experiments using different lots oftest material (Dr Martha Moore, Division of Genetic andMolecular Toxicology, Federal Drug Administration, personal
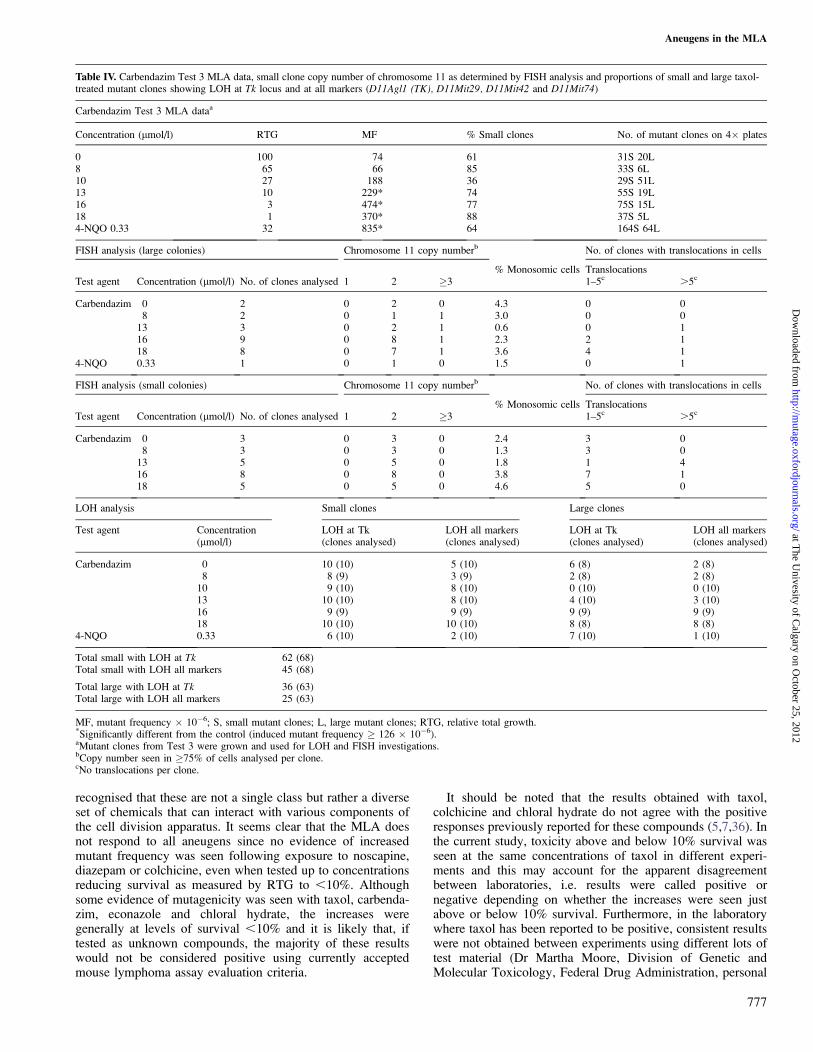
Table IV. Carbendazim Test 3 MLA data, small clone copy number of chromosome 11 as determined by FISH analysis and proportions of small and large taxol-treated mutant clones showing LOH at Tk locus and at all markers (D11Agl1 (TK), D11Mit29, D11Mit42 and D11Mit74)
Carbendazim Test 3 MLA dataa
Concentration (lmol/l) RTG MF % Small clones No. of mutant clones on 4� plates
0 100 74 61 31S 20L8 65 66 85 33S 6L10 27 188 36 29S 51L13 10 229* 74 55S 19L16 3 474* 77 75S 15L18 1 370* 88 37S 5L4-NQO 0.33 32 835* 64 164S 64L
FISH analysis (large colonies) Chromosome 11 copy numberb No. of clones with translocations in cells
Test agent Concentration (lmol/l) No. of clones analysed 1 2 �3% Monosomic cells Translocations
1–5c .5c
Carbendazim 0 2 0 2 0 4.3 0 08 2 0 1 1 3.0 0 0
13 3 0 2 1 0.6 0 116 9 0 8 1 2.3 2 118 8 0 7 1 3.6 4 1
4-NQO 0.33 1 0 1 0 1.5 0 1
FISH analysis (small colonies) Chromosome 11 copy numberb No. of clones with translocations in cells
Test agent Concentration (lmol/l) No. of clones analysed 1 2 �3% Monosomic cells Translocations
1–5c .5c
Carbendazim 0 3 0 3 0 2.4 3 08 3 0 3 0 1.3 3 0
13 5 0 5 0 1.8 1 416 8 0 8 0 3.8 7 118 5 0 5 0 4.6 5 0
LOH analysis Small clones Large clones
Test agent Concentration(lmol/l)
LOH at Tk(clones analysed)
LOH all markers(clones analysed)
LOH at Tk(clones analysed)
LOH all markers(clones analysed)
Carbendazim 0 10 (10) 5 (10) 6 (8) 2 (8)8 8 (9) 3 (9) 2 (8) 2 (8)
10 9 (10) 8 (10) 0 (10) 0 (10)13 10 (10) 8 (10) 4 (10) 3 (10)16 9 (9) 9 (9) 9 (9) 9 (9)18 10 (10) 10 (10) 8 (8) 8 (8)
4-NQO 0.33 6 (10) 2 (10) 7 (10) 1 (10)
Total small with LOH at Tk 62 (68)Total small with LOH all markers 45 (68)
Total large with LOH at Tk 36 (63)Total large with LOH all markers 25 (63)
MF, mutant frequency � 10�6; S, small mutant clones; L, large mutant clones; RTG, relative total growth.*Significantly different from the control (induced mutant frequency � 126 � 10�6).aMutant clones from Test 3 were grown and used for LOH and FISH investigations.bCopy number seen in �75% of cells analysed per clone.cNo translocations per clone.
Aneugens in the MLA
777
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from
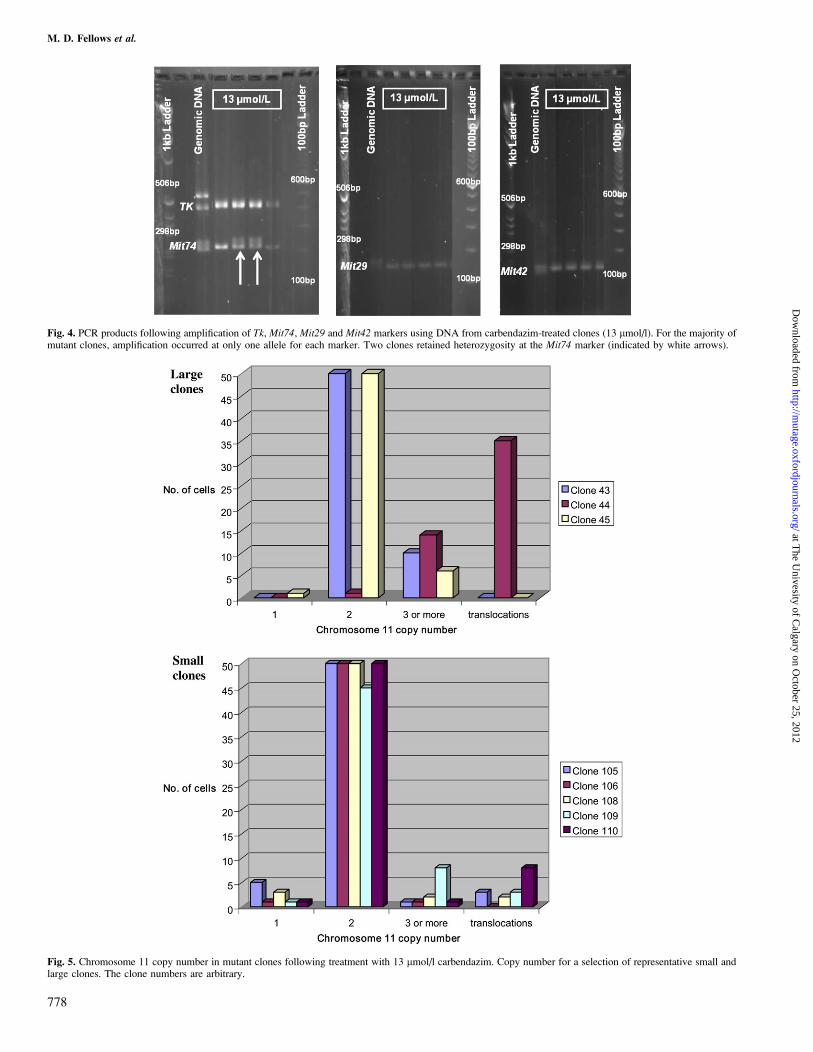
Fig. 5. Chromosome 11 copy number in mutant clones following treatment with 13 lmol/l carbendazim. Copy number for a selection of representative small andlarge clones. The clone numbers are arbitrary.
Fig. 4. PCR products following amplification of Tk, Mit74, Mit29 and Mit42 markers using DNA from carbendazim-treated clones (13 lmol/l). For the majority ofmutant clones, amplification occurred at only one allele for each marker. Two clones retained heterozygosity at the Mit74 marker (indicated by white arrows).
M. D. Fellows et al.
778
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from

communication). For colchicine, concentration-related increasesin mutant frequency from 0.01 to 0.1 lmol/l have been reported(5) but, again, these were most apparent at concentrations giving,20% survival. However, it is possible that the karyotype ofcells used may also have influenced the response; the cellsreported to give positive results contained a population of 2.7%with three copies of chromosome 11 (5), but no such cells wereseen in 600 metaphases in this laboratory. It is theoreticallypossible that loss of a single chromosome 11b from a trisomic(11a, 11a and 11b) could give rise to a viable TFT-resistantclone, whereas there are data to indicate that cells monosomicfor chromosome 11, which would obviously result from loss ofa single chromosome from a disomic, are not viable (9,10).
Although the increases in mutant frequency with taxol andcarbendazim were seen at high levels of toxicity, they didrepresent increases in real numbers of mutant colonies notsimply a calculated increase resulting from reduced cloningefficiency. Therefore, TFT-resistant clones surviving treat-ment with concentrations of each aneugen were analysed forLOH and chromosome 11 karyotype. All the clones treatedwith high concentrations of either compound showed LOH atthe tk locus, as would be expected for TFT-resistant mutants.FISH analysis showed that the majority of cells in every clonehad two or more copies of chromosome 11. Although all themutant clones did contain a small proportion of cellsmonosomic for chromosome 11 (,5%), it is unlikely thatthey were related to the induction of TFT resistance by theaneugens. The clones were analysed after growth for at least 3weeks after treatment, with much of this time in 96-well platesin medium containing TFT. It is probable that the non-diploidcells represent terminal events occurring with time in culture
because the incidence was similar in both control andaneugen-treated clones. Hence, there was no evidence tosuggest any TFT-resistant clone arose from monosomy ofchromosome 11a.
It was notable that at the highest concentration tested oftaxol and carbendazim, 95% of small and 100% of largemutant clones analysed had LOH at all markers, although theyhad two copies of chromosome 11. It also appears that theLOH was a consequence of treatment with the aneugensbecause only 35% (18/51) control and 18% (7/40) 4-NQO-induced clones showed LOH at all four markers. Honma et al.found similar differences between the LOH for spontaneousand treatment-induced clones with colchicine and vinblastin(5). The mechanism by which this occurs has not beenelucidated, but this may contribute to the difficulty inidentifying responses to anuegens in the MLA. The increasedlevel of LOH following aneugen treatment was consistentwith loss of chromosome 11b with concomitant duplication ofchromosome 11a, which has been proposed as the most likelymechanism responsible for the induction of TFT-resistantmutants by aneugens in the MLA (5,7,11). If the mutantclones showing LOH at all markers had lost chromosome 11band duplicated 11a, it would be expected that all would havecentromeres of the same size, i.e. the small centromere from11a. However, the large majority (93/120) of taxol-treatedclones showed both small and large centromeres, and onlytwo had the expected two small centromeres. The reasons forthis are not known but could represent complex chromosomerearrangements such as whole-arm translocations.
Since it is clear that all aneugens are not readily detected inthe MLA, it is relevant to consider possible alternative tests.The in vitro micronucleus assay (MN(vit)) is accepted inregulatory guidance as being able to detect aneugens (37) andprevious work in this laboratory confirmed colchicine inducesmicronuclei in L5178Y cells. However, it was necessary to testup to highly toxic concentrations (�50% survival as measuredby Relative Increase in Cell Counts or Relative PopulationDoublings) before a positive response was seen (38) and datafrom other laboratories confirm this requirement for colchicine,diethylstilboestrol and vinblastin (39,40). Hence, while theMN(vit) is capable of detecting aneugenic compounds, testingup to levels of toxicity bordering on the unacceptable may bea pre-requisite for some of them.
In conclusion, some aneugens such as taxol and carbendazimappear to be able to induce real increases in numbers of TFT-resistant mutants in mouse lymphoma L5178Y cells but only athighly toxic concentrations (�17% RTG). Furthermore, thesemutants appear to have undergone complex genetic changes
Fig. 6. Chromosome 11 demonstrated by FISH in colonies treated with Carbendazim. (A) Cell from a clone with metaphase plate with two copies of chromosome11 painted green (FITC). (B) Diagram of metaphase plate with painted copies of chromosome 11. (C) Tetraploid cell with a reciprocal translocation in one of thecopies of chromosome 11.
Table V. Chromosome 11 centromere size assessment for selected clonesshowing LOH at all markers
Treatment Clone size Centromere size
S/L L/L S/S ? Total
Taxol 0.015 lmol/l Small 19 0 0 1 20Taxol 0.018 lmol/l Small 20 0 0 0 20Taxol 0.023 lmol/l Small 16 1 0 3 20Taxol 0.023 lmol/l Small 18 1 0 1 20Vehicle control Large 16 2 0 2 20Vehicle control Large 17 0 0 3 20Taxol 0.018 lmol/l Large 16 1 0 3 20Taxol 0.023 lmol/l Large 14 0 2 4 204 NQO 0.33 lmol/l Large 18 0 2 0 20
S, small centromere; L, large centromere; ?, indeterminable sized centromere.
Aneugens in the MLA
779
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from

that are not apparent in TK-deficient clones arising spontane-ously or after treatment with 4-NQO. However, whenconsidered together, these data show that the MLA cannot beused routinely as a screen to detect aneugens.
Acknowledgements
The author wishes to thank Katie Clare, Jenny Molloy and Katie Wood for theirexcellent technical assistance and Joseph Ryan for his work at AstraZeneca asa BSc placement student.
Conflict of interest statement: None declared.
References
1. Aardema, M. J., Albertin, S., Arni, P., Henderson, L. M., Kirsch-Volders, M.,Mackay, J. M., Sarrif, A. M., Stringer, D. A. and Taalman, R. D. (1998)Aneuploidy: a report of an ECETOC task force. Mutat. Res., 410, 3–79.
2. Pacchierotti, F. and Ranaldi, R. (2006) Mechanisms and risk of chemicallyinduced aneuploidy in mammalian germ cells. Curr. Pharm. Des., 12,1489–1504.
3. Applegate, M. L., Moore, M. M., Broder, C. B., Burrell, A., Juhn, G.,Kasweck, K. L., Lin, P. F., Wadhams, A. and Hozier, J. C. (1990)Molecular dissection of mutations at the heterozygous thymidine kinaselocus in mouse lymphoma cells. Proc. Natl Acad. Sci. U S A, 87, 51–55.
4. ICH Topic S2B. (1997) Genotoxicity: A Standard Battery for GenotoxicityTesting for Pharmaceuticals. http://www.ich.org/fileadmin/Public_Web_Site/ICH_Products/Guidelines/Safety/S2_R1/Step4/S2B_step_4.pdf (accessedJanuary 14, 2011).
5. Honma, M., Momose, M., Sakamoto, H., Sofuni, T. and Hayashi, M.(2001) Spindle poisons induce allelic loss in mouse lymphoma cellsthrough mitotic non-disjunction. Mutat. Res., 493, 101–114.
6. O’Donovan, M. R., Clare, M. G., Clements, J., Fellows, M. and Oliver, J.(1999) Are aneugens mutagenic in the mouse lymphoma TK assay?Abstracts of the United Kingdom environmental mutagen society 23(rd)annual general meeting, June 23-25, university of Ulster, Coleraine,Northern Ireland, UK. Mutagenesis, 14, 639–656.
7. Wang, J., Sawyer, J. R., Chen, L., Chen, T., Honma, M., Mei, N. andMoore, M. M. (2009) The mouse lymphoma assay detects recombination,deletion, and aneuploidy. Toxicol. Sci., 109, 96–105.
8. Evans, H. H., Mencl, J., Horng, M.-F., Ricanati, M., Sanchez, C. andHozier, J. (1986) Locus specificity in the mutability of L5178Y mouselymphoma cells: the role of multilocus lesions. Proc. Natl Acad. Sci. U S A,83, 4379–4383.
9. Liechty, M. C., Scalzi, J. M., Sims, K. R., Crosby, H., Jr., Spencer, D. L.,Davis, L. M., Caspary, W. J. and Hozier, J. C. (1998) Analysis of large andsmall colony L5178Y tk-/- mouse lymphoma mutants by loss ofheterozygosity (LOH) and by whole chromosome 11 painting: detectionof recombination. Mutagenesis, 13, 461–474.
10. Fellows, M., Doherty, A., Hayes, J., Thompson, A. and O’Donovan, M. R.(2005) In the mouse lymphoma Tk assay, are Tk�/� mutant clones formedfollowing treatment with the aneugen Carbendazim monosomic for
chromosome 11? Abstracts of the 28th Annual Meeting of the UnitedKingdom Environmental Mutagen Society, August 3–5, at the University ofBradford, UK. Mutagenesis, 20, 469–488.
11. de Nooij-van Dalen, A. G., van Buuren-van Seggelen, V. H. A.,Lohman, P. H. M. and Giphart-Gassle, M. (1998) Chromosome loss withconcomitant duplication and recombination both contribute most to loss ofheterozygosity in vitro. Genes Chromosomes Cancer, 21, 30–38.
12. Wijnhoven, S. W., Sonneveld, E., Kool, H. J., van Teijlingen, C. M. andVrieling, H. (2003) Chemical carcinogens induce varying patterns of LOHin mouse T-lymphocytes. Carcinogenesis, 24, 139–144.
13. Kirsch-Volders, M., Vanhauwaert, A., Eichenlaub-Ritter, U. andDecordier, I. (2003) Indirect mechanisms of genotoxicity. Toxicol. Lett.,140–141, 63–74.
14. Schiff, P. B., Fant, J. and Horwitz, S. B. (1979) Promotion of microtubuleassembly in vitro by taxol. Nature, 277, 665–667.
15. Faust, J. B. and Salmon, A. (2003) Evidence on the Carcinogenicty ofChloral hydrate (2003 Update). Reproductive and Cancer HazardAssessment Section, Office of environmental health Hazard assessment,California environmental Protection Agency, http://oehha.ca.gov/prop65/hazard_ident/pdf_zip/chloralhyDhid.pdf. (accessed March 22, 2011).
16. Diguiseppi, J., Gawdi, G. M. and Herman, B. (1987) Chloral hydrate disruptsmitosis by increasing intracellular free calcium. J. Cell Sci., 88, 603–612.
17. Schuler, M., Muehlbauer, P., Guzzie, P. and Eastmond, D. A. (1999)Noscapine hydrochloride disrupts the mitotic spindle in mammalian cellsand induces aneuploidy as well as polyploidy in cultured humanlymphocytes. Mutagenesis, 14, 51–56.
18. Aneja, R., Dhiman, N., Idnani, J., Awasthi, A., Arora, S. K., Chandra, R.and Joshi, H. C. (2007) Preclinical pharmacokinetics of bioavailability ofnoscapine, a tubulin-binding anticancer agent. Cancer Chemother.Pharmacol., 60, 831–839.
19. Andersson, L. C., Lehto, V. P., Stenman, S., Badley, R. A. and Virtanen, I.(1981) Diazepam induces mitotic arrest at prometaphase by inhibitingcentriolar separation. Nature, 291, 247–248.
20. Parry, J. M. and Sors, A. (1993) The detection and assessment of aneugenicpotential of environmental chemicals: the European Community Aneu-ploidy Project. Mutat. Res., 287, 3–15.
21. Wallin, M., Friden, B. and Billger, M. (1988) Studies of the interaction ofchemicals with microtubule assembly in vitro can be used as an assay fordetection of cytotoxic chemicals and possible inducers of aneuploidy.Mutat. Res., 201, 303–311.
22. Jordan, M. A. and Wilson, L. (2004) Microtubules as a target for anticancerdrugs. Nat. Rev. Cancer, 4, 253–265.
23. Gudi, R., Xu, J. and Thilagar, A. (1992) Assessment of the in vivoaneuploidy/micronucleus assay in mouse bone marrow cells with 16chemicals. Environ. Mol. Mutagen., 20, 106–116.
24. Hozier, J., Sawyer, J., Moore, M. M., Howard, B. and Clive, D. (1981)Cytogenetic analysis of the L5178Y/TKþ/- leads to TK-/- mouselymphoma mutagenesis assay system. Mutat. Res., 84, 169–181.
25. Sawyer, J., Moore, M. M., Clive, C. and Hozier, J. (1985) Cytogeneticcharacterization of the L5178Y TKþ/� 3.7.2C mouse lymphoma cell line.Mutat. Res., 147, 243–253.
26. Clive, D., Flamm, W. G., Machesko, M. R. and Bernheim, N. J. (1972) Amutational assay system using the thymidine kinase locus in mouselymphoma cells. Mutat. Res., 16, 77–87.
Fig. 7. Chromosome 11 showing centromere size in clone with LOH at all markers. Cell from a colony treated with 0.015 lmol/l taxol showing size difference incentromeres of chromosome 11 (highlighted in green in the left image and in pink in the right inverted image).
M. D. Fellows et al.
780
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from

27. Clive, D. and Spector, J. F. S. (1975) Laboratory procedures for assessingspecific locus mutations at the TK locus in cultured L5178Y mouselymphoma cells. Mutat. Res., 31, 17–29.
28. Clements, J. (2000) The mouse lymphoma assay. Mutat. Res., 455, 97–110.29. Honma, M., Zhang, L. S., Sakamoto, H., Ozaki, M., Takeshita, K.,
Mamose, M., Hayashi, M. and Sofuni, T. (1999) The need for long-termtreatment in the mouse lymphoma assay. Mutagenesis, 14, 23–29.
30. Moore, M. M., Honma, M., Clements, J. et al. (2006) Mouse lymphomathymidine kinase gene mutation assay: follow-up meeting of theInternational Workshop on Genotoxicity Testing—Aberdeen, Scotland,2003—assay acceptance criteria, positive controls, and data evaluation.Environ. Mol. Mutagen., 47, 1–5.
31. Singh, S. P., Chen, T., Chen, L., Me, N., McLain, E., Samokyszyn, V.,Thaden, J. J., Moore, M. M. and Zimniak, P. (2005) Mutagenic effects of 4-hydroxynonenal triacetate, a chemically protected form of the lipidperoxidation product 4-hydroxynonenal, as assayed in L5178Y/Tkþ/�
mouse lymphoma cells. J Pharmacol. Exp. Ther., 313, 855–861.32. NCBI. (2008) Primer-BLAST [online]. http://www.ncbi.nlm.nih.gov/tools/
primer-blast/ (accessed March 22, 2011).33. Sawyer, J. R., Binz, R. L., Wang, J. and Moore, M. M. (2006) Multicolor
spectral karyotyping of the L5178Y Tkþ/- -3.7.2C mouse lymphoma cellline. Environ. Mol. Mutagen., 47, 127–131.
34. Liechty, M. C., Hassanpour, Z., Hozier, J. C. and Clive, D. (1994) Use ofmicrosatellite DNA polymorphisms on mouse chromosome 11 for in vitroanalysis of thymidine kinase gene mutations. Mutagenesis, 9, 423–427.
35. Liechty, M. C., Crosby, H., Jr., Murthy, A., Davis, L. M., Caspary, W. J.and Hozier, J. C. (1996) Identification of a heteromorphic microsatellitewithin the thymidine kinase gene in L5178Y mouse lymphoma cells.Mutat. Res., 371, 265–271.
36. Harrington-Brock, K., Doerr, C. L. and Moore, M. M. (1998) Mutagenicityof three disinfection by-products: di- and trichloroacetic acid and chloralhydrate in L5178Y / TKþ/�- 3.7.2C mouse lymphoma cells. Mutat. Res.,413, 265–276.
37. OECD Test Guideline 487. http://www.oecd-ilibrary.org/docserver/download/fulltext/9748701e.pdf?expires51305722935&id5id&accname5freeContent&checksum579BAA1F9D18C26EED3D78AE07B3107E2(accessed May 20, 2011).
38. Fellows, M. D. and O’Donovan, M. R. (2010) Etoposide, cadmiumchloride, benzo[a]pyrene, cyclophosphamide and colchicine tested in the invitro mammalian cell micronucleus test (MNvit) in the presence andabsence of cytokinesis block using L5178Y mouse lymphoma cells and 2-aminoanthracene tested in MNvit in the absence of cytokinesis block usingTK6 cells at AstraZeneca UK, in support of OECD draft Test Guideline487. Mutat. Res., 702, 163–170.
39. Kirchner. S. and Zeller, A. (2010) Comparison of different cytotoxicitymeasures for the in vitro micronucleus test (MNVit) in L5178Y tkþ/� cells:summary of 4 compounds (Mitomycin C, Cyclophosphamide, Colchicineand Diethylstilboestrol) with clastogenic and aneugenic mode of action.Mutat. Res., 702, 193–198.
40. Lorge, E. (2010) Comparison of different cytotoxicity measurements for thein vitro micronucleus assay using L5178Y and TK6 cells in support ofOECD draft Test Guideline 487. Mutat. Res., 702, 199–207.
Aneugens in the MLA
781
at The U
nivesity of Calgary on O
ctober 25, 2012http://m
utage.oxfordjournals.org/D
ownloaded from





























