Embed Size (px)
Citation preview

Insect Biochem. Molec. Biol. Vol. 23, No. 4, pp. 439M47, 1993 0965-1748/93 $6.00 + 0.00 Printed in Great Britain. All rights reserved Copyright © 1993 Pergamon Press Ltd
The cDNA and Deduced Protein Sequence of House Fly NADPH-Cytochrome P450 Reductase J. F. KOENER,* F. A. CARIiqO,I" R. FEYEREISEN*~
Received 11 August 1992; revised and accepted 10 November 1992
Antisera to purified house fly NADPH-cytochrome P450 reductase were used to select eDNA clones from an expression library of abdomens of phenobarbital-treated house flies. A partial eDNA of 1841 bp containing a TAG termination codon, a consensus polyadenylation site and 269 bp of Y untranslated sequence was obtained. Sequencing of a genomic clone coupled with mRNA sequencing yielded the complete coding sequence including the starting ATG. The resulting open reading frame of 2013 nucleotides codes for a protein of 671 residues. The native reductase apoprotein has a molecular weight of 76,366 and the deduced molecular weight of the holoenzyme (i.e. with 1 mol of FAD and FMN) is 77,608. The sequence of the house fly P450 reductase protein is highly similar to that of rabbit fiver, the overall amino acid positional identity is 54.5% and the overall identity among eukaryotic P450 reductases is about 25%. The P450 reductase gene of 19-23 kb was located on chromosome III, as shown by comparison of RFLP-patterns of the P450 reductase gene in two house fly strains and their hybrids.
Musca domestica NADPH-cytochrome P450 reductase Flavoprotein Sequence Molecular evolution Chromosome III
INTRODUCTION
NADPH-cytochrome P450 reductase (E.C. 1.6.2.4, here- after called P450 reductase) is a microsomal flavoprotein that plays an essential role in the transfer of reducing equivalents from NADPH to the various microsomal P450 proteins. The N-terminal hydrophobic domain of the enzyme is responsible for anchoring the enzyme to the membrane and its tryptic cleavage abolishes the capacity of the enzyme to reduce P450 proteins in reconstituted systems. The larger C-terminal domain contains 1 mol each of FAD and FMN and is catalyti- cally competent in the reduction of cytochrome c and other artificial acceptors. The reduction of P450 proteins proceeds in several steps, the reduction of FAD by NADPH, the transfer of electrons from FAD to FMN, followed by the reduction of P450 proteins in two one-electron steps (review by Masters and Okita, 1980). In addition, this enzyme also transfers reducing equiva- lents to cytochrome b5 and to microsomal enzymes involved in fatty acid elongation, desaturation and heme oxygenation. Analysis of the protein sequence of rat liver P450 reductase led Porter and Kasper (1986) to
*Department of Entomology and Center for Insect Science, University of Arizona, Tucson, AZ 85721, U.S.A.
tDSIR Plant Protection, Canterbury Agriculture Centre, Lincoln, Christchurch, New Zealand.
:~Author for correspondence.
propose that the enzyme arose through the fusion of two ancestral bacterial genes for flavodoxin (FMN domain) and for ferredoxin-NADP ÷ reductase (FAD domain). In a strain of Bacillus megaterium, a fusion protein between P450am (CYP102) and P450 reductase operates as a soluble NADPH-dependent monooxygenase (Ruettinger et al., 1989). Recently, the C-terminal por- tion of mammalian nitric oxide synthases was shown to be homologous to P450 reductase (Bredt et al., 1991; Xie et al., 1992).
The insect P450 reductase has been solubilized and purified in its native form from the house fly, Musca dornestica (Mayer and Durrant, 1979; Vincent et al., 1983; Ronis et al., 1988) and from the blow fly (Vincent and Terriere, 1985). Its function in microsomal P450- linked reactions has been demonstrated by immuno- inhibition studies (Mayer and Prough, 1977; Crankshaw et al., 1981; Feyereisen and Vincent, 1984; Baldridge and Feyereisen, 1986; Halliday et al., 1986). Other functions of the insect P450 reductase have not been documented to date. As part of our study of the cytochrome P450 monooxygenase system in the house fly and its role in insecticide resistance we have recently cloned and se- quenced a P450 cDNA (Feyereisen et al., 1989). The present paper describes the isolation and sequence of the house fly P450 reductase by a similar strategy, namely the screening of a cDNA expression library with anti- bodies to the house fly enzyme. We also document the
439

440 J.F. KOENER et al.
sequence conservation between vertebrate, yeast, bac- terial and insect P450 reductases, and we report the chromosomal localization of the P450 reductase gene in the house fly.
MATERIALS AND METHODS
Insects
The R-Diazinon (Rutgers) strain and the S- stw ;bwb ;ocra (sbo) strain of M. domestica were provided by F. W. Plapp Jr (Texas A&M University). The mutant sbo strain carries visible recessive markers on chromosomes II (stubby wing), III (brown body) and V (ocra eyes). The flies were reared as described (Feyereisen and Vincent, 1984). Strains that are homozygous for chromosomes II, III or V from the R strain in a sbo background, and backcrosses between male (R × sbo) F1 and female sbo flies were used to determine the chromosomal localization of the P450 reductase gene.
Enzymes and chemicals
Restriction endonucleases were purchased from Bethesda Research Laboratories, New England Biolabs, Promega or Boehringer-Mannheim. Oligonucleotides were synthesized on an Applied Biosystems 380A DNA synthesizer in the Central Service Laboratory, Center for Gene Research and Biotechnology, Oregon State University. Sequenase sequencing kits were obtained from United States Biochemical. 2 DASH vector and Gigapack Gold packaging mix were obtained from Stratagene. A nick translation kit was obtained from Boehringer-Mannheim. Radioactive compounds were obtained from New England Nuclear or Amersham.
Antibodies, cDNA library and screening
Rabbit anti-house fly P450 reductase antisera were obtained previously (Feyereisen and Vincent, 1984). The sera from two rabbits immunized with affinity purified P450 reductase were treated with ammonium sulfate to precipitate the immunoglobulins (IgG fraction), dia- lyzed and stored at -80°C. The cDNA expression library from phenobarbital-treated house flies was con- structed and screened as described (Feyereisen et al., 1989). Screening of the cDNA expression library was essentially as described (Davis et al., 1986) and positive clones were purified through three rounds of plaque purification.
DNA sequencing
Double-stranded DNA from pBluescript clones was sequenced by the dideoxynucleotide method of Sanger et aL (1977) with [~-35S]thio-dATP, dlTP, dGTP and Sequenase. The reactions were primed with Stratagene SK or KS primers, or with custom synthesized oligonu- cleotides of 15-17 bases. Sequences obtained were ana- lyzed by the MicroGenie program of Queen and Korn (1984).
Genomic library screening
A genomic library of M. domestica R-Diazinon (Rutgers) strain was constructed in the vector 2 DASH (Stratagene). Genomic DNA was partially digested with the enzyme MboI and sized by sucrose gradient centrifu- gation. Fragments of c. 20 kb in size were ligated in the Bam HI site of the vector and packaged in vitro with Gigapack Gold in E. coli P2392 cells. A library of 0.9 x 1 0 6 plaque-forming units (clones) was obtained. The library was screened with a nick-translated EcoRI-XbaI fragment of the cDNA clone.
Northern (RNA) blot and dot blots
Poly(A)-enriched RNA from control or phenobarbi- tal-treated flies was separated by electrophoresis and transferred to nitrocellulose as described previously (Feyereisen et al., 1989). Dilution dot blots, hybridiz- ation and analysis were done as described (Carifio et al., 1992).
Restriction fragment length polymorph&m
Genomic DNA was extracted from house fly eggs of the parental R- and S-strains. After digestion with restriction endonucleases, electrophoresis and blotting on Zetaprobe, the Southern blots were hybridized at high stringency with nick-translated DNA of the P450 reductase cDNA (Davis et al., 1986). Restriction frag- ment length polymorphisms found for ClaI and PstI were then analyzed in the backcrosses between male (R × sbo) F1 and female sbo flies.
mRNA sequencing
Oligonucleotides complementary to the Y-end of the reductase cDNA clone were synthesized and end-labeled with [y-32p]ATP via polynucleotide kinase (Davis et al., 1986). After purification on a Nensorb (New England Nuclear) column, 5 ng of the labeled oligonucleotide was annealed in siliconized tubes to 12gg of poly(A)- enriched RNA from phenobarbital-treated house flies. The mRNA was then sequenced by the dideoxy- nucleotide method with AMV reverse transcriptase (Geliebter, 1987).
RESULTS
Cloning and sequencing
The house fly eDNA expression library was screened with two sets of purified IgG from rabbit antisera against purified house fly P450 reductase. One clone that was positive with both antibodies was found (HFR1). Partial sequence analysis of the Y-end of this 0.8 kb clone revealed that it was an authentic P450 reductase clone, as judged by the high degree of similarity between its open reading frame and the rat P450 reductase sequence (Porter and Kasper, 1985). Nick-translated DNA insert from clone HFR1 was then used to rescreen the eDNA library. Six additional clones were obtained. The clone with the longest insert (HFR621) was found to overlap

HOUSE FLY P450 REDUCTASE 441
with clone HFR1. The HFR621 insert was rescued into pBluescript plasmids, and used for sequencing. The insert was also cut with the restriction enzymes ApaI, BamHI and XbaI and the fragments obtained were subcloned into pBluescript plasmids and sequenced in both directions. The clone HFR621 cDNA was 1841 nucleotides long and had an open reading frame fol- lowed by a T A G stop codon and a 3'-untranslated region of 266 nucleotides. A putative polyadenylylation signal A A T A A A was located 21 nucleotides upstream of the 3' end of the clone (Fig. 1). However its 5' end did not include the initiation codon ATG. In order to obtain the 5' end of the P450 reductase sequence, a genomic library was screened with the 5' E c o R I - X b a I fragment of clone HFR621 as probe.
A genomic clone (G6) of 16 kb was obtained and its D N A was digested with restriction enzymes and ana- lyzed by Southern blotting. An Xba I fragment of 5.2 kb was found to hybridize to the cDNA and to an oligonu- cleotide representing the 5' end of the c D N A clone. This XbaI fragment (G6-Xba 5.2) was subcloned into pBlue- script and its restriction map is shown on Fig. 1. The 5' end of the reductase gene was sought by a combination of sequencing of this subclone and m R N A sequencing with five different oligonucleotides selected to confirm putative exon-intron junctions. The position of introns 2 and 3 and the 3' end of intron 1 were thus confirmed by alignment of the RNA sequence and genomic se- quence, and these introns conformed to the G T - A G rule (Fig. 2). The A T G translation initiation site was located in exon 2 (166 bp in length). The sequence flanking the A T G was in excellent agreement with the consensus for Drosophila and other invertebrate translation initiation codons compiled by Cavener and Ray (1991). Intron 1 was more than 1500 bp long; the junction to the untrans- lated exon 1 was not found on clone G6-Xba 5.2 or on the 5'-adjacent subclone from the parent G6 clone. We cannot exclude the possibility that one or more introns interrupt exon 1 but we assume here that it is not, for simplicity of numbering. This is also the case for the rat
P450 reductase gene for which exon 1 has not been found (Porter et al., 1990). The extent of R N A sequence seen on the gels indicated that exon 1 would be at least 60 bp long. Exon 3 and 4 are 193 and 150bp in length, respectively. Exon 4 contains an EcoRI site which corresponds to the 5' end of the cDNA. It is clear therefore that the protection of the EcoRI sites by methylation of the cDNA prior to cloning in 2 ZAP must have been unsuccessful as noted also by Fournier et aL (1992). The 2013 bp open reading frame (Fig. 2) thus obtained by a combination of cDNA, m R N A and genomic sequencing allowed us to deduce the amino acid sequence of a protein of 671 residues (Fig. 2) with a molecular weight of 76,366.
Northern blot analysis and developmental expression
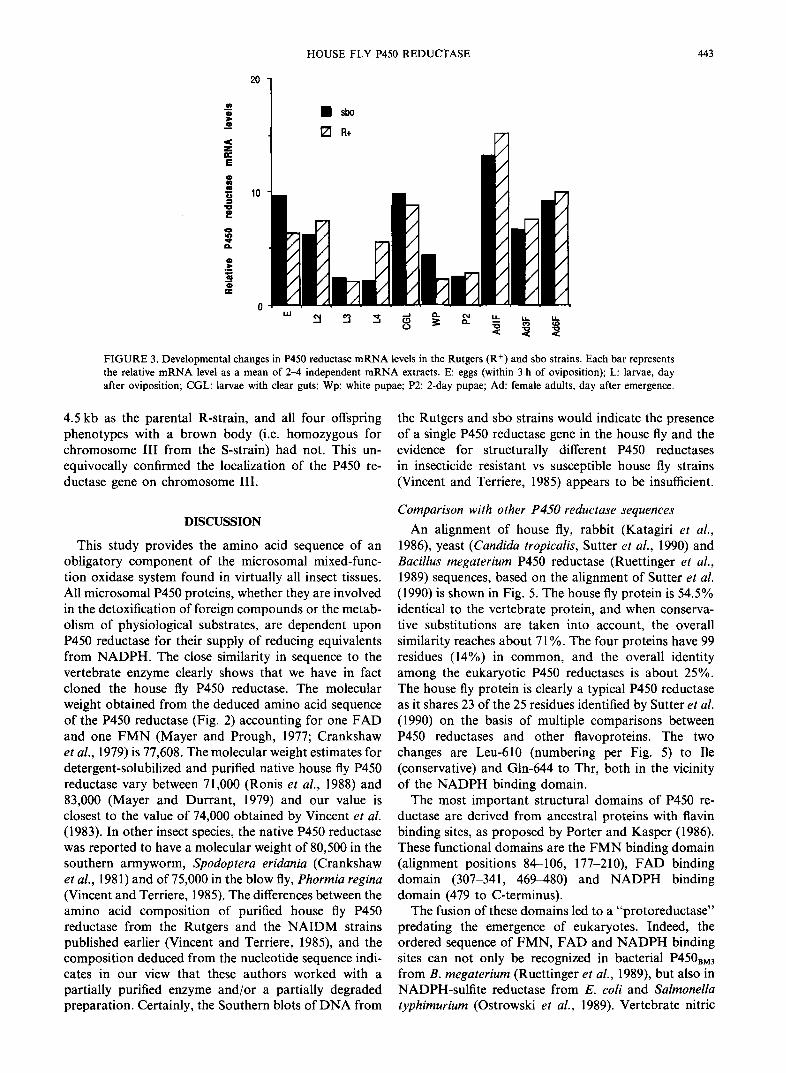
Northern blot analysis of house fly adult and larval m R N A indicated that the nick-translated HFR621 probe hybridized to a m R N A species of 3.15 kb in length (not shown). Dot blots of serially diluted m R N A from different stages showed that the P450 reductase gene was expressed throughout development, with high levels before metamorphosis (in clear gut larvae) and low levels in pupae. The significance of the relatively high levels of expression in eggs needs to be analyzed at the level of the protein and its activity. No significant differences in the levels of P450 reductase m R N A were seen between the Rutgers and sbo strains (Fig. 3).
Chromosomal localization
We analyzed the D N A from the parental R- and S-strains. Restriction fragment length polymorphisms (RFLPs) were sought with the nick-translated insert from clone HFR621 as a probe. Southern analysis showed only one, two or three bands for each of the restriction enzymes with six-base palindromic sequences (BamHI, ClaI, EcoRI, HindIII , KpnI , PstI, SalI, SmaI), suggesting the existence of a single gene c. 19-23 kb long. Clear differences in the restriction fragment pattern between the R- and S-strain were detected with
exon2 exon3 exon4 XC E E H HE X L..J liT ~ ,. ir ~ . - ]
ATG
IO0--bp G6 - Xba 5.2
I II TAG A ATA A A
HFR 621
FIGURE 1. Map of the genomic and cDNA clones for NADPH cytochrome P450 reductase. The top line shows the map of the 5.2 kb XbaI subclone of the genomic clone G6 with boxes representing exons. The three interrupted sequences are 0.4, 1.0 and 1.0 kb long respectively. Only the middle portion of intron 3 (about 1 kb) was not sequenced. The bottom line shows the map of the cDNA clone HFR 621. The EcoRI site in exon 4 represents the 5' end of the cDNA sequence (see also Fig. 2). The abbreviations used for the restriction endonuclease sites are X: XbaI; C: ClaI; E: EcoRI; H: HindlII; B: BamHI;
A: ApaI. The scale is identical for the two maps.

442 J.F. KOENER et al.
gaattctaaaatttaaaagttctttttcagATTTATATAAAATGAGCGCGGAACACGTCGAAGAAGTGGTTTCGGAAGAACCATTTTTGGGC ...... intron 1"...>1,5 kb .... ] MetSerAlaGluHisValGluGluValValSerGluGluProPheLeuGly 17
ACACTGGATATTGCATTATTAGTAGTCCTCTTAGTCGGTGCAACATGGTACTTTATGAGAAGTCGTAAGAAGGAAGAAGCTCCAATTCGTTCA ThrLeuAspI•eA•aLeuLeuva•va•LeuLeuVa•G•yA•aThrTrpTyrPheMetArgserArgLysLysG•uG•uA•aPr•I•eArgSer 48
TATT•TATAcAgtaagtttgttgcatatcctcaagtacaagctgattttctaaattttattctcagA•CCACTACAGTCAGCACCGTTTCAAcG
TyrSerIleGl[ ................... intron 2 ...55bp .................. ]nProThrThrValSerThrValSerThr 61
AcAGAGAATTCGTTCATAAAAAAATTGAAAGCTTCAGGTCGTAGTTTAGTTGTGTTTTATGGTTCCCAGACCGGAACAGCCGAAGAATTTGcC ThrG•uAsnSerPheI•eLysLysLeuLysA•aSerG•yArgSerLeuva•Va•PheTyrG•ySerG•nThrG•yThrA•aG•uG•uPheA•a 92
GGACGATTGGCCAAGGAAGGTTTACGCTATCGCATGAAGGGAATGGTGGCGGACCCCGAAGAATGTGACATGgtaagtatactatccctattt GlyArgLeuAlaLysGluGlyLeuArgTyrArgMetLysGlyMetValAlaAspProGluGluCysAspMet [ ........ intron 3 ... 116
...atattttttattactttttagGAAGAATTGTTACAAATGAAGGATATTCCTAAcTCATTAGccGTTTTCTGTTTGG•TACATATGGTGAA
...1,430 bp ............ ]GluGluLeuLeuGlnMetLysAspIleProAsnSerLeuAlaValPheCysLeuAlaThrTyrGlyGlu 139
GGAGATC•CA•TGATAACGCTATGGAATTCTATGAATGGATTA•AAATGG•GAGGTTGATTTGACGGGTTTAAACTAcGc•gtaagtaaaata G•yAspPr•ThrAspA$nA•aMetG•uPheTyrG•uTrpI•eThrAsnG•yG•uva•AspLeuThrG•yLeuAsnTyrA•a[...intr•n 4 166
>cDNA sequence
......... GTATTTGGcTTAGGCAACAAGAcTTATGAACATTACAAcAAAGTTGCAATTTATGTGGACAAGAGGTTGGAGGAACTCGGTGCT
.>2,8 kb]ValPheG•yLeuG•yAsnLysThrTyrG•uHisTyrAsnLysValA•aI•eTyrva•AspLysArgLeuG•uG•uLeuG•yA•a 194
ACGAGAGTGTTcGAGTTAGGTcTTGGAGAcGATGATGcAAATATCGAAGATGACTTCATTAcGTGGAAAGACAGATTTTGGCCATCTGTTTGT ThrArgVa•PheG•uLeuG•yLeuG•yAspAspAs•A•aAsnI•eG•uAspAspPheI•eThrTrpLysAspArgPheTrpPr•SerVa•cys 225
GATTTcTTCGGAATCGAAGGAAGTGGTGAGGAGGTATTGATGCGTCAATTCCGACTTTTGGAAcAGCcGGATGTTCAACcCGATCGTATTTAC AspPhePheG•yI•eG•uG•ySerG•yG•uG•uvalLeuMetArgG•nPheArgLeuLeuG•uG•nPr•AspvalG•nPr•AspArgI•eTyr 256
ACCGGCGAAATAGCTcGGTTGCACTcAATGCAAAATcAAAGAcCTCcATTTGATGCTAAAAATcCATTTTTGGcTTCAGTCATAGTTAATCGA ThrG•yG•uI•eA•aArgLeuHisSerMetGlnAsnG•nArgPr•Pr•PheAspA•aLysAsnPr•PheLeuA•aSerVa•I•eVa•AsnArg 287
GAGCTGCATAAAGGcGGCGGACGGTCATGTATGcATATTGAGTTGGATATTGATGGATCAAAAATGcGGTATGATGcAGGCGACCATATAGcA G•uLeuHisLy•G•yG•yG•yArgSercysMetHisI•eG•uLeuAs•I•eAspG•ySerLysMetArgTyrAspA•aG•yAspHisI•eA•a 318
ATGTATCCGATTAATGATAAAATCCTGGTAGAAAAATTGGGTAAAcTATGTGATGCAAATCTAGATACAGTcTT•TcTTTGATCAATACGGAT MetTyrPr•I•eAsnAspLysI•eLeuva•G•uLysLeuG•yLysLeuCysAspA•aAsnLeuAspThrVa•PheSerLeuI•eAsnThrAsp 349
ACGGATAGCAGTAAAAAACATCCATTCCCCTGTCCCACCACATACcGTACCGCcCTGACACATTACTTGGAGATTACTGCAATACCGAGAACA ThrAspSerSerLysLysHisPr•PhePr•CysPr•ThrThrTyrArgThrA•aLeuThrHisTyrLeuG•uI•eThrA•aI•ePr•ArgThr 380
CATATTCTGAAGGAGTTAGCAGAGTATTGTTCCGATGAAAAGGATAAGGAATTTTTACGCAACATGGcGTCTATTACTcCGGAAGGCAAAGAG HisI•eLeuLysG•uLeuAlaG•uTyrCysSerA•pG•uLysAspLysGluPheLeuArgAsnMetA•aSerI•eThrPr•G•uG•yLysG•u 411
AAGTATcAGAATTGGATCCAGAACTcTTCAAGAAATATTGTGCATATTTTGGAGGATATCAAATCTTGTAGACCACCAATTGATcATATTTGT LysTyrG•nAsnTrpI•eG•nAsnSerSerArgAsnI•eva•HisI•eLeuG•uAspI•eLysSercysArgPr•Pr•I•eAspHisI•eCys 442
GAACTGTTGCCAAGACTTCAA•cGCGTTATTACTCGATTTCATcTTCTTCTAAGCTCTATcCAACTAATGTTCATATTACAGCAGTTCTGGTT G•uLeuLeuPr•ArgLeuG•nPr•ArgTyrTyrSerI•eserSerSerserLysLeuTyrPr•ThrAsnva•HisI•eThrA•ava•LeuVa• 473
CAATATGAAA•A•CTA•TGGCCGTGTAAACAAAGGTGTTGCTA•GTCATA•ATGAAGGAAAAAAATCCTTCGGTTGGAGAGGTTAAAGTG•CT GlnTyrGluThrPr•ThrGlyArgVa•AsnLysG•y•alAlaThrSerTyrMetLysGluLysAsnPr•SerValGlyGluVa•LysVa•Pr• 504
GTATTTATTCGCAAATCTCAATTTAGGTTGCCAA•TAAATCCGAGATTCCAAT•ATAATGGTGGG•CCAGGAACGGGTCTAG•ACCTTTCAGA valPheIleArgLysSerGlnPheArgLeuPr•ThrLys•erGluIlePr•IleIleMetValGlyPr•GlyThrGlyLeuAlaPr•PheArg 535
GGATTCATACAGGAAAGA•AATTTTTA•GAGATGGAGGAAAAGTTGTCGGCGATACCATATTGTATTTCGGGTGTAGAAAGAAGGA•GAAGAC GlyPheIleGlnGluArgG•nPheLeuArgAspG•yGlyLysValValGlyAspThrIleLeuTyrPheG•yCysArgLysLysAspGluAsp 566
TTCATTTATAGAGAAGAACTAGAGCAATATGTACAAAATGGAACTTTAACCTTGAAAACGGCATTTT•AAGAGACCAACAAGAAAAAATTTAT PheIleTyrA•gGluGluLeuGluGlnTyrValGlnAsnGlyThrLeuThrLeuLysThrAlaPheSerArgAspGlnG•nG•uLysIleTyr 597
GTTA•T•AT•TAATCGAACAAGATG•TGATTTAATTTGGAAAGTAATTGGCGAA•AAAAGGGA•A•TTTTATATTTGCGGGGATGCTAAAAAT •alThrHisLeuIleGluGlnAspAlaAspLeuIleTrpLysValIleGlyGluG•nLysGlyHisPheTyrIleCysGlyAspAlaLysAsn 628
ATGGCTGTGGATGTTAGAAATATTTTAGTTAAAATTTTGT•AA•CAAAGGCAATATGAACGAATCAGATGCTGTGCAATATATAAAAAAGATG MetA•aVa•AspVa•ArgAsnI•eLeuValLysI•eLeuSerThrLysG•yAsnMetAsnG•uSerAspA•aVa•GlnTyrI•eL•sLysMet 659
GAGGCAcAAAAGCGTTATTcCGCGGACGTTTGGAGCTAGAAAAAAACGTCATTTGTACAACACTAGTAAAACCAATAATTCCTTCGTGTCTTGT GluAlaGlnLysArgTyrSerAlaAspValTrpSerTer 671
TTCCTAAAATAAGAAAATTTGCAATTCCATGCAAAAACAAGATTATTGATATTTTTTATAATATTTTTGTACAGAAAAcAAAGAAGT•TTCCGG TTTGGGAATATTTTCCACGTGcTTACTAATGTTTTATTGTATGCATTTTTAcccATCAccACTTTTGTcAGAATAATTTATAGAcAAATAAAAA AGTATCTTTCGAATCAAT
FIGURE 2. Nucleotide (top) and deduced amino acid sequence (bottom) of NADPH cytochrome P450 reductase. The asterisk attached to the designation of the first intron indicates that the numbering of introns is tentative in the absence of sequence information on the first exon(s). The sequence of the cDNA starts at the underlined EcoRI site. The 3' end of intron 4 was not found on genomic clone G6. The oligonucleotides used for mRNA sequencing are underlined. The putative polyadeny-
lylation site is also underlined. Numbering on the right indicates the amino acid residue position.
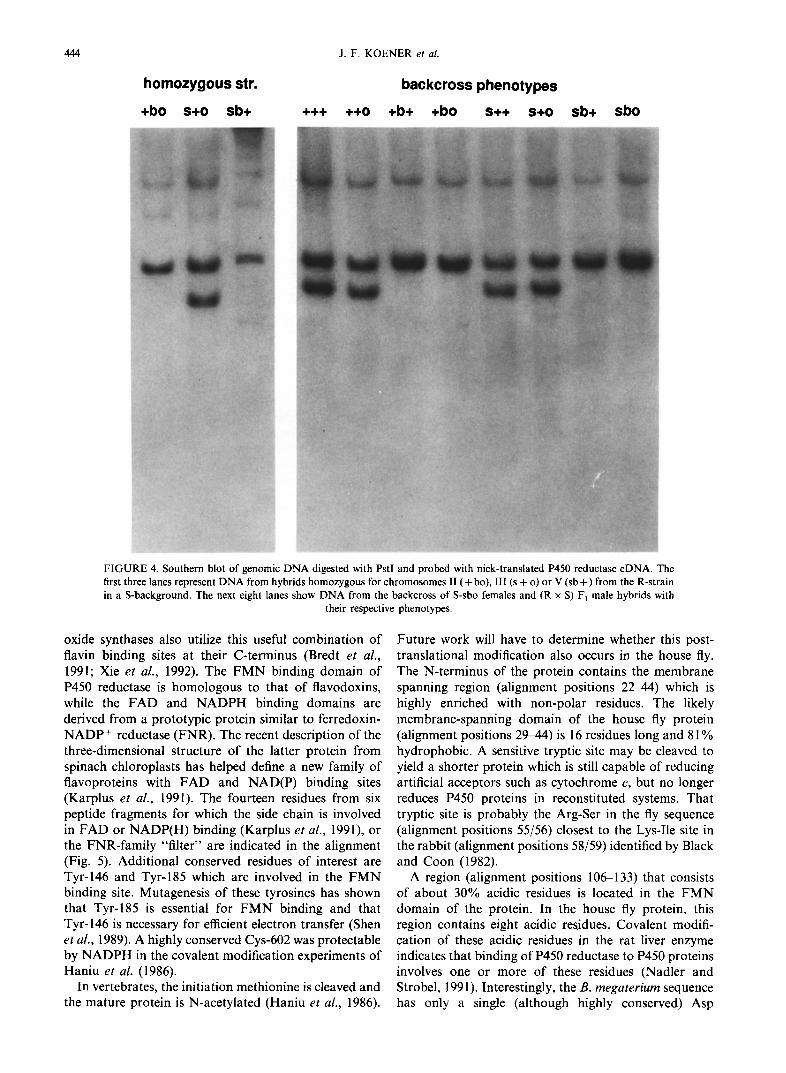
the enzymes ClaI and PstI. These R F L P s were then used to determine the local izat ion o f the P450 reductase gene by analyzing the D N A o f hybrid S-flies h o m o z y g o u s for the II ( + bo), III (s + o) or V (sb + ) c h r o m o s o m e from the R-strain in an S background. Southern blot hybrid- ization analysis l inked c h r o m o s o m e III from the R-
strain to the restriction fragment pattern o f the R-strain. Analysis o f the R F L P s o f backcrosses between (R × S) F1 male hybrids and S-sbo females, in which all four offspring phenotypes lacking the brown body marker (i.e. with one copy o f c h r o m o s o m e III from the R-strain) had a PstI band o f 4.2 kb (Fig. 4) and a ClaI band o f
HOUSE FLY P450 REDUCTASE 443
o
,< Z h- E
1
Q Iv-
20
10
r ' ~ o .
FIGURE 3. Developmental changes in P450 reductase mRNA levels in the Rutgers (R +) and sbo strains. Each bar represents the relative mRNA level as a mean of 2-4 independent mRNA extracts. E: eggs (within 3 h of oviposition); L: larvae, day after oviposition; CGL: larvae with clear guts; Wp: white pupae; P2: 2-day pupae; Ad: female adults, day after emergence.
4.5 kb as the parental R-strain, and all four offspring phenotypes with a brown body (i.e. homozygous for chromosome III from the S-strain) had not. This un- equivocally confirmed the localization of the P450 re- ductase gene on chromosome III.
DISCUSSION
This study provides the amino acid sequence of an obligatory component of the microsomal mixed-func- tion oxidase system found in virtually all insect tissues. All microsomal P450 proteins, whether they are involved in the detoxification of foreign compounds or the metab- olism of physiological substrates, are dependent upon P450 reductase for their supply of reducing equivalents from NADPH. The close similarity in sequence to the vertebrate enzyme clearly shows that we have in fact cloned the house fly P450 reductase. The molecular weight obtained from the deduced amino acid sequence of the P450 reductase (Fig. 2) accounting for one FAD and one FMN (Mayer and Prough, 1977; Crankshaw et al., 1979) is 77,608. The molecular weight estimates for detergent-solubilized and purified native house fly P450 reductase vary between 71,000 (Ronis et al., 1988) and 83,000 (Mayer and Durrant, 1979) and our value is closest to the value of 74,000 obtained by Vincent et al. (1983). In other insect species, the native P450 reductase was reported to have a molecular weight of 80,500 in the southern armyworm, Spodoptera eridania (Crankshaw et al., 1981) and of 75,000 in the blow fly, Phormia regina (Vincent and Terriere, 1985). The differences between the amino acid composition of purified house fly P450 reductase from the Rutgers and the NAIDM strains published earlier (Vincent and Terriere, 1985), and the composition deduced from the nucleotide sequence indi- cates in our view that these authors worked with a partially purified enzyme and/or a partially degraded preparation. Certainly, the Southern blots of DNA from
the Rutgers and sbo strains would indicate the presence of a single P450 reductase gene in the house fly and the evidence for structurally different P450 reductases in insecticide resistant vs susceptible house fly strains (Vincent and Terriere, 1985) appears to be insufficient.
Comparison with other P450 reductase sequences
An alignment of house fly, rabbit (Katagiri et al., 1986), yeast (Candida tropicalis, Sutter et al., 1990) and Bacillus megaterium P450 reductase (Ruettinger et al., 1989) sequences, based on the alignment of Sutter et al. (1990) is shown in Fig. 5. The house fly protein is 54.5% identical to the vertebrate protein, and when conserva- tive substitutions are taken into account, the overall similarity reaches about 71%. The four proteins have 99 residues (14%) in common, and the overall identity among the eukaryotic P450 reductases is about 25%. The house fly protein is clearly a typical P450 reductase as it shares 23 of the 25 residues identified by Sutter et al. (1990) on the basis of multiple comparisons between P450 reductases and other flavoproteins. The two changes are Leu-610 (numbering per Fig. 5) to Ile (conservative) and Gin-644 to Thr, both in the vicinity of the NADPH binding domain.
The most important structural domains of P450 re- ductase are derived from ancestral proteins with flavin binding sites, as proposed by Porter and Kasper (1986). These functional domains are the FMN binding domain (alignment positions 84-106, 177-210), FAD binding domain (307-341, 469-480) and NADPH binding domain (479 to C-terminus).
The fusion of these domains led to a "protoreductase" predating the emergence of eukaryotes. Indeed, the ordered sequence of FMN, FAD and NADPH binding sites can not only be recognized in bacterial P450Bm3 from B. megaterium (Ruettinger et al., 1989), but also in NADPH-sulfite reductase from E. coli and Salmonella typhimurium (Ostrowski et al., 1989). Vertebrate nitric
444 J. F. KOENER et al.
homozygous str. backcross phenotypes
+bo s+o sb+ +++ ++0 +b+ +bo s++ s+o sb+ sbo
FIGURE 4. Southern blot of genomic DNA digested with PstI and probed with nick-translated P450 reductase cDNA. The first three lanes represent DNA from hybrids homozygous for chromosomes II (+ bo), III (s + o) or V (sb +) from the R-strain in a S-background. The next eight lanes show DNA from the backcross of S-sbo females and (R x S) F~ male hybrids with
their respective phenotypes.
oxide synthases also utilize this useful combination of flavin binding sites at their C-terminus (Bredt et al., 1991; Xie et al., 1992). The FMN binding domain of P450 reductase is homologous to that of flavodoxins, while the FAD and NADPH binding domains are derived from a prototypic protein similar to ferredoxin- NADP + reductase (FNR). The recent description of the three-dimensional structure of the latter protein from spinach chloroplasts has helped define a new family of flavoproteins with FAD and NAD(P) binding sites (Karplus et al., 1991). The fourteen residues from six peptide fragments for which the side chain is involved in FAD or NADP(H) binding (Karplus et al., 1991), or the FNR-family "filter" are indicated in the alignment (Fig. 5). Additional conserved residues of interest are Tyr-146 and Tyr-185 which are involved in the FMN binding site. Mutagenesis of these tyrosines has shown that Tyr-185 is essential for FMN binding and that Tyr-146 is necessary for efficient electron transfer (Shen et al., 1989). A highly conserved Cys-602 was protectable by NADPH in the covalent modification experiments of Haniu et al. (1986).
In vertebrates, the initiation methionine is cleaved and the mature protein is N-acetylated (Haniu et al., 1986).
Future work will have to determine whether this post- translational modification also occurs in the house fly. The N-terminus of the protein contains the membrane spanning region (alignment positions 22-44) which is highly enriched with non-polar residues. The likely membrane-spanning domain of the house fly protein (alignment positions 29-44) is 16 residues long and 81% hydrophobic. A sensitive tryptic site may be cleaved to yield a shorter protein which is still capable of reducing artificial acceptors such as cytochrome c, but no longer reduces P450 proteins in reconstituted systems. That tryptic site is probably the Arg-Ser in the fly sequence (alignment positions 55/56) closest to the Lys-Ile site in the rabbit (alignment positions 58/59) identified by Black and Coon (1982).
A region (alignment positions 106-133) that consists of about 30% acidic residues is located in the FMN domain of the protein. In the house fly protein, this region contains eight acidic residues. Covalent modifi- cation of these acidic residues in the rat liver enzyme indicates that binding of P450 reductase to P450 proteins involves one or more of these residues (Nadler and Strobel, 1991). Interestingly, the B. megater ium sequence has only a single (although highly conserved) Asp

HOUSE FLY P450 REDUCTASE 445
1 60 fly ....... msaehveevvseepflgtlDialLvvLiVgatwY.Fmrsrkkeeapirsysiq rab madshgdtgatmpeaaaqeasvfsmtDv.vLfsLiVglitYwFlfrkkkeevpeft.kiq yea .................... maldklDlyvIitLvVaiaaY.Faknqfld...qqqdtgf BM3 ............................................................
61 120 fly ptTvStvsttensfikklKasgrslvVfYGSQTGT~efAgrLakegl.RyrnKGnv/~p rab apTsSsv..kessfvekmKktgrnivVfYGSQTGTAEefAnrLskdah.RygmrGmaADp yea InTdSgdg.nsrdilqalKknnkntlllfGSQTGTAEdyAnkLsrelhsRfglKtmvADf BM3 .............. kkaenahntpllVlYGSnmGTAEgtArdLadiams .... KGfapqv
** **** . .
121 180 fly eecDmeellqmkdipnsLAVFclATYGEGdPTDNAmeFyeWitng.evdLtGlnYaVFGL rab eeyDladlsslpeinnaLAVFcmATYGEGdPTDNAqdFydWLqet.dvdLsGvkYaVFGL yea adyDfenfgdite..diLvfFivATYGEGePTDNAdeFhtWLtee.adtLstlkYtVFGL BM3 atlD .... shagnlpregAVlivtasynGhPpDNAkqFvdWLdqasadevkGvrYsVFGc
, , , *** , • , ***
181 240 fly GNKTYE.hyNkvaiyvDkrLeelGAtRvfEiGiGDDd.aniEdDFitWkdrfWpsVcdfF tab GNKTYE.hfNamgkyvDqrLeqlGAqRifEIGmGDDd.anlEeDFitWreqfWpaVcehF yea GNsTYE.ffNaigrkfDrlLgekGgdRfaEyGeGDDgtgtldeDFlaWkdnvfdslkndl BM3 GdKnwattyqkvpafiDetLaakGAeniadrGeaDas.ddfEgtyeeWrehmWsdVaayF
, ** , ** ,
241 300 fly giEgsgEEvlmrqfrLleqpDv..qpdriytGEia..rlhs..mqnqrpPFDaknPFLAs tab gvEatgEEssirqyeLvlhtDi..dvakvyqGEmg..rlks..yenqkpPFDaknPFLAt yea nfEek.ElkyepnvkLterdDlsgndpdvslGEpnvkyiksegvdltkgPFDhthPFLAr BM3 nldiensEdnkstlsLqfvdsaadmplakmhG ....................... aFstn
301 360 fly VivnrELhk. GggRscn~'IiE1DIdgSkmrYdaGDHi~'nyPiNdkilVekigklcd..anL rab VttnrkLnq.GteRhlmHiEiDIsdSkirYesGDHvAvyPaNdsalVnqlgeilG..adL yea ivktkELft.sedRhcvHvEfDIseSnlkYttGDHiAiwPsNsdenikqfakcfGledkL BM3 VvaskELqqpGsaRstrHiEielp.keasYqeGDHlgviPrNyegiVnrvtarfG .... L
. . , . . *** . , .
361 420 fly DtVfsLintDtdSskkHPFPcPTtYrtaLthYLEItaip~thiLkelAeycsDekdNefL rab DvVmsLnnlDeeSnkkHPFPcPTsYrtaLtyYLdItnPprtnvLyelAqyaaDpaeqeqL yea DtVieLkalD..StysiPFPnPitYgavirhhLEIsgPvsrqfflsiAgfapDeetKksf BM3 DasqqirleaeeeklaHlplakTvsveeLlqYvElqdPvtrtqLramAaktvcpphKveL
421 . o oo 480
rnmasitpEgKekYqnwiqnssrnivhiLediksclpPidhicELLPrLqPRYYSISSSS rkmasssgEgKelYlswvvearrhiLaiLqdyPslrpPidhlcELLPrLqaRYYSIaSSS triggdkqEfaskvtrrkfniadalLfasnnrPwsdvPfefliEnvqhLtPRYYSISSSS ealle ..... EqaYkeqvlakrltmLelLekyPacemkfsefiaLLPsirPRYYSISSSp
***** **
481 o o . 540
klyptnvhITAVlVqyEtptGRvn.KGVATsymkeknps[..vGEv . . . . . . . . . . . . . . kvhpnsvhIcAVaVeyEtkaGRln.KGVATswLrakep .... aGEnggra .......... isekqtinvTAVveaeEeadGRpv.tGVvTnlLknieieqnktGEtpmvhydlngprgkf rvdekqasITvsvVsgEawsGygeyKGiAsnyLaelqegdtitcf ...............
541 oo . . 600 ...kvPvfiRkSqFrLPtkseiPiIMVGPGTGIAPFRGFiQERqfLrdgGkvVGdTiLYf ...IvPmfvRkSqFrLPfkatTPvIMVGPGTGVAPFiGFiQERawLrqqGkeVGeTILYy skfrlPvhvRrSnFkLPknstTPvIliGPGTGVAPiRGFvrERvqqvknGvnVGkTvLfy ...... istpqSeFtLPkdpeTPIIMVGPGTGVAPFRGFvQaRkqLkeqGqslGeahLYf
. . ** . • ***** ** ** . . • .
601 . oo o o o . 660 GCRkkdEDfiYrEELeqyvqn.Gtlt.LktAFSRdqqe.KiYVtHlieqDadliwkvige GCRraaEDyLYrEELagfqkd.GtlsqLnvAFSReqaq.KvYVQHllrrDkehlwrLihe GCRnseqDfLYkqEwseyasvlGenfemfnAFSRqdptkKvYVQdkilensalvdeLl.s GCRsphEDyLYqEELenaqse.Giit.LhtAFSRmpnqpKtYVQHvmeqDgkklieLl.d *** , • . . **** . **
661 o . . . o 717 qkgHfYiCGDAknMAvDVrnilvKIlstkgnmnEsdAvqyiKkmeaqkRYsaDVWs. gGAHiYvCGDArnMArDVqntfydIvAelgamehaqAvdyvKklmtkgRYslDVWs. sGAiiYvCGDAsrMArDVqaaiaKIvAksrdihEdkAaelvKswkvqnRYqeDVW.. qGAHfYiCGDgsqMApaVeatlmKsyAdvhqvsEadArlwlqqleekgRYakDVWag
. *** ** • , ** ***
fly rab
yea BM3
fly rab yea BM3
fly tab yea BM3
fly rab
yea BM3
fly rab
yea BM3
FIGURE 5. Alignment of house fly, rabbit, yeast and B. megaterium NADPH cytochrome P450 reductases. Residues in common in at least three sequences are indicated in upper case letters. Residues in common in all four sequences are marked with an asterisk, fly: M. domestica (this work); rab: rabbit (Katagiri et al., 1986); yea: C. tropicalis (Sutter et al., 1990); BM3: B. rnegaterium P450 BM3 (Ruettinger et al., 1989). The conserved residues which constitute the core of the FNR-family filter
(Karplus et al., 1991) are marked by "O" on top of the alignment.
residue in this region as well as two gaps comprising eight residues. The interaction between the reductase domain and P450 domain of that fusion protein may therefore prove to be quite different from the electro- static interaction between basic residues on P450 pro- teins (Shen and Strobel, 1992) and acidic residues of P450 reductase in microsomal systems. Another acidic region (alignment positions 148-157) was postulated by Porter and Kasper (1986) to interact with P450 proteins,
based on the homology with flavodoxins and their interaction with c-type cytochromes. A third region of six clustered acidic residues in the rat enzyme (alignment positions 215-224) was shown by Nisimoto (1986) to cross-link with Lys-13 of cytochrome c.
The high overall identity between P450 reductase proteins may be caused by the evolutionary constraints of a protein with multiple interacting domains. In addition to the FMN, FAD and N A D P H binding

446 J .F . KOENER et al.
domains interacting with their ligand, the protein also needs to be anchored to the membrane and to interact with a stoichiometric excess of diverse P450 proteins (Feyereisen et al., 1990). This high similarity explains why the insect P450 reductase can reduce mammalian P450 (CYP2B 1) in transgenic Drosophila (Jowett et al., 1991).
Although the gene structure of the house fly P450 reductase is not known in its entirety, some comparisons can be made with the gene structure of the rat P450 reductase. The rat gene spans at least 20 kb of D N A and at least 16 exons, 15 of which have been characterized (Porter et al., 1990). Based on the alignment of ver- tebrate and house fly enzymes, it appears that exon 2 starts 13 and 11 bp upstream from the initiation codon in the rat and house fly respectively. House fly exons 2 and 3 are separated by a 55 bp intron which is not found in the rat gene, and conversely, introns 2 and 3 of the rat gene are not found in the house fly gene. However, house fly exons 3 and 4 are interrupted by a 1430 bp intron which is located exactly at the position of intron 4 (710 bp) in the rat gene. The 3' end of exon 4 in the house fly also corresponds exactly to the 3' end of exon 5 in the rat. The similarity between vertebrate and insect en- zymes is therefore not limited to their amino acid sequence, but extends to the organization of their genes, and a full characterization of the house fly gene would be of interest.
The localization of the P450 reductase gene on chromosome III in the house fly can be discussed in the light of Scott and Georghiou's data (1986) on resistance to permethrin in the Learn-PyR strain. This strain is highly resistant to pyrethroids as a result of multiple resistance mechanisms including elevated P450- dependent metabolism. An elevated level of P450 re- ductase activity in this strain (when compared to two susceptible strains) was mapped to chromosomes III and V. A single copy of chromosome III from the Learn-PyR strain contributed about half the difference in activity observed between the (homozygous) resistant and sus- ceptible strains. This would suggest that a cis-acting element on chromosome III from Learn-PyR contributes to the elevated P450 reductase activity. Moreover, chromosome V also contributes somewhat to the elev- ated P450 reductase activity, suggesting a complex regu- latory mechanism, not unlike that of P450 enzymes themselves. The availability of a molecular probe for the P450 reductase gene should allow a dissection of the genetic control of this gene and its role in insecticide resistance.
REFERENCES
Baldridge G. D. and Feyereisen R. (1986) Blood meal and cytochrome P-450 monooxygenases in the northern house mosquito, Culex pipiens. Pest. Biochem Physiol. 25, 407-413.
Black S. D. and Coon M. J. (1982) Structural features of liver microsomal NADPH-cytoehrome P-450 reductase. J. biol. Chem. 257, 5929-5938.
Bredt D. S., Hwang P. M., Glatt C. E., Lownstein C., Reed R. R. and Snyder S. H. (1991) Cloned and expressed nitric oxide synthase structurally resembles cytochrome P-450 reductase. Nature 351, 714-718.
Carifio F., Koener J. F., Plapp F. W. Jr and Feyereisen R. (1992) Expression of the cytochrome P450 gene CYP6AI in the house fly, Musca domestiea. ACS Syrup. Ser. 505, 31-40.
Cavener D. R. and Ray S. C. (1991) Eukaryotic start and stop translation sites. Nucl. acids Res. 19, 3185-3192.
Crankshaw D. L., Hetnarski K. and Wilkinson C. F. (1979) Purifi- cation and characterization of NADPH-cytochrome c reductase from the midgut of the southern armyworm (Spodoptera eridania). Biochem. J. 181, 593~05.
Crankshaw D. L., Hetnarski K. and Wilkinson C. F. (1981) The functional role of NADPH-cytochrome c reductase in southern armyworm (Spodoptera eridania) midgut microsomes. Insect Bio- chem. 11, 515-522.
Davis L. G., Dibner M. D. and Battey J. F. (1986) Basic Methods in Molecular Biology, p. 388. Elsevier, New York.
Feyereisen R. and Vincent D. R. (1984) Characterization of antibodies to house fly NADPH-Cytochrome P-450 reductase. Insect Biochem. 14, 163-168.
Feyereisen R., Koener J. F., Carifio F. A. and Daggett A. S. (1990) Biochemistry and molecular biology of insect cytochrome P450. Molecular Insect Science (Edited by Hagedorn H. H., Hildebrand J. G., Kidwell M. G. and Law J. H.), pp. 263 272. Plenum Press, New York.
Feyereisen R., Koener J. F., Farnsworth D. E. and Nebert D. W. (1989) Isolation and sequence of cDNA encoding a cytochrome P-450 from an insecticide-resistant strain of the house fly, Musca domestica. Proc. natn. Acad. Sci. U.S.A. 86, 1465-1469.
Fournier D., Bride J. M., Poirie M., Berg~ J. B. and Plapp F. W. Jr (1992) Insect glutathione S-transferases. J. biol. Chem. 267, 1840-1845.
Geliebter J. (1987) Dideoxynucleotide sequencing of RNA and un- cloned cDNA. Focus 9, 5-8.
Halliday W. R., Farnsworth D. E. and Feyereisen R. (1986) Hemo- lymph ecdysteroid titer and midgut ecdysone 20-monooxygenase activity during the last larval stage of Diploptera punctata. Insect Biochem. 16, 627~i34.
Haniu M., Iyanagi T., Miller P., Lee T. D. and Sbively J. E. (1986) Complete amino acid sequence of NADPH-cytochrome P-450 reductase from porcine hepatic microsomes. Biochemistry 25, 7906-7911.
Jowett T., Wajidi M. F. F., Oxtoby E. and Wolf C. R. (1991) Mammalian genes expressed in Drosophila: a transgenic model for the study of mechanisms of chemical mutagenesis and metabolism. EMBO J. 10, 1075 1081.
Karplus P. A., Daniels M. J. and Herriott J. R. (1991) Atomic structure of ferredoxin-NADP+ reductase: prototype for a struc- turally novel flavoenzyme family. Science 251, 60456,
Katagiri M., Murakami H., Yabusaki Y., Sugiyama T., Okamoto M., Yamano T. and Ohkawa H. (1986) Molecular cloning and sequence analysis of full-length cDNA for rabbit liver NADPH-cytochrome P-450 reductase mRNA. J. Biochem. 100, 945-954.
Masters B. S. S. and Okita R. T. (1980) The history, properties, and function of NADPH-cytochrome P-450 reductase. Pharmac. Ther. 9, 227-244.
Mayer R. T. and Durrant J. L. (1979) Preparation of homo- geneous NADPH cytochrome c (P-450) reductase from house flies using affinity chromatography techniques. J. biol. Chem. 254, 756-761.
Mayer R. T. and Prough R. A. (1977) Purification and characterization of NADPH-cytochrome c (P-450) reductase from the house fly, Musca domestica. Comp. Biochem. Physiol. 57B, 81-87.
Nadler S. T. and Strobel H. W. (1991) Identification and characteriz- ation of an NADPH-cytochrome P-450 reductase derived peptide involved in binding to cytochrome P-450. Archs. Biochem. Biophys. 290, 277-284.

HOUSE FLY P450 REDUCTASE 447
Nisimoto Y. (1986) Localization of cytochrome c-binding domain on NADPH-cytochrome P-450 reductase. J. biol. Chem. 261, 14232-14239.
Ostrowski J., Barber M. J., Rueger D. C., Miller B. E., Siegel L. M. and Kredich N. M. (1989) Characterization of the flavoprotein moieties of NADPH-sulfite reductase from Salmonella typhimurium and Escherichia coli. J. biol. Chem. 264, 1579~15808.
Porter T. D. and Kasper C. B. (1985) Coding nucleotide sequence of rat NADPH-cytochrome P-450 oxidoreductase cDNA and identifi- cation of flavin-binding domains. Proc. natn. Acad. Sci. 82, 973-977.
Porter T. D. and Kasper C. B. (1986) NADPH-cytochrome P-450 oxidoreductase: flavin mononucleotide and flavin adenine dinucle- otide domains evolved from different flavoproteins. Biochemistry 25, 1682-1687.
Porter T. D., Beck T. W. and Kasper C. B. (1990) NADPH- cytochrome P-450 oxidoreductase gene organization correlates with structural domains of the protein. Biochemistry 29, 9814-9818.
Queen C. and Korn L. J. (1984) A comprehensive sequence analysis program for the IBM Personal Computer. Nucl. acids Res. 12, 581-599.
Ronis M. J. J., Hodgson E. and Dauterman W. C. (1988) Character- ization of multiple forms of cytochrome P-450 from an insecticide resistant strain of house fly (Musca domestica). Pest. Biochem. Phsiol. 32, 74-90.
Ruettinger R. T., Wen L.-P. and Fulco A. J. (1989) Coding nucleotide, 5' regulatory, and deduced amino acid sequences of P-450~M_ 3, a single peptide cytochrome P-450:NADPH-P-450 reductase from Bacillus megaterium. J. biol. Chem. 264, 10987-10995.
Sanger F., Nicklen S. and Coulson A. R. (1977) DNA sequencing with chain-terminating inhibitors Proc. natn. Acad. Sci, 74, 5463-5467.
Scott J. G. and Georghiou G. P.(1986) The biochemical genetics of
permethrin resistance in the Learn-PyR strain of house fly. Biochem. Genet. 24, 25-37,
Shen S. and Strobel H. W. (1992) The role of cytochrome P450 lysine residues in the interaction between cytochrome P450IA1 and NADPH-cytochrome P450 reductase. Archs Biochem. Biophys. 294, 83-90.
Shen S., Porter T. D., Wilson T. E. and Kasper C. B. (1989) Structural analysis of the FMN binding domain of NADPH-cytochrome P-450 oxidoreductase by site-directed mutagenesis. J. biol. Chem. 264, 7584-7589.
Sutter T. R., Sanglard D. and Loper J. C. (1990) Isolation and Characterization of the alkane-inducible NADPH-cytochrome P-450 oxidoreductase gene from Candida tropicalis. J. biol. Chem. 265, 16428-16436.
Vincent D. R. and Terriere L. C. (1985) Characterization of NADPH- cytochrome P-450 reductase from house flies (Musca domestica, L.) susceptible and resistant to insecticides, and the blow fly (Phormia regina, Meigen). Insect Biochem. 15, 299 307.
Vincent D. R., Moldenke A. F. and Terriere L. C. (1983) NADPH- cytochrome P-450 reductase from the house fly, Musca domestica. Insect Biochem. 13, 559 566.
Xie Q , Cho H. J., Calaycay J., Mumford R. A., Swiderek K. M., Lee T. D., Ding A., Troso T. and Nathan C. (1992) Cloning and characterization of inducible nitric oxide synthase from mouse macrophages. Science 256, 225-228.
Acknowledgements--This work was supported by NIH grant GM39014. We thank A. S. Daggett for technical assistance, and Dr J. F. Andersen for reading the manuscript.





























