Embed Size (px)
Citation preview

Journal of Social Issues, Vol. 41 , No. I , 1985, p p . 127-148
The Changing Genetics/Socialization Balance
Lois Wladis Hoffman University of Michigan
My talk today has to do with a basic psychological issue, but one that inevitably has social implications. I am referring to the old heredity-environ- ment controversy which, like Frankenstein’s monster, often seems to be buried beneath tons of rubble, but rises again for a sequel.
There is occurring in developmental psychology a shift in the extent to which heredity or environment is seen to play the major role in the child’s development. The importance of the family environment, in particular, has been questioned. In several recent journal articles, integrative research reviews, and in one entire issue of Child Development, data have been reported that provide an impressive amount of evidence for the genetic influence on intelligence, personality, and even on the environment itself-that is, the child’s genetic makeup plays a major role in both eliciting the parent response and in selectively processing the environment (Scarr & McCartney, 1983). Taken at face value, the case for environmental effects, except for real deprivation, looks very much weakened.
The social implications of this are considerable, for the new work is not only suggesting the presence of a genetic influence but also the absence of a general family effect. The role of the parents and of early family experience is seen as relatively unimportant. This new research is a blow for those of us who think that there is such a thing as good parenting, that good parenting can be
This presidential address to the Society for the Psychological Study of Social Issues (Division 9) was delivered at the annual meeting of the American Psychological Association in Toronto, Canada. August 1984. The first draft was prepared while I was Scholar in Residence at the Bellagio Study and Conference Center. 1 want to express my appreciation to the Rockefeller Foundation and the staff at the Center for providing this opportunity for uninterrupted work. I was also aided by my colleagues at Temple University, particularly Marsha Weinraub and Nora Newcornbe, who provided helpful comments on the earlier draft, and Susan A n d , who served as my research assistant.
Correspondence regarding this article should be addressed to Lois W. Hoffman, Department of Psychology, University of Michigan, Ann Arbor, MI 48109.
127
~ 2 2 - 4 5 3 7 / 8 5 / 0 3 ~ - 0 1 2 7 ~ 501 I 0 19x5 The Society for the Psychological Study of Social Issues

128 Hoffman
taught, and that while parents do not treat all their children the same, there are general patterns that characterize a parenting style and children are affected by this.
Greater support has emerged for the genetic than for the environmental positior., 1 think, because the bulk of the relevant research is being conducted by behavioral geneticists who, though very sophisticated in their own area, are not familiar with environmental theory or with socialization models.
1 also feel, however, that part of the strength of their position lies in the fact that research on the effects of the family and parenting has not kept pace with other developments in psychology. I think we have become overwhelmed with the complexities of these questions, disillusioned with the oversimplified classi- cal theories, and discouraged by the problems of adequate measurement. Most of us no longer trust parent interviews as the source of data. and behavioral observa- tion techniques are expensive to use, time consuming, and involve so much of the subjects’ cooperation that samples are often small and selective. Work with infants has been more vigorous than with older children, but here there are problems of knowing whether the measures of infant development actually pre- dict to enduring behavior. And as for theory, with a few exceptions, there has been considerable rejection of the old, but little effort to build the new.
In this paper I want to describe the case that the behavioral geneticist has built and to criticize this work from the standpoint of a developmental psychol- ogist who has studied the effects of the family and child-rearing practices. Most of all, however, I would like to refire enthusiasm for undertaking studies that will investigate the process by which family life and parent behavior affect the child. I would like to see this new work by the behavioral geneticists serve as a challenge to develop theories, research designs, and measures that will clarify the nature of family socialization and show its effect.
Research Designs in Behavioral Genetics Studies
The recent research by the behavioral geneticists on genetic influences on intelligence, personality, and interests have typically used one of three designs: One design involves comparing the similarities between monozygotic or identi- cal twins to the similarities between dizygotic or fraternal twins. Since the former have developed from a single fertilized egg, they have identical genetics. The latter have developed from two eggs separately fertilized and thus are simply siblings born at the same time. To the behavioral geneticist, greater similarity in intelligence test or personality scale scores between identical than fraternal twins indicates a genetic effect.
A second design, a much more powerful one, obtains measures from chil- dren adopted at or near birth and compares their scores to those of biological parents, who did not interact with them, and those of adoptive parents, who

Genetics / Socialization Balance I29
reared them. Similarlity in scores between biological parents and the offspring they did not rear suggests a genetic effect. Similarlity between the adoptive parent and the child would indicate an environmental effect-with the matching by adoption agencies sometimes having to be taken into account.
A third design is a variation of the second, but no data are obtained from the biological parents of the adopted offspring. Instead, two samples are used-onc sample of adopted families and one of biological families who are not related to the adopted families but are matched with respect to certain demographic vari- ables. Here a comparison is made between the two sets of correlations to see whether there is more parent-child and sibling similarity in the biological or in the adoptive family.
A design not used in the recent work. though it still excites popular interest, is the study of identical twins reared apart. As many of us felt even before the exposure of Cyril Burt’s fraudulent reports, this occurs too rarely and the sam- ples are too peculiar to make such subjects useful for research. This view is currently shared by behavioral geneticists (Scarr & Kidd. 1983).
A Problem in the Twin-Study Design: thc Assiirription qf Eyurrll? Similur Environments
My present focus will be on the recent adoption research because I believe that studies comparing identical and fraternal twins are seriously flawed. In these studies the assumption is made that identical and fraternal twins have equally similar environments, and i t is only their genetics that are not equally similar. Thus, if the identical twins are more like one another than are fraternal twins on intelligence or personality measures, their greater similarity is taken as proof of a genetic effect and the difference in the size of the correlations is used for the heritability estimate. But identical twins look exactly alike. They are often mis- taken for one another. How can it be that their environment is not more similar than that of fraternal twins who may not resemble each other at all? There is abundant evidence (Hartup, 1983; Sore11 & Nowak, 1981) that adults and peers, parents and siblings, respond different!y to different appearances. Good-looking children get more positive responses from their environment than do unattractive ones; height and body build influence social responses; physical similarity to one parent more than to the other affects parental responses and the child’s identifica- tion. Thus, from this difference alone, same-sex fraternal twins cannot have as similar an environment as do identical twins.
In some ways, fraternal twins may actually have a less similar environment than do ordinary siblings. It is true that they share an ordinal position and have arrived at the same point in the family cycle-that is, to inexperienced or experienced parents, when the family is having economic struggles or during a period of relative financial security, during a time of stress or a period of

130 Hoffman
harmony-and in this sense have a more similar environment than more widely spaced siblings. But they are also more subject to the contrast effect that parents and other siblings impose on them. It is common for parents to think of closely spaced siblings in contrast-e.g., “this is the smart one, this is the dumb one”; “this is the mischievous one, this is the passive one;” the closeness in age leads to a comparison that results in exaggerated labels of contrast. And the children, too, partly in response and partly to establish their own sense of identity, often perceive themselves as different; they may exaggerate minor differences and may seek to develop separate interests.
In any case, there is abundant evidence that the environments of mono- zygotic twins are more similar than those of dizygotic, and most behavioral geneticists acknowledge this (Lytton, 1977; Plomin, Willerman, & Loehlin, 1976; Scarr & Carter-Saltzman, 1979). They hold, however, that the assumption of equally similar environments is nevertheless a valid one, and some empirical work has been undertaken to support this defense. Loehlin & Nichols (1976), for example, have argued that greater environmental similarity does not increase trait similarity. In a sample of monozygotic twins, they found that certain en- vironmental variables-such as whether or not the twins shared a room, played together, or had the same teacher-did not relate to the similarity of their scores on the California Personality Inventory. Plomin, Willerman, and Loehlin ( 1976), focusing more on the degree of physical resemblance between the twins, also studied monozygotic twins (identified as monozygotic by the mother’s report of their physical similarity). They found that within that group, parental reports of the twins’ degree of physical similarity and “confusability” did not relate to the parental reports of the twins’ degree of personality similarity.
These data, however, suggesting that differences in personality do not relate to differences in certain environmental variables or differences in physical ap- pearance for a sample of monozygotic twins, cannot be generalized to the dif- ferences between monozygotic and dizygotic twins. These same studies also show that the differences in environment and physical appearance within the monozygotic sample are much less than the differences within a dizygotic sam- ple. Failure to find an effect for the former does not mean that effects do not exist in the latter group where the environment and physical appearance differences between pairs are much greater. Furthermore, even within the monozygotic sample, the failure to disprove the null hypothesis of no difference does not mean it has been proved. It is possible that the environmental variables examined in the Loehlin and Nichol’s study were not the crucial ones, and total reliance in the Plomin, Willerman, and Loehlin study on parental reports is a particular weak- ness in that design.
Both Lytton (1977) and Scarr and Carter-Saltzman (1979), on the other hand, argue that while it is true monozygotic twins have more similar environ- ments than dizygotic twins, this may be because of their greater genetic sim-

Genetics / Socialization Balance 131
ilarity. Scarr and Carter-Saltzman have attempted to defend the questioned as- sumption in the twin studies by examining the belief in zygosity, the similarity in appearance of the twins, and the similarity in cognitive and personality mea- sures. Actual zygosity related to similarity, but so did belief in zygosity, particu- larly for the personality measures. Dizygotic twins who bore a particularly close physical resemblance to each other were likely to believe they were monozygotic and were more likely to be similar on personality measures. The authors argue that the personality similarity between dizygotic twins, like the physical sim- ilarity between them, may be the cause of their error in thinking they are mono- zygotic. This interpretation is, of course, a possibility, but it is also possible that the greater physical similarity leads to a more similar environment which in turn leads to more similar personalities. Thus, this evidence, which the authors in- terpret as being supportive of the behavioral-genetics assumption, could also be seen as evidence against the assumption. That is, the environmentalist position is that monozygotic twins, because they resemble each other physically, have more similar environments and thus they are more similar in personality and possibly cognitive ability. By the same logic, dizygotic twins who also happen to resem- ble each other closely would have similar environments and therefore more similar personalities. The Scarr and Carter-Saltzman data are consistent with this possibility. Since monozygotic twins are more likely to look like each other than are dizygotic, as a group they are likely to have more similar environments. And, as already indicated, they do (Cohen, Dibble, Grawe, & Pollin, 1973; Jones, 1955; Scarr, 1968). Thus, while the physical similarity is genetic, the effect of genetics on personality and cognition may be only indirect, through its effects on environment.
In summary, in the view of the environmentalists (Kamin, 1974; Lewontin, Rose, & Kamin, 1984), it is because monozygotic twins look alike that they are treated alike, and because they are treated alike, they develop similar person- alities. By the same logic, fraternal twins who look alike would be treated alike, and they too would develop similar personalities. The fact that it is much more probable that monozygotic twins will look alike is exactly why their environ- ments are expected to be more similar. The assumption of equally similar en- vironments for monozygotic and dizygotic twins is thus not a sound one and the empirical efforts to defend it are not convincing. Nevertheless, the entire design of the twin studies rests on this assumption. Since the critics of the assumption believe that identical twins are more similar than fraternal twins not only in genes but also in environment, the many demonstrations that they are also more similar on cognitive abilities, personality traits, and interests do not seem to support a genetic explanation any more than an environmental explanation. As one might expect, the heritability extimates yielded by the twin studies are much higher than those yielded by the adoption studies-particularly with respect to the personality variables (Scarr, Webber, Weinberg, & Wittig, 198 1).

132 Hoffman
Problems in the Adoptive-Study Designs
Criticisms can also be made of the adoptive-family design, but they are less fundamental than the criticisms of the various twin studies. One criticism is that because the adoptive families know they are not biologically related, the parents will expect and seek less similarity with their adopted children; thus there would be less socialization effort in this direction. Scarr and her colleagues (Scarr, Scarf, & Weinberg, 1980) have attempted to dispose of this criticism by a study suggesting that perceived similarity between parents and children was not related to similarity as measured by personality inventories and cognitive tests. Howev- er, they did find greater perceived similarity in biological than in adoptive families. The absence of a relationship between perceived and actual similarity is not proved by the study, however, and, in any case, perceived similarity may not be the crucial variable. Simply knowing the children are adopted may lessen the parents’ efforts to mold the child to their own image either because the parents’ identification with the child is less or because the parents feel more of an obligation to let the adoptive child develop independently.
Other considerations have to do with special characteristics of adoptive families. On one hand, there is evidence that adoptive parents are more highly selected than are nonadoptive parents. To adopt a child, parents must be screened. They are typically of higher socioeconomic status and IQ, and represent a more narrow range than either the biological parents or than a population of nonadoptive families. They have also been screened for marital stability and personal qualities. Furthermore, adoptive parents are likely to be older than parents who have conceived their children. These differences present a number of problems in the comparison between adoptive and natural families that might reduce the size of the correlations for the adoptive families. (See Lewontin, Rose, & Kamin, 1984, for a data-based discussion of this criticism.)
On the other hand, it is also possible that, despite the prescreening, adoptive families are less psychologically healthy than nonadoptive ones. This has been suggested in several psychoanalytic reports (Blum, 1983; Grotevant, 1984), and, in one empirical study (Scarr et al. 1981), adoptive parents scored higher on neuroticism and anxiety scales. Child-rearing practices and their effectiveness could be affected by these differences.
With respect to the intelligence test scores in particular, of course, it should also be noted that, as Scarr (Scan-Salapatek, 1971) has pointed out in her excellent criticism of Jensen’s work, differences among individuals are more likely to be genetic when the environment is an enriched one, as it is in all of the adoption studies. Furthermore, the fact that the adoptive families represent a more homogenous group than the biological families with respect to IQ tends to lower the parent-child correlation in the adoptive families.
These criticisms, however, would not dispose of findings that indicated

Genetics / Socialization Balance I33
significant correlations on intelligence test or personality inventory scores be- tween biological parents and offspring they did not see after birth. Such correla- tions would be impressive evidence for a genetic influence. The criticisms, though, may operate to depress the size of the correlations among the nonrelated members of the adoptive family. The real problem with this particular design is that the correlations obtained among the nonbiological family members under- estimate the effects of socialization both because of these methodological prob- lems and because they involve a distortion of socialization theory. As I will take up later, the behavioral geneticists are using the wrong independent variables for testing the influence of environment, and they may be neglecting important dependent variables as well.
The adoption studies that do not have data on the biological parents have a less powerful test of the genetic hypothesis. They must rely on the differences between biological families and adoptive families to infer a genetic effect, but there may be other differences between these two kinds of families, such as those already mentioned, that are involved. The socialization patterns may be different in these two kinds of families, and different degrees of similarity need not retlect simply genetics.
The Recent Adoption Studies
There are three major adoption studies that have provided most of the recent data on the heredity-environment issue. The most impressive investigation is one being carried out in Texas (Horn, 1983). It is a considerable improvement over any previous adoption study, including the well-known one by Skodak and Skeels (1949). First, in the Texas study there are IQ scores for the biological mothers, the adoptive parents, and the adopted children. This is the most com- plete set of IQ data ever provided by an adoption study. Second, unlike most previous work, data were obtained from the siblings of the adopted child, includ- ing both other adopted children reared in the same family and children born to the adoptive parents. Third, personality tests were administered as well. In addition to the completeness of the data set, the Texas study offers two other improve- ments over previous designs: all adopted children were placed in their adopted families by two weeks of age, and the biological mothers are a more homoge- neous group than in previous studies. (The Beta IQ scores for these women averaged 108.7 with a standard deviation of 8.67.) When the children were tested they were, on the average, eight years of age.
The second adoption study that has contributed heavily to the recent liter- ature is a study of adolescent adopted children conducted by Scarr and her colleagues in Minnesota (Scarr & Weinberg, 1983). In this study there were no data on the biological parents of the adopted children. Instead, the resemblances among family members of adoptive families were compared to resemblances in a

134 Hoffman
sample of biological families matched on various demographic variables to the adoptive families. The instruments administered to all family members included Wechsler intelligence tests and various measures of personality, interests, and attitudes. All of the adopted children were placed within the first year.
The third study is a transracial adoption study also conducted by Scarr and her colleagues (Scarr & Weinberg, 1976). It was undertaken primarily to test the hypothesis-subsequently confirmed-that black and interracial children, reared in the culture of the tests and schools, perform as well as white adopted children in similar families. This study was not designed to test the same hypotheses as the other two, but it has provided some relevant data. In this study, 1 1 1 children were adopted during the first year and 65 after that. The analyses consider these two groups separately. There were no IQ data on biological parents; educational levels were used instead. Both IQ and personality-scale scores were obtained for the adoptive parents, the child, and the child's siblings in his family of adoption. The adopted children ranged in age from 4 to 16 and averaged about 7 years of age when the data were collected.
The Data
In the tables reproduced here from the three studies, Tables 1, 2, 3, 6, and 7, it can be seen that there are moderate to high correlations between biological parents and their children-even when the adopted child was placed within two
Table 1. Correlation of Parent's Beta 10 with Child's I 0 Tests"." ~ ~~~
Correlational pairing
~~ ~~ ~ ~
Child test
Wechsler Wechsler or Performance IQ Binet total IQ
r n r n
Adoptive father and biological child .29 I44 .28 I63 adopted child . I2 405 . I4 462
biological child .21 I43 .20 162 Adoptive mother and
adopted child . I S 40 1 . I7 459
her child .28 297 .31 345 other adopted child in same family . I5 202 .I9 233 biological child in same family .06 I43 .08 161
Unwed mother and
~~~~~ ~ ~ ~~
aData from the Texas Adoption Study (Horn, Loehlin, & Willerman, 1979) bns refer to the number of pairings (= the number of children)-the same parent may enter more than one pairing In the case of twins, the second twin was excluded from the unwed mother-other child comparisons
Genetics / Socialization Balance 135
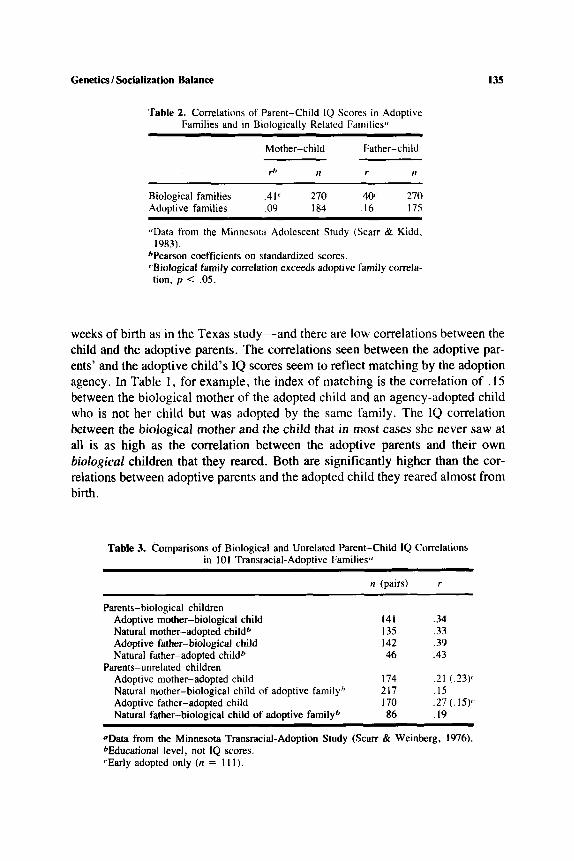
'Table 2. Correlations of Parent-Child IQ Scores in Adoptive Families and in Biologically Related Families"
Mother-child Father-child
z I 1 r I1
Biological families .41( 270 .40< 270 AdoDtive families . 09 I84 .I6 175
~~
"Data from the Minnesota Adolcscent Study (Scarr & Kidd.
bPearson coefficients on standardized scores. cBiological family correlation exceeds adoptive family correla-
1983).
tion, p < .05.
weeks of birth as in the Texas study-and there are low correlations between the child and the adoptive parents. The correlations seen between the adoptive par- ents' and the adoptive child's 1Q scores seem to reflect matching by the adoption agency. In Table 1 , for example, the index of matching is the correlation of . I5 between the biological mother of the adopted child and an agency-adopted child who is not her child but was adopted by the same family. The 1Q correlation between the biological mother and the child that in most cases she never saw ar all is as high as the correlation between the adoptive parents and their own biological children that they reared. Both are significantly higher than the cor- relations between adoptive parents and the adopted child they reared almost from birth.
Table 3. Comparisons of Biological and Unrelated Parent-Child IQ Correlations in 101 Transracial-Adoptive Familiesc'
n (pairs)
Parents-biological children Adoptive mother-biological child 141 Natural mother-adopted childb I35 Adoptive father-biological child I42 Natural father-adopted childh 46
Adoptive mother-adopted child 174 Natural mother-biological child of adoptive family" 217 Adoptive father-adopted child I70 Natural father-biological child of adoptive familyb 86
Parents-unrelated children
r
.34
.33
.39
.43
.21 (.23)['
.I5
.27 (. 15)~'
.19
UData from the Minnesota Transracial-Adoption Study (Scam & Weinberg, 1976). bEducationa1 level, not IQ scores. "Early adopted only ( n = I I I ).

136 Hoffman
Many different data analyses have been reported from these studies, only a few of which are included in the tables presented here. I want to summarize the conclusions that have been reached from these data, and then discuss why I think some of these conclusions are invalid.
There are five general conclusions that have been drawn by the behavioral geneticists from this new body of research (Scarr & Kidd, 1983). First, the case for a genetic influence on 1Q scores has been strengthened. These studies, particularly the Texas study with its improved design, reaffirm that within a reasonably advantaged environment (that of middle-class white Americans) ge- netic factors account for a considerable part of the individual differences. Sec- ond, personality and interests, as measured by the various personality scales and the Strong-Campbell Interest Inventory, also show a genetic influence. This conclusion comes primarily from the Minnesota adolescent study. Third, be- tween-family differences do not explain individual personality or interest dif- ferences. That is, people who are in the same family are not more similar than nonfamily members with respect to personality or interests except for their com- mon genes. Fourth, environmental differences result primarily not from the situation but from genes; that is, individual genotypes evoke and experience different responses from their environment. Fifth, the effects of the family en- vironment diminish as the child matures; the effects of genes increase. This conclusion is based on contrasts that will be discussed between the Minnesota adolescent study and the two studies of younger children.
After an extensive integrative study of this literature, Scarr and Kidd (1983) conclude:
greater genetic resemblance is associated with greater behavioral resemblance, quite strongly in the intellectual domain and to a lesser extent in personality and interests. (p. 419)
and
Being reared in the same family accounts for some portion of the intellectual differences among people in studied populations, but common rearing accounts for little of the variance among individuals in personality or interests. (p. 419)
This new body of data and the conclusions drawn from it constitute a different attack on the environmentalist position than we have previously en- countered. These researchers are not making inferences about differences be- tween groups whose life circumstances are very different. They are careful to clarify that their findings can be generalized only to the white middle-class populations they study. Nevertheless, one implication is that socialization expe- riences, child-rearing patterns in particular, are not very important in shaping children intellectually or socially: Differences in parenting do not have mean- ingful effects, and the early childhood environment washes out with develop- ment.

Genetics / Socialization Balance 137
The main issue I want to address is the geneticist’s conclusion that family environment is not significant in explaining individual differences. 1 feel it is not justified because these researchers have not employed appropriate models and concepts in testing the socialization hypothesis.
The IQ Data
First, let us consider the IQ studies. Although similarities in 1Qs between biological parents and adopted-away children might be an appropriate test to demonstrate a genetic hypothesis, the parents’ IQs are not an appropriate mea- sure of the richness and intellectual stimulation of the home environment. In all three studies, as in all adoption studies, the adoptive parents have above-average 1Qs with a very narrow range in each sample. What socialization theory would suggest that a parent with an IQ of 112 provides a more stimulating environment for her child than a parent with an IQ of 110, but less stimulating than a parent with an IQ of 1141 Researchers in developmental psychology have studied and identified various aspects of family life that affect the child’s cognitive develop- ment. These include the amount of parent-child interaction, the kind of interac- tion, the variety of the stimulation provided, the complexity of the language, whether inductive or power-assertive discipline is used, the degree of indepen- dence training provided, the physical environment, and a host of other variables. It is not known whether parental IQ is related to these factors at all, but it is doubtful that a relationship would be found for a population with as narrow a range of 1Q as that in the adoptive parent samples. In these groups, IQ is probably not even a distal variable estimating the cognitive stimulation that a home provides, and it is certainly not a sufficiently sensitive measure of the environment for a correlational study; but it i s the measure used in all three studies as well as in the previous adoption studies.’
If someone conducted research using the Caldwell Home Inventory (Elardo, Bradley, & Caldwell, 1977), a measure of the stimulation of the home environ- ment, to see if scores obtained in the biological parents’ home were more or less correlated to the child’s IQ than were scores from the adoptive parents’ home, we would have an equally inappropriate test of whether genetics or family environ- ment is more important; but this time, I think environment and not genetics would be the winner. To me, the only fair design would involve not only IQ scores from the biological parents, but some measure of the richness of the cognitive environment in the adoptive home, and it seems likely that, for this restricted population, a new measure would need to be developed. I do not think
‘The importance of home environment rather than maternal IQ. in affecting the child’s develop- ment by 48 months of age, has been shown in a sample of children at risk for mental retardation (Yeates, Mac Phee, Campbell, & Ramey, 1983).

138 Hoffman
such a test would yield figures on how much of a child's IQ score may be attributed to heredity and how much to environment, because these influences interact in too complex a way for figures of this sort to be meaningful. It would, however, provide a better test of the behavioral geneticists' question, and I think they would find that the family environment is much more potent than they now believe.
How do the correlations among unrelated siblings reared in the same family fit into this picture? Examining their degree of similarity would seem to be a more legitimate test of the effects of the home environment because, while there is no measure of the quality of this environment, they have shared it. These data are interesting and are provided by all three studies. In both the Texas study and the Transracial study, biologically unrelated children reared together had sub- stantial correlations in their IQ scores, suggesting the influence of their shared environment. In the Texas study, genetically unrelated siblings reared together had IQ scores that correlated .26. This was lower than the similarity among the biological siblings born to the adoptive parents, .35, but it is a substantial cor- relation. In the Transracial study, the comparable figures were .33 and .42, showing a similar pattern (Horn, Loehlin, & Willerman, 1979). In the Trans- racial study, however, interesting differences emerge when the distinction is made between whether the unrelated sibling pairs involve two adopted children or an adopted child and a biological offspring. These data are shown in Table 4. In this analysis, the unrelated siblings reared together who were all adopted showed a very high correlation of .44. This was just as high as the correlation for the biological siblings in these families. When a separate correlation was com- puted for the sibling pairs that involved an adopted child and a nonadopted child, the correlation was between .25 and .30, depending on the child's age at the time of adoption. This raises the possibility that when parents have both adopted and natural children, they treat them differently. Thus, the Transracial study suggests that the highest correlation among siblings appears when all the children have the same status-either all adopted or all biological. This particular dif-
Table 4. Sibling IQ Correlations, Natural and All Adopted Children of Adoptive Families"
n (pairs) r
Natural siblings I07 .42 Natural siblings-adopted siblings 230 .25 Natural siblings-early adopted siblings 34 .30 All adopted siblings 140 .44 Early adopted siblings 53 .39
UData from the Minnesota Transracial-Adoption Study (Scarr & Kidd, 1983).
Genetics / Socialization Balance 139
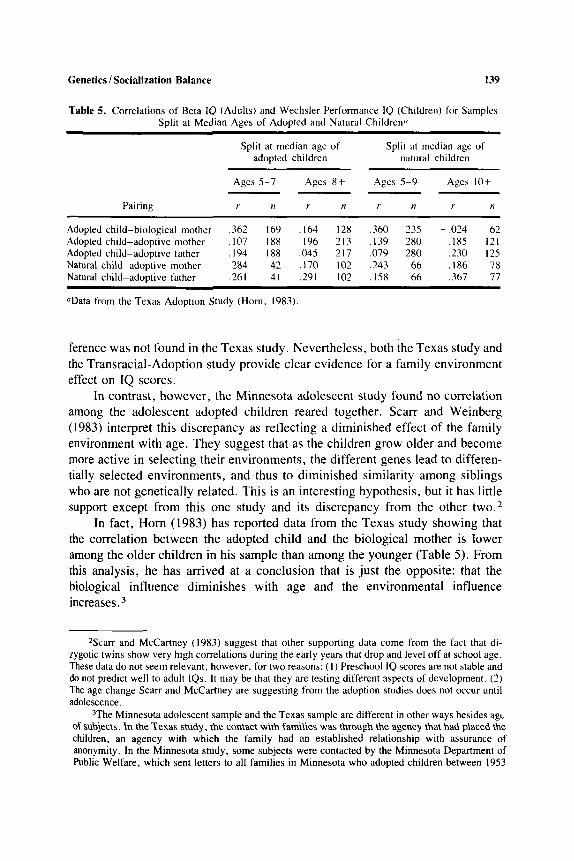
Table 5. Correlations of Beta IQ (Adults) and Wechsler Performance IQ (Children) for Samples Split at Median Ages of Adopted and Natural Children"
Split at median age of Split at median age of adopted children natural children
Agea 5-7 Ages 8 + Ages 5-9 Ages 10+
Pairing r I 1 r n r n r n ~~ ~ ~~ ~
Adopted child-biological mother ,362 169 .I64 128 ,360 235 -.024 62 Adopted child-adoptive mother .I07 188 . I96 213 .I39 280 . IX5 121 Adopted child-adoptive father .I94 188 .045 217 ,079 280 .230 125 Natural child-adoptive mother -284 42 .I70 102 ,243 66 ,186 78 Natural child-adoptive father .261 41 .291 102 .I58 66 ,367 77
~-
"Data from the Texas Adoption Study (Horn, 1983)
ference was not found in the Texas study. Nevertheless, both the Texas study and the Transracial-Adoption study provide clear evidence for a family environment effect on IQ scores.
In contrast, however, the Minnesota adolescent study found no correlation among the adolescent adopted children reared together. Scarr and Weinberg (1983) interpret this discrepancy as reflecting a diminished effect of the family environment with age. They suggest that as the children grow older and become more active in selecting their environments, the different genes lead to differen- tially selected environments, and thus to diminished similarity among siblings who are not genetically related. This is an interesting hypothesis, but it has little support except from this one study and its discrepancy from the other two.*
In fact, Horn (1983) has reported data from the Texas study showing that the correlation between the adopted child and the biological mother is lower among the older children in his sample than among the younger (Table 5). From this analysis, he has arrived at a conclusion that is just the opposite: that the biological influence diminishes with age and the environmental influence increases.
2Scarr and McCartney (1983) suggest that other supporting data come from the fact that di- zygotic twins show very high correlations during the early years that drop and level off at school age. These data do not seem relevant, however, for two reasons: ( I ) Preschool IQ scores are not stable and do not predict well to adult IQs. It may be that they are testing different aspects of development. (2) The age change Scarr and McCartney are suggesting from the adoption studies does not occur until adolescence.
3The Minnesota adolescent sample and the Texas sample are different in other ways besides agt of subjects. ln the Texas study, the contact with famihes was through the agency that had placed the children, an agency with which the family had an established relationship with assurance of anonymity. In the Minnesota study, some subjects were contacted by the Minnesota Department of Public Welfare, which sent letters to all families in Minnesota who adopted children between 1953

140 Hoffman
Thus, the data are not yet clear on whether or not unrelated siblings who share a family environment are similar in IQ. There seems to be a substantial correlation up to adolescence. Whether the Minnesota adolescent study failed to find a relationship because of the age of the subjects or because of some other aspect of the design has not yet been determined.
To sum up the IQ findings, I believe that, despite the conclusions drawn from the data, the behavioral geneticists have not shown that family environment fails to affect intelligence, nor even that it has less effect than genetics. The IQ scores of the adoptive parents do not represent the environment they provide and the sibling data provide mixed evidence.
Personality and Interest Data
Turning now to the personality and interest studies, it should first be noted that the evidence for even a genetic effect is somewhat equivocal and limited to only a few variables. The typical twin studies yield a heritability estimate for the personality variables they examine as high as their heritability estimate for intel- ligence. That is, although the correlations between both identical and fraternal twins are lower for personality measures than for IQ scores, the diference between the two kinds of twins is the same in the personality and IQ studies, thus yielding the same heritability estimate. (Examination of these data reveals that the estimate of heritability from twin studies is much higher when the measure of the twins’ personalities is based on the mother’s report than when it is more independently assessed; Goldsmith, 1983 .)
The heritability estimate for personality measures that is based on twin studies, at best, appears to be exaggerated for two reasons. First, the assumption, already discussed, that identical twins and fraternal twins have equally similar environments seems most untenable when personality traits are considered. Physical appearance has been particularly linked to personality development (Hartup, 1983; Sore11 & Nowak, 1981). The greater difference in the appearance of fraternal twins would elicit greater differences in the responses of both family and nonfamily members, and this would have especial significance for person- ality development. Second, the adoption studies do not confirm the twin studies. The Texas study provides no evidence for a genetic influence on personality; the data showed no similarity between the personalities of biological parents and adopted-out offspring. The Minnesota adolescent study did find evidence, but the heritability estimates yielded by these results are considerably lower than those yielded by the twin studies, .22 as compared to .52 (Scarr and Kidd, 1983).
and 1959; the children had been placed by various agencies or privately. Other subjects were recruited through newspaper advertisements and personal referrals (Grotevant, Scarr, & Weinberg, 1977). It seems likely, then, that the Minnesota families were more self-selected than the Texas families.
Genetics / Socialization Balance 141
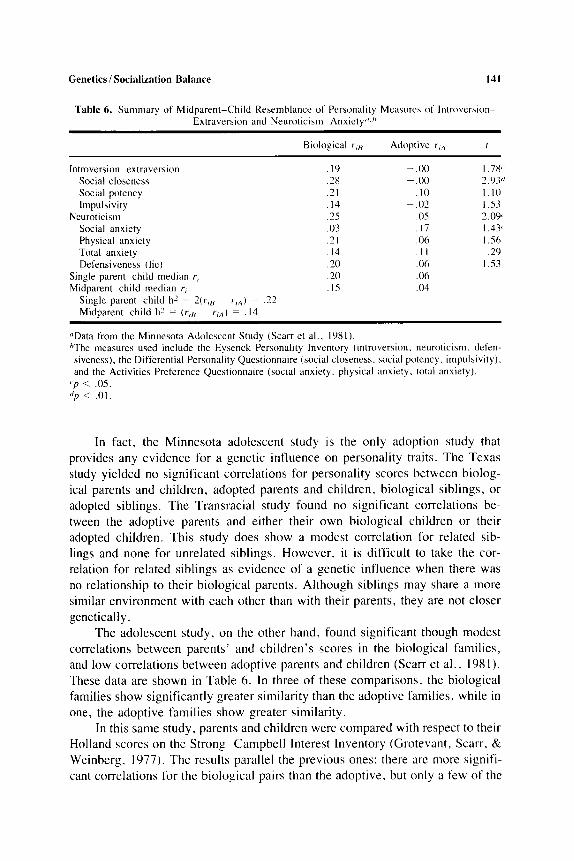
Table 6. Summary of Midparent-Child Resemblance of Personality Measures of Introvcrsion Extraversion and Neurotici~ni- Anxiety"."
Biological r,/< Adoptive t-,,, I
Introversion-extraversion Social closeness Social potcncy Impulsivity
Social anxiety Physical anxiety Totdl anxiety Defensiveness (l ic)
Neuroticism
Single parent-child median r , Midparent-child median r,
. I9
.28
.21
. I4
.25
.03
.21
. I 4
.20
.20
. I5
- .oo ~ .00
. I 0 - .O?
.05
. I 7
.06
. I I
.Oh
.Oh
.04
1.78' 2.93" I. 10 1.53 2.0"
~ 1.431 I .56
.?Y I .s3
Single parent-child h' = 2 ( r r 8 - r-,,,) = .22 Midparent-child h' = (r,{] ~ r,,,) = . I4
"Data from the Minnesota Adolescent Study (Scarr et al . . 1981 ). bThe measures used include the Eysenck Personality Inventory (introversion. neuroticism. defen- siveness). the Differential Personality Questionnaire (social closeness, social potency, impulsivity). and the Activities Preference Questionnaire (social anxiety, physical anxiety. total anxiety).
' p < .0s. </p < .01.
In fact, the Minnesota adolescent study is the only adoption study that provides any evidence for a genetic influence on personality traits. The Texas study yielded no significant correlations for personality scores between biolog- ical parents and children, adopted parents and children, biological siblings, or adopted siblings. The Transracial study found no significant correlations be- tween the adoptive parents and either their own biological children or their adopted children. This study does show a modest correlation for related sib- lings and none for unrelated siblings. However. it is difficult to take the cor- relation for related siblings as evidence of a genetic influence when there was no relationship to their biological parents. Although siblings may share a more similar environment with each other than with their parents, they are not closer genetically.
The adolescent study, on the other hand, found significant though modest correlations between parents' and children's scores in the biological families, and low correlations between adoptive parents and children (Scarr et al., 1981). These data are shown in Table 6. In three of these comparisons, the biological families show significantly greater similarity than the adoptive families. while in one, the adoptive families show greater similarity.
In this same study, parents and children were compared with respect to their Holland scores on the Strong-Campbell Interest Inventory (Grotevant. Scarr. & Weinberg. 1977). The results parallel the previous ones; there are more signifi- cant correlations for the biological pairs than the adoptive. but only a few o f the
142 Hoffman
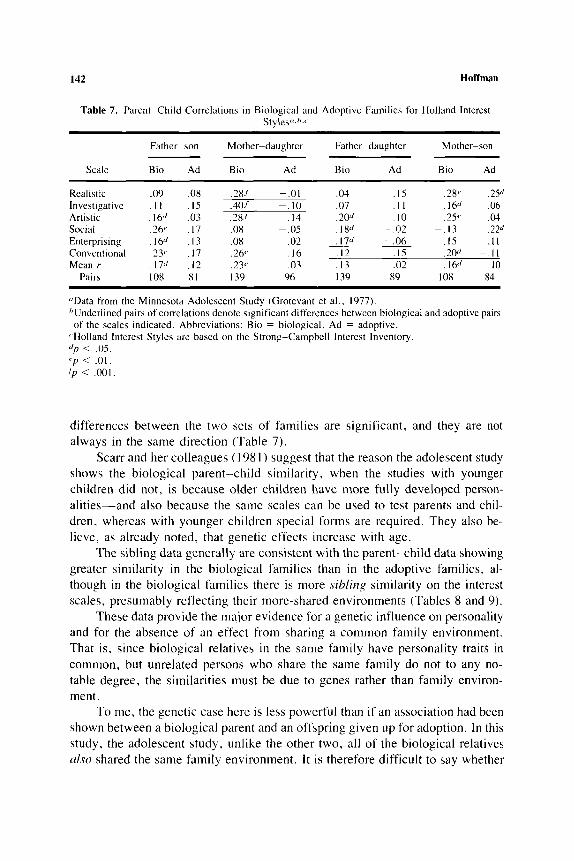
Table 7. Parent-Child Correlations in Biological and Adoptive Families for Holland Interest S t y l e s ~ ~ . h . ~
Father-son Mother-daughter Father-daughter Mother-son
Scale Bio Ad Bio Ad Bio Ad Bio Ad _ _ _ _ _ _ _ _ _ _ _ _ _ _ _ _____ ~ ~~
Realistic .09 .OX .28/ - . O l .04 . IS .2@' .25d Investigative . I I . I 5 .40f - . 10 .07 . I I .16" .06
. I0 .25<' .04 Artistic . I W .03 .28/ . I 4 .& 10" Social ,261' . I7 .ox - .05 . I 8" -.02 - . I3 ,221'
Enterprising .16" -13 .OX .02 . 17" - .06 .I5 . I I Conventional . 2 3 ~ . I7 ,261' .I6 . I 2 - . I S .I 70d - . I I Mean r . 1 7 " . 1 2 .b. 1 ? P .03 . I3 .02 . I & / .I0
Pairs 108 81 I39 96 139 89 I OX 84 ~ ~~ ~ ~~
"Data from the Minnesota Adolescent Study (Grotevant ct a l . , 1977). "Underlined pairs of correlations denote significant differences between biological and adoptive pairs
'Holland Interest Styles are based o n thc Strong-Campbell Interest Inventory. <//I < .os. ep < .01. ' p < ,001.
of the scales indicated. Abhrcviations: Bio = biological. Ad = adoptive.
differences between the two sets of families are significant, and they are not always in the same direction (Table 7).
Scarr and her colleagues ( I98 1 ) suggest that the reason the adolescent study shows the biological parent-child similarity, when the studies with younger children did not, is because older children have more fully developed person- alities-and also because the same scales can be used to test parents and chil- dren, whereas with younger children special forms are required. They also be- lieve, as already noted, that genetic effects increase with age.
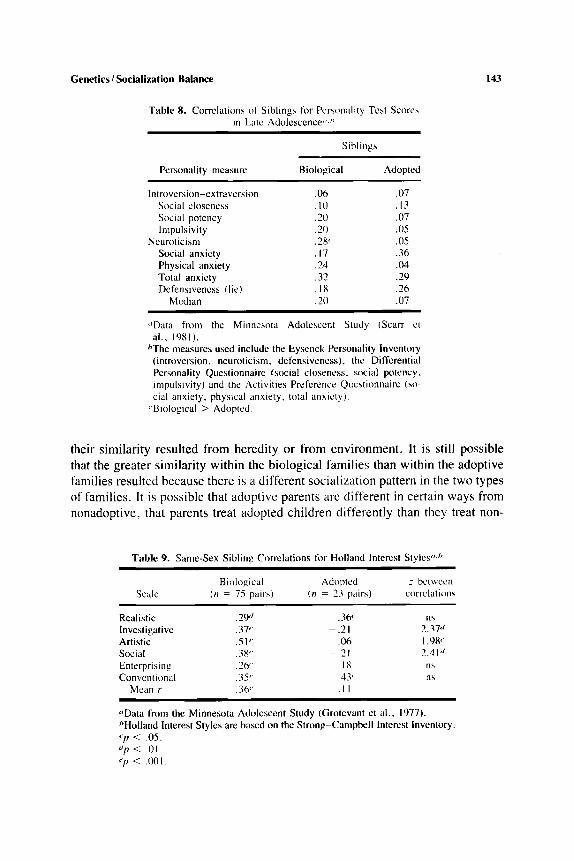
The sibling data generally are consistent with the parent-child data showing greater similarity in the biological families than in the adoptive families, al- though in the biological families there is more sibling similarity on the interest scales, presumably reflecting their more-shared environments (Tables 8 and 9).
These data provide the major evidence for a genetic influence on personality and for the absence of an effect from sharing a common family environment. That is. since biological relatives in the same family have personality traits in common, but unrelated persons who share the same family do not to any no- table degree, the similarities must be due to genes rather than family environ- ment.
To me, the genetic case here is less powerful than if an association had been shown between a biological parent and an offspring given up for adoption. In this study, the adolescent study. unlike the other two, all of the biological relatives d.so shared the same family environment. I t is therefore difficult to say whether
Genetics / Socialization Balance 143
Table 8. Correlations of Siblings for Personality Teat Scores i n Late Adolescencetr ''
~~
Personality measure Biological Ado p t e d
Introversion-ex traversion Social closeness Social potency Impulsivity
Strial anxicty Physical anxiety Total anxiety Defensivcncss (lie)
Median
Neuroticism
.06
. I 0
. 20
.20
. x i
. I7
.24
.32
. I X
. 2 0
.07
. I 3
.07
.05
.05
.36
.04
.2')
.26
.07
<'Data froni the Minnesota Adolescent Study (Sc;irr ct al., 1981).
"The measures used include the Eyscnck Personality Invcntory (introversion. ncuroticism. dcfcnsivencss). the Diffcrentid Personality Questionnaire (sociel closeness. social potency. impulsivity) and the Activities Prcfcrence Qucationnairc (so- cial anxiety. physical anxiety, total anxiety).
1 Biological > Adopted.
their similarity resulted from heredity or from environment. It is still possible that the greater similarity within the biological families than within the adoptive families resulted because there is a different socialization pattern in the two types of families. It is possible that adoptive parents are different in certain ways from nonadoptive, that parents treat adopted children differently than they treat non-
Table 9. Same-Sex Sibling Correlations for Holland lntcrcsl Styles".~'
Biological Adopted : between Scale ( ) I = 75 pairs) ! n : 23 pairs) correlation5
Realistic .- 79d .36< ns Investigative .37r -.21 2.37" Artistic .51<' .06 I .9x. Social .3XC - . ? I -. 3 41" Enterprising .7@ 18 I1 s Conventional ,351' ,331 11s
Mean r .36" .II
"Data from the Minnesota Adolescent Study (Grotcvant ct al.. 1977). "Holland Interest Styles are hascd on thc Strong-Campbell Interest Inventory. ' p < -05. <'p < .o I " p < .oo I .

144 Hoffman
adopted, and that knowing one is an adopted child is itself an environmental variable affecting the impact of the parent’s influence (Grotevant, 1984). The design of the Minnesota adolescent study allows for the possibility that other factors besides genetics explain the differences found.
These same criticisms also pertain to whether or not these data indicate the absence of the effects of a common family environment. That is, the lesser similarity among the members of the adoptive family might result from the special aspects of the adoption situation. But in addition, some of the data, particularly the parent-child comparisons, are inappropriate as tests of an en- vironmental influence. If I were looking for a family effect on personality, I would not seek trait similarity between parents and children. 1 would study child- rearing patterns and the family environment, and see if these predicted to the child variables. Children do not develop as clones of the parent. To explain dependency in a child, I would not expect to find a dependent parent but rather an overprotective one. Anxiety might develop from having an anxious parent, but it might also result because the parent creates a frightening world. A sense of powerlessness is more likely to develop in a child when the parents are all powerful and domineering than when they also feel powerless (Strodtbeck, 1958). Having an extremely competent and self-assured parent may make a child feel inadequate in contrast. So the relative absence of parent-child similarity on such traits does not provide for me an indication of the absence of an environ- mental effect in the adopted families.
Again, the sibling similarities are more relevant to the environment question because the study does not measure environment directly at all. Parent charac- teristics do not describe the child’s relevant environment any more than IQ scores describe the child’s cognitive environment. But siblings share the family en- vironment and so, even if it is unmeasured, commonality among siblings might be expected.
Clearly there is more sibling similarity in the biological than in the adoptive families, and this provides the major evidence for the conclusion “common rearing accounts for little of the variance among individuals in personality or interests” (Scarr and Kidd, 1983, p. 419). It is not true, however, that there is no similarity among the adopted siblings. Similarity is shown for some variables but not for others. Furthermore, family similarity increased in both sets of families in a subsequent analysis by Grotevant ( 1979) of the Strong-Campbell measures when parental agreement on these measures was considered. In any case, at most, statements about the importance of the family environment can only be made about the particular variables that were examined in this study. These variables were not randomly selected. The investigators chose personality di- mensions for which there was some previous evidence of a genetic effect or a temperament influence. With different variables, a family influence and not a genetic influence might be demonstrated. For example, Plomin, Foch, and Rowe

Genetics / Socialization Balance 145
(198 I ) , who are themselves important contributors to the behavioral-genetics literature, recently concluded from a study of BoBo Clown aggression that the family environment, and not genetics, was the major source of individual dif- ferences. And many socialization studies have shown the effects of discipline styles (Baumrind, 1967), family structure (Hetherington, 1965), stress, and other general family variables on children’s personality where it would be quite unrea- sonable to assume that the results could be explained away by a genetic explana- tion or one that viewed the child’s characteristics as the evoker of the family style. The behavioral-geneticists’ generalization from their data that individual differences in personality cannot be explained by between- family differences (Goldsmith, 1983; Scan & Kidd, 1983) shows a lack of familiarity with these studies.
They also fail to understand the complexities of the socialization models current today. Most of us in the socialization area are quite aware that charac- teristics in the child evoke different parental responses and that children differ in their reactions to the same parent behavior. Some of these differences between children may be genetic and some may be based on previous socialization experi- ences. There are enormous within-family differences in the child’s experience. Ordinal position, spacing intervals, sibling interactions, the sex of the child, whether the child resembles one parent or the other, the different ages of each child when a specific family event occurs, individual experiences-a fall, an illness, a specific encounter-all of these operate to create within-family dif- ferences. But while children may be different as elicitors of parent behavior, not all parents respond the same to the same child behavior, so even when the child is the elicitor, parenting styles come through. Personality results from many differ- ent inputs. For some qualities, the general family environment might be the most important; for others, more child-specific experiences might be important; for most, an interaction between the two is probably involved. Thus, to take a particular selected group of personality traits and show that biological siblings are more alike than adopted siblings could not lead to the conclusion that the difSeerence between families cannot explain individual differences in personality. It could not explain all, but no child developmentalist studying family effects would claim it could; it still might explain some.
In summary of the personality and interest studies, I feel the case for genetics is a weak one and the conclusion that differences between families in socialization patterns are not important is an overgeneralization from limited data. Comparing a sample of biological families to a sample of adopted families ignores the possibility that the two kinds of families are different in other ways besides their genetic relatedness. Seeking trait similarity between parents and children is not an appropriate test of socialization theory. And, at best, conclu- sions must be confined to the particular variables that showed the predicted patterns.

146
A Challenge for the Social Developmentalist
Hoffman
I do think, however, that the recent research by the behavioral geneticists and their criticisms of the socialization studies pose a challenge. The best answer to the behavioral-geneticist position will come from research. The need for improved measuring instruments, research designs, and theory is highlighted by their work. For example, while there are measures of the degree of cognitive stimulation provided by the family environment (Clarke-Stewart, 1973; Elardo, Bradley, & Caldwell, 1977; Wachs, 1978; Wachs & Gruen, 1982; Wohlwill & Heft, 1975), many of these are targeted for the infant or preschool child. The development of measures that take into account the shifts in the child’s needs occurring with maturation, and tap not only the quality of the early childhood environment but also the quality of the environment provided during middle childhood and adolescence, would greatly facilitate the investigation of issues raised here.
As for research designs, any study showing a link between a parent behavior and a child quality must consider the possibility of reciprocal causality. It is always possible, for example, that parents use more reasoning in disciplining a reasonable child and this needs to be considered before the conclusion of a parenting effect can be drawn. Studying parent-discipline techniques in relation to different children in the same family would be a valuable addition to the more common design of only comparing different families. Conducting such research using a sample of adopted families would be particularly valuable. The behav- ioral geneticists’ criticism of socialization studies-that what are reported to be effects of parent behavior may actually show responses to the child or manifesta- tions of their common genes-may not apply generally, but it does remind the socialization researcher of a problem that must often be considered. This may require more sophisticated research designs than are now generally used.
Perhaps most important, developmental psychology is in need of new theo- ries that spell out the process by which the family socializes the child. Which parent behavior is important for which effects? What are the ways in which the general family environment is important and when do the more specific aspects of the child’s situation come into play? How do individual differences in children mediate the effects of parental behavior and how do they influence what this behavior is? Although these various patterns of influence are recognized, there has been little systemic effort to specify when one is more predominant than another, which variables are most affected by each, and how the different pat- terns interact.
If the behavioral geneticists have undermined the environmentalist position by using an over-simplified version of socialization theory, part of the problem is because we have not made an adequate effort to discover and demonstrate the complex process of socialization. The best counter-argument can be made by renewed attention to socialization theory and research.

Genetics / Socialization Balance
References
147
Baumrind, D. (1967). Child care practices anteceding three patterns of preschool behavior. Genetcc. Psvchologv Monographs. 75, 43-88.
Blum, H. (1983). Adoptive parents: Generative conflict and generational continuity. In The psvcho- unulytic study OJ the child, Vol. 38 (pp. 167-208). New Haven, CT: Yale Univerjity Presj.
Clarke-Stewart, K. A. ( 1973). Interactions between mothers and their young children: Characteristics and consequences. Monographs of the Societv for Research i n Child Development. 38, 6-7. (Serial No. 153.)
Cohen, D., Dibble, E. , Grawe. J . . & Pollin, W. (1973). Separating identical from fraternal twins. Archives ($Genetic P.yychiatrv. 2Y, 465-469.
Elardo. R . , Bradley. R. . & Caldwell, B. M. (1977). A longitudinal study of the relation of infants' home environments to language development at age three. Child Development. 48. 595-603.
Goldsmith, H. H. (1983). Genetic influences on personality from infancy to adulthood. Chiid Development. 54, 331-355.
Grotevant, H. D. (1979). Environmental influences on vocational interest development in adoles- cents from adoptive and biological families. Child Dcvelopmetir. 50. 854-860.
Grotevant, H. D. (1984, August). Exploration und negoriarion of differences withinfamilies during adolescence. Invited address, annual meeting of the American Psychological Association, Toronto, Canada.
Grotevant, H. D., Scam, S., & Weinberg, R. A. (1977). Patterns of interest similarity in adoptive and biological families. Journal of Personality and Soc.iul Psyhology, 35, 667-676.
Hartup, W. W. (1983). The peer system. In P. H. Mussen (Ed.), Handbook ofchildpsvchologv. 41h edition, Vol. /V (pp. 91-152). New York: Wiley.
Hetherington, E. M. (1965). Developmental study of the effects of sex of the dominant parent on sex role preference, identification and imitation in children. Journal ($ P ersonuliry und Sociul
Horn, J . M. (1983). The Texas adoption project: Adopted children and their intellectual resemblance
Horn, J. M. , Loehlin, J . C., & Willerman, L. (1979). Intellectual resemblance among adoptive and
Jones, H. E. (1955). Perceived differences among twins. Eugenics Quurterlv. 5 . 98-102. Kamin, L. J . (1974). The science and politics oj'lQ. Potomac. MD: Erlbaum. Lewontin, R. C . , Rose, S. , & Kaniin. L. J . (1984). N o t in our genes. New York: Pantheon. Loehlin. J . C . , & Nichols, R. (1976). Heredity, environment. andpersonulitv: A stud! qfN50 xetx qf
Lytton, H. (1977). Do parents create, or respond to, differences in twins'? Developmental Psvchol-
Plomin, R. , Foch, T . T . , & Rowe, D. C. (1981). Bobo clown aggression in childhood: Environment.
Plomin, R.. Willerman, L . . & Loehlin. J . C . (1976). Resemblance in appearance and the equal
Scam, S . (1968). Environmental bias in twin studies. Eugenics Quarterly. 15. 34-40. Scam, S.. & Carter-Saltzman, L. (1979). Twin method: Defense of a critical assumption. Behui?or
Genetics. 9. 527-542. Scarr, S. , & Kidd, K . K. (1983). Developmental behavior genetics. In P. H. Mussen (Ed.). Hund-
book qf child ps.vchologJ3, 4th e d . , Vol. 11. Infancv and developmental p.swhobiolog.v, (pp. 345-433). New York: Wiley.
Scam, S. , & McCartney, K. (1983). How people make their own environments: A theory of genotype + environment effects. Child Development. 54, 424-435.
Scam, S . , & Weinberg, R. A. (1976). IQ test performance of black children adopted by white families. American Psychologist, 31, 726-739.
Scarr, S., & Weinberg, R. A. (1983). The Minnesota adoption studies: Genetic differences and malleability. Child Development, 54, 260-267.
Scam, S. , Scarf, E.. & Weinberg, R . A . (1980). Perceived and actual similarities in biological and adoptive families: Does perceived similarity bias genetic inferences? Behavior Genetics, 10. 445-458.
Psvcholog~. 2, 188-194.
to biological and adoptive parents. Child Development. 54, 268-275.
biological relatives: The Texas adoption project. Behavior Generics. 9. 177-207.
twins. Austin, TX: University of Texas Press.
ORV. 13. 456-459.
not genes. Journal of Research in Persondim, 15. 331-342.
environments assumption in twin studies of personality. Behavior Genetics. 6 . 43-52.

148 Hoffman
Scam, S. , Webber, P. L., Weinberg, R. A. , & Wittig, M. A. (1981). Personality resemblance among adolescents and their parents in biologically-related and adoptive families. Journal of Person- ality and Social Psvchologv, 40. 885-898.
Scam-Salapatek, S. (1971). Race, social class, and IQ. Science. 174. 1285.1295. Skodak, M., & Skeels, H. M. (1949). A final follow-up study of one-hundred adopted children.
Journal of Genetic Psychology, 75, 85- 125. Sorell, G. T., & Nowak, C. A. (1981). The role of physical attractiveness as a contributor to
individual development. In R. M. Lerner & N. A. Bush-Rossnagel (Eds.), Individuals as producers qf their development. (pp. 389-446). New York: Academic Press.
Strodtbeck, F. L. (1958). Family interaction, values, and achievement. In D. C. McClelland, A. L. Baldwin, U. Bronfenbrenner, & F. L. Strodtbeck (Eds.), Talenr and society. Princeton, NJ: Van Nostrand.
Wachs, T. D. (1978). The relationship of infants' physical environment to their Binet performance at 2-4 years. International Journal of Behavioral Development. I , 5 1-56.
Wachs, T. D.. & Gruen, G . (1982). Earlv experience and human development. New York: Plenum. Wohlwill, J . , & Heft, H. (1975, July). Environmenfsfit,for the developing child. Paper presented to
the International Society for the Study of Behavioral Development, Surrey, England. Yeates, K. O., Mac Phee, D., Campbell, F. A, , & Ramey. C. T. (1983). Maternal I.Q. and home
environment as determinants of early childhood intellectual competence: A developmental analysis. Developmental Psvchologv, 19. 73 1-739.





























