Embed Size (px)
Citation preview

The Fibronectin Extra Domain A Activates MatrixMetalloproteinase Gene Expression by anInterleukin-1-dependent Mechanism*
(Received for publication, April 5, 1999, and in revised form, July 22, 1999)
Shigeki Saito‡, Noboru Yamaji, Kunio Yasunaga, Tetsu Saito, Shun-ichiro Matsumoto,Masao Katoh, Seiji Kobayashi, and Yasuhiko Masuho
From the Institute for Drug Discovery Research, Yamanouchi Pharmaceutical Co., Ltd., 21 Miyukigaoka, Tsukuba,Ibaraki 305-8585, Japan
The extra domain-A (EDA), present in fibronectin (FN)molecules arising from alternatively spliced transcripts,appears only during specific biological and pathogenicprocesses. However, its function is poorly understood.To define the physiologic role of this domain in jointconnective tissue, the biological effects on rabbit carti-lage explants, chondrocytes, and synovial cells werestudied. A recombinant EDA protein (rEDA) increasedproteoglycan release (3.6-fold) in cartilage explant cul-tures and markedly induced production of matrix met-alloproteinase (MMP)-1 in chondrocytes. In addition,rEDA induced MMP-1, MMP-3, and MMP-9 in synovialcells. These effects were elicited only by rEDA, while itsneighboring type III repeats, III11 or III12, scarcely hadany such effects. Interestingly, reorganization of F-actinstress fibers accompanied MMP-1 expression in synovialcells treated with rEDA, suggesting alteration of cellularphenotype. Subsequent Northern blotting revealed ex-pression of pro-inflammatory cytokines, including inter-leukin (IL)-1a and IL-1b, was induced by rEDA prior toMMP-1 expression. Delayed MMP-1 expression suggeststhat rEDA-induced IL-1s promote MMP-1 expression inan autocrine manner. This hypothesis is supported bythe reduction of EDA-induced MMP-1 production byIL-1 receptor antagonist. The effect of EDA on MMP-1production was reduced by connection with an adjacenttype III repeat on either the NH2 or COOH side of EDAand was abolished by connection on both sides of EDA,suggesting that exposure of either the NH2 or COOHterminus of EDA domain by proteolytic cleavage re-leases the inducing activity. In agreement with theseresults, full-length cellular FN did not induce MMP-1production. Furthermore, a 160-kDa EDA-positive FNfragment, which was purified from human placental tis-sue and corresponds to the region from NH2 terminusthrough the EDA, induced MMP-1 production. Takentogether, these results suggest that the EDA in FN frag-ments triggers alterations of cell physiology and plays arole in matrix degradation in joint connective tissue.
Fibronectin (FN)1 is a multifunctional glycoprotein abun-
dant in plasma and widely distributed in the extracellularmatrix (1). It is a dimer of subunits cross-linked by disulfidebonds. Each FN monomer is comprised of three types of repeat-ing units designated type I, II, and III (2). Some of theserepeats bind to cell surface and extracellular matrix compo-nents such as integrins, collagens, heparin, and fibrin. Severalof these binding activities have been assigned to the motifsequences in FN, including the Arg-Gly-Asp (RGD) motif in thetype III10 domain (3), the Pro-His-Ser-Arg-Asn (PHSRN) motifin the type III9 domain (4), and the CS-1 sequence in the III-CSregion (5). Consequently, this multifunctional glycoprotein me-diates a variety of cellular functions including cell adhesion,cell migration, and cell differentiation.
FN molecules have multiple isoforms generated from a singlegene by alternative splicing of combinations of 3 exons: extradomain-A (EDA), extra domain-B (EDB), and III-CS. Both EDAand EDB exons are type III repeating units (6). Plasma fi-bronectin (pFN), produced by hepatocytes and abundant inplasma, lacks both the EDA and EDB domains. However, cel-lular FNs (cFNs), many of which are insoluble and incorpo-rated into the pericellular matrix, contain the EDA and EDBsegments in various combinations. The EDA domain is presentin FN molecules produced during embryonic development (7).However, its presence in adults is minimal except in somedisease states such as rheumatoid arthritis (8), wound healing(9), epithelial fibrosis (10), and vascular intimal proliferation(11). Thus, the highly regulated splicing of the EDA domaininto transcripts suggests significantly different FN functions.It was reported that pFN and cFN differ in structural andphysical properties such as glycosylation (12) and solubility(13) as well as presence of EDA domain. However, the biolog-ical functions the EDA domain confers upon these moleculesare still poorly understood (7).
In this study, the biological functions of the FN EDA domainwere studied in synovial cells and chondrocytes, which areintegral components of joint connective tissue metabolic regu-lation. The results show induction of MMPs and pro-inflamma-tory cytokines occurs in response to exposure to the EDA do-main. This observation, that a distinct domain of anextracellular matrix protein can trigger gene expression, hasprofound implications on biological and pathologic processes.
EXPERIMENTAL PROCEDURES
Preparation of Recombinant FN Type III Repeat Proteins—cDNAsencoding individual human fibronectin type III repeats were amplified
* The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked“advertisement” in accordance with 18 U.S.C. Section 1734 solely toindicate this fact.
‡ To whom correspondence should be addressed: Product PlanningDept., Yamanouchi Pharmaceutical Co., Ltd., 3-17-1, Hasune, Itabashi,Tokyo 174–8612, Japan. Tel.: 81-3-5916-5543; Fax: 81-3-5916-5616.
1 The abbreviations used are: FN, fibronectin; EDA, extra domain-A;EDB, extra domain-B; pFN, plasma fibronectin; cFN, cellular fibronec-tin; MMP, matrix metalloproteinase; IL-1, interleukin-1; VEGF, vascu-
lar endothelial cell growth factor; IL-1RA, interleukin-1 receptor antag-onist; FBS, fetal bovine serum; ELISA, enzyme-linked immunosorbentassay; RT-PCR, reverse transcription-polymerase chain reaction;DMEM, Dulbecco’s modified Eagle’s medium; mAb, monoclonal anti-body; PAGE, polyacrylamide gel electrophoresis; G3PDH, glyceralde-hyde-3-phosphate dehydrogenase.
THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 274, No. 43, Issue of October 22, pp. 30756–30763, 1999© 1999 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U.S.A.
This paper is available on line at http://www.jbc.org30756
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from

by reverse transcription-polymerase chain reaction (RT-PCR) of humanliver mRNA (CLONTECH, Palo Alto, CA), using Takara RNA PCR kit(AMV) version 2.1 (Takara, Kyoto, Japan) according to the manufac-turer’s instructions. The PCR profile consisted of 40 cycles of 94 °C for30 s, 55 °C for 30 s, and 72 °C for 1 min. After the final cycle, thereaction was maintained at 72 °C for an additional 7 min. The primersfor EDA (from Asn1600 through Thr1689) are the sense primer EDA-s,59-CCATATGAACATTGATCGCCCTAAAGGACT-39 and the antisenseprimer EDA-a, 59-AGCGGCCGCTGTGGACTGGGTTCCAATCAGGG-G-39. The primers for III11 (from Glu1510 through Thr1599) are the senseprimer III11-s, 59-CCATATGGAAATTGACAAACCATCCCA-39 and theantisense primer III11-a, 59-AGCGGCCGCGGTTACTGCAGTCTGAAC-CA-39. The primers for III12 (from Ala1690 through Thr1779) are the senseprimer III12-s, 59- CCATATGGCTATTCCTGCACCAACTGA-39 and theantisense primer III12-a, 59- AGCGGCCGCAGTGGTGACAACACCCT-GAG-39. The NdeI site on the sense primers and the NotI site on theantisense primers are underlined. Amino acids are numbered from theamino-terminal pyroglutamate of the mature protein (GenBank acces-sion number X02761). cDNAs encoding FN fragment proteins withseveral permutations of type III repeats consisting of III11, EDA, andIII12, as depicted in Fig. 9A, were also amplified by RT-PCR as de-scribed above. PCR was performed using primer sets, III11-s and EDA-afor the III11-EDA protein, and EDA-s and III12-a for the EDA-III12
protein. cDNAs encoding the III11-EDA-III12 protein and the III11-III12
protein were amplified using the common primer set, III11-s and III12-a,and they were then separated on agarose electrophoresis according totheir molecular sizes. PCR-amplified cDNA was subcloned into thepCRII plasmid (Invitrogen, Carlsbad, CA), digested with NdeI and NotI,then subcloned into the bacterial expression vector pET-22b (Novagen,Madison, WI), which enables expression of fusion proteins carrying sixadditional histidine residues (63His tags) at the carboxyl terminus.Individual FN type III protein genes with 63His tags were expressed inEscherichia coli BL21(DE3) and purified by TALON Metal AffinityResin column (CLONTECH). Endotoxin levels in the highest rEDAconcentration used were under 0.2 ng/ml, which are insufficient toaffect MMP biosynthesis in the culture systems subsequently described.
Proteoglycan Degradation Assay—Rabbit articular cartilage was ob-tained as described previously (14). The cartilage was cut into slices,whose weight was adjusted to approximately 5 mg and placed in thewells of 24-well plates. Explants were cultured in 0.5 ml of Dulbecco’smodified Eagle’s medium (DMEM) containing 5% fetal bovine serum(FBS) for 48 h, with or without exogenous recombinant FN type IIIproteins. Glycosaminoglycan in the culture supernatants and papain-digested cartilage were determined by the dimethylmethylene blueassay (15). Results are expressed as the percentage of glycosaminogly-can in the culture supernatant to the sum of glycosaminoglycan con-tents in the culture supernatants and cartilage digested with papain.
Cell Culture—Synovial cells and chondrocytes were prepared fromJapanese white rabbits. Synovial cells were isolated from synovialtissue incubated with 0.4% collagenase A (Roche Molecular Biochemi-cals, Indianapolis, IN) for 3 h at 37 °C followed by incubation with0.25% trypsin and 1 mM EDTA for 1 h at 37 °C. Chondrocytes wereisolated from cartilage slices incubated with 0.25% trypsin and 1 mM
EDTA for 1 h at 37 °C followed by incubation with 0.15% collagenase Afor 3 h at 37 °C. Cells were cultured to a subconfluent monolayer inDMEM containing 10% FBS. To assess effects of rEDA, cells werestimulated with different recombinant FN type III proteins in DMEMcontaining 5% FBS. Synovial cells were used for assays after just 1passage. Chondrocytes were used without passage.
Measurement of Marix Metalloproteinase-1 (MMP-1) by Enzyme-linked Immunosorbent-Assay (ELISA)—MMP-1 in the culture superna-tants was measured using a double-antibody sandwich ELISA systemthat recognizes pro and active forms of MMP-1, as described previously(14). Mouse polyclonal antibodies generated against rabbit pro-MMP-1were used as both the primary trapping antibody and the secondarybiotinylated antibody. Briefly, plates were coated with the first anti-body and blocked with 1% bovine serum albumin. The plates were thenincubated with the culture supernatants of rabbit synovial cells, fol-lowed by incubation with the secondary biotinylated antibody. Theplates were developed using peroxidase-conjugated streptavidin (Am-ersham Pharmacia Biotech) at a dilution rate of 1:2000, followed byaddition of tetramethylbenzidine. Absorbance at 450 nm was deter-mined using a plate reader.
Western Blot Analysis of MMP-1 and MMP-3—The culture superna-tants of rabbit synovial cells were heat-denatured with 2-mercaptoeth-anol and subjected to SDS-PAGE. Proteins were transferred onto apolyvinylidene difluoride membrane and blocked with Block Ace (Snow
Brand, Sapporo, Japan). The membrane was then incubated with 1mg/ml mouse anti-rabbit pro-MMP-1 polyclonal antibodies or the mousemonoclonal antibody MP1807, which was generated against rabbitMMP-3 as described previously (16). Both antibodies recognize pro andactive forms. After washing, the membrane was incubated with perox-idase-conjugated rabbit anti-mouse IgG antibodies (Zymed Laborato-ries Inc., San Francisco, CA) at a dilution rate of 1:2000, followed bystaining using an enhanced chemiluminescent detection kit (NEN LifeScience Products Inc., Boston, MA) according to the manufacturer’sinstructions.
Gelatin Zymography—The culture supernatants of rabbit synovialcells were applied without reduction to a 10% polyacrylamide slab gelimpregnated with 1 mg/ml gelatin. Gel electrophoresis was performedat 4 °C. After electrophoresis, the gel was incubated in 2.5% (v/v) TritonX-100 for 1 h and then for 18 h at 37 °C in 50 mM Tris-HCl, pH 7.5,containing 200 mM NaCl, 10 mM CaCl2, 10 mM ZnCl2, and 0.02% Brij-35.The gel was then stained with Coomassie Brilliant Blue.
Northern Blotting—Northern blotting was performed as describedpreviously (17). Total RNA was isolated from rabbit synovial cells usingan ISOGEN kit (WAKO, Osaka, Japan), according to the manufactur-er’s instructions. Total RNA (20 mg) was separated on agarose gels,transferred to Hybond-N nylon membrane(Amersham Pharmacia Bio-tech) and hybridized to 32P-labeled cDNA probes. Probes for rabbitMMP-1, rabbit MMP-3, rabbit MMP-2, rabbit MMP-9, and glyceralde-hyde-3-phosphate dehydrogenase (G3PDH) were prepared using aBcaBEST Labeling Kit (Takara, Kyoto, Japan), as described previously(18). The probe for rabbit vascular endothelial cell growth factor(VEGF) was prepared from cDNA fragments obtained according to themethod of Maniscalco et al. (19). Other probes were generated fromcDNA fragments corresponding to nucleotide 6–496 for rabbit interleu-kin-1a (IL-1a) (GenBank accession number X02852) and nucleotide227–778 for rabbit IL-1b (GenBank accession number M26295).
Cytohistochemistry—Rabbit synovial cells were cultured using a Lab-Tek Chamber Slide system (Nunc, Naperville, IL) in DMEM containing10% FBS. Subconfluent synovial cells were treated with different re-combinant FN type III proteins for 48 h. Slides were then washed withphosphate-buffered saline, fixed with 3.7% paraformaldehyde for 20min, permeabilized with ice-cold acetone, and stained with fluorescein-phalloidin (Molecular Probes, Eugene, OR) and propidium iodide(WAKO, Osaka, Japan). The coverslips were then mounted using aSlowFade-Light Antifade kit (Molecular Probes) and examined withconfocal laser scanning microscopy (Olympus, Tokyo, Japan).
Preparation and Characterization of FN Proteins and an FN ProteinFragment—pFN was purified from human plasma as described previ-ously (17). cFN, obtained from Upstate Biotechnology (Lake Placid,NY), was treated with the Kurimover II (Kurita, Tokyo, Japan) columnaccording to the manufacturer’s instructions to remove endotoxin. TheEDA-positive FN protein fragment was prepared from human placen-tas. Placental tissue was homogenized and extracted in 4 M urea in 20mM Tris-HCl (pH 7.5) containing 1 mM phenylmethylsulfonyl fluorideaccording to the method of Laine et al. (20). The extract was diluted20-fold with 20 mM Tris-HCl (pH 7.5), 1 mM phenylmethylsulfonylfluoride and applied to a Gelatin-Sepharose CL-4B column. The gelatin-column was washed with 20 mM Tris-HCl (pH 7.5), 0.15 M NaCl (TBS),1 mM phenylmethylsulfonyl fluoride, then eluted with 1.5 M KBr-TBS,followed by 8 M urea-TBS. The EDA-positive FN fragment was found infractions eluted with 8 M urea by Western blotting using rabbit anti-EDA polyclonal antibodies, which were raised against rEDA. Thesefractions were dialyzed against 10 mM phosphate buffer (pH 7.0) andfurther purified by hydroxyapatite chromatography (CHT-10, BioLogicsystem, Bio-Rad) with a 10–500 mM linear gradient of phosphate buffer(pH 7.0).
The binding activities of anti-EDA monoclonal antibodies (mAbs)against different epitopes of EDA-positive FN fragment were deter-mined by ELISA, as follows. Plates were coated with a single type ofEDA-positive FN fragment, blocked with 1% bovine serum albumin,then were incubated with the anti-FN mAb being tested, followed byhorseradish peroxidase-conjugated rabbit anti-mouse IgG (Zymed Lab-oratories Inc., San Francisco, CA). Anti-FN mAbs, FN9-1, FNC4-4,FN12-8, FNH3-8, and FN8-12, which were obtained from Takara(Kyoto, Japan), specifically recognize the amino-terminal heparin/fi-brin-binding domain, the gelatin-binding domain, the cell-binding do-main, the second heparin-binding domain, and the COOH-terminalfibrin-binding domain, respectively.
EDA-induced Expression of MMP and IL-1 30757
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from
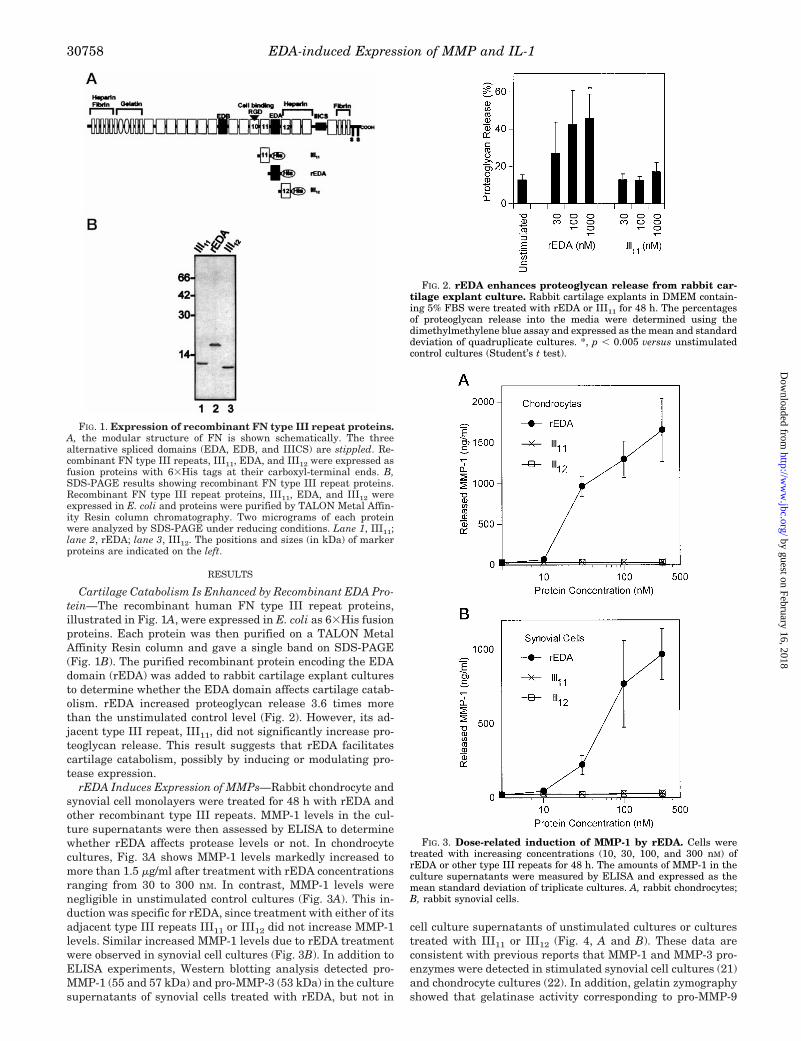
RESULTS
Cartilage Catabolism Is Enhanced by Recombinant EDA Pro-tein—The recombinant human FN type III repeat proteins,illustrated in Fig. 1A, were expressed in E. coli as 63His fusionproteins. Each protein was then purified on a TALON MetalAffinity Resin column and gave a single band on SDS-PAGE(Fig. 1B). The purified recombinant protein encoding the EDAdomain (rEDA) was added to rabbit cartilage explant culturesto determine whether the EDA domain affects cartilage catab-olism. rEDA increased proteoglycan release 3.6 times morethan the unstimulated control level (Fig. 2). However, its ad-jacent type III repeat, III11, did not significantly increase pro-teoglycan release. This result suggests that rEDA facilitatescartilage catabolism, possibly by inducing or modulating pro-tease expression.
rEDA Induces Expression of MMPs—Rabbit chondrocyte andsynovial cell monolayers were treated for 48 h with rEDA andother recombinant type III repeats. MMP-1 levels in the cul-ture supernatants were then assessed by ELISA to determinewhether rEDA affects protease levels or not. In chondrocytecultures, Fig. 3A shows MMP-1 levels markedly increased tomore than 1.5 mg/ml after treatment with rEDA concentrationsranging from 30 to 300 nM. In contrast, MMP-1 levels werenegligible in unstimulated control cultures (Fig. 3A). This in-duction was specific for rEDA, since treatment with either of itsadjacent type III repeats III11 or III12 did not increase MMP-1levels. Similar increased MMP-1 levels due to rEDA treatmentwere observed in synovial cell cultures (Fig. 3B). In addition toELISA experiments, Western blotting analysis detected pro-MMP-1 (55 and 57 kDa) and pro-MMP-3 (53 kDa) in the culturesupernatants of synovial cells treated with rEDA, but not in
cell culture supernatants of unstimulated cultures or culturestreated with III11 or III12 (Fig. 4, A and B). These data areconsistent with previous reports that MMP-1 and MMP-3 pro-enzymes were detected in stimulated synovial cell cultures (21)and chondrocyte cultures (22). In addition, gelatin zymographyshowed that gelatinase activity corresponding to pro-MMP-9
FIG. 1. Expression of recombinant FN type III repeat proteins.A, the modular structure of FN is shown schematically. The threealternative spliced domains (EDA, EDB, and IIICS) are stippled. Re-combinant FN type III repeats, III11, EDA, and III12 were expressed asfusion proteins with 63His tags at their carboxyl-terminal ends. B,SDS-PAGE results showing recombinant FN type III repeat proteins.Recombinant FN type III repeat proteins, III11, EDA, and III12 wereexpressed in E. coli and proteins were purified by TALON Metal Affin-ity Resin column chromatography. Two micrograms of each proteinwere analyzed by SDS-PAGE under reducing conditions. Lane 1, III11;lane 2, rEDA; lane 3, III12. The positions and sizes (in kDa) of markerproteins are indicated on the left.
FIG. 2. rEDA enhances proteoglycan release from rabbit car-tilage explant culture. Rabbit cartilage explants in DMEM contain-ing 5% FBS were treated with rEDA or III11 for 48 h. The percentagesof proteoglycan release into the media were determined using thedimethylmethylene blue assay and expressed as the mean and standarddeviation of quadruplicate cultures. *, p , 0.005 versus unstimulatedcontrol cultures (Student’s t test).
FIG. 3. Dose-related induction of MMP-1 by rEDA. Cells weretreated with increasing concentrations (10, 30, 100, and 300 nM) ofrEDA or other type III repeats for 48 h. The amounts of MMP-1 in theculture supernatants were measured by ELISA and expressed as themean standard deviation of triplicate cultures. A, rabbit chondrocytes;B, rabbit synovial cells.
EDA-induced Expression of MMP and IL-130758
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from
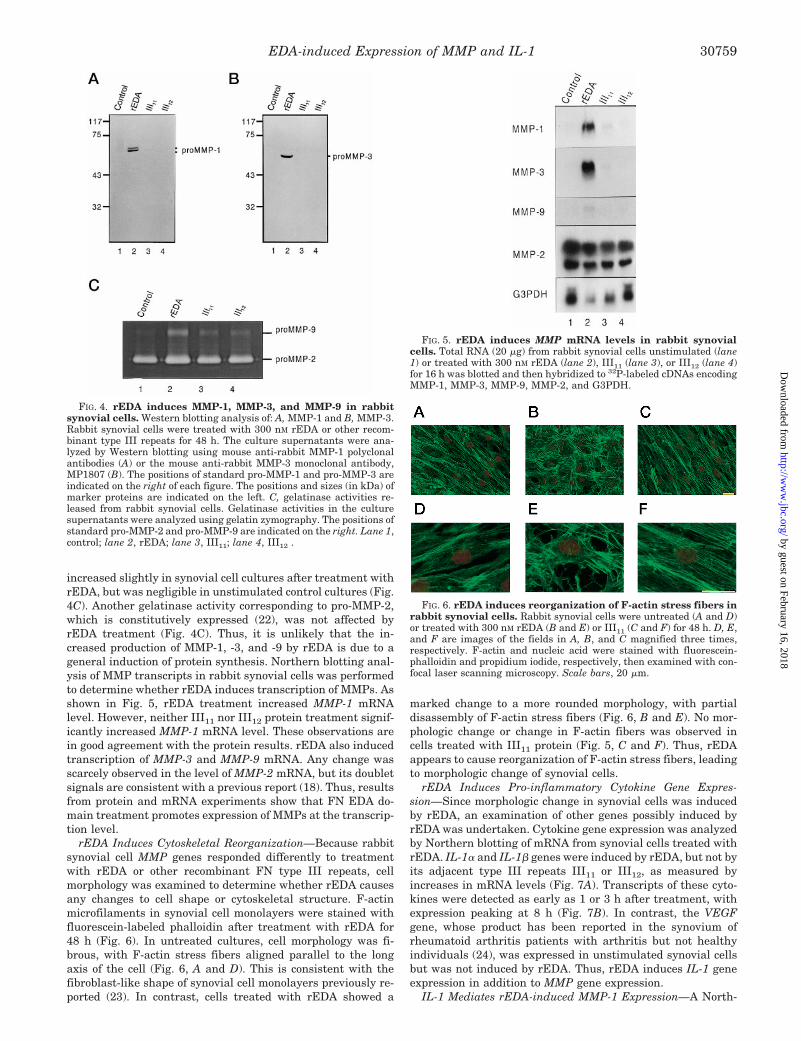
increased slightly in synovial cell cultures after treatment withrEDA, but was negligible in unstimulated control cultures (Fig.4C). Another gelatinase activity corresponding to pro-MMP-2,which is constitutively expressed (22), was not affected byrEDA treatment (Fig. 4C). Thus, it is unlikely that the in-creased production of MMP-1, -3, and -9 by rEDA is due to ageneral induction of protein synthesis. Northern blotting anal-ysis of MMP transcripts in rabbit synovial cells was performedto determine whether rEDA induces transcription of MMPs. Asshown in Fig. 5, rEDA treatment increased MMP-1 mRNAlevel. However, neither III11 nor III12 protein treatment signif-icantly increased MMP-1 mRNA level. These observations arein good agreement with the protein results. rEDA also inducedtranscription of MMP-3 and MMP-9 mRNA. Any change wasscarcely observed in the level of MMP-2 mRNA, but its doubletsignals are consistent with a previous report (18). Thus, resultsfrom protein and mRNA experiments show that FN EDA do-main treatment promotes expression of MMPs at the transcrip-tion level.
rEDA Induces Cytoskeletal Reorganization—Because rabbitsynovial cell MMP genes responded differently to treatmentwith rEDA or other recombinant FN type III repeats, cellmorphology was examined to determine whether rEDA causesany changes to cell shape or cytoskeletal structure. F-actinmicrofilaments in synovial cell monolayers were stained withfluorescein-labeled phalloidin after treatment with rEDA for48 h (Fig. 6). In untreated cultures, cell morphology was fi-brous, with F-actin stress fibers aligned parallel to the longaxis of the cell (Fig. 6, A and D). This is consistent with thefibroblast-like shape of synovial cell monolayers previously re-ported (23). In contrast, cells treated with rEDA showed a
marked change to a more rounded morphology, with partialdisassembly of F-actin stress fibers (Fig. 6, B and E). No mor-phologic change or change in F-actin fibers was observed incells treated with III11 protein (Fig. 5, C and F). Thus, rEDAappears to cause reorganization of F-actin stress fibers, leadingto morphologic change of synovial cells.
rEDA Induces Pro-inflammatory Cytokine Gene Expres-sion—Since morphologic change in synovial cells was inducedby rEDA, an examination of other genes possibly induced byrEDA was undertaken. Cytokine gene expression was analyzedby Northern blotting of mRNA from synovial cells treated withrEDA. IL-1a and IL-1b genes were induced by rEDA, but not byits adjacent type III repeats III11 or III12, as measured byincreases in mRNA levels (Fig. 7A). Transcripts of these cyto-kines were detected as early as 1 or 3 h after treatment, withexpression peaking at 8 h (Fig. 7B). In contrast, the VEGFgene, whose product has been reported in the synovium ofrheumatoid arthritis patients with arthritis but not healthyindividuals (24), was expressed in unstimulated synovial cellsbut was not induced by rEDA. Thus, rEDA induces IL-1 geneexpression in addition to MMP gene expression.
IL-1 Mediates rEDA-induced MMP-1 Expression—A North-
FIG. 4. rEDA induces MMP-1, MMP-3, and MMP-9 in rabbitsynovial cells. Western blotting analysis of: A, MMP-1 and B, MMP-3.Rabbit synovial cells were treated with 300 nM rEDA or other recom-binant type III repeats for 48 h. The culture supernatants were ana-lyzed by Western blotting using mouse anti-rabbit MMP-1 polyclonalantibodies (A) or the mouse anti-rabbit MMP-3 monoclonal antibody,MP1807 (B). The positions of standard pro-MMP-1 and pro-MMP-3 areindicated on the right of each figure. The positions and sizes (in kDa) ofmarker proteins are indicated on the left. C, gelatinase activities re-leased from rabbit synovial cells. Gelatinase activities in the culturesupernatants were analyzed using gelatin zymography. The positions ofstandard pro-MMP-2 and pro-MMP-9 are indicated on the right. Lane 1,control; lane 2, rEDA; lane 3, III11; lane 4, III12 .
FIG. 5. rEDA induces MMP mRNA levels in rabbit synovialcells. Total RNA (20 mg) from rabbit synovial cells unstimulated (lane1) or treated with 300 nM rEDA (lane 2), III11 (lane 3), or III12 (lane 4)for 16 h was blotted and then hybridized to 32P-labeled cDNAs encodingMMP-1, MMP-3, MMP-9, MMP-2, and G3PDH.
FIG. 6. rEDA induces reorganization of F-actin stress fibers inrabbit synovial cells. Rabbit synovial cells were untreated (A and D)or treated with 300 nM rEDA (B and E) or III11 (C and F) for 48 h. D, E,and F are images of the fields in A, B, and C magnified three times,respectively. F-actin and nucleic acid were stained with fluorescein-phalloidin and propidium iodide, respectively, then examined with con-focal laser scanning microscopy. Scale bars, 20 mm.
EDA-induced Expression of MMP and IL-1 30759
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from
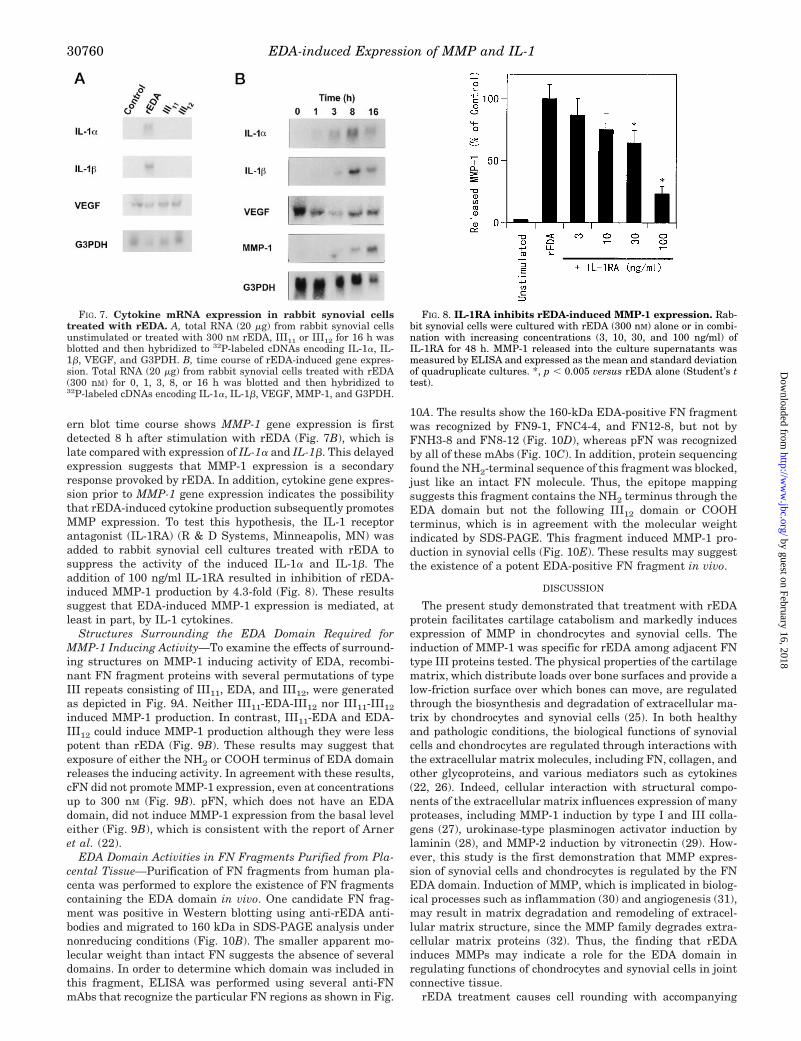
ern blot time course shows MMP-1 gene expression is firstdetected 8 h after stimulation with rEDA (Fig. 7B), which islate compared with expression of IL-1a and IL-1b. This delayedexpression suggests that MMP-1 expression is a secondaryresponse provoked by rEDA. In addition, cytokine gene expres-sion prior to MMP-1 gene expression indicates the possibilitythat rEDA-induced cytokine production subsequently promotesMMP expression. To test this hypothesis, the IL-1 receptorantagonist (IL-1RA) (R & D Systems, Minneapolis, MN) wasadded to rabbit synovial cell cultures treated with rEDA tosuppress the activity of the induced IL-1a and IL-1b. Theaddition of 100 ng/ml IL-1RA resulted in inhibition of rEDA-induced MMP-1 production by 4.3-fold (Fig. 8). These resultssuggest that EDA-induced MMP-1 expression is mediated, atleast in part, by IL-1 cytokines.
Structures Surrounding the EDA Domain Required forMMP-1 Inducing Activity—To examine the effects of surround-ing structures on MMP-1 inducing activity of EDA, recombi-nant FN fragment proteins with several permutations of typeIII repeats consisting of III11, EDA, and III12, were generatedas depicted in Fig. 9A. Neither III11-EDA-III12 nor III11-III12
induced MMP-1 production. In contrast, III11-EDA and EDA-III12 could induce MMP-1 production although they were lesspotent than rEDA (Fig. 9B). These results may suggest thatexposure of either the NH2 or COOH terminus of EDA domainreleases the inducing activity. In agreement with these results,cFN did not promote MMP-1 expression, even at concentrationsup to 300 nM (Fig. 9B). pFN, which does not have an EDAdomain, did not induce MMP-1 expression from the basal leveleither (Fig. 9B), which is consistent with the report of Arneret al. (22).
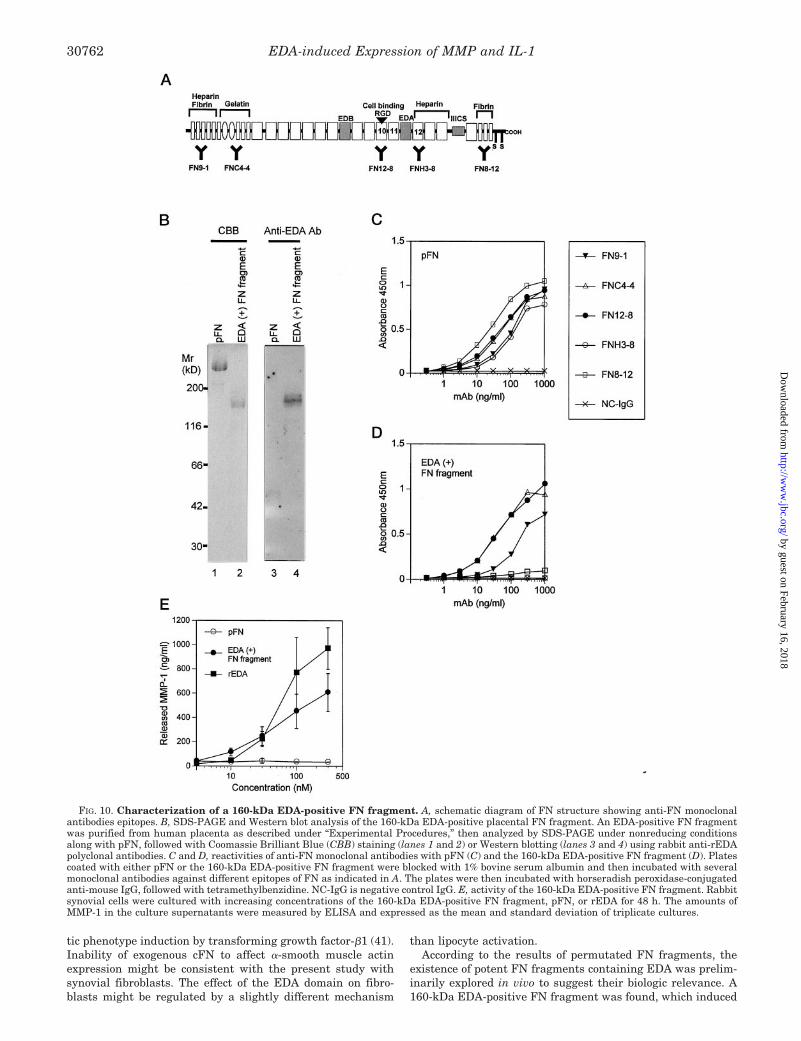
EDA Domain Activities in FN Fragments Purified from Pla-cental Tissue—Purification of FN fragments from human pla-centa was performed to explore the existence of FN fragmentscontaining the EDA domain in vivo. One candidate FN frag-ment was positive in Western blotting using anti-rEDA anti-bodies and migrated to 160 kDa in SDS-PAGE analysis undernonreducing conditions (Fig. 10B). The smaller apparent mo-lecular weight than intact FN suggests the absence of severaldomains. In order to determine which domain was included inthis fragment, ELISA was performed using several anti-FNmAbs that recognize the particular FN regions as shown in Fig.
10A. The results show the 160-kDa EDA-positive FN fragmentwas recognized by FN9-1, FNC4-4, and FN12-8, but not byFNH3-8 and FN8-12 (Fig. 10D), whereas pFN was recognizedby all of these mAbs (Fig. 10C). In addition, protein sequencingfound the NH2-terminal sequence of this fragment was blocked,just like an intact FN molecule. Thus, the epitope mappingsuggests this fragment contains the NH2 terminus through theEDA domain but not the following III12 domain or COOHterminus, which is in agreement with the molecular weightindicated by SDS-PAGE. This fragment induced MMP-1 pro-duction in synovial cells (Fig. 10E). These results may suggestthe existence of a potent EDA-positive FN fragment in vivo.
DISCUSSION
The present study demonstrated that treatment with rEDAprotein facilitates cartilage catabolism and markedly inducesexpression of MMP in chondrocytes and synovial cells. Theinduction of MMP-1 was specific for rEDA among adjacent FNtype III proteins tested. The physical properties of the cartilagematrix, which distribute loads over bone surfaces and provide alow-friction surface over which bones can move, are regulatedthrough the biosynthesis and degradation of extracellular ma-trix by chondrocytes and synovial cells (25). In both healthyand pathologic conditions, the biological functions of synovialcells and chondrocytes are regulated through interactions withthe extracellular matrix molecules, including FN, collagen, andother glycoproteins, and various mediators such as cytokines(22, 26). Indeed, cellular interaction with structural compo-nents of the extracellular matrix influences expression of manyproteases, including MMP-1 induction by type I and III colla-gens (27), urokinase-type plasminogen activator induction bylaminin (28), and MMP-2 induction by vitronectin (29). How-ever, this study is the first demonstration that MMP expres-sion of synovial cells and chondrocytes is regulated by the FNEDA domain. Induction of MMP, which is implicated in biolog-ical processes such as inflammation (30) and angiogenesis (31),may result in matrix degradation and remodeling of extracel-lular matrix structure, since the MMP family degrades extra-cellular matrix proteins (32). Thus, the finding that rEDAinduces MMPs may indicate a role for the EDA domain inregulating functions of chondrocytes and synovial cells in jointconnective tissue.
rEDA treatment causes cell rounding with accompanying
FIG. 7. Cytokine mRNA expression in rabbit synovial cellstreated with rEDA. A, total RNA (20 mg) from rabbit synovial cellsunstimulated or treated with 300 nM rEDA, III11 or III12 for 16 h wasblotted and then hybridized to 32P-labeled cDNAs encoding IL-1a, IL-1b, VEGF, and G3PDH. B, time course of rEDA-induced gene expres-sion. Total RNA (20 mg) from rabbit synovial cells treated with rEDA(300 nM) for 0, 1, 3, 8, or 16 h was blotted and then hybridized to32P-labeled cDNAs encoding IL-1a, IL-1b, VEGF, MMP-1, and G3PDH.
FIG. 8. IL-1RA inhibits rEDA-induced MMP-1 expression. Rab-bit synovial cells were cultured with rEDA (300 nM) alone or in combi-nation with increasing concentrations (3, 10, 30, and 100 ng/ml) ofIL-1RA for 48 h. MMP-1 released into the culture supernatants wasmeasured by ELISA and expressed as the mean and standard deviationof quadruplicate cultures. *, p , 0.005 versus rEDA alone (Student’s ttest).
EDA-induced Expression of MMP and IL-130760
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from
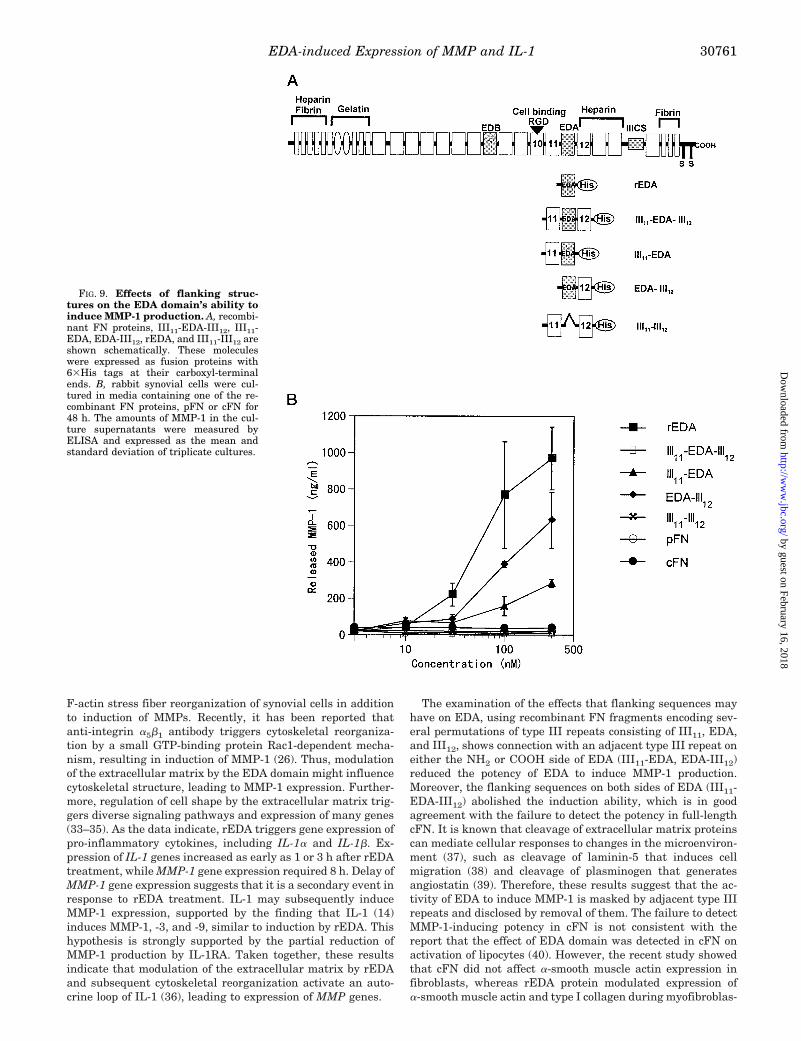
F-actin stress fiber reorganization of synovial cells in additionto induction of MMPs. Recently, it has been reported thatanti-integrin a5b1 antibody triggers cytoskeletal reorganiza-tion by a small GTP-binding protein Rac1-dependent mecha-nism, resulting in induction of MMP-1 (26). Thus, modulationof the extracellular matrix by the EDA domain might influencecytoskeletal structure, leading to MMP-1 expression. Further-more, regulation of cell shape by the extracellular matrix trig-gers diverse signaling pathways and expression of many genes(33–35). As the data indicate, rEDA triggers gene expression ofpro-inflammatory cytokines, including IL-1a and IL-1b. Ex-pression of IL-1 genes increased as early as 1 or 3 h after rEDAtreatment, while MMP-1 gene expression required 8 h. Delay ofMMP-1 gene expression suggests that it is a secondary event inresponse to rEDA treatment. IL-1 may subsequently induceMMP-1 expression, supported by the finding that IL-1 (14)induces MMP-1, -3, and -9, similar to induction by rEDA. Thishypothesis is strongly supported by the partial reduction ofMMP-1 production by IL-1RA. Taken together, these resultsindicate that modulation of the extracellular matrix by rEDAand subsequent cytoskeletal reorganization activate an auto-crine loop of IL-1 (36), leading to expression of MMP genes.
The examination of the effects that flanking sequences mayhave on EDA, using recombinant FN fragments encoding sev-eral permutations of type III repeats consisting of III11, EDA,and III12, shows connection with an adjacent type III repeat oneither the NH2 or COOH side of EDA (III11-EDA, EDA-III12)reduced the potency of EDA to induce MMP-1 production.Moreover, the flanking sequences on both sides of EDA (III11-EDA-III12) abolished the induction ability, which is in goodagreement with the failure to detect the potency in full-lengthcFN. It is known that cleavage of extracellular matrix proteinscan mediate cellular responses to changes in the microenviron-ment (37), such as cleavage of laminin-5 that induces cellmigration (38) and cleavage of plasminogen that generatesangiostatin (39). Therefore, these results suggest that the ac-tivity of EDA to induce MMP-1 is masked by adjacent type IIIrepeats and disclosed by removal of them. The failure to detectMMP-1-inducing potency in cFN is not consistent with thereport that the effect of EDA domain was detected in cFN onactivation of lipocytes (40). However, the recent study showedthat cFN did not affect a-smooth muscle actin expression infibroblasts, whereas rEDA protein modulated expression ofa-smooth muscle actin and type I collagen during myofibroblas-
FIG. 9. Effects of flanking struc-tures on the EDA domain’s ability toinduce MMP-1 production. A, recombi-nant FN proteins, III11-EDA-III12, III11-EDA, EDA-III12, rEDA, and III11-III12 areshown schematically. These moleculeswere expressed as fusion proteins with63His tags at their carboxyl-terminalends. B, rabbit synovial cells were cul-tured in media containing one of the re-combinant FN proteins, pFN or cFN for48 h. The amounts of MMP-1 in the cul-ture supernatants were measured byELISA and expressed as the mean andstandard deviation of triplicate cultures.
EDA-induced Expression of MMP and IL-1 30761
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from
tic phenotype induction by transforming growth factor-b1 (41).Inability of exogenous cFN to affect a-smooth muscle actinexpression might be consistent with the present study withsynovial fibroblasts. The effect of the EDA domain on fibro-blasts might be regulated by a slightly different mechanism
than lipocyte activation.According to the results of permutated FN fragments, the
existence of potent FN fragments containing EDA was prelim-inarily explored in vivo to suggest their biologic relevance. A160-kDa EDA-positive FN fragment was found, which induced
FIG. 10. Characterization of a 160-kDa EDA-positive FN fragment. A, schematic diagram of FN structure showing anti-FN monoclonalantibodies epitopes. B, SDS-PAGE and Western blot analysis of the 160-kDa EDA-positive placental FN fragment. An EDA-positive FN fragmentwas purified from human placenta as described under “Experimental Procedures,” then analyzed by SDS-PAGE under nonreducing conditionsalong with pFN, followed with Coomassie Brilliant Blue (CBB) staining (lanes 1 and 2) or Western blotting (lanes 3 and 4) using rabbit anti-rEDApolyclonal antibodies. C and D, reactivities of anti-FN monoclonal antibodies with pFN (C) and the 160-kDa EDA-positive FN fragment (D). Platescoated with either pFN or the 160-kDa EDA-positive FN fragment were blocked with 1% bovine serum albumin and then incubated with severalmonoclonal antibodies against different epitopes of FN as indicated in A. The plates were then incubated with horseradish peroxidase-conjugatedanti-mouse IgG, followed with tetramethylbenzidine. NC-IgG is negative control IgG. E, activity of the 160-kDa EDA-positive FN fragment. Rabbitsynovial cells were cultured with increasing concentrations of the 160-kDa EDA-positive FN fragment, pFN, or rEDA for 48 h. The amounts ofMMP-1 in the culture supernatants were measured by ELISA and expressed as the mean and standard deviation of triplicate cultures.
EDA-induced Expression of MMP and IL-130762
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from

MMP-1 production. The COOH terminus of the 160-kDa FNfragment was missing just posterior to the EDA domain asdetermined by mAb epitope mapping. Therefore, the exposureof COOH terminus of EDA domain by proteolytic cleavagewould give the 160-kDa EDA-positive FN fragment this po-tency. In synovial fluids of patients with arthritis, EDA-posi-tive FN proteins were detected as fragments weighing 220, 200,180, 170, 110, and 100 kDa (42). Expression of FN containingthe EDA domain is increased in cartilage and synovium ofpatients with arthritis (8, 42). In addition, proteases includingMMPs increase in synovial fluids of patients with arthritis (43).Taken together, increase of the EDA domain and proteasesmay result in generation of the potent FN fragments contain-ing EDA domain, leading to further deterioration of pathogen-esis, by induction of proinflammatory cytokines and subse-quent MMPs.
In summary, treatment with rEDA protein facilitated carti-lage catabolism and markedly induced expression of MMP inchondrocytes and synovial cells. In addition, rEDA inducedcytoskeletal reorganization and expression of IL-1 in synovialcells, leading to subsequent expression of MMPs in an auto-crine manner. This activity of EDA domain was masked byconnection with adjacent type III repeats and revealed by ex-posure of either side of the EDA domain. Consistently, MMP-1production was not induced by full-length cFN, but a 160-kDaEDA-positive fragment did induce MMP-1 production. Thesefindings suggest that the EDA domain in FN fragments trig-gers alterations of cell physiology and plays a role in matrixdegradation in joint connective tissue.
Acknowledgments—We thank Steven E. Johnson for editing themanuscript. We also thank Dr. Jun Takasaki for helpful discussions.
REFERENCES
1. Hynes, R. O. (1990) Fibronectins, Springer-Verlag, New York2. Petersen, T. E., Thogersen, H. C., Skorstengaard, K., Vibe Pedersen, K., Sahl,
P., Sottrup Jensen, L., and Magnusson, S. (1983) Proc. Natl. Acad. Sci.U. S. A. 80, 137–141
3. Pierschbacher, M. D., and Ruoslahti, E. (1984) Nature 309, 30–334. Aota, S., Nomizu, M., and Yamada, K. M. (1994) J. Biol. Chem. 269,
24756–247615. Humphries, M. J., Komoriya, A., Akiyama, S. K., Olden, K., and Yamada,
K. M. (1987) J. Biol. Chem. 262, 6886–68926. Bennett, V. D., Pallante, K. M., and Adams, S. L. (1991) J. Biol. Chem. 266,
5918–59247. Ffrench-Constant, C. (1995) Exp. Cell Res. 221, 261–2718. Hino, K., Shiozawa, S., Kuroki, Y., Ishikawa, H., Shiozawa, K., Sekiguchi, K.,
Hirano, H., Sakashita, E., Miyashita, K., and Chihara, K. (1995) ArthritisRheum. 38, 678–683
9. Ffrench-Constant, C., Van de Water, L., Dvorak, H. F., and Hynes, R. O. (1989)J. Cell Biol. 109, 903–914
10. Kuhn, C., Boldt, J., King, T. E., Jr., Crouch, E., Vartio, T., and McDonald, J. A.(1989) Am. Rev. Respir. Dis. 140, 1693–1703
11. Glukhova, M. A., Frid, M. G., Shekhonin, B. V., Vasilevskaya, T. D., Grunwald,J., Saginati, M., and Koteliansky, V. E. (1989) J. Cell Biol. 109, 357–366
12. Fukuda, M., Levery, S. B., and Hakomori, S. (1982) J. Biol. Chem. 257,6856–6860
13. Yamada, K. M., Schlesinger, D. H., Kennedy, D. W., and Pastan, I. (1977)Biochemistry 16, 5552–5559
14. Saito, S., Katoh, M., Masumoto, M., Matsumoto, S., and Masuho, Y. (1997)J. Biochem. (Tokyo) 122, 49–54
15. Andrews, H. J., Plumpton, T. A., Harper, G. P., and Cawston, T. E. (1992)Agents Actions 37, 147–54
16. Matsumoto, S., Kobayashi, T., Katoh, M., Saito, S., Ikeda, Y., Kobori, M.,Masuho, Y., and Watanabe, T. (1998) Am. J. Pathol. 153, 109–119
17. Matsumoto, S., Katoh, M., Saito, S., Watanabe, T., and Masuho, Y. (1997)Biochim. Biophys. Acta 1354, 159–170
18. Matsumoto, S., Katoh, M., Watanabe, T., and Masuho, Y. (1996) Biochim.Biophys. Acta 1307, 137–139
19. Maniscalco, W. M., Watkins, R. H., Finkelstein, J. N., and Campbell, M. H.(1995) Am. J. Respir. Cell Mol. Biol. 13, 377–386
20. Laine, R. A., Fisher, S. J., and Zhu, B. C. R. (1987) Methods Enzymol. 144,420–429
21. Werb, Z., Tremble, P. M., Behrendtsen, O., Crowley, E., and Damsky, C. H.(1989) J. Cell Biol. 109, 877–889
22. Arner, E. C., and Tortorella, M. D. (1995) Arthritis Rheum. 38, 1304–131423. Zvaifler, N. J., Tsai, V., Alsalameh, S., von Kempis, J., Firestein, G. S., and
Lotz, M. (1997) Am. J. Pathol. 150, 1125–113824. Koch, A. E., Harlow, L. A., Haines, G. K., Amento, E. P., Unemori, E. N., Wong,
W. L., Pope, R. M., and Ferrara, N. (1994) J. Immunol. 152, 4149–415625. Bonassar, L. J., Frank, E. H., Murray, J. C., Paguio, C. G., Moore, V. L., Lark,
M. W., Sandy, J. D., Wu, J. J., Eyre, D. R., and Grodzinsky, A. J. (1995)Arthritis Rheum. 38, 173–183
26. Kheradmand, F., Werner, E., Tremble, P., Symons, M., and Werb, Z. (1998)Science 280, 898–902
27. Shapiro, S. D., Kobayashi, D. K., Pentland, A. P., and Welgus, H. G. (1993)J. Biol. Chem. 268, 8170–8175
28. Khan, K. M. F., and Falcone, D. J. (1997) J. Biol. Chem. 272, 8270–827529. Bafetti, L. M., Young, T. N., Itoh, Y., and Stack, M. S. (1998) J. Biol. Chem.
273, 143–14930. Zhang, Y., McCluskey, K., Fujii, K., and Wahl, L. M. (1998) J. Immunol. 161,
3071–307631. Vu, T. H., Shipley, J. M., Bergers, G., Berger, J. E., Helms, J. A., Hanahan, D.,
Shapiro, S. D., Senior, R. M., and Werb, Z. (1998) Cell 93, 411–42232. Woessner, J. F., Jr. (1991) FASEB J. 5, 2145–215433. Ingber, D. (1991) J. Cell. Biochem. 47, 236–24134. Boudreau, N., Werb, Z., and Bissell, M. J. (1996) Proc. Natl. Acad. Sci. U. S. A.
93, 3509–351335. Jones, P. L., Crack, J., and Rabinovitch, M. (1997) J. Cell Biol. 139, 279–29336. West-Mays, J. A., Strissel, K. J., Sadow, P. M., and Fini, M. E. (1995) Proc.
Natl. Acad. Sci. U. S. A. 92, 6768–677237. Werb, Z. (1997) Cell 91, 439–44238. Giannelli, G., Falk-Marzillier, J., Schiraldi, O., Stetler-Stevenson, W. G., and
Quaranta, V. (1997) Science 277, 225–22839. O’Reilly, M. S., Holmgren, L., Shing, Y., Chen, C., Rosenthal, R. A., Moses, M.,
Lane, W. S., Cao, Y., Sage, E. H., and Folkman, J. (1994) Cell 79, 315–32840. Jarnagin, W. R., Rockey, D. C., Koteliansky, V. E., Wang, S. S., and Bissell, M.
(1994) J. Cell Biol. 127, 2037–204841. Serini, G., Bochaton-Piallat, M. L., Ropraz, P., Geinoz, A., Borsi, L., Zardi, L.,
and Gabbiani, G. (1998) J. Cell Biol. 142, 873–88142. Chevalier, X., Claudepierre, P., Groult, N., Zardi, L., and Hornebeck, W. (1996)
J. Rheumatol. 23, 1022–103043. Walakovits, L. A., Moore, V. L., Bhardwaj, N., Gallick, G. S., and Lark, M. W.
(1992) Arthritis Rheum. 35, 35–42
EDA-induced Expression of MMP and IL-1 30763
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from

Masao Katoh, Seiji Kobayashi and Yasuhiko MasuhoShigeki Saito, Noboru Yamaji, Kunio Yasunaga, Tetsu Saito, Shun-ichiro Matsumoto,
Expression by an Interleukin-1-dependent MechanismThe Fibronectin Extra Domain A Activates Matrix Metalloproteinase Gene
doi: 10.1074/jbc.274.43.307561999, 274:30756-30763.J. Biol. Chem.
http://www.jbc.org/content/274/43/30756Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
http://www.jbc.org/content/274/43/30756.full.html#ref-list-1
This article cites 42 references, 21 of which can be accessed free at
by guest on February 16, 2018http://w
ww
.jbc.org/D
ownloaded from





























