Embed Size (px)
Citation preview

[CANCER RESEARCH 53, 4701-4714, October 1, 1993]
Investigation of the Subcellular Distribution of the bcl-20ncoprotein: Residence in
the Nuclear Envelope, Endoplasmic Reticulum, and Outer Mitochondrial
Membranes I
S t a n i s l a w K r a j e w s k i , 2 S h i g e k i T a n a k a , 2'3 S h i n i c h i T a k a y a m a , M a t t h e w J. Schib ler , W a y n e F e n t o n , a n d J o h n C. R e e d 4
Oncogene and Tumor Suppression Gene Program, La Jolla Cancer Research Foundation, La Jolla, California 92037 [J. C. R.], and Department of Human Genetics, Yale University School of Medicine, New Haven, Connecticut 06510 [~. F.]
ABSTRACT
A multidisciplinary approach was taken to investigate the intracellular locations of the 26-kDa integral membrane protein encoded by the bcl-2 gene. Subcellular fractionation analysis of a t(14;18)-containing lym- phoma cell line revealed the presence of Bcl-2 protein in nuclear, heavy- membrane, and light-membrane fractions but not in cytosol. Sedimenta- tion of heavy-membrane fractions in Nycodenz and Percoll continuous gradients demonstrated comigration of p26-Bcl-2 with mitochondrial but not other organelle-associated proteins. Fractionation of light-membrane fractions using discontinuous sucrose-gradients revealed association of Bcl-2 protein primarily with lighter-density microsomes (smooth endo- plasmic reticulum) as opposed to heavy-density microsomes (rough endo- plasmic reticulum). Immune microscopy studies using laser-scanning mi- croscopy, pre- and postembedding electron microscopic methods, and six different anti-Bcl-2 antibodies demonstrated Bcl-2 immunoreactivity in the nuclear envelope and outer mitochondrial membrane in a patchy distribution. Furthermore, anti-Bci-2 antibody immunoreactivity gener- ally appeared to directly overlie the nuclear envelope in high magnifica- tion electron microscopic studies, reminiscent of nuclear pore complexes. Addition of in vitro translated p26-Bcl-2 to isolated translocation-compe- tent mitochondria revealed transmembrane domain-dependent associa- tion of Bcl-2 protein with mitochondria but provided no evidence for import into a protease-resistant compartment, consistent with immuno- microscopic localization to the outer mitochondrial membrane. Taken together, the findings demonstrate that p26-Bcl-2 resides primarily in the nuclear envelope, endoplasmic reticulum, and outer mitochondrial mem- brane in a nonuniform distribution suggestive of participation in protein complexes perhaps involved in some aspect of transport.
INTRODUCTION
The bcl-2 gene was first discovered because of its involvement in the t(14;18) chromosomal translocations commonly found in non- Hodgkin's lymphomas (1). This cytogenetic abnormality places the bcl-2 gene from chromosome 18 into juxtaposition with the transcrip- tionally active immunoglobulin heavy-chain locus on chromosome 14, resulting in inappropriately high levels of bcl-2 gene expression in malignant B-lymphocytes (reviewed in Ref. 2). The protein encoded by this gene is structurally and functionally unique in that it bears little or no significant homology with other known cellular proteins (3, 4) and because it contributes to malignant cell expansion primarily by prolonging cell survival rather than by increasing the rate of cellular proliferation (5). The specific mechanism by which the Bcl-2 protein extends cell survival remains enigmatic, inasmuch as no biochemical activity has thus far been ascribed to this oncoprotein, but high levels
Received 6/4/93; accepted 7/28/93. The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1 This work was supported by NIH Grant CA-47956, American Cancer Society Grant DHP-71185, and a Scholar Award to J. C. R. from the Leukemia Society of America.
2 S. K. and S. T. contributed equally to the data presented. 3 Present address: Department of Pediatrics, Mie University School of Medicine, Mie,
Japan. 4 To whom requests for reprints should be addressed, at Oncogene and Tumor Sup-
pressor Gene Program, La Jolla Cancer Research Foundation, 10901 N. Torrey Pines Road, La Jolla, CA 92037.
of Bcl-2 protein have been shown to delay or block programmed cell death ("apoptosis") in a variety of circumstances (6-9).
Apoptosis is an active form of cell death that often requires new RNA and protein synthesis (10). One of the hallmarks of programmed cell death is degradation of the nuclear DNA of the cell into oligo- nucleosomal-length fragments, presumably because of the activation of endonucleases that occurs relatively early in the apoptotic process. Although apoptosis plays an important role in normal physiological situations where it helps to ensure that the rate of new cell production in proliferative tissues is offset by a commensurate rate of cell death, it is also important clinically because many chemotherapeutic drugs appear to ultimately kill cancer cells by activating undefined bio- chemical pathways leading to programmed cell death (reviewed in Ref. 11). In this regard, high levels of Bcl-2 protein production have been shown to protect lymphoid cells from death and DNA fragmen- tation induced by a wide variety of antineoplastic drugs (12, 13).
The human Bcl-2 protein is an intracellular, integral membrane protein with a molecular mass of - 2 6 kDa (14). The ability of p26-Bcl-2 to associate with membranes has been attributed to a hy- drophobic stretch of amino acids located near its carboxyl terminus (15). The topic of the intracellular membranes in which p26-Bcl-2 resides, however, is controversial. Previously, Hockenbery et aL (6) reported that Bcl-2 is found primarily in the inner mitochondrial membrane, based on subcellular fractionation experiments and immu- nofluorescence confocal microscopy. In this regard, however, the Bcl-2 protein does not contain a positively charged amphiphilic, a-he- lical NH2-terminal leader sequence typical of many nuclear-encoded proteins that undergo transport into mitochondria (16, 17). In another study, Chen-Levy et aL (14) used crude subcellular fractionation methods and conventional immunofluorescence microscopy and re- ported that Bcl-2 resides mostly in perinuclear membranes, with about one-half of the protein being associated with nuclei and the rest mostly with other intracellular membranes. Bcl-2 is also found primarily in association with the nuclear fraction when expressed in Sf9 insect cells using the baculovirus system (18). Liu et al. (19) examined the issue using two-color immunofluorescence confocal microscopy and reported that about one-half of the immunodetectable Bcl-2 protein is found in association with mitochondria; the remainder of the protein apparently resided in an undefined nonnuclear compartment. Hamil- ton et al. (20) reached the same conclusion using immunofluorescence confocal microscopy but relied on only single-color methods. Re- cently, Jacobson et al. (21) obtained evidence from 2-color immuno- confocal microscopy suggesting that Bcl-2 protein can enter the mem- branes of the endoplasmic reticulum, nuclear envelope, and mitochondria when overexpressed in fibroblast-like cells. In a sepa- rate study involving primarily immunoelectromicroscopic methods, Monaghan et al. (22) reported that Bcl-2 resides in the nuclear enve- lope and outer mitochondrial membranes.
Taken together, the currently available data concerning the intra- cellular locations of the Bcl-2 protein paint a confusing picture. There are several potential reasons for the disparate results obtained by various groups. For example, in most of these reports the data were based upon observations obtained with a single antibody reagent or
4701
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
with ant ibodies directed against a single region of the Bcl-2 protein.
Ideally, one wou ld like to see equivalent results obtained using at least
two different ant ibodies that recognize distinct nonover l app ing epit-
opes on the Bcl-2 protein. Another potential factor of course is the
particular exper imenta l me thods employed , since var ious techniques
have different l imitat ions and advantages.
In an effort to clarify some of the present controvers ies sur rounding
the subcel lular locat ions of the Bcl-2 oncoprote in , we under took a
mul t id isc ipl inary invest igat ion of this issue. For these studies, we
uti l ized four polyclonal and two monoc lona l ant ibody reagents raised
against synthet ic pept ides cor responding to two nonover lapp ing seg-
ments o f the h u m an Bcl-2 protein. A m o n g the exper imenta l ap-
proaches that were used to examine the locat ion of the Bcl-2 protein
were: (a) i m m u n o m i c r o s c o p y using laser-scanning and electron mi-
c roscopic methods ; (b) subcellular fractionation; (c) studies of Bcl-2
protein interact ion in vitro with intact freshly isolated mi tochondr ia ;
and (d) gene transfer to produce the h u m a n Bcl-2 protein in cellular
backgrounds in which it is not normal ly found. On the basis o f these
invest igat ions, we conc lude that the Bcl-2 protein resides primari ly in
3 intracellular m e m b r a n e compar tments : nuclear envelope; endoplas-
mic re t iculum; and outer mi tochondr ia l membrane . The impl icat ions of these f indings are d iscussed below.
the heavy-membrane fraction (pellet), and this supernatant was then centri- fuged for 60 min at 150,000 • g (SW50.1) to obtain the light-membrane (pellet) and cytosolic (supernatant) fractions. The HM and LM fractions were resuspended in 1% Triton-containing lysis buffer (26) and either analyzed immediately for enzyme activities or frozen at -80~ for subsequent immu- noblot assays.
Separation of Organelles by Density Gradient Centrifugation
Sucrose, Percoll, and Nycodenz gradients were prepared as described pre- viously and used to separate organelles based on buoyant density (28-30) using either the HM fraction or the postnuclear low-speed supernatants derived from 5-10 • 107 cells. Briefly, for sucrose, the postnuclear low-speed supernatants or the HM fraction was resuspended in 0.2-0.3 ml of MES buffer and layered onto 4 ml of a 1-2 M linear sucrose gradient and centrifuged at 110,000 • g (SW50.1) for 3 h at 4~ For Nycodenz, a 9-35% w/v linear gradient was used and centrifugation was at 140,000 • g for 2 h. Percoll gradients were formed in situ by resuspending cell fractions in 30% Percoll (final concentration) containing 0.25 M sucrose, 42 mM KC1, 10 mM 4-(2-hydroxyethyl)-l-pipera- zineethanesulfonic acid (pH 7.4), and 5 mM MgC12 and centrifuging at 48,000 • g for 60 min at 4~ Protease inhibitors were included in all solutions. Sequential fractions of 0.2 ml were collected from the gradients and either assayed immediately for enzyme activities or mixed with an equal volume of 2• Triton lysis buffer (26) and stored at -80~
MATERIALS A N D M E T H O D S
Cells and Culture Methods
Cell lines used for these studies included the t[14;18]-containing B-cell lymphoma lines RSl1846 and SU-DHL-4 (23, 24). These cells were main- tained at 105-106 cells/ml in RPMI 1640 with 10% (v/v) heat-inactivated fetal bovine serum at 37~ in 5% COJ95% air. 32D-NEO and 32D-BCL-2 cells are interleukin 3-dependent murine myeloid cells that had been stably infected with Zip-NEO (negative control) and Zip-BCL-2 (encodes human Bcl-2 pro- tein) recombinant retroviruses. The characteristics and conditions for mainte- nance of these cells in culture have been described elsewhere (25).
Preparation of Cellular Subfractions
Isolation of Microsomal Membranes
Microsomal membranes were prepared from 107 RSl1846 cells by centrifu- gation of the LM fraction on discontinuous sucrose step gradients by the method of Meyer and Dobberstein (31). The material recovered from the 0.8 M--1.23 M and the 1.23 M--1.31 M sucrose interfaces was pooled (smooth ER). The heavier-density microsomes were recovered from the 1.31 M--2.0 M inter- face (rough ER). Microsomal membranes were mixed with 3 volumes of 50 mM Tris (pH 7.5)-5 mM MgCI2 and centrifuged at 105,000 • g for 60 rain. The resulting membrane pellet was solubilized in 1.0% Triton X-100-containing lysis buffer and Bcl-2 protein was immunoprecipitated using polyclonal rabbit antisera (26). Proteins remaining in the lysates after preclearing of p26-Bcl-2 were then evaluated by immunoblot assays using mouse monoclonal antibodies to mitochondrial and ER proteins.
RSl1846 cells (1-10 • 107 cells) were washed twice with ice-cold Dul- becco's phosphate-buffered saline (pH 7.4) (Gibco, Inc.) and resuspended in 0.5-1.5 ml of MES 5 buffer containing protease inhibitors as described previ- ously (26). After homogenization for 10-20 strokes with a Dounce homog- enizer and checking that >99% of cells were lysed based on trypan blue dye uptake, samples were transferred to Eppendorf centrifuge tubes (1 ml maxi- mum/1.5-ml tube) and centrifuged at 500 • g for 5 min in a swinging-bucket rotor at 4~ to pellet nuclei. The nuclear pellet was then resuspended in 0.5-2 ml of 1.6 M sucrose containing either 50 mM Tris-HCl (pH 7.5), 25 mM KCI, and 5 mM MgCI2 according to the method of Blobel and Potter (27) or 17 mM morpholinopropanesulfonic acid (pH 7.4) and 2.5 mM EDTA. After underlay- ering with 1-2 ml of 2.0-2.3 M sucrose containing the same buffer and centrifugation at 150,000 • g for 60 min in a Beckman SW50.1 rotor with or without adaptors for 1.5-ml tubes depending on the volume, the resulting nuclear pellets were again resuspended in 1.6 M sucrose solution and the procedure was repeated prior to lysing nuclei in 0.1-0.3 ml of a 1% Triton X-100-containing buffer that has been shown previously to extract essentially all p26-Bcl-2 from cells (26). Similar results were obtained with both nuclei preparation methods.
The supernatant resulting from the initial low-speed centrifugation was subjected to centrifugation at 10,000 • g (average) for 15 min at 4~ to obtain
5 The abbreviations used are: MES, 17 mM morpholinopropanesulfonic acid (pH 7.4)- 2.5 mM EDTA-250 mM sucrose; HM, heavy-membrane; LM, light-membrane; ER, endo- plasmic reticulum; HRPase, horseradish peroxidase; LDH, lactate dehydrogenase; SDS, sodium dodecyl sulfate; PAGE, polyacrylamide gel electrophoresis; Mab, monoclonal antibody; mPBS, modified PBS (120 mM NaCI-11.5 mM NaH2 PO4-37.3 mM K2HPO4, pH 7.6); TNK-BTM, 0.1 M Tris- (pH 7.6-7.8) -0.55-0.6 M NaCI-I0 mM KC1-2% bovine serum albumin; TNK, 0.1 M Tris (pH 7.6-7.8)-0.55-0.6 M NaCI-10 mM KC1; dH20, deionized water; EM, electron microscopy; IUCR, ubiquinol-cytochrome c reductase; AL, annulate lamellae.
Cellular Uptake and Assay for Horseradish Peroxidase
RSl1846 cells were cultured for 16-18 h at 0.25 • 106 cells/ml in RPMI 1640 containing 10% fetal bovine serum, antibiotics, 1 mM L-glutamine, and 100/xg/ml HRPase to label the lysosomal compartment (32). The cells were then washed 3 times with Dulbecco's phosphate-buffered saline to remove HRPase that had not entered cells by fluid-phase pinocytosis, and the heavy- membrane fraction was prepared for loading onto Nycodenz and Percoll gra- dients. Fractions collected from gradients were then assayed for HRPase ac- tivity by the method of Steinman and Cohn (33), using o-dianisidin as the substrate and measuring the absorbance at 450 nm after 3 rain.
Enzyme Assays for LDH,/]-Hexosaminidase, and Galactosyltransferase
The lysosomal marker /3-hexosaminidase was assayed by measuring the hydrolysis ofp-nitrophenyl-/3-N-acetylglucosamine to yield free p-nitrophenol (34). The Golgi marker UDP-galactose:N-acetylglucosamine-galactosyltrans- ferase was assayed by the method of Morr6 (35) using UDP-o-[laC]galactose from Amersham, Inc. (CFB129) and measuring by liquid scintillation counting the amount of [14C]lactose generated in 20 or 30 min at room temperature, where unhydrolyzed substrate was removed on an anion exchange resin (AG1- X2/CI- form; Bio-Rad). LDH activity was measured using a kit from Sigma (No. 500). Data from enzyme assays were normalized for total protein content where indicated.
Antibodies, Immunobiot, and Immunoprecipitation Assays
Polyclonal rabbit antisera specific for the human Bcl-2 protein were raised against synthetic peptides corresponding to amino acids 41-54 or 61-76 of p26-Bcl-2 and were used for immunoprecipitations under conditions that re- sulted in recovery of essentially all immunodetectable Bcl-2 protein from cell
4702
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
lysates (26). In some cases, 5 jxg of competing peptide were added to lysates prior to antibody, to block binding to p26-Bcl-2. Samples were normalized for total protein content using the bicinchoninic acid method (36) where indicated. Antibodies used for immunoblot assays included the polyclonal rabbit antisera 9716-10 [anti-Bcl-2(41-54) used at 0.1% v/v]; 9718-8 [anti-Bcl-2(61-76), 0.1%], anti-Fl-13-ATPase [0.05%] (37); anti-2H1 which identifies an unchar- acterized 60-kDa endoplasmic reticulum protein (0.025%) (38); anti-mannose 6-phosphate receptor (0.025%) (39). Mouse monoclonal antibodies included 7H8.2C12, specific for cytochrome c (ascites at 0.1%) (40), and an antibody specific for the p70-o~-docking protein of the rough ER (41). SDS-PAGE and electroblotting to nitrocellulose filters were performed as described previously (26, 42), except when assessing mannose 6-phosphate receptor, in which case proteins were not reduced to preserve immunoreactivity. Antibodies were detected on filters using 0.25 /xCi/ml ~25I-protein. A (Amersham No. IM144) directly or after first incubating blots with 1/xg/ml rabbit anti-mouse IgG when monoclonal antibodies were used. In some cases, detection of rabbit anti-Bcl-2 antibodies was accomplished with biotinylated goat anti-rabbit IgG serum followed by horseradish peroxidase-avidin-biotin complex reagent (Vector Laboratories, Inc.) and 3-amino-9-ethylcarbazole; and detection of mouse monoclonals was achieved with alkaline phosphatase-conjugated rabbit anti- mouse IgG serum (Dako, Inc.) followed by the chemiluminescent substrate Lumiphos (Bohringer-Mannheim, Inc.) according to the manufacturer's rec- ommendations.
In Vitro Transcription, Translation, and Mitochondriai Transport Assays
Bcl-2~ and Bcl-2/3 complementary DNAs, cloned into the pBluescript (Stratagene, Inc.) vector pSK-II (43), were transcribed in vitro using T3 or T7 RNA polymerase, respectively, and a Riboprobe kit from Promega according to the supplier's instructions. Approximately 2/xg of the resulting RNAs were used without capping for in vitro translation with a rabbit reticulocyte lysate translation kit (Promega) supplemented with [35S]methionine according to the supplier's protocol. The translation products were used directly in the mito- chondrial transport experiments.
Mitochondria were prepared from the livers of male Sprague-Dawley rats according to the procedure of Conboy and Rosenberg (44). They were sus- pended at a protein concentration of 20 mg/ml in 220 mM mannitol-70 mM sucrose-2 mM 4-(2-hydroxyethyl)-l-piperazineethanesulfonic acid (pH 7.4; 4~ for use in transport experiments. Mitochondrial transport was assessed as described previously (45), except that 3 volumes of translated protein mixture were combined with 2 volumes of mitochondria. After a 20-min incubation at 27~ the mitochondria were recovered by a 3-min centrifugation in an Ep- pendorf microcentrifuge. The mitochondria were resuspended in an equal volume of isolation buffer (above), and an aliquot was treated with trypsin at a final concentration of 10 /xg/ml for 20 min at 4~ An aliquot of the supernatant was treated with the same concentration of trypsin. The reactions were terminated with SDS-PAGE loading buffer, the samples were heated at 95~ for 5 min, and the proteins were separated on a SDS-10% polyacrylamide gel. The gels were fixed, treated with Autofluor (National Diagnostics), and exposed to XAR-5 film at -70~ Similar results were obtained when import assays were performed at 37~ and for longer times (up to 60 min), except that in this case translation of intramitochondrial RNAs occurred, resulting in the production of additional 35S-labeled proteins the appearance of which could be blocked by addition of chloramphenicol.
Microscopy
Immune staining of cells was accomplished using a variety of anti-Bcl-2 antibody reagents, including: (a) two different rabbit antisera against the 41-54 peptide described above (9716-10, 1631-8); (b) two antisera raised against the 61-76 peptide (9718-8; 1633-8); (c) Mab 124, a murine monoclonal antibody against the 41-54 peptide (purchased from DAKO, Inc.); and (d) ascites from the hybridoma 4D7 which produces a monoclonal antibody directed against the 61-76 peptide (46). All data presented were reproduced with a minimum of 2 different antibody reagents directed against nonoverlapping epitopes on Bcl-2.
Laser-scanning Microscopy. Immunostaining of cells for evaluation by laser-scanning microscopy was accomplished by both pre- and postembedding methods. In the postembedding method, cells were pelleted by centrifugation and resuspended in fixative, either 4% neutral-buffered formalin or Bouin's solution for 5 min, and then washed twice for 5 min each with gentle agitation
in mPBS. Fixed cells were then resuspended in 1.5% agarose in mPBS and embedded in paraffin. Sections 5 ~m thick were mounted on poly-c-lysine- coated slides and deparaffinized by heating at 90~ followed by xylene ex- traction. After rinsing in absolute ethanol for 15 s, endogenous peroxidase activity was quenched by incubation for 30-45 min in 2% HzO2 in methanol. After sequentially washing in absolute ethanol (20 s), 70% ethanol (20 s), dH20 (1 min) and mPBS for 5 rain, cells were incubated for 2 h in 20 mM sodium citrate (pH 3.2). Cells were washed for 5 min in mPBS and then preblocked for 2-3 h using 1% goat serum in TNK-BTM. Slides were then incubated overnight with various anti-Bcl-2 antibodies at optimal concentra- tions in TNK-BTM [usually 1:2000 (v/v) for polyclonal antisera], washed 3 time for 5 min each in mPBS, and incubated for 1 h with appropriate biotin- ylated secondary antibodies [either 2.8/~g/ml goat anti-rabbit IgG or 2.0 txg/ml goat anti-mouse IgG (Vector Laboratories, Inc.)] also in TNK-BTM. After 2 washes for 5 min each in mPBS, slides were incubated for 45 min with HRPase-avidin-biotin complex reagent (prepared according to the manufac- turer's protocol; Vector Laboratories, Inc.) in TNK. Following 2 washes for 5 min each in mPBS, slides were treated for 10 min with TNK containing 10 mM imidazole, 0.01% H202, and 0.5 mg/ml 3,3'-diaminobenzidine, with or without methyl green counterstaining. The slides were washed in water and then dehydrated by successive incubations in graded solutions of acetone and xy- lenes before mounting with cover slips and Acrytol (Surgipath, Inc.).
For the preembedding method, cells were fixed in 2% glutaraldehyde in mPBS for 5 min and then washed in mPBS, and treated with 0.01% H202 in mPBS for 15 min. After 2 washes for 5 rain each in phosphate-buffered saline, cells were preblocked and immunostained in suspension as described above, except that most of the incubations with antibodies and washes were performed at 37~ with agitation, and the concentration of the anti-Bcl-2 antibodies was: 1:1500-1:2000 (v/v) for 9716-10 and 9718-8; 1:800-1:1000 (v/v) for 1631-8 and 1633-8; 1:300-1:500 (v/v) for 4D7 ascites; and 1:150-1:200 for Mab 124 supernatant. The stained cells were then immobilized in 1% agarose/mPBS, postfixed with either 1% OsO4 in phosphate buffer (81 mM NazHPO4 and 16 mM NaHzPO4, pH 7.3-7.4) for i h or in a solution containing 1% OsO4, 1.25% K2Cr207, and 150 mM NaC1 at pH 7.4 for 0.5 h before dehydrating in a series of alcohols and embedding in propylene oxide:plastic resin [25% (w/v) TAAB, 25% Epon 812, 30% dodecenylsuccinic anhydride, 20% nadic methyl anhy- dride, and 1.5% (v/v) DMP-30 (1:1)]. Sections 1 /~m thick were mounted on poly-L-lysine-coated slides.
Laser-scanning microscopy was performed using a Zeiss inverted LSM-410 laser-scanning confocal microscope equipped with an argon ion laser (488 nm) and transmitted bright-field mode, similar to the method of Itoh et al. (47). A Planapochromat (X63) objective lens with a numerical aperture of 1.4 was used. The images were derived through electronic processing methods, which enhance the positive reaction signals by a combination of contrast normaliza- tion and scaling density adjustments. Final images were printed or photo- graphed using a Sony UP-D7000 Digital Color Printer or a Lasergraphics LFR Mark II film graphics recorder. These data were representative of findings obtained by routine light microscopy with regard to relative signal intensities.
Electron Microscopy. For preembedding EM studies, cells were prepared as exactly described above. For postembedding EM studies, cells were fixed in glutaraldehyde as above, immobilized in 1.5% agarose, postfixed in 1% OsO4, and washed extensively with water prior to incubating for 1 h in 1% uranyl acetate on ice in the dark. Cells in agarose were then dehydrated by sequential 0.5-1 h incubations in 30, 50, 70, 80, 96, and 100% ethanol followed by three 5-min incubations in propylene oxide, several h in propylene oxide:plastic resin (1:1), and finally overnight in resin alone. After embedding in plastic resin, ultrathin sections (200--400 A,) were mounted on 200 mesh nickel grids. In some cases, sections for postembedding EM were treated for 0.5-1 h with either 10% H202 in mPBS or a saturated solution of m-periodate in dH20. Preblocking, primary, and secondary antibody incubations were performed as described above, with the exception that the concentrations of anti-Bcl-2 antibodies used were: 0.1-0.2% for rabbit antisera; 0.3% for 4D7 ascites; and 1% for Mab 124 supernatant. After 1 h of incubation with biotinylated sec- ondary antibodies, the epitopes were detected using 1% streptavidin/15 nM colloidal gold conjugate (American Qualex, Inc.) in either TNK-BTM buffer adjusted to pH 6.6-6.8 or in 2.5 mM acetic acid (1.75 mM Na § (pH 5.5). After incubation in a solution containing colloidal gold for 3 h at 37~ 4 washes in mPBS, and 3 washes in dH20, the grids were imaged and photographed using a Hitachi 600 electron microscope.
4703
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
RESULTS
Subcellular Fractionation Analysis of Bci-2 Protein Distribution
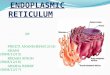
Evidence for Association of p26-Bcl-2 with Mitochondria. The RSl1846 cell line contains a t[14;18] translocation involving bcl-2 and was established from a patient with non-Hodgkin's B-cell lym- phoma. Crude fractionation of these cells to yield nuclei, heavy mem- branes, light membranes, and cytosol revealed p26-Bcl-2 protein pri- marily in the nuclear and HM fractions, although a small amount of 26-kDa material reacting with anti-Bcl-2 antibodies was detected also in light membranes during these immunoblot assays where subcellular fractions were normalized for total protein content (Fig. 1). Similar results were obtained using two different anti-Bcl-2 antisera directed against nonoverlapping epitopes (not shown). The presence of Bcl-2 protein in the nuclear compartment could not be attributable to re- sidual nonlysed cells, based on LDH measurements, and persisted despite centrifugation of nuclei twice through 2.0-2.3 M sucrose. Despite attempts at rigorous purification, however, nuclear prepara- tions from these malignant B-lymphocytes frequently appeared to be contaminated with mitochondria, since p50-Fl-/3-ATPase and p12- cytochrome c were also detected by immunoblotting using specific antibodies (Fig. 1). However, we often observed disproportionate amounts of p26-Bcl-2 in the nuclear fraction relative to mitochondrial markers such as F~-/3-ATPase and cytochrome c (Fig. 1), suggesting that mitochondrial contamination cannot entirely account for the pool of Bcl-2 found in this fraction.
N HM LM C
pso--~ - - 50 kD F1 -[3-ATPase
p26--I~ Bcl-2
--33 - - 2 8
- - 1 9
p12 ..-.1~ cyto-c
Fig. 1. Subcellular distribution of p26-Bcl-2 in RS 11846 lymphoma cells. Nucleus (N), HM, LM, and cytosolic (C) fractions were prepared from RSl1846 cells and resuspended in lysis buffer containing 1% Triton X-100 (26). Detergent lysates were normalized for protein content (100/xg/lane) and subjected to SDS-PAGE using 12% gels. An aliquot of each fraction was also assayed for LDH activity. Proteins in gels were transferred to nitrocellulose filters which were cut into sections for immunoblot assays using antibodies specific for p50-F1-/3-ATPase, p26-Bcl-2, and pl2-cytochrome c. The relative positions of prestained molecular weight markers (Bio-Rad, Inc.) on blots are shown in kilodaltons (Uga).
Because the HM fraction typically contains not only mitochondria but also lysosomes, Golgi, and some of the rough endoplasmic re- ticulum, we next subjected HM material obtained from RSl1846 cells to centrifugation in continuous Nycodenz and Percoll gradients to achieve separation of various organelles based on differences in their buoyant density. Analysis of fractions recovered from these gradients was accomplished by either immunoblotting [using antibodies spe- cific for p26-Bcl-2, mitochondrial p50-F1-/3-ATPase, and p12-cyto- chrome c or an uncharacterized 60-kDa protein found only in the ER (antibody 2H1)] or by performing standard enzymatic assays for ly- sosomal/3-hexosaminidase and Golgi galactosyltransferase. In some experiments, RSl1846 cells were cultured with HRPase to allow pinocytotic marking of the lysosomal compartment (32).
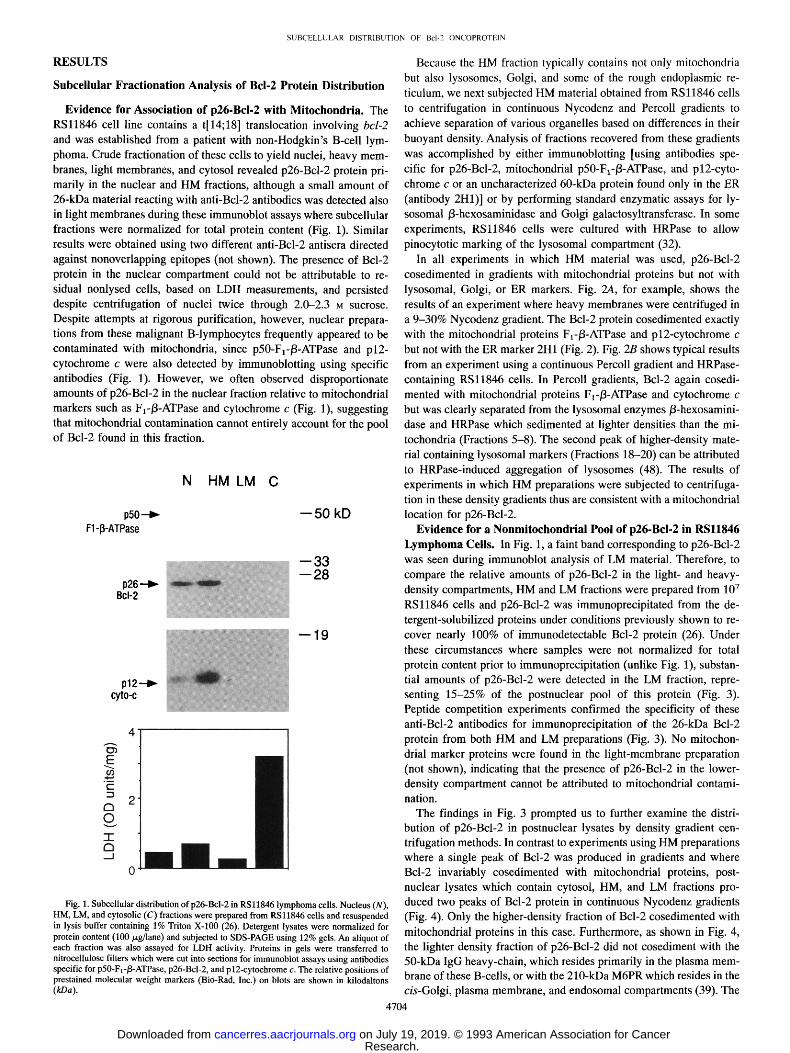
In all experiments in which HM material was used, p26-Bcl-2 cosedimented in gradients with mitochondrial proteins but not with lysosomal, Golgi, or ER markers. Fig. 2A, for example, shows the results of an experiment where heavy membranes were centrifuged in a 9-30% Nycodenz gradient. The Bcl-2 protein cosedimented exactly with the mitochondrial proteins F~-I3-ATPase and p12-cytochrome c but not with the ER marker 2H1 (Fig. 2). Fig. 2B shows typical results from an experiment using a continuous Percoll gradient and HRPase- containing RSl1846 cells. In Percoll gradients, Bcl-2 again cosedi- mented with mitochondrial proteins F1-/3-ATPase and cytochrome c but was clearly separated from the lysosomal enzymes/3-hexosamini- dase and HRPase which sedimented at lighter densities than the mi- tochondria (Fractions 5-8). The second peak of higher-density mate- rial containing lysosomal markers (Fractions 18-20) can be attributed to HRPase-induced aggregation of lysosomes (48). The results of experiments in which HM preparations were subjected to centrifuga- tion in these density gradients thus are consistent with a mitochondrial location for p26-Bcl-2.
Evidence for a Nonmitochondrial Pool of p26-Bcl-2 in RSl1846 Lymphoma Cells. In Fig. 1, a faint band corresponding to p26-Bcl-2 was seen during immunoblot analysis of LM material. Therefore, to compare the relative amounts of p26-Bcl-2 in the light- and heavy- density compartments, HM and LM fractions were prepared from 10 7
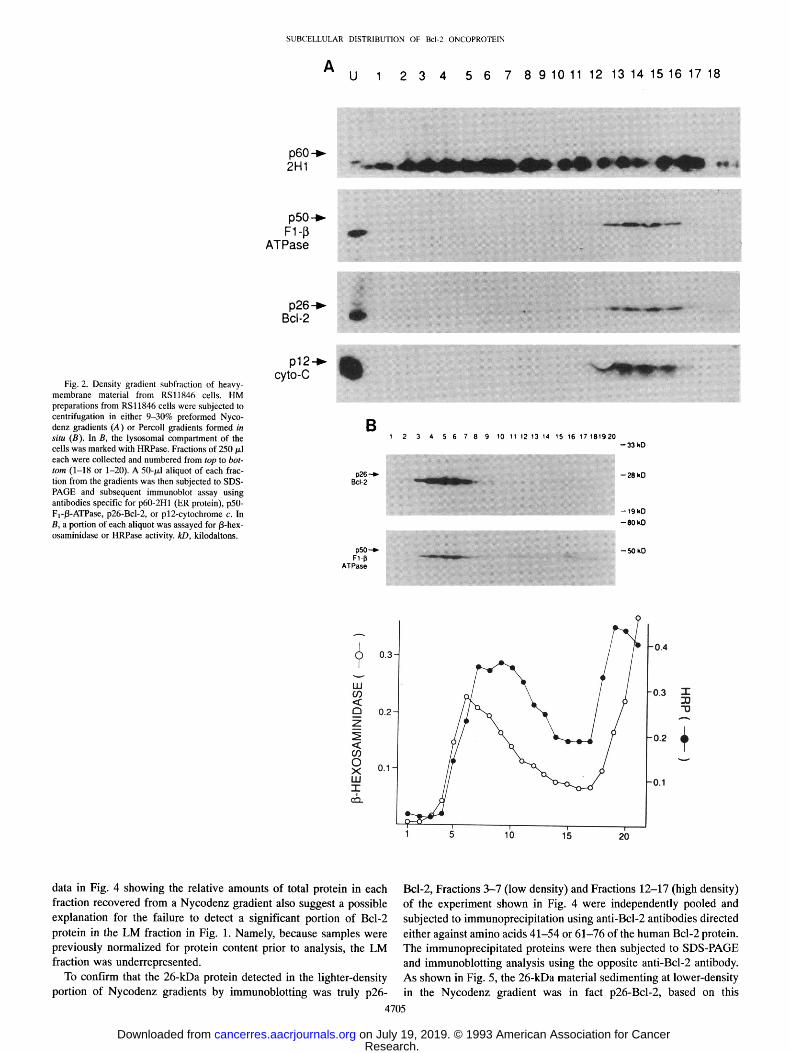
RSl1846 cells and p26-Bcl-2 was immunoprecipitated from the de- tergent-solubilized proteins under conditions previously shown to re- cover nearly 100% of immunodetectable Bcl-2 protein (26). Under these circumstances where samples were not normalized for total protein content prior to immunoprecipitation (unlike Fig. 1), substan- tial amounts of p26-Bel-2 were detected in the LM fraction, repre- senting 15-25% of the postnuclear pool of this protein (Fig. 3). Peptide competition experiments confirmed the specificity of these anti-Bcl-2 antibodies for immunoprecipitation of the 26-kDa Bcl-2 protein from both HM and LM preparations (Fig. 3). No mitochon- drial marker proteins were found in the light-membrane preparation (not shown), indicating that the presence of p26-Bcl-2 in the lower- density compartment cannot be attributed to mitochondrial contami- nation.
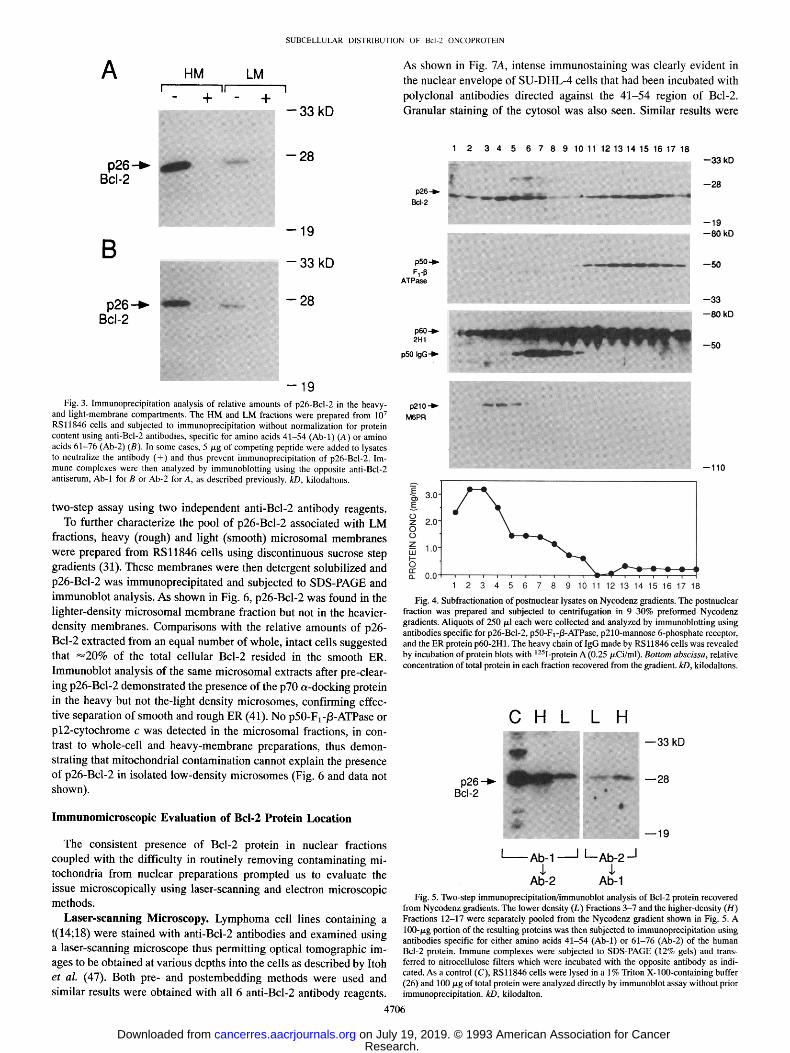
The findings in Fig. 3 prompted us to further examine the distri- bution of p26-Bcl-2 in postnuclear lysates by density gradient cen- trifugation methods. In contrast to experiments using HM preparations where a single peak of Bcl-2 was produced in gradients and where Bcl-2 invariably cosedimented with mitochondrial proteins, post- nuclear lysates which contain cytosol, HM, and LM fractions pro- duced two peaks of Bcl-2 protein in continuous Nycodenz gradients (Fig. 4). Only the higher-density fraction of Bcl-2 cosedimented with mitochondrial proteins in this case. Furthermore, as shown in Fig. 4, the lighter density fraction of p26-Bcl-2 did not cosediment with the 50-kDa IgG heavy-chain, which resides primarily in the plasma mem- brane of these B-cells, or with the 210-kDa M6PR which resides in the cis-Golgi, plasma membrane, and endosomal compartments (39). The
4704
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
A U 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
p60 -I~ 2H1
p50-1~ Fl-p
ATPase
p26-~ Bcl-2
Fig. 2. Density gradient subfraction of heavy- membrane material from RSl1846 cells. HM preparations from RS11846 cells were subjected to centrifugation in either 9-30% preformed Nyco- denz gradients (A) or Percoll gradients formed in situ (B). In B, the lysosomal compartment of the cells was marked with HRPase. Fractions of 250/~1 each were collected and numbered from top to bot- tom (1-18 or 1-20). A 50-/zl aliquot of each frac- tion from the gradients was then subjected to SDS- PAGE and subsequent immunoblot assay using antibodies specific for p60-2H1 (ER protein), p50- FI-/3-ATPase, p26-Bcl-2, or pl2-cytochrome c. In B, a portion of each aliquot was assayed for/3-hex- osaminidase or HRPase activity, kD, kilodaltons.
p12--~- cyto-C
B
p 2 6 - -~ Bcl-2
p 5 0 - -~ F1-I]'
ATPase
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 181920 - - 33 k D
- - 28 kD
- 19 kD
- - 8 0 kO
- -50 kO
data in Fig. 4 showing the relative amounts of total protein in each fraction recovered from a Nycodenz gradient also suggest a possible explanation for the failure to detect a significant portion of Bcl-2
protein in the LM fraction in Fig. 1. Namely, because samples were previously normalized for protein content prior to analysis, the LM fraction was underrepresented.
To confirm that the 26-kDa protein detected in the lighter-density portion of Nycodenz gradients by immunoblotting was truly p26-
Bcl-2, Fractions 3-7 (low density) and Fractions 12-17 (high density) of the experiment shown in Fig. 4 were independently pooled and subjected to immunoprecipitation using anti-Bcl-2 antibodies directed
either against amino acids 41-54 or 61-76 of the human Bcl-2 protein. The immunoprecipitated proteins were then subjected to SDS-PAGE and immunoblotting analysis using the opposite anti-Bcl-2 antibody. As shown in Fig. 5, the 26-kDa material sedimenting at lower-density in the Nycodenz gradient was in fact p26-Bcl-2, based on this
4705
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
S U B C E L L U L A R D I S T R I B U T I O N O F B c l - 2 0 N C O P R O T E I N
A
p26. -~ Bcl-2
B
HM LM 11
- +
: r
+ I
- - 3 3 k D
- - 2 8
- -19
- 3 3 k D
p26 B c l - 2
................ :i!i?i'~
- 2 8
i!ii~!ii~i~i:; �84 ! - 1 9
Fig. 3. Immunoprecipitation analysis of relative amounts of p26-Bcl-2 in the heavy- and light-membrane compartments. The HM and LM fractions were prepared from 10 7
RSl1846 cells and subjected to immunoprecipitation without normalization for protein content using anti-Bcl-2 antibodies, specific for amino acids 41-54 (Ab-1) (A) or amino acids 61-76 (Ab-2) (B). In some cases, 5/xg of competing peptide were added to lysates to neutralize the antibody (+) and thus prevent immunoprecipitation of p26-Bcl-2. Im- mune complexes were then analyzed by immunoblotting using the opposite anti-Bcl-2 antiserum, Ab-1 for B or Ab-2 for A, as described previously, kD, kilodaltons.
two-step assay using two independent anti-Bcl-2 antibody reagents. To further characterize the pool of p26-Bcl-2 associated with LM
fractions, heavy (rough) and light (smooth) microsomal membranes were prepared from RSl1846 cells using discontinuous sucrose step gradients (31). These membranes were then detergent solubilized and p26-Bcl-2 was immunoprecipitated and subjected to SDS-PAGE and immunoblot analysis. As shown in Fig. 6, p26-Bcl-2 was found in the lighter-density microsomal membrane fraction but not in the heavier- density membranes. Comparisons with the relative amounts of p26- Bcl-2 extracted from an equal number of whole, intact cells suggested that ~--20% of the total cellular Bcl-2 resided in the smooth ER. Immunoblot analysis of the same microsomal extracts after pre-clear- ing p26-Bcl-2 demonstrated the presence of the p70 a-docking protein in the heavy but not the-light density microsomes, confirming effec- tive separation of smooth and rough ER (41). No p50-F1-/3-ATPase or pl2-cytochrome c was detected in the microsomal fractions, in con- trast to whole-cell and heavy-membrane preparations, thus demon- strating that mitochondrial contamination cannot explain the presence of p26-Bcl-2 in isolated low-density microsomes (Fig. 6 and data not shown).
l m m u n o m i c r o s c o p i c Evaluat ion o f Bcl-2 Protein Locat ion
The cons is ten t p resence o f Bcl -2 protein in nuc lear f rac t ions
coup led wi th the d i f f icu l ty in rou t ine ly r e m o v i n g c o n t a m i n a t i n g mi-
tochondr ia f rom nuc lear p repara t ions p rompted us to eva lua te the
issue mic roscop ica l ly us ing lase r - scanning and e lect ron mic roscop ic
methods .
Laser-scanning Microscopy. L y m p h o m a cell l ines con ta in ing a
t (14;18) were s ta ined wi th ant i -Bcl-2 ant ibodies and e x a m i n e d us ing
a l ase r - scanning mic roscope thus pe rmi t t ing opt ical t o m o g r a p h i c im-
ages to be ob ta ined at va r ious depths into the cells as descr ibed by I toh
et al. (47). Both pre- and p o s t e m b e d d i n g m e t h o d s were used and
s imi lar resul ts were ob ta ined wi th all 6 ant i -Bcl-2 an t ibody reagents .
As shown in Fig. 7A, intense immunostaining was clearly evident in the nuclear envelope of SU-DHL-4 cells that had been incubated with polyclonal antibodies directed against the 41-54 region of Bcl-2. Granular staining of the cytosol was also seen. Similar results were
p26 - I~
Bcl-2
p50 F 1 -~
ATPase
p60.-~. 2H1
p50 tgG'-~
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
- - 3 3 kD
~ a t ~' - - 2 8
--19 - - 8 0 kD
,,.,., : : : - - : - ~" , . . . . . 50
--33
- - 8 0 kD
--50
p210"1~ M6PR
E 3.0'
0 ~ 2.0" 0 7 ,'n 1.0
~ 0.0
--110
_ 7 ,
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
Fig. 4. Subfractionation of postnuclear lysates on Nycodenz gradients. The postnuclear fraction was prepared and subjected to centrifugation in 9-30% preformed Nycodenz gradients. Aliquots of 250 p,1 each were collected and analyzed by immunoblotting using antibodies specific for p26-Bcl-2, p50-F1-/3-ATPase, p210-mannose 6-phosphate receptor, and the ER protein p60-2H1. The heavy chain of IgG made by RSl1846 cells was revealed by incubation of protein blots with 125I-protein A (0.25 p, Ci/ml). Bottom abscissa, relative concentration of total protein in each fraction recovered from the gradient, kD, kilodaltons.
C H L L H ~ ~ 3 3 kD
p 2 6 . - ~ i,
I Ab- 1 I L_ Ab-2 / $ $
Ab-2 Ab-1 Fig. 5. Two-step immunoprecipitation/immunoblot analysis of Bcl-2 protein recovered
from Nycodenz gradients. The lower density (L) Fractions 3-7 and the higher-density (H) Fractions 12-17 were separately pooled from the Nycodenz gradient shown in Fig. 5. A 100-/xg portion of the resulting proteins was then subjected to immunoprecipitation using antibodies specific for either amino acids 41-54 (Ab-1) or 61-76 (Ab-2) of the human Bcl-2 protein. Immune complexes were subjected to SDS-PAGE (12% gels) and trans- ferred to nitrocellulose filters which were incubated with the opposite antibody as indi- cated. As a control (C), RSl1846 cells were lysed in a 1% Triton X-100-containing buffer (26) and 100/xg of total protein were analyzed directly by immunoblot assay without prior immunoprecipitation, kD, kilodalton.
4706
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
gg,% g ggg , --11~a-
50 ..I-
i -
- - 2 8 + ~ ~ ~1- p l 2-cyto-C p26.-~-
J Bcl-2 - - 19
A B Fig. 6. Association of p26-Bcl-2 with microsomal membranes. In A, microsomal mem-
branes were prepared from the LM fraction of 107 RSl1846 cells (27). The HM and nuclear fraction (N) membranes were also prepared from 107 cells. Membranes were solubilized in 1% Triton X-100 lysis buffer and Bcl-2 was immunoprecipitated using Ab-1. The immune complexes were subjected to SDS-PAGE and the resulting blots were incubated anti-Bcl-2 antibody (Ab-2) for detection of p26-Bcl-2 using biotinylated goat anti-rabbit lgG serum followed by HRPase-avidin-biotin complex reagent and then 3-amino-9-ethylcarbazole for color development. Comparisons were made with extracts prepared from 107 whole, intact cells. The ribosomal (Ribo) pellet resulting from prepa- ration of microsomes was included as a negative control. In B, aliquots of the same lysates precleared of Bcl-2 protein were analyzed by immunoblotting for relative levels of pl2-cytochrome c, and p70-ot-docking protein (DP). Mouse monoclonal antibodies were detected on filters by incubation with alkaline phosphatase-conjugated rabbit anti-mouse IgG serum followed by Lumiphos. The blot was then exposed to X-ray film for 10-20 min to optimize independently the exposures for the p12 and p70 proteins. MW, molecular weight; kD, kilodaltons; SER, smooth ER; RER, rough ER.
obtained for SU-DHL-4 and other lymphoma cell lines using other anti-Bcl-2 antibodies. Fig. 7D, for example, shows the results obtained when SU-DHL-4 cells were stained using a monoclonal anti-Bcl-2 antibody directed against the 61-76 region of Bcl-2. Interestingly, in some lymphoma cell lines such as RSl1846 that contain large mito- chondria (confirmed by EM), intense anti-Bcl-2 antibody immuno- staining was observed on the surface of cytoplasmic organelles re- sembling mitochondria, when cross-sectional images were obtained (Fig. 7B). Preabsorption of anti-Bcl-2 antibodies with competing pep- tide markedly diminished the immunoreactivity in all areas of the cells (Figs. 7, C and E), but residual areas of nuclear reactivity suggested that this represented nonspecific staining.
The specificity of the anti-human Bcl-2 antibody reactivity with the nuclear envelope was further confirmed by immunocytochemical analysis of a pair of murine cell lines, 32D-NEO and 32D-BCL-2, that had been stably transfected with control or human Bcl-2-producing constructs, thus exploiting the fact that none of our anti-Bcl-2 anti- body reagents cross-reacts with the mouse Bcl-2 protein (26, 42, 49). When 32D-BCL-2 cells were subjected to the immunostaining pro- cedure, reactivity with the nuclear envelope was readily detected (Fig. 8A). Furthermore, in some cells where the plane of sectioning oc- curred more tangentially, the immunostaining of the nuclear envelope appeared as two closely spaced concentric circles around the nucleus, suggestive of inner and outer nuclear membranes (Fig. 8B). In con- trast, 32D-NEO cells which harbor a control virus lacking human bcl-2 sequences displayed no immunoreactivity (Fig. 8C), thus dem- onstrating the specificity of the results with anti-Bcl-2 antibodies. As in the t(14;18)-containing lymphomas described above, some images of 32D-BCL-2 cells revealed circumferential immunostaining of cy- tosolic organelles (Fig. 8, D and E), suggestive of mitochondria. Again, the specificity of these results was confirmed by peptide com- petition (Fig. 8F).
Electron Microscopy (EM). The presence of Bcl-2 in the nuclear envelope of lymphoma cells was also confirmed at the EM level. For these experiments, immunostaining was performed by both preem- bedding and postembedding methods to minimize risks of artifactual immunolocalization results. Fig. 9 shows some typical results ob- tained by the preembedding method where anti-Bcl-2 antibody detec- tion was accomplished using HRP/3,3'-diaminobenzidine. At the
higher magnification afforded by EM, it was appreciated that the immunostaining of the nuclear envelope occurred in a patchy, non- uniform distribution. Furthermore, the immunoreactivity appeared to directly overlie the nuclear envelope in many cases (Fig. 9, D and E), reminiscent of nuclear pore complexes. In some views, membrane structures resembling tubes were evident in the cytosol of cells, par- ticularly towards the periphery of the cell (see Fig. 9, C and D). In these membranes structures, anti-Bcl-2 immunostaining again oc- curred in discrete patches and appeared to span the lumen between these membranes (Fig. 9D). In fact, in some cells these membrane structures occurred in parallel sheets of flattened membrane cisternae (Fig. 9D), resembling the annulate lamellae (reviewed in Ref. 50) which are known to containly densely packed nuclear pore complexes (51).
These immunoelectron microscopic data also provided evidence that the Bcl-2 protein is located exclusively in the outer membrane of mitochondria (Fig. 9, A, C, and F), consistent with a previous report (22). The immunoreactivity seen in association with the mitochondrial outer membrane also occurred in a nonuniform distribution (Fig. 9A). Patches of antibody reactivity were also seen in the cytosol (Fig. 9C), perhaps representing the pool of Bcl-2 associated with the ER. In Fig. 9F, for example, a loop of ER that appears to be contiguous with the nuclear envelope can be seen winding into the cytosol. Patches of intense anti-Bcl-2 antibody immunoreactivity were found associated with this membrane structure. Peptide competition, use of control antibodies, and evaluations of the 32D-NEO and 32D-BCL-2 cell lines described above confirmed the specificity of these results. In Fig. 9, A and B, for example, where whole-cell views are provided, com- parisons of cells stained with anti-Bcl-2 antibody with and without peptide preabsorption revealed essentially complete abrogation of spe- cific immunoreactivity when competing peptide was included.
When postembedding methods were used in conjunction with im- munogold detection of anti-Bcl-2 antibodies, again the majority of the immunoreactivity was associated with the nuclear envelope and outer mitochondrial membranes (Fig. 10). Fig. 10B, for example, shows gold particles localized along the nuclear envelope and the circum- ference of a mitochondrion in SU-DHL-4 cells. Gold particles were also seen in the cytosol, perhaps reflecting Bcl-2 associated with ER or annulate lamella, but there was little deposition of particles over the nucleus. No clear evidence for association of Bcl-2 with the plasma membrane or with lysosomes or other intracellular orgenelles was obtained. Fig. 10C shows a high-power image of a mitochondrion, revealing two areas of gold particle deposition over the outer mem- brane. In several high-power images, the gold particles localized to the nuclear envelope appeared to cluster directly over nuclear membranes (Fig. 10D), suggestive of nuclear core complexes. The specificity of these results was confirmed by peptide competition experiments (Fig. 10A), use of preimmune serum, and evaluations of 32D-NEO and 32D-BCL-2 cells, as well as in preliminary experiments involving cryo-EM methods (not shown).
Studies of in Vitro Association of Bcl-2 Protein with Isolated Mitochondria
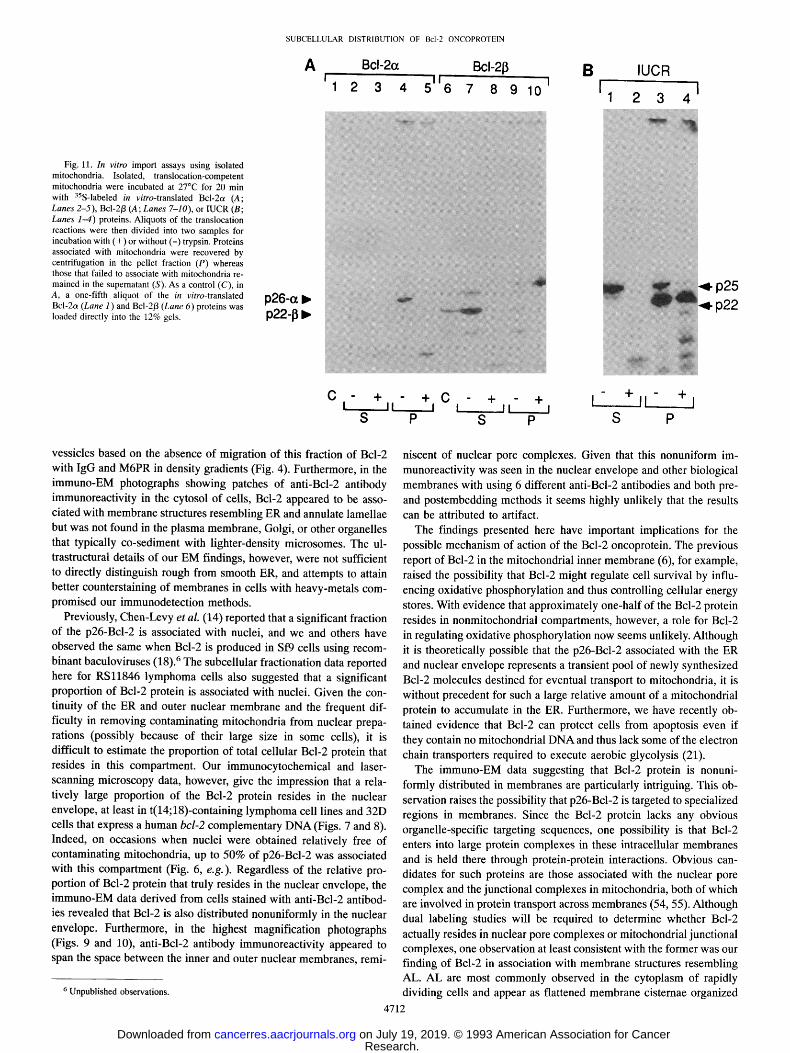
To further investigate the association of Bcl-2 protein with mito- chondria, we performed experiments in which the in vitro-translated Bcl-2 product was added to intact, isolated mitochondria under con- ditions where typical mitochondrial proteins will undergo import into this organelle (45). As shown in Fig. l lA, when 35S-labeled p26-Bcl-2 protein (referred to as Bcl-2o 0 was added to mitochondria, most of the protein became associated with mitochondria as determined by its cosedimentation with these organelles upon centrifugation (compare Lanes 2 and 4). Essentially all of the mitochondria-associated Bcl-2 protein, however, appeared to remain on the surface of the organelle,
4707
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
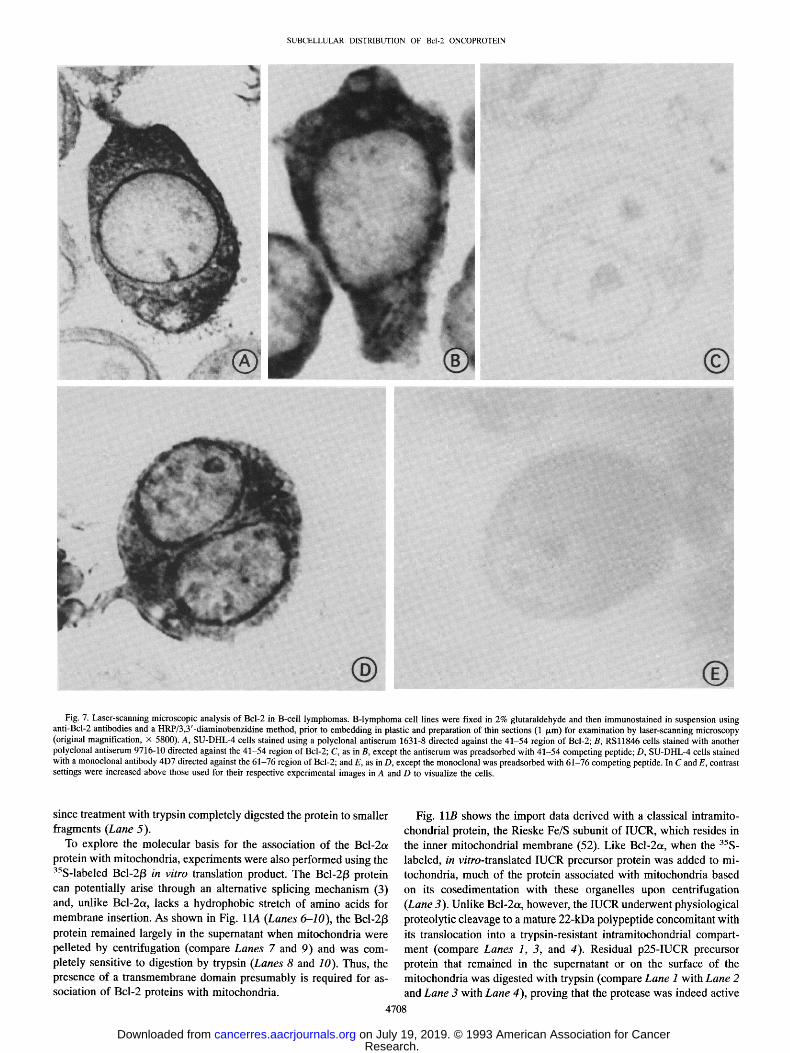
SUBCELLULAR DISTRIBUTION OF Bc l -20NCOPROTEIN
Fig. 7. Laser-scanning microscopic analysis of Bcl-2 in B-cell lymphomas. B-lymphoma cell lines were fixed in 2% glutaraldehyde and then immunostained in suspension using anti-Bcl-2 antibodies and a HRP/3,3'-diaminobenzidine method, prior to embedding in plastic and preparation of thin sections (1 /~m) for examination by laser-scanning microscopy (original magnification, • 5800). A, SU-DHL-4 cells stained using a polyclonal antiserum 1631-8 directed against the 41-54 region of Bcl-2; B, RSl1846 cells stained with another polyclonal antiserum 9716-10 directed against the 41-54 region of Bcl-2; C, as in B, except the antiserum was preadsorbed with 41-54 competing peptide; D, SU-DHL-4 cells stained with a monoclonal antibody 4D7 directed against the 61-76 region of Bcl-2; and E, as in D, except the monoclonal was preadsorbed with 61-76 competing peptide. In C and E, contrast settings were increased above those used for their respective experimental images in A and D to visualize the cells.
since treatment with trypsin completely digested the protein to smaller fragments (Lane 5).
To explore the molecular basis for the association of the Bcl-2ot protein with mitochondria, experiments were also performed using the 35S-labeled Bcl-2/3 in vitro translation product. The Bcl-2/3 protein
can potentially arise through an alternative splicing mechanism (3) and, unlike Bcl-2ot, lacks a hydrophobic stretch of amino acids for membrane insertion. As shown in Fig. l lA (Lanes 6-10), the Bcl-213
protein remained largely in the supernatant when mitochondria were pelleted by centrifugation (compare Lanes 7 and 9) and was com- pletely sensitive to digestion by trypsin (Lanes 8 and 10). Thus, the presence of a transmembrane domain presumably is required for as- sociation of Bcl-2 proteins with mitochondria.
Fig. l lB shows the import data derived with a classical intramito- chondrial protein, the Rieske Fe/S subunit of IUCR, which resides in the inner mitochondrial membrane (52). Like Bcl-2ot, when the 35S-
labeled, in vitro-translated IUCR precursor protein was added to mi- tochondria, much of the protein associated with mitochondria based on its cosedimentation with these organelles upon centrifugation (Lane 3). Unlike Bcl-2ot, however, the IUCR underwent physiological
proteolytic cleavage to a mature 22-kDa polypeptide concomitant with its translocation into a trypsin-resistant intramitochondrial compart- ment (compare Lanes 1, 3, and 4). Residual p25-IUCR precursor protein that remained in the supernatant or on the surface of the mitochondria was digested with trypsin (compare Lane I with Lane 2 and Lane 3 with Lane 4), proving that the protease was indeed active
4708
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
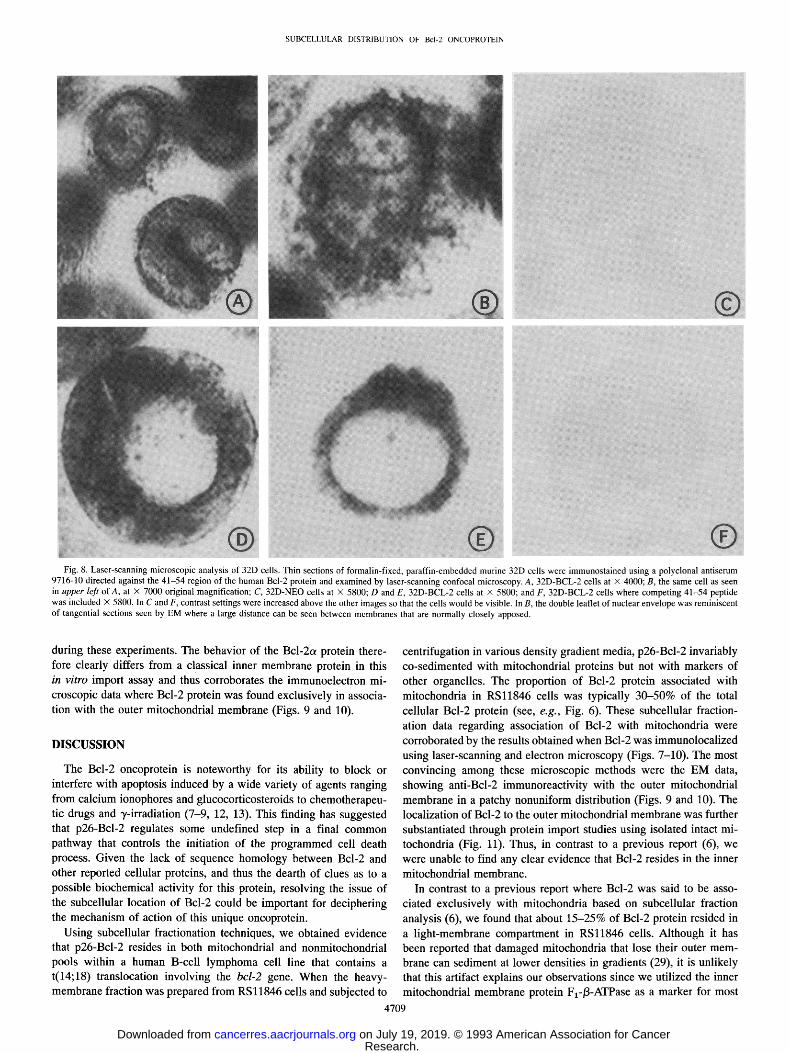
SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
Fig. 8. Laser-scanning microscopic analysis of 32D cells. Thin sections of formalin-fixed, paraffin-embedded murine 32D cells were immunostained using a polyclonal antiserum 9716-10 directed against the 41-54 region of the human Bcl-2 protein and examined by laser-scanning confocal microscopy. A, 32D-BCL-2 cells at x 4000; B, the same cell as seen in upper left of A, at X 7000 original magnification; C, 32D-NEO cells at X 5800; D and E, 32D-BCL-2 cells at x 5800; and F, 32D-BCL-2 cells where competing 41-54 peptide was included x 5800. In C and F, contrast settings were increased above the other images so that the cells would be visible. In B, the double leaflet of nuclear envelope was reminiscent of tangential sections seen by EM where a large distance can be seen between membranes that are normally closely apposed.
during these experiments. The behavior of the Bcl-2o~ protein there- fore clearly differs from a classical inner membrane protein in this in vitro import assay and thus corroborates the immunoelectron mi- croscopic data where Bcl-2 protein was found exclusively in associa- tion with the outer mitochondrial membrane (Figs. 9 and 10).
DISCUSSION
The Bcl-2 oncoprotein is noteworthy for its ability to block or interfere with apoptosis induced by a wide variety of agents ranging from calcium ionophores and glucocorticosteroids to chemotherapeu- tic drugs and T-irradiation (7-9, 12, 13). This finding has suggested that p26-Bcl-2 regulates some undefined step in a final common pathway that controls the initiation of the programmed cell death process. Given the lack of sequence homology between Bcl-2 and other reported cellular proteins, and thus the dearth of clues as to a possible biochemical activity for this protein, resolving the issue of the subcellular location of Bcl-2 could be important for deciphering the mechanism of action of this unique oncoprotein.
Using subcellular fractionation techniques, we obtained evidence that p26-Bcl-2 resides in both mitochondrial and nonmitochondrial pools within a human B-cell lymphoma cell line that contains a t(14;18) translocation involving the bcl-2 gene. When the heavy- membrane fraction was prepared from RSl1846 cells and subjected to
centrifugation in various density gradient media, p26-Bcl-2 invariably co-sedimented with mitochondrial proteins but not with markers of other organelles. The proportion of Bcl-2 protein associated with mitochondria in RSl1846 cells was typically 30-50% of the total cellular Bcl-2 protein (see, e.g., Fig. 6). These subcellular fraction- ation data regarding association of Bcl-2 with mitochondria were corroborated by the results obtained when Bcl-2 was immunolocalized using laser-scanning and electron microscopy (Figs. 7-10). The most convincing among these microscopic methods were the EM data, showing anti-Bcl-2 immunoreactivity with the outer mitochondrial membrane in a patchy nonuniform distribution (Figs. 9 and 10). The localization of Bcl-2 to the outer mitochondrial membrane was further substantiated through protein import studies using isolated intact mi- tochondria (Fig. 11). Thus, in contrast to a previous report (6), we were unable to find any clear evidence that Bcl-2 resides in the inner mitochondrial membrane.
In contrast to a previous report where Bcl-2 was said to be asso- ciated exclusively with mitochondria based on subcellular fraction analysis (6), we found that about 15-25% of Bcl-2 protein resided in a light-membrane compartment in RSl1846 cells. Although it has been reported that damaged mitochondria that lose their outer mem- brane can sediment at lower densities in gradients (29), it is unlikely that this artifact explains our observations since we utilized the inner mitochondrial membrane protein F1-/3-ATPase as a marker for most
4709
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
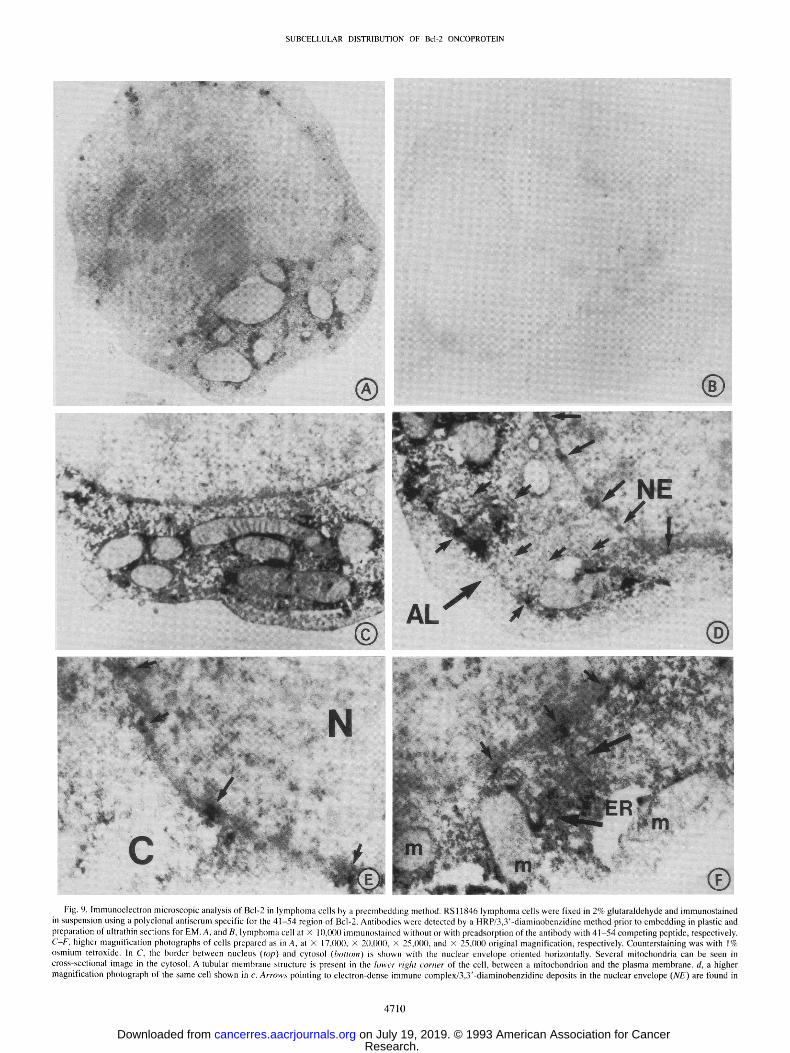
SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
Fig. 9. lmmunoelectron microscopic analysis of Bcl-2 in lymphoma cells by a preembedding method. RSl1846 lymphoma cells were fixed in 2% glutaraldehyde and immunostained in suspension using a polyclonal antiserum specific for the 41-54 region of Bcl-2. Antibodies were detectcd by a HRP/3,3'-diaminobenzidine method prior to embedding in plastic and preparation of ultrathin sections for EM. A, and B, lymphoma cell at • 10,000 immunostained without or with preadsorption of the antibody with 41-54 competing peptide, respectively. C-F, higher magnification photographs of cells prepared as in A, at • 17,l)00, • 20,000, • 25,0lj0, and • 25,000 original magnification, respectively. Counterstaining was with 1% osmium tetroxide. In C, the border between nucleus (top) and cytosol (bottom) is shown with the nuclear envelope oriented horizontally. Several mitochondria can be seen in cross-sectional image in the cytosol. A tubular membrane structurc is present in the lower right corner of the cell, between a mitochondrion and the plasma membrane, d, a higher magnification photograph of the same cell shown in c. Arrows pointing to electron-dense immune complex/3,3'-diaminobenzidine deposits in the nuclear envelope (NE) are found in
4710
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
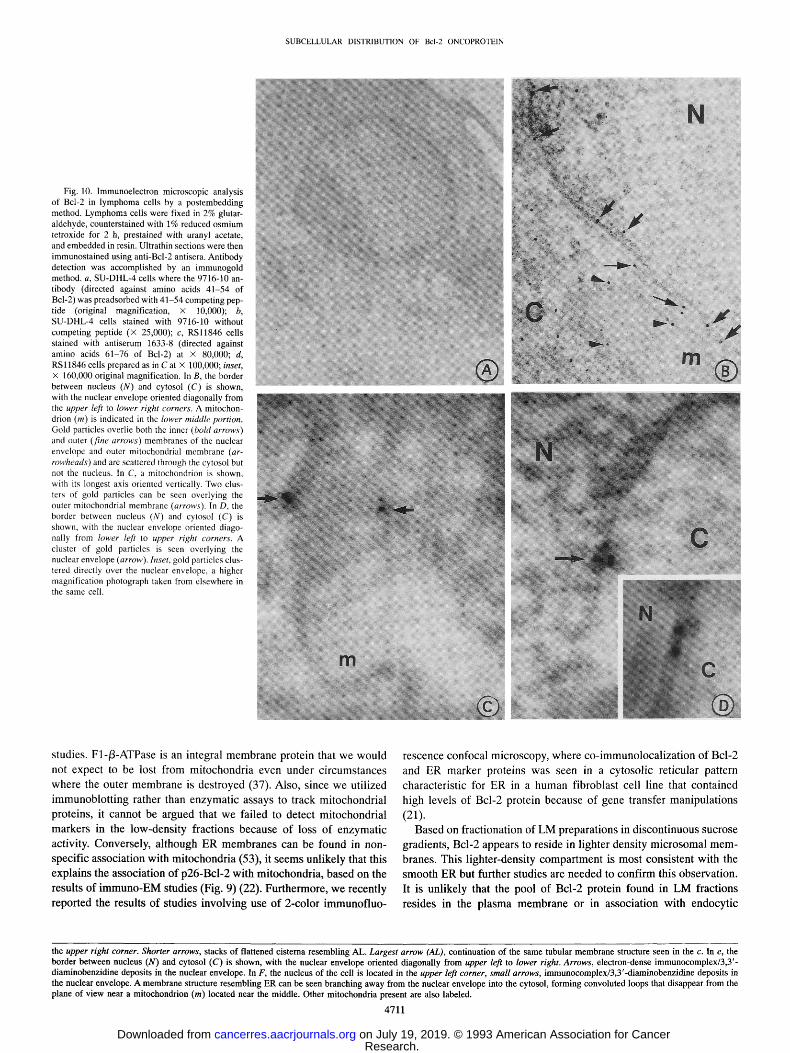
SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
Fig. 10. Immunoelectron microscopic analysis of Bcl-2 in lymphoma cells by a postembedding method. Lymphoma cells were fixed in 2% glutar- aldehyde, counterstained with 1% reduced osmium tetroxide for 2 h, prestained with uranyl acetate, and embedded in resin. Ultrathin sections were then immunostained using anti-Bcl-2 antisera. Antibody detection was accomplished by an immunogold method, a, SU-DHL-4 cells where the 9716-10 an- tibody (directed against amino acids 41-54 of Bcl-2) was preadsorbed with 41-54 competing pep- tide (original magnification, x 10,000); b, SU-DHL-4 cells stained with 9716-10 without competing peptide (x 25,000); c, RSl1846 cells stained with antiserum 1633-8 (directed against amino acids 61-76 of Bcl-2) at x 80,000; d, RSl1846 ceils prepared as in C at X 100,000; inset, x 160,000 original magnification. In B, the border between nucleus (N) and cytosol (C) is shown, with the nuclear envelope oriented diagonally from the upper left to lower right corners. A mitochon- drion (m) is indicated in the lower middle portion. Gold particles overlie both the inner (bold arrows) and outer (fine arrows) membranes of the nuclear envelope and outer mitochondrial membrane (ar- rowheads) and are scattered through the cytosol but not the nucleus. In C, a mitochondrion is shown, with its longest axis oriented vertically. Two clus- ters of gold particles can be seen overlying the outer mitochondrial membrane (arrows). In D, the border between nucleus (N) and cytosol (C) is shown, with the nuclear envelope oriented diago- nally from lower left to upper right corners. A cluster of gold particles is seen overlying the nuclear envelope (arrow). Inset, gold particles clus- tered directly over the nuclear envelope, a higher magnification photograph taken from elsewhere in the same cell.
s tudies. F1-/3-ATPase is an integral m e m b r a n e prote in that we w o u l d
not expect to be lost f rom mi tochondr i a even under c i r cums tances
where the outer m e m b r a n e is des t royed (37). Also , s ince we ut i l ized
i m m u n o b l o t t i n g rather than enzyma t i c assays to t rack mi tochondr ia l
proteins , it canno t be argued that we fa i led to detect mi tochondr i a l
markers in the l ow-dens i ty f rac t ions because o f loss o f e n z y m a t i c
activity. Converse ly , a l though E R m e m b r a n e s can be found in non-
speci f ic assoc ia t ion wi th m i t o c h o n d r i a (53), it s eems un l ike ly that this
expla ins the assoc ia t ion o f p26-Bcl -2 wi th mi tochondr i a , based on the
results o f i m m u n o - E M studies (Fig. 9) (22). Fur the rmore , w e recent ly
repor ted the results o f s tudies invo lv ing use o f 2-color i m m u n o f l u o -
rescence confoca l microscopy, whe re c o - i m m u n o l o c a l i z a t i o n o f Bcl -2
and E R marker proteins was seen in a cy toso l ic re t icular pat tern
character is t ic for ER in a h u m a n f ibroblas t cell l ine that con ta ined
h igh levels o f Bcl-2 protein because o f gene t ransfer man ipu la t ions
(21).
Based on f rac t iona t ion of L M prepara t ions in d i scon t inuous sucrose
gradients , Bcl-2 appears to reside in l ighter dens i ty m i c r o s o m a l m e m -
branes. This l ighter -dens i ty c o m p a r t m e n t is mos t cons is ten t wi th the
s m o o t h E R but fur ther s tudies are needed to con f i rm this observat ion.
It is un l ike ly that the pool o f Bcl -2 prote in f o u n d in L M frac t ions
res ides in the p la sma m e m b r a n e or in assoc ia t ion wi th endocy t i c
the upper right corner. Shorter arrows, stacks of flattened cisterna resembling AL. Largest arrow (AL), continuation of the same tubular membrane structure seen in the c. In e, the border between nucleus (N) and cytosol (C) is shown, with the nuclear envelope oriented diagonally from upper left to lower right. Arrows, electron-dense immunocomplex/3,3'- diaminobenzidine deposits in the nuclear envelope. In F, the nucleus of the cell is located in the upper left corner, small arrows, immunocomplex/3,3'-diaminobenzidine deposits in the nuclear envelope. A membrane structure resembling ER can be seen branching away from the nuclear envelope into the cytosol, forming convoluted loops that disappear from the plane of view near a mitochondrion (m) located near the middle. Other mitochondria present are also labeled.
4711
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from
SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
A Bcl-2a Bcl-213
1 2 3 4 5 6 7 8 9 10
B I
IUCR
1 2 3 4
Fig. 11. In vitro import assays using isolated mitochondria. Isolated, translocation-competent mitochondria were incubated at 27~ for 20 min with aSS-labeled in vitro-translated Bcl-2a (A; Lanes 2-5), Bcl-2/3 (A; Lanes 7-10), or IUCR (B; Lanes 1-4) proteins. Aliquots of the translocation reactions were then divided into two samples for incubation with (+) or without (-) trypsin. Proteins associated with mitochondria were recovered by centrifugation in the pellet fraction (P) whereas those that failed to associate with mitochondria re- mained in the supernatant (S). As a control (C), in A, a one-fifth aliquot of the in vitro-translated Bcl-2a (Lane 1) and Bcl-2/3 (Lane 6) proteins was loaded directly into the 12% gels.
p26-0~ �9 p22-1], �9
o
D
p25 p22
C - + t II
S - + C - + - + L- + + j
I t It ~ I [ ' P S P S P
vessicles based on the absence of migration of this fraction of Bcl-2 with IgG and M6PR in density gradients (Fig. 4). Furthermore, in the immuno-EM photographs showing patches of anti-Bcl-2 antibody immunoreactivity in the cytosol of cells, Bcl-2 appeared to be asso- ciated with membrane structures resembling ER and annulate lamellae but was not found in the plasma membrane, Golgi, or other organelles that typically co-sediment with lighter-density microsomes. The ul- trastructural details of our EM findings, however, were not sufficient to directly distinguish rough from smooth ER, and attempts to attain better counterstaining of membranes in cells with heavy-metals com- promised our immunodetection methods.
Previously, Chen-Levy et al. (14) reported that a significant fraction of the p26-Bcl-2 is associated with nuclei, and we and others have observed the same when Bcl-2 is produced in Sf9 cells using recom- binant baculoviruses (18). 6 The subcellular fractionation data reported here for RSl1846 lymphoma cells also suggested that a significant proportion of Bcl-2 protein is associated with nuclei. Given the con- tinuity of the ER and outer nuclear membrane and the frequent dif- ficulty in removing contaminating mitochondria from nuclear prepa- rations (possibly because of their large size in some cells), it is difficult to estimate the proportion of total cellular Bcl-2 protein that resides in this compartment. Our immunocytochemical and laser- scanning microscopy data, however, give the impression that a rela- tively large proportion of the Bcl-2 protein resides in the nuclear envelope, at least in t(14;18)-containing lymphoma cell lines and 32D cells that express a human bcl-2 complementary DNA (Figs. 7 and 8). Indeed, on occasions when nuclei were obtained relatively free of contaminating mitochondria, up to 50% of p26-Bcl-2 was associated with this compartment (Fig. 6, e.g.) . Regardless of the relative pro- portion of Bcl-2 protein that truly resides in the nuclear envelope, the immuno-EM data derived from cells stained with anti-Bcl-2 antibod- ies revealed that Bcl-2 is also distributed nonuniformly in the nuclear envelope. Furthermore, in the highest magnification photographs (Figs. 9 and 10), anti-Bcl-2 antibody immunoreactivity appeared to span the space between the inner and outer nuclear membranes, remi-
6 Unpublished observations.
niscent of nuclear pore complexes. Given that this nonuniform im- munoreactivity was seen in the nuclear envelope and other biological membranes with using 6 different anti-Bcl-2 antibodies and both pre- and postembedding methods it seems highly unlikely that the results can be attributed to artifact.
The findings presented here have important implications for the possible mechanism of action of the Bcl-2 oncoprotein. The previous report of Bcl-2 in the mitochondrial inner membrane (6), for example, raised the possibility that Bcl-2 might regulate cell survival by influ- encing oxidative phosphorylation and thus controlling cellular energy stores. With evidence that approximately one-half of the Bcl-2 protein resides in nonmitochondrial compartments, however, a role for Bcl-2 in regulating oxidative phosphorylation now seems unlikely. Although it is theoretically possible that the p26-Bcl-2 associated with the ER and nuclear envelope represents a transient pool of newly synthesized Bcl-2 molecules destined for eventual transport to mitochondria, it is without precedent for such a large relative amount of a mitochondrial protein to accumulate in the ER. Furthermore, we have recently ob- tained evidence that Bcl-2 can protect cells from apoptosis even if they contain no mitochondrial DNA and thus lack some of the electron chain transporters required to execute aerobic glycolysis (21).
The immuno-EM data suggesting that Bcl-2 protein is nonuni- formly distributed in membranes are particularly intriguing. This ob- servation raises the possibility that p26-Bcl-2 is targeted to specialized regions in membranes. Since the Bcl-2 protein lacks any obvious organelle-specific targeting sequences, one possibility is that Bcl-2 enters into large protein complexes in these intracellular membranes and is held there through protein-protein interactions. Obvious can- didates for such proteins are those associated with the nuclear pore complex and the junctional complexes in mitochondria, both of which are involved in protein transport across membranes (54, 55). Although dual labeling studies will be required to determine whether Bcl-2 actually resides in nuclear pore complexes or mitochondrial junctional complexes, one observation at least consistent with the former was our finding of Bcl-2 in association with membrane structures resembling AL. AL are most commonly observed in the cytoplasm of rapidly dividing cells and appear as flattened membrane cisternae organized
4712
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
either singly or in stacks of parallel sheets (reviewed in Ref. 50). Typically, AL are located towards the periphery of cells and contain densely spaced nuclear pore complexes. It has been speculated that AL serve as a reservoir of nuclear envelope precursor material that can be mobilized quickly in tumors and other rapidly dividing cells to ensure that their postmitotic chromosomes become enclosed by a nuclear envelope containing functional pore complexes (50, 51).
Although we can only speculate on a function for Bcl-2 based on the data presented here, one potential explanation lies with the Ca 2+ dependence of at least some of the endonucleases that have been implicated in the internucleosomal DNA fragmentation that typically accompanies apoptosis (56, 57). The major intracellular storage site for Ca 2+ is the lumen of the ER, which is contiguous with the space between the outer and inner nuclear membranes. Furthermore, mito- chondria can also sequester Ca 2+ under some circumstances (58). Recently, we have shown that apoptosis is accompanied by reparti- tinning of intracellular stores of sequestered Ca 2+ in an interleukin 3-dependent hematopoietic cell line when deprived of lymphokine, and we have observed that high levels of Bcl-2 protein production interfere with apoptosis and prevent these alterations in Ca 2§ homeo- stasis from occurring (25). Among the alterations in Ca 2+ found in these cells as they initiated apoptosis were reductions in the amounts of Ca 2§ stored in the ER and increases in mitochondrial Ca 2+ pools. Although highly speculative, therefore, Bcl-2 could potentially regu- late the activity of Ca 2+ pumps or channels in the ER, mitochondrial, and nuclear envelope and thereby influence the activity of Ca 2+- dependent endonucleases or other enzymes involved in the apoptotic process. Yet another possibility with relevance to Ca 2+ concerns recent data suggesting that a form of the Ca 2+- and Mg2+-dependent endonuclease DNase I may be responsible in part of the genomic digestion that typically occurs during apoptosis and the possibility that this enzyme is sequestered in the lumen of the ER (59). It is conceiv- able therefore that Bcl-2 could somehow preclude both the exit of Ca z+ and DNase I or other proteins from the lumen of the ER and thus forestall apoptosis. With regard to the possible association of Bcl-2 with nuclear pore and mitochondrial junctional complexes, although no connection between apoptosis and regulation of protein transport has been made heretofore, it is at least conceivable that alterations in this process could be casually involved in the mechanisms that lead to programmed cell death. For example, Eastman (11) and Ucker (60) have independently speculated that cdc-2 or related kinases could be involved in phosphorylating histones, lamins, or other nuclear pro- teins, thus leading to some of the characteristic morphological charges seen in the nuclei of apoptotic cells. Since the activities of this family of serine/threonine-specific kinases are dependent on association with cyclins, it is theoretically possible that Bcl-2 could prevent transport of newly translated cyclins or other proteins from the cytosol into the nucleus and thus block some of the biochemical events associated with apoptosis. Regardless of the accuracy of these speculations, the data presented here help to provide additional insights into the poten- tial mechanisms of action of this unique oncoprotein.
ACKNOWLEDGMENTS
We thank W. Neupert, N. Gonaitis, L. Smith, D. Meyer, and W. Brown for
providing antibodies; D. Meyer for technical advice; S. Delgado and M.
Hasham for photographic artwork; C. Stephens for manuscript preparation, and
D. Newmeyer for helpful discussions and critical reading of the manuscript.
Special thanks goes to Mildred Ostrander for donation of the laser-scanning
confocal microscope.
4713
REFERENCES
1. Tsujimoto, Y., Cossman, J., Jaffe, E., and Croce, C. Involvement of the bcl-2 gene in human follicular lymphoma. Science, (Washington, D.C.), 228: 1440-1443, 1985.
2. Korsmeyer, S. J. bcl-2 initiates a new category of oncogenes: regulators of cell death. Blood, 80: 879--886, 1992.
3. Tsujimoto, Y., and Croce, C. M. Analysis of the structure, transcripts and protein products of bcl-2, the gene involved in human follicular lymphomas. Proc. Natl. Acad. Sci. USA, 83: 5214-5218, 1986.
4. Cleary, M. L., Smith, S. D., and Sklar, J. Cloning and structural analysis of cDNAs for bcl-2 and a hybrid bcl-2/immunoglobulin transcript resulting from the t[14;18] translocation. Cell, 47: 19-28, 1986.
5. Vaux, D., Cory, S., and Adams, J. BCL-2 gene promotes haemopoietic cell survival and cooperates with c-myc to immortalize pre-B cells. Nature (Lond.), 335: 440-442, 1988.
6. Hockenbery, D., Nunez, G., Milliman, C., Schreiber, R. D., and Korsmeyer, S. J. Bci-2 is an inner mitochondrial membrane protein that blocks programmed cell death. Nature (Lond.), 348: 334, 1990.
7. Sentman, C. L., Shutter, J. R., Hockenbery, D., Kanagawa, O., and Korsmeyer, S. J. BCL-2 inhibits multiple forms of apoptosis but not negative selection in thymocytes. Cell, 67: 879, 1991.
8. Strasser, A., Harris, A. W., and Cory, S. BCL-2 transgene inhibits T cell death and perturbs thymic self-censorship. Cell, 67." 889, 1991.
9. Siegel, R. M., Katsumata, M., Miyashita, T., Louie, D. C., Greene, M. I., and Reed, J. C. Inhibition of thymocyte apoptosis and negative antigenic selection in bcl-2 transgenic mice. Proc. Natl. Acad. Sci. USA, 89: 7003-7007, 1992.
10. Wyilie, A. H., Morris, R. G., Smith, A. L., and Dunlop, D. Chromatin cleavage in apoptosis: association with condensed chromatin morphology and dependence on macromolecular synthesis. J. Pathol., 153: 313, 1987.
11. Eastman, A. Activation of programmed cell death by anticancer agents: cisplatin as a model system. Cancer Cells, 2: 275-280, 1990.
12. Miyashita, T., and Reed, J. C. BCL-2 gene transfer increases relative resistance of $49.1 and WEH 17.2 lymphoid cells to cell death and DNA fragmentation induced by glucocorticoids and multiple chemotherapeutic drugs. Cancer Res., 52: 5407-5411, 1992.
13. Miyashita, T., and Reed, J. C. Bcl-2 oncoprotein blocks chemotherapy-induced apop- tosis in a human leukemia cell line. Blood, 81: 151-157, 1993.
14. Chen-Levy, Z., Nourse, J., and Cleary, M. Z. The bcl-2 candidate proto-oncogene product is a 24-KD integral membrane protein highly expressed in lymphoid cell lines and lymphomas carrying the t[14;18] translocation. Mol. Cell. Biol., 9: 701-710, 1989.
15. Chen-Levy, S., and Cleary, M. L. Membrane topology of the Bcl-2 proto-oncogenic protein demonstrated in vitro. J. Biol. Chem., 265: 4929-4933, 1990.
16. von Heijne, G., Steppuhn, J., and Herrmann, R. G. Domain structure of mitochondrial and chloroplast targeting peptides. Eur. J. Biochem., 180: 535-545, 1989.
17. Cazals-Hatem, D., Louie, D., Tanaka, S., and Reed, J. C. Molecular cloning and DNA sequence analysis of cDNA encoding chicken homologue of the Bcl-2 oncoprotein. Biochim. Biophys. Acta, 1132: 109-113, 1992.
18. Alnemri, E. S., Robertson, N. M., Fernandes, T. F., Croce, C. M., and Litwack, G. Overexpressed full-length human BCL-2 extends survival of baculovirus-infected SF9 insect cells. Proc. Natl. Acad. Sci. USA, 89: 7295-7299, 1992.
19. Liu, Y-J., Mason, D. Y, Johnson, G. D., Abbott, S., Gregory, C. D., Hardie, D. L., Gordon, J., and MacLennan, I. C. M. Germinal center cells express bcl-2 protein after activation by signals which prevent their entry into apoptosis~ Eur. J. Immunol., 21: 1905, 1991.
20. Hamilton, M. S., Barker, H. E, Bail, J., Drew, M., Abbott, S. D., and Franklin, I. M. Normal and neoplastic human plasma cells express bcl-2 antigen. Leukemia (Baltimore), 5: 768-771, 1991.
21. Jacobson, M. D., Burne, J. F., King, M. P., Miyashita, T., Reed, J. C., and Raft, M. C. Bcl-2 blocks apoptosis in cells lacking mitochondrial DNA. Nature (Lond.), 361: 365-368, 1993.
22. Monaghan, P., Robertson, D., Amos, T., Dyer, M., Mason, D., and Greaves, M. Ultrastructural localization of Bcl-2 protein. J. Histochem Cytochem., 40: 1819-1825, 1992.
23. Reed, J., Tsujimoto, Y., Epstein, S., Cuddy, M., Slabiak, T., Nowell, P., and Croce, C. Regulation of bcl-2 gene expression in lymphoid cell lines containing t(14;18) or normal #18 chromosomes. Oncogene Res., 4: 271-282, 1989.
24. Winter, J., Variakojis, S., and Epstein, A. L. Phenotypic analysis of established diffuse histiocytic lymphoma cell lines utilizing monoclonal and cytochemical techniques. Blood, 63: 140-146, 1984.
25. Baffy, G., Miyashita, T., Williamson, J. R., and Reed, J. C. Apoptosis induced by withdrawal of interleukin-3 (IL-3) from an IL-3-dependent hematopoietic cell line is associated with repartitioning of intracellular calcium and is blocked by enforced Bcl-2 oncoprotein production. J. Biol. Chem., 268: 6511-6519, 1993.
26. Reed, J. C., Meister, L., Tanaka, S., Cuddy, M., Yum, S., Geyer, C., and Pleasure, D. Differential expression of bcl2 protooncogene in neuroblastoma and other human tumor cell lines of neural origin. Cancer Res., 51: 6529-6538, 1991.
27. Blobei, G., and Potter, V. R. Nuclei from rat liver: isolation method that combines purity with high yield. Science (Washington DC), 154: 1662-1165, 1966.
28. Ali, N., Aligue, R., and Evans, W. H. Highly purified bile-canalicular vesicles and lateral plasma membranes isolated from rat liver on Nycodenz gradients. Biochem. J., 271: 185-192, 1990.
29. Evans, H. W. Organelles and membranes. In: J. B. C. Findlay and W. H. Evans (eds.), Biological Membranes: a Practical Approach, pp. 1-35. Oxford, United Kingdom: RL Press, 1987.
30. Record, M., Bes, J. C., Chap, H., and Donste-Blazy, L. Isolation and characterization of plasma membranes from Krebs II ascite cells using Percoll gradient. Biochim.
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

SUBCELLULAR DISTRIBUTION OF Bcl-20NCOPROTEIN
Biophys. Acta, 688: 57, 1982. 31. Meyer, D. I., and Dobberstein, B. Proteins mediating vectorial translocation: purifi-
cation of the active domain of the endoplasmic reticulum docking protein. Methods Enzymol., 96: 692-700, 1983.
32. Fritsch, J. E., Buckmaster, M. J., and Storrie, B. Fibroblasts maintain a complete endocytic pathway in the presence of lysosomotrophic reagents. Exp. Cell Res., 175: 277-285, 1988.
33. Steinman, R. M., and Cohn, Z. A. The interaction of soluble horse radish peroxidase with mouse peritoneal macrophages in vitro. J. Cell Biol., 55: 186-204, 1972.
34. Hall, C., Liebaers, I., Di Natale, P., and Neufeld, E. E Enzymatic diagnosis of genetic mocopolysaccharide storage disorders. Methods Enzymol., 50: 439--456, 1978.
35. Morr6, D. J. Isolation of Golgi apparatus. Methods Enzymol., 22: 130-143, 1971. 36. Smith, P., Krohn, R., Hermanson, G., Mallia, K., Gartner, E, Prozenzano, M., Fuji-
moto, E., Goeke, N., Olson, B., and Klenk, D. Measurement of protein using bicin- choninic acid. Anal. Biochem., 150: 76-85, 1985.
37. Schwaiger, M., Herzog, V., and Neupert, W. Characterization of translocation contact sites involved in the import of mitochondrial proteins. J. Cell Biol., 105: 235-246, 1987.
38. Chen, Y., Hickey, W., Mezitis, S., Stieber, A., Lavi, E., Gonatas, J., and Gonatas, N. Monoclonal antibody 2H1 detects a 60--65 KD membrane polypeptide of the rough endoplasmic reticulum of neurons and selectively stains cells of several rat tissues. J. Histochem. Cytochem., 39: 635-643, 1991.
39. Brown, W., and Farquhar, M. G. The mannose-6-phosphate receptor for lysosomal enzymes is concentrated in cis-Golgi cisternae. Cell, 36: 295-307, 1984.
40. Kuo, L. M., Davies, H. C., and Smith, L. Monoclonal antibody to human cytochrome c: effect on electron-transfer reactions. Biochim. Biophys. Acta, 848: 247-255, 1986.
41. Hortsch, M., Avossa, D., and Meyer, D. I. A structural and functional analysis of the docking protein. Characterization of active domains by proteolysis and specific an- tibodies. J. Biol. Chem., 260: 9137-9145, 1985.
42. Reed, J., Yum, S., Cuddy, M., Turner, B., and Rapp, U. Differential regulation of the p72-74 RAF-1 kinase by in 3T3 fibroblasts expressing ras or src oncogenes. Cell Growth & Differ., 2: 235-243, 1991.
43. Tanaka, S., Saito, K., and Reed, J. C. Structure-function analysis of the apoptosis- suppressing Bcl-2 oncoprotein: substitution of a heteroiogous transmembrane domain restores function to truncated Bcl-2 proteins. J. Biol Chem, 268: 10920-10926, 1993.
44. Conboy, J. G., and Rosenberg, L. E. Posttranslational uptake and processing of in vitro synthesized transcarbamoylase precursor by isolated rat liver mitochondria. Proc. Natl. Acad. Sci. USA, 78: 3073-3077, 1981.
45. Fenton, W. A., Hack, A. M., Helfgott, D., and Rosenberg, L. E. Biogenesis of the mitochondrial enzyme methylmalonyl-CoA mutase. J. Biol. Chem., 259: 6616--6621, 1984.
46. Reed, J., Tanaka, S., Cuddy, M., Cho, D., Smith, J., Kallen, R., Sargovi, U., and Torigoe, T. A strategy for generating monoclonal antibodies against recombinant Baculovirus-produced proteins. Anal. Biochem., 205: 70-76, 1992.
47. Itoh, J., Osamura, Y., and Watanabe, K. Subcellular visualization of light microscopic specimens by laser scanning microscopy and computer analysis: a new application of image analysis. J. Histochem. Cytochem., 40: 955-967, 1992.
48. Courtoy, P. J., Quintart, J., and Baudhuim, P. Shift of equilibrium density induced by 3,3'-diaminobenzidine cytochemistry: a new procedure for the analysis and purifica- tion of peroxidase-containing organelles. J. Cell Biol., 98: 870-876, 1984.
49. Pezella, E, Tse, A. G. D., Cordell, J. L., Pulford, K. A. F., Gatter, K. C., and Mason, D. Y. Expression of the bcl-2 oncoprotein is not specific for the t(14;18) chromosomal translocation. Am. J. Pathol., 137: 225-230, 1990.
50. Kessel, R. G. The annulate lamellae-from obscurity to spotlight. Electron Microsc. Rev., 2: 257-348, 1989.
51. Dabauvalle, M-C., Loos, K., Merket, H., and Sheer, U. Spontaneous assembly of pore complex-containing membranes ("annulate lamellae") in Xenopus egg extract in the absence of chromatin. J. Cell Biol., 112: 1073-1082, 1991.
52. Hartl, F-U., Schmidt, B., Wachter, E., Weiss, H., and Neupert, W. Transport into mitochondria and intramitochondrial sorting of the Fe/S protein of ubiquinol-cyto- chrome c reductase. Cell, 47." 939-951, 1986.
53. Meier, P. J., Spycher, M. A., and Meyer, U. A. Isolation and characterization of rough endoplasmic reticulum associated with mitochondria from normal rat liver. Biochim. Biophys. Acta, 646: 283-287, 1981.
54. Neumeyer, D. The nuclear pore complex and nucleocytoplasmic transport. Curr. Opinion Cell Biol., in press, 1993.
55. Planner, N., and Neupert, W. The mitochondrial import apparatus. Annu. Rev. Bin- chem., 59: 331-353, 1990.
56. Wyllie, A. H., Kerr, J. E R., and Cumi, A. R. Cell death: the significance of apoptosis. Int. Rev. Cytol., 68: 251-306, 1980.
57. Caron-Leslie, L-A. M., Schwartzman, R. A., Gaido, M. L., Compton, M. M., and Cidiowski, J. A. Identification and characterization of glucocorticoid-regulated nucle- ase(s) in lymphoid cells undergoing apoptosis. J. Steroid Biochem. Mol. Biol., 40: 661-671, 1991.
58. Carafoli, E. IntraceUular calcium homeostasis. Annu. Rev. Biochem., 56: 395-433, 1987.
59. Peitsch, M. C., Polzar, B., Stephan, H., Crompton, T., MacDonald, H. R., Mannherz, H. G., and Tschopp, J. Characterization of the endogenous deoxyribonuclease in- volved in nuclear DNA degradation during apoptosis (programmed cell death). EMBO J., 12: 371-377, 1993.
60. Ucker, D. S. Death by suicide: one way to go in mammalian cellular development? New Biologist, 3: 103-109, 1991.
4714
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from

1993;53:4701-4714. Cancer Res Stanislaw Krajewski, Shigeki Tanaka, Shinichi Takayama, et al. Endoplasmic Reticulum, and Outer Mitochondrial MembranesOncoprotein: Residence in the Nuclear Envelope,
bcl-2Investigation of the Subcellular Distribution of the
Updated version
http://cancerres.aacrjournals.org/content/53/19/4701
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/53/19/4701To request permission to re-use all or part of this article, use this link
Research. on July 19, 2019. © 1993 American Association for Cancercancerres.aacrjournals.org Downloaded from









![Endoplasmic reticulum[1]](https://img.pdfslide.net/doc/110x75/58ed5fc71a28aba1678b4611/endoplasmic-reticulum1.jpg)