Embed Size (px)
Citation preview

New Phytol. (1990), 114, 173-182
The role of iron in nodulation and nitrogenfixation in Lupinus angustifolius L.
BY C A I X I A N T A N G \ ALAN D. ROBSON^AND MICHAEL J. DILWORTH'^
^ Soil Science Mid Plant Nutrition, School of Agriculture, The University of WesternAustralia, Nedlands, W.A. 6009, Australia
School of Biological a^id Environmental Sciences, Murdoch University, MurdochW.A. 6150, Australia
{Received 5 June 1989; accepted 16 October 1989)
SUMMARY
The effects of iron concentration in solution on nodulation and symbiotic N, fixation in lupins (Lupinusangustifolius L. cv. Yandee) were studied in solution culture in the glasshouse by comparing plants supplied withNH^NO.j with those reliant on N., fixation.
At very low iron supply, adding NHjNOj did not increase shoot growth. Under moderate rron deficiency,where plants were pale green, adding NHjNOj increased shoot growth, implying that moderate iron deficiencywas, operationally, nitrogen deficiency. Higher ir-on concentrations in solution were required for maximum growthof plants reliant on symbiotic Nj fixation than for those supplied with NH^NO,.
Iron deficiency depressed nodule initiation earlier than host plant growth, resulting in decreased nodule numberand mass. Alleviating iron deficiency enhanced leghaemoglobin production in nodules and increased nitrogenconcentrations in the shoots. Iron concentration in the youngest fully expanded leaves provided a good indicationof the severity of iron deficiency-caused chlorosis.
The results suggest that iron is involved in symbiotic N, fixation through effects on both nodule initiation andnodule function, and that the symbiosis has a higher iron requirement than that needed for host plant growth.
Key words: Lupinus angustifotius, iron deficiency, nodulation, nitrogen fixation, mineral interaction.
INTRODUCTION changes in nodule number, size, weight or dis-tribution if the nutrient is involved in nodulation.
Deficiencies of mineral nutrients may limit symbiotic Ir-on is an essential nutrient for both host legumenrtrogen fixation in legumes through specific effects and root nodule bacteria. Iron deficiency, whichon survival and gr-owth of rhizobia in the external occurs mainly on calcareous or alkaline soils, is amedium, on nodule initiation and development, on worldwide problem in production of crops, includingnodule function and on host plant growth (Loner- legumes. Correcting the deficiency by rron applica-agan, 1972; Robson, 1978; 1983). Several criteria tions to soil is impractical as applied iron quicklyhave been recommended for assessment of the becomes unavailable and chelated iron compoundsrelative requirements of nutrients by host legume are expensive (Chen & Barak, 1982). The alternative,growth and symbiotic nitr-ogen fixation (Robson, seleetion of iron-efficient plant cultivars, requires an1978; 1983). Frrstly, if symbiotic nitr-ogen fixation understanding of whether symbiotic nitrogen fixa-has a hrgher nutrient r-equir-ement than host plant tion is more sensitive than growth of the host legumegrowth, a negative interaction is expected between a to iron deficieney.nutrrent and combined nitrogen for legume growth. The role of iron in nodulation and symbioticSecondly, alleviating the deficiency of a nutrient that nitr-ogen fixation is not clearly understood. Poorrs requrred for symbiotie nitrogen fixation should nodulation caused by iron deficiency has beenmcrease the nitrogen concentration in tops. Thirdly, reported on chickpea (Rai, Singh & Prasad, 1982),alleviation of nutrient deficiency should result in lentil (Rai et al., 1984) and French bean (Hemanta-

174 C. Ta7ig, A. D. Robson and M.J. Dilworth
ranjan & Garg, 1986), but these reports did notdifferentiate bet'ween limitation of nodule initiationand subsequent development. Iron is important innodule nitrogen fixation as it is a component ofseveral key proteins such as nitrogenase, leghaemo-globin and ferredoxin. In Arachis hypogaea L. grownon a calcareous soil, iron deficiency resulted inarrested nodule development shortly after initiationand in delay or preventioti of nitrogenase production(O'Hara et al., 1988a). On an alkaline soil poorgrowth of Lupinus angustifolius L. was identified asdue to limittng iron supply (White & Robson, 1989)and could have resulted from poor nodulation. Irondeficiency in legumes may therefore manifest itselfas nitrogen deficiency in at least some circumstances.
The first aim of the present experiment was toassess the role of iron in nodule formation, nodulefunction and host plant growth in L. angustifolius.The second aim was to compare the effect of ironsupply on the growth of plants reliant on symbioticnitrogen fixation with that of plants supplied withmineral nitrogen.
MATERIALS AND METHODS
Experimental design
Plants were grown in a glasshouse, in solutionculture with roots maintained at 20-22 °C. Theexperimental design was a complete factorial withsix iron levels (0-20 /IM) and two nitrogen treat-ments, nil (-NH.NO^) and added 0-5 mM NH4NO3(+ NH4NO3). All plants were inoculated with Rrady-rhizobium. lupini WU425. Treatments were repli-cated three times and randomized within replicates.
Bacterial growth
B. lupini (WU425) was grown at 28 °C on slopes ofYGMA which contained (per litre): mannitol, 2 g ;glucose, 3 g ; yeast extract, 1 g; KjHPO^, 0-2 g;MgSO4.7H2O, 0-2 g; NaCl, 0-1 g; CaCl2.2H2O,30 mg; NH.NOg, 0-1 g; FeClg, 3 mg; MnSO, .4H2O, 2 mg and agar, 15 g, with a final pH of 70.Cells to be used as inoculum were suspended indeionized water; bacterial numbers were estimatedfrom a curve relating cell numbers to absorbance at600 nm.
Plant growth
Uniform seeds (mean Fe content 5-4/yg seed"') of L.angustifolius cv. Yandee (a cultivar sensitive to irondeficiency; White & Robson, 1989) were germinatedfor 6 d on stainless steel screens covered with cottongauze above an aerated solution of 06 mM CaSO^ and2 /iM H3BO3 until the rootlets were about 5 cm long.Nine seedlings held by lids were then transferred to5-1 plastic pots containing nutrient solution of thefollowing composition (//M) : KHjPO^, 20; KjSO,,
600; MgSO^, 200; CaCl^, 600; H3BO.J, 5; Na.,MoOj,0-03; ZnSO,, 0-75; MnSO,, 1-0; CoSO,, 0-2; andCuSO^, 0-2. Iron was added as ferric monosodiumsalt of ethylenediamine tetraacetic acid (FeNa-EDTA). A dense suspension of B. lupini WU425was added to all pots at a rate of about 3x10-^ cellsm r \ This solution was kept unchanged for 4 d andthen replaced by a similar solution with addedrhizobia. After another 2 d, plants were placed insolutions without added rhizobia and solutionschanged every second day. After 24 d followinggermination, when iron-adequate nodules beganfunctioning, NH^NO^ was added to +NH4NO3treatments to give a concentration of 0-5 niM. ThepH of the solution was adjusted daily to 5-5 withKOH.
Harvests
Three plants were taken from each pot at 20, 32 and45 d after germination. Plants were separated intothe youngest fully expanded leaf blades (YFB), tops,roots and nodules. To remove dust, plant topsincluding YFB were rinsed in deionized water,washed in 0-1 M HCl and again rinsed in deionizedwater. Roots were soaked in 0-01 M CaCl^ solutionfor 5 min and then rinsed in deionized water toremove external iron. One leaflet of the YFB wassampled for measurement of chlorophyll concen-tration. Other plant tissues and remaining noduleswere oven-dried at 68 °C for iron analysis.
Measurements
Nodule initials (foci of infected cortical cells, notvisible macroscopically) were counted using thefollowing procedure. Water rinsed roots were clearedin 10% KOH at room temperature for 8 h, followedby three rinses with water until the brown colour haddisappeared. Roots were acidified by soaking in0-25 M HCl for 5 min and stained in 0-1 % (w/v)Brilliant green (No. B-6756, Sigma) for 30 min andthen destained overnight in water. Nodule initialswere counted under a dissecting microscope at x 20magnification (Riley & Dilworth, 1985 a). Initialsrepresent an index of successful infection andinitiation separate from subsequent development(Riley & Dilworth, 1985 a).
Chlorophyll concentration in YFB was deter-mined by the method of Hiscox & Israelstam (1979).Leghaemoglobin concentration in nodules was de-termined by the procedure of Riley & Dilworth(1985 6). Total iron concentrations in tissues weredetermined using atomic absorption spectro-photometry after digesting plant materials in a 4:1mixture by volume of concentrated nitric andperchloric acids (Johnson & Ulrich, 1959). Nitrogenwas determined by a micro-Kjeldahl method(McKenzie & Wallace, 1954).
Nodulation and nitrogeti fixation in Lupinus angustifolius 175
RESULTSSymptoms
Chlorosis of the younger leaves, a ger-reral symptomof iron deficiency, occurred in lupir-rs about 2 weeksafter gern-rination for the 0 and 0-02 /lyi Fe treat-ments and after 4 weeks at 0'2 pw Fe. Plants suppliedwith 1 //M Fe or more showed no deficiencysymptoms.
The new leaves turned pale green and eventuallybright yellow. In contrast to the interveir-ral chlorosisassoerated with iron deficiency rn other plants,younger lupin leaves were uniformly yellow. Brownnecrotic spots then developed along the middlesection of the yellow leaves. Under very severe irondeficiency, the leaves curved inwards giving a claw-like appearance (Snowball & Robson, 1986), afterwhich leaves were shed and growing points died.
Adding NH.,NO.j char-rged symptom expression.Compared with plants reliant on N.^, plants given
were more chlorotic at lower iron supplies(0 and 0-02 pm) and less chlorotic at 0-2 p.M iron(Table 1).
Slight nitrogen deficiency occurred in all plantsabout 2 weeks after germination. Plants showed
paleness ii-i old leaves, especially at higher iron levels.By 24 d after germination, plants with 5 and 20 //Miron in solution began to recover. By 26 d aftergermination, a similar recovery occurred in plantswith 1 //M iron, and in NH^NO^-fed plants, butthose with low iron concentrations continued toappear nitrogen-deficient and growth was stunted.
Plant growth
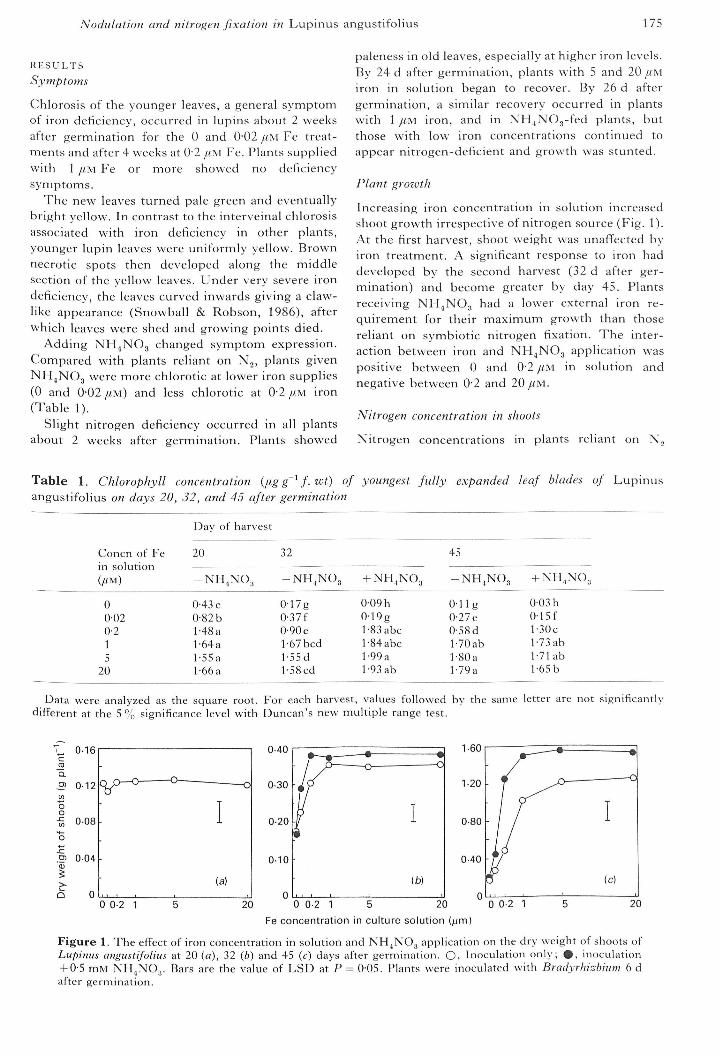
Increasing iron concentration in solution increasedshoot growth irrespective of nitrogen source (Fig. 1).At the first harvest, shoot weight was unafTected byiron treatment. A significant response to iron haddeveloped by the second harvest (32 d after ger-mination) and becon-re greater by day 45. Plantsreceivir-rg NH^NOg had a lower external iron re-quirement for their maximum growth than thosereliant on symbiotic nitrogen fixation. The inter-action between iron and NH^NOg application waspositive between 0 and 0-2 //M in solution andnegative between 0-2 and 20//M.
Nitrogen concentration in shoots
Nitrogen concentrations in plar-rts reliant on N.
Table 1. Chlorophyll concentration (fig g ^ f. wt) of youngest fully expanded leaf blades of Lupinusangustifolius on days 20, 32, and 45 after germination
Concn of Fe
(/IM)
00-020-215
20
Day of harvest
20
-NH4NO3
0-43 c0-82b1-48 al-64al-55a1-66 a
32
- N H . N O ,
0-17gO-37f0-90 el-67bcdl-55dl-58cd
+ NH,NO3
0-09 h0-19gl-83abcl-84abc1-99 al-93ab
45
-NHjNO. ,
0-llg0-27e0-58 dl-70abl-80al-79a
+ NH,NO3
0-03 h0-15 fl-30cl-73ab1-71 abl-65b
Data were analyzed as the square root. For each harvest, values followed by the same letter are not significantlydifferent at the 5 % significance level with Duncan's new multiple range test.
"L 0-16
0 0-2 1 20
0 4 0
0 30 -
0-20 -I
0-10 •
0 0 2 1 5 20
Fe concentration in culture solution
1-60
1-20 -
0-80 -
0-40 -
0 0 2 1
Figure 1. The efTect of iron concentration in solution and NH^NOj application on the dr-y weight of shoots ofLupinus angustifolius at 20 (a), 32 (b) and 45 (c) days after germination. O, Inoculation only; 0 , inoculatior-i+ 0-5 mM NHjNO.,. Bars are the value of LSD at P = 0-05. Plants were inoculated with Bradyrhizbium 6 dafte r germination.
176 C. Tang, A. D. Robson and M.J. Dilworth
Table 2. Effects of iron concentration in solution and application of 0-5 niMNH^NO.^ on nitrogen concentration ("/„ dry weight) in shoots of Lupinusangustifolius
Concn of Fe
ijlM)
00-020-215
20
LSD (P = 0'05)FeNFexN
Day of
32
- N H ,
2-121-631-543-263-253-13
harvest
NO,,
0-360-210-52
+ NH,NO
4-174-204-324-314-324-36
45
- N H ,
1-671-262-484-064-213'97
NO,,
0-350-200-49
+ NH,NO3
5-204'714-775-014-914-92
increased with increasing iron concentrations insolution up to 1 //M for harvests at day 32 and day 45(Table 2). Plants given no iron had higher nitrogenconcentrations than plants receiving low iron (0-02and 0-2 fiM at day 32; 0-02 /iM at day 45). Applicationof NH^NOg enhanced nitrogen concentration inshoots at all iron levels but nitrogen concentration inshoots was not affected by iron application in thepresence of NH^NO.,.
Iron concentration in plant tissues
Increasing supplies of iron increased iron concen-trations in YEB, tops, roots, nodules and wholeplants in both nitrogen-fixing and nitrogen-fertilizedplants (Table 3). Iron concentrations in YEB andshoots on plants given NH4NO3 were greater thanthose in the plants reliant on Nj ; the same was truefor roots at day 32.
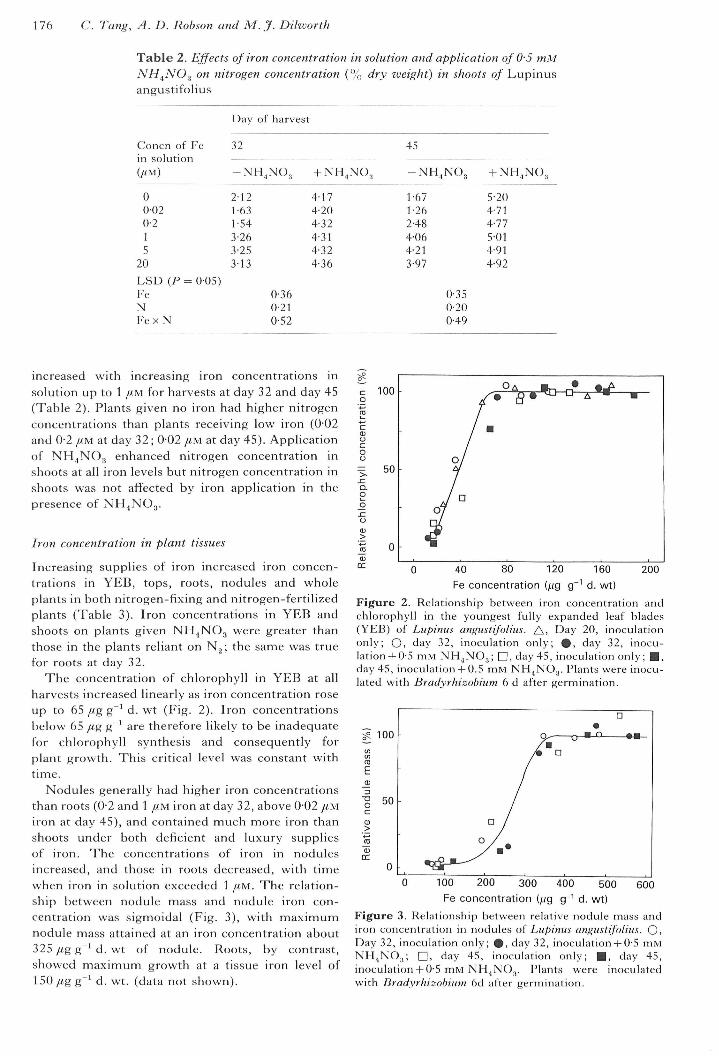
The concentration of chlorophyll in YEB at allharvests increased linearly as iron concentration roseup to 65/<g g^'d. wt (Fig. 2). Iron concentrationsbelow 65 fig g ' are therefore likely to be inadequatefor chlorophyll synthesis and consequently forplant growth. This critical level was constant withtime.
Nodules generally had higher iron concentrationsthan roots (0-2 and 1 /<M iron at day 32, above 0-02 //Miron at day 45), and contained much more iron thanshoots under both deficient and luxury suppliesof iron. The concentrations of iron in nodulesincreased, and those in roots decreased, with timewhen iron in solution exceeded 1 fim. The relation-ship between nodule mass and nodule iron con-centration was sigmoidal (Fig. 3), with maximumnodule mass attained at an iron concentration about325 ^g g ' d. wt of nodule. Roots, by contrast,showed maximum growth at a tissue iron level of1 50 /tg g ^ d. wt. (data not shown).
0 40 80 120 160 200
Fe concentration (^g g"^ d. wt)
Figure 2. Relationship between iron concentration andchlorophyll in the youngest fully expanded leaf blades(YEB) of Lupinus angustifolius. /\, Day 20, inoculationonly; O, day 32, inoculation only; %, day 32, inocu-lation+0-5 mM NH^NO.,; D, day 45, inoculation only; • ,day 45, inoculation + 0.5 mM NH^NO,. Plants were inocu-lated with Bradyrhizobium 6 d after germination.
100
T3O 50
100 200 300 400 500 600Fe concentration {,i/g g ^ d. wt)
Figure 3. Relationship between relative nodule mass andiron concentration in nodules oi Lupinus angustifolius. O,Day 32, inoculation only; %, day 32, inoculation+ 0-5 mMNHjNO.,; n , day 45, inoculation only; • , day 45,inoculation + 0-5 mM NH^NO.j. Plants were inoculatedwith Bradyrliizohium 6d after germination.

Nodulation and nitrogen fixation in Lupinus angustifolius 177
_«
s
So
6fl
S
KO
o
I
K C
•2
21 «
il11
o
rt -^
o
o
7, - . 5
C 3
Q i
O^ 0000 IT)
T 1
GO U• ^ CO
\O !>•• '—'
.,—(
ONr^
T 3
0000O J
u
oooI O
o[ .r o' ^
ca- ^• ^
00
ONOMO
o00
( J _
(U
00
a;
O
! _,r o
-0CJ
l OTJ-1—1
"Ou
CO
-Q ra!>• VO^ I.O(N CO
CJ
X ra(^ , 1T—I O
raONONl O
ra
oo
11
IIa.
c
'noc
J O C O'— r o ^ O C O r o T—i
CO r-i
T— O4 • . — . T — i \ O - - — '
\D O oo O CO CX)CsJ ro in 00 "^ ^O
O fN0 0 0
(N (N
O O O ^ i n o O O O ^ in 0
g.
E
c3
Q
O
a ra
oj ra
Qra

178 C. Tang, A. D. Robson and M. J. Dilworth
Nodulation
The number of nodule initials at day 20 increasedmore than ten-fold over the range from 0 to 5 //,Miron, but was no higher at 20//M than at 5 /iM (Fig.4). The concentration of chlorophyll in YEB,however, had almost reached the maximum value inthe presence of 0-2//M iron. Clearly, the stimulatoryeffect of iron on the number of nodule initials wasmore rapid than the effect of iron on host plantgrowth (Fig. 1 a).
Increasing iron concentration in solution increasednodule number at all harvests (Table 4), reaching amaximum at about 5 //,M iron. The effect of iron inincreasing nodule number was evident at day 20, butwas less marked than that on the number of noduleinitials (Fig. 4). At the two later harvests (days 32and 45), nodule number increased even more rapidlyas iron level increased. Plants with 5 or 20/iM ironhad visible nodules 15 d after germination (9 d afterinoculation), whereas nodules did not appear untilday 17 at 1 //,M and even later for lower ironconcentrations. Addition of NH^NO., did notaffect nodule numbers, probably because NH^NO.jwas added after plants had completed noduleestablishment.
The effects of iron on nodule fresh weight perplant followed a similar pattern to those on nodulenumber (Table 4). Irrespective of iron treatments,however, nodule weight per plant and nodule sizewere decreased where NH^NOg was added.
0 0-2 1 5 20Fe concentration in culture solution
Figure 4. Effects of iron concentration in solution on thenumber of nodule initials ( • ) and visible nodules (D) onroots, and on chlorophyll concentration (%) in theyoungest fully expanded leaf blades (YEB) of Lupinusangustifolius at day 20. Bars are LSD values (P = 0-05) ofchlorophyll (a), nodule number (b) and the number ofnodule initials (c). Plants were inoculated with Brady-rhizobium 6 d after germination.
No effect of iron levels on nodule size was seen atthe first harvest, but the few nodules on plants grownwith 0'02 or 0'2 fiM iron grew much larger at the twolater harvests (Table 4).
Leghaemoglobin
The production of leghaemoglobin was depressedunder conditions of iron deficiency (Table 5).
Table 4. Effects of iron concentration in solution and application of 0-5 mMweight and size for Lupinus angustifolius inoculated with Bradyrhizobium
on nodule number, fresh
Day ofharvest
20
32
45
Concn of Eein solution(/i.M)
00-020-215
20
00-020-215
2000-020-215
20
Nodule number(No plant
-NH.NC
OdOdl c7 b
lOab12a
l cl c3b
42 a54 a41a
l c< l c
3 b39 a44 a41a
')
), +NH,NO
2 beOd2 be
35 a43 a40 a
l cl c3 b
41a38 a45 a
F'resh weight(mg plant"')
3 - N H . N O . ,
OcOc3 b
18a28 a37a
16ef28 de
105 be502 a497 a504 a
21 cd46 be
597 a1492 a1687 a1983a
+ NH,NO,
lOfOg
46 cd230 ab288 ab264 ab
4d48 be
119b833 a896 a890 a
Nodule size(mg nodule"'
, -NH.NO.,
n.a.n.a.
2-8 a2-7 a2-8 a2-7 a
11 be32 a35 a13 be9 be
13 be
17e98 ab
191a38ede42 cd51bc
)
+ NH,NO3
6 cn.a.18ab
8c7c7 c
6f34ede36cde21 de27cde20 de
Data excluding nodule size at day 20, were analyzed as logj,,. Eor each harvest, values followed by the same letter arenot different at P = 0-05 with Duncan's new multiple range test,
n.a., not available.

Nodulation and nitrogen fixation in Lupinus angustifolius 179
Table 5. Effects of iron concentration in solution and application of 0-5 mM NH^NO.^ on leghacmoglobinconcentration {nmol g~^ fresh zveight) in nodules of Lupinus angustifolius
Concn of Fe
i/IM)
00-020-215
20
LSD (P = 0-05)FeNFex N
Day of harvest
20
- N H j N O ,
n.a.n.a.18172739
n.s.
32
- N H
211824SO6449
4NO3
117
16
+ NH.,NO,
272748656087
45
- ^
202629545453
m,NO,
7n.s.n.s.
+ NH,NO,
183228485343
n.a., not available,n.s., not significant.
Nodules of iron-adequate plants (1 to 20//M iron)contained more than double the leghaemoglobinconcentration of iron-deficient plants. NHjNOgslightly increased leghaemoglobin concentration atday 32 but not at day 45.
Iron uptake and distribution
Total iron uptake by plants increased with time atidincreased iron concentrations in solution for bothnitrogen treatments. The addition of NH^NOg whichincreased plant growth, increased iron uptake (Table6).
Iron distribution within the plants was markedlyinfluenced by iron and tiitrogen treatments (Table6). Adding NH^NO., decreased iron content innodules and increased iron cotitent in shoots.
DISCUSSIONEffects of iron on nodulation a?id nitrogen fixation
In the present experiment, iticreasitig iron con-centration in solution markedly increased the num-ber of nodule initials. The effect of iron in etihaticitignodule initiation was prior to that on host plantgrowth and may indicate that nodule initiation isdirectly affected by iron. The external iron con-centration needed to give maximal nodule initiationappears to be much higher than that required forniaximal chlorophyll biosynthesis.
The effect of iroti on nodule number and weightper plant showed a similar pattern to that of iron onthe number of nodule initials. Depression of nodulenumber by iron deficiency may therefore occurthrough effects on nodule initiation. In this experi-nient, where rhizobia were added to plant roots foronly a short period, nodule initiation did not seem tooccur at later stages. In addition, 20 d after ger-
mitiation, increasitig iron concentration in solutiotiincreased nodule number less than it did noduleinitials. Poor nodule initiation therefore appears tobe the major cause of depressed nodule tiumber.
The few nodules on iron-deficient plants grewmuch larger than those oti iron-sufBcietit platits, butthis compensatioti occurred otily under moderateiroti deficiency. Where no iron was supplied, plantsgrew so poorly that tiodules were small, since otilyvery limited metabolites would have been availableto support nodule development. It is possible thatiron indirectly affects subsequent nodule devel-opment by restricting metabolite supply from thehost plant.
Iron deficiency has previously beeti reported tocause a decrease in tiodule number and mass inchickpea (Rai et al., 1982), lentil (Rai et al., 1984),French bean (Hemantaratijan & Garg, 1986) andpeanut (O'Hara et al, 1988a; 1988Z)). Sexeral ofthese studies did not distinguish direct effects of irondeficiency on nodule itiitiation and developmentfrom itidirect effects mediated through host plantgrowth. Hetnantaratijan & Garg (1986) found thatiroti application to Fretich bean grown on an iroti-deficient soil increased nodule number and postu-lated that at! effect on nodule initiation is importatit.O'Hara et o/.,(1988(j) concluded that the tnostsensitive stage of the peatiut symbiosis to irotideficiency was early nodule development and notnodule initiatioti, since they observed a similarnumber of tiodule initials in both control and foliariron-supplemented plants. However, as shown here,in lupitis the stage most sensitive to iron deficieticyappears to be nodule initiation rather than noduledevelopment.
Iticreasing external iron concentration increasedthe tiitrogen coticentration in shoots of lupin plantsreliant on symbiotically fixed nitrogen. This effect of

180 C. Tang, A. D. Robson and M. J. Dilworth
O
cT
I
5
5.
-ce
X)
H
Q
*
O W)
" c
C 3
c3.E
Q
00 O LO 00
V V
OOOOOrn'o 00 in in
O rvlO O CD
Oo
Q
C3
Q
oo
• a
O
-a0)
c
* d

Nodulation and nitrogen fixation in Lupinus angustifolius 181
iron was not observed in platits supplied withNHjNOg, which suggests that the efTect of ironsupply on the nitrogen concentration in N,, fixingplants does tiot result from either changes in plantgrowth patterns or in the metabolism of nitrogenwithin the plants. It was also evident that iron-defieient plants appeared nitrogen deficient at earlystages of growth. Low nitrogen concentrations andcontents due to iroti deficiency have also beenreported for French bean (Hemantaranjan & Garg,1986; Hemantaranjan, 1988), peanut (O'Hara et al.,19886) and mungbean (Chahal & Chahal, 1988).
As in French beati (Hemantaranjan & Garg, 1986)and peanut (O'Hara et al., 1988c7), iron deficiencydepressed the concentration of leghaemoglobin inlupin nodules. As leghaemoglohin is a key protein inNj fixation in nodules, its concentration getierallyeorrelating with nitrogen fixed (Graham & Parker,1963), any effect of iron deficiency on leghaemo-glohin production is likely to depress N, fixation.
Nodules had higher iron concentrations than rootsand shoots. Moreover, the internal iron concen-tration required for maximal nodule mass was muchhigher than that for host roots, implying that noduleformation may require higher internal iron than hostgrowth.
The nature of the interaction with combined nitrogen
When iron supply was extremely limited, plantscontained a higher percentage nitrogen than mod-erately iron-deficient plants and did not respond toNH^NOg. Growth of the plants was apparentlystunted hy iron deficiency rather than nitrogendeficiency. As iron concentratioti in solution in-ereased, a positive interaction with NH4NO3 oc-curred. Under conditions of moderate iron defici-ency, the symbiosis was still itihibited, platitsappearing pale green with low nitrogen concen-trations. Nitrogen fertilization increased theirgrowth and nitrogen coticentration. A negativeinteraction with NH^NO., existed with furtherincrease of iron supplies, indicating that the efTect ofiron on nitrogen fixation is direct and symbiosis hasa greater iron requirement than host platit growth(Robson, 1978; 1983). The response of lupins to ironapplication is thus similar to that of subterraneanclover to calcium, which is directly itivolved insymbiotic nitrogen fixation (Loneragan, 1959).
NH |NO.,-fertilized plants had higher iroti conceti-trations in YEB and shoots than nitrogen-fixingplants. This is partly due to the depression of nodulemass by NH,NO., so that more iron could betranslocated to shoots instead of to nodules. In workon molybdenum deficiency in soybean, Chotech-aungmanirat (1988) found a similar phenomenon;plants given combined nitrogen had less nodule massand higher molvbdenum concentrations iti the
youngest fully expanded leaves atid shoots than didplants dependent on fixed nitrogen.
Iron concentration in YEB provided a goodindication of the severity of iron deficiency-causedchlorosis on lupiti plants. The concentration in-dicative of iron deficiency in these leaves remainedconstatit at all harvests. An iron concentration itiYEB of 65 //g g~' dry weight appears to be a criticallevel below which lupiti plants show iroti deficiencysymptoms, and is consistent with that found byWhite & Robson (1989).
.ACKNOWLEDGEMENTS
Grateful acktiowledgement is made to Dr G. W. O'Hara,Mr W. J. Simmons and Mr K. Snowball for technicalassistatice and Dr P. F. White for ptoviditig seed.
REFERENCES
CHAHAL, P. P. K. & CHAHAL, V. P. S. (1988). Interaction ofRhizobium and Meloidogyne incognita oti sytnbiotic nitrogetifixation in mungbean under difTerent concentrations of iron.Journal of Plant Nutrition 11, 821 828.
CHEN, Y . & BARAK, P. (1982). Iron nuttition of plants incalcareous soils. Advances in Agronomy 35, 217 240.
Ct-tOTECHAiTNGMANtRAT, S. (1988). Diagnosis of nmtybdcnum def-iciency in soybeans by plant analysis. M.Sc. Thesis, School ofAgriculture, The University of Western Australia.
GRAHAM, P. H. & P.^RKER, C . .A. (1963). Leghaemoglobin andsymbiotic nitrogen fixation. Australian Journal of Science 23,231-232.
HEMANTARANJAN, .\. (1988). iroti fettilization tn relation tonodulation and nitrogen fixation in French bean {PhaseolusT'l/lgaris L.). Journal of Plant Nutrition 11, 829-842.
HEMANTARANJAN, A. & GARG, O . K. (1986).' Introduction ofnitrogeti-fixing nodules through iron atid zinc fertilizatioti itithe non nodule-forming Fretich beati {Phaseolu.t fiitgaris L.).Journal of Plant Nutrition 9, 281-288.
Htscox, J. D. & isRAELSTAM, G. F. (1979). A method for theextractioti of chlorophyll from leaf tissue without rnaceration.Canadian Journal of Botany 57, 1332-1334.
JOHNSON, C . M . & Ut.RicH, A. (1959). Analytical methods for usein plant analysi.s. California Agriculture Experiment StationBulletin \ o . 766.
LONERAGAN, J. F. (1959). Calcium in the nitrogen metabolistii ofsubterranean clover. Australian Journal of Biological Scienee 12,26-39.
LONERAGAN, J. F. (1972). The soil chemical environment inrelation to symbiotic nitrogen fixation. In: Use of Isotopes forStudy of Fertiliser Utilisation by Legume Crops. TechnicalReport No. 149. FAO/IAEA, Vienna.
McKt:NZtE, H. A. & WAt.LACti, H. .S. (1954). The Kjehdahldetermination of nitrogen : a critical study of digestioti con-ditions-temperature, catalyst, and oxidising agent. AustralianJournal of Chemistry 7, 55 70.
O'HARA, G . W . , Dtt.woRTH, M. J., BOONKERD, N . & PARKTMAN, P.(1988(j). Iron deficiency specilically litnits nodule de\elopmentin peanut inoculated with tiradvrhizobium sp. New PItvtologist108, 51 57.
O'HARA, G . W . , HARTZOOK, A., BEt.L, R. W. & LONERAGAN, J. F.(1988i). Response to Bradyrhizobium strains of peanut cultivarsgrown under iron atress. Journal of t^lnnt Nutrition 11, 843-852.
RAI, R. , PtiASAD, \'., C H O I D H I R V , S. K . & StNHA, N. P. (1984).Iron tiutrition and symbiotic N.2-fixation of lentil {Lens eulinaris)genotypes in calcareous soil. Journal of Plant Nutrition 7399-405.
RAt, R., StNGH, S. \ . & PRA.SAD, V. (1982). Efleet of presstnudatnetid pyrite on symbiotic N'.^-fixation, active iron contents ofnodules, grain yield and quality of chickpea {Cicer arietimimLinn) genotypes in calcareous soil. Journal of Ptant Nutrition 5,905 913.

182 C. Tang, A. D. Robson and M. J. Dilworth
RlI.EY, I . T . & Dli.wOKTH, M.J . (1985a). Cobalt status andeffects on soil populations of Rhizohiiim lupim. rhizospherecolonization and nodule initiation. Soil Biology and Biochemistry17, 81 8.S.
Rii.i-Y, I. T. & DILWORTH, M . J. (198.S/)). Cobalt requirement fornodule formation and function in Lupinus angustifolius L., NewPhytologist 100, 347-359.
RoBS(JN, A. D. (1978). Mineral nutrition limiting nitrogen fixationin legumes. In: The Mineral Nutrition of Legumes on Tropicaland Subtropical Soils (Ed. by C. S. Andrew & E. J. Kamprath),pp. 277-293. CSIRO, Melbourne.
RoiiSON, A. D. (1983). Mineral nutrition. In; Nitrogen Fixation,vol. 3 (Ed. by W. J. Broughton), pp. 36-55. Oxford.
SNOWBALL, K . & ROBSON, A. D. (1986). Symptoms of NutrientDeficieney: Lupins, pp. 59-60. Soil Science and Plant Nutrition,University of Western .Australia.
WHITE, P. F. & ROBSON, A. D. (1989). Lupin species and peasvary widely in their sensitivity to iron deficiency. AustralianJournal of Agricultural Research 40, 539-547.






























