Embed Size (px)
Citation preview

ANNALS OF CLINICAL AND LABORATORY SCIEN CE, Vol. 16, No. 4 Copyright © 1986, Institu te for Clinical Science, Inc.
The Selective Suppression of Immunogenicity by Hyaluronic Acid*J. M ICHAEL D ELM AGE, P h .D . , | DARLEEN R. POWARS, M .D .,f PATRICK K. JAYNES, P h .D .,§ and SAMUEL E. ALLERTON, Ph .D."
fEstelle Doheny Eye Foundation, Los Angeles, CA 90033
fDepartment of Pediatrics, School of Medicine, University of Southern California,
Los Angeles, CA 90033 §DeYor Laboratories, Inc.,
Youngstown, OH 44512 NDepartment o f Basic Sciences,
University of Southern California School of Dentistry,Los Angeles, CA 90089
ABSTRACTA hyaluronidase-sensitive com ponent of human peritoneal fluid from a
patient with W ilms’ tum or when injected into rabbits has been shown to suppress the form ation of hum oral precipitating antibodies to certain major classes of proteins present in the fluid.
Furtherm ore, it has been found that hyaluronic acid, when included with certain test antigens (serum albumin, fetuin) or antigen mixtures (tumor isolates or mixtures of albumin, immunoglobulin G and im m unoglobulin M), produces a m arked distortion or complete blockage of immu- noelectrophoresis precipitin arcs, as well as altered gel chromatography elution profiles.
These findings that hyaluronic acid can interfere profoundly with both the elicitation of a com plete antibody response and the formation of “normal” patterns of antigen-antibody precipitates in laboratory tests supports the possibility that this polysaccharide may play an im m uno-regulatory role by masking potential immunogens. Consideration of the mechanisms for these in vivo and in vitro effects suggests that there may be some common basis in an “excluded volume” property of the hyaluronate, but this does not appear sufficient to explain the complexity and selectivity of the observed phenom ena.
IntroductionAmong the major classes of vertebrate
glycosaminoglycans (acid m ucopolysac
* Address reprint requests to J. Michael Deimage, Ph.D ., Estelle Doheny Eye Foundation, 1355 San Pablo Street, Los Angeles, CA 90033.
charides), the hyaluronic acids are rather d is tin c tiv e b e c au se of (a) th e ir h igh molecular weights, (b) their ubiquitous distribution among many tissues, and (c) th e ir o ccu rren ce as u n b ran ch ed and unsulfated polysaccharide chains generally free of covalently bound p ro te in .17 Increased levels of hyaluronic acid, and
3030091-7370/86/0700-0303 $01.20 © Institute for Clinical Science, Inc.

3 0 4 D E L M A G E , P O W A R S , JA Y N E S , A N D A L L E R T O N
of the o th er glycosaminoglycans, have been noted in association with a variety o f tu m o rs .7 In r e c e n t y ea rs , m any repo rts have ap p eared com paring the glyeosaminoglycan profiles of neoplastic cells and tissues with those of their normal counterparts.1014 Although the differences observed are often striking, it has b een d ifficu lt to in te rp re t th em because the biological roles of these substances in normal and aberrant developm ental processes are no t fully u n d e rstood.^!,3 ,13,20 ,25,27
This study was initiated by our discovery that a sample of a viscous peritoneal fluid from a patient with W ilms’ tum or elicited an incom plete antibody response in rabbits, i.e ., antibodies were not produced against protein antigens known to be present in the specimen. Subsequent experim ents im plicated hyaluronic acid as the likely suppressive agent. O ther tests with conventional im m unoelectro- phoresis procedures showed that hyaluronic acid is capable of altering immu- n o p rec ip itin arcs (and in som e cases preventing the ir formation) w hen it is present in test antigen sources. Studies with similar mixtures of hyaluronate and these test proteins w ere perform ed with molecular sieve column chromatography in order to examine the selectivity and the extent of binding betw een the polysaccharide and the protein antigens.
M aterials and M ethodsHum an peritoneal fluid was obtained
from a pa tien t (J.M.) during an operation for surgical rem oval of a prim ary W ilm s’ tum or. H um an um bilical cord hyaluronic acid, crystalline bovine albumin, and hum an immunoglobulins (IgG, IgM) w ere purchased* and Spiro’s purified calf fetuin and antiserum to whole human sera w ere obtained, f Electropho-
retically p u re hum an m ercap ta lbum in was prepared from fresh plasma by e th anol p rec ip ita tion and tre a tm e n t w ith cysteine according to th e m ethods of Allerton et al.2 Varidase (streptokinase, s trep todornase) was ob tained^ as was F reund’s com plete adjuvant. §
P r e p a r a t io n o f A n t is e r a
Two groups of four young female New Zealand white rabbits that w ere used to develop antisera w ere inoculated with injectâtes consisting of 1.5 ml antigen solution containing 15 to 50 mg protein mixed with 1.5 ml F reu n d ’s com plete ad juvan t. This p re p a ra tio n was th o r oughly emulsified and was injected sub- cutaneously in 0.5 ml portions to six sites at the base of each animal’s neck. Four booster injections w ere given at approximately 20-day intervals, and the animals were bled via cardiac puncture w ithin 15 days of the last booster. All of the blood samples w ere collected in sterile Vacu- tainer tubes and allowed to clot. Following centrifugation, the serum was tested for sp ec ific ity ag a in st w ho le h um an serum with O uchterlony and im m uno- e le c tro p h o re s is te c h n iq u e s . F o r th e experim ents p resen ted here , the an tisera samples from animals w ithin each group w ere found to b e qualitatively in d is tin g u ish ab le and w ere th e re fo re pooled. The antisera w ere aliquotted and stored frozen.
I m m u n o e l e c t r o p h o r e s is
The M illip o re-W o rth in g to n Phoro- slide electrophoresis system was used for all im m unoelectrophoresis analyses as re c o m m e n d e d by th e m a n u fac tu re r,
* Sigma Corporation, St. Louis, MO. t Gibco Grand Island, NY.
$ L ederle L abora tories, A m erican C yanam id Company, Pearl River, NY.
§ Calbiochem Corporation, San Diego, CA.

S E L E C T IV E S U P P R E S S IO N O F IM M U N O G E N IC IT Y 3 0 5
except that Coomassie Brilliant Blue was substituted for the Buffalo Black stain. A sample volume of two (xl and an antisera volum e of 50 jjuI w ere used. Sam ples were electrophoresed at 100 volts for 35 minutes, incubated with antisera, extensively washed, and finally stained with 0.4 percent Coomassie Brilliant Blue in five percen t m ethanol-acetic acid solution.
“ R o c k e t” im m u n o e le c tro p h o re s is plates w ere used to examine the effect of h y a lu ro n ic acid on th e in te ra c tio n s betw een bovine serum album in (BSA) and rabbit anti-BSA. These plates were p repared by mixing 0.5 ml of antisera with liquefied Ionagar11 at 45°C followed by cooling to room tem perature. Sample wells of 1.5 mm w ere in troduced into the gels to accommodate 15 jxl of sample. Following an equilibration electrophoresis at 100 volts for 15 m inutes, specimens with and without added hyaluronic acid w ere electrophoresed for two hours at 100 volts. After washing, the gels w ere stained as indicated.
I m m u n o d i f f u s io n
O u ch te rlo n y d o u b le diffusion agar gelsll w ere filled w ith e ith e r 10 |xl of antigen or an tisera, incuba ted for 48 hours, then deproteinized and stained as indicated for the im m unoelectrophoresis.
D ig e s t io n s w it h V a r id a s e
Hum an peritoneal fluid and other test sam ples w ere enzym atically degraded with Varidase contain ing p o ten t hyal- uronidase activity (0.1 g p er dl in calcium and m agnesium -free p h o sp h a te buffered isotonic saline) for 24 hours at 37°C.26 C ontrol specim ens w ere incubated in parallel w ithout Varidase.
G e l F il t r a t io n
Molecular sieve chromatography, used to explore the interactions betw een hyal- u ronate and se lec ted serum p ro te in s, was conducted in glass columns (2.5 cm X 50 cm) packed with Sephacryl S-200. * The columns w ere equilibrated with isotonic buffered saline at pH 7.2, and the samples w ere loaded in a like buffer. The columns w ere m aintained at a flow of 0.5 ml per m inute, m onitored with a spec- tropho tom etert (at 280 nanom eters), and the effluent was collected in 0.7 ml samples.
ResultsThe im m unoelectrophoresis profile of
peritoneal flu id from the pa tien t w ith W ilm s’ tum or p roduced w ith an tisera against this m aterial (generated in our rabbits) is shown in figure 1. For comparison, the profile produced with this same antigen source and antisera made against Varidase-treated peritoneal fluid is illustrated in figure 2. It is apparent from these examples that after four to six w eeks p o s t- in o c u la tio n , th e an im als treated with the “intact” peritoneal fluid generated antibodies only against five proteins of beta electrophoretic mobility, whereas the digested samples generated a w ide spectrum of an tibodies w hich reacted with antigens of the test specimens. The la tte r of these provided an im m unoelectrophoresis profile analogous to that made by commercial antisera to whole hum an sera (figure 3). O f particular in te rest was the dem onstration of precipitin arcs against major protein antigens such as albumin and IgG only with the commercial antisera or our antisera to the digested m aterial. Also w orthy of note was the distortion and elongation of precipitin lines in all of the
II Consolidated Laboratories, Chicago Heights, IL. H Hyland Laboratories, Costa Mesa, CA.
* Pharmacia, Uppsala, Sweden, t Beckman DB, Brea, CA.
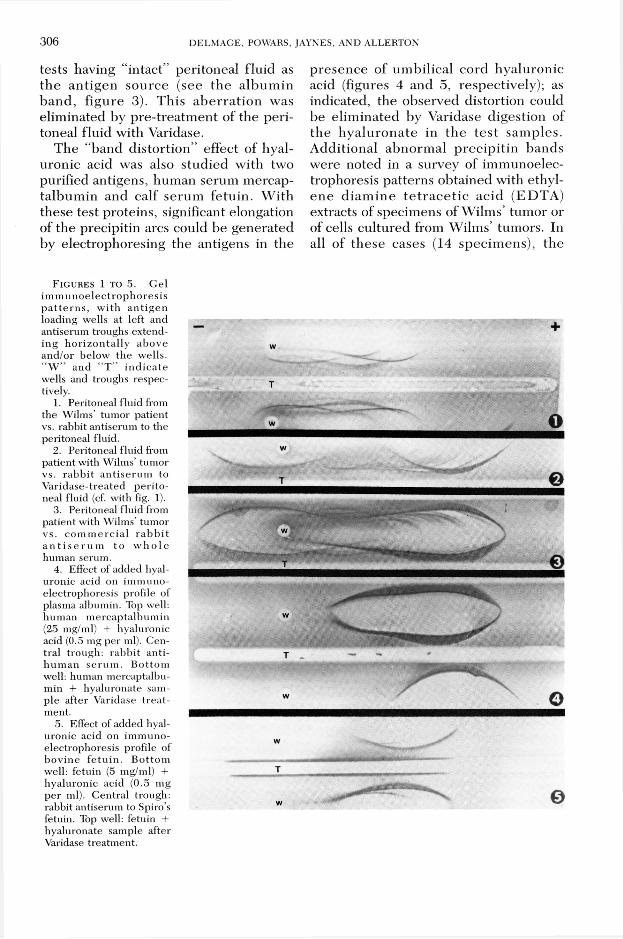
306 D E L M A G E , P O W A R S , JA Y N E S , A N D A L L E R T O N
tests having “intact” peritoneal flu id as th e an tigen source (see th e a lbum in b and , figure 3). T his ab e rra tio n was elim inated by p re-treatm ent of the peritoneal fluid with Varidase.
The “band d isto rtion” effect of hyaluronic acid was also stud ied with two purified antigens, hum an serum mercap- talbum in and calf serum fetu in . W ith these test proteins, significant elongation of the precipitin arcs could be generated by electrophoresing the antigens in the
p resence of um bilical cord hyaluronic acid (figures 4 and 5, respectively); as indicated, the observed distortion could be elim inated by Varidase digestion of th e h y a lu ro n a te in th e te s t sam ples. A dditional abnorm al p rec ip itin bands w ere noted in a survey of im m unoelec- trophoresis patterns obtained with ethylen e d iam in e te tr a c e t ic ac id (ED TA ) extracts of specim ens of W ilm s’ tum or or of cells cultured from W ilm s’ tumors. In all of these cases (14 specim ens), the
F i g u r e s 1 t o 5. G el im m u n o e lec tro p h o re s is p a t te rn s , w ith an tig en loading wells at left and antiserum troughs extending h o rizo n ta lly above and/or below the wells. “ W ” and “ T ” in d ic a te wells and troughs respectively.
1. Peritoneal fluid from the Wilms’ tum or patient vs. rabbit antiserum to the peritoneal fluid.
2. Peritoneal fluid from patient with Wilms’ tumor vs. rab b it an tise rum to V aridase-treated p e rito neal fluid (cf. with fig. 1).
3. Peritoneal fluid from patient with Wilms’ tumor vs. co m m erc ia l ra b b it a n t i s e r u m to w h o le human serum.
4. Effect of added hyaluronic acid on im m unoelectrophoresis profile of plasma albumin. Top well, hum an m ercaptalbum in (25 mg/ml) + hyaluronic acid (0.5 mg per ml). Central trough: rabb it an tihu m an se ru m . B ottom well: human mercaptalbumin + hyaluronate sample after Varidase tre a tment.
5. Effect of added hyaluronic acid on im m unoelectrophoresis profile of bov ine fe tu in . B ottom well: fetuin (5 mg/ml) + hyaluronic acid (0.5 mg per ml). C entral trough: rabbit antiserum to Spiro’s fetuin. Top well: fetuin + hyaluronate sample after Varidase treatm ent.
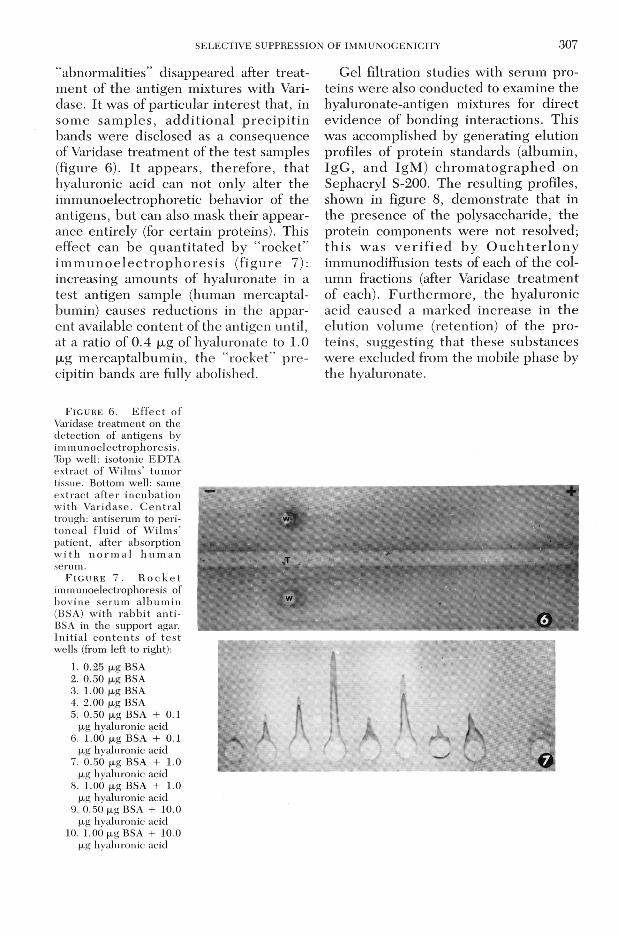
S E L E C T IV E S U P P R E S S IO N O F IM M U N O C E N IC IT Y 307“abnorm alities” disappeared after trea tm ent of the antigen mixtures with Vari- dase. It was of particular in terest that, in som e sam p les , a d d itio n a l p re c ip it in bands w ere disclosed as a consequence of Varidase treatm ent of the test samples (figure 6). I t appears, therefo re , tha t hyaluronic acid can not only alter the im m unoelectrophoretie behavior of the antigens, bu t can also mask their appearance entirely (for certain proteins). This effect can be q u a n tita ted by “ ro ck e t” im m u n o e le c tro p h o re s is (f ig u re 7): increasing am ounts of hyaluronate in a test antigen sample (human mercaptal- bumin) causes reductions in the apparent available content of the antigen until, at a ratio of 0.4 |xg of hyaluronate to 1.0 |xg m ercaptalbum in, the “rocket” p re cipitin bands are fully abolished.
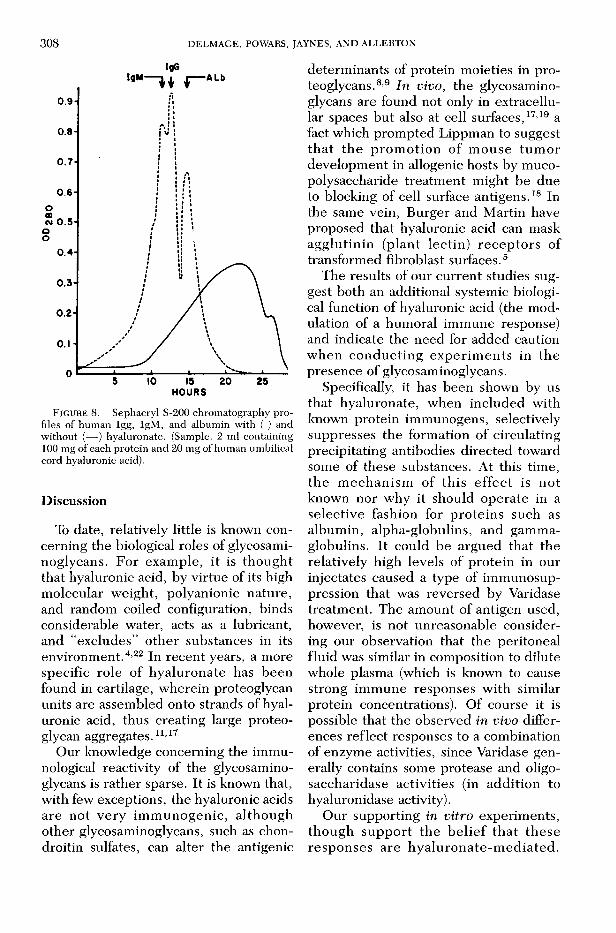
Gel filtration studies with serum proteins w ere also conducted to examine the hyaluronate-antigen mixtures for direct evidence of bonding interactions. This was accomplished by generating elution profiles of p ro tein standards (album in, IgG , and IgM ) c h ro m a to g ra p h e d on Sephacryl S-200. The resulting profiles, shown in figure 8, dem onstrate that in the presence of the polysaccharide, the protein com ponents w ere not resolved; th is w as v e r i f ie d by O u c h te r lo n y immunodiffusion tests of each of the colum n fractions (after Varidase treatm ent of each). F u rtherm ore , the hyaluronic acid caused a m arked increase in th e elution volum e (retention) of the p ro teins, suggesting that these substances w ere excluded from the mobile phase by the hyaluronate.
F i g u r e 6 . E f f e c t o f Varidase treatm ent on the detection of antigens by im m unoelectrophoresis. Top well: isotonic EDTA extract of W ilm s’ tum or tissue. Bottom well: same ex tract a fte r incubatio n w ith V aridase. C e n tra l trough: antiserum to perito nea l f lu id o f W ilm s’ patient, after absorption w i th n o r m a l h u m a n serum.
F i g u r e 7 . R o c k e t immunoelectrophoresis of bov ine se ru m a lb u m in (BSA) w ith rab b it an ti- BSA in the support agar. In itia l c o n te n ts o f te s t wells (from left to right):
1. 0.25 fxg BSA2. 0.50 fxg BSA3. 1.00 fjig BSA4. 2.00 fjig BSA5. 0.50 ^g BSA + 0.1
(jig hyaluronic acid6. 1.00 fxg BSA + 0.1
|xg hyaluronic acid7. 0.50 (jig BSA + 1.0
jxg hyaluronic acid8. 1.00 ^g BSA + 1.0
(jig hyaluronic acid9. 0.50 |xg BSA + 10.0
fxg hyaluronic acid10. 1.00 p,g BSA + 10.0
(jig hyaluronic acid
a |
f t k 1 ■ V X
- 0
3 0 8 D E L M A G E , PO W A R S, JA Y N E S , A N D A L L E R T O N
IgG
F IG U R E 8. Sephacryl S-200 chromatography profiles of human Igg, IgM, and albumin with ( ) and without (— ) hyaluronate. (Sample: 2 ml containing 100 mg of each protein and 20 mg of human umbilical cord hyaluronic acid).
DiscussionTo date, relatively little is known con
cerning the biological roles of glycosami- noglycans. For exam ple, it is though t that hyaluronic acid, by virtue of its high m olecular w eight, polyanionic nature, and random coiled configuration, binds considerable water, acts as a lubricant, and “excludes” o ther substances in its environm ent.4,22 In recent years, a more specific role of hyalu ronate has been found in cartilage, w herein proteoglycan units are assem bled onto strands of hyaluronic acid, thus creating large proteoglycan aggregates.11,17
Our knowledge concerning the im m unological reactivity of the glycosamino- glycans is rather sparse. It is known that, w ith few exceptions, the hyaluronic acids are no t very im m unogen ic , a lthough other glycosaminoglycans, such as chon- droitin sulfates, can alter the antigenic
determ inants of protein moieties in proteoglycans.8,9 In vivo , the glycosaminoglycans are found not only in extracellular spaces but also at cell surfaces,17,19 a fact which prom pted Lippm an to suggest th a t th e p ro m o tio n of m ouse tu m o r developm ent in allogenic hosts by mucopolysaccharide treatm ent m ight be due to blocking of cell surface antigens.18 In the same vein, Burger and M artin have proposed that hyaluronic acid can mask ag g lu tin in (p lan t lectin ) re c e p to rs of transform ed fibroblast surfaces.5
The results of our curren t studies suggest both an additional systemic biological function of hyaluronic acid (the modulation of a hum oral im m une response) and indicate the need for added caution w hen co n d u c tin g ex p erim en ts in th e presence of glycosaminoglycans.
Specifically, it has been shown by us that hyaluronate, w hen included with known protein immunogens, selectively suppresses the form ation of circulating precipitating antibodies d irected toward some of these substances. At this time, th e m ech an ism of th is effec t is no t known nor why it should operate in a selective fashion for p ro te in s such as album in, alpha-globulins, and gam m aglobulins. It could be argued that the relatively high levels of protein in our injectâtes caused a type of im m unosuppression that was reversed by Varidase treatm ent. The am ount of antigen used, however, is not unreasonable considering our observation that the peritoneal fluid was similar in composition to dilute whole plasma (which is known to cause strong im m une responses w ith similar protein concentrations). O f course it is possible that the observed in vivo differences reflect responses to a combination of enzym e activities, since Varidase generally contains some protease and oligo- saccharidase activities (in addition to hyaluronidase activity).
O ur supporting in vitro experim ents, th ough su p p o rt th e b e lie f th a t th ese resp o n ses are h y a lu ro n a te -m ed ia ted .

S E L E C T IV E S U P P R E S S IO N O F IM M U N O G E N IC IT Y 3 0 9
Collectively, our observations appear to indicate that hyaluronic acid interacts with albumin and other proteins so as to alter their immunogenicity. O ne possible explanation for such interaction is a so- c a l le d “ e x c lu d e d v o lu m e ” e f f e c t described by Balazs,4 w hereby the large effective volume of hyaluronate in solution reduces the domain of w ater availab le to o th e r so lu tes (w hich in tu rn , becom e less soluble and te n d to seek accessible com partm ents of higher water activity). An example of such a reduced solubility consequence was described in Hellsing’s study of the effect of dextrans which increased the am ount of precipitate form ed in an albumin-antialbumin reaction.12
L auren t15,16 also has review ed many examples of the com partm ental redistribution phenom enon, including the classical study of O gston,23 in which albumin was p re fe ren tia lly d is tr ib u te d to th e hyaluronic acid-free cham ber in a transfilter experim ent. O ur own gel chrom atography study may fu rther exemplify th is p h e n o m e n o n ; th e h y a lu ro n a te increased th e partition ing of th e test proteins toward the gel phase, thereby causing delays in elution and mixing of protein components. This same kind of phenom enon may account for distortion o f im m u n o e le c t r o p h o r e s is b a n d s , although it does not seem adequate to explain the selective loss of antigen and im m u n o g e n ic ity . H y a lu ro n ic ac id - m ediated electrophoretic band distortion has also b een rep o rte d in association with the production of hyaluronic acid by n e o p la s tic c e lls .1,13,28 A seco n d , no t m utually exclusive, m echanism m ight in v o lv e p r e f e r e n t i a l b in d in g , as d e s c r ib e d by P ig m an an d h is a sso ciates.21,24 In their studies, new moving b o u n d aries and in c reased m obilities w ere observed w hen intact hyaluronic acid was present with proteins of synovial fluids in m oving-boundary electrophoresis. Such associations of hyaluronic acid w ith various proteins may be too
weak to de tec t by conventional means such as gel chrom atography or sedim entation velocity studies, b u t they could have marked effects on immunogenicity, shapes of im m unoprecip itin bands and solubility of antigen-antibody complexes.
Additionally, the formation of charge- m ediated complexes betw een glycosami- noglycans and particular proteins could significantly alter the im m unogenicity of the proteins in question. The possible ability of hyaluronate to mask the im m unogenicity of particular p ro teins com bined with the observation that certain neoplasm s p roduce excess am ounts of hyaluronate1,3,13,20,25,27 suggests a functional role. Indeed, it is tem pting to postulate that neoplastic cells may produce glycosaminoglycans as an immunogenic mask to reduce or elim inate the recognition of tum or cell surface proteins by circulating antibodies. This m ight be partic u la r ly r e le v a n t in v iru s - in d u c e d neoplasms w here viral proteins are in teg ra ted in to th e cell m em b ran e upon v irus re le a se . A lth o u g h o u r c u r re n t understanding of the possible role glycosaminoglycans play in vivo is speculative, our stud ies do affect th e way in which experim ents should be designed.
Hyaluronate clearly affects the availability of particular proteins to the surrounding environm ent. Specifically, it is ap p aren t th a t th e search for and the characterization of “tum or-associated” p ro te in a n tig e n s6 m u st c o n s id e r th e potential interfering effects of hyaluronic acid and possibly o ther glycosaminoglycans.
References1 . A l l e r t o n , S . E . , B e i e r l e , J . W ., P o w a r s ,
D. R. and B a v e t t a , L . A.: Abnormal extracellular components in Wilms’ tumor. Cancer Res. 30:679-683, 1970.
2. A l l e r t o n , S . E . , E l w y n , D ., E d s a l l , J . T ., and S p a h r , P. F.: Isolation and amino acid composition of dog plasma albumin. J . Biol. Chem. 237:85-88, 1962.
3. A l l e r t o n , S . E . , P o w a r s , D. R., B e i e r l e , J. W ., C h e n , C . , a n d W i s e , K .: W i l m s ’ t u m o r

3 1 0 D E L M A G E , P O W A R S , JA Y N E S , A N D A L L E R T O N
associated proteoglycans and glycoprotein antigen. Proc. Am. Soc. Clin. Oncol. 27:292, 1976.
4. B a l a z s , E. A. and G lB B S , D. A.: The rheological properties and biological function of hyaluronic acid. Chemistry and Molecular Biology of the Intercellular Matrix, vol. 3. Balazs, E. A., ed. N ew York: A c ad e m ic P re s s , 1970, pp . 1241-1253.
5. B u r g e r , M . M . and M a r t i n , G. S.: Agglutination of cells transformed by Rous sarcoma virus by w heat germ agglutinin, Concanavalin A. Nature New Biol. 237:9-12 , 1972.
6. B u r t i n , P. and G e n d r o n , M. C.: A tumor-associated antigen in human nephroblastomas. Proc. Nat. Acad. Sci., USA, 270:2051-2054, 1973.
7. C a m e r o n , E .: H y aluron idase and C ancer. Oxford: Pergamon Press, 1966.
8. C a s t o r , C. W . and N a y l o r , B . : Acid mucopolysaccharide com position of serous effusions. Cancer 20:462-466, 1967.
9. D i F e r r a n t e , N.: Antigenicity of protein-poly- saccharide complexes from cartilage. The Chemical Physiology of Mucopolysaccharides. Quin- ta re lli, G ., ed. Boston: L ittle , Brown and Company, 1968, pp. 81-89.
10 . F o r r e s t e r , J . V. and W i l k i n s o n , P. C.: Inhibition of leukocyte locomotion by hyaluronic acid. J . Cell Science 4 8 : 3 1 5 —3 3 1 , 1 9 8 1 .
11. H A SCA LL, V. C.: Interactions of cartilage proteoglycans w ith hyaluronic acid. J. Supramo- lecular Structure 7:101—120, 1977.
12. H e l l s i n g , K.: Im m une reactions in polysaccharide media. I. The effect of dextran on the reaction betw een Iodine-125 labelled human serum albumin and gamma-globulin from rabbit anti-albumin sera. Acta Chem. Scand. 20:1251- 1262, 1966.
13. J a y n e s , P. K. and A l l e r t o n , S. E.: Glycosami- noglycan in the blood and renal tissue of a patient with nephroblastom atosis. Ann. Clin. Lab. Sci. 25:45-50, 1985.
1 4 . I s h i m o t o , N . , T e m i n , H . M . , and S t r o m i n g e r , J. L. : Studies of carcinogenesis by avian sarcoma viruses. II. Virus-induced increase in hyaluro- nate synthetase in chicken fibroblasts. J. Biol. Chem. 241:2052-2057, 1966.
15. L a u r e n t , T. L. : The exclusion of macromolecules from polysaccharide media. The Chemical Physiology of Mucopolysaccharides. Quintarelli,G., ed. Boston: Little, Brown and Company, 1968, pp. 153-170.
16 . L a u r e n t , T. C.: Structure of hyaluronic acid.E. A. Balazs (ed.), Chem istry and molecular
biology of the intercellular matrix, vol. 2. Balazs,E. A., ed. New York, Academic Press, 1970, pp. 703-732.
17. L i n d a h l , U . and H o o k , M.: Glycosaminogly- cans and their binding to biological macromolecules. Ann. Rev. Biochem. 47:385-417, 1978.
18. LlPPMAN, M.: Transplantation and cytotoxicity changes induced by acid mucopolysaccharides. Nature 219:33-36 , 1968.
19. L u f t , J. H . : Fine stru ctu re of capillary and endocapillary layers as revealed by ruthenium red. Fed. Proc. 25:1773-1783, 1966.
20. M o r s e , B. S., a n d N u s s b a u m , M .: T h e d e t e c t i o n o f h y a l u r o n i c a c id i n t h e s e r u m a n d u r i n e o f a p a t i e n t w i t h n e p h r o b l a s t o m a . A m . J . M e d . 42:996—1002, 1967.
21. N i e d e r m e i e r , W., G r a m l i n g , E., and P i g m a n , W .: Interaction of hyaluronic acid and bovine p lasm a a lb u m in . B iochem . B iophys. Acta 230:143-149, 1966.
22. O g s t o n , A. G . : Biological functions of the gly- cosam inoglycans. C hem istry and M olecular Biology o f the In te rce llu la r M atrix, vol. 3. Balazs, E. A., ed. New York, Academic Press, 1970, pp. 1231-1240.
23. O g s t o n , A . G . and P h e l p s , C. F.: The partition of solutes between buffer solutions and solutions containing hyaluronic acid. Biochem. J. 78:827— 833, 1961.
24. P i g m a n , W ., G r a m l i n g , E ., an d H o l l e y ,H . L .: In terac tions of hyaluronic acid with serum a lb u m in . B iochem . B iophys. Acta 46:100-107, 1961.
25. P o w a r s , D. R., A l l e r t o n , S. E ., B e i e r l e , J. W., and B u t l e r , B . B . : Wilms’ tumor clinical correlation with circulating mucin in three cases. Cancer 29:1597-1605, 1972.
26. SlLBERT, J. E.: Incorporation of carbon-14 and tritium from labeled nucleotide sugars into a polysaccharide in the presence of cell-free prepa ra tio n s from c a r ti la g e . J. B iol. C h em . 239:1310-1315, 1964.
27. T o o l e , B. P., O k a y a m a , M . , O r k i n , R. W., Y o s h i m u r a , L. M ., M u t o , M . , and K a j i , A.: Developmental roles of hyaluronate and chon- droitin sulfate proteoglycans. Cell and Tissue Interactions. Lash, J . W. and Burger, M . M . ed s. N ew York, R aven P re s s , 1977, pp . 139-154.
28. W u, A . H . B., P a r k e r , S. O . and Ford, L.: Hyperviscosity caused by hyaluronic acid in serum in a case of Wilms’ tumor. Clin. Chem. 30:914-916, 1984.





























