Embed Size (px)
Citation preview

The Systematics of Free Living Heterotrophic Flagellates of Beytepe Pond
Beytepe Göleti’nde Serbest Yaşayan Heterotrof Flagellatların Sistematiği
Research Article
E. E. Aydin, A. İ. Demirsoy / Hacettepe J. Biol. & Chem., 2012, Special Issue, 337-342
Esra Elif Aydin* and Ali DemirsoyDepartment of Biology, Hacettepe University, Ankara, Turkey
ÖZ E T
Bu çalışma, Türkiye tatlı sularında yaşayan heterotrof flagellat tür kompozisyonunu tespit etmek için Beytepe Göleti’nde Mayıs 2004 –Mayıs 2005 tarihleri arasında gerçekleştirilmiş bir pilot çalışmadır. Örnekler ekolojik
farklılıklarına gore tespit edilmiş dört ayrı istasyondan örnekleme şişeleri kullanılarak alınmıştır. Örnekler Leica marka, DIC ataçmanlı bir DMR mikroskop ile canlı incelenmiş ve tüm kayıtlar bu mikroskoba bağlı DFC 280 marka dijital görüntüleme sistemi ile yapılmıştır. 9’u tür, 4’ü cins düzeyinde olmak üzere toplamda 13 takson teşhis edilmiştir. Bu 13 türün 12’si Türkiye tatlı suları için yeni kayıtken, sadece 1’i fitoplankton çalışmalarında bildirilmiştir.
Anahtar KelimelerBeytepe Göleti, Serbest yaşayan Heterotrof Flagellatlar, Sistematik
A B S T R AC T
In order to investigate the species composition of the heterotrophic flagellate communities in freshwater locations in Turkey, we carried out this pilot study at Beytepe Pond from May 2004 to May 2005. Water
samples were taken from four stations determined according to ecological differences with sampling bottles. Species observed alive using a Leica DMR microscope equipped with a digital camera Leica DFC 280. As a result, 13 species were identified. Of this 13 species 9 identified to species level, 4 identified to genus level . Of the 13 species, 12 of them are new to Turkey freshwater algae, but with 1 species, having been reported from pyhtoplankton studies of Turkey Freshwaters.
Key Words Beytepe Pond, Free-Living Heterotrophic Flagellates, Systematics.
Article History: Received November 18, 2011; Revised April 01, 2012; Accepted May 3, 2012; Avaliable Online: August 10, 2012.
Correspondence to: Esra Elif Aydın, Hacettepe University, Department of Biology, Ankara, 06800, Turkey
Tel: +90 312 297 67 49 Fax: +90 312 299 20 28 E-Mail: [email protected]

E. E. Aydin, A. İ. Demirsoy / Hacettepe J. Biol. & Chem., 2012, Special Issue, 337-342338
INTRODUCTION
The term “Free-Living Heterotrophic Flagellates” refers to a group of free-living protists moving
and/or feeding through the use of flagella exclusively by heterotrophic means or, if with plastids, also capable of ingesting particles [1]. Heterotrophic flagellates form an important and diverse component of the communities living in aquatic ecosystems [2-5] as they are the predators on bacteria and small phytoplankton, prey for larger zooplankton, and facilitate remineralization and recycling of elements essential for phytoplankton and microbial growth [6-8].
Heterotrophic flagellates hold the key to the evolution of protozoa and higher organisms, and also this group plays an important role in planktonic processes, such as biochemical cycling of nutrients and carbon and the flow of energy [4]. In addition, they are the primary consumers of bacterial products in aquatic systems. Besides their special characteristics mentioned above, heterotrophic flagellates had not been taken into consideration until the last decade because of both, intrinsic and extrinsic factors [9].
The aim of this study was to conduct a pilot study to investigate the species composition of the heterotrophic flagellate communities in freshwater locations in Turkey.
MATERIALS AND METHODS
This study was carried out between May 2004 and May 2005 at Beytepe Pond, an artificial pond located at the 10th km of Ankara-Eskişehir motorway (39° 52.8’ N–32° 44.5’ E). The maximum depth of the pond is 16 m and the total surface area is 68.260 m2 changing due to irrigation. Four sampling stations were chosen according to ecological differences with the first located at the southwestern region of the pond is reedy and also where Maslak Stream flows through the pond. The second station is a bay located at the western part of the pond. The base of the pond at this station is covered with gravel and coarse stones. The third is at the sluice gate north of the pond. The fourth one is at where rain water collector is connected to the pond and is located at the southern part of the pond.
Water samples were taken with sampling bottles, placed into flasks, and kept at approximately 3°C during transportation to the laboratory. The initial volume of the sample was 250 ml, containing 200 ml of water and 50 mm3 of bottom substrate (or moss).
Heterotrophic flagellates were identified through the observation of living cells as described by Kudo [10]. Microscopic observations were made under a Leica brand DMR microscope equipped with digital interference contrast and oil immersion objectives with a total magnification of +1000. The microscope was equipped with a digital camera Leica DFC 280, connected to a computer. To take the micrographs and measurements, IM50 and QWin programs were used, respectively.
RESULTS
The organisms were reported in two categories: [1] taxa identified on species level and/or with adequate documentation, and [2] organisms which could not be identified on species level, but were nonetheless distinctive.
Category 1. Organisms identified on species level1.Hexamita inflata Dujardin, 1838 (Figure 1i-j, 2m)Cells are roundishly oval, 6–10 microns in length. Several contractile vacuoles were seen. The posterior end of the cell is rounded. There are two cytostomal tubes formed by the depression of the longitudinal cytostom and cytophariynx tubes visible below the equator and on the ventral side of the cell. Cell possesses two karyomastigonts anteriorly, each karyomastigont with 4 pairs of flagella (totally eight flagella); two medium and four short flagella about the cell length. These flagella insert anterio-laterally into a small depression on the anterior part of the cell with the long flagella normally crossing each other. The cells moved by skidding or swimming.
2. Bodo saltans Ehrenberg, 1831 (Figure 1f,2d)Cells are about 10 microns long, somewhat elliptical, and bean-shaped from lateral view. Cytostome is located subapically, with two flagella emerging from a pocket-like structure at the posterior region of

E. E. Aydin, A. İ. Demirsoy / Hacettepe J. Biol. & Chem., 2012, Special Issue, 337-342 339
the cytostome, with the anterior flagellum slightly shorter than the cell and usually directed backwards, with the non–acronematic posterior flagellum about 3 times the cell length. They are often attached to the substrate by the tip of the posterior flagellum. While attached, the flagellum bends quickly so that the cells jerk. The nucleus is located at the posterior region of the flagella pocket while the kinetoplastid is found at the postero–lateral region.
3. Rhyncomonas nasuta Klebs, 1893 (Figure 1l, 2e)Cells are 4 to 7 microns long, flattened and flexible with a bulbous motile snout which is 3 microns in length. The snout, which contains a mouth, beats slowly. The anterior flagellum lies along the snout and is hard to see, with a trailing flagellum of about 2 to 2.5x the cell length, and is acronematic. The cells feed on attached bacteria and move by gliding.
4. Entosiphon sulcatum Dujardin 1841 (Figure 1d, 2h)Cells are 20–25 µm long and similar to that of a truncated egg apically with a flagellum pocket bearing two flagella. One of the flagella is of the same length with the body and beating with undulating motion anteriorly. The other flagellum is more than twice the length of the anterior one and deflected posteriorly. Cytopharinyx is located at the lateral side of the flagellum pocket. At the posterior side of the cytopharinyx, a single contractile vacuole is located which empties into the base of the flagellum pocket. At the dorsal side of the cytopharynx and the pocket, there is an ingestion organelle composed of rods fused together to make a protrusible tube (siphon) which can make pumping movement and a longitudinal fluting and ridge.
5. Entosiphon obliquum Dujardin, 1841 (Figure 1g, 2b)Cells are about 15 µm long and similar to that of a tapered egg, both at anterior and posterior regions, with a flagellum pocket located apically bearing two flagella. The anterior flagellum is of the same length with the body and beating with undulating motion anteriorly while the other flagellum is 1.5x the length of the body and provides rotational movement. There is an ingestion tube running along the body and a contractile vacuole at the posterior region of the flagellum pocket.
6. Astasia lagenula Schew (Figure 1a, 2a)Cells are about 20 µm in length with the cell body being elongate with a rounded posterior end and plastic with euglenoid movement. A cell has a flagellum pocket located apically bearing a flagellum almost of the cell length. The flagella emerge in a sharp-edged region. A cell is with ovoidal paramylum granules and somewhat with a large reservoir. The nucleus is more or less central.
7. Astasia conica Matvienko, 1938(Figure 1b,2c)Cells are 17–25 µm in length, with the body like a wedge or comma shaped while swimming, and cylindrical tapering posteriorly and ending stubbly but not sharp like a needle while resting. The cell has a flagellum pocket located apically bearing a flagellum which is of the same length with the cell. The flagella emerge from a region similar to a bent slit, apical roundishly oval. At the anterior end of the cell, large ovoidal paramylum granules are located.
8. Distigma curvatum Pringsh, 1936 (Figure 1c, 2g)Cells are 20–25 µm in length; rounded at cross section, anterior end also rounded and posterior is pointed. Flagellar canal opens apically. The anterior flagellum is about 1.0 cell length and the recurrent flagellum is about 0.2 cell length visible only on movement. The anterior part of the cells contains large, roundish or cylindrical refractile granules while the posterior part contains a large number of very small refractile granules.
9. Chilomonas paramecium Ehrenberg, 1831 (Figure 1e, 2i)Cells are 19–30 µm in length and elongate-ovate in shape. Two subequal flagella which are shorter than the length of the body are located in an anterior cage surrounded by rows of extrusomes. Small extrusomes are also visible under the cell membrane. The nucleus is located posteriorly to the cage. A contractile vacuole is located anteriorly and empties into the cage. Cells contain refractile stored substances and often two refractile Maupas bodies. Cells rotate while swimming, and can also swim backwards.
Category 2: Taxa unassignable to species Organisms from four other genera were observed but they could not be identified on species level.
E. E. Aydin, A. İ. Demirsoy / Hacettepe J. Biol. & Chem., 2012, Special Issue, 337-342340
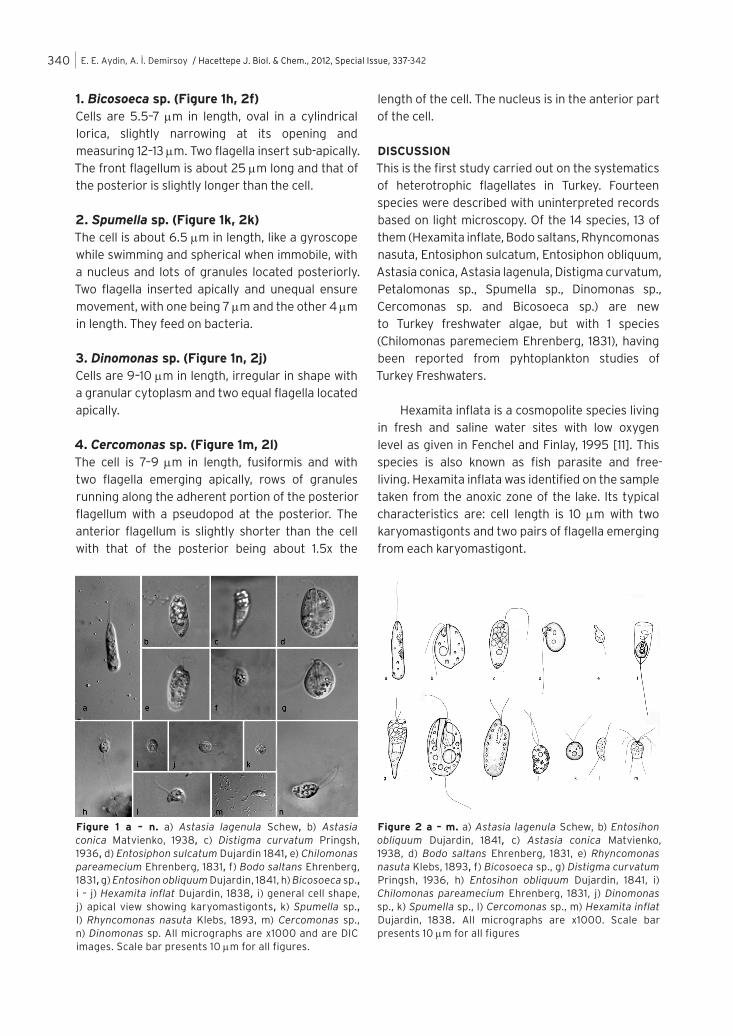
1. Bicosoeca sp. (Figure 1h, 2f)Cells are 5.5–7 µm in length, oval in a cylindrical lorica, slightly narrowing at its opening and measuring 12–13 µm. Two flagella insert sub-apically. The front flagellum is about 25 µm long and that of the posterior is slightly longer than the cell.
2. Spumella sp. (Figure 1k, 2k)The cell is about 6.5 µm in length, like a gyroscope while swimming and spherical when immobile, with a nucleus and lots of granules located posteriorly. Two flagella inserted apically and unequal ensure movement, with one being 7 µm and the other 4 µm in length. They feed on bacteria.
3. Dinomonas sp. (Figure 1n, 2j)Cells are 9–10 µm in length, irregular in shape with a granular cytoplasm and two equal flagella located apically.
4. Cercomonas sp. (Figure 1m, 2l)The cell is 7–9 µm in length, fusiformis and with two flagella emerging apically, rows of granules running along the adherent portion of the posterior flagellum with a pseudopod at the posterior. The anterior flagellum is slightly shorter than the cell with that of the posterior being about 1.5x the
length of the cell. The nucleus is in the anterior part of the cell.
DISCUSSION
This is the first study carried out on the systematics of heterotrophic flagellates in Turkey. Fourteen species were described with uninterpreted records based on light microscopy. Of the 14 species, 13 of them (Hexamita inflate, Bodo saltans, Rhyncomonas nasuta, Entosiphon sulcatum, Entosiphon obliquum, Astasia conica, Astasia lagenula, Distigma curvatum, Petalomonas sp., Spumella sp., Dinomonas sp., Cercomonas sp. and Bicosoeca sp.) are new to Turkey freshwater algae, but with 1 species (Chilomonas paremeciem Ehrenberg, 1831), having been reported from pyhtoplankton studies of Turkey Freshwaters.
Hexamita inflata is a cosmopolite species living in fresh and saline water sites with low oxygen level as given in Fenchel and Finlay, 1995 [11]. This species is also known as fish parasite and free-living. Hexamita inflata was identified on the sample taken from the anoxic zone of the lake. Its typical characteristics are: cell length is 10 µm with two karyomastigonts and two pairs of flagella emerging from each karyomastigont.
Figure 1 a – n. a) Astasia lagenula Schew, b) Astasia conica Matvienko, 1938, c) Distigma curvatum Pringsh, 1936, d) Entosiphon sulcatum Dujardin 1841, e) Chilomonas pareamecium Ehrenberg, 1831, f) Bodo saltans Ehrenberg, 1831, g) Entosihon obliquum Dujardin, 1841, h) Bicosoeca sp., i – j) Hexamita inflat Dujardin, 1838, i) general cell shape, j) apical view showing karyomastigonts, k) Spumella sp., l) Rhyncomonas nasuta Klebs, 1893, m) Cercomonas sp., n) Dinomonas sp. All micrographs are x1000 and are DIC images. Scale bar presents 10 µm for all figures.
Figure 2 a – m. a) Astasia lagenula Schew, b) Entosihon obliquum Dujardin, 1841, c) Astasia conica Matvienko, 1938, d) Bodo saltans Ehrenberg, 1831, e) Rhyncomonas nasuta Klebs, 1893, f) Bicosoeca sp., g) Distigma curvatum Pringsh, 1936, h) Entosihon obliquum Dujardin, 1841, i) Chilomonas pareamecium Ehrenberg, 1831, j) Dinomonas sp., k) Spumella sp., l) Cercomonas sp., m) Hexamita inflat Dujardin, 1838. All micrographs are x1000. Scale bar presents 10 µm for all figures

E. E. Aydin, A. İ. Demirsoy / Hacettepe J. Biol. & Chem., 2012, Special Issue, 337-342 341
Bodo saltans is the most common species of the bodo genus. It is noticed that this species lives in fresh, marine and brackish waters. Despite the poor initial description [12], Bodo saltans is a kinetoplastid, distinguished with its typical kicking movement ensured by the long flagellum while feeding [13].
Rhynchomonas nasuta had previously been reported from marine, soil and freshwater habitats in a wide variety of locations worldwide [14-17]. It is distinguished from other gliding flagellates by its mobile bulbous snout. Only a single, thickened flagellum is easily visible on light microscopy [15].
Species belonging to the genus Entosiphon are distinguished by the body shape, the rate between the long flagellum and the length of the body. Entosiphon sulcatum is distinguished from the Entosiphon obliquum with its truncated egg-like shape and pointed posterior region. Its actively swimming flagellum is the same length as the body and trailing posteriorly and is longer than the body length. That is why it differs from the Entosiphon striatum and Entosiphon ovatum. Entosiphon obliquum is distinguished from Entosiphon polyaulax with its active flagellum being as long as the body length and the flagellum trailing posteriorly’s length being 1.5x of the body length. [18]. Species belonging to the genus Entosiphon have a great radiation at freshwater sites and are confused with the species of the genus Anisonema. But they differ by the presence of the digestion tube on Entosiphon genus.
Species of genus Astasia are distinguished from each other with the shape of the region from where flagellum emerges, shape of the body, length of flagellum, the presence of starch granules in the cell and, if present, the shape and the number of these granules. The region from where flagellum emerges in Astasia lagenula and Astasia conica is not parallel but in both species, this region is like a roundish crest and without bends. Both species have starch granules. The starch granules of Astasia lagenula are small droplets while Astasia conica’s located anteriorly, are considerably large [18]. Species of the genus Astasia are often identified with freshwaters.
A rain collector was connected with the pond by ASKI (Ankara Water and Sewage Management) and as confirmed by Ponds Department Authority, the water brought by this collector causes fecal pollution. According to the analyses done, Escherichia coli has been identified in the pond. Chilomonas paramecium, an organic material indicator, exists in aquatic systems densely polluted as notified by Calaway and Lackey, 1962 [19].
Until now, 20 species belonging to the genus Chilomonas mostly living in freshwaters has been identified. Descriptions of these 20 species are insufficient as they differ from each other according to body length and shape. The species identified are mostly synonyms of Chilomonas paramecium [20].
The genus Bicosoeca, consist of 38 species, 30 of which live only in freshwaters, 2 in both brackish and freshwaters, 1 marine and fresh waters [21]. Species of the genus Bicosoeca, are distinguished from each other by the shape and the length of lorica [22]. Since the whole literature of the 33 species could not be reached, it was not possible to identify these taxa at species levels.
Spumella genus consists of 6 freshwater species. The characteristics of the genus are, lack of chloroplast, naked body surface and the possession of two flagella; one long and the other too short [19]. It is one of the most common species in freshwaters. [21]. As sufficient descriptions could not be reached, it was not possible to identify this taxa at species level.
Species of the genus Cercomonas have great radiation and also very common in marine, freshwater and soil. All are bacteriovorus [23]. The species of the genus are distinguished from each other by the arrangement of granules in the body and also, the length of the body, too [5]. Since the whole literature of the 33 species could not be reached, it was not possible to identify this species at species level.
It is noticed that the species of the Dinomonas genus, both live in marine and freshwaters [23]. There is no comprehensive study on this genus. That was why it could not be identified at species level.

E. E. Aydin, A. İ. Demirsoy / Hacettepe J. Biol. & Chem., 2012, Special Issue, 337-342342
There have been intensive taxonomic studies on free-living heterotrophic flagellates in marine ecosystems [5, 15-17, 19, 22, 24, 25] in sharp contrast to freshwater flagellates. Moreover, species-based literature is distributed in original papers and focused on reviews (e.g. 18) with some of the literature not reviewed for over a century. As a result of this, some of the descriptions of heterotrophic flagellate species insufficient and identifying them at species level is difficult. More literature must be collected to make a more effective systematic study.
Thus, this study therefore, becomes important, being the pilot work for further systematic studies of heterotrophic flagellates for more detailed ecological research of aquatic systems in the future.
R E F E R E N C E S
1. D.J. Patterson, J. Larsen (eds.), The Biology of Free-living Heterotrophic Flagellates, Systematic Association, Oxford, (1991) 503.
2. F. Azam, T. Fenchel, J.G. Field, J.S. Gray, L.A. Meyer-Reil, F. Thingstad, The ecological role of water-column microbes in the sea, Marine Ecology Progress Series, 10 (1983) 257.
3. D.J. Patterson, J.Larsen, J.O. Corliss, The ecology of heterotrophic ciliates and flagellates living in marine sediments,Prog. Protistol., 3 (1989) 185.
4. J. Laybourn-Parry, J. Parry, Flagellates and the microbial loop. In: Leadbeater BSC, Green JC (eds) The flagellates, Taylor & Francis, London, (20009) 216.
5. W.J. Lee, D.J. Patterson, Heterotrophic flagellates (Protista) from marine sediments of Botany Bay, Australia, J. Nat. Hist., 34 (2000) 483 .
6. E.B.B.F Sherr, B.F. Sherr, Roles of micropes in pelagic food webs: a revides concept, Limnol. Oceanogr., 33 (1988) 1225.
7. D.L. Kirchman, Particulate detritus and bacteria in marine environments. In Microbiology–An ecological approach, T. E. Ford., Editor , Blackwell Scientific Publications, Boston, (1994) 321.
8. M.L. Pace, D. Vaqué, The importance of Daphnia in determining mortality rates of protozoans and rotifers in lakes, Limnology, 39 (1994) 985.
9. W.J. Lee, D.J. Patterson, Abundance and biomass of heterotrophic flagellates, and factors controlling their abundance and distribution in sediments of Botany Bay., Microbial Ecology, 43 (2002) 467.
10. R.R. Kudo, Protozoology, Fifth Edition, Charles C. Thomas Publisher, USA, (1966).
11. G.A. Biagini, K. Kirk, P.J. Schofield, Role of K+ and amino acids in osmoregulation by the free-living microaerophilic protozoon Hexamita inflata, Microbiology, 146 (2000) 427.
12. C.G. Ehrenberg, Über die Entwickelung und Lebensdauer der Infusionsthiere; nebst ferneren Beiträgen zu einer Vergleichung ihrer organischen Systeme. Abhandlungen der Königlichen Akademie Wissenschaften zu Berlin, Physikalische Klasse, (1831)1.
13. B.F. Zhukov, The diversity of bodonids, in The Biology of Free-living Heterotrophic Flagellates, J.D. Patterson, J. Larsen (eds.), Syst. Assoc., Oxford, 1991, 117.
14. M.L. Pace, D. Vaqué, The importance of Daphnia in determining mortality rates of protozoans and rotifers in lakes, Limnology, 39 (1994) 985.
15. J. Larsen, D.J. Patterson, Some flagellates (Protista) from tropical sediments, J. Nat. Hist., 24 (1990) 801.
16. D.J. Patterson, A.G.B. Simpson, Heterotrophic flagel-lates from costal marine and hypersalines sediments in Western Australia, Europ. J. Potistol., 32 (1996) 423.
17. N. Vørs, Heterotrophic amoebae, flagellates and Heliozoa from Arctic marine waters (North West Teritories, Canada and West Greenland), Polar Biology, 13 (1992) 113.
18. G. Huber–Pestalozzi, Das Phytoplankton des Süswassers, 4. Teil, Euglenophyceen, in Die Binnengewasser, Schweizerbart, Stuttgart, (1955) 607.
19. C. Bernard, A.G.B. Simpson, D.J. Patterson, Some free-living flagellates (Protista) from anoxic habitats, Ophelia, 52 (2000) 113.
20. D.R.A. Hill, Diversity of heterotrophic crytomonads, in The Biology of Free-living Heterotrophic Flagellates, D.J. Patterson, J. Larsen (eds.), Systematic Association, Oxford, (1991) 235.
21. H.R. Preisig, N. Vørs, G. Hällfors, Diversity of heterotrophic heterokonts, in The Biology of Free-living Heterotrophic Flagellates, D.J. Patterson, J. Larsen (eds.), Systematic Association, Oxford, (1991) 361.
22. M.S. Tong, Heterotrophic flagellates and other protists from Southhampton Water, U.K., Ophelia, 47 (1997) 71.
23. D.J. Patterson, M. Zölffel, Heterotrophic flagellates of uncertain taxonomic position, in The Biology of Free-living Heterotrophic Flagellates, D.J. Patterson, J. Larsen (eds.), Systematic Association, Oxford, (1991) 427.
24. S.M. Tong, Heterotrophic flagellates from the water column in Shark Bay, Western Australia. Mar. Biol., 128 (1997) 517.
25. S.M. Tong, N. Vørs, D. J. Patterson, Heterotrophic flagellates, centrohelid heliozoa and filose amoebae from marine and freshwater sites in the Antarctic. Polar Biol., 18 (1997) 91.





























