Embed Size (px)
Citation preview

J. Cell Sci. 3 ) 161-174 (1968) 161
Printed in Great Britain
THE ULTRASTRUCTURE AND ONTOGENY OF
POLLEN IN HELLEBORUS FOETIDUS L.
I. THE DEVELOPMENT OF THE TAPETUM ANDUBISCH BODIES
P. ECHLIN AND H. GODWINElectron Microscope Laboratory, Botany School, University of Cambridge
SUMMARY
The ontogeny of the tapetum and Ubisch bodies in Helleborus foetidus L. has been examined atthe ultrastructural level, and their development has been closely linked with that of the sporo-genous cell and pollen grains. During development the tapetum passes through successivephases of synthesis, maturity and senescence, ending in complete dissolution. During theanabolic phase of growth, precursors of the Ubisch bodies are formed as spheroidal vesicles ofmedium electron density within the tapetal cytoplasm; they are associated with a zone of radiat-ing ribosomes, which, as development proceeds, can clearly be seen to be situated on strands ofendoplasmic reticulum. The callose special wall round the microspores and the tapetal cell wallnow disintegrate and the pro-Ubisch bodies are extruded through the cell membrance of thetapetal cells, where they remain on the surface of the anther cavity and soon become irregularlycoated with sporopollenin. Deposition of sporopollenin continues on the Ubisch bodies at thesame time as upon the exines of the developing pollen grains. In both cases, the later stages ofsporopollenin deposition are associated with electron-transparent layers of unit-membranedimensions appearing in section as white lines of uniform thickness. Continuing deposition ofsporopollenin leads to the formation of compound or aggregate Ubisch bodies. It is conjecturedthat the sporopollenin is synthesized from the compounds of low molecular weight released intothe anther loculus by the breakdown of the callose special wall and the tapetal cell wall. Thefinal stages of tapetal autolysis involve the disappearance of all the cell organelles. An attemptis made to relate the findings to those described in other recent studies on Ubisch body forma-tion and to combine them in a common ontogenetic pattern.
INTRODUCTION
In the developing anthers of vascular plants the microspores are surrounded by thetapetum, which consists of one or more cell layers characterized by densely stainingprotoplasts and prominent nuclei. The tissue plays an essential nutritive role in theformation of the pollen grains and attains its maximal development at the tetrad stageof microspore growth. Two principal types of tapetum are recognized: the glandularor secretory, in which the cells give up their contents to the microspores, and theamoeboid or plasmodial type, in which the cell walls break down and the protoplastsform a peri-plasmodium around the developing pollen grains.
The works of Schnarf (1923), Clausen (1927), Cooper (1933), Witkus (1945),Maheshwari (1950) and Carniel (1963) contain numerous references to both the de-velopment of the tapetum and the aberrant nuclear divisions in this tissue. Thispresent study will be concerned with the ultrastructure of the tapetum in relation to

162 P. Echlin and H. Godwin
nutrition of the developing pollen grains, including the ontogeny of their exines, andthe concomitant development of the Ubisch bodies.
Ubisch bodies are spheroidal structures found in the anthers of many genera ofangiosperms, both monocotyledons and dicotyledons, and of some gymnosperms.They occur in large numbers on the walls of the tapetal cells, especially those liningthe anther loculus. They are generally only a few microns in diameter, but frequentlyfuse into larger compound aggregates in the later stages of their development. In someinstances such aggregates constitute platelets extending far across the tapetal cellsurface and crossing the cell boundaries. The walls of the Ubisch bodies apparentlyconsist of sporopollenin, the main constituent of the mature microspore exine, amaterial of conspicuous durability and characteristic resistance to acetolysis. Themature wall is usually thick in proportion to the total size of the Ubisch body, andalthough generally uniform in thickness, it is sometimes verrucose or spinose, soexhibiting greater or less similarity to the pollen grain exine. Extremely useful generalaccounts of the morphology and distribution of Ubisch bodies in the plant kingdomhave been given by Ubisch (1927), Kosmath (1927) and more recently by Davis (1966).From their work it seems that Ubisch bodies are produced by the glandular or secre-tory type of tapetum and not by the amoeboid. With the final autolysis of the tapetalcells in the later stages of sporogenesis, the Ubisch bodies tend to lie irregularly uponthe remnants of the tapetum next to the fibrous endothecium, and amongst thematuring pollen-grains, especially on the outer side of the thecae.
The deposition of patterned sporopollenin on the walls of sterile pollen grainsdeficient or lacking in nuclear material had already attracted attention. Tischler (1908)found that in spite of a degeneration of the protoplasm in pollen grains of Mirabilisjalapa the exine patterning developed normally, and more recently Heslop-Harrison &Heslop-Harrison (1958) demonstrated normal exine sculpturing in male-sterile Silenependula. It would seem that the Ubisch bodies represent the capacity for organizingsimilar, but less complex exinous systems by smaller cytoplasmic units. The Ubischbodies were inconveniently small for investigation solely by light microscopy but withthe advent of the electron microscope they were easily re-identified and it becamepossible to consider the details of their morphology and ontogeny along with that ofmicrospore exine. Several morphological descriptions have been made at the electron-microscopical level (see, for example, Rowley, Miihlethaler & Frey-Wyssling, 1959;Rowley, 1962, 1964; Chambers & Godwin, 1961; Yamazaki & Takeoka, 1962; Dahl &Rowley, 1965), but the only ontogenetic studies so far available are for Cannabis sativaand Silene pendula (Heslop-Harrison, 1962, 1963), Poa annua (Rowley, 1963), Zeamays (Skvarla & Larsen, 1966), and Zea mays (J. Skvarla, private communication): tothese accounts we shall refer in the general discussion.
Our own study describes the fine structure and development of the tapetal cellsand Ubisch bodies during microsporogenesis in Helleborus foetidus L., making use oforganic aldehydes to obtain as high a standard of fixation of protoplast structures aspossible. A preliminary report on this work was presented elsewhere (Echlin, Godwin,Chapman & Angold, 1966a).

Pollen of Helleborus. I 163
MATERIALS AND METHODS
Intact flowers of Helleborus foetidus L. (Stinking Hellebore) were removed fromplants grown in a Cambridge garden, and the stage of development of the anthersestablished by light-microscopic examination of aceto-carmine squashes. Intactanthers of the required stage were removed from the flowers, immediately placed in 5 %glutaraldehyde in o-i M Sorenson's phosphate buffer (pH 7-0) and cut in half. Theanthers remained in the fixative for 15 h at 4 °C. The tissue was then thoroughlywashed in Palade's veronal-acetate buffer (pH 7-0) and post-fixed for 2 h at 4 °C in 1 %osmium tetroxide in Palade's veronal-acetate buffer (pH 7-0). The specimens weredehydrated in a graded ethanol series, passed through three changes of 1,2-epoxy pro-pane and embedded in Araldite without plasticizer. Thin sections were briefly stainedwith either lead citrate or potassium permanganate, dried, coated with a thin layer ofevaporated carbon, and examined in an AEI EM 6 electron microscope.
RESULTS
In describing the ontogeny of the tapetum and Ubisch bodies it will be necessary torelate the development to a known time sequence. Study of microsporogenesis inH. foetidus has revealed that development proceeds through a series of stages recogniz-able at both the light- and electron-microscope levels. The sequence of events observedin the tapetum will be related to the development of the pollen grains, including thestages of construction of the mature exine, with its bacula, tectum and foot layer(see Echlin et al. 1966a).
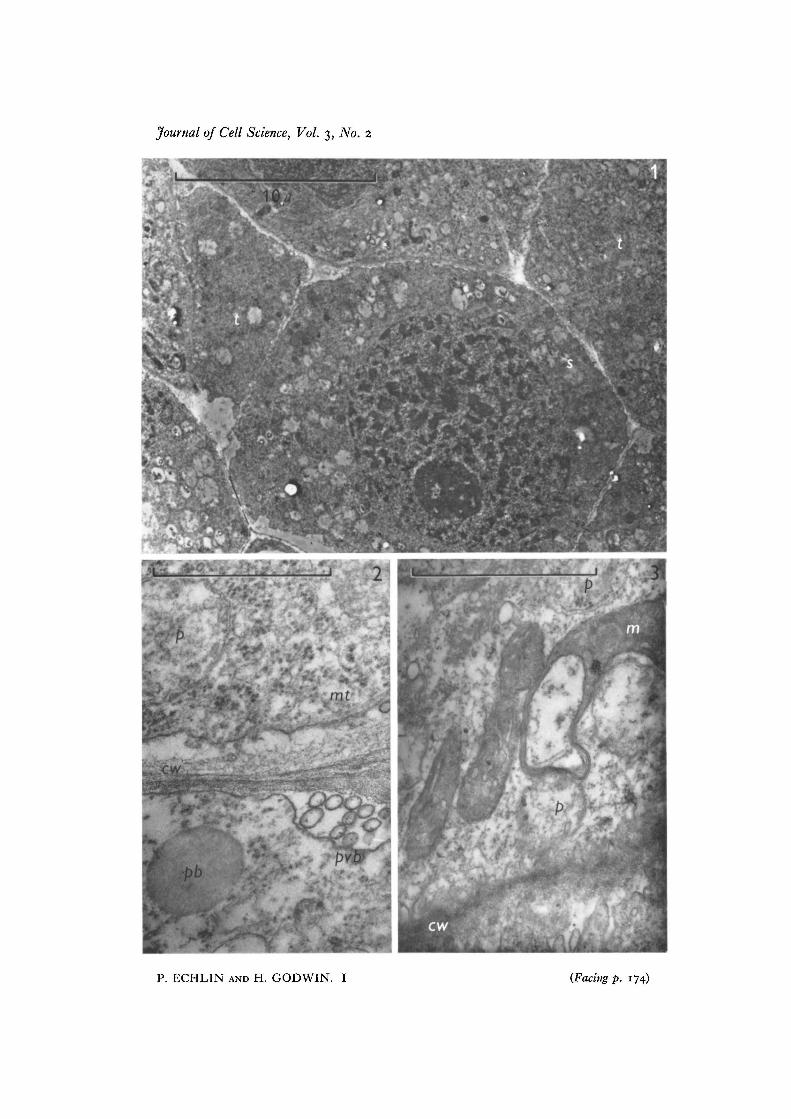
The earliest recognizable stage is of sporogenous tissue surrounded by activelydividing tapetal cells (Fig. 1). The tapetal cells, which form a layer one or two cellsthick, have prominent nuclei and nucleoli which tend to be in the half of the cellnearest to the developing sporogenous tissue. Cell organelles are not readily recogniz-able at this stage, but it is possible to see mitochondria (Fig. 3) which are characterizedby electron-dense contents, plastids with relatively electron-transparent contents(Figs. 2, 3) and a number of 'grey bodies', or pro-Ubisch bodies (Fig. 2). The tapetalcytoplasm also contains dictyosomes with only a few peripherally associated vesicles,electron-dense ribosomes and occasionally a few profiles of smooth endoplasmicreticulum. Starch grains are transitorily present in the tapetal cells at this stage, thoughthey are a more permanent feature of the endothecial cells which form a layer betweenthe tapetum and the anther epidermis. Reserve proteins in the form of 'proteoplasts'as described by Py (1932) have not been seen in the tapetum. The tapetal cell walls arerelatively thin at this stage and appear to be composed of middle lamella with a smallamount of cellulosic primary wall. Microtubules are frequently seen in the corticalcytoplasm close to the developing cell walls (Figs. 2, 3) and polyvesicular bodies (seeEchlin, Godwin, Chapman & Angold, 19666) may be found associated with the tapetal/sporogenous tissue cell walls (Fig. 2) and with the tapetal/endothecium cell walls.
The sporogenous tissue gives rise to the pollen mother cells, and by this stage thetapetal/pollen mother cell wall has become thicker and the tapetal cytoplasm appears

164 P. Echlin and H. Godwin
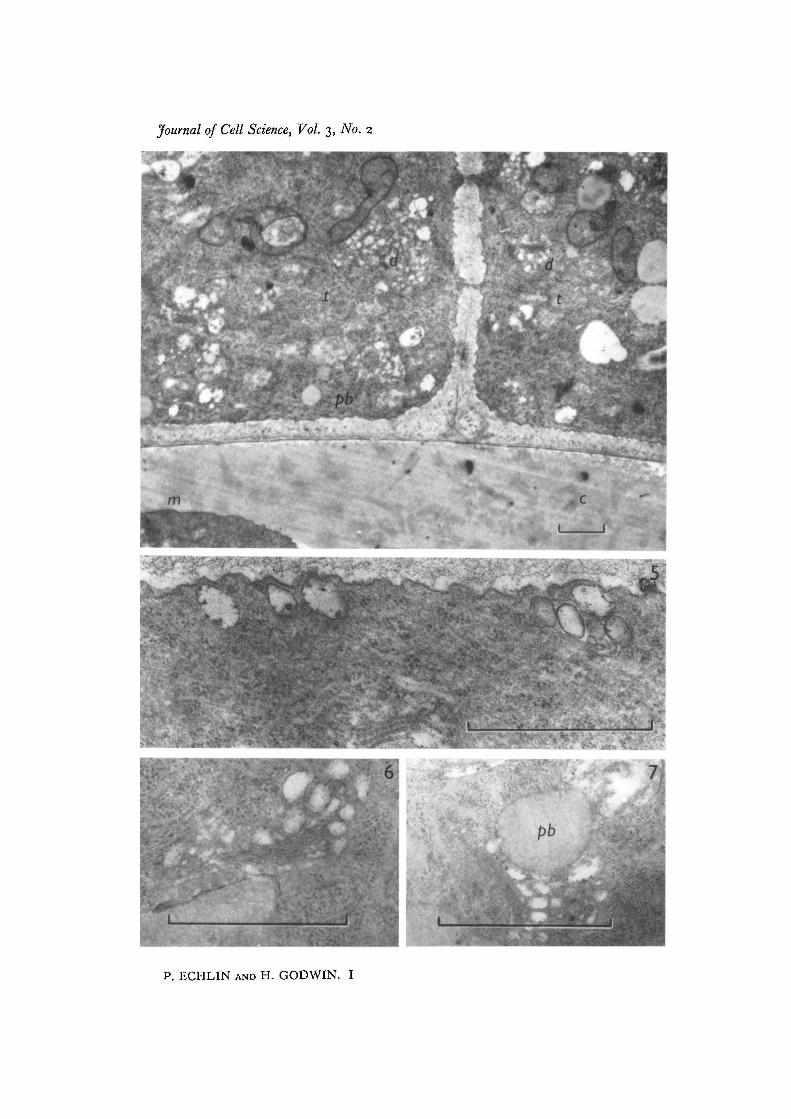
more dense due to the increased number of ribosomes and pro-Ubisch bodies. Thethickening of the tapetal/anther cavity cell wall is irregular, more cellulose being de-posited on the tapetal side of this wall. The material deposited on the tapetal side hasa loosely fibrous appearance, whereas the wall components nearest the anther cavityare far more compacted (Fig. 4). The pollen mother cell undergoes meiotic division toform the tetrad of 4 haploid microspores which briefly remain closely apposed to eachother. At this stage in microspore development, the tapetal nuclei and cell walls havefurther increased in size and there has been an increase in the number of pro-Ubischbodies in the tapetal cytoplasm. The pro-Ubisch bodies, although distributed through-out the tapetal cytoplasm, appear to be in greater numbers in the sector of the cellnearest the anther cavity. The endoplasmic reticulum, which until this time has beenonly just visible, is now more apparent and may be seen with and without ribo-somes.
At the time of late tetrad formation the 4 microspores, which by now are completelyseparate, are enveloped in thick layers of callose (Fig. 4), and the tapetal/anther cavitycell wall is now thicker by a factor of 2-5 than the original tapetal/sporogenous cell wall.Numerous plasmodesmata are found between adjoining tapetal cells (Fig. 4). No suchconnexions have been seen between the tapetum and the anther cavity. There appearto be fewer plastids and microtubules and, although not immediately apparent, anincreased number of the ribosomes appear to be associated with the endoplasmicreticulum (Fig. 7). We find the endoplasmic reticulum is particularly difficult todemonstrate adequately at this stage, an experience shared with other workers usingorganic aldehydes as fixatives. It is thought that the preservation of some of the morelabile cellular constituents has caused them to mask the endoplasmic reticulum. Otherfixatives have been used, and although they clearly reveal endoplasmic reticulum theydo so at the expense of the less robust cell components.
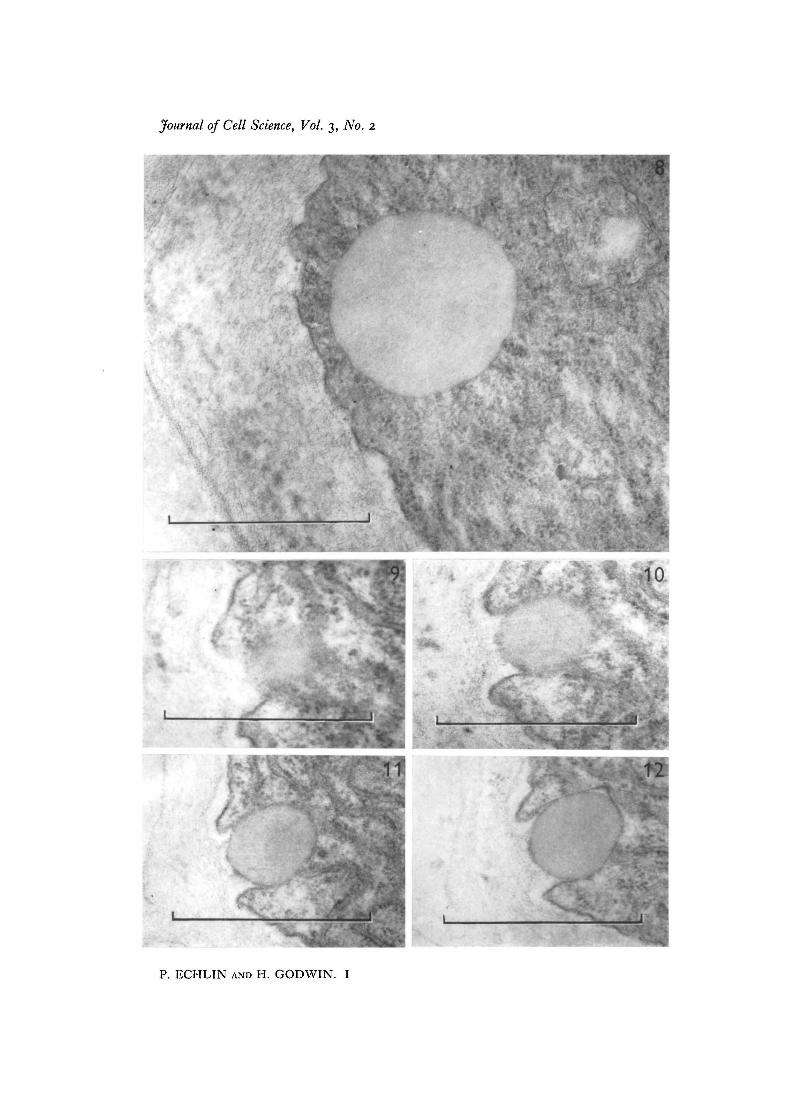
At this stage, two components of the tapetal cytoplasm merit special attention.Concomitant with the development of the microspore, the pro-Ubisch bodies whichhave appeared in increasing numbers in the tapetal cytoplasm now appear to be sur-rounded by a zone of ribosomes, which radiate from the pro-Ubisch bodies like thespokes of a wheel (Figs. 4, 7, 8). The limiting membrane of the pro-Ubisch body isvisible only when cut in median section, and is usually discontinuous at those placeswhich correspond to the insertion of the rays of radiating ribosomes (Fig. 8). The pro-Ubisch bodies are of various sizes and are more frequently found in the cortical cyto-plasm, either singly or in groups. The other cellular organelles which warrant attentionare the dictyosomes, which, until this stage of development, have been relativelyinactive. There now appear to be a large number of vesicles at the periphery of thedictyosomes (Figs. 5-7) and in some views they appear to radiate around the organelle(Fig. 4). In some instances the vesicles associated with the periphery of the dictyosomesare themselves enclosed within a larger vesicle. The dictyosomes and peripheralvesicles have not been found associated with the pro-Ubisch bodies. The tapetal cellmembrane is now highly convoluted and many of the enclosures made by the convolu-tions are equivalent in size and appearance to the electron-transparent vesicles im-mediately below the cell surface (Fig. 5). This would suggest that these vesicles, which

Pollen of Helleborus. I 165
appear morphologically identical to the vesicles at the periphery of the dictyosome,are liberated into the anther cavity by a process of membrane fusion and extrusion.
Soon after the microspores have separated from one another, and while they arestill enclosed within their special callose walls, the first signs of primexine (the pre-cursor or template of the main exinous wall component) appear on the surface of thedeveloping pollen grains.
The ribosomal zone surrounding the pro-Ubisch bodies is now even more apparent,and the cortical regions of the tapetal cytoplasm contain abundant dictyosomes. Thetapetum/anther cavity cell wall is now less electron-dense and the fibrous componentshave begun to disappear, leaving an irregular array of electron-dense granules in theouter regions of the wall (Fig. 13).
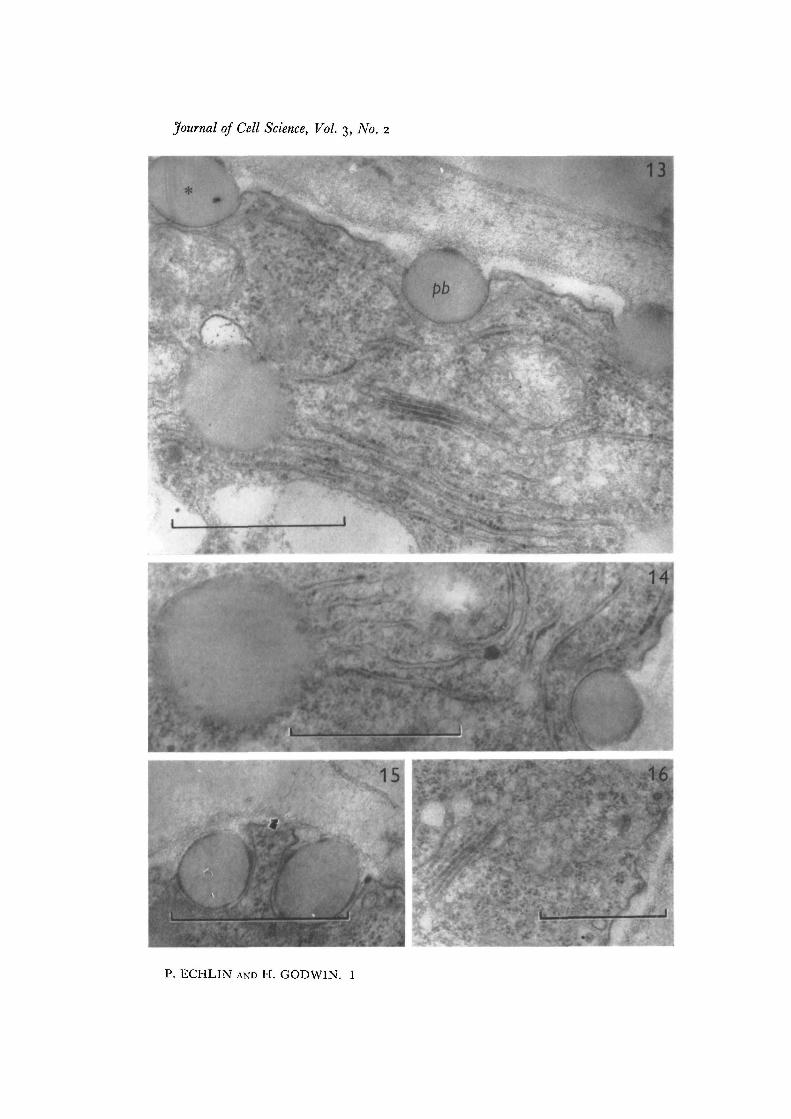
During the formation of recognizable exinous components (bacula and tectum) inthe pollen grain wall, the tapetum shows further distinctive changes. The endoplasmicreticulum is now recognizable, and both smooth and rough profiles may be seenrunning approximately parallel to the cell surface, in close association with the pro-Ubisch bodies (Fig. 13). There appears to be open continuity between the lumen ofthe pro-Ubisch bodies and the cisternae of the endoplasmic reticulum; the materialof both is of similar electron density and structure (Fig. 14). This appearance suggeststhat both the ribosomes and endoplasmic reticulum are involved in the metabolism ofthe material of the pro-Ubisch body. There are many more pro-Ubisch bodies at thisstage than previously and they can be seen to become incorporated into, and eventuallyto pass through, the tapetal cell membranes (Figs. 9-14). In no instance has it beenpossible to find a break in the tapetal cell membrane, and it is suggested that duringextrusion the membrane surrounding the pro-Ubisch body fuses with the tapetal cellmembrane. Once the pro-Ubisch bodies have been extruded from the tapetal cyto-plasm into the space between the membrane and the cell wall, they become investedwith a layer of electron-dense material which appears in some instances to be depositedon both sides of an electron-transparent layer of unit-membrane dimensions (Figs. 13-15). This electron-dense material is identical with the appearance and structure of theexinous material being formed at this time at the surface of the pollen grain. The layeris deposited only after the pro-Ubisch body has been released from the cytoplasm, andhas never been observed while the body is still within the cell. Some micrographs haveshown material of similar electron density apparently within the cytoplasm, but in allinstances this can be attributed either to non-specific, electron-dense lipid material, orto tangential sections through the surface of the tapetal cell.
The endoplasmic reticulum and ribosomes appear to be the only organelles asso-ciated with the pro-Ubisch bodies, although the mitochondria are abundant in thisregion and often closely apposed to the endoplasmic reticulum (Figs. 13, 18). Noelectron-dense material has been found accumulating in the mitochondria, and, on thecontrary, as development of the tapetum proceeds there is a gradual diminution indensity of this organelle. The ribosomes at this stage are abundant and are either asso-ciated in groups or arranged along the endoplasmic reticulum (Figs. 16-18). Thedictyosomes continue to appear active and numerous vesicles are seen at the end of thecisternae (Fig. 16). Vesicles of similar size and electron density may also be found at

166 P. Echlin and H. Godwin
the tapetal cell surface, either singly or in groups contained within a common mem-brane (Figs. 16,17). The larger electron-transparent vesicles or regions found randomlydistributed throughout the tapetal cytoplasm are thought to be a result of the gradualdissolution of either mitochondria, plastids, or both. Plastids have become pro-gressively less evident as development has proceeded.
The common cell wall between tapetum and anther cavity now appears considerablyless electron-dense and there is a space between it and the cell membrane. Theperipheral tapetal cytoplasm is now devoid of microtubules and the nucleus still re-mains intact, although there is considerable lobing of the nuclear membrane.
By this stage in development the exine of the pollen grain has reached a stage atwhich the first formed bacula (columellae) are being supplemented by the depositionof either a foot layer, or incipient endexine of similar density. In some instances theformation of this layer coincides with the appearance in the developing pollen-grainwall of an electron-transparent region of unit-membrane dimension—the 'whitelines' referred to by Godwin, Echlin & Chapman (1967). The pro-Ubisch bodiescontinue to be formed within the tapetal cytoplasm and to be extruded from the cells(Fig. 19), more electron-dense material is deposited on them, the deposition initiallybeing greater on the side nearest the anther loculus (Fig. 20). The increasing densityof the deposits on the surface of the pro-Ubisch bodies parallels the increase in densityof the exine of the pollen-grain wall (Fig. 19). The morphology of the spheroids at thesurface of the tapetum is very similar to that of the Ubisch bodies described by otherworkers and seen by ourselves in other species so that it is now apparent that the pro-Ubisch bodies are truly the precursors of Ubisch bodies. The electron-dense materialcan be shown to be resistant to acetolysis and is in this regard identical to sporopolleninwhich has already been established as being one of the major components of the pollen-grain wall exine. What may be called 'multiple' Ubisch bodies (Figs. 21, 23) may beseen where a series of small pro-Ubisch bodies begins to form around a large pro-Ubisch body, the whole complex becoming progressively enveloped with electron-dense material.
All the tapetal cell walls have by this stage virtually disappeared and only a thinlayer remains (Figs. 21, 24). The rest of the tapetal cytoplasm appears normal, withthe prominent nucleus and nucleoli occupying a more central position in the cell andsurrounded by endoplasmic reticulum, ribosomes and dictyosomes (Fig. 21). Thepollen-grain wall is now nearly fully formed, and intine begins to appear below thepores.
Further development of the Ubisch bodies mainly involves an increase in thicknessof the electron-dense material around the pro-Ubisch bodies (Fig. 22). In no instanceshave any narrow electron-transparent channels been found in the dense walls of theUbisch bodies, although a number of different fixatives have been tried, includingpermanganate, which other workers have found specially apt to reveal the presence ofnarrow channels. The density and apparent rate of deposition of sporopollenin con-tinues closely to parallel the deposition of exinous material on the pollen grain. At thisstage in the development of the pollen-grain wall, the method of exine depositionchanges and proceeds around those elements of unit-membrane dimensions described

Pollen of Helleborus. I 167
by ourselves (Godwin et al. 1967) and by Rowley & Southworth (1967). A closeexamination of Ubisch bodies at this stage also shows similar structures associatedwith the electron-dense material and although lines of similar size and density may befound immediately below the tapetal cell membrane it is not presently clear how theyarise (Figs. 25, 26).
The deposition of sporopollenin continues unevenly and there is a suggestion thatthe overall cross-section of the simple Ubisch body superficially resembles the overallcross-section of the tricolpate pollen grain (Figs. 21, 27) in the localization of distinctareas of heavy and light thickening. The multiple Ubisch bodies continue to develop,and even the small peripheral vesicles gradually become coated with sporopollenin(Fig. 23). As development of the Ubisch bodies proceeds, the central area, whichcorresponded to the original pro-Ubisch body, becomes smaller and smaller, and atmaturity has nearly disappeared (Figs. 23, 25). At no stage in development have wefound Ubisch bodies free in the anther cavity; they invariably remain associated withall the surface faces of the tapetal cell. It is interesting to note that during the earlystages of Ubisch body development, from about the time of the emergence of greybodies from the tapetum, it has been possible to see on a single micrograph variousdegrees of sporopollenin deposition around the Ubisch bodies. In the later stages, allUbisch bodies appear to be fully formed, and although the pro-Ubisch bodies are stillpresent in the tapetal cytoplasm, they appear no longer to emerge from the cell(Fig. 21).
At this time a number of smaller granules and, more particularly, vesicles, occurimmediately to the outside of the tapetum. It is not clear whether these vesicles areidentical or not to those found associated with the dictyosomes at an earlier stage indevelopment (Figs. 21, 23, 24).
The tapetum gradually becomes more and more disorganized. The mitochondriaand plastids appear to be the first organelles to disappear, and this is associated withan apparent increase in vesiculation in the cytoplasm (Figs. 21, 22). The loss ofplastids and mitochondria is closely followed by a gradual diminution in the numberof dictyosomes and ribosomes in the cell, although even at this late stage the latter maystill be found closely associated with the pro-Ubisch bodies, which continue to aboundin the tapetal cytoplasm (Fig. 25). The cell wall has now virtually disappeared, yet thecell membrane, together with the nucleus and endoplasmic reticulum, remains intactfor some considerable time.
In the final stages of anther development, at about the time of anther dehiscence,the tapetal cytoplasm is completely disorganized. The limiting cell membrane is rup-tured, and very few recognizable organelles may be found in the residue. The Ubischbodies and the remaining pro-Ubisch bodies are liberated into the anther cavity(Fig. 27), and some may accumulate on and around the mature pollen grains. There isa close similarity in electron density and structure between the pro-Ubisch bodiesreleased from the tapetum at this final stage and the lipo-proteinaceous tryphine whichbecomes deposited on the pollen-grain wall during these closing phases of development.
Cell Sci. 3

168 P. Echlin and H. Godwin
DISCUSSION
The pro-Ubisch bodies are initially surrounded by a zone of radiating ribosomes butlater seem to lie at the confluence of several strands of endoplasmic reticulum: theirprecise mode of origin is still obscure. It might perhaps be conjectured that the pro-Ubisch bodies have something in common with the much smaller spherosomes thatappear to be involved in the synthesis of lipids (Walek-Czernecka, 1962; Frey-Wyssl-ing, Grieshaber & Miihlethaler, 1963; Matile, Balz, Semadini & Jost, 1966) and whichhave been shown to represent a differentiated fragment of the endoplasmic reticulum.
Once the pro-Ubisch bodies have been extruded through the plasmalemma theyrapidly become coated with sporopollenin. What little is known of the biochemistry ofthis substance suggests that it may be formed either from the polymerization ofhydroxycarboxylic acids which have several esterifiable groups or from the polymeriza-tion of terpenoids. Shaw & Yeadon (1966), using mild degradative techniques, foundevidence for an undefined xylan, lignin, cellulose and a large lipid fraction that onoxidative degradation gave compounds containing 16 carbon atoms or less. Thesuggestion is that the sporopollenin precursors are likely to be osmophilic, lipoidalsubstances of low molecular weight that would be expected to have the mediumelectron-density characteristic of the pro-Ubisch bodies.
Ubisch bodies outside the tapetal cells appear to be all of much the same size, yetpro-Ubisch bodies in the tapetal cytoplasm are frequently much larger. This might beexplained by assuming that it is only during a limited period in development of theanther that pro-Ubisch bodies are extruded, and that those subsequently formedincrease in size, sometimes coalesce and are only released from the cell at the time oftapetal disintegration.
From the beginning of meiosis in the microsporocyte until well after its completionthe microspores are separated from each other and from other tissues of the anther bya series of special callose walls (Waterkeyn, 1962). Our experience of poor preservationof the sporogenous cells at this stage indicates that callose is relatively impermeableto fixatives, just as the recent investigations of Heslop-Harrison (1966a, b) have shownit to be impermeable to metabolites derived from the tapetum. It is possible that inH. foetidus the source of the hydrolytic enzymes that cause the breakdown and eventualdissolution of both the tapetal cell wall and the callose special wall is the dictyosomeswhich become particularly abundant in the tapetum at this time, which correspondsapproximately also with primexine deposition on the microspores. The loss of thecallose wall must of course have the double consequence of greatly facilitating trans-port of materials through the anther loculus to the developing microspores and ofproducing carbohydrates of low molecular weight, which are available for resynthesisby the microspores along with the products of hydrolysis of the cellulosic walls of thetapetum. It remains a striking fact that the two sites of condensation of sporopollenin,the microspore exines and the Ubisch bodies, both lie outside the limiting cell mem-branes of microspores and tapetum. Sporopollenin is never seen to be depositedintracellularly and its accumulation appears to proceed simultaneously at both sites,as one would expect were they both drawing upon the same supplies of substrate

Pollen of Helleborus. I 169
available in the thecal fluid to surfaces or structures with suitable synthetic com-petence. There seems good reason to think that both Ubisch bodies and microsporeexines have structures of this kind in the shape of electron-transparent layers of unit-membrane dimensions.
Such layers have already been described by several workers as associated with exineformation in the pollen grain (Godwin et al. 1967; Skvarla & Larsen, 1966; Rowley &Dunbar, 1967; Rowley & Southworth, 1967). In the ontogeny of Ipomoea purpureapollen (Godwin et al. 1967) these layers, recognizable in section as 'white lines',appeared not to be concerned with the deposition of sporopollenin in the bacula,tectum or footlayer of the ektexine, but to play an essential role in formation of thethick, heavily stranded endexine which is also made of sporopollenin. Other observershaving also found the 'white lines' characteristically associated with the endexine(sometimes especially in the annulus of the germ pore), it has been proposed that itmay be possible to distinguish a ' primary' exine formation in which the ' white lines'are not recognizable, and a ' secondary' exine ontogeny that is dependent upon them.As soon as the endexine formation begins in Helleborus foetidus abundant white linesare found associated with it and they can be seen also in the Ubisch bodies which bythis time have heavy coats of sporopollenin. It is still not clear where the white linesoriginate, although Rowley & Dunbar (1967) have discussed the matter at some length.In the pro-Ubisch bodies of Helleborus it seems that they come initially from the endo-plasmic reticulum, although in later stages they show a very close relationship withthe tapetal cell membrane.
It has long been realized that plant tissues experience during their cycle of inception,growth, maturity and senescence what may be called, following F. F. Blackman, an'ontogenetic metabolic drift'. It was shown by de Vries (1877-1879), in his studies ofthe physiology of clover and sugar beet, that all plant tissues, in whatever region of thewhole plant they may be, follow a similar sequence first of protein condensation, thenof starch (and perhaps oil). After maturity there follows hydrolysis of starch andfinally of protein as autolysis preponderates in the ageing tissue. What we clearlyexperience in the maturing anther is a tapetal tissue that passes rapidly through itsontogenetic metabolic phases, after the stage of nuclear multiplication quickly enteringupon a state of general senescence and autolysis in which not only are reserve andskeletal substances hydrolysed but enzymic breakdown of the cytoplasm itself ensues.Whilst this is in train the microspores are at the other end of the ontogenetic metabolicdrift, in a stage of the most active anabolism. With the liberation of the products ofautolysis into the vicinity of the microspores resynthesis is inevitable, and exine forma-tion is one expression of it. It is possible to consider the Ubisch bodies in these termssimply as the consequence of the liberation from the tapetum of fragments of cellcytoplasm sufficiently organized to have escaped autolysis and still capable of limitedsynthesis given substrates adequate in kind and concentration. This conception, it willbe seen, is simply of a mechanistic kind: it makes no attempt to attribute a 'function'to the Ubisch bodies (such as a transport mechanism for sporopollenin), nor does itspeculate upon them in phylogenetic terms. At least for the present this seems to goas far as the evidence warrants.

170 P. Echlin and H. Godwin
There seems no doubt that future studies of the fine structure of the tapetum couldwell be directed to consideration in general terms of senescing tissues. Wrischer (1965)has already made a detailed study of changes in cell ultrastructure during necrobiosisand several of the features he reports—deformation of mitochondria, vacuolation ofplastid stromata, etc.—we also encounter in the tapetum of Helleborus foetidus wheremitochondria and plastids are the first organelles to disappear. Before this stage, how-ever, there has taken place the breakdown of the tapetal cell walls and the callosespecial wall in the thecal sac. At this stage many profiles of dictyosomes are present,together with abundant small vesicles derived from them. These tend to accumulatenear the surface of the tapetal cell. They are considerably smaller than the cyto-somes or microbodies described by Mollenhauer, Morre & Kelley (1966) and areclearly not the same as the spherosomes described by Frey-Wyssling et al. (1963):they are clearly distinct from the pro-Ubisch bodies that originate in endoplasmicreticulum.
In so far as the present study may be taken to have established (at the ultrastructurallevel) a complete developmental history of Ubisch bodies in Helleborus foetidus, fromthe grey vesicles within the tapetal cytoplasm to maturity on the surfaces of the thecalsac, one must naturally inquire whether a similar mechanism might not be found(given sufficiently close study) to be present in other plants. We venture to reconsiderhitherto published accounts with this in mind, remembering of course the likelihoodof differences between species and the difficulty of reproducing detail in the half-tonereproductions of electron micrographs.
It now seems permissible to reinterpret some of the figures published by Heslop-Harrison for Cannabis sativa (figs. 1, 2a, b in Heslop-Harrison, 1962; and pi. 7,figs. 1-4 in Heslop-Harrison, 1963). What he there described as a 'cup' of electron-dense material (sporopollenin) presented at the plasmalemma, base outwards, seemsto correspond closely with the early stages of sporopollenin deposition which takesplace upon the outer surfaces of the grey vesicles of Helleborus foetidus as soon as theyhave broached the plasmalemma of the tapetal cells. If this is so the ontogeny ofUbisch bodies may well be similar in both genera, although we have seen no evidencein Helleborus that the precursor bodies are mitochondria, as was suggested to be thecase in Cannabis, The Helleborus material also yields no trace of the discrete electron-dense materials in the tapetum reported in Silene as particles originating in organellesof the tapetal cells and discharged at the cell surfaces. The plaques figured at the sur-faces of the tapetal cells in Silene pendula (pi. 6, figs. 1, 2 and pi. 11, fig. 1 in Heslop-Harrison, 1963) appear to correspond very closely to the compound or aggregateUbisch bodies which we find very late in the ontogeny of Helleborus foetidus and whichhave been previously recorded from other genera, such as Oxalis where their dimen-sions may sometimes exceed the diameter of tapetal cells. Rowley (1963) has givenwhat might at first sight appear to be a very full account of the origin of Ubisch bodiesin Poa annua, but, useful though the descriptions and electron micrographs of laterstages may be, the earlier stages are less than fully represented. The difficulty inrelating Rowley's results to those of other workers lies chiefly in the fact that he givesno account of a stage in which the microsporocyte, and subsequently the four micro-

Pollen of Helleborus. I 171
spores, are invested in the special callose wall, a stage that investigations in manyother species makes highly probable to have existed also in Poa annua.
It is during this stage in Helleborus foetidus, when a considerable thickness of callosestill intervenes between the microspores and tapetum, that the primexine of themicrospores is first being formed and the first condensation of sporopollenin occurson the surfaces of the extruded grey bodies. It is difficult to understand how therecomes about the spatial apposition of pro-Ubisch bodies and the primexine of themicrospores that is figured for Poa annua by Rowley (1963, figs. 1a, c, 2-7). Theelectron micrograph (fig. 1 c) stated to be at stage 5 of his developmental series showsthe spaces between the microspores to be full of vesicles and other organelles, togetherwith considerable numbers of dispersed Ubisch bodies, whilst the tapetal cell seennext to the endothecium has lost its cellulose walls. It is true that Rowley reports atransient stage in which 'the tapetal cells enlarge radially into the locula to envelopearly microspores', subsequently retracting to a peripheral position. It seems alter-natively possible that either Rowley is dealing with a late stage in breakdown of thetapetal cells in which their contents have been released to fill the thecal sac, or, perhapsless likely, the preparative processes have caused premature rupture of the tapetalcells. In either case it is clear that the contact between Ubisch bodies and microsporesurfaces seems late or fortuitous. The earliest stage shown (Rowley, 1963, fig. id)might indeed be held to represent a very early Ubisch body (stage 2), but even heretapetal cell walls are absent and tapetal cytoplasm apparently reaches the surface ofthe young microspore at a developmental stage (the earliest primexine) when bothHeslop-Harrison (1962, 1963) and ourselves find the microspores still surrounded bythick callose. Likewise figs. 6 and 7 of Rowley (1963), representing Ubisch bodies instage 5 or thereabouts, show naked tapetal cytoplasm next to Ubisch bodies anddeveloping exine. Poa annua may of course differ ontogenetically from the plants wehave examined, but one cannot help feeling that there is need for re-examination of theearlier stages of Ubisch body formation in this species. It may be emphasized that thethin-walled, medium-density vesicles drawn by Rowley (figs. 2-6) as the centresround which Ubisch bodies grow have much in common with the grey vesicles that wehave described as Ubisch body precursors in Helleborus. Indeed the appearance of suchbodies in stage 4 (Fig. 4) recalls the grey vesicles with radiating ribosomes that occurfreely in the Helleborus tapetum (Figs. 8, 13, 14).
With the doubts expressed above we naturally find it difficult to agree with Rowleywhen he suggests that the ' lines of transport to the male gametophyte effect the patternof exine sculpturing', and we prefer to adhere to the view that this pattern is geneticallydetermined by the haploid microspores where in at least several species it expressesitself in a clear primexine whilst still within the callose special wall.
Skvarla & Larsen (1966) conclude that Ubisch bodies 'develop concomitantly withthe exine' and represent sporopollenin deposition upon membranes aggregated atthe tapetal surface. They also consider that 'a strong possibility exists that thesemembranes are derived from ruptured microspores' and point to the presence oforganelles in the fluids of the anther loculus. In Helleborus the phase of liberation oforganelles from the senescing tapetum comes far too late in Ubisch-body ontogeny for

172 P. Echlin and H. Godwin
it to be the normal source of originating membranes, and it may be held that theconsiderable uniformity of size and shape indicates a less fortuitous origin than thedebris of ruptured cells. Skvarla & Larsen continue, that' Ubisch bodies resemble theektexine in that they are channelled, but they are without definitive organization intotectum, columellae and footlayer, and lack the endexine'. They conclude, 'thus it ispossible to explain Ubisch bodies and aborted pollen grains within the model we havesuggested'. It is, of course, agreed that it would be wise to seek a general ontogenticscheme that brings Ubisch body and exine development into a basically similar pattern,and we believe this increases the possibility that the Helleborus system may prove tobe a general one. Finally it may be said that the same authors refer to Heslop-Harri-son's (1963) view that the substrate for sporopollenin is elaborated in the mitochondriaof tapetal cells, and comment that sporopollenin 'platelets' originating thus wouldhave to be depolymerized and then repolymerized along the microspore surfaceswhere condensation could occur without active participation of the microspore proto-plasts. However, Heslop-Harrison (1963) did suggest that the sporopollenin 'platelets'might represent by-products, resulting from the intracellular polymerization of theprecursors of sporopollenin, and recognized the difficulty in supposing the plateletsto represent a temporary storage pool. Skvarla & Larsen, like ourselves, find no evi-dence of platelets in the mitochondria of the tapetum and adopt the more general viewthat sporopollenin substrates are elaborated both from the tapetum and the locularfluid, an opinion close to our own.
The authors wish to express their most grateful thanks to Mr B. Chapman whose technicalassistance throughout has been invaluable, and to Mr Paul Curtis for assistance with thephotography.
REFERENCES
CARNIEL, K. (1963). Das Antherentapetum. Ost bot. Z. n o , 145.CHAMBERS, T. C. & Godwin, H. (1961). The fine structure of the pollen wall of Tilia platy-
phyllos. New Phytol. 60, 393-399.CLAUSEN, P. (1927). Ober das Verhalten des Antheren-Tapetums bei einigen Monokotylen und
Ranales. Bot. Arch. 18, 1-27. ••COOPER, D. C. (1933). Nuclear divisions in the tapetal cells of certain angiosperms. Am. J. Bot.
20, 358-364-DAHL, A. O. & ROWLEY, J. R. (1965). Pollen of Degeneria vitiensis.J. Arnold Arbor. 46, 308-323.DAVIS, G. L. (1966). Systematic Embryology of the Angiosperms, p. 14. New York: John Wiley.ECHLIN, P., CHAPMAN, B., GODWIN, H. & ANGOLD, R. (1966a). The fine structure and de-
velopment of the pollen of Helleborus foetidus L. Vlth Int. Congr. Electron Microsc, Kyoto,1966, vol. 11, pp. 315-316.
ECHLIN, P., GODWIN, H., CHAPMAN, B. & ANGOLD, R. (19666). The structure of polyvesicularbodies associated with cell walls in the developing anthers of Ipomoea purpurea (L.) Roth.Vlth Int. Congr. Electron Microsc, Kyoto, 1966, vol. 11, pp. 317-318.
FREY-WYSSLING, A., GRIESHABER, E. & MOHLETHALER, K. (1963). Origin of Spherosomes inplant cells, jf. Ultrastruct. Res. 8, 506-516.
GODWIN, H., ECHLIN, P. & CHAPMAN, B. (1967). The development of the pollen grain wall inIpomoea purpurea (L.) Roth. Review of Palaeobotany and Palynology 3, 181-195.
HESLOP-HARRISON, J. (1962). Origin of exine. Nature, Lond. 195, 1069-1071.HESLOP-HARRISON, J. (1963). Ultrastructural aspects of differentiation in sporogenous tissue.
Symp. Soc. exp. Biol. 17, 315-340.

Pollen of Helleborus. I 173
HESLOP-HARRISON, J. (1966a). Cytoplasmic continuities during spore formation in floweringplants. Endeavour 25, 65-72.
HESLOP-HARRISON, J. (19666). Cytoplasmic connections between angiosperm meiocytes. Ann.Bot. 30, 221-230.
HESLOP-HARRISON, J. & HESLOP-HARRISON, Y. (1958). Long-day and auxin-induced malesterility in Silene pendula. Port. Ada biol. 5, 79-94.
KOSMATH, L. (1927). Studien iiber das Antherentapetum. Ost. bot. Z. 76, 235-241.MAHESHWARI, P. (1950). An Introduction to the Embryology of Angiosperms. Chapter 2. The
microsporangium. New York: McGraw Hill.MATILE, M., BALZ, J. P., SEMADINI, E. & JOST, M. (1966). Isolation of spherosomes with
lysosome characteristics from seedlings. Z. Naturf. 20b, 693-698.MOLLENHAUER, H. H., MORRE, D. J. & KELLEY, A. G. (1966). The widespread occurrence of
plant cytosomes resembling animal microbodies. Protoplasma 62, 44-52.PY, G. (1932). Recherches cytologiques sur l'assise nourriciere des microspores et les micro-
spores des plantes vasculaires. Rev. gin. Bot. 44, 316-512.ROWLEY, J. R. (1962). Nonhomogenous sporopollenin in microspores of Poa annua L. Grana
palynol. 3, 3-19.ROWLEY, J. R. (1963). Ubisch body development in Poa annua. Grana palynol. 4, 25-36.ROWLEY, J. R. (1964). Formation of the pore in pollen of Poa annua. In Pollen Physiology and
Fertilization (ed. H. F. Linskens). Amsterdam: North Holland Publishing Company.ROWLEY, J. R. & DUNBAR, A. (1967). Sources of membranes for exine formation. Svensk. bot.
Tidskr. 61, 49-64.ROWLEY, J. R., MOHLETHALER, K. & FREY-WYSSLING, A. (1959). A route for the transfer of
materials through the pollen grain wall. jf. biophys. biochem. Cytol. 6, 537—538.ROWLEY, J. R. & SOUTH WORTH, D. (1967). Deposition of sporopollenin on lamellae of unit
membrane dimensions. Nature, Lond. 213, 703.SCHNARF, K. (1923). Kleine Beitrage zur Entwicklungsgeschichte der Angiospermen. IV. Uber
das Verhalten des Antheren-tapetums einiger Pflanzen. Ost. bot. Z. 72, 242-245.SHAW, G. & YEADON, A. (1966). Chemical studies of the constitution of some pollen and spore
membranes. J. chem. Soc. (C) pp. 16-22.SKVARLA, J. J. & LARSEN, D. A. (1966). Fine structural studies of Zea mays pollen. I. Cell
membranes and exine ontogeny. Am. jf. Bot. 53, 1112-1125.TISCHLER, G. (1908). Zellstudien an sterilen Bastardpflanzen. Arch. Zellforsch. 1, 33-51.UBISCH, G. V. (1927). Zur Entwicklungsgeschichte der Antheren. Planta 3, 490-495.VRIES, H. DE (1877-79). Reprinted in Hugo de Vries Opera e periodicis collata (1918), vol. Ill,
pp. 31-523. Utrecht: Oosthoek.WALEK-CZERNECKA, A. (1962). Mise en evidence de la phosphatase acide (monophosphoesterase
II) dans les spherosomes des cellules dpidermiques des 6scailles bubaires d'Allium cepa. AdaSoc. Bot. Pol. 31, 539-546-
WATERKEYN, L. (1962). Les parois microsporocytaires de nature callosique. Cellule 62, 225-255.WITKUS, E. R. (1945). Endomitotic tapetal cell divisions in Spinacia. Am. J. Bot. 32, 326-330.WRISCHER, M. (1965). Elektronenmikroskopische Untersuchungen der Zellnekrobiose. Proto-
plasma 60, 355-400.YAMAZAKI, T. & TAKEOKA, M. (1962). Electron microscopic investigations of the fine details of
the pollen grain surface in Japanese gymnosperms. Grana Palynol. 3 (2), 3-12.
{Received 21 July 1967—Revised 18 October 1967)

174 P- Echlin and H. Godwin
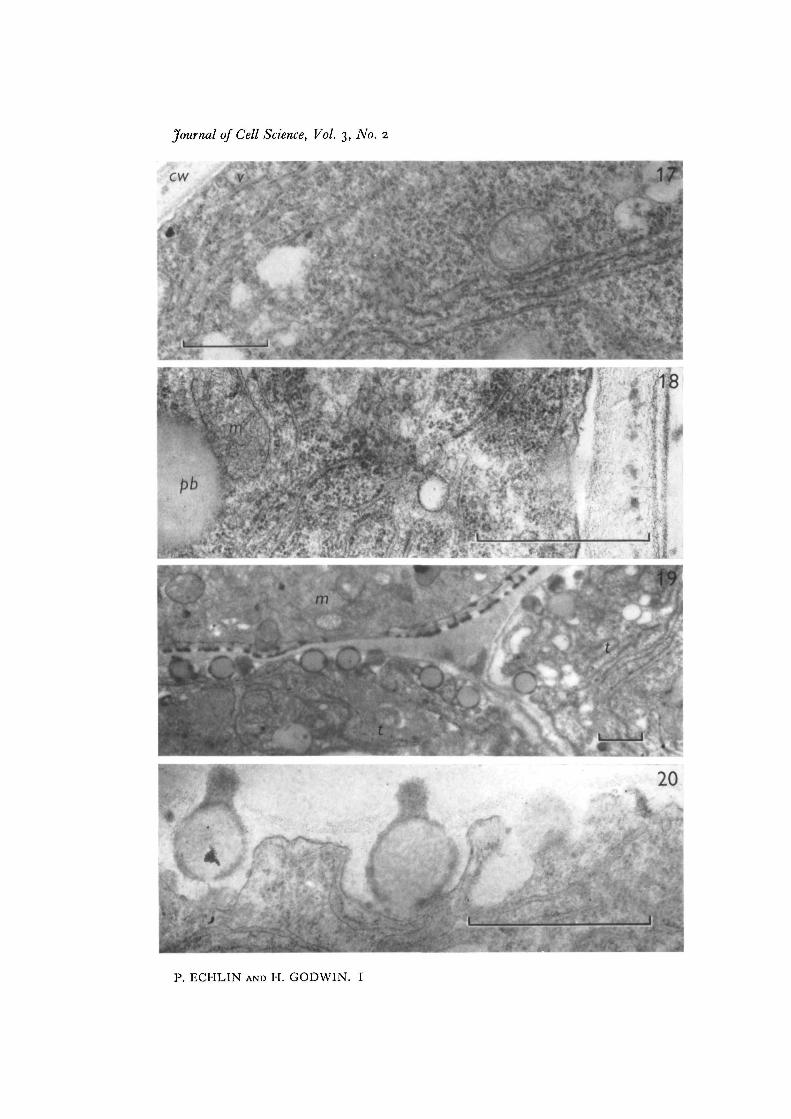
The scale on the micro, 'ise indicated.Fig. i. Sporogenous cell (s) surrounded by a number of tapetal cells (t). x 5000.Fig. 2. Polyvesicular body (pvb) at the junction of two tapetal cells of the same stageas Fig. 1. The cell wall (cw) is relatively thin, and is associated with microtubules (mi)in the peripheral cytoplasm. Plastids (p) and pro-Ubisch bodies (pb) are seen in thetapetal cytoplasm, x 46500.Fig. 3. Details of the tapetal cytoplasm taken at the same stage as Figs. 1 and 2. Theelongate mitochondria (m) are closely associated with the plastids (p). Microtubulesmay be seen associated with both sides of the cell wall (cw). There are few ribosomes inthe cytoplasm, x 48000.
Journal of Cell Science, Vol. 3, No. 2
P. ECHLIN AND H. GODWIN. I {Facing p. 174)

Fig. 4. Tapetal cells (t) at the time of early tetrad formation. The microspores (m) areinvested with a heavy layer of callose (c) and plasmodesmata may be seen connectingadjoining tapetal cells. Pro-Ubisch bodies (pb), mitochondria and dictyosomes (d)are present in the tapetal cytoplasm and the cell wall between the tapetum and anthercavity shows a considerable increase in thickness, x 12000.Fig. 5. Edge of tapetal cell at the junction with the anther cavity, showing vesicleslying below the convoluted tapetal cell membrane. Developmental stage as in Fig. 4.x 48000.Fig. 6. Dictyosome and associated vesicles at periphery of tapetal cytoplasm. Develop-mental stage as in Fig. 4. x 48000.Fig. 7. Dictyosomes and vesicles in close proximity to developing pro-Ubisch bodies(pb) in tapetal cells. Note the increased number of ribosomes in the cytoplasm, someof which appear to be associated with the endoplasmic reticulum. Developmentalstage as in Fig. 4. x 48000.
Journal of Cell Science, Vol. 3, No. 2
P. ECHLIN AND H. GODWIN. I

Fig. 8. Pro-Ubisch body surrounded by a zone of ribosomes. The microspores are atthe late tetrad stage, x 52000.Figs. 9-12. Four consecutive sections through the periphery of the tapetal cytoplasmshowing the extrusion of the pro-Ubisch body from the cell, x 52000.
Journal of Cell Science, Vol. 3, No. 2
12
P. ECHLIN AND H. GODWIN. I

Fig. 13. Periphery of the tapetum at the time of primexine formation in the microspore.The cell wall is less electron-dense and begins to show a more open fibrous appearancecontaining irregularly shaped electron-dense granules. The extruded pro-Ubischbodies (pb) have a thin coating of electron-dense exinous material. The tapetal cyto-plasm is filled with recognizable organelles, and the endoplasmic reticulum can clearlybe seen to be associated with the developing pro-Ubisch bodies. Note regions of unit-membrane dimensions surrounding the extruded pro-Ubisch body (asterisk), x 45 000.Fig. 14. Details of the close association between endoplasmic reticulum and pro-Ubisch bodies, x 45 000.Fig. 15. Pro-Ubisch bodies with thin coating of sporopollenin on outer surface.Developmental stage as in Fig. 13. x 45000.Fig. 16. Dictyosome and associated vesicles in the peripheral tapetal cytoplasm.Developmental stage as in Fig. 13. x 32000.
Journal of Cell Science, Vol. 3, No. 2
P. ECHLIN AND H. GODWIN. I

Fig. 17. Tapetal cell showing ribosomes free in the cytoplasm and attached to theendoplasmic reticulum. The cell wall (cw) has lost much of its electron density.Electron-transparent vesicles (v) are located beneath the tapetal cell membrane,x 22500.
Fig. 18. The tapetal cell wall shows further signs of dissolution, and there is an increasein the deposit of electron-dense material along the middle of the cell wall. The pro-Ubisch bodies (pb) are in close association with the ribosomes and mitochondria (m).x 45 000.
Fig. 19. General appearance of the developing microspore (»//) and tapetal cells (t).The pollen grain wall now contains recognizable exinous components which are of thesame density as the material surrounding the pro-Ubisch bodies. The tapetal cell wallshave now virtually disappeared, x 11 000.Fig. 20. Uneven deposition of sporopollenin on pro-Ubisch bodies. The tapetal cellwall is now reduced to a narrow finely fibrous layer, x 48000.
Journal of Cell Science, Vol. 3, No. 2
CW
P. ECHLIN AND H. GODWIN. I

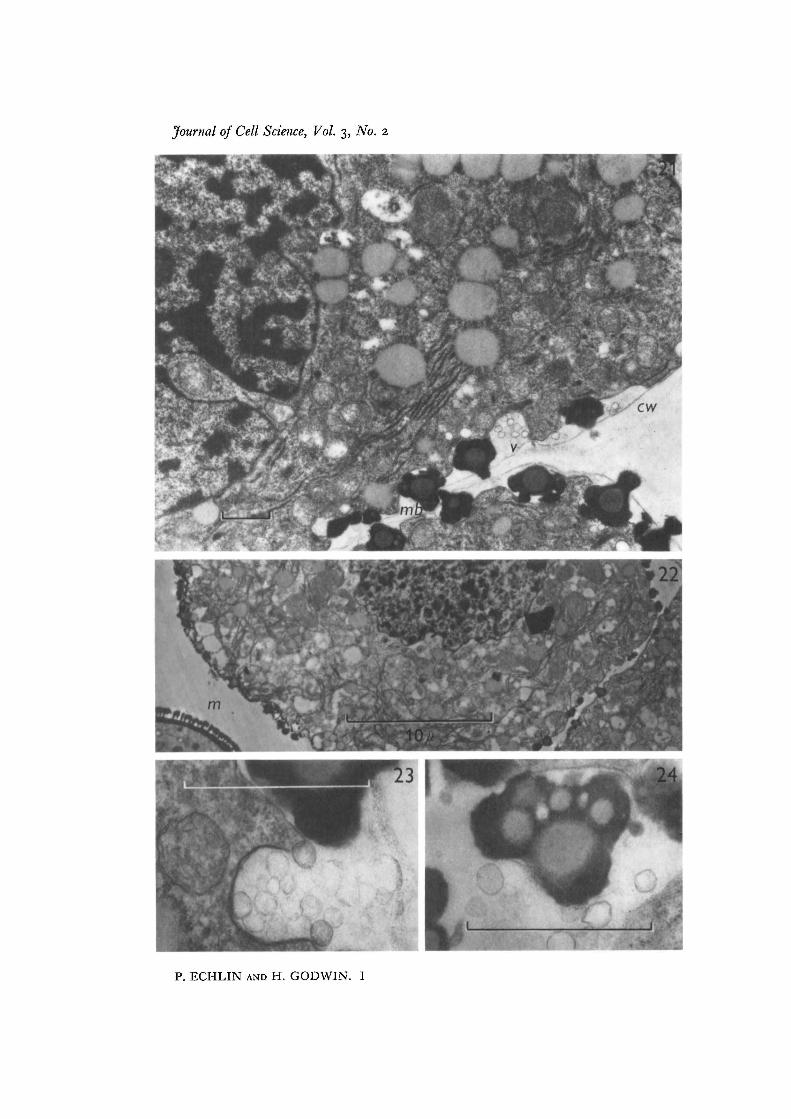
Fig. 21. Tapetum with large lobate nucleus, and a number of pro-Ubisch bodies ofdifferent sizes. The cell wall (czu) is reduced to a thin layer and multiple Ubisch bodies(mb) are forming outside the tapetum, along both the anther cavity wall and along thetapetal side walls. Electron-transparent vesicles (v) collect along the outside of thetapetal cell, while there is an apparent increase in the number of electron-transparentregions in the main body of the tapetum. x 12000.
Fig. 22. Tapetum in the phase of senescence containing many regions of low electron-density with many pro-Ubisch bodies remaining within the cytoplasm. The micro-spore (in) wall is now nearly fully formed, x 3750.Fig. 23. Details of the electron-transparent vesicles on the outside of the tapetal cell,at the stage of the final dissolution of the tapetal cell wall and the special callose walls.Developmental stage as in Figs. 21, 23. x 48000.Fig. 24. Details of a multiple Ubisch body. Note the thin 'white lines' or electron-transparent regions of unit-membrane dimensions associated with the sporopollcnindeposition in the outer regions of the Ubisch body, x 48000.
Journal of Cell Science, Vol. 3, No. 2
P. ECHLIN AND H. GODWIN. I

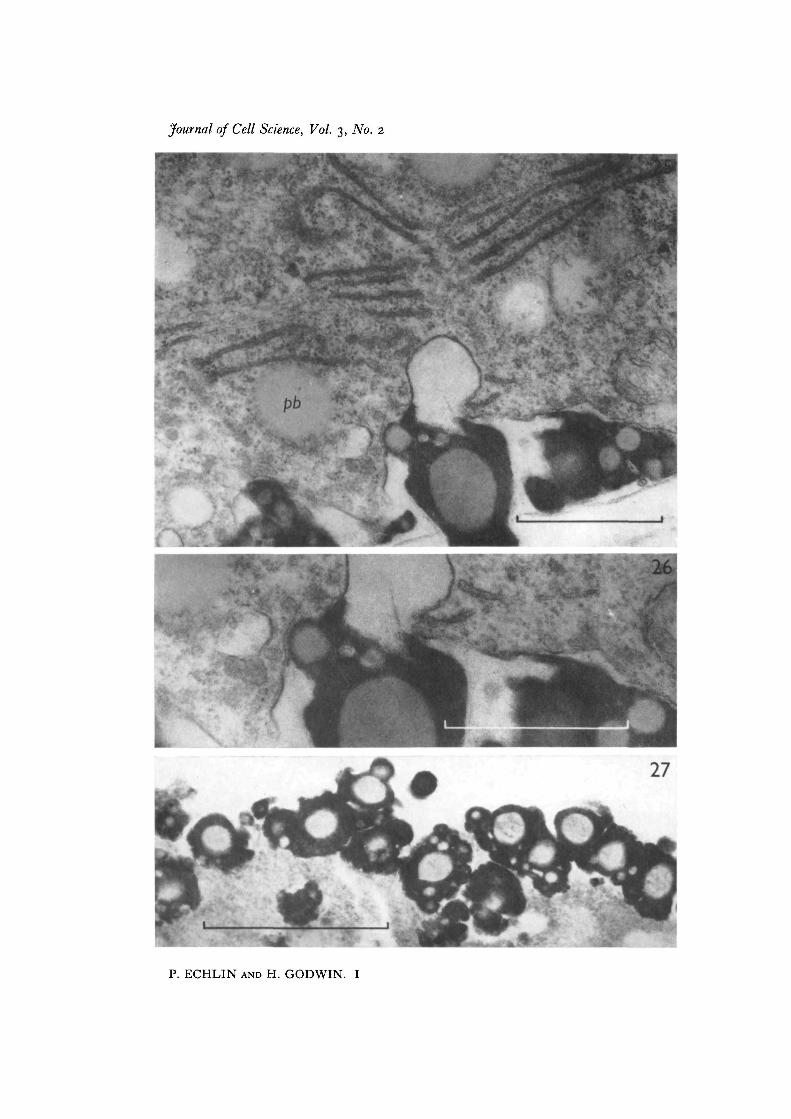
Fig. 25. Nearly mature Ubisch body with 'white lines'. The tapetal cytoplasm,although in a phase of decline, still shows profiles of endoplasmic reticulum andelectron-dense ribosomes. Pro-Ubisch bodies (pb) are still present in the tapetalcytoplasm, x 39000.Fig. 26. Detail of Fig. 25, showing the close association of the 'white lines' with thedeposition of sporopollenin. There is some evidence that the ' white lines' may be con-tiguous with the tapetal cell membrane, x 48000.Fig. 27. The final stage in the dissolution of the tapetum. The cytoplasm is now com-pletely disorganized, and aggregated Ubisch bodies line the outer region of the remainsof this tissue, x 48000.
Journal of Cell Science, Vol. 3, No. 2
P. ECHLIN AND H. GODWIN. I






























