Embed Size (px)
Citation preview

© 1970 by Academic Press, I ll f'.
J. ULTRASTRUCTURE RESEARC H 30, 317- 327 ( 1970)
The Ultrastructure of the Nuclear Envelope of Amphibian
Oocytes: A Reinvestigotion
11. The Immature Oocyte and Dynamic Aspects
W ERNER W . FRANKE AND U LRICH SCHEER
317
Divisiol1 of Cell Biology, Institute of Biology If, University of Freiburg i. Br. , Germany
Received May 27, 1969, and in revised form July 7, 1969
Different stages o f oogenesis of the Alpine newt were compared with respect to the nuclear envelope structure, em ploying negative staining and sectional work: la rva l eggs, ea rly and later lampbrush stages, and mature eggs. While in a ll the stages the genera l components of the nuclear pore complexes cou ld be observed, differences were found concerning qua ntitative structural data. In particular th e frequency of pores containing a central granule decreased from 61 % in the la rva l to 36 % in the mature egg. Some new information on the mode of nucleocyto plasmic transit of nucleolus-derived ma terial could be obtained from the lampbrush stage eggs. In this stage, fibrillar strands spinning out from the nucleo lar periphery can be seen in contact with the nuclear pore complex, expecially with the inner annular granules . Furthermore, dense spheres of nucleolar material migrate through the central channel of the pore in a rodlike configuration in the very same mode that has been describ:;:d for sa li vary glands of Chironomus thummi (69). Alternative relationships of nucleocytoplasmic RNP transport to the const ituents of the nuclear pore complex are discussed.
There ex ist drast ic changes in qua lity a nd quantity of RNA synthesis during the oogenesis of the a mphibian oocyte (e.g., 11- 15, 18- 20). Since the nuclear pore complexes are widely thought of as either regulating or being functionally correlated to the efflux of ribonucleoprotein-containing particles from the nucleoplasm into the cytoplasm, it appears reasonable to investigate whether any changes in nuclear envelope structure and composition occur during oogenesis . Merriam (43) has compared the structure of osmicated, a ir-dried nuclear envelopes after isolation from different stages of Rana pipiens oogenesis by using the Callan- Tomlin technique. It is the purpose of the present study to extend this work and to compare the structure of the mature amphibia n oocyte which has been described in detai l in a foregoing article (23) with that of the lampbrush and the la rval stage by combining section and negative sta ining electron microscopy.

318 FRANKE AND SCHEER
MATERIALS AND METHODS
For negative staining preparations, nuclear envelopes were isolated from oocytes or the Alpine newt, Tri/urus aipes/ris Laur. , according to the methods previously described (23). For section work the total isolated nuclei as well as intact oocytes were prepared as described in that article. Electron micrographs were made with Siemens Elmiskop lA and 101.
The investigated stages were (a) larval stage (transparent oocytes with a diameter of 140 ± 25 Jim) ; (b) lampbrush stage (slightly pigmented oocytes with a diameter of 750 ± 30 Jim ; comparable to stage 4- 5 sensu Duryee (22), to stage C sensu Grant (25), to stage Y3 sensu Kemp (33, 34), to stage 3 sensu Scharrner (59), to stage J[ a sensu Warten berg (75»; and (c) the mature stage (oocyte diameter about 1600 Itm). J n some cases late lampbrush stages (oocyte diameters about 900 Jim) were also examined. Since the structure of the nuclear envelope depends strongly on the special isolation conditions used, particular care was taken to evaluate comparatively only material that had been prepared in the same way. For quantitative evaluation of central granule frequency, every distinct centrally located particle larger than 30 A has been considered.
RESULTS AND DlSCUSS[ON
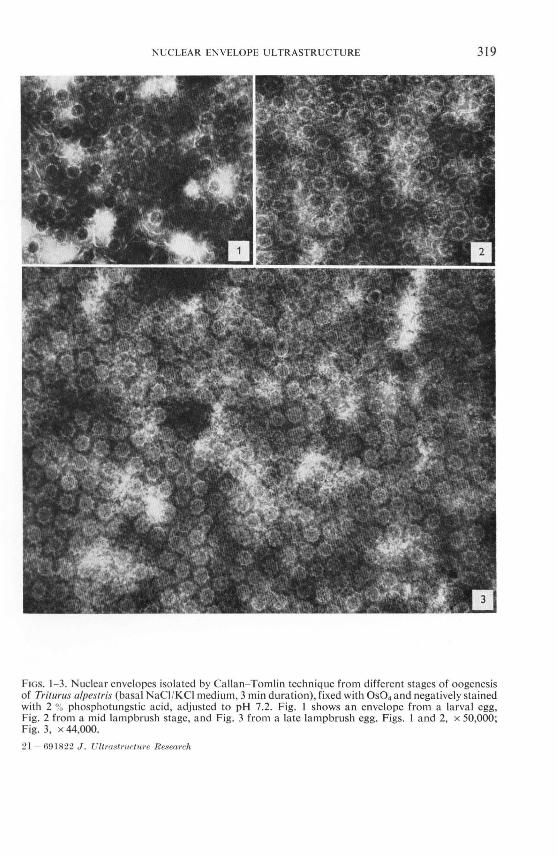
As can be seen from Figs. 1- 3 no remarkable differences in the structural properties of the nuclear pore complex of the isolated envelopes could be observed between the different stages of oogenesis. All the granular and fibrillar structures known from the
nuclear pore complex of the mature newt egg (23) occur also in the earlier stages of oogenesis.
Comparative sectional studies on different aspects of egg development are numerous
(1, 3, 5, 7,8, 10, 16, 29, 30, 34, 36, 45, 51 , 52, 67,71 , 74- 78). With respect to nuclear pore complex structure and function , however, a new interesting finding could be obtained from the lampbrush stage. In this stage, in which exceedingly high RNAsynthesis takes place (e.g., 19), the nucleoli are preferentially located in the periphery
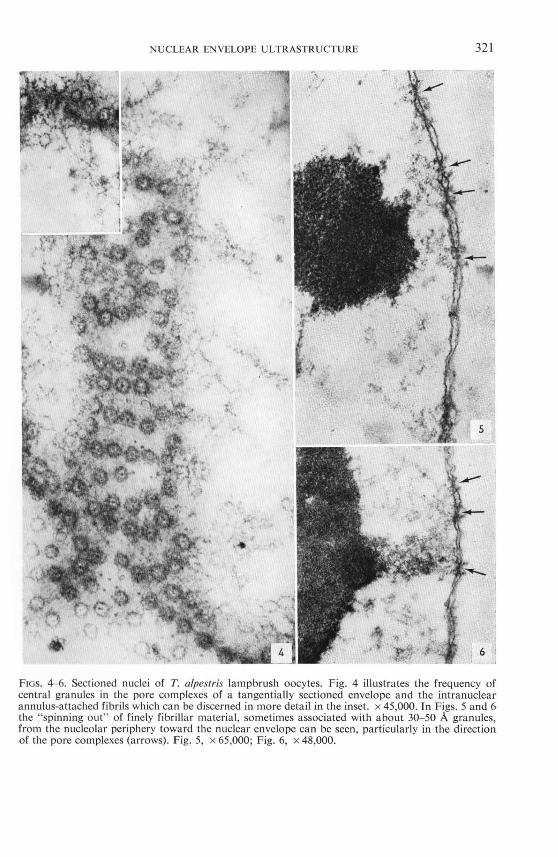
of the nucleus beneath the nuclear envelope (e.g., 39, 78). Strands consisting of finely filamentous (about 30 A) and granular material of various size can be seen extending
from the nucleolar periphery toward the nuclear pore complexes (Figs. 5 and 6). While some of these filaments appear to be associated with granules of diameters of about 30- 60 A, arrangements of the filaments with dense spheres with diameters from 200 to 700 A also can be observed (Figs. 7- 10; cf. "the streams of granules"
of 39). Similar "spinning out" processes of nucleolar fibrils in the direction of the nuclear envelope have been reported from other oocytes (36- 38). Observations suggesting a structural transformation of coarse RNP granules into rods or fibrils when
these approach the nuclear pore complex were recently reported by Monneron and Bernhard (47). Images like that shown, e.g., in Figs. 5 and 7, suggest that at least some of these nucleolar-derived filaments might be continuous with and identical to
NUCLEAR ENVELOPE ULTRASTRUCTURE 319
FIGs. 1- 3. Nuclear envelopes isolated by Callan- Tomlin technique from different stages of oogenesis of Tritlll"lIs alpeslris (basa l NaCI /KCl medium, 3 min duration), fixed with OS04 and negatively stained with 2 % phosphotungstic acid, adjusted to pH 7.2. Fig. J shows an envelope from a larval egg, Fig. 2 from a mid lampbrush stage, and Fig. 3 from a late lamp brush egg. Figs. I and 2, x 50,000; Fig. 3, x 44,000.
21 - 69 J 822 J. UltrastrllctuTe R esea,·"h
320 FRANKE AND SCHEER
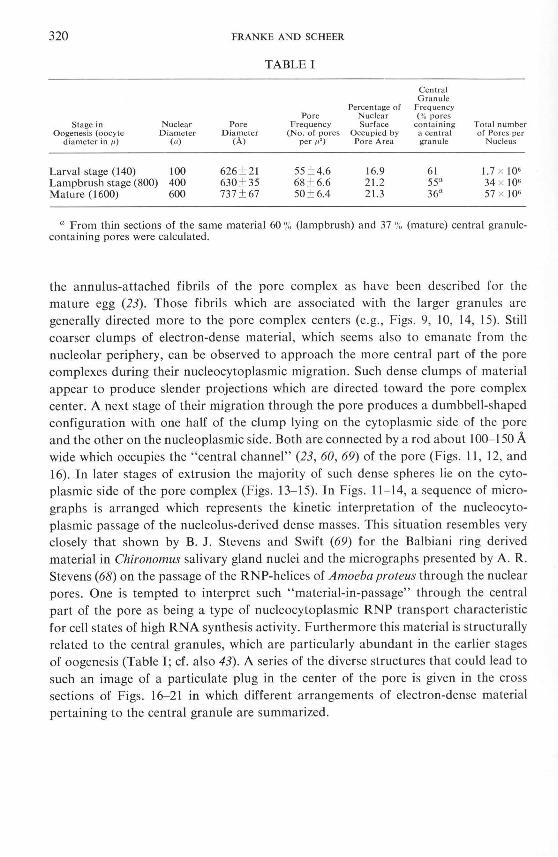
TABLE I
Central Granule
Percentage of Frequency Pore Nuclear ( % pores
Stage in Nuclear Pore Frequency Surface co ntaining Total number Oogenes is (oocyte Diameter Diameter (No. of pores Occupied by a cen tral of Pores per
diameter in /1) (1') (A) per tt2) Pore A rea granu le Nucleus
Larval stage (140) 100 626 ± 21 55 ± 4.6 16.9 61 1.7 x 10· Lampbrush stage (800) 400 630 ± 35 68 ± 6.6 21.2 55a 34 x 10· Mature (1600) 600 737 ± 67 50 ± 6.4 21.3 36a 57 x ]0·
a From thin sections of the same material 60 % (lamp brush) and 37 % (mature) central granulecontaining pores were calculated.
the annulus-attached fibrils of the pore complex as have been descri bed for the mature egg (23). Those fibrils which are associated with the larger granules are generally directed more to the pore complex centers (e.g., Figs. 9, 10, 14, 15). Still coarser clumps of electron-dense material , which seems also to emanate from the nucleolar periphery, can be observed to approach the more central part of the pore complexes during their nUcleocytoplasmic migration. Such dense clumps of material appear to produce slender projections which are directed toward the pore complex center. A next stage of their migration through the pore produces a dumbbell-shaped configuration with one half of the clump lying on the cytoplasmic side of the pore and the other on the nucleoplasmic side. Both are connected by a rod about 100- 150 A wide which occupies the "central channel" (23, 60, 69) of the pore (Figs. 11, 12, and 16). In later stages of extrusion the majority of such dense spheres lie on the cytoplasmic side of the pore complex (Figs. 13- 15). In Figs. 11- 14, a sequence of micrographs is arranged which represents the kinetic interpretation of the nucleocytoplasmic passage of the nucleolus-derived dense masses. This situation resembles very closely that shown by B. J. Stevens and Swift (69) for the Balbiani ring derived material in Chironomus salivary gland nuclei and the micrographs presented by A. R . Stevens (68) on the passage of the RNP-helices of Amoeba proteus through the nuclear pores. One is tempted to interpret such "material-in-passage" through the central part of the pore as being a type of nucleocytoplasmic RNP transport characteristic for cell states of high RNA synthesis activity. Furthermore this material is structurally related to the central granules, which are particularly abundant in the earlier stages of oogenesis (Table I; cf. also 43). A series of the diverse structures that could lead to such an image of a particulate plug in the center of the pore is given in the cross sections of Figs. 16-21 in which different arrangements of electron-dense material pertaining to the central granule are summarized.
NUCLEAR ENVELOPE ULTRASTRUCTURE 321
FIGs. 4-6. Sectioned nuclei of T. alpesfris lampbrush oocytes. Fig. 4 illustrates the frequency of central granules in the pore complexes of a tangentially sectioned envelope and the intranuclear annulus-attached fibrils which can be discerned in more detail in the inset. x 45,000. In Figs. 5 and 6 the " spinning out" of finely fibrillar material, sometimes associated with about 30- 50 A granules, from the nucleolar periphery toward the nuclear envelope can be seen, particularly in the direction of the pore complexes (arrows). Fig. 5, x 65,000; Fig. 6, x 48,000.

322 FRANKE AND SCHEER
In a few cases it could be recognized that slender strands of nucleolar-derived material extending through the pore center were as thin as 30- 50 A (Fig. 17). This corresponds to the class of "small" central granules occasionally found in negative staining preparations of lampbrush as well as of later oocytes (cr. 23). Although there is evidently no doubt about the RNA-content of such nucleocytoplasmic migrating dense material (e.g., 4,7,9,35,44,49,53,63, 68,69), it is not clear what kind of RNA they contain, i.e., either mRNA (dRNA) or rRNA or both . While so me authors have favored an interpretation of messenger ribonucleoprotein (69, 72), the nucleolar origin of these particles seems to indicate a ribosomal nucleoprotein (cf. 2,6,49,74). In this connection, however, it should be kept in mind that extrusion of nucleolar material into the cytoplasm need not necessarily to take place via the nuclear pores since other mechanisms have been reported: e.g., budding off of parts of the perinuclear cisterna into vesicles including the nucleolar material (e.g., 70) or partial dissolution of nuclear envelope (73).
In a previous article it was shown that there exist no interspecific differences with respect to the structural properties of the nuclear pore complex (23). Interesting quantitative intraspecific differences, however, were observed in the course of oocyte development (Table I). Thus, a comparison of the three stages of oogenesis in Triturus
alpesfris reveals that during egg growth the portion of central granule containing pore complexes declines from an average of 61 % in the larval eggs down to 36 %
in the mature oocytes. This finding is in general agreement with the remark by Merriam (43) that the centraL dots were more frequent in immature frog oocytes than in mature ones. The fact that this author found in both stages an absolutely lower percentage of central granule-containing pores (l6 % and 2 %) seems to be explained by the lower resolution and contrast difference of the preparation method he employed.
The relative pore area seems to keep stable during oogenesis. The importance of the question how pore complexes are formed may be emphasized by the calculation that
FIGs. 7- 15. Details of the structural relationship of the nucleolus-derived dense material with the nuclear pore complexes. In Figs. 7 and 8 the fibrillar connections of such nucleolar clumps to the inner annulus of pore complexes (arrows) can be seen. In Fig. 9 a cluster of dense spheres and thin fibrils is in contact with a pore complex (arrow). Note also the outer annular granules. Fig. 10 presents a line of such spheres, which seemingly is in transit through the central part of a pore complex (arrow). In Fig. J I a large clump of nucleolar material lies attached to the pore still on the nucleoplasmic side while in Fig. 12 such material reveals a dumbbell-shaped configuration with one half intranuclear and the other half lying already in the cytoplasm, both halves connected by a slender rod extending through the central channel of the pore. In Figs. 13 and J 4 such clumps of nucleolar material lie in the cytoplasm adjacent to the nuclear envelope, but other smaller particulate material seems to be " en route. " Note the thin 30 A filament (Fig. 13, arrow) connecting two such dense spheres through a nuclear pore complex. Fig. J 5 presents a survey which demonstrates how frequent such nucleocytoplasmic migration of nucleolus-derived dense material (nm) is in the lampbrush stage. N, nucleoplasm; M, mitochondrion. Figs. 7, 8, and 10, x 55,000; Figs. 9 and 12, x 70,000; Fig. 11, X 100,000; Figs. 13 and 14, X 62,000; Fig. 15, X 50,000.
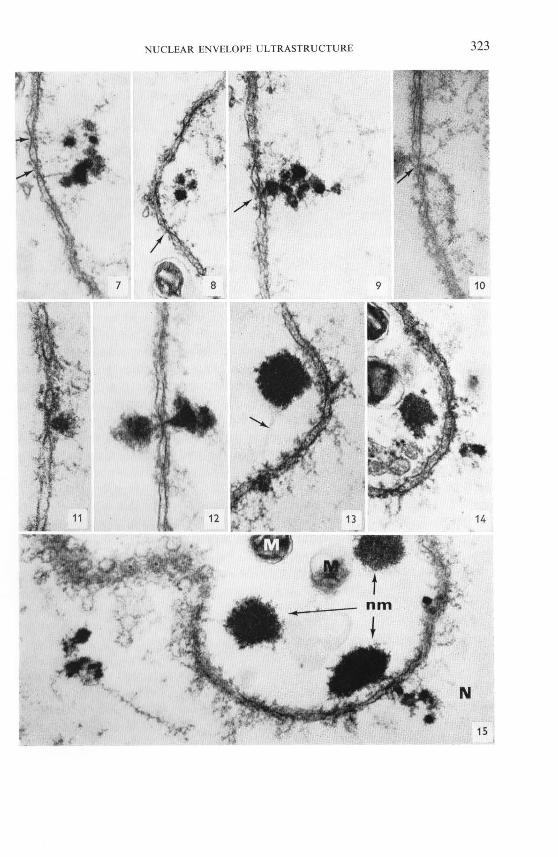
NUCLEAR ENVELOPE ULTRASTRUCTURE 323

324 FRANKE AND SCHEER
during the described interval of oogenesis of the Alpine newt the total number of pores, as a general consequence of nuclear growth , must increase from 1.7 million pores to 57 million pores per nucleus (Table I) .
Apart from the central granule which seemingly represents a variable structure (Figs. 16-21; cf. 23), it is apparent that two basic structural components chiefly make up the particulate pore complex materia l, namely the 30 A-broad fibrils, sometimes associated with about 40- 60 A granules, a nd the 100- 200 A large annul ar granules. Consideri ng the present background of biochemical knowledge in this field , the following a lternative speculations on the possible biological meaning of the pore complex seem to be reasonable and might serve as st imuli for further experi ments:
l. The inner annulus-attached fibrils are identica l with the 30 A ribonucleoprotein (RNP) strands described by Spirin and hi s colleagues (review 64). Thus, nascent riboso mal RNP strands coil up at the pore margin into granules equiva lent to ribosomal subunits. This would be consistent with the general predominance of rRNA synthesis in such oocytes (e.g., 20), with their origin from the nucleolar periphery as can be seen particularly well in the Jampbrush stage, with their high stai nability with lead and uranyl stainings, with the existence of an electron-transparent core in the annular subunits and with ~he coiling tendency of these fibrils (23) . This evidence fits into the general concept of the pore complex as a site where ribosomal and/or polysomal constituents are accumulated and assembled. Then the final maturation of the ribosomal precursor particles to the effective ribosome subunits, e.g., the completingaddition of ribosomal protein , could also occur within the pore complex region.
2. The annulus-attached fibrils represent initiator co mplexes (recent review, 27), i.e., mRNA plus smaller ribosomal subunits (/7, 28, 32, 40) . This could explain the small pa rticles observed as attached to these fibrils within the pore complex and studded a long the fibrils extending into the nucleoplasm. In this concept the pore complex would act as a gateway in which the polyso me is completed ready to translation by adding simply the la rge subunits. Thus, in thi s case the rate of polysome formation within the pore complex could control the time of the messenger's stay at the pore complex. This means that under conditions of slow polysome formation, as is the case in the fully mature oocyte, the pore complex functions by piling up informational RNA in the mode of a structure-bound pool, thus providing a sto rage of "stable messengers" known to be present in egg cells (e.g., 18,26,66). In this concept the annulate lamellae could serve as a greatly enhanced structural basis of pore complex-bound messenger capacity (cf. 60).
3. The annulus-attached fibrils represent "informator complexes" ("informosomes" sensu 58,65; review, 24), i.e., mRNA bound by nonriboso mal "i nformofer" protein (31, 46, 54, 57; cf. , however, 50) . The dimensions of such complexes as have been reported by Samarina et al. (58) and by Monneron and M oule (48), however,

NUCLEAR ENVELOPE ULTRASTRUCTURE 325
FIGS. 16- 21. Sectional study on the diversity of structures which can produce the image of a central granule in tangential sections or in isolated envelopes: A central about 100 A thick rodlike waist of the nucleolar material penetrating the pore complex (Fig. 16), an about 30 A thin filament directed to the pore center (Fig. 17, upper pore), a 100-150 A large plug in the pore center (Fig. 18), two 100-150 A granules lying on either side of the pore equator (Fig. 19, both pores), one 100- .\ 50 A la rge granule lying on the nucleoplasmic (Fig. 20) or on the cytoplasmic side of the pore equator (Fig. 21). All, x .\ 10,000.
exceed those of the 30 A fibril s by far and could otherwise rather correspond to the central dot material.
4. The pore complex itself is some sort of a special polysome in that the 16 annular granules, and possibly also the central granule, represent the ribosomes of one or several sets of nuclear pore complex polysomes at which the start of cytoplasmic protein synthesis takes place (cf. 42, 56).
5. The nuclear pore complex is a site of special ribonucleotidase activity thus cleaving either the polycistronic 40 S- 30 S RNA containing RNP strands (55) to the smaller RNA's (reviewed, e.g., J J) of the ribosomal subunits or cleaving a polycistronic messenger as known from bird erythroblastic cells (6J, 62) into the smaller effective messengers.
6. Despite some unlikelihood it can nevertheless so far not be excluded that the annulus-attached fibrils are highly disperse chromatin strands in a relatively active state (21). DNA-containing masses associated with nuclear pores were recently described for an insect ovarian nurse cell (41).
Further experiments including labeling, cytochemical, and autoradiographic techniques are in progress in our laboratory to determine which of these functional alternatives approach reality.
The authors thank Miss Sigrid Krien and Miss Mari;mne Winter for accurate technical assistance and Drs. H. Falk, H. Kleinig, and F. Wunderlich for helpful discussions. The work was supported in part by the Deutsche Forschungsgemeinschaft.

326 FRANKE AND SCHEER
REFERENCES
J. AOAMS, E. C. and HERTlG, A. T., J. Cell Bioi. 21 , 397 Cl 964). 2. ALLEN, E. R . and CAVE, M. D., Z. Zel/Jorsch. Mikroskop. Ana!. 92, 477 (1968). 3. ANOERSON, E., J. Cell Bioi. 37, 514 (1968). 4. ANOERSON, E. and BEAMS, H. W., J. Biophys. Biochem. Cylol. 2, Suppl. , 439 (1956). 5. BAKER, T. G. and FRANCHI, L. L., J. Cell Sci. 2, 213 (1967). 6. BAL, A. K., JUBINVILLE, F., COUSINEAU, G. H . and JNOUl~, S., J. Ullraslrucl. Res. 25, 15
(1968). 7. BALlNSKY, B. 1. and D Evls, R. J ., Acla Embryc>/. Morphol. Expll. 6, 55 (1963). 8. BEAMS, H. W. and KESSEL, R. G., J. Cell Bioi. 17, 621 (1963). 9. BEERMANN, W., J. Expll. Zool. 157,49 (1964).
10. BORuM, K., Expll. Cell R es. 45, 39 Cl 966). 1 J. BROWN, D. D. , in MoscoNA, A. A. and MONROY, A. (Eds.), Current Topics in Develop-
mental Biology, Vol. 2, p. 47. Academic Press, New York, J967. 12. BROWN, D. D. and LITTNA, E., J. Mol. Bioi. 8, 669 (1964). 13. - ibid. 8, 688 (1964). ] 4. -- ibid. 20, 81 Cl 966). J 5. -- ibid. 20, 95 (1966). 16. CONE, M. V. and SCALZI, H. A., J. Cell Bioi. 35, J63A (1967). 17. MCCON KEY, E. H. and HOPKINs, J. W., J. Mol. BioI. 14, 257 (1965). 18. CRIPPA, M. and GROSS, P. R ., Proc. Nail. Acad. Sci. U.S. 62, 120 (1969). J9. DAVIDSON, E. H ., ALLFREY, V. G. and MIRSKY, A. E., Proc. Nail. A cad. Sci. U.S. 52,
50J (1964). 20. DAVIDSON, E. H ., CRIPPA, M., KRAMER, F. R. and MIRSKY, A. E., Proc. Nail. Acad. Sci.
U.S. 56, 856 (1966). 21. Du PRAW, E. J., Proc. Nail. A cad. Sci. U.S. 53,161 (1965). 22. DURYEE, W. R ., Ann. N. Y. A cad. Sci, 50, 920 (I 950). 23. FRANKE, W. W. and SCHEER, U., J. Ullraslrucl. Res. 30, 288 (1969). 24. GEORGIEV, G. P., in DAVIDSON, M . and COHN, W. E. (Eds.), Progress in Nucleic Acid
Research and Molecular Biology, Vol. 6, p. 259. Academic Press, New York , 1967. 25. GRANT, P ., J. Expll. Zool. 124, 513 (1953). 26. GROSS, P. R ., in MOSCONA, A. A. and MONROY, A. (Eds.), Current Topics in Develop-
mental Biology, Vol. 2, p. I. Academic Press, New York, 1967. 27. GUTHRIE, C. H. and NOMuRA, M., Nalure 219,232 (1968). 28. HENSHAW, E. c., REVEL, M. and HIATT, H. H., J. Mol. Bioi. 14, 241 (1965). 29. HOPE, J ., HUMPHRI ES, A. A. and BouRNE, G. H., J. Ullraslrucl. Res. 10, 547 ( 1964). 30. -- ibid. 10, 557 (1964). 31. INFANTE, A. A. and NEMER, M. , J. Mol. BioI. 32, 543 (1968). 32. JOKLlK, W. K. and BEcKER, Y., J. Mol. Bioi. 13, 511 (1965). 33. KEMP, N. E., J. Morphol. 92, 487 Cl 952). 34. -- J. Biophys. Biochem. Cytol. 2, Suppl., J 87 (1956). 35. KESSEL, R. G., J. Ultrastrucl. Res. 15, J81 (1966). 36. -- ibid. 22, 63 (1968). 37. -- J. Cell Bioi. 36, 658 (1968). 38. KESSEL, R. O. and BEAMS, H. W., J. Cell BioI. 39, 735 (J 968). 39. LANE, N. J ., J. Cell Bio!. 35, 421 (1967).

NUCLEAR ENVELOPE ULTRASTRUCTURE 327
40. LATHAM, H. an d DARNELL, J . E. , J . Mol. BioI. 14, 13 (1965). 41. LUZZATTO, C. A. R. a nd URBA NI, E. , J. Submicroscopic Cytol. 1, 85 (1969). 42. MEPHAM, R. H . and LANE, G. R. , Nature 221 , 288 (1969). 43 . M ERRIAM, R . W., J. Cell BioI. 12, 79 (1962). 44. MILLER, O. L. , Proc. 5th intern. Congr. Electron Microscopy , Philadelphia, i962, Vol. 2,
NN-8. Academic Press, New York, 1962. 45. MILLONIG, G ., Bosco, M . and GIAMBERTONE, L., J. Exptl. Zool. 169, 293 (1968). 46. MOLNAR, J. , SAMARI NA, O. P. a nd GEORGIEV, G. P. , Mol. BioI. (Moscow) 2, 795 ( 1968). 47. MONNERON, A. and BERN HARD, W., J. Ultrastruct. R es. 27, 266 (1969). 48. MONNERON, A. and MOULI~ , Y., Exptl. Cell Res. 51 , 531 (1968). 49. MORITA, M., BEST, J . B. a nd NOEL, J., J. Ultrastruct. Res. 27, 7 ( 1969). 50. MOULI~, Y. and CHAUVEAU, J. , J. Mol. BioI. 33, 465 (1968). 51. N0RREVANG, A., Vidensk. Medd. Dansk Naturh. Foren. 128, 1 (1965). 52. -- intern. Rev. Cytol. 23, 113 (1968). 53 . ORNSTEIN, L., J. Biophys. Biochem. Cytol. 2, Suppl. , 351 ( 1956). 54. PERRY, R. P. a nd KELLEY, D . E. , J. M ol. BioI. 35, 37 (1968). 55. ROGERS, M. E., J. Cell BioI. 36, 421 (1968). 56. SADOWSKI, P. D . and How DEN, J . A., J. Cell BioI. 37, 163 ( 1968). 57. SAMAR INA, O. P. , LUKANIDIN, E. M., MOLNAR, J . a nd GEORGIEV, G. P ., J. Mol. BioI. 33,
25 1 ( 1968). 58. SAMARINA, O. P., MOLNAR, J ., LUKANIDIN, E. M. , BR USKOV, V. 1. , KRICHEVS KAYA , A. A
and G EORGIEV, G. P., J. Mol. BioI. 27, 187 (1967). 59. SCHA FFNER, H. , Histochemie 13, 346 (1968). 60. SCHEER, U. a nd FRANKE, W. W., J. Cell BioI., 42, 5J9 (1969) . 61. SCHERRER, K. and MARCAUD, L. , J. Cellular Physiol. 72, Suppl. l (1968). 62. SCHERRER, K. , MARCAUD, L., ZAJDELA, F., LONDoN, I. M. and GROS, F. , Proc. Natl.
A cad. Sc i. U.S. 56, 1571 (1966). 63. SENTEIN, P. a nd TEMPLE, D ., Compt. Rend. A cad. Sci. 268, 540 (1969). 64. SPIRI N, A. S. , Macromolecular Structure of Ribonucleic Acids. Reinhold , New York,
1964. 65. SPIRI N, A. S., BELlTSINA, N. V. a nd AJTKHOSCHI N, M. A. , Zh. Obshch. BioI. 25, 321
(1964). 66. STAVY, L. and GROSS, P. R ., Proc. Natl. Acad. Sci. 57, 735 (1967). 67. STEGNER, H .-E., Arch. Gyniikol. 199, 151 (1963). 68. STEVENS, A. R. , in GOLDSTEIN, L. (Ed.), The Control of Nuclear Activity, p. 189. Pren-
tice-Ha ll , Englewood Cliffs, New Jersey, 1967. 69. STEVENS, B. J . and SWIFT, H ., J . Cell BioI. 31 , 55 ( 1966). 70. SZOLLOSI, D ., J. Cell BioI. 25, 545 (1965). 71. T AKAMOTO, K., Japan. J. Exptl. Morphol. 18, 50 (1964). 72. TAKAMOTO, T. , Nature 211, 772 (1966). 73. TASHIRO, Y., MATSUU RA , S., MORIMOTO, T. a nd N AGATA, S., J. Cell BioI. 36, C5 (1968). 74. VER HEY, C. A. a nd MOYER, F. H. , J. Exptl. Zool. 164, 195 (1967). 75. WARTENBERG , H. , Z. Z ellJorsch. Mikroskop. Anat. 58, 427 (1962). 76. WISCHNITZER, S., Cellule 62, 133 (1962). 77. - - in ABERCROMBIE, M. and BRACHET, J. (Eds.), Advances in Morphogenesis, Vol. 5,
p. 131. Academic Press, New York, 1966. 78. - ibid. Vol. 6, p. 173 (1967).