Embed Size (px)
Citation preview

Photosynthesis Research 6, 193-199 ~1985 Martinus Nijhoff/Dr W. Junk Publishers,
Dordrecht, Printed in the Netherlands.
193
The use of polyclonal antibodies to identify peptides exposed on the
stroma side of the spinach thylakoid
STAN IVEY and STEVEN P. BERG
Department of Biological Sciences, University of Denver, Denver, CO 80208, USA
Kay Words: Thylakoids, antibodies, photosynthesis, photosystem II, membrane topography.
Abstract : We have raised polyclonal antibodies against an oxygen-evolving photosystem II preparatio~ Western Blot analysis of the whole serum reveals antibodies specific for at least 15 Coomassie visible bands ranging from 59 to 11 kD~ These antibodies are specific for proteins located on both sides of the membran~ Included are antibodies specific for Tris-removable peptides (33, 25 and 18 kda], which are thought to be exposed on the lumen surface of the PS II comple~ Since the whole serum agglutinates thylakoids, antibodies specific for the stroma side of the PS II complex are also present. A sub-popula~on of antibodies can be isolated by allowing the antibodies in whole serum to bind to EDTA- treated thylakoid membrane~ The antibodies which specificatly bind are cross-reactive with peptides with Mr of 59, 57, 34, 28, 27, 2B, end 23 kO~ Our data indicate that these peptides have antigenic determinants exposed on the stroma side of the thylakoid membrane
I n t r o d u c t i o n
The proteins which participate in photosynthetic electron transport and
photophosphorylation are now known to exist in discrete transmembrane
protein complexes. A remaining research goal is to understand the
topography of these various complexes in detail sufficient to explain the
structure/function relationships which exist among these protelns.
Efforts to understand the topography of the Photosystem II (PS II)
protein complex have resulted in a number of widely disparate models
[4,5,12,15]. We describe here an experimental protocol for the isolation
of antibodies specific for the stroma side of the thylakoid membrane.
These antibodies have been used to identify PS II peptides which are
exposed to the stroma side of the PS II complex. This technique has
allowed us to identify seven bands which seem to have stromal exposure.
The three extrinsic proteins associated with water oxidation, which are
proposed to be located on the lumenal side of the thylakoid [I]° do not
react significantly with antibodies specific for proteins exposed to the
stroma side of the membrane.
M a t e r i a l s a n d N e t h o d s
Highly purified PS II complexes were isolated from spinach by the
protocol of Kuwabara and Murata [i0], as we have described elsewhere [7].

194
The preparation had no Coomassie detectable PSI or cytochrome f/b6
complex. A 1.6 ml PS II preparation (2.7 mg chl/ml and 23 mg protein/ml)
was split into eight 200 ~I aliquots each of which was injected into a
different subcutaneous site on a rabbit's back. No adjuvant was employed
because the aggregated PS II complexes were already particulate.
Identical secondary injections of the same material were given after
eight weeks. The rabbits were bled and the whole serum was frozen at
-8~C until needed. Western Blotting was done as described in [17].
Prior to electrophoresis, samples were dissolved in a solution containing
35% glycerol, 6% lithium dodecylsulfate (LDS), 5% mercaptoethanol and 0.5
mM TRIS, pH 6.8. LDS-PAGE (4.5 mA/cm 2) was done on 10 to 15% gradient
gels containing 2% lithium dodecylsulfate at 4°C. Other conditions were
hke those described elsewhere [6]. Antibodies specific for the stroma
side of the thylakoid membrane were isolated by exposing whole PS II
anti-serum to unstacked thylakoids which had been prepared by washlng
stacked thylakoids three times in 200 mM sucrose containing 1 mM EDTA (pH
7.5) at 20°C. The resulting unappressed thylakoids were resuspended to
0.3 mg chl/ml in 200 mM sucrose containing I0 mM NaCI, i0 mM TES (pH 7.5)
and 3% gelatin (BioRad, Richmond, CA). The gelatin was found to
significantly reduce non-specific antibody binding to thylakoids. After
30 min the gelatin-coated thylakoids were collected by centrifugation and
the pellet was resuspended to 1 mg chl/ml in 200 mM sucrose containing i0
mM NaCI, i0 mM TES (pH 7.5), and 1% gelatin. Three ml of whole serum
were brought to 200 mM sucrose before the serum was added dropwise to 65
ml of the thylakoid suspension. After a 14 h incubation at 20°C, the
thylakoids were collected by centrifugation at 40,000Xg. The thylakoids
with bound antibody were washed three times with 200 mM sucrose
containing 20 mM Tris (pH 7.5) and 500 mM NaCI. The pellets were
stripped of specifically bound antibody by resuspension in 40 ml of 200
mM sucrose containing I00 mM glycine (pH 2.3) at 0°C. After stirring
slowly tot 25 minutes at 0°C, the stripped thylakoids were removed by
centrifugation. The antibody-containing supernatant was adjusted to I%
gelatin, the pH was adjusted to 7.5 with 1M Tris (pH 9.2) and NaCI was
added to a final concentration of 0.i M. This solution contains the
putative stroma-side antibodies.
R e s u l t s and D i s c u s s i o n
The process of Western Blotting begins with the electrophoretlc
separation of peptides in polyacrylamide gels. The peptides are then
transferred electrophoretically to nitrocellulose paper, where they bind
tightly. When the polyacrylamide gels were stained with Coomassie after
the electrophoretic transfer (data not shown), no Coomassie visible bands
remained, implying that peptide transfer from the gel was complete.
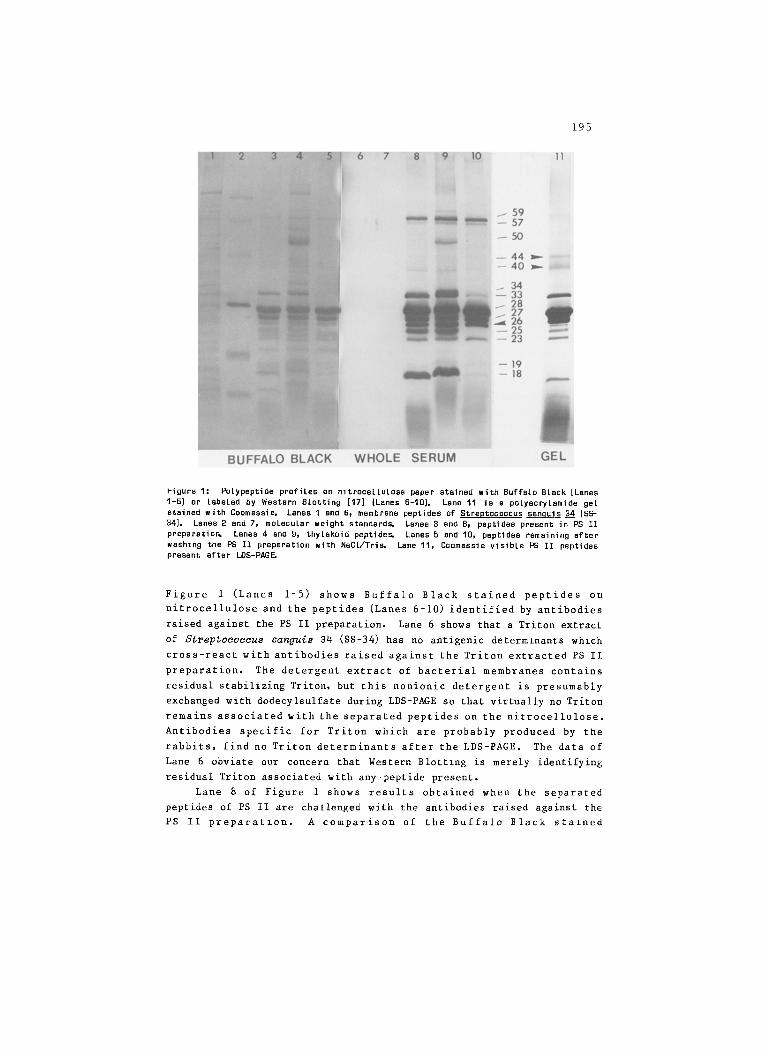
195
Figure I: Polypeptide profiles on nitrocellulose paper stained with Buffalo Black [Lanes 1-5) or labeled by Western Blotting [17] (Lanes 6-I0]. Lane 11 is s polyacrylsmide gel stained with Coomassie. Lanes 1 and 6, membrane peptides of Streptococcus san~uis 34 (SS- 34]. Lanes 2 and 7, molecular weight standard~ Lanes 3 and 8, peptides present in PS II preparatio~ Lanes 4 and g, thylakoid peptide~ Lanes 5 and 10, peptides remaining after washing the PS II preparation with NaC1/Tri~ Lane 11, Coomassie visible PS II peptides present after LDS-PAG~
Figure 1 (Lanes 1-5) shows Buffalo Black stained peptides on
nitrocellulose and the peptides (Lanes 6-10) identified by antibodies
raised against the PS II preparation. Lane 6 shows that a Triton extract
of Streptococcus sanguis 34 (SS-34) has no antlgenic determlnants which
cross-react with antibodies raised against the Triton extracted PS II
preparation. The detergent extract of bacterial membranes contains
residual stabilizing Triton, but this nonionic detergent is presumably
exchanged with dodecylsulfate during LDS-PAGE so that virtually no Triton
remains associated with the separated peptides on the nitrocellulose.
Antibodies specific for Triton which are probably produced by the
rabbits, find no Triton determinants after the LDS-PAGE. The data of
Lane 6 obviate our concern that Western Blottzng is merely identifying
residual Triton associated with any peptide present.
Lane 8 of Figure 1 shows results obtained when the separated
peptides of PS II are challenged with the antibodies raised against the
PS II preparatzon. A comparison of the Buffalo Black stazned

196
nitrocellulose (Lane 3) and the antibody stained profile of Lane 8,
indicates that antibodies have been raised against most of the Buffalo
Black visible peptides of the PS II preparation. When the separated
peptides from thylakoids are challenged with the same antibodies (Lane 9)
the peptides identified are very similar to those of the isolated PS II
complex. The most notable difference between the thylakoid profile (Lane
9) and the PS II profile (Lane 8) is the identification of a 50 kDa
peptide in the thylakoid profile. This band is never seen with
thylakoids which have been washed with EDTA at low ionic strength, or
with isolated PS II preparations. Thus, we are certain that this 50 kDa
peptide is neither a component of PSI or PS II. Together these results
indicate that antibodies raised against PS II are not cross reactive with
other thylakoid peptides (except the 50 kDa), including those associated
with PS I.
Lanes 5 and 10 of Figure 1 show the separated peptides of a PS II
preparation which has been washed with 2 M NaCI and 1M Tris. Tris
treatment is known to remove at least three peptides from PS II
preparations [19], and these three peptides are absent in the Buffalo
Black (Lane 5) and the antibody labeled (Lane i0) profiles. After the
NaCl/Tris treatment a diffuse doublet remains at 34 and 33 kDa. The
diffuse 33 kDa band appears to comigrate with the Tris-removable 33 kDa
peptide. These diffuse bands might be any one of several proteins known
to migrate between 30-35 kDa [18,11,14,16J. Since we can make no
definitive assignments for any of these bands, we can only conclude that
we might have antibodies specific for any of HBP32 [18], D2 [14,16J or
the 34 kDa intrinsic protein [11]. Further experiments are underway to
allow us to make these band assignments. When Lane 8 is compared with
Lane 10, we conclude that the whole serum contains antibodies speclfic
for the lumen side of the PS II complex. This conclusion is based on the
observation (Lane 8) of antigenically identifiable bands at 18, 25 and 33
kDa which can be removed from the PS II preparation by treatment with
NaCI and Tris [19,20]. These peptides have been shown to be extrinsic
proteins on the lumen side of the thylakoid membrane [1,20]. We have
also observed that our whole serum strongly agglutinates EDTA-treated
thylakoids suggesting that antlbodies are present whlch are specific for
the stroma side of the PS II complex as well.
Once the agglutinated EDTA-treated thylakoids mentloned above are
washed to remove nonspecifically bound antibody, the specifically bound
antibody can be removed with a low pH wash [8,13]. The thylakoids are
removed by centrifugation and the pH of the supernatant containing the
released antibodies can be titrated back to pH 7.5, thus reactivating the
antibodies. These reactivated antibodies should be those specific for
the stroma side of the PS II complex. If none of the EDTA-treated
thylakoids are turned inside-out during their isolation, there should be
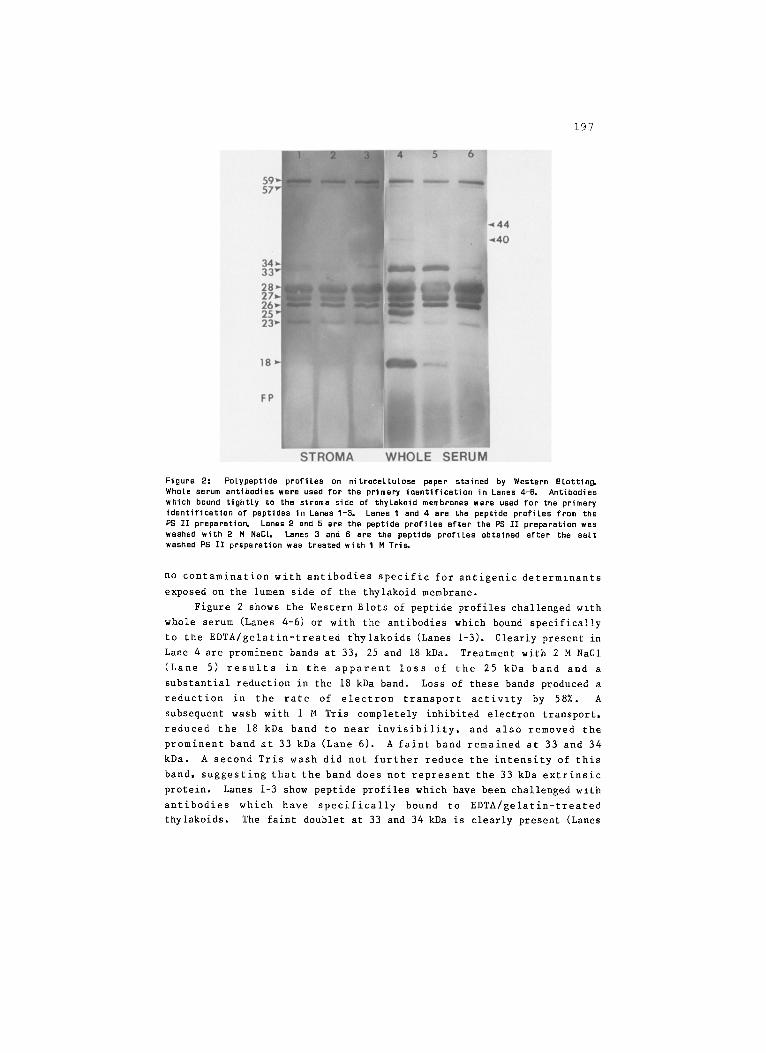
197
Figure 2: Pslypeptide profiles on nitrocellulose paper stained by Western Blotting. Whole serum antibodies were used for the primary identification in Lanes 4-6, Antibodies which bound tightly to the stroma side of thylaksid membranes were used for the primary identification of peptides in Lanes I-3. Lanes I and 4 are the peptide profiles from the PS II preparation Lanes 2 and 5 are the peptide profiles after the PS II preparation was washed with 2 M NaC1. Lanes 3 and 6 are the peptide profiles obtained after the salt washed PS II preparation was treated with 1M Tri~
no contamination with antibodies specific for antigenic determlnants
exposed on the lumen side of the thylakoid membrane.
Figure 2 shows the Western Blots of peptide profiles challenged wlth
whole serum (Lanes 4-6) or with the antibodies which bound specifically
to the EDTA/gelatin-treated thylakoids (Lanes 1-3). Clearly present in
Lane 4 are prominent bands at 33, 25 and 18 kDa. Treatment with 2 M NaCI
(Lane 5) results in the apparent loss of the 25 kDa band and a
substantial reduction in the 18 kDa band. Loss of these bands produced a
reduction in the rate of electron transport activlty by 58%. A
subsequent wash with 1 M Tris completely inhibited electron transport,
reduced the 18 kDa band to near invisibility, and also removed the
prominent band at 33 kDa (Lane 6). A faint band remained at 33 and 34
kDa. A second Tris wash did not further reduce the intensity of this
band, suggesting that the band does not represent the 33 kDa extrinsic
protein. Lanes 1-3 show peptide profiles which have been challenged with
antibodies which have specifically bound to EDTA/gelatin-treated
thylakoids. The faint doublet at 33 and 34 kDa is clearly present (Lanes

198
I-2) except in Lane 3 where only the 34 kDa band is visible. Very faint
bands at 25 and 18 kDa appear to be present, suggesting that there may be
very alight whole serum contamination. The faint band at 34 kDa which
appears to comigrate with the 33 kDa extrinsic protein [11] could be
HBP32 [18J, the 34 kDa intrinsic protein [11] or D2 [14,16J. If we
ignore any faint bands which might be due to contamination with whole
serum antibodies, we would conclude that the major bands identlfied in
Lanes 1-3 are proteins which have antigenic determinants exposed on the
stroma side of the PS II complex. These bands include peptides with Mr
of 59, 57, 34, 28, 27, 26 and 23 kDa.
Stromal exposure of PS II peptides with similar molecular weights
have been suggested by others. For instance, Andersson et al. [3] raised
antlbodies against purified light-harvesting complex. They found
antibodies specific for peptides of 28, 27, 25, and 23.5 kDa. They also
found that the antiserum would agglutinate unstacked right-side-out and
inside-out thylakoids, allowing them to conclude that the light-
harvestlng complex was transmembraneous. However, they did not
definitively show which, if any, of the peptides actually had antigenic
determinants on both sides of the membrane. Our data identlfied peptides
which are exposed on the stromal surface of the thylakoid membrane, but
at this time we are unable to definitely assign any of these visible
bands or to correlate them with the observations ot Andersson et al.
However, work is underway with "green'" gels, urea gels, and secondary
LDS-PAGE to make these assignments. Since there is evidence that the
herbicide binding protein has a stromal exposure [21] and since Vermaas
et al. L18] has shown that HBP32 comigrates with the Tris removable
LRP32, we would tentatively identify the 34 kDa band as HBP32.
Figure i (Lane 11) shows the Coomassie visible peptides in a
polyacrylamide gel after LDS-PAGE. Clearly visible are peptides at 40
and 44 kDa which are probably CP43 and CP47 respectively [5]. Western
Blot analysis (Lanes 8-10) shows that these bands are visible when the
peptides are challenged with whole serum, indicating that these peptides
have been transferred to the nitrocellulose and that antlbodies to these
peptides are present. In spite of their presence, the 40 and 44 kDa
peptldes are not stained with Buffalo Black (Figure i, Lanes 3 and 5),
and these peptides are not identified with stroma-side antibodies (Figure
2, Lanes 1-3). This suggests that there is not significant direct
exposure of the PS II reaction center to the aqueous stroma.
With our PS II antibodies, we routinely observe an antlgenically
identified doublet at about 59 and 57 kDa. As with the peptides
discussed above we can not yet offer a clear assignment for these bands,
but we believe that they are probably two oligomeric species of LHCPI or
LHCP2 which have been described by Anderson [2] in her re-electrophoresis
studies. Andersson et al. found a similar high molecular weight band

199
when their nitrocellulose was challenged with antibodies specific for LHC
[3]. In summary, we report here that immunization of rabbits wzth
purified PS II complexes results in the production of antibodies specific
for many of the peptides present in the complex. From these antlbodies,
it is possible to select those which are specific for antigenic
determinants exposed on the stromal surface of the thylakoid membrane.
By coupling the technique of Western Blotting to the selection of stroma-
side-specific antibodies, we can identify peptides which are exposed on
the stroma surface of the PS II complex. Further work is underway to
identzfy the peptides exposed on the lumen surface of the thylakoid, and
to unambiguously assign the identity of the antibody-identified peptides
exposed on both surfaces.
Acknowledgement
This work was supported by a grant (82-CRCR-I-1125) from the Competitive
Grants Office of the USDA to SPB. The authors are grateful to Dr. Victor
Reusch for generously supplying the SS-34 membrane preparation used in
this study.
~ f e ~ n ~ 8
I. Akerlund, H-E and Jsnsson, C, (1981) FEBS Letters 124:229-232. 2. Anderson, JM, (1980) Biochim. Biophys. Acta 581:113-126. 3. Andersson, 8, Anderson, JM, and Ryrie, IJ, (1982) Eur. J. 8iochem. 123:465-472. 4. Barber, J, (1984) Trends in Biochem. Sci. 5:79-80. 5. Carom, EL, and Green, BR, (1983) J. Cell Biochem. 23:171-179. 6. Chua, N, and Bennoun, P, (1975) Proc. Natl. AcacL Sci. (USA) 72:2175-2179. 7. Dunahay, TG, Staehelin, LA, Seibert, M, Ogilvie, PD, and Berg, SP, I19B4) Bioshim.
Biophys. Acts 764:179-193. 8. Eisen, HN, Aronson, N, end Simms, ES, (1964) in Methods in Medical Research, Vcl 10,
pp 91-99, (HN Eisen, ed.) Year Book Medical Publishers, NY. 9. Guikema, JA, and Sherman, LA, (1983) Arch. Biochem. 8iophys. 220:155-166.
10. Kuwabera, T, and Murats, N, (1982] Plant Cell Physiol. 23:533-539. 11. Metz, JG and Seibert, M, (1984) Plant Physiol. 76;829-832. 12. Murate, N, Miyao, M, end Kuwsbara, T, (1983) in "The Oxygen Evolving System of
Photosynthesis", Academic Press (Japan) pp 213-222. 13. Onoue, K, Yagi, Y, and Pressman, O, (1966) Immunochem. 2:181-196. 14. Rasmussen, OF, Boskjans, G, Stummann, BM, and Henningsen, KW, [1984) Plant Mol. Biol.
3:181-199. 15. Renger, G, and Akerlund, R-E, (1983) in "The Oxygen Evolving System of
Photosynthesis", Academic Press (Japan) pp 209-212. 16. Rochaix, JO, (1981) Experientie 37:329-340. 17. Towbin, H, Btaehelin, T, and Gordon, J, (1979) Pros. Natl. Acad. Sci. (USA) 76:4350-
4354. 18. Vermaas, WFJ, Steinback, KE, nd Arntzen, CJ, (1984) Arch. Biochem. Biophys. 231:226-
232 •
19. Yamamoto, Y, Osi, M, Tamure, N, and Nishimura, M, [1881} FEBS Letters 193:265-268. 20. Akerlund, H~E, Jansson, C, and Andersson, B, (1982) Biochim. Biophys. Acta 681:1-10. 21. Steinback, KE, Pfiste, K, and Arntzen, CJ, [1981) Z. Naturforsch. 38C:98-108.





![Multiphoton imaging to identify grana, stroma thylakoid ...cellstudio.org/.../BMCPlant2014_Multiphoton-grana.pdf · and grana in plant [11,16-27]. Inside a chloroplast, both starch](https://img.pdfslide.net/doc/110x75/5fb8149a7316217d4776dc5c/multiphoton-imaging-to-identify-grana-stroma-thylakoid-and-grana-in-plant-1116-27.jpg)























