Embed Size (px)
Citation preview

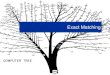
Tree Searching MethodsTree Searching Methods
• Exhaustive search (exact)
• Branch-and-bound search (exact)
• Heuristic search methods (approximate)– Stepwise addition– Branch swapping– Star decomposition

Exhaustive Search
12
12
11
12
13
13
13
13
13
13
12
13
13
13
13

Searching for trees
• Generation of all possible trees
B
C
A
D
D
D
B
CD
A
B
CD
B C
DB
A
1.Generate all 3 trees for first 4 taxa:

Searching for trees
B
C
D
AE
EE
C
DE
AB
C
DE
BA
C
DB
AE
D
EB
AC
C
EB
AD
2. Generate all 15 trees for first 5 taxa:
(likewise for each of the other two 4-taxon trees)

Searching for trees
3. Full search tree:
EA
CB
D
DA
CB
E
DA
EB
C
DA
EC
B
CB
ED
A
CA
DB
E
CA
EB
D
CA
ED
B
DB
EC
A
EA
DC
BE
B
DC
A
BA
DC
E
BA
EC
D
BA
ED
C
D
A
B
C
B
A
C
D
A
B
C
C
A
B
D
DB
EA
C

Searching for trees
Branch and bound algorithm:
The search tree is the same as for exhaustive search, with tree lengths for a hypothetical data set shown in boldface type. If a tree lying at a node of this search tree has a length that exceeds the current lower bound on the optimal tree length, this path of the search tree is terminated (indicated by a cross-bar), and the algorithm backtracks and takes the next available path. When a tip of the search tree is reached (i.e., when we arrive at a tree containing the full set of taxa), the tree is either optimal (and hence retained) or suboptimal (and rejected). When all paths leading from the initial 3-taxon tree have been explored, the algorithm terminates, and all most-parsimonious trees will have been identified. Asterisks indicate points at which the current lower bound is reduced. Circled numbers represent the order in which phylogenetic trees are visited in the search tree.
1
*229
EA
CB
D
DA
CB
E
DA
EB
C
DA
EC
B
CB
ED
A
CA
DB
E
CA
EB
D
DB
EC
A
D
A
B
C
A
B
C
233
235
237 237245
251258
C
A
B
D
280
221 213
B
A
C
D
234
*241
*242
242245
246247
249
268C
A
ED
B
245
241
241
244248
251
232
226
233
235
251
262
243
227
2
3
11
12
13-19
4-10
DB
EA
C
20
21
22
26
23
24
25
27
28-34

Stepwise Addition (in a nutshell)
3
2
1
42
31
43
21
34
21

Searching for trees
Stepwise addition
A greedy stepwise-addition search applied to the example used for branch-and-bound. The best 4-taxon tree is determined by evaluating the lengths of the three trees obtained by joining taxon D to tree 1 containing only the first three taxa. Taxa E and F are then connected to the five and seven possible locations, respectively, on trees 4 and 9, with only the shortest trees found during each step being used for the next step. In this example, the 233-step tree obtained is not a global optimum. Circled numbers indicate the order in which phylogenetic trees are evaluated in the stepwise-addition search.
EA
CB
D
DA
CB
E
DA
EB
C
DA
EC
B
CB
ED
A
D
A
B
C
A
B
C
233*
235
237 237245
251258
C
A
B
D
280
221 213
B
A
C
D
235
251
262
243
227
2
1
2
3
5
6
7
8
4
9
10-16

Stepwise Addition Variants
• As Is– add in order found in matrix
• Closest– add unplaced taxa that requires smallest increase
• Furthest– add unplaced taxa that requires largest increase
• Simple– Farris’s (1970) “simple algorithm” uses a set of pairwise
reference distances
• Random– random permutation of taxa is used to select the order

Branch swappingNearest Neighbor Interchange (NNI)
E
A
C
B
D
A
D
E
CB
DA
CB
E

Branch swappingSubtree Pruning and Regrafting (SPR)
D
AB
C
GF
E
D
GF
E
AB
C
G
DE
F
BA
C
a

Branch swappingTree Bisection and Reconnection (TBR)
D
AB
C
GF
ED
GF
E
AB
C
G
DE
F
BC
A
G
DE
F
BA
C
G
DE
F
CA
B

Reconnection limits in TBR
1
2 3 45
6
x zy
r
s
t u v
w
1
2 3 45
6
x zx'
u v
w1
2 4 3 5
6
1
2 3 45
6
0 01
1
2
2
Reconnection distances:

(D)
1
2 3 45
6
y
r
s
v
wy'
3
1 2 54
6
01
1
2 3 45
6
1
1
1
0Reconnection distances:
In PAUP*, use “ReconLim” to set maximum reconnection distanceIn PAUP*, use “ReconLim” to set maximum reconnection distance
Reconnection limits in TBR

Star-decomposition search
1
2
3
4
5
1
3
2
4
5
3
5
1
2
4
•••
4
5
1
2
3
1
2
3
4
5
14
3
2
5
12
3
4
5
15
3
2
4
Step 1
Step 2 Step 3

Overview of maximum likelihood as used Overview of maximum likelihood as used in phylogeneticsin phylogenetics
• Overall goal: Find a tree topology (and associated parameter Overall goal: Find a tree topology (and associated parameter estimates) that maximizes the probability of obtaining the observed estimates) that maximizes the probability of obtaining the observed data, given a model of evolutiondata, given a model of evolution
Likelihood(hypothesis) Likelihood(hypothesis) Prob(dataProb(data||hypothesis)hypothesis)
Likelihood(tree,model) = k Prob(observed sequences|tree,model)Likelihood(tree,model) = k Prob(observed sequences|tree,model)
[[notnot Prob(tree Prob(tree||data,model)]data,model)]

Computing the likelihood of a single treeComputing the likelihood of a single tree
1 1 jj NN(1) C…GGACA…(1) C…GGACA…CC…GTTTA…C…GTTTA…C(2) C…AGACA…(2) C…AGACA…CC…CTCTA…C…CTCTA…C(3) C…GGATA…(3) C…GGATA…AA…GTTAA…C…GTTAA…C(4) C…GGATA…(4) C…GGATA…GG…CCTAG…C …CCTAG…C
(1)(1)
(2)(2)
(3)(3)
(4)(4)
CCCC AA GG
(6)(6)
(5)(5)

Computing the likelihood of a single treeComputing the likelihood of a single tree
ProbProb
CCCC AA GG
AA
AA
Likelihood at site Likelihood at site jj = =
+ Prob+ Prob
CCCC AA GG
AA
CC
ProbProb
CCCC AA GG
TT
TT+ … ++ … +
But use Felsenstein (1981) pruning algorithmBut use Felsenstein (1981) pruning algorithm

Computing the likelihood of a single treeComputing the likelihood of a single tree
€
€
L = L1L2L LN = L jj=1
N
∏
€
lnL = lnL1 + lnL2 +L lnLN = lnL1
j=1
N
∑
Note: PAUP* reports -ln Note: PAUP* reports -ln LL, so lower -ln , so lower -ln LL implies higher likelihood implies higher likelihood

Finding the maximum-likelihood treeFinding the maximum-likelihood tree(in principle)(in principle)
• Evaluate the likelihood of each possible Evaluate the likelihood of each possible tree for a given collection of taxa.tree for a given collection of taxa.
• Choose the tree topology which Choose the tree topology which maximizes the likelihood over all maximizes the likelihood over all possible trees.possible trees.

Probability calculations require…Probability calculations require…• An explicit model of substitution that specifies change An explicit model of substitution that specifies change
probabilities for a given branch lengthprobabilities for a given branch length
“Instantaneous rate matrix”“Instantaneous rate matrix”
Jukes-CantorJukes-CantorKimura 2-parameterKimura 2-parameterHasegawa-Kishino-Yano (HKY)Hasegawa-Kishino-Yano (HKY)Felsenstein 1981, 1984Felsenstein 1981, 1984General time-reversibleGeneral time-reversible
€
Q =
π ArAA π CrAC π GrAG π TrAT
π ArCA π CrCC π GrCG π TrCT
π ArGA π CrGC π GrGG π TrGT
π ArTA π CrTC π GrTG π TrTT
⎛
⎝
⎜ ⎜ ⎜ ⎜
⎞
⎠
⎟ ⎟ ⎟ ⎟
€
P(v) = eQν
• An estimate of optimal branch lengths in units of An estimate of optimal branch lengths in units of expected amount of change (expected amount of change ( = rate x time) = rate x time)

For example:For example:
€
Q =
− α α α
α − α α
α α − α
α α α −
⎛
⎝
⎜ ⎜ ⎜ ⎜
⎞
⎠
⎟ ⎟ ⎟ ⎟
Jukes-Cantor (1969)Jukes-Cantor (1969)
€
Q =
− β α β
β − β α
α β − β
β α β −
⎛
⎝
⎜ ⎜ ⎜ ⎜
⎞
⎠
⎟ ⎟ ⎟ ⎟
Kimura (1980) “2-parameter”Kimura (1980) “2-parameter”
€
Q =
− π Cβ π Gα π Tβ
π Aβ − π Gβ π Tα
π Aα π Cβ − π Tβ
π Aβ π Cα π Gβ −
⎛
⎝
⎜ ⎜ ⎜ ⎜
⎞
⎠
⎟ ⎟ ⎟ ⎟
Hasegawa-Kishino-Yano (1985)Hasegawa-Kishino-Yano (1985)
€
Q =
π ArAA π CrAC π GrAG π TrAT
π ArCA π CrCC π GrCG π TrCT
π ArGA π CrGC π GrGG π TrGT
π ArTA π CrTC π GrTG π TrTT
⎛
⎝
⎜ ⎜ ⎜ ⎜
⎞
⎠
⎟ ⎟ ⎟ ⎟
General-Time ReversibleGeneral-Time Reversible

E.g., transition probabilities forE.g., transition probabilities forHKY and F84:HKY and F84:
Pij t( ) =
π j +π j1
Π j
−1⎛
⎝ ⎜ ⎜
⎞
⎠ ⎟ ⎟ e
−μν +Π j −π j
Π j
⎛
⎝ ⎜ ⎜
⎞
⎠ ⎟ ⎟ e
−μνA (i= j)
π j +π j
1Π j
−1⎛
⎝ ⎜ ⎜
⎞
⎠ ⎟ ⎟ e
−μν −π j
Π j
⎛
⎝ ⎜ ⎜
⎞
⎠ ⎟ ⎟ e
−μνA (i≠ j, transition)
π j 1−e−μν( ) (i≠ j, transversion)
⎧
⎨
⎪ ⎪ ⎪ ⎪ ⎪
⎩
⎪ ⎪ ⎪ ⎪ ⎪

A Family of Reversible Substitution ModelsA Family of Reversible Substitution Models
GTR
SYMTrN
F81
JC
K3ST
K2P
HKY85F84
Equal base frequencies
3 substitution types(transitions,2 transversion classes)
2 substitution types(transitions vs. transversions)
3 substitution types(transversions, 2 transition classes)
2 substitution types(transitions vs.transversions)
Single substitution type
Equal basefrequencies
Single substitution typeEqual base frequencies
(general time-reversible)
(Tamura-Nei)
(Hasegawa-Kishino-Yano)
(Felsenstein)
Jukes-Cantor
(Kimura 2-parameter)
(Kimura 3-subst. type)
(Felsenstein)

The Relevance of Branch LengthsThe Relevance of Branch LengthsC C A A A A A A A A
A
C
C C A A A A A A A A
CA

When does maximum likelihood work When does maximum likelihood work better than parsimony?better than parsimony?
• When you’re in the “Felsenstein Zone”When you’re in the “Felsenstein Zone”
AA CC
BB DD
(Felsenstein, 1978)(Felsenstein, 1978)

In the Felsenstein ZoneIn the Felsenstein Zone
AA CC GG TTAA -- 55 66 22CC 55 -- 33 88GG 66 33 -- 11TT 22 88 11 --
Substitution rates:Substitution rates:
Base frequencies:Base frequencies: A=0.1A=0.1 C=0.2C=0.2 G=0.3G=0.3 T=0.4T=0.4
AA BB
CC DD
0.10.1
0.10.1 0.10.1
0.80.8 0.80.8

In the Felsenstein ZoneIn the Felsenstein Zone
0
0.2
0.4
0.6
0.8
1
0 5000 10000
Sequence Length
parsimonyML-GTR
Pro
port
ion
corr
ect

The long-branch attraction (LBA) problemThe long-branch attraction (LBA) problem
Pattern typePattern type
11 44AA I = Uninformative (constant)I = Uninformative (constant) AA
A AA A 22 33
The true phylogeny ofThe true phylogeny of1, 2, 3 and 41, 2, 3 and 4
(zero changes required on any (zero changes required on any tree)tree)

The long-branch attraction (LBA) problemThe long-branch attraction (LBA) problem
Pattern typePattern type
11 44AA I = Uninformative (constant)I = Uninformative (constant) AAAA II = UninformativeII = Uninformative GG
A AA A 22 33
The true phylogeny ofThe true phylogeny of1, 2, 3 and 41, 2, 3 and 4
(one change required on any tree)(one change required on any tree)

The long-branch attraction (LBA) problemThe long-branch attraction (LBA) problem
Pattern typePattern type
11 44AA I = Uninformative (constant)I = Uninformative (constant) AAAA II = UninformativeII = Uninformative GGCC III = UninformativeIII = Uninformative GG
A AA A 22 33
The true phylogeny ofThe true phylogeny of1, 2, 3 and 41, 2, 3 and 4
(two changes required on any tree)(two changes required on any tree)

The long-branch attraction (LBA) problemThe long-branch attraction (LBA) problem
Pattern typePattern type
11 44AA I = Uninformative (constant)I = Uninformative (constant) AAAA II = UninformativeII = Uninformative GGCC III = UninformativeIII = Uninformative GGG G IV = IV = MisinformativeMisinformative GG
A AA A 22 33
The true phylogeny ofThe true phylogeny of1, 2, 3 and 41, 2, 3 and 4
(two changes required on true tree)(two changes required on true tree)

The long-branch attraction (LBA) problemThe long-branch attraction (LBA) problem
GG 44
AA 22
AA 33
GG 11
… … but this tree needs only one stepbut this tree needs only one step

Concerns about statistical properties Concerns about statistical properties and suitability of models and suitability of models
(assumptions)(assumptions)
ConsistencyConsistency
If an estimator converges to the true value of a If an estimator converges to the true value of a parameter as the amount of data increases toward parameter as the amount of data increases toward infinity, the estimator is infinity, the estimator is consistentconsistent..

When do both methods fail?When do both methods fail?• When there is insufficient phylogenetic signal...When there is insufficient phylogenetic signal...
22
11 33
44

When does parsimony work “better” When does parsimony work “better” than maximum likelihood?than maximum likelihood?
• When you’re in the Inverse-Felsenstein (“Farris”) zoneWhen you’re in the Inverse-Felsenstein (“Farris”) zone
AA
BB
CC
DD
(Siddall, 1998)(Siddall, 1998)

Siddall (1998) parameter space Siddall (1998) parameter space
a
a
b
b
b
Both methods do poorly
Parsimony has higheraccuracy than likelihood
Both methods do well
pa
pb0 0.75
0.75

Parsimony vs. likelihood in the Inverse-Felsenstein ZoneParsimony vs. likelihood in the Inverse-Felsenstein Zone
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
20 100 1,000 10,000 100,000
Sequence length
ParsimonyML/JC
15%67.5%
67.5%
(expected differences/site)
Acc
ura
cy

Why does parsimony do so well in theWhy does parsimony do so well in theInverse-Felsenstein Inverse-Felsenstein zone?zone?
A
A
C
C
AC
A
A
C
C
AG
A
C G
C
A
A
C
CAC
AC
True synapomorphyTrue synapomorphy
Apparent synapomorphiesApparent synapomorphiesactually due toactually due tomisinterpreted homoplasymisinterpreted homoplasy

Parsimony vs. likelihood in the Felsenstein ZoneParsimony vs. likelihood in the Felsenstein Zone
15%
67.5% 67.5%
Acc
ura
cy
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
20 100 1,000 10,000 100,000
ParsimonyML/JC
(expected differences/site)
Sequence length

From the Farris Zone to the Felsenstein ZoneFrom the Farris Zone to the Felsenstein Zone
CC
DD
AA
BB
CC
DD
AA
BB
CC
DD
AA
BB
BB
CC
DD
AA
BB
DD
CC
AA
External branches = 0.5 or 0.05 substitutions/site, Jukes-Cantor model of nucleotide substitutionExternal branches = 0.5 or 0.05 substitutions/site, Jukes-Cantor model of nucleotide substitution

0
0.2
0.4
0.6
0.8
1.0
0.05 0.04 0.03 0.02 0.01 0 0.01 0.02 0.03 0.04 0.05
100 sites
1,000 sites
10,000 sites ML/JC
Length of internal branch ( d)Farris zone Felsenstein zone
0
0.2
0.4
0.6
0.8
0.05 0.04 0.03 0.02 0.01 0 0.01 0.02 0.03 0.04 0.05
Length of internal branch ( d)Farris zone Felsenstein zone
100 sites
1,000 sites
10,000 sites
1.0
Acc
ura
cyA
ccu
racy
ParsimonyParsimony
LikelihoodLikelihood
SimulationSimulationresults:results:

Maximum likelihood models are Maximum likelihood models are oversimplifications of reality. If I assume the oversimplifications of reality. If I assume the
wrong model, won’t my results be meaningless?wrong model, won’t my results be meaningless?
• Not necessarily (maximum likelihood is pretty robust)Not necessarily (maximum likelihood is pretty robust)

Model used for simulation...Model used for simulation...
AA CC GG TTAA -- 55 66 22CC 55 -- 33 88GG 66 33 -- 11TT 22 88 11 --
Substitution rates:Substitution rates:
Base frequencies:Base frequencies: A=0.1A=0.1 C=0.2C=0.2 G=0.3G=0.3 T=0.4T=0.4
AA BB
CC DD
0.10.1
0.10.1 0.10.1
0.80.8 0.80.8

Performance of ML when its model is Performance of ML when its model is violated (one example)violated (one example)
0
0.2
0.4
0.6
0.8
1
100 1000 10000
Sequence Length
parsimonyML-JCML-K2PML-HKYML-GTR

Among site rate heterogeneity
• Proportion of invariable sites– Some sites don’t change do to strong functional or structural constraint (Hasegawa et
al., 1985)
• Site-specific rates– Different relative rates assumed for pre-assigned subsets of sites
• Gamma-distributed rates– Rate variation assumed to follow a gamma distribution with shape parameter
Lemur AAGCTTCATAG TTGCATCATCCA …TTACATCATCCAHomo AAGCTTCACCG TTGCATCATCCA …TTACATCCTCATPan AAGCTTCACCG TTACGCCATCCA …TTACATCCTCATGoril AAGCTTCACCG TTACGCCATCCA …CCCACGGACTTAPongo AAGCTTCACCG TTACGCCATCCT …GCAACCACCCTCHylo AAGCTTTACAG TTACATTATCCG …TGCAACCGTCCTMaca AAGCTTTTCCG TTACATTATCCG …CGCAACCATCCT
equal rates?

Performance of ML when its model is Performance of ML when its model is violated (another example)violated (another example)
...
0
0.02
0.04
0.06
0.08
0 1 2
Rate
=50
=200
Modeling among-site rate variation with a gamma distribution...Modeling among-site rate variation with a gamma distribution...
……can also estimate a proportion of “invariable” sites (pcan also estimate a proportion of “invariable” sites (p invinv))
=2
=0.5
Fre
quen
cy

Performance of ML when its model is Performance of ML when its model is violated (another example)violated (another example)
Sequence Length
Proportion Correct
Tree a = 0.5, =0.5pinv a = 1.0, =0.5pinv a = 1.0, =0.2pinv
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
GTRigGTRgHKYgGTRiHKYiGTRerHKYerparsimony
HKYigGTRigGTRgHKYgGTRiHKYiGTRerHKYerparsimony
HKYig
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
GTRigGTRgHKYgGTRiHKYiGTRerHKYerparsimony
HKYig
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
GTRigHKYigGTRgHKYgGTRiHKYiGTRerHKYerParsimony
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
GTRigHKYigGTRgHKTgGTRiHKYiGTRerHKYerparsimony
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
GTRigHYYigGTRgHKYgGTRiHKYiGRTerHKYerparsimony
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
GTRigGTRgHKYgGTRiHKYiGTRerHKYerparsimony
HKYig
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
GTRigGTRgHKYgGTRiHKYiGTRerHKYerparsimony
HKYig
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000

“MODERATE”–Felsenstein zone
= 1.0, pinv=0.5
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
JCerJC+GJC+IJC+I+GGTRerGTR+GGTR+IGTR+I+Gparsimony

“MODERATE”–Inverse-Felsenstein zone
00.10.20.30.40.50.60.70.80.91
100 1000 10000 100000
JCerJC+GJC+IJC+I+GGTRerGTR+GGTR+IGTR+I+Gparsimony

Bayesian Inference in Phylogenetics
• Uses Bayes formula:
Pr(|D) = Pr(D|) Pr() Pr(D)
Pr(D|) Pr()
L() Pr()
• Calculation involves integrating over all tree topologies and model-parameter values, subject to assumed prior distribution on parameters
(( =tree topology, =tree topology, branch-lengths, and branch-lengths, and substitution-model substitution-model parameters)parameters)

Bayesian Inference in Phylogenetics
• To approximate this posterior density (complicated multidimensional integral) we use Markov chain Monte Carlo (MCMC)– Simulated Markov chain in which transition probabilities are
assigned such that the stationary distribution of the chain is the posterior density of interest
– E.g., Metropolis-Hastings algorithm: Accept a proposed move from one state to another state * with probability min(r,1) where
r = Pr(*|D) Pr(| *)Pr(|D) Pr(*| )
– Sample chain at regular intervals to approximate posterior distribution
• MrBayes (by John Huelsenbeck and Fredrik Ronquist) is most popular Bayesian inference program

AB
C D
AB
C D
Like
lihoo
d
Iterations
A brief intro to Markov chain Monte Carlo (MCMC)
A
B
C D
...
If the chain is run “long enough”, the stationary distribution of states in the chain will represent a good approximation to the target distribution (in this case, the Bayesian posterior)
1. Initialize the chain, e.g., by picking a random state X0 (topology,branch lengths, substitution-model parameters) from the assumed prior distribution
A
B
C
D
AB|CD
A
B
C
D
AB|CD
AB
C D
BC|AD
AB
C D
BC|AD
AB
C D
BC|AD
AB
C D
BC|AD
B
CD
A
AC|BDAB|CD
A
B
C
D
€
(X,Y ) = min 1,Pr Y | D( )q(X |Y )
Pr X | D( )q(Y | X)
⎛
⎝ ⎜
⎞
⎠ ⎟= min 1,
π (Y )
π (X)×
Pr(D |Y )
Pr(D | X)×q(X |Y )
q(X |Y )
⎛
⎝ ⎜
⎞
⎠ ⎟
2. For each time t, sample a new candidate state Y from some proposal distribution q(.|X t) (e.g., change branch lengths or topology plus branch lengths)
Calculate acceptance probability
3. If Y is accepted, let Xt+1 = Y; otherwise let Xt+1 = Xt
“burn in”

Model-based distancesModel-based distances• Can also calculate pairwise distances based on these modelsCan also calculate pairwise distances based on these models• These distances estimate the number of substitutions per site These distances estimate the number of substitutions per site
that have accumulated since the two sequences shared a that have accumulated since the two sequences shared a common ancestor, allowing for superimposed substitutions common ancestor, allowing for superimposed substitutions (“multiple hits”)(“multiple hits”)
• E.g.:E.g.:– Jukes-Cantor distanceJukes-Cantor distance– Kimura 2-parameter distanceKimura 2-parameter distance– General maximum-likelihood distances available for other General maximum-likelihood distances available for other
modelsmodels

1 3
42
a d
e
c
b
€
−d12 −
d13 d23 −
d14 d24 d34 −
1
2
3
4
1 2 3 4
p12 = a+bp13 = a+c+dp14 = a+c+ep23 = b+c+dp24 = b+c+ep34 = d+e
pij = dij for all i and j if the treetopology is correct and distancesare additive
Distance-based optimality criteria“Additive trees”

Distances in general will not be additive, sochoose optimal tree according to one of the
following criteria (objective functions):
"Goodness - of - fit" : minimize wij pij−diji < j∑
r
Typicall , y r = 2 (least-squares) and wij = 1/dij2 ("Fitch-
Margoliash" method)
"Minimum- "evolution : minimize vkk=1
#branches
∑ or vkk=1
#branches
∑

Distance-based optimality criteriaMinimum evolution and least-squares
Pongo
Lemur catta
Pan
Homo sapiens
Gorilla
0.044
0.085
0.286
0.015
0.0500.045
0.050
0.39646 0.39021 0.0000390.39838 0.39602 0.0000060.09506 0.09507 0.0000000.37222 0.38084 0.0000740.11172 0.11011 0.0000030.11431 0.11592 0.0000030.37096 0.37096 0.0000000.18107 0.18894 0.0000620.19399 0.19475 0.0000010.18820 0.17958 0.000074
0.000261
pijdij SS
Least-Squares
0.286110.044360.015110.044630.050440.050380.084850.57588
Minumumevolution(ME)
LS branch lengths




![Appendix A. Exact Search Strings...A-1 Appendix A. Exact Search Strings PubMed® search strategy (January 12, 2016) #1 "Dystocia"[Mesh] OR "Dystocia"[tiab] OR "Dystocias"[tiab] OR](https://img.pdfslide.net/doc/110x75/5fe2bbac07a41052623b8649/appendix-a-exact-search-strings-a-1-appendix-a-exact-search-strings-pubmed.jpg)