Embed Size (px)
Citation preview

J. Physiol. (1987), 391, pp. 209-218 209With 7 text-figuresPrinted in Great Britain
FEVER AND ACUTE PHASE RESPONSE INDUCED IN RABBITS BYHUMAN RECOMBINANT INTERFERON-y
BY AKIO MORIMOTO, NAOTOSHI MURAKAMI, MITSUAKI TAKYADA*,SHYUICHI TESHIROGI* AND TATSUO WATANABE
From the Department of Physiology, Yamaguchi University School of Medicine,Ube, Yamaguchi 755, Japan
(Received 15 December 1986)
SUMMARY
1. Intravenous (i.v.) and intracerebroventricular (I.c.v.) injections of human re-combinant interferon-y (IFN-y) produced dose-dependent fevers in rabbits. The feverinduced by i.v. injection was monophasic and the maximum elevation occurred80-110 min after injection. The fever induced by i.c.v. injection was observed fromabout 20 min after injection and was remarkably prolonged over 4 h.
2. The development of pyrogenic tolerance to IFN-y was observed when rabbitswere given i.v. injections on 3 successive days. Furthermore, the pyrogenicity ofIFN-y was significantly attenuated by heating at 60 'C for 40 min. The i.v. injectionof IFN-y enhanced the febrile response induced by endotoxin but had no effect onthat induced by endogenous pyrogen.
3. The i.v. injection of a large dose of IFN-y (6 x 106 units/kg) induced an acutephase response, which included a reduction in plasma concentration of iron and zinc.
4. The present results suggest that IFN-y released from lymphocytes is one of theendogenous mediator proteins responsible for producing fever and acute phase re-sponse.
INTRODUCTION
It is well known that animals with infection or inflammation exhibit severalresponses, among them the elevation of body temperature (fever) (Atkins, 1960),changes in cardiovascular response (Shido & Nagasaka, 1986) and changes in adeno-hypophyseal hormone release (Moberg, 1971; Kasting & Martin, 1982; Kasting,Mazurek & Martin, 1985). In addition, during such conditions changes in the plasmalevel of certain trace metals, activation of hepatic protein synthesis and an increasein circulating leucocyte count are also induced. As a group, these changes are termedthe acute phase response (Gordon & Koj, 1985). The acute phase response is nowgenerally recognized as representing a primary host defence response (Kluger,Ringler & Anver 1975; Kluger & Rothenberg, 1979). Increasing evidence has beenadduced that these responses are mediated by endogenous pyrogen (Kampschmidt,1980), a substance recently known as interleukin-1 (IL-1) (Dinarello, 1984), which is
* Present address: Sogo Biomedical Laboratories, Kawagoe Matoba, Saitama 350, Japan.
A. MORIMOTO AND OTHERS
released by phagocytic leucocytes in response to such pathogenic stimuli as bacterialendotoxin.
In recent advances of research in immunology, immunoregulatory functions ofmacrophages have been extensively described for a variety of lymphocyte functions(see the text by Bellanti, 1985; review: Pick, 1985). As shown in Fig. 1, endogenous
.',: Antigen
Macrophage
Endogenous pyrogenW o ~ ~~~~~~(L-1)
\ * ^ Helper T lymphocyte
IFN-ly ,
\ . IL-2
O T lymphocyte
IFN-,y A /
Qi) Natural killer cell
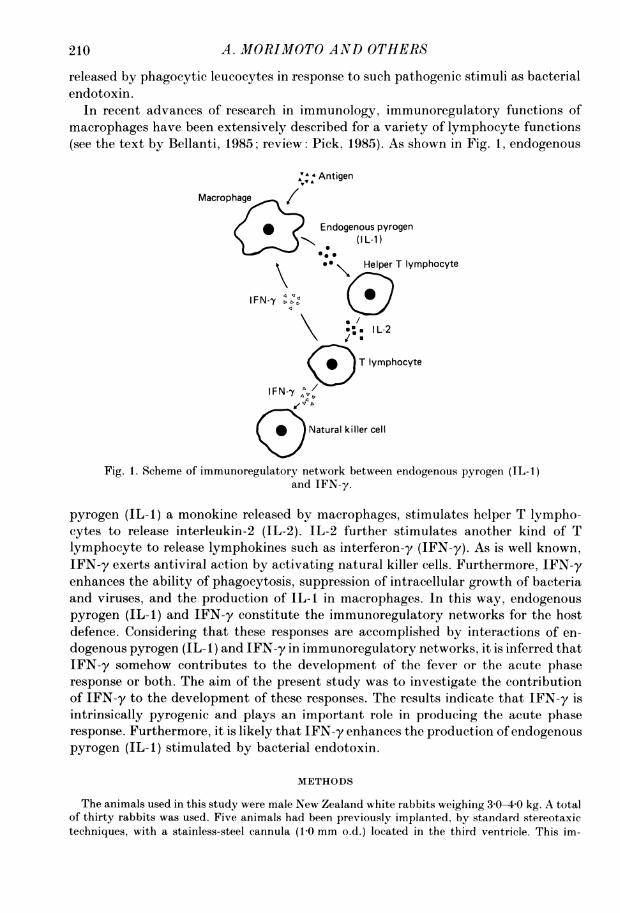
Fig. 1. Scheme of immunoregulatory network between endogenous pyrogen (IL-1)and IFN-y.
pyrogen (IL-1) a monokine released by macrophages, stimulates helper T lympho-cytes to release interleukin-2 (IL-2). IL-2 further stimulates another kind of Tlymphocyte to release lymphokines such as interferon-y (IFN-y). As is well known,IFN-y exerts antiviral action by activating natural killer cells. Furthermore, IFN-yenhances the ability of phagocytosis, suppression of intracellular growth of bacteriaand viruses, and the production of IL-1 in macrophages. In this way, endogenouspyrogen (IL-1) and IFN-y constitute the immunoregulatory networks for the hostdefence. Considering that these responses are accomplished by interactions of en-dogenous pyrogen (IL- 1) and IFN-y in immunoregulatory networks, it is inferred thatIFN-y somehow contributes to the development of the fever or the acute phaseresponse or both. The aim of the present study was to investigate the contributionof IFN-y to the development of these responses. The results indicate that IFN-y isintrinsically pyrogenic and plays an important role in producing the acute phaseresponse. Furthermore, it is likely that IFN-y enhances the production of endogenouspyrogen (IL-1) stimulated by bacterial endotoxin.
METHODS
The animals used in this study were male New Zealand white rabbits weighing 3 0-4 0 kg. A totalof thirty rabbits was used. Five animals had been previously implanted. by standard stereotaxictechniques, with a stainless-steel cannula (1 0 mm o.d.) located in the third ventricle. This im-
210

INTERFERON AND FEVER 211
plantation was done under general anaesthesia (sodium pentobarbitone, 20 mg/kg, I.v.). at least2 weeks before the start of the experiment. Interferon was injected directly through the cannulainto the third ventricle.Human recombinant IFN-y was supplied by the Kyowa Hakko Kogyo Co. Ltd., and had been
produced by recombinant strains of Escherichia coli. This IFN-y was highly purified (99 %O) and wasendotoxin-free, as restrictively confirmed by Limulus amoebocyte lysate test. The molecularweight was 17 100. For i.v. injection, one vial containing 2 x 106 antiviral units of human IFN-yand human serum albumin was used in the experiment, while human serum albumin dissolved insterile saline was used as the control. For intracerebroventricular (i.c.V.) injection, pure IFN-y,without human serum albumin, was dissolved in sterile saline at a concentration of 5 x 106 units/ml and was frozen at -20 °C, because the serum albumin had a strong pyrogenic action (Morimoto,Murakami, Nakamori & Watanabe, 1987b) when it was injected into the cerebral ventricle. Weused the pure IFN-y within 2 days after thawing since it became remarkably unstable after thattime.The endogenous pyrogen (IL- 1) was prepared from white blood cells of male rabbits (New Zealand
white strain). The white blood cells were stimulated by lipopolysaccharide of Salmonella typhosaendotoxin (Difco). The general procedures for preparing endogenous pyrogen (IL-1) have beendescribed in detail elsewhere (Morimoto, Watanabe, Ono, Sakata & Murakami, 1986). Partialpurification was achieved by ultrafiltration using two types of membranes (lOYM10 and 1OXM50,Amicon), which removed all substances of molecular weight outside the range 10000-50000. Thus1-0 ml of this partially purified endogenous pyrogen solution was derived from approximately1-5 x 107 white blood cells. We confirmed that an i.v. injection of heat-treated endogenous pyrogen(IL-1) (1 ml/kg), which had been inactivated by heating in a hot water bath of 60 °C for 40 min,did not cause fever in rabbits. This result demonstrated that the partially purified endogenouspyrogen (IL- 1) was not contaminated by endotoxin. The bacterial endotoxin used in this study wasthe lipopolysaccharide of Salmonella typhosa endotoxin (Difco; LPS), dissolved in sterile saline.Injection doses in each experimental group are described in the Results, including appropriatecontrol injections.On the day of the experiment animals were minimally restrained in conventional stocks at an
ambient temperature of 21 + 1 °C between 08-30 and 18-00. All had been well trained to adapt tothe stocks for 6 h every other day, from at least 10 days before the start of the experiment.Throughout the experiment the rectal temperature was measured every minute with a copper-constantan thermocouple. All injections of pyrogenic substances or saline control were performedat the time of 12-00. Intravenous injections were made into the marginal ear vein through a sterileneedle (25 gauge). Intraventricular injections were made through a stainless-steel needle (0-6 mm,o.d.) attached to a polyethylene tube, and the volume infused was always 20 ,ul. Intraventricularinjections were performed with a microsyringe pump (Infors AG. CH-4015) over a period of 5 min.For measuring the blood cell counts and the plasma concentration of iron, zinc, copper and
fibrinogen, about 5 ml of blood was withdrawn through the marginal ear vein. Blood samples weretaken three times: 1 h before, 8 and 24 h after injections of IFN-y or the control. Immediatelyafter collecting the blood, the numbers of white blood cells and red blood cells were measured withan automatic cell counter (Coulter, Model S plus II). In addition, by preparing Wright-stainedsmears of the blood, the differential leucocyte count was microscopically measured and the per-centage composition of each cell of the neutrophil, lymphocyte, monocyte, basophil and eosinophilwas calculated. The remaining blood was centrifuged at 2000 r.p.m. for 15 min at 4 'C. Only theplasma was collected in the test tube and this was stored at -20 'C until measurement of iron, zinc,copper and fibrinogen concentrations. The plasma concentration of iron was measured by anautoanalyser (Olympus Super-Multi ASA8000), and those of zinc and copper were measured by anatomic absorption spectrophotometer (Hitachi model 180-80). Fibrinogen concentration was mea-sured by the method of von Clauss (1975). Data were analysed for statistical significance byStudent's t test.
RESULTS
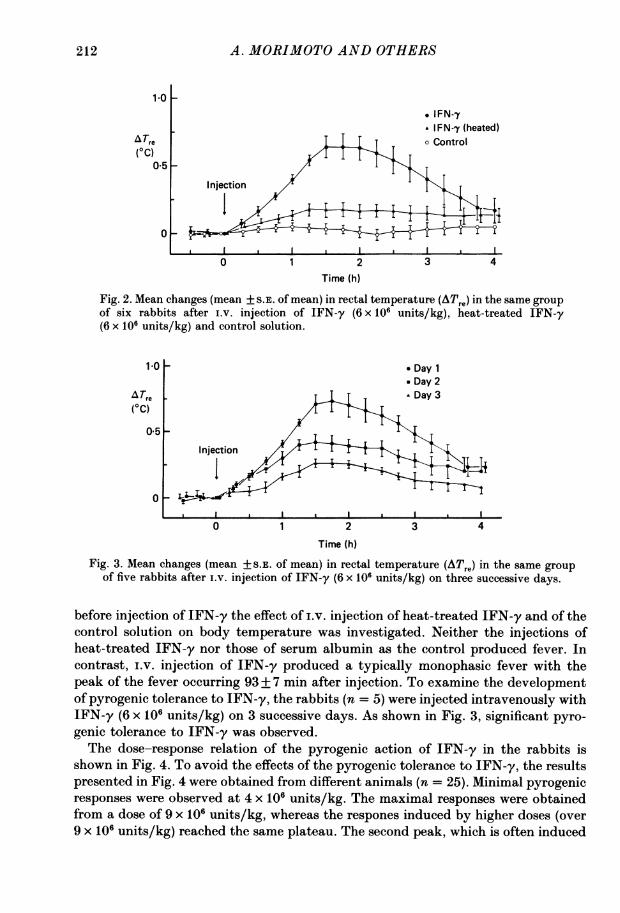
The effect of intravenous (i.v.) injection of IFN-y (6 x 106 units/kg) on rectaltemperature is shown in Fig. 2. Changes in the rectal temperature (ATre) are ex-pressed as a deviation from the base line recorded at the injection time. Several days
212 A. MORIMOTO AND OTHERS
1.0IFN-'yI F N-,y (heated)
ATre o Control(OC)
0*5
I njection /
0I . .0 1 2 3 4
Time (h)
Fig. 2. Mean changes (mean + S.E. of mean) in rectal temperature (ATre) in the same groupof six rabbits after i.v. injection of IFN-y (6 x 106 units/kg), heat-treated IFN-y(6 x 106 units/kg) and control solution.
1.0 * Day 1Day 2
05
0 1 2 3 4Time (h)
Fig. 3. Mean changes (mean +S.E. of mean) in rectal temperature (ATre) in the same groupof five rabbits after i.v. injection of IFN-y (6 x 106 units/kg) on three successive days.
before injection of IFN-y the effect of i.v. injection of heat-treated IFN-y and of thecontrol solution on body temperature was investigated. Neither the injections ofheat-treated IFN-y nor those of serum albumin as the control produced fever. Incontrast, i.v. injection of IFN-y produced a typically monophasic fever with thepeak of the fever occurring 93+ 7 min after injection. To examine the developmentof pyrogenic tolerance to IFN-y, the rabbits (n = 5) were injected intravenously withIFN-y (6 x 106 units/kg) on 3 successive days. As shown in Fig. 3, significant pyro-genic tolerance to IFN-y was observed.The dose-response relation of the pyrogenic action of IFN-y in the rabbits is
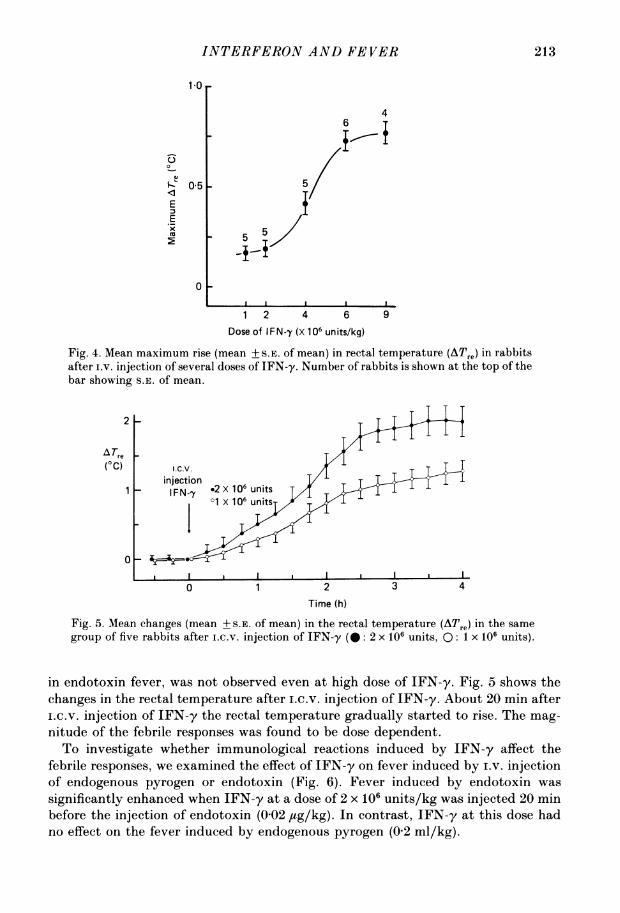
shown in Fig. 4. To avoid the effects of the pyrogenic tolerance to IFN-y, the resultspresented in Fig. 4 were obtained from different animals (n = 25). Minimal pyrogenicresponses were observed at 4 x 106 units/kg. The maximal responses were obtainedfrom a dose of 9 x 106 units/kg, whereas the respones induced by higher doses (over9 x 106 units/kg) reached the same plateau. The second peak, which is often induced
INTERFERON AND FEVER
1-0r
5
1 2 4 6
Dose of IFN-y (X 106 units/kg)
9
Fig. 4. Mean maximum rise (mean + S.E. of mean) in rectal temperature (ATre) in rabbitsafter i.v. injection of several doses of IFN-y. Number of rabbits is shown at the top of thebar showing S.E. of mean.
21-
ATre
(O C) I.C.V.
injection .2 X 106 unitsFN-y
01~~~x 106 units
I I I I I
0 2
Time (h)
3 4
Fig. 5. Mean changes (mean + S.E. of mean) in the rectal temperature (ATre) in the same
group of five rabbits after i.c.V. injection of IFN-y (@: 2 x 106 units, 0: 1 x 106 units).
in endotoxin fever, was not observed even at high dose of IFN-y. Fig. 5 shows thechanges in the rectal temperature after i.c.v. injection of IFN-y. About 20 min afteri.c.v. injection of IFN-y the rectal temperature gradually started to rise. The mag-
nitude of the febrile responses was found to be dose dependent.To investigate whether immunological reactions induced by IFN-y affect the
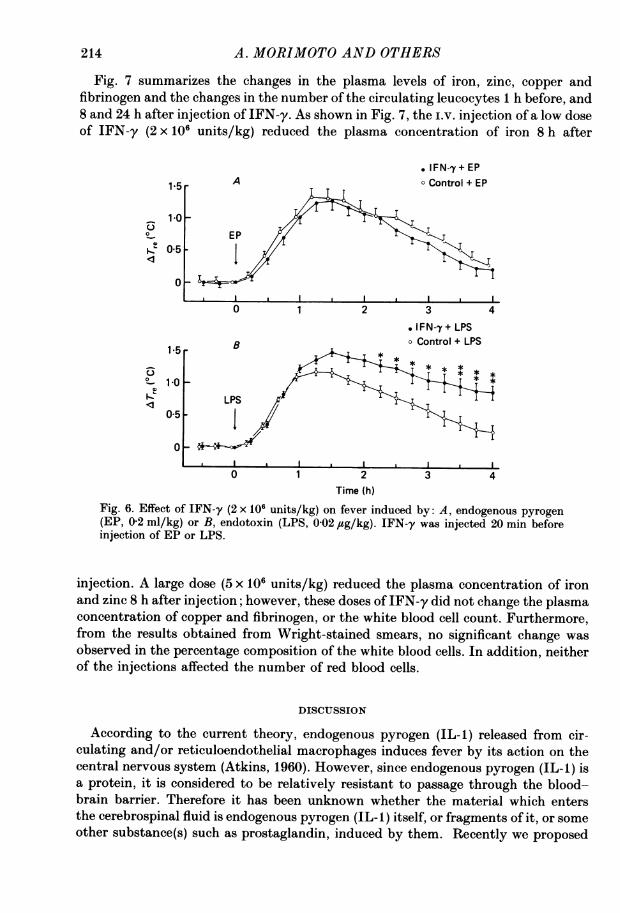
febrile responses, we examined the effect of IFN-y on fever induced by i.v. injectionof endogenous pyrogen or endotoxin (Fig. 6). Fever induced by endotoxin was
significantly enhanced when IFN-y at a dose of 2 x 106 units/kg was injected 20 minbefore the injection of endotoxin (0-02 ,tg/kg). In contrast, IFN-y at this dose hadno effect on the fever induced by endogenous pyrogen (0-2 ml/kg).
0*5pC1EEx(U
0
213
46
5T
1
214 A. MORIMOTO AND OTHERS
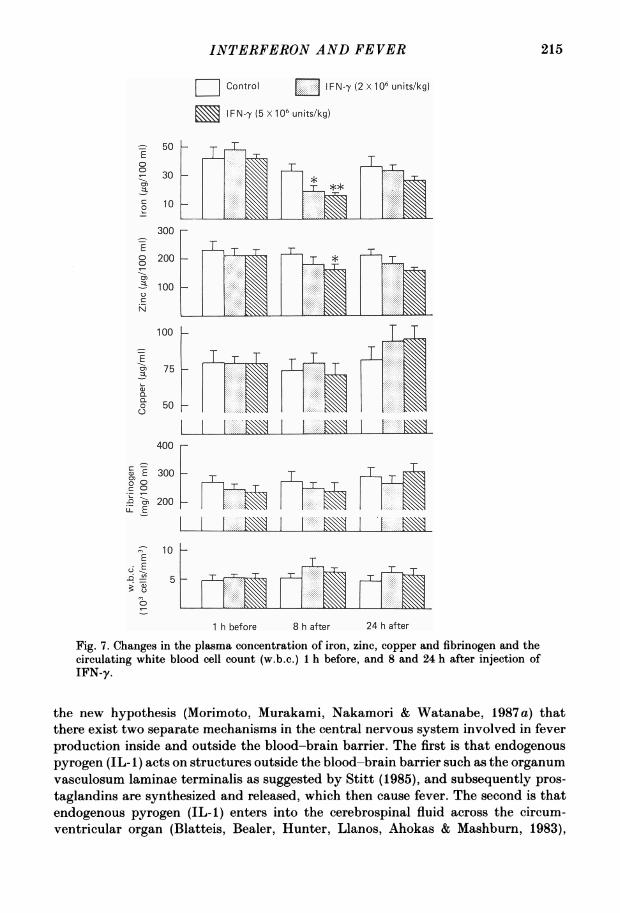
Fig. 7 summarizes the changes in the plasma levels of iron, zinc, copper andfibrinogen and the changes in the number of the circulating leucocytes 1 h before, and8 and 24 h after injection of IFN-y. As shown in Fig. 7, the i.v. injection of a low doseof IFN-y (2 x 106 units/kg) reduced the plasma concentration of iron 8 h after
. IFN-y+ EPo Control + EP
2 3. IFN- + LPSo Control + LPS
4
LPS
I a al
2Time (h)
3 4
Fig. 6. Effect of IFN-y (2 x 106 units/kg) on fever induced by: A, endogenous pyrogen(EP, 0-2 ml/kg) or B, endotoxin (LPS, 0-02 jug/kg). IFN-y was injected 20 min beforeinjection of EP or LPS.
injection. A large dose (5 x 106 units/kg) reduced the plasma concentration of ironand zinc 8 h after injection; however, these doses of IFN-y did not change the plasmaconcentration of copper and fibrinogen, or the white blood cell count. Furthermore,from the results obtained from Wright-stained smears, no significant change wasobserved in the percentage composition of the white blood cells. In addition, neitherof the injections affected the number of red blood cells.
DISCUSSION
According to the current theory, endogenous pyrogen (IL-1) released from cir-culating and/or reticuloendothelial macrophages induces fever by its action on thecentral nervous system (Atkins, 1960). However, since endogenous pyrogen (IL-1) isa protein, it is considered to be relatively resistant to passage through the blood-brain barrier. Therefore it has been unknown whether the material which entersthe cerebrospinal fluid is endogenous pyrogen (IL-1) itself, or fragments of it, or someother substance(s) such as prostaglandin, induced by them. Recently we proposed
1 5r A
1 01'ak~I1
0*5 -EP
0I I I
0
B15r
10FIio
k~I1
05 F0
0
1
1
INTERFERON AND FEVER 215
LI Control [II IFN- (2 x 106 units/kg)
m IFN- (5 x 106 units/kg)
50 ,-X1L
3003
ZLl 7 ....1
0 10 1200 _
- 100
1 1 1: 11V 11S
100
E) KCo 50
400
E300
200 rLr0r0ri
1 h before 8 h after 24 h after
Fig. 7. Changes in the plasma concentration of iron, zinc, copper and fibrinogen and thecirculating white blood cell count (w.b.c.) 1 h before, and 8 and 24 h after injection ofIFN-y.
the new hypothesis (Morimoto, Murakami, Nakamori & Watanabe, 1987a) thatthere exist two separate mechanisms in the central nervous system involved in feverproduction inside and outside the blood-brain barrier. The first is that endogenouspyrogen (IL-i1) acts on structures outside the blood-brain barrier such as the organumvasculosum laminae terminalis as suggested by Stitt (1985), and subsequently pros-taglandins are synthesized and released, which then cause fever. The second is thatendogenous pyrogen (IL-i) enters into the cerebrospinal fluid across the circum-ventricular organ (Blatteis, Bealer, Hunter, Llanos, Ahokas & Mashburn, 1983),

A. MORIMOTO AND OTHERS
where the blood-brain barrier is absent, and acts on structures within the blood-brain barrier to release prostaglandins which then cause fever. The present resultsshow that fever was produced by i.v. and i.c.v. injection of IFN-y. Furthermore, thepatterns of febrile responses induced by i.v. and i.c.v. injections of IFN-y respect-ively resembled those induced by injections of endogenous pyrogen (IL- 1) (Morimotoet al. 1986). Considering that the molecular weight of IFN-y is almost the same asthat of endogenous pyrogen (IL-1) (Dinarello, 1984), it is suggested that IFN-yinduces the fever by both peripheral and central actions, as does endogenous pyrogen(IL-1). Fever in rabbits induced by a human recombinant interferon-a (IFN-a) wasrecently reported by Dinarello, Bernheim, Duff, Le, Nagabhushan, Hamilton &Coceani (1984). The characteristics of pyrogenic action found by them following i.v.injection of IFN-a were almost the same as those which are obtained with IFN-y;however, the pyrogenicity per unit of IFN-a seems to be slightly higher than that ofIFN-y. Dinarello et al. (1984) reported that i.c.v. injection of IFN-a produced abiphasic response consisting of an initial hypothermia followed by hyperthermia incats. However the control solution containing albumin and glycine also producedhypothermia. Therefore, it seems that in their results, hypothermia induced by i.c.v.injection of IFN-a solution was caused by glycine (Glyn & Lipton, 1980). As for theacute phase response the present results show that the i.v. injection of a large doseof IFN-y induces some of the acute phase response induced by endogenous pyrogen(IL-1) (Kampschmidt, 1980; Gordon & Koj, 1985; Morimoto, Murakami, Myogin,Takada, Teshirogi & Watanabe, 1987 a). It is still unknown whether or not otherkinds of IFN induce the acute phase response. Recently, we also suggested thatendogenous pyrogen (IL-1) induces the acute phase response, in addition to fever, byits action on both the peripheral target organ and the central nervous system(Morimoto et al. 1987 c). Therefore it is inferred that IFN-y induced fever and theacute phase response by both peripheral and central actions, although the effect ofthe central action of IFN-y on the acute phase response is still unknown. Based onthe present results, IFN-y is suggested to be one of the endogenous mediator proteinswhich produces the fever and the acute phase response.
Table 1 summarizes some properties of endogenous pyrogen (IL-1), IFN-a andIFN-y, as established in the present experiments and also as reported in the previousliterature. As noted in Table 1, the cells producing endogenous pyrogen (IL-1) andIFN are quite different. This result indicates the possibility that fever is processednot only by macrophages but also by other kinds of white blood cells. Moreover, asshown in Fig. 1, because of the close interactions of the immunoregulatory networksbetween macrophages and lymphocytes mediated by interleukins and interferons, itis possible that endogenous pyrogen (IL-1) and IFN-y may contribute mutually tothe induction of fever and the acute phase response. In fact, the present results showthat IFN-y enhanced the febrile response induced by endotoxin but had no effect onthat induced by endogenous pyrogen (IL-1). This phenomenon may suggest thepossibility that IFN-y enhances synthesis and release of endogenous pyrogen (IL-1)when macrophages are exposed to endotoxin. This suggestion is supported by therecent experiments in vitro showing that IFN-y enhances secretion of endogenouspyrogen (IL-1) by human monocytes (Arenzana-Seisdedos & Virelizier, 1983;Newton, 1985).
216

INTERFERON AND FEVER
Other characteristics ofIFN, such as molecular weight and heat lability, are similarto those of endogenous pyrogen (IL-1). However, one significant difference is thatpyrogenic tolerance can exist with IFN but not with endogenous pyrogen (IL-1). In
TABLE 1. Summary of biological characteristics of endogenous pyrogen and interferonEndogenous Interferonpyrogen(IL-1) IFN-ac IFN-y
Chemical nature Protein Protein Protein(molecular weight) (15000-45000) (15000-50000) (15000-50000)
Origin cells Macrophage Lymphocyte LymphocyteThermal stability Heat-labile Heat-labile Heat-labilePyrogenic response tosuccessive daily injections Constant Decrease Decrease
the results from private reports by the Kyowa Hakko Kogyo Co. Ltd. (unpublisheddata of phase I), human IFN-y (2 x 106 units/person) produced remarkable fever(>1 C) in humans. Pyrogenic tolerance was also observed when IFN-y was used totreat viral infections or tumours. The pyrogenicity per unit of IFN-y in humans issignificantly greater than that in rabbits. Thus, it is at present unknown whetherthere exist structural or functional differences in the action of IFN between differentspecies. Moreover, the reason for an induction of tolerance to IFN still remains to beelucidated.
We are grateful to the Kyowa Hakko Kogyo Co. Ltd for supplies of human recombinantIFN-y.
REFERENCES
ARENZANA-SEISDEDOS, F. & VIRELIZIER, J. L. (1983). Interferons as macrophage-activatingfactors. II. Enhanced secretion of interleukin 1 by lipopolysaccharide-stimulated human mono-cytes. European Journal of Immunology 13, 437-440.
ATKINS, E. (1960). Pathogenesis of fever. Physiological Reviews 40, 580-646.BELLANTI, J. A. (1985). Immunology III. U.S.A.: Saunders.BLATTEIS, C. M., BEALER, S. L., HUNTER, W. S., LLANOS, Q. J., AHOKAS, R. A. & MASHBURN JR,
T. A. (1983). Suppression of fever after lesions of the anteroventral third ventricle in guinea pigs.Brain Research Bulletin 11, 519-526.
DINARELLO, C. A. (1984). Interleukin-1. Review of Infectious Diseases 6, 51-95.DINARELLO, C. A., BERNHEIM, H. A., DUFF, G. W., LE, H. V., NAGABHUSHAN, T. L., HAMILTON,
N. C. & COCEANI, F. (1984). Mechanisms of fever induced by recombinant human interferon.Journal of Clinical Investigation 74, 906-913.
GLYN, J. R. & LIPTON, J. M. (1980). Comparison of the effects of central administration of serineand glycine on body temperature of rabbit. Brain Research Bulletin 5, 661-665.
GORDON, A. H. & KoJ, A. (1985). The acute phase-response to injury and infection. ResearchMonographs in Cell and Tissue Physiology, vol. 10. New York: Elsevier.
KAMPSCHMIDT, R. F. (1980). Metabolic alterations elicited by endogenous pyrogen. In Fever, ed.Lipton, J. M., pp. 49-56. New York: Raven.
KASTING, N. W. & MARTIN, J. B. (1982). Altered release of growth hormone and thyrotropininduced by endotoxin in the rat. American Journal of Physiology 243, E332-337.
KASTING, N. W., MAZUREK, M. F. & MARTIN, J. B. (1985). Endotoxin increases vasopressin releaseindependently of known physiological stimuli. American Journal of Physiology 248, E420-424.
217

A. MORIMOTO AND OTHERS
KLUGER, M. J., RINGLER, D. H. & ANVER, M. R. (1975). Fever and survival. Science 188, 166-168.
KLUGER, M. J. & ROTHENBERG, B. A. (1979). Fever and reduced iron: their interaction as a hostdefense response to bacterial infection. Science 203, 374-376.
MOBERG, G. P. (1971). Site of action of endotoxins on hypothalamic-pituitary-adrenal axis.American Journal of Physiology 220, 397-400.
MORIMOTO, A., MURAKAMI, N., NAKAMORI, T. & WATANABE, T. (1987a). Evidence for separatemechanisms of induction of biphasic fever inside and outside the blood-brain barrier in rabbits.Journal of Physiology 383, 629-637.
MORIMOTO, A., MURAKAMI, N., NAKAMORI, T. & WATANABE, T. (1987b). Fever induced in rabbitsby intraventricular injection of rabbit and human albumin. Journal of Physiology 390,137-144
MORIMOTO, A., MURAKAMI, N., MYOGIN, T., TAKADA, M., TESHIROGI, S. & WATANABE, T. (1987c).Separate mechanisms of induction of the metabolic changes inside and outside the blood-brainbarrier in febrile rabbits. Journal of Physiology (in the Press).
MORIMOTO, A., WATANABE, T., ONO, T., SAKATA, Y. & MURAKAMI, N. (1986). Rat endogenouspyrogen and fever. American Journal of Physiology 250, R776-782.
NEWTON, R. C. (1985). Effect of interferon on the induction of human monocyte secretion ofinterleukin- 1 activity. Immunology 56, 441-449.
PICK, E. (1985). Lymphokines, a Forum for Immunoregulatory Cell Products, vol. 11. New York:Academic.
SHIDO, 0. & NAGASAKA, T. (1986). Cardiovascular and thermal responses to intravenous endotoxinin guinea pigs. Japanese Journal of Physiology 36, 543-554.
STITT, J. T. (1985). Evidence for the involvement of the organum vasculosum laminae terminalisin the febrile response of rabbits and rats. Journal of Physiology 368, 501-511.
VON CLAUSS, A. (1957). Gerinnungsphysiologische Schnellmethode zur Bestimmung des Fibrino-gens. Acuta haematologie 17, 237-246.
218





























