Embed Size (px)
Citation preview

ENZIMI KOJI
UČESTVUJU U
"PLANT
GROWTH
PROMOTING"
POENCIJALU
BTAKTERIJA

ZAŠTO JE OVO VAŽNO➢Klimatske promene
➢Porast broja ljudi na planeti
➢Veći zahtevi za poizvodnjom hrane, hrane za životinje, goriva posebno biodizela
➢Sve to na zemljištu koje ima povećan sadržaj metala (i teških metala), soli, smanjenu koncentraciju vode
➢Interdisciplinarni pristup je jedini koji može da da odgovore
➢ Izučavanje “Plant growth promoting bacteria – PGPB”
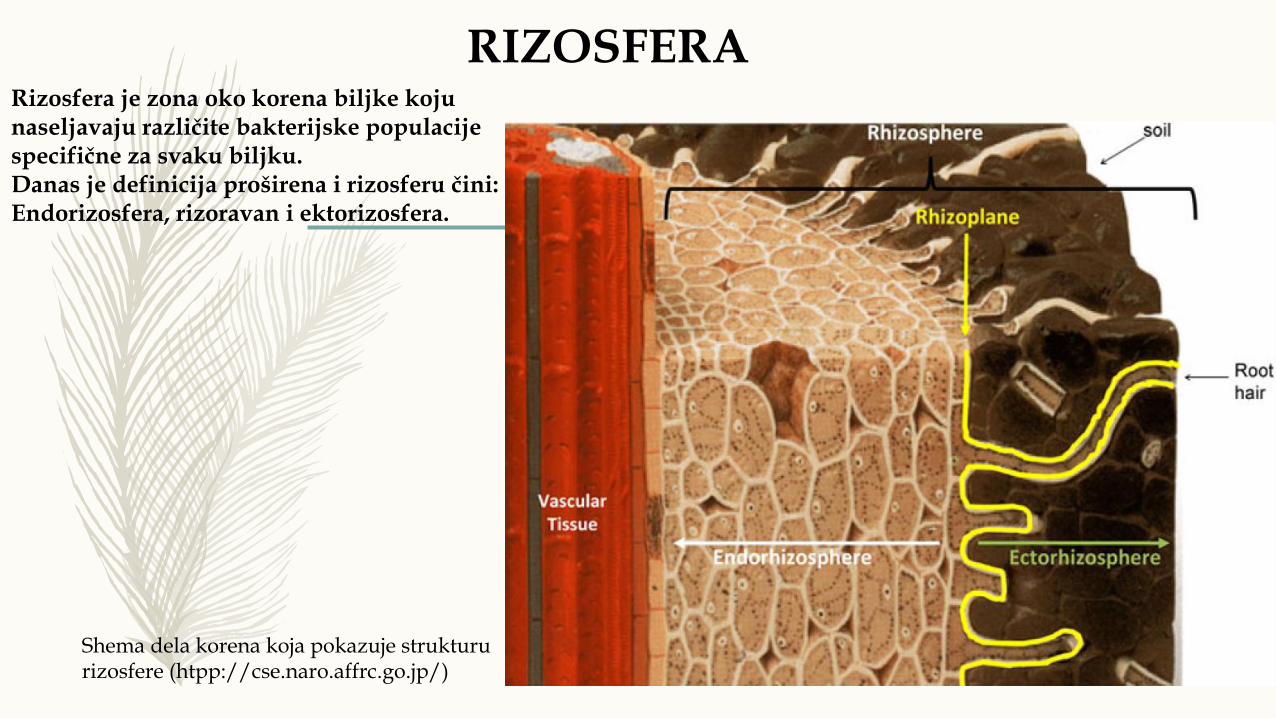
Shema dela korena koja pokazuje struktururizosfere (htpp://cse.naro.affrc.go.jp/)
Rizosfera je zona oko korena biljke kojunaseljavaju različite bakterijske populacijespecifične za svaku biljku.Danas je definicija proširena i rizosferu čini:Endorizosfera, rizoravan i ektorizosfera.
RIZOSFERA
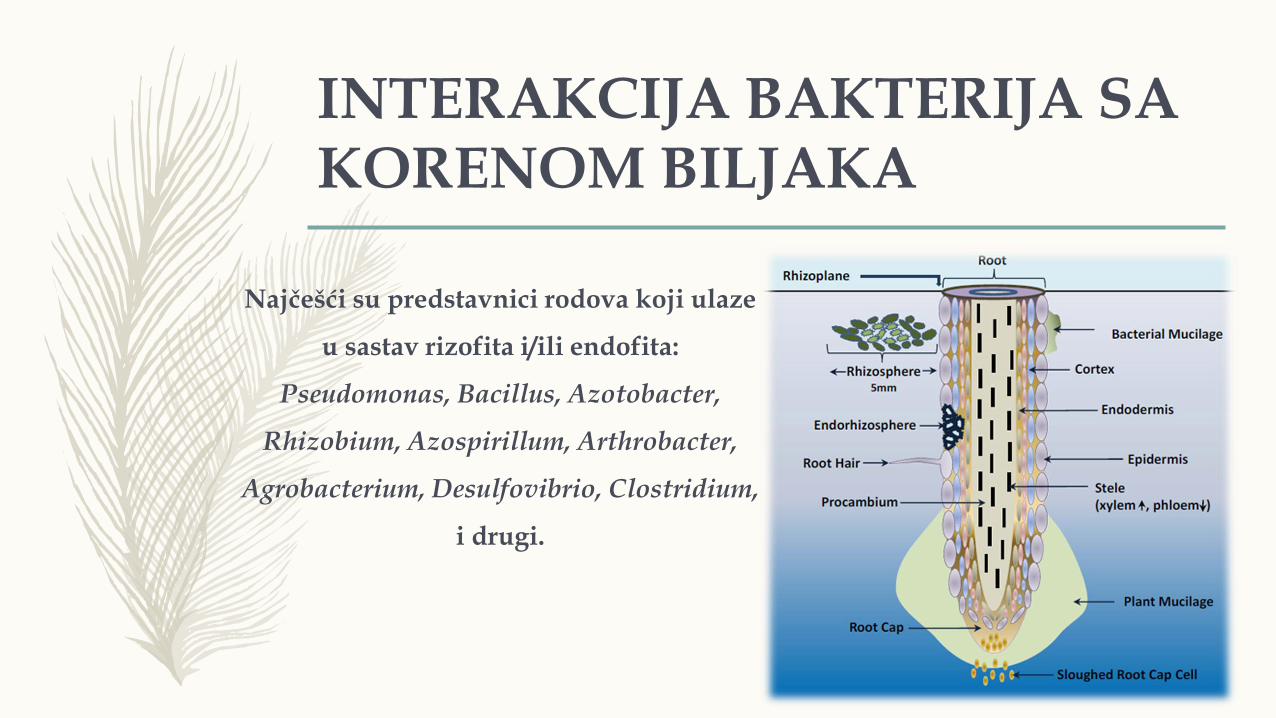
INTERAKCIJA BAKTERIJA SA KORENOM BILJAKA
Najčešći su predstavnici rodova koji ulaze
u sastav rizofita i/ili endofita:
Pseudomonas, Bacillus, Azotobacter,
Rhizobium, Azospirillum, Arthrobacter,
Agrobacterium, Desulfovibrio, Clostridium,
i drugi.
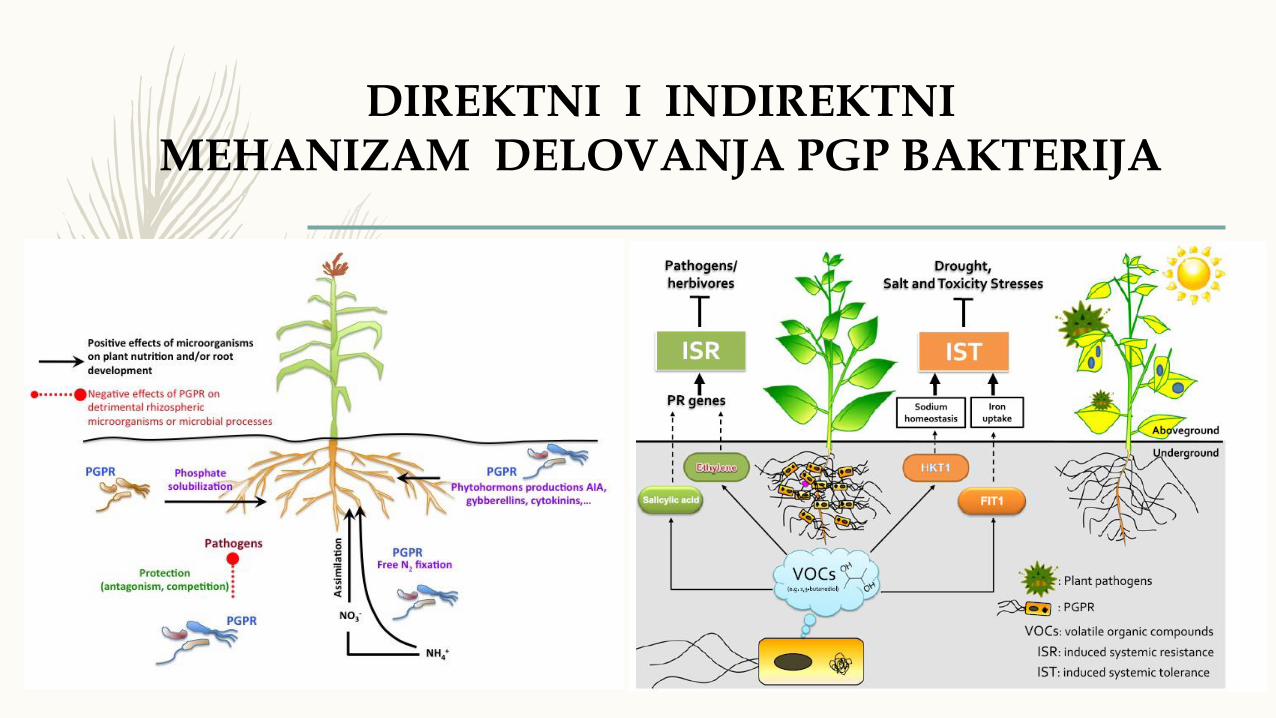
DIREKTNI I INDIREKTNIMEHANIZAM DELOVANJA PGP BAKTERIJA
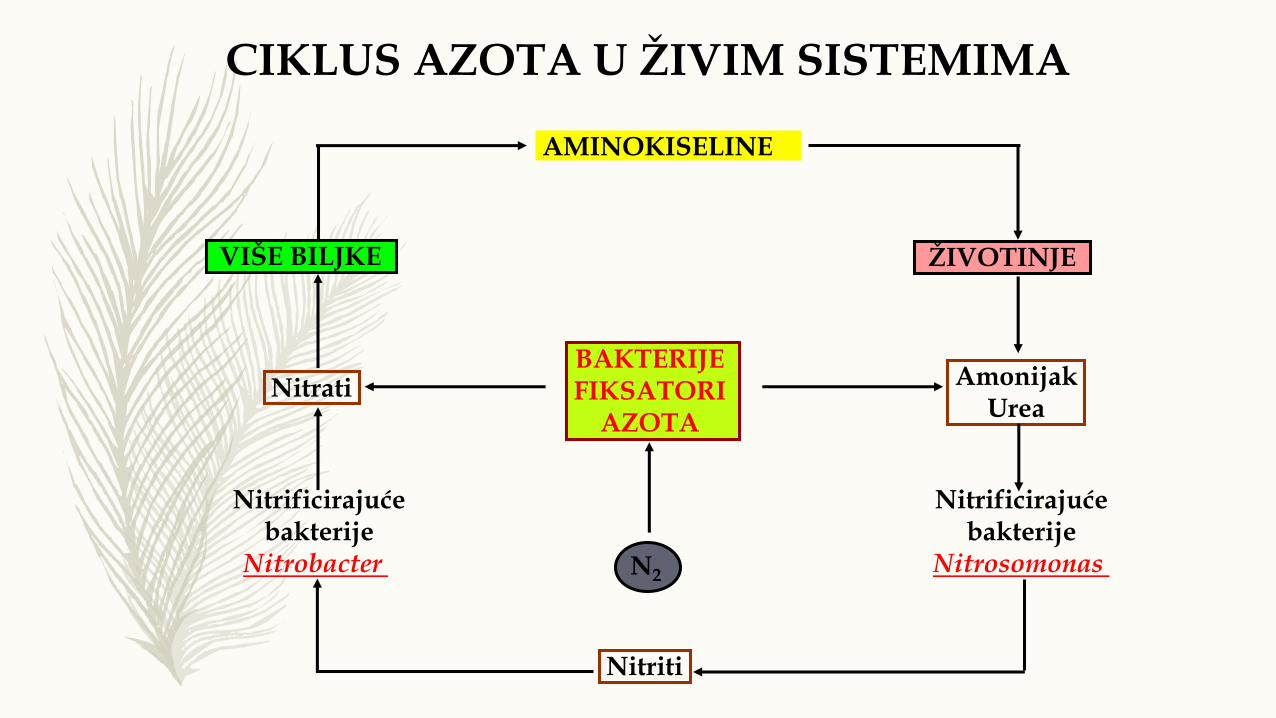
VIŠE BILJKE ŽIVOTINJE
BAKTERIJE FIKSATORI
AZOTA
N2
Nitrati
AMINOKISELINE
Amonijak Urea
Nitrificirajuće bakterije
Nitrobacter
Nitrificirajućebakterije
Nitrosomonas
Nitriti
CIKLUS AZOTA U ŽIVIM SISTEMIMA

Azotofiksacija
» Samo u prokariotima (bakterije i arhee)
» Proces osetljiv na kiseonik
» Slobodnoživeći i simbionti
» Tri tipa simbioze:
Rhizobium Frankia Nostoc

Azotofiksacija
» DIAZOTROFI je termin za sve bakterije i arhee koje imaju sposobnostfiksacije azota (fototrofi, hemoorganotrofi i hemolitotrofi). Diazotrofi mogubiti slobodnoživeći i simbionti. Diazotrofi simbionti su bakterije (familijaRhizobiaceae, rodovi Rhizobium, Bradyrhizobium i Azorhizobium),aktinomicete (rod Frankia) i cijanobakterije (rod Nostoc) koje žive u simbiozisa višim biljkama.
Rhizobium Frankia Nostoc
Faze u azotofiksaciji
1. •Signal i prepoznavanje
2. •Formiranje kvržice
3. •Redukcija azota

1.•Signal i prepoznavanje
» Razmena signala i prepoznavanje simbiotskih partnera
» Deformacija i uvijanje korenskih dlačica
» Ekxpreija nod gena i sinteza Nod faktora (Nodulini)
» Mogući uzroci: depolarizacija plazmaleme, promene u fluksu Ca2+,
promene protonskog efluksa, rearanžmani aktinskih filamenata
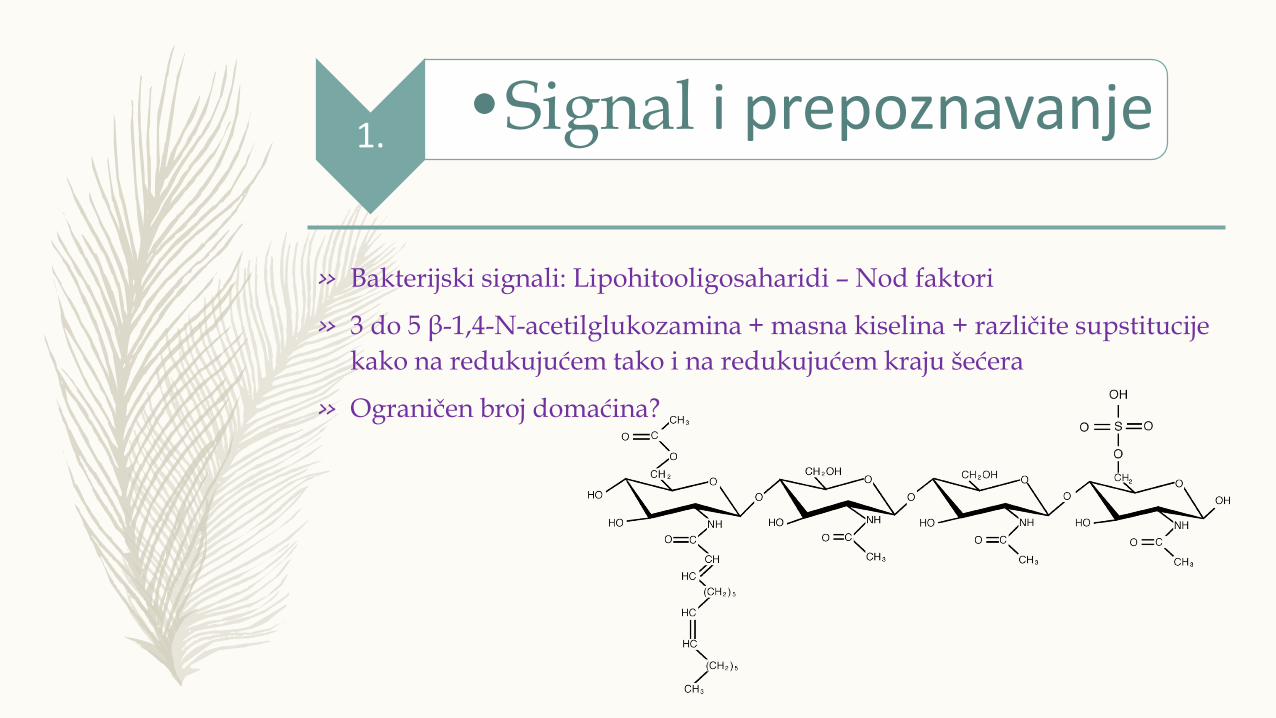
» Bakterijski signali: Lipohitooligosaharidi – Nod faktori
» 3 do 5 β-1,4-N-acetilglukozamina + masna kiselina + različite supstitucije
kako na redukujućem tako i na redukujućem kraju šećera
» Ograničen broj domaćina?
1. •Signal i prepoznavanje
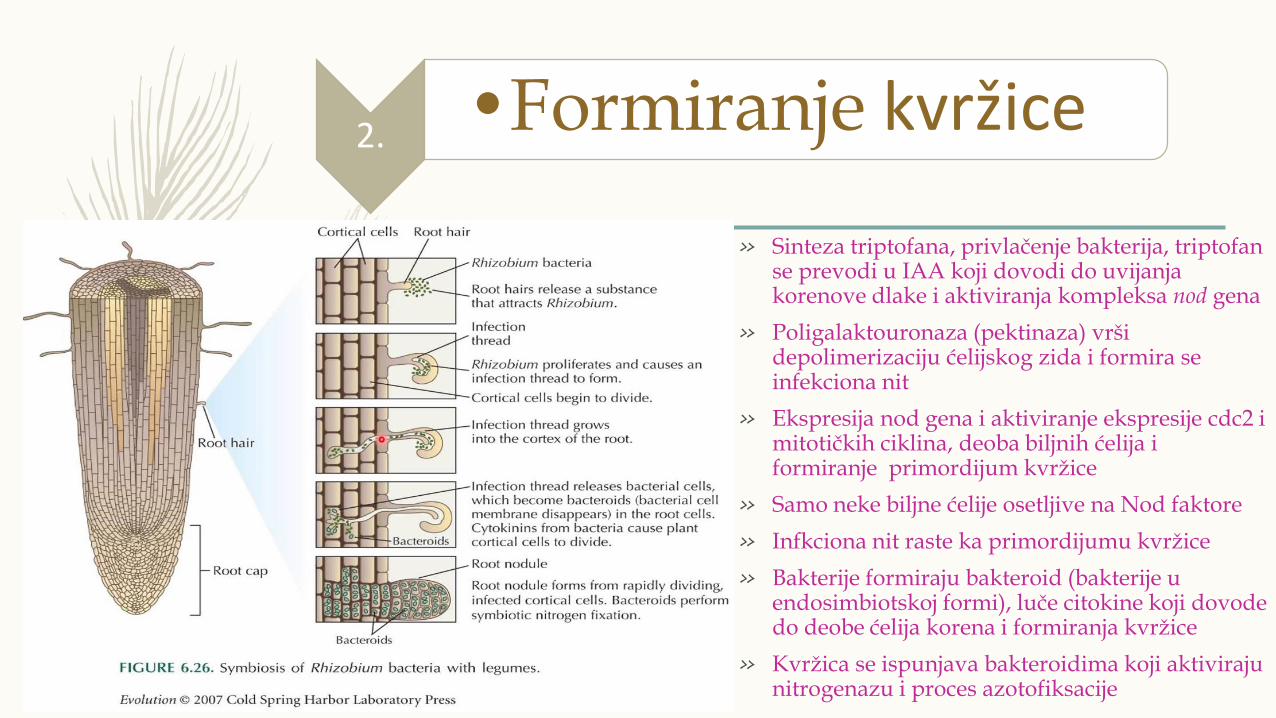
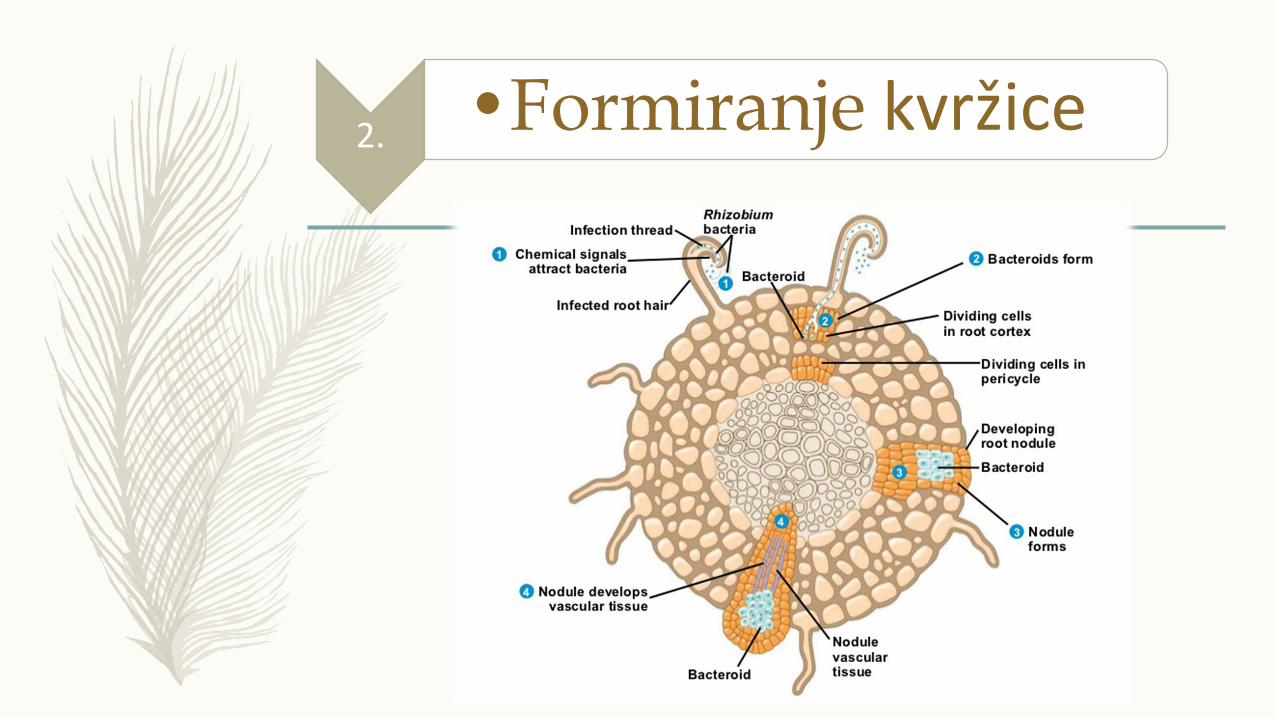
» Sinteza triptofana, privlačenje bakterija, triptofan se prevodi u IAA koji dovodi do uvijanja korenove dlake i aktiviranja kompleksa nod gena
» Poligalaktouronaza (pektinaza) vrši depolimerizaciju ćelijskog zida i formira se infekciona nit
» Ekspresija nod gena i aktiviranje ekspresije cdc2 i mitotičkih ciklina, deoba biljnih ćelija i formiranje primordijum kvržice
» Samo neke biljne ćelije osetljive na Nod faktore
» Infkciona nit raste ka primordijumu kvržice
» Bakterije formiraju bakteroid (bakterije u endosimbiotskoj formi), luče citokine koji dovode do deobe ćelija korena i formiranja kvržice
» Kvržica se ispunjava bakteroidima koji aktiviraju nitrogenazu i proces azotofiksacije
2. •Formiranje kvržice
2. •Formiranje kvržice
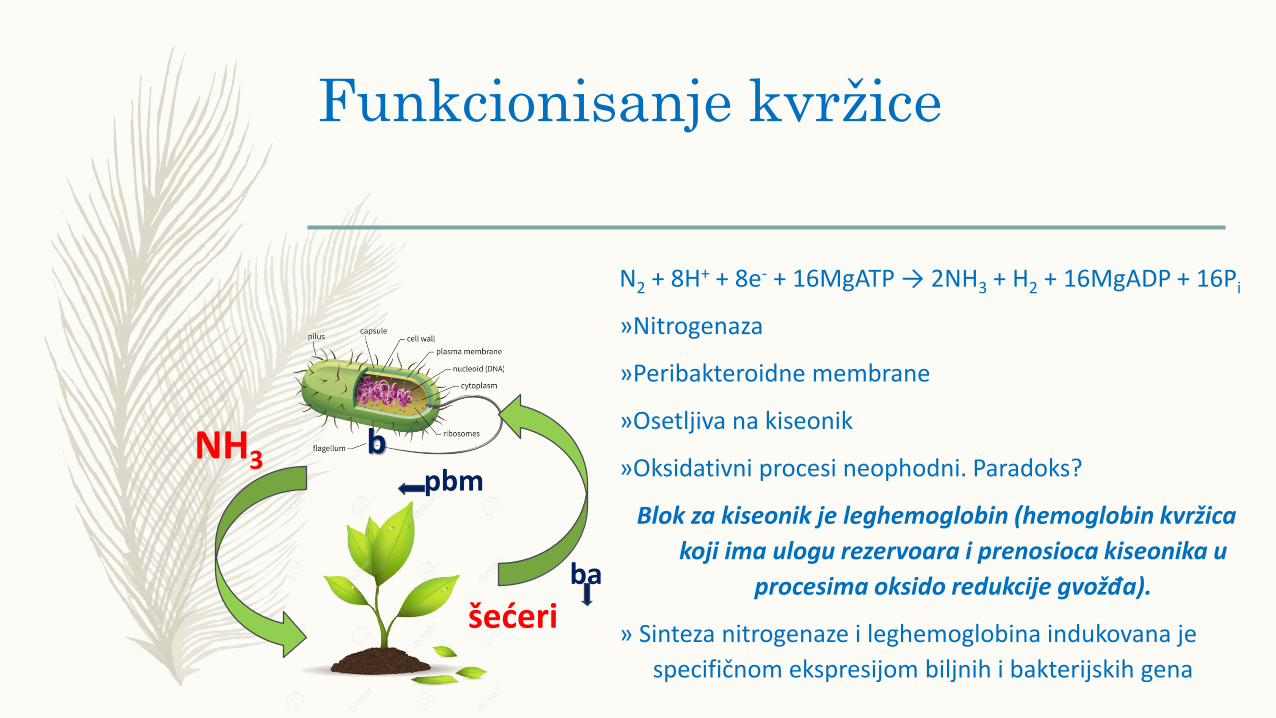
Funkcionisanje kvržice
N2 + 8H+ + 8e- + 16MgATP → 2NH3 + H2 + 16MgADP + 16Pi
»Nitrogenaza
»Peribakteroidne membrane
»Osetljiva na kiseonik
»Oksidativni procesi neophodni. Paradoks?
Blok za kiseonik je leghemoglobin (hemoglobin kvržica
koji ima ulogu rezervoara i prenosioca kiseonika u
procesima oksido redukcije gvožđa).
» Sinteza nitrogenaze i leghemoglobina indukovana je
specifičnom ekspresijom biljnih i bakterijskih gena
NH3
šećeri
bpbm
ba


Nitrogenazni kompleks
Fe-protein (dinitrogenazna reduktaza)
» nifH
» Homodimer
» [4Fe-4S] klaster
» ATP vezujuća mesta na svakoj
subjedinici
MoFe-protein (dinitrogenaza)
» nifD i nifK
» 2β2-tetramer
» P-klaster [8Fe-7S]
» M-klaster [Mo-7Fe-9S-X-
homocitrat], X = C, N, O
»3 visoko homologne klase – Mo-, V- i Fe-nitrogenaza, najbolje proučena Mo nitrogenaza Azotobacter vinelandii
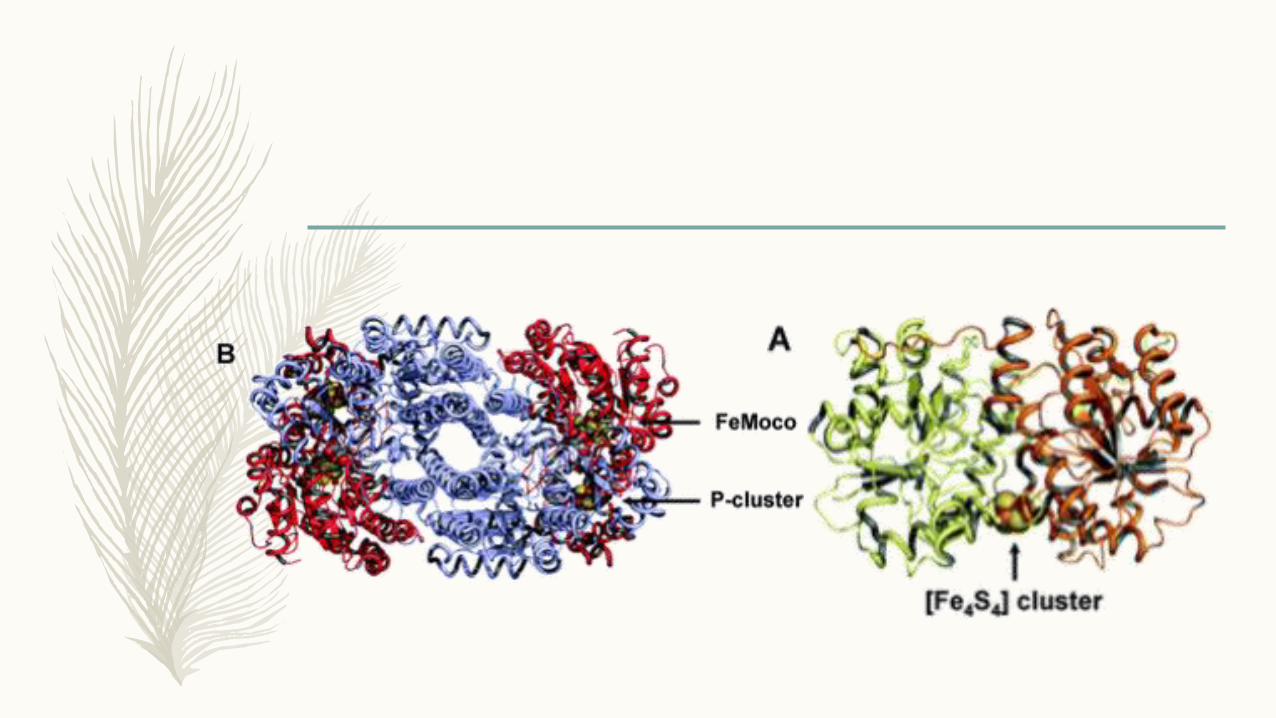
Nitrogenazni kompleks
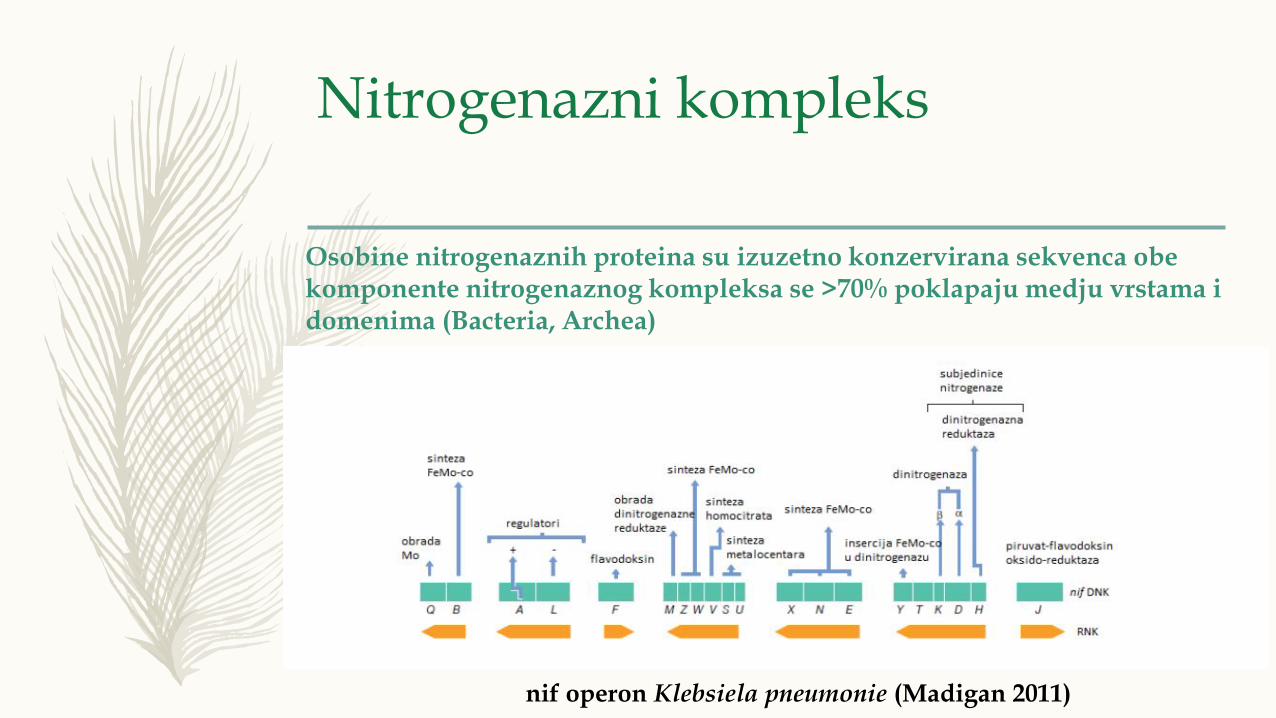
nif operon Klebsiela pneumonie (Madigan 2011)
Osobine nitrogenaznih proteina su izuzetno konzervirana sekvenca obekomponente nitrogenaznog kompleksa se >70% poklapaju medju vrstama idomenima (Bacteria, Archea)
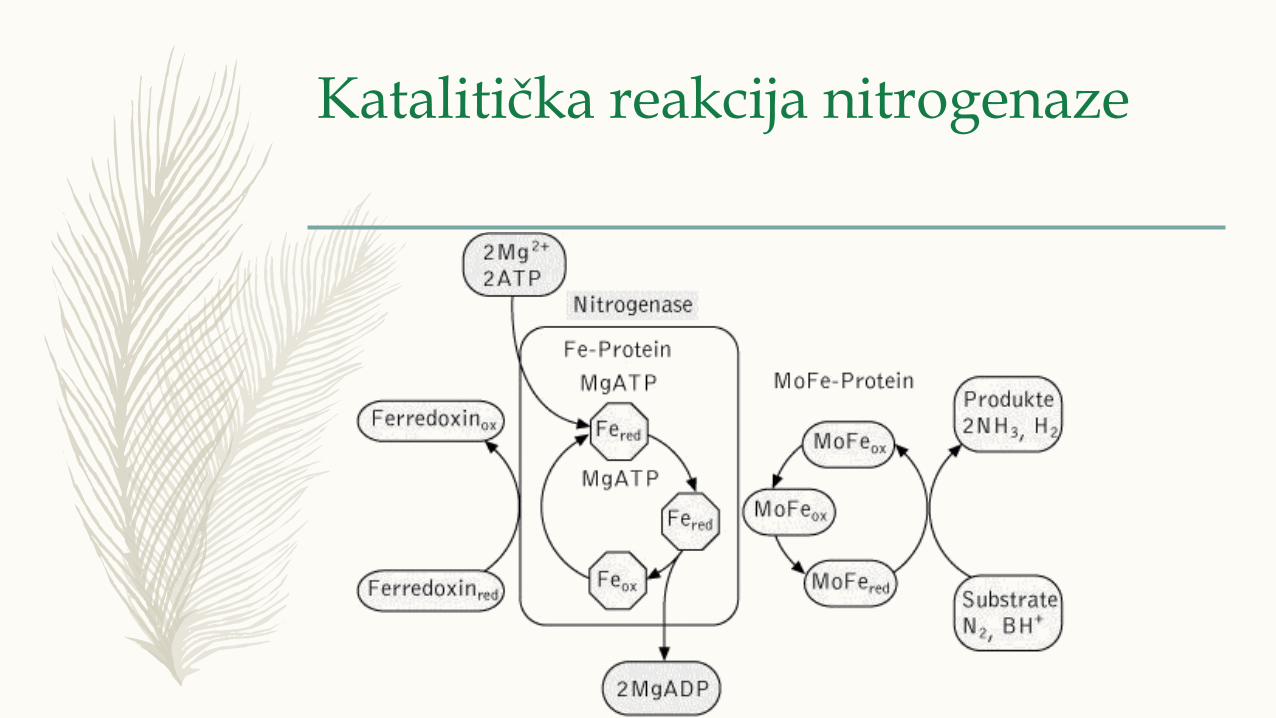
Katalitička reakcija nitrogenaze
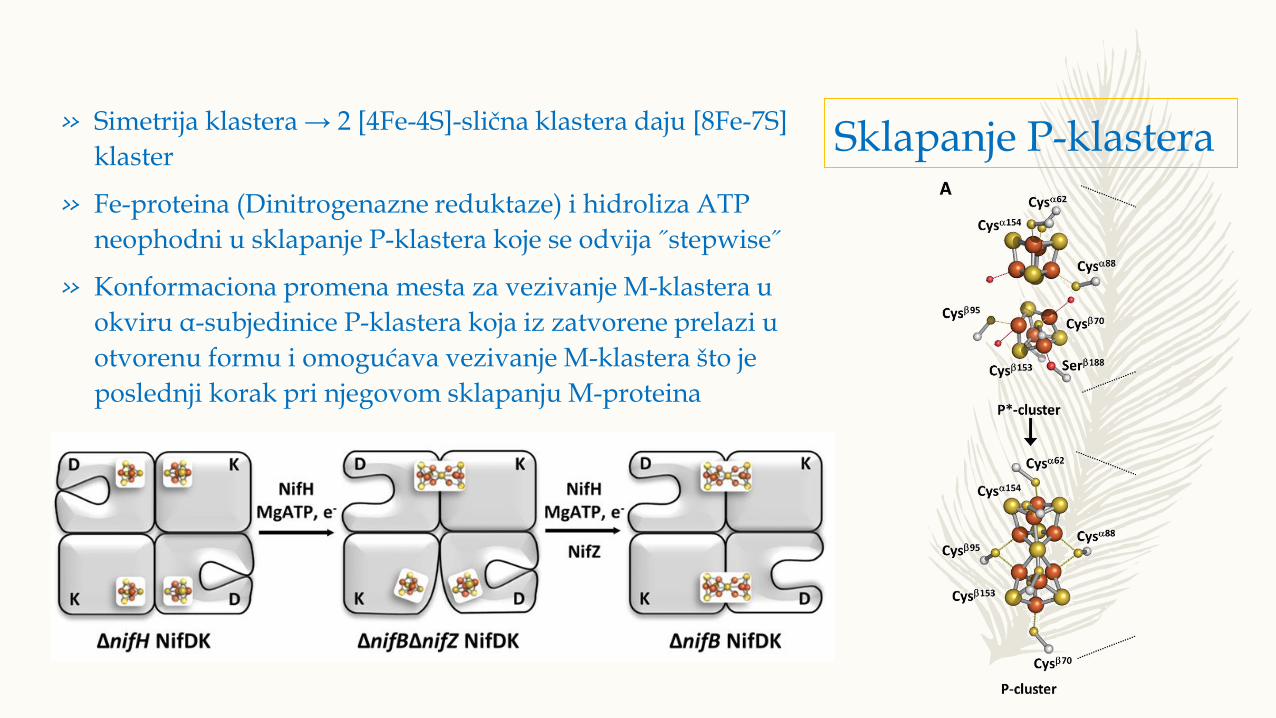
Sklapanje P-klastera» Simetrija klastera → 2 [4Fe-4S]-slična klastera daju [8Fe-7S]
klaster
» Fe-proteina (Dinitrogenazne reduktaze) i hidroliza ATP
neophodni u sklapanje P-klastera koje se odvija ˝stepwise˝
» Konformaciona promena mesta za vezivanje M-klastera u
okviru α-subjedinice P-klastera koja iz zatvorene prelazi u
otvorenu formu i omogućava vezivanje M-klastera što je
poslednji korak pri njegovom sklapanju M-proteina
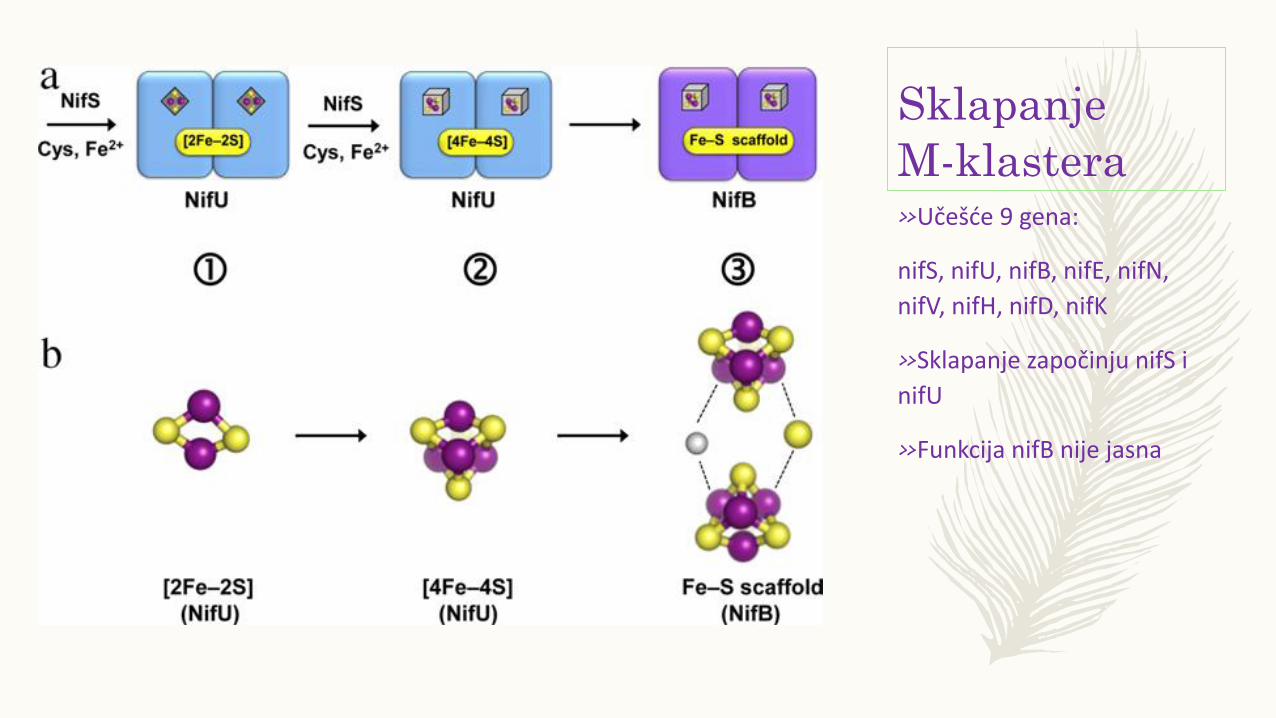
Sklapanje
M-klastera»Učešće 9 gena:
nifS, nifU, nifB, nifE, nifN,
nifV, nifH, nifD, nifK
»Sklapanje započinju nifS i
nifU
»Funkcija nifB nije jasna
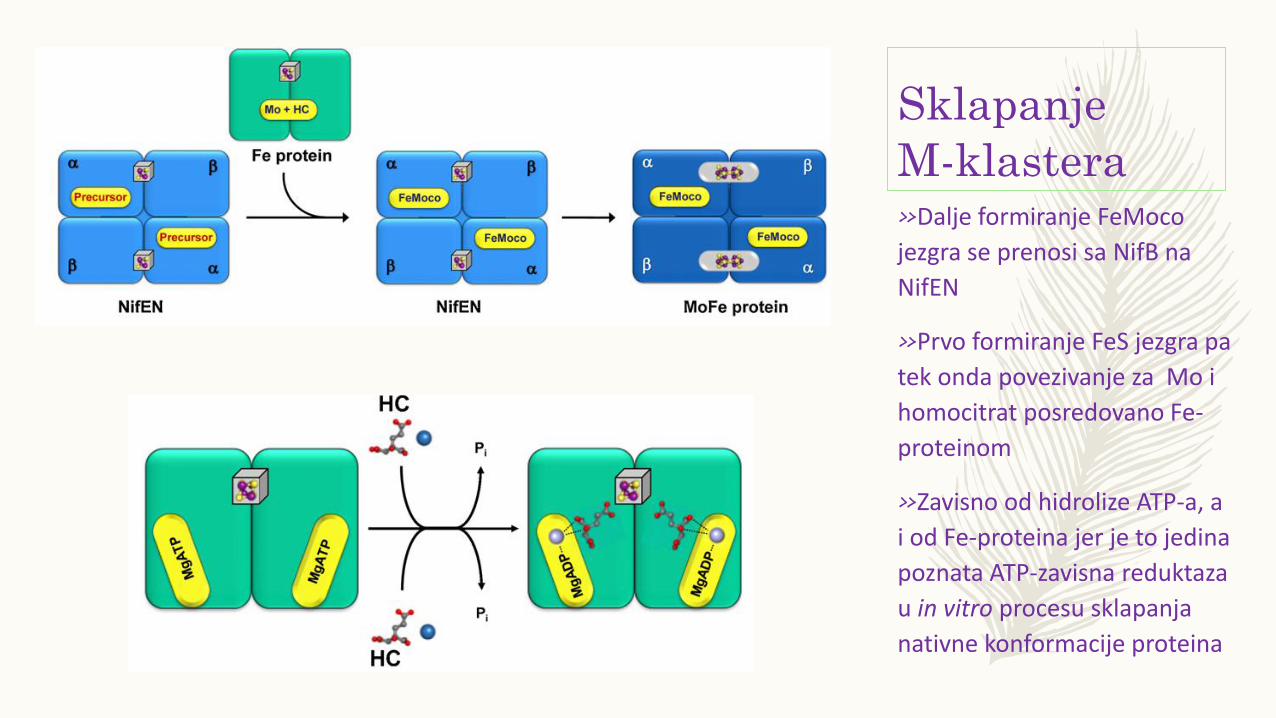
Sklapanje
M-klastera»Dalje formiranje FeMoco
jezgra se prenosi sa NifB na
NifEN
»Prvo formiranje FeS jezgra pa
tek onda povezivanje za Mo i
homocitrat posredovano Fe-
proteinom
»Zavisno od hidrolize ATP-a, a
i od Fe-proteina jer je to jedina
poznata ATP-zavisna reduktaza
u in vitro procesu sklapanja
nativne konformacije proteina
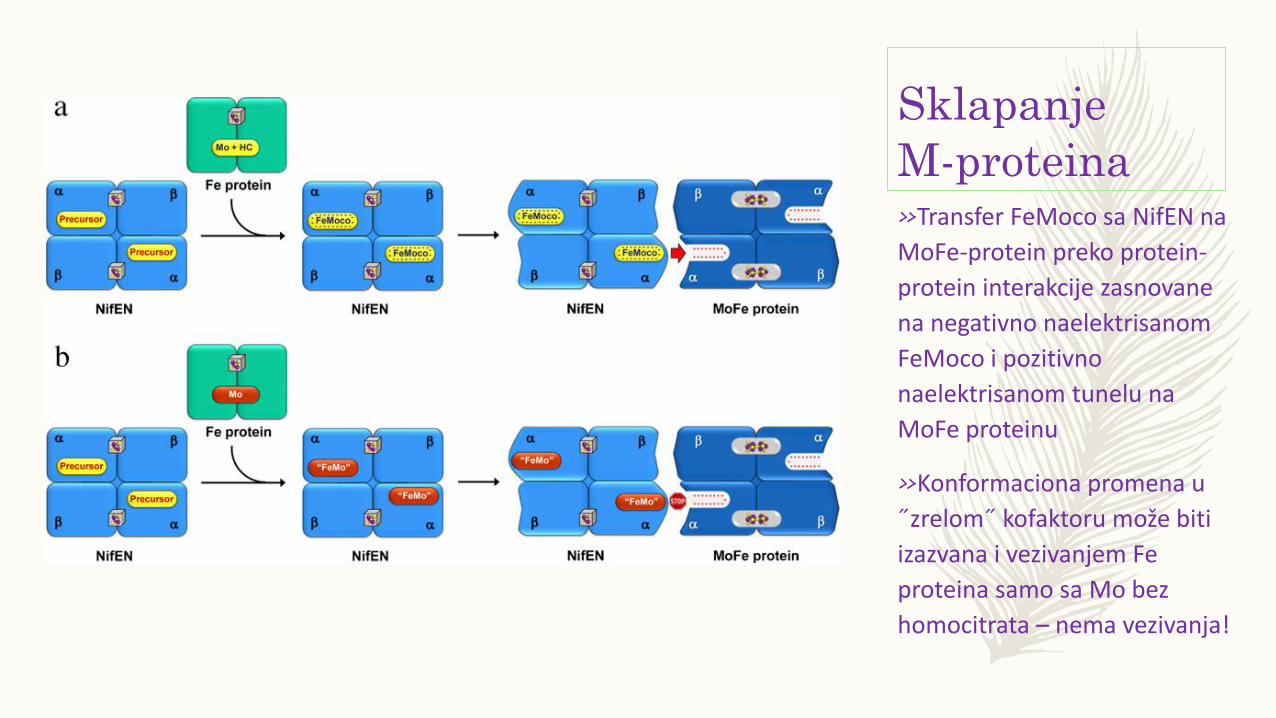
Sklapanje
M-proteina»Transfer FeMoco sa NifEN na
MoFe-protein preko protein-
protein interakcije zasnovane
na negativno naelektrisanom
FeMoco i pozitivno
naelektrisanom tunelu na
MoFe proteinu
»Konformaciona promena u
˝zrelom˝ kofaktoru može biti
izazvana i vezivanjem Fe
proteina samo sa Mo bez
homocitrata – nema vezivanja!

Regulacija ekspresije gena za
nitrogenazu
Transkripciona regulacija
» O2 i nivo NH4+
» Da li su simbionti osetljivi na NH4+?
Post-translaciona regulacija
» Kod slobodnoživećih
» Ograničena E, dovoljno N2
» Rhodospirillum rubrum, Rhodobacter capsulatus
*DRAT – dinitrogenazna reduktaza ADP-riboziltransferaza**DRAG – dinitrogenazna reduktaza-aktivirajuća glikohidrolazaR101 – Arg101 – N iz gvanido grupe Arg formira N-glikozidnu vezu sa ribozom iz ADP
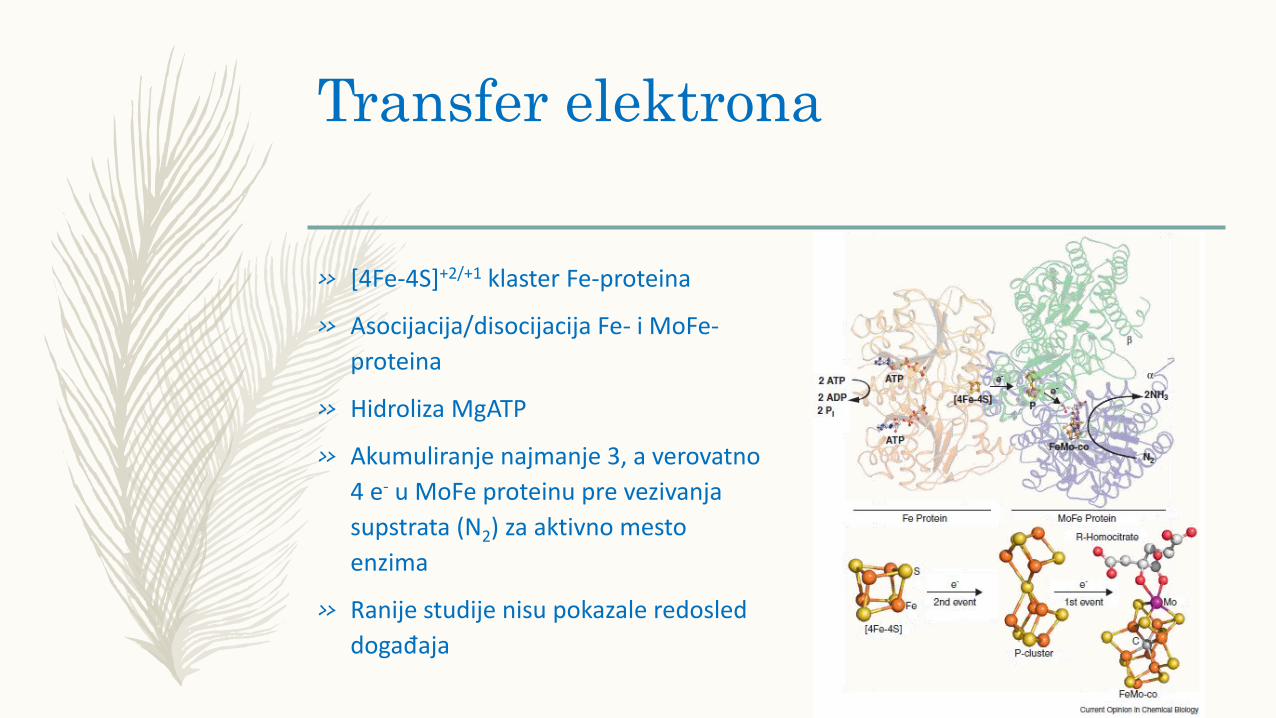
Transfer elektrona
» [4Fe-4S]+2/+1 klaster Fe-proteina
» Asocijacija/disocijacija Fe- i MoFe-
proteina
» Hidroliza MgATP
» Akumuliranje najmanje 3, a verovatno
4 e- u MoFe proteinu pre vezivanja
supstrata (N2) za aktivno mesto
enzima
» Ranije studije nisu pokazale redosled
događaja
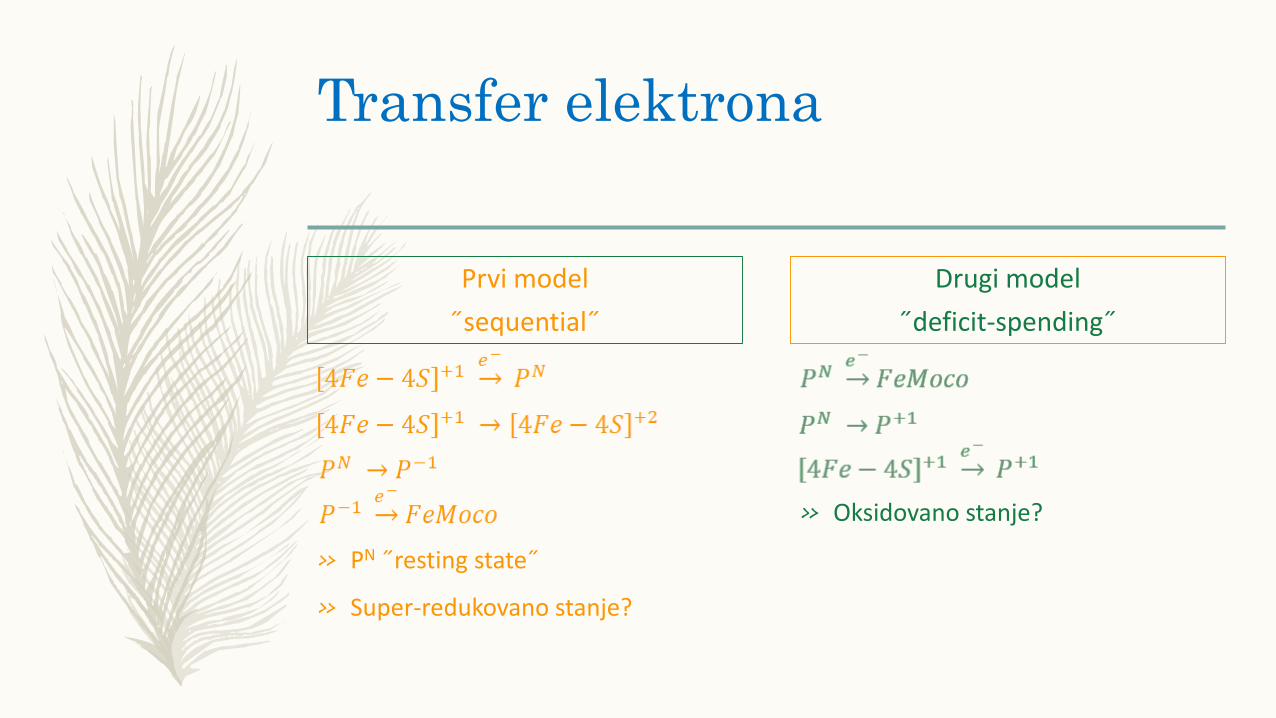
Transfer elektrona
Prvi model
˝sequential˝
» PN ˝resting state˝
» Super-redukovano stanje?
Drugi model
˝deficit-spending˝
» Oksidovano stanje?

» Trostruka veza u molekulu N2 ga čini inertnim, izuzetno skupa kataliza i
jedna od najtežih kataliza u biološkim sistemima
» Svi koraci redukcije direktno na nitrogenazi bez slobodnih intermedijera
» Može katalizovati redukciju različitih supstrata sa dvostrukom i
trostrukim vezama (acetilena do etilena, azida do N2 i amnijaka, CO2 do
CO, itd.)
» Više redoks stanja
» Unidirekciono kretanje e-
» Gde i kako se elektroni akumuliraju?
3. •Redukcija supstrata

Značaj fiksacije azota
Lucerka300 kg/ha/god
Bob600 kg/ha/god
Azotobacter7 kg/ha/god
Cyanobacteria80 kg/ha/god

Značaj fiksacije azota
» Pionirske zajednice
» Nova istraživanja – simbioza sa kukuruzom i pšenicom?
» Više sreće sa slobodnim diazotrofima?
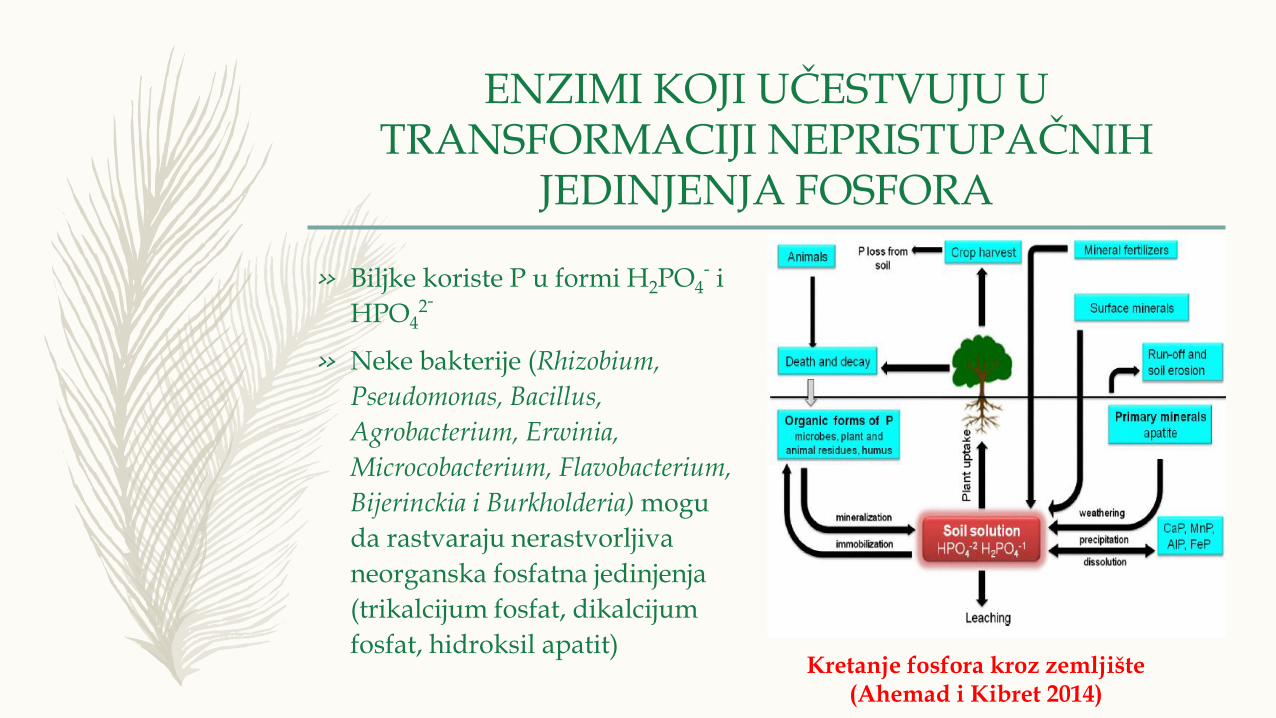
» Biljke koriste P u formi H2PO4- i
HPO42-
» Neke bakterije (Rhizobium,
Pseudomonas, Bacillus,
Agrobacterium, Erwinia,
Microcobacterium, Flavobacterium,
Bijerinckia i Burkholderia) mogu
da rastvaraju nerastvorljiva
neorganska fosfatna jedinjenja
(trikalcijum fosfat, dikalcijum
fosfat, hidroksil apatit)Kretanje fosfora kroz zemljište
(Ahemad i Kibret 2014)
ENZIMI KOJI UČESTVUJU U TRANSFORMACIJI NEPRISTUPAČNIH
JEDINJENJA FOSFORA
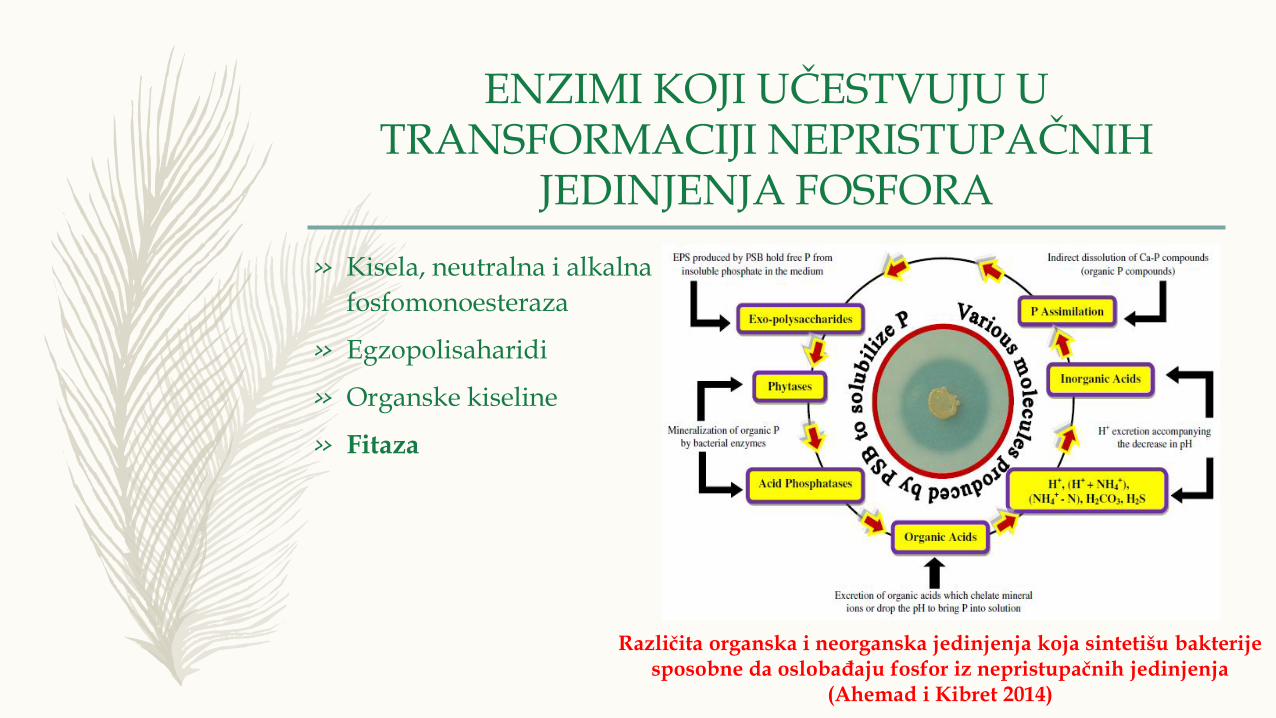
» Kisela, neutralna i alkalna
fosfomonoesteraza
» Egzopolisaharidi
» Organske kiseline
» Fitaza
ENZIMI KOJI UČESTVUJU U TRANSFORMACIJI NEPRISTUPAČNIH
JEDINJENJA FOSFORA
Različita organska i neorganska jedinjenja koja sintetišu bakterijesposobne da oslobađaju fosfor iz nepristupačnih jedinjenja
(Ahemad i Kibret 2014)
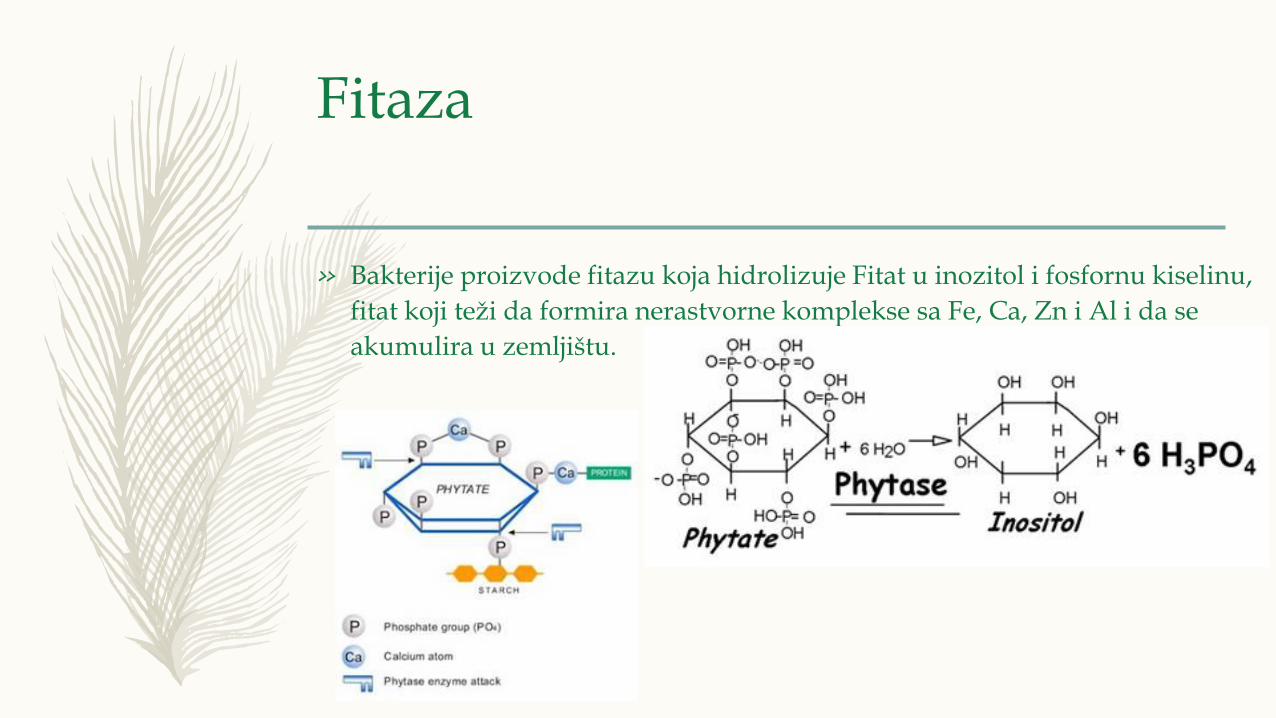
Fitaza
» Bakterije proizvode fitazu koja hidrolizuje Fitat u inozitol i fosfornu kiselinu,
fitat koji teži da formira nerastvorne komplekse sa Fe, Ca, Zn i Al i da se
akumulira u zemljištu.

BAKTERIJSKI ENZIMI KOJI UČESTVUJU U BIOSINTEZI AUKSINA
– Prva grupa biljnih hormona koji su otkriveni na osnovu toga što stimulišu
rastenje biljnih ćelija.
– Naziv potiče od grčke reči auxein – rasti.
– Po hemijskom sastavu glavna grupa auksina su indolna jedinjenja, ali kod
nekih je prisutna i fenil sirćetna kiselina.
– Pored biljaka, mnoge bakterije proizvode auksin kao što su bakterije
rizosfere fitopatogene (Agrobacterium tumefaciens) i PGPB, ali nije dokazan
uticaj auksina na rast i razmnožavanje bakterija.
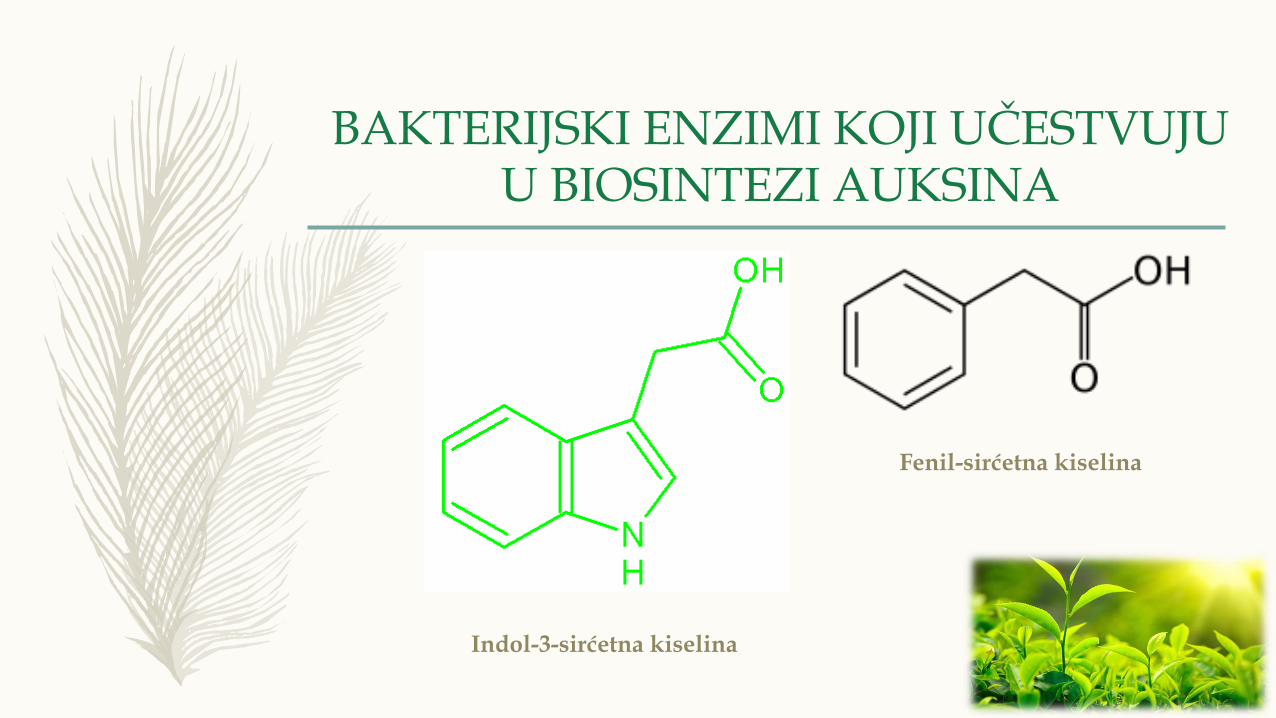
BAKTERIJSKI ENZIMI KOJI UČESTVUJU U BIOSINTEZI AUKSINA
Indol-3-sirćetna kiselina
Fenil-sirćetna kiselina

BIOSINTEZA AUKSINA –INDOL-3-SIRĆETNE KISELINE (IAA)
– Kod viših biljaka postoje dva glavna puta biosinteze auksina:
1. triptofan-nezavisan put biosinteze (indol-3-glicerol fosfat je najverovatniji
prekursor)
2. triptofan-zavisan put biosinteze u kojoj je definisano više puteva:
- put indol-3-acetoamida
- put indol-3-pirogrožđane kiseline
- put triptamina
- put indol-3-acetaldoksima...
– Neke bakterije mogu sintetisati auksine zasebnim putem sličnim triptofan-
zavisnim putevima viših biljaka.
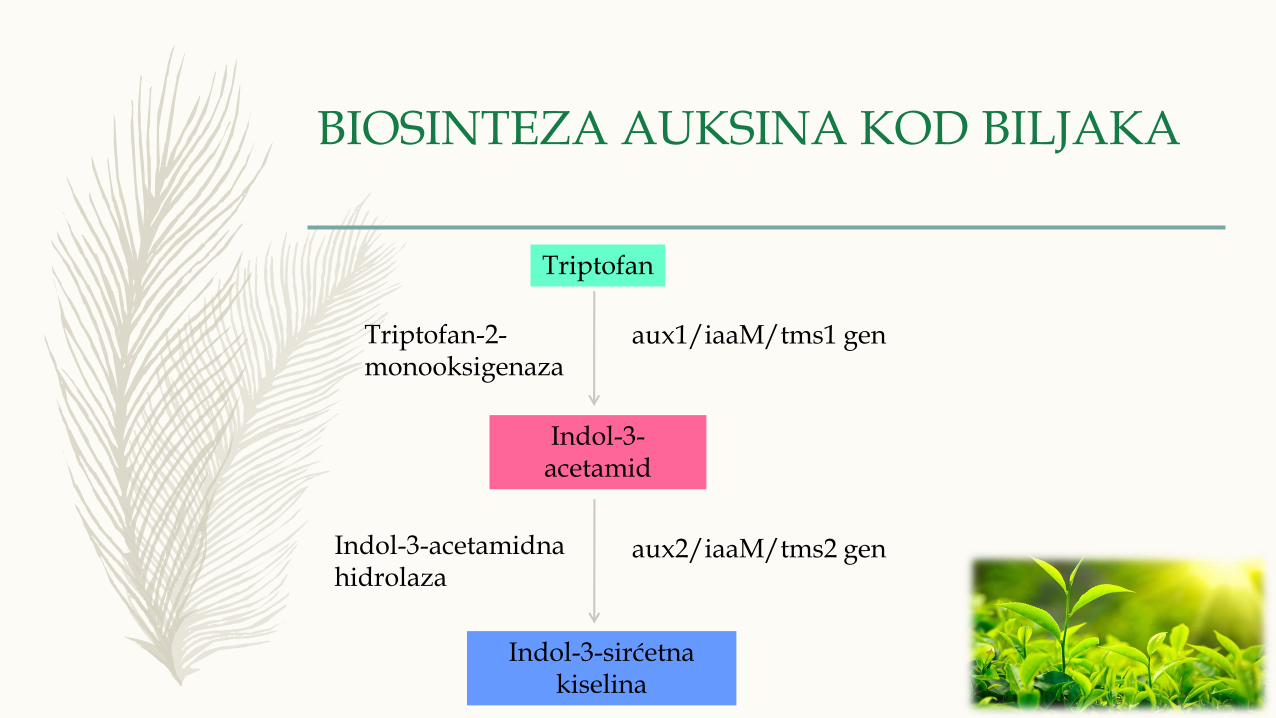
BIOSINTEZA AUKSINA KOD BILJAKA
Triptofan
Indol-3-acetamid
Indol-3-sirćetna kiselina
Triptofan-2-monooksigenaza
Indol-3-acetamidna hidrolaza
aux1/iaaM/tms1 gen
aux2/iaaM/tms2 gen
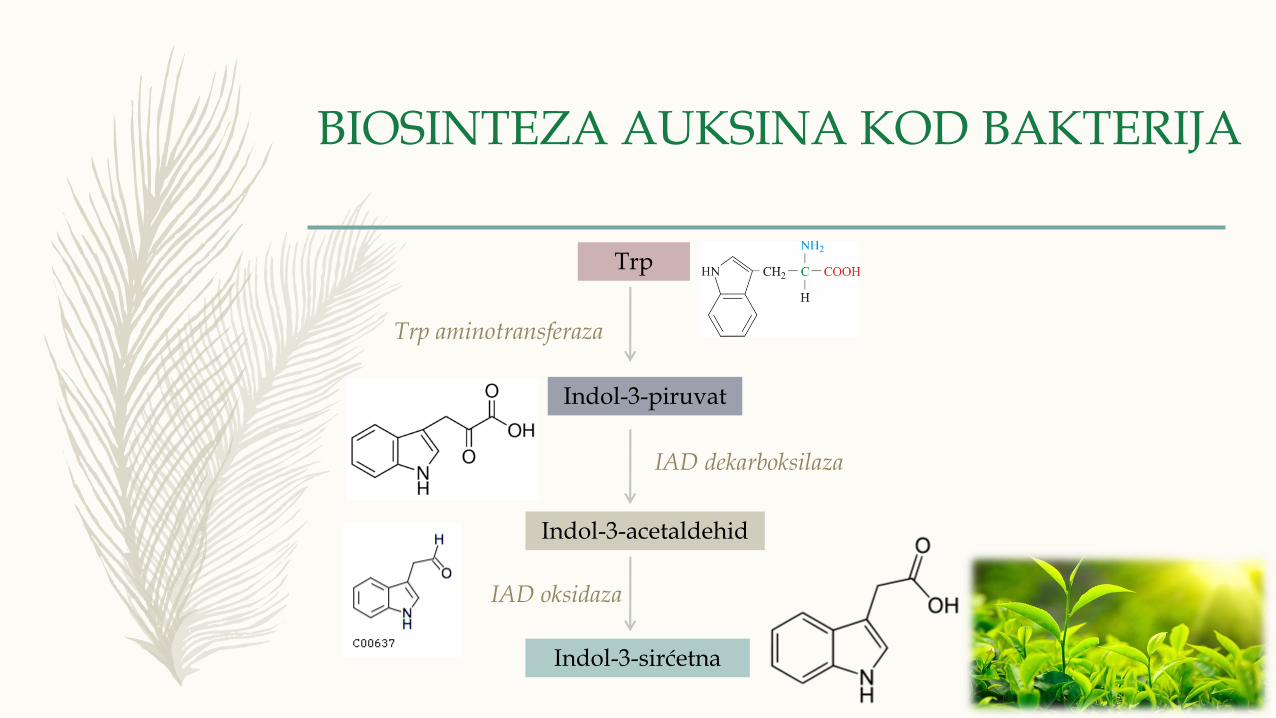
BIOSINTEZA AUKSINA KOD BAKTERIJA
Trp
Indol-3-piruvat
Indol-3-acetaldehid
Indol-3-sirćetna
Trp aminotransferaza
IAD dekarboksilaza
IAD oksidaza
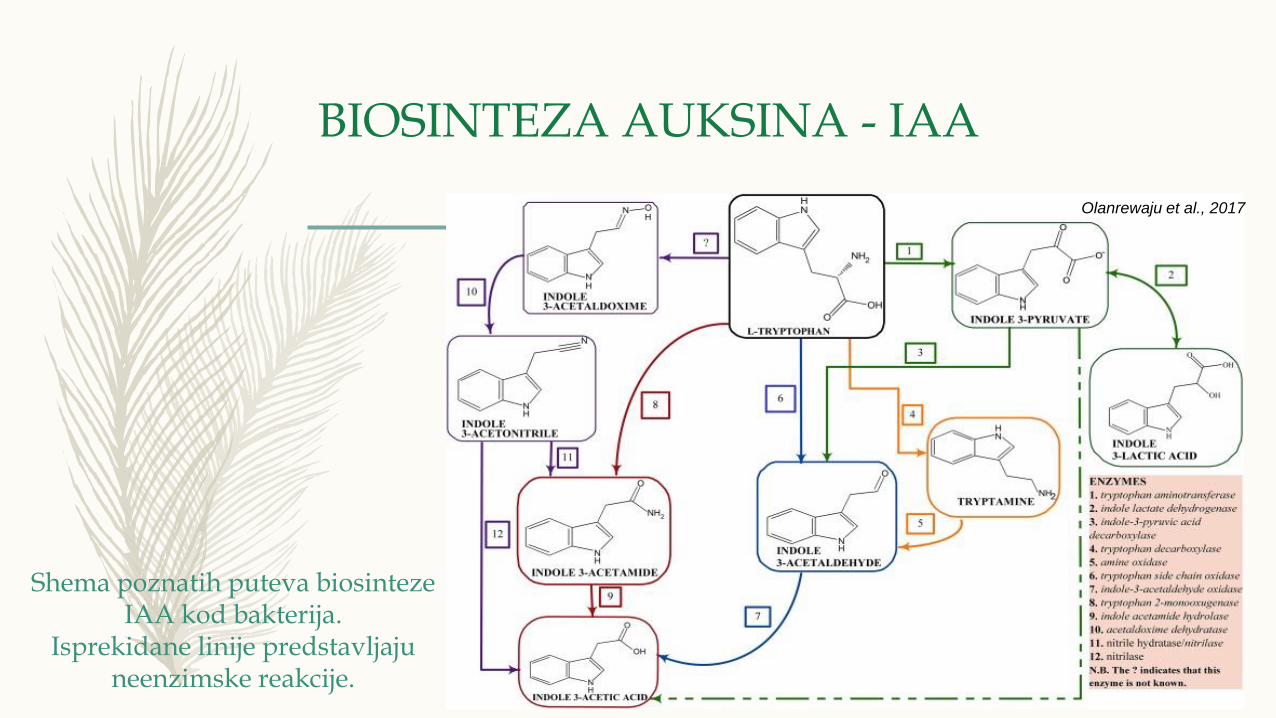
BIOSINTEZA AUKSINA - IAA
Olanrewaju et al., 2017
Shema poznatih puteva biosinteze IAA kod bakterija.
Isprekidane linije predstavljaju neenzimske reakcije.
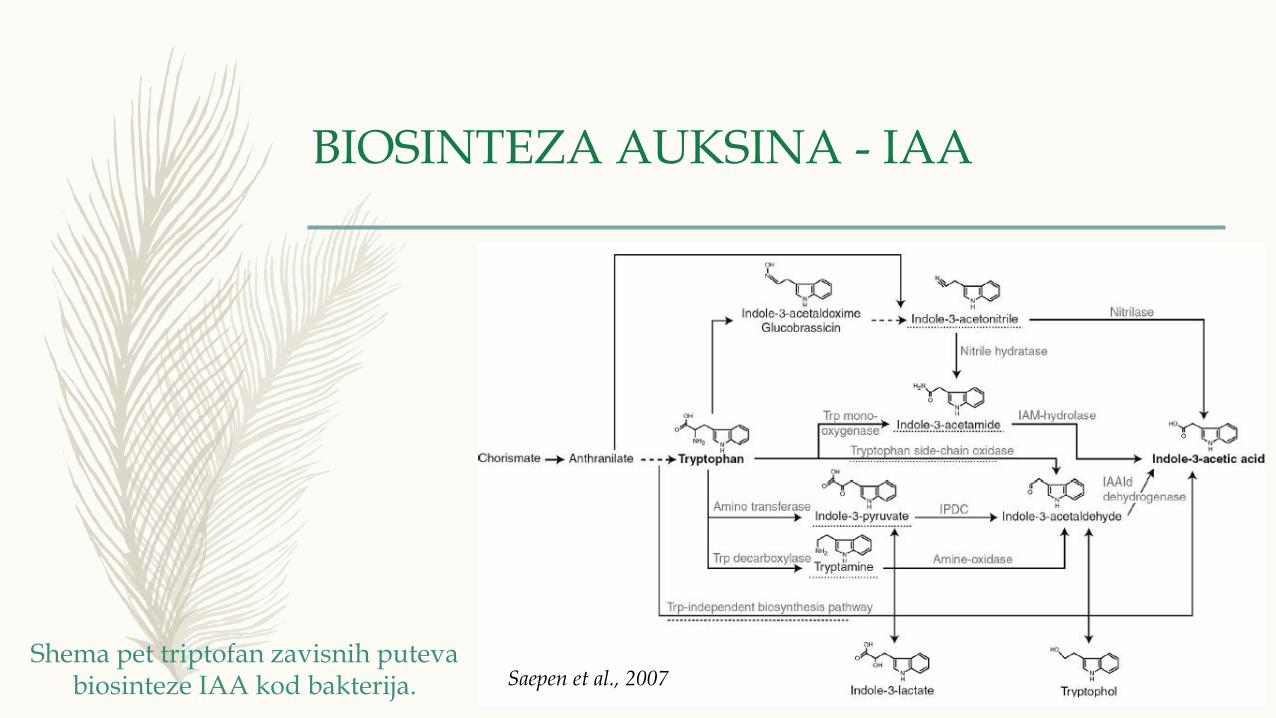
BIOSINTEZA AUKSINA - IAA
Shema pet triptofan zavisnih puteva biosinteze IAA kod bakterija. Saepen et al., 2007

1-aminociklopropan-1-karboksilat deaminazna (ACC)
➢ Etilen je jedini gasoviti biljni hormon kojisintetišu biljke usled delovanja različitihbiotičkih i abiotičkih procesa. Poznat je i kao hormon stresa jer pomaže biljkama da prežive sušu, visoku koncentraciju soli, toksičnihmetala (Ni, Pb, Zn i Cd), poplave, prisustvo patogenih bakterija.
(Glick 2014)

➢ ACC deaminaza katalizuje razgradnju ACC na α-ketobutirat i amonijak i na taj način smanjuje nivo etilena, a dobijeni metaboliti se koriste za rast bakterija. Ovaj enzim je pronađen samo kod mikroorganizama.
➢ Usled izlaganje biljke stesnim situacijama primećena su dva pika količine etilena. Jedan je manji i veruje se da on odgovoran za transkripciju gena koji kodiraju baketijske zaštitne proteine. A drugi pik je mnogo veći i javlja se u situacijama koje štete biljci i izazivaju senescenciju, hlorozu i abscisiju...
Olanrewaju et al., 2017
1-aminociklopropan-1-karboksilat deaminazna (ACC)
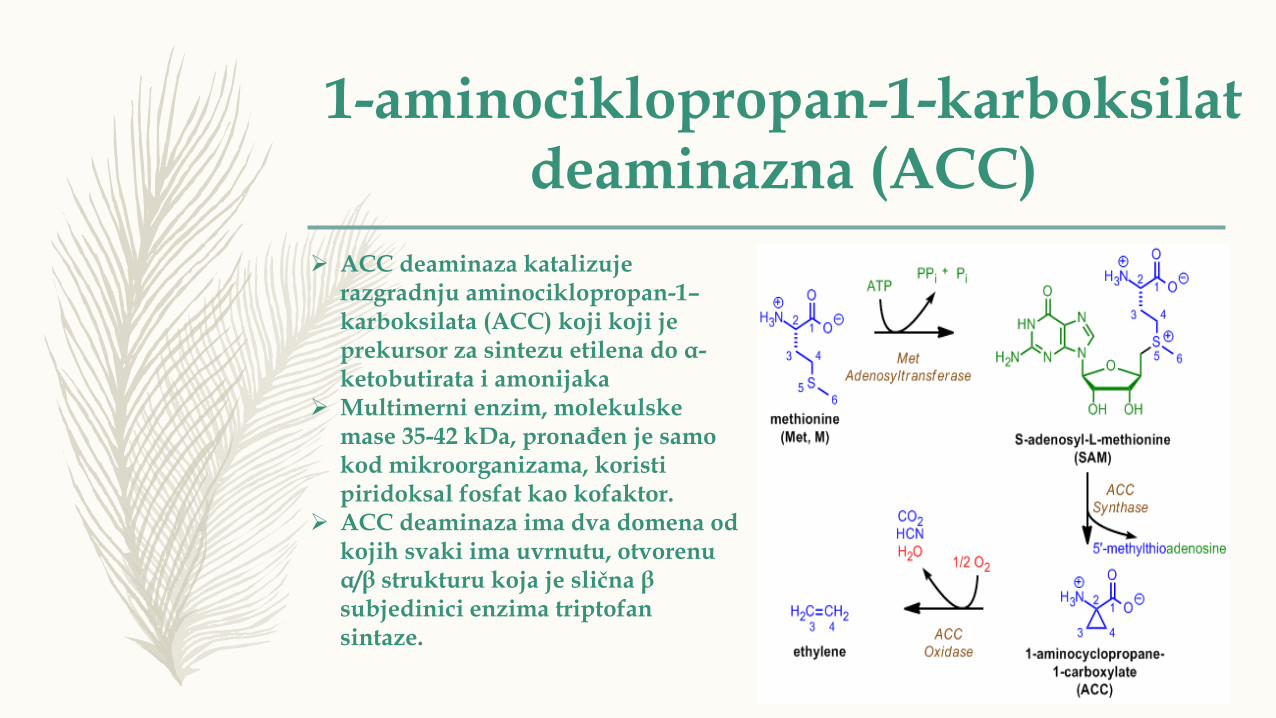
➢ ACC deaminaza katalizuje razgradnju aminociklopropan-1–karboksilata (ACC) koji koji je prekursor za sintezu etilena do α-ketobutirata i amonijaka
➢ Multimerni enzim, molekulske mase 35-42 kDa, pronađen je samo kod mikroorganizama, koristi piridoksal fosfat kao kofaktor.
➢ ACC deaminaza ima dva domena od kojih svaki ima uvrnutu, otvorenu α/β strukturu koja je slična β subjedinici enzima triptofan sintaze.
1-aminociklopropan-1-karboksilat deaminazna (ACC)

➢ Dobijeni 5-metil-tioadenozin ulazi u proces regeneracije metionina koji se zove Jangov ciklus.
➢ Sledeći korak je prevođenje ACC-a u etilen pomoću ACC-oksidaze. Međutim da bi se održao optimalan nivo etilena i kako bi i u stresnim uslovima biljka imala mogućnost da obavlja neke procese potrebno je da se smanji nivo etilena odnosno ACC-a, pomoću enzima ACC deaminaze.
➢ Otvaranje ciklopropanskog prstena je glavna karakteristika reakcije katalizovane od strane ACC deaminaze.
1-aminociklopropan-1-karboksilat deaminazna (ACC)

➢ Geni acdS ACC deaminaze su povezani sa genima fiksacije azota i moze biti regulisana pomocu nifA za koji je poznato da regulise i ekspresiju gena nif.
Geni koji kodiraju ACC deaminazu
➢ Jedan od glavnih reglutornih proteina koji učestvuju u ekspresiji acdS gena je LRP protein koji se sa AcdB proteinom vezujeza ACC i grade trinarni kompleks koji aktivira transkripciju acdS gena tako što se vezuje za njegov promotor. Takođe, transkripcija ovog gena je negativno regulsana leucinom koji se sintetiše od α-ketobutirata koji je produkt reakcije katalizovane ACC deaminazom, tako da leucin utice na isključivanje transkripcije acdS gena.

➢ Poboljšanje rasta biljke pomoću ACC deaminaze bakterija je motivisalo naučnike da prenesu gen koji kodira ovaj enzim u biljke. To bi predstavljalo jedan od načina da se smanji štetni efekat etilena kod biljaka podložnim stresnim situacijama. Transgena Petunia održava znatno smanjenu količinu etilena u ćelijama polena. Slično tome postojanje transgene uljane repice je doprinelo boljem rastu u uslovima povećane količine soli. Konstruisane su i dve sorte paradajza i duvana u kojima je zbog smanjene količine etilena odloženo starenje.
Transgene biljke sa aktivnošću ACC deaminaze
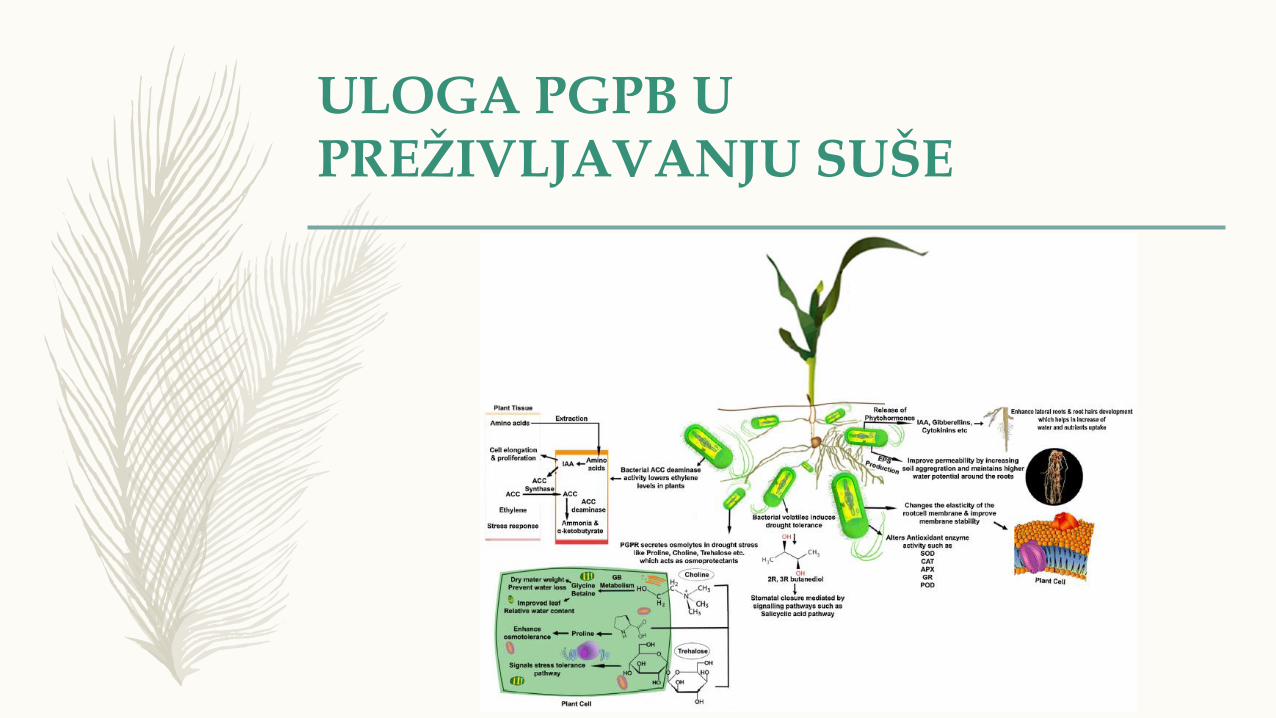
ULOGA PGPB UPREŽIVLJAVANJU SUŠE

ZAKLJUČAK
➢ Primena PGPB kao biofertilizatora je našla široku primenu u poljoprivredi
➢ Prednosti: - smanjuje se upotreba skupih mineralnih đubriva- biljke lakše usvajaju hemijske elemente- utiče se na dinamiku i pravac mikrobioloških procesa koji posredno
utiču na određivanje i povećanje plodnosti zemljišta- biološka kontrola
➢ U cilju odabira optimalnih sojeva bakterija, ubuduće je potrebno jasno definisati koje bakterijsko svojstvo je korisno i neophodno za određene uslove sredine i različite biljke, a u skladu sa tim i koja enzimska aktivnost ovih bakterija je korisna u datim uslovima.

Teme za prezentacije
» Fiksacija azota – primer Frankia
» Enzimi uključeni u sintezu citokina i giberelina kod bakterija
» Uticaj prisustva teških metala na sintezu IAA i ACC deaminaze
» Fitaze – nove uloge za stare enzime

Hvala na pažnji!












![[PPT]Slide 1 · Web viewTuliskan persamaan reaksi untuk produksi Etil Alkohol dengan cara penambahan air kepada Etilena. Berapa banyak Etil Alkohol dapat dibuat dari 14 ton Etilena?](https://img.pdfslide.net/doc/110x75/5b3fa74f7f8b9a91078c61bc/pptslide-1-web-viewtuliskan-persamaan-reaksi-untuk-produksi-etil-alkohol-dengan.jpg)
















