Embed Size (px)
Citation preview

Eur. J. Biochem. 233, 458-466 (1905) 0 FEBS 1995
UDP-N-acetylglucosamine : dolichyl-phosphate N-acetylglucosamine- 1 - phosphate transferase is amplified in tunicamycin-resistant soybean cells Yucheng ZENG and Alan D. ELBEIN Department of Biochemistry and Molecular Biology, University of Arkansas for Medical Sciences, Little Rock AR. USA
(Received 2 March/X May 1995) - EJB 95 0338/5
A tunicamycin-resistant soybean cell line was developed by gradually increasing the concentration of tunicamycin in the growth medium. At the final stage, the resistant cells could survive in media containing 60 pg/ml of tunicamycin, whereas normal cells show a greatly retarded growth rate at 0.5 pg/ml of antibiotic. The tunicamycin-resistant cells had a greater than 40-fold increase in the activity of the enzyme UDP-GlcNAc :dolichyl-P GlcNAclP transferase, a 2-3-fold increase in the activity of dolichyl-P-man- nose synthase, but no increase in the activities of other enzymes of the lipid-linked saccharide pathway such as dolichyl-P-glucose synthase or mannosyl transferases. There was also no change in the activities of the glycoprotein-processing enzymes, glucosidase I or glucosidase 11, as compared to wild-type cells. The increase in GlcNAclP transferase was due to an increased production of enzyme, as seen by a dramatic increase in the amount of a 39-kDa protein, which is presumed to be this enzyme protein. The GlcNAcl P transferase from tunicamycin-resistant cells was equally sensitive to tunicamycin as was the wild-type enzyme, but was considerably more labile to temperatures above 30°C. The activity in tuni- camycin-resistant cells was greatly stimulated by exogenous dolichyl-P. The spectrum of oligosaccharides from labeled lipid-linked oligosaccharides was similar in wild-type and tunicamycin-resistant soybean cells, but the resistant cells had significantly greater amounts of the shorter and much lower amounts of the larger-sized oligosaccharides.
Keyvtmds: tunicamycin-resistant ; glycosylation-altered soybean cells; amplification of GlcNAclP trans- ferase ; N-linked glycosylation ; complex carbohydrates.
In plant cells, the biosynthesis of the asparagine-linked oli- gosaccharide chains of the N-linked glycoproteins begins with the transfer of GlcNAc 1 P from UDP-GlcNAc to dolichyl-P to form GlcNAc-PP-Do1 [l 1. This reaction is followed by the addi- tion of a second GlcNAc, also from UDP-GlcNAc, to GlcNAc- PP-Do1 giving GlcNAc-GlcNAc-PP-Do1 [2, 31. Nine mannose and three glucose residues are then added to produce a large lipid-linked oligosaccharide, i.e. Glc,Man,(GlcNAc),-PP-Do1 [4, 51. The initial seven reactions of this cycle have been pro- posed to occur on the cytoplasmic face of the endoplasmic retic- ulum membrane. since they involve the utilization of the nucleo- side diphosphate sugars UDP-GlcNAc and GDP-Man [6]. On the other hand, the last four Man and three Glc residues are added via the activated lipid-linked sugars, Dol-P-Man [7-9] and Dol-P-Glc [ I 0. 11 1 : these reactions are believed to occur in the lumen of the endoplasmic reticulum. The final step in this assembly pathway involves the transfer of the completed oligo- saccharide, Glc,Man,,(GlcNAc)l, from the lipid carrier to spe- cific asparagine residues on the nascent polypeptide chain. This reaction also occurs in the endoplasmic reticulum as the protein is being synthesized and inserted into the lumen [32, 131.
Correspor7det~e to A. D. Elbein. Department of Biochemistry and Molecular Biology, University of Arkansas for Medical Sciences, 4301 W. Markham, Slot 5 16. Little Rock, Arkansas 72205, USA
Ahhwviorioti. CHO, Chinese hamster ovary. En;ynzes. liDP-N-acetylg1ucosaniine:dolichyl-phosphate N-acetyl-
glucosarnine-I-phoaphate transferase. GlcNAclP transferase (EC 2.7.8.15); Dol-P-Man synthase (EC 2.4.1.83); Dol-P-Glc synthase (EC 2.4.1 . I 17).
The GlcNAclP transferase has been the subject of consider- able study since this enzyme catalyzes the first reaction in the assembly of the lipid-linked oligosaccharides and is therefore a prime site for regulation [14- 161. Thus, a number of attempts have been made to purify this protein from plants and animals but in most cases the enzyme has proved to be too unstable, and only a partial purification was achieved. Purification of the enzyme to homogeneity from mammary glands has been re- ported, but no information on regulation has been published [17]. In animal cell membranes 118, 191 and with the purified enzyme [15, 171, the activity is greatly stimulated by the addi- tion of Dol-P-Man to the incubation mixtures.
The antibiotic, tunicamycin, has been a valuable tool in the study of GlcNAclP transferase [20]. The structure of this com- pound somewhat resembles the two substrates UDP-GlcNAc and Dol-P, and tunicamycin has been postulated to be a bi-sub- strate analog that mimics the transition state [14]. Tunicamycin is a potent and fairly specific inhibitor of GlcNAcl P transferase 114, 21, 221 and does not inhibit other GlcNAc transferases [23, 241. As a result of its specificity and potent inhibitory activity, this antibiotic has been utilized in a novel way to isolate the gene for GlcNAclP transferase. Thus, long-term culture of Chinese hamster ovary (CHO) cells in increasing concentrations of tuni- camycin results in the isolation of a population of cells that are resistant to this inhibitor. These cells show a great increase in the activity of GlcNAcl P transferase, suggesting an increase in the copy number of the structural gene due to amplification [25, 261. Amplification of the gene was shown to be responsible for this resistance, and a full-length cDNA was cloned and se-

Zeng and Elbein (Eur: J. Biochem. 233) 4.59
quenced 127, 281. Overexpression of GlcNAcl P transferase has also been achieved in trypanosomastid parasites of the genus Leishmaniu [29, 301 by gradual acclimatization of cells to increasing concentrations of the drug. In yeast, overexpression of the gene for GlcNAclP transferase was accomplished by preparing a Saccharomyces cerevisiae genomic library in a multicopy plasmid vector and isolating tunicamycin-resistant transformants [3 11.
In the work described in the present paper, we have isolated a tunicamycin-resistant soybean cell line and demonstrate that this culture has a 40-fold elevation in the activity of GlcNAcl P transferase, as well as a small but significant increase in Dol-P- Man synthase. When the total membrane proteins from tuni- camycin-resistant soybean cells were subjected to SDS/PAGE, there was a dramatic increase in one protein with a migration of about 39 kDa, which is probably the GlcNAclP transferase pro- tein. In addition, the resistant cells show significant alterations in the profiles of the lipid-linked oligosaccharides as compared to wild-type cells.
MATERIALS AND METHODS
Materials. Tunicamycin was kindly supplied by Eli Lilly and Co. Dol-P (grade 1111, UDP-Glc and GDP-Man were from Sigma Chemical Co. UDP-['HIGlcNAc (28 Ci/mmol), UDP- ['HIGlc (60 Ci/mmol) and D-[2-3H]mannose (1 5 Ci/mmol) were from American Radiolabeled Chemical Co. GDP-['4C]mannose (284 Ci/mol) was from New England Nuclear Co. Silica gel (GHL) plates were from Analtech. All other chemicals were from reliable suppliers, and were of the highest grade available.
Isolation of tunicamycin-resistant soybean cell line. Soy- bean cells were routinely grown for 7 days at 28°C in suspen- sion culture in 500-ml conical flasks containing 100 ml B-5 me- dium [32]. This medium has the following composition (mg/l) : KNO,, 3000; (NH,),SO,, 134; MgSO, . 7 H,O, 500; CaCl, ' 7 H,O, 150; NaH,PO,. H20, 150; MnSO, . H,O, 100; KI, 0.75; H,BO,, 3 ; ZnSO, . 7 H,O, 2 ; CuSO,, 0.025; Na,MoO, . 2 H,O, 0.25 ; CoCI, . 6 H20, 0.025 ; EDTA-ferric sodium salt, 28; m y - inositol, 100; thiamin . HCI, 10; nicotinic acid, 1 ; pyridoxine . HCI, 1 ; sucrose, 20000; 2,4-dichlorophenoxyacetic acid, 0.5. The final solution was adjusted to pH 5.5.
For the selection of resistant cells, wild-type soybean cells were grown in 50-ml flasks containing 10 ml B-5 medium that contained increasing amounts of tunicamycin. Initially, the tuni- camycin concentration in the medium was 0.5 pg/ml, and this flask was innoculated with wild-type cells. These cells reached the normal level of growth in about 10 days. At that point, an aliquot of these cells were transferred to fresh medium that con- tained 1 pg/ml of tunicamycin, and the cells were allowed to grow until normal growth was again attained. This method was continued over a period of several months until a resistant cell line was obtained that could grow well in 60 pg/ml of tuni- camycin. The tunicamycin-resistant cells were kept in the same medium containing 60 pg/ml of the antibiotic. For maintenance of this cell line, the cells were transferred to solid medium (B- 5 medium containing 1.5 5% agar plus 60 yg/ml of tunicamycin) contained in flasks or petri dishes.
Preparation of cell membranes. Membrane fractions were prepared from three different soybean cells as follows : wild-type cells, tunicamycin-resistant cells grown for five passes in the absence of tunicamycin and tunicamycin-resistant cells grown in medium containing 60 pg/ml of antibiotic. In each case, the cells were in the log phase of growth when harvested. Cells were harvested by filtering through a funnel filled with glasswool. The cells were then washed well with buffer A (50 mM Tris/
HCl pH 7.2, containing 0.2.5 M sucrose, 1 mM EDTA, I mM MgCI,, 1 mM dithiothreitol, and 0.5 % polyvinylpyrrolidone). The washed cells (about 20 ml packed pellet) were suspended in 70 ml ice-cold buffer A and disrupted by sonic oscillation for 5 min with a Fisher model 300 sonic oscillator at 80% relative output with the large probe. Sonic oscillation was repeated twice more with a 10-min cooling period between each sonication. The lysate was subjected to low-speed centrifugation to remove unbroken cells and then to high-speed centrifugation to isolate membranes. The membrane fraction was resuspended in buffer B (20 mM Tris/HCI pH 7.2, containing 0.15 M NaCI, 10% glyc- erol, 5 mM MgCI,, 0.1 mM EDTA and 0.1 mM dithiothreitol), and stored in the freezer at -80°C until used. The GlcNAclP transferase activity was found to be stable under these conditions for at least two months.
Assays for the various enzymatic activities. GlcNAclP transferase. The assay mixture contained the following compo- nents in a final volume of 200 1-11: 50 mM Tris/HC1 pH 7.2, 0.14 M NaCI, 10 mM MgCIZ, 2 mM ATP, 2 pg DoI-P, 0.1 pCi UDP-[3H]GlcNAc and an appropriate amount of the particulate enzyme preparation. The reaction was initiated by the addition of enzyme, and incubations were usually for 15 min at 30°C. The reactions were stopped by the addition of CHCl,/CH,OH (2: 1) and the amount of radioactivity in the CHCI, phase was assayed as previously described [33].
Dol-P-Man synthase and Dol-P-Glc synthase. The assay mixtures for these two enzymes were essentially the same as that described above for the GlcNAclP transferase, except that the glycosyl donor was changed from UDP-GlcNAc to either GDP-['TC]Man (50000 cpm), or UDP-['HIGlc (80000 cpm). In- cubations were usually for 15 min at 30°C. Reactions were stopped by the addition of CHCl,/CH,OH and the radioactivity in the CHCI, phase was measured [34].
Glucosidase I. Glucosidase I was assayed by addition of appropriate amounts of the membrane enzyme fraction to a reac- tion mixture of 200 1-11 of the following components: 100 mM Mes pH 6.5, 0.1 5% Triton X-100, 2 mM EDTA, and 15000 cpm ['HIGlc-labeled Glc,Man,GlcNAc. Incubations were at 30°C for 1 h, after which time the reaction mixture was passed through a column of concanavalin-A- Sepharose to remove the radio- active oligosaccharides; the ['Hlglucose in the effluent was mea- sured 1351.
Glucosidase II. Glucosidase I1 was assayed in reaction mixtures that contained the following components in a volume of 200 1-11: 100 mM Mes pH 6.8, 0.1 % Triton X-100, 10 mM p-nitrophenyl a-D-glucopyranoside. Reactions were initiated by the addition of appropriate amounts of the particulate enzyme and the incubations were usually for 1 h at 30°C. The reactions were stopped by the addition of 2.5 ml 0.4 M glycine pH 10.4 and, after removal of insoluble material by centrifugation, the liberated p-nitrophenol was measured at 410 nm 1361.
In all of these cases, the protein content was measured by the method of Bradford [37].
Preparation of 'H-labeled Dol-PP-oligosaccharides in in- tact cells. Wild-type and tunicamycin-resistant soybean cells were harvested in the log phase of growth and washed several times with culture medium in which the sucrose had been re- placed with 0.14 M NaCl. An aliquot of the cell pellet (1.5 ml packed cells) was resuspended in 2.4 ml culture medium (with NaCl rather than sucrose), and 100 yCi [2-'Hlmannose was added. The cells were allowed to incubate for 5 - 50 min as indi- cated in the figures. The reactions were stopped by heating the cells for 5 min at 75°C and then 4 rnl 5 mM EDTA pH 8.0 was added. The cells were harvested by centrifugation and the cell pellets were washed several times with 5 mM EDTA to remove unreacted [2-'H]mannose.
460 Zeng and Elbein ( E M J . Biochern. 233)
The cells were then lyophilized to dryness and extracted with various CHCl,/CH,OH/H,O solutions to obtain the different lipid-linked saccharides. The pellet was first extracted three times, each with 5 ml CHCl,/CH,OH (2: I), to isolate the lipid- linked monosaccharides and short-chain oligosaccharides. The cell pellet was then washed three times, each with 8 ml water to remove free sugars and especially any ['Hlmannose.
Cell pellets were next suspended in 5 ml CHCl,/CH,OH/H,O (10:10:3) and sonicated three times, with a 5-min pause each time, to extract the larger-sized lipid-linked oligosaccharides. After standing for 1 h in this solvent, the cell pellet was isolated by centrifugation and the supernatant was removed and saved. Cell pellets were extracted twice more with 5-ml aliquots of CHCl,/CH,OH/H,O. The pooled 10: 30: 3 phase was analyzed directly by chromatography on DEAE-cellulose in CHCI,/ CH,OH/H,O, or the solvent could be removed by rotary evapo- ration, and the remainder suspended in 50% propanol containing 0.025 M HC1 and heated at 100°C for IS min to release the oligosaccharides. After hydrolysis, the solvent was removed un- der a stream of nitrogen, and CHClJwater was added with vigor- ous mixing to dissolve the sugars and lipids. The aqueous phase, containing the liberated oligosaccharides, was removed, taken to dryness, resuspended in 0.5 ml H20, and analyzed by gel filtra- tion on columns of Bio-Gel P-4.
Methods of purification and characterization. Thin-layer chromatography. The products of GlcNAcl P transferase activ- ity, after extraction into the CHC1, solvent, were applied directly to silica gel plates that were developed in CHCl ,/CH,OH/H,O (60: 25: 4). The radiolabeled glycolipids were located by cutting the plates into 1-cm strips and measuring their radioactive content in the scintillation counter.
HPLC separution of monosaccharides. The isolated GlcNAcl P transferase radioactive products were subjected to mild acid hydrolysis in 0.5 M HCl in 50% propanol. The lipids were removed by extraction with CHCl,, and the aqueous phase containing the liberated sugars was treated with mixed-bed resin to remove salt or other charged compounds. A mixture of stan- dard sugars was added to the aqueous phase from this treatment and the mixture was then subjected to Dionex chromatography on a Carbopak PA1 column. The column was cleaned with 200 mM NaOH and then equilibrated with 16 mM NaOH prior to application of the sample. After loading the sample, the col- umns were eluted with the same solvent. Eluted standard sugars were detected by pulsed amperometric detection and labeled sugars were detected by collecting fractions of the eluate and measuring the radioactivity in the scintillation counter.
DEAE-cellLilose chromatography. DEAE-cellulose (C1- ) was converted to the acetate form by treatment with 2 M acetic acid. After washing the resin with water until neutral, and then with methanol, the resin was suspended in CHCl,/CH,OH/H,O (10: 10:3). Glycolipid samples, dissolved in the same solvent, were applied to the column and samples were eluted in a step- wise manner with increasing concentrations of ammonium ace- tate. An aliquot of each elution was assayed for radioactive content.
Bio-Gel P-4 chrr,tnatography. Oligosaccharides liberated from the lipids by mild acid hydrolysis were applied to a column (1.2X200 cm) of Bio-Gel P-4 (200-400 mesh) which had been equilibrated with 1% acetic acid. The column was eluted with the same solvent, and 1 -ml fractions were collected and analyzed for radioactivity. Oligosaccharide standards of various sizes were used to calibrate the column.
RESULTS Selection of a tunicamycin-resistant soybean cell line. Wild- type soybean cells were initially inoculated into flasks contain-
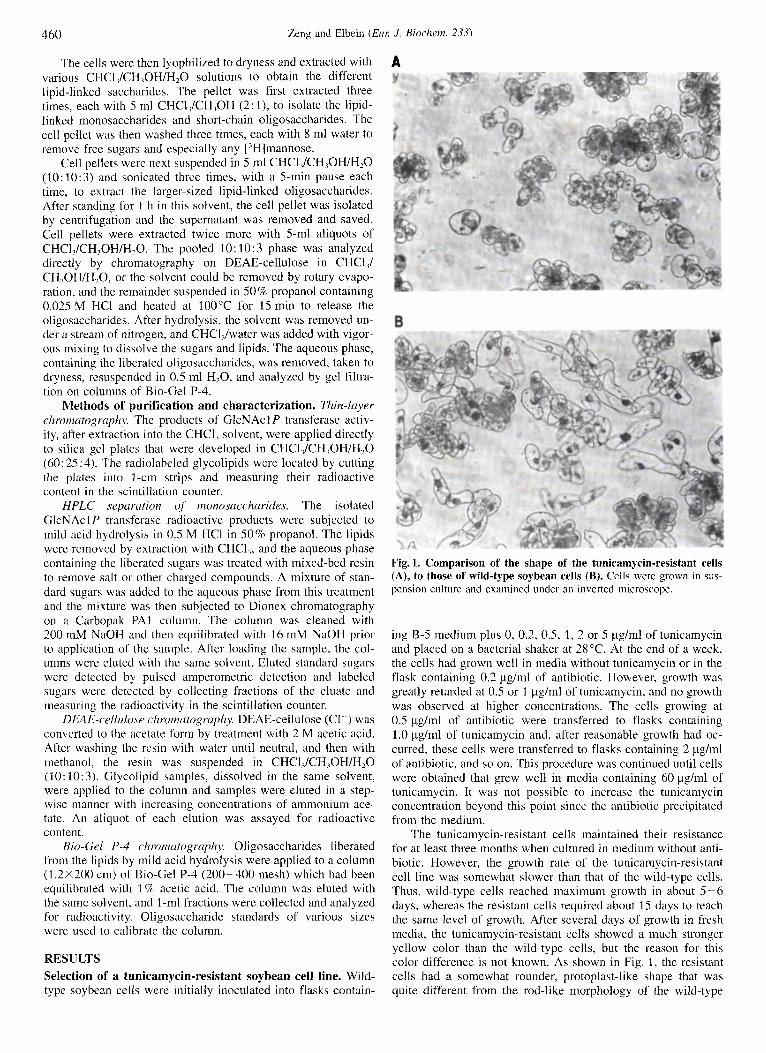
Fig. 1. Comparison of the shape of the tunicamycin-resistant cells (A), to those of wild-type soybean cells (B). Cells were grown in sus- pension culture and examined under an inverted microscope.
ing B-5 medium plus 0, 0.2, 0.5, 1, 2 or 5 pg/ml of tunicamycin and placed on a bacterial shaker at 28°C. At the end of a week, the cells had grown well in media without tunicamycin or in the flask containing 0.2 pg/ml of antibiotic. However, growth was greatly retarded at 0.5 or 1 pg/ml of tunicamycin, and no growth was observed at higher concentrations. The cells growing at 0.5 pg/ml of antibiotic were transferred to flasks containing 1.0 pg/ml of tunicamycin and, after reasonable growth had oc- curred, these cells were transferred to flasks containing 2 pg/ml of antibiotic, and so on. This procedure was continued until cells were obtained that grew well in media containing 60 pg/ml of tunicamycin. It was not possible to increase the tunicamycin concentration beyond this point since the antibiotic precipitated from the medium.
The tunicamycin-resistant cells maintained their resistance for at least three months when cultured in medium without anti- biotic. However, the growth rate of the tunicamycin-resistant cell line was somewhat slower than that of the wild-type cells. Thus, wild-type cells reached maximum growth in about 5-6 days, whereas the resistant cells required about IS days to reach the same level of growth. After several days of growth in fresh media, the tunicamycin-resistant cells showed a much stronger yellow color than th,e wild-type cells, but the reason for this color difference is not known. As shown in Fig. 1, the resistant cells had a somewhat rounder, protoplast-like shape that was quite different from the rod-like morphology of the wild-type
Zeng and Elbein (ELK J. Aiochem. 233) 461
0 20 40 60 80 100 Membrane Protein ( pg)
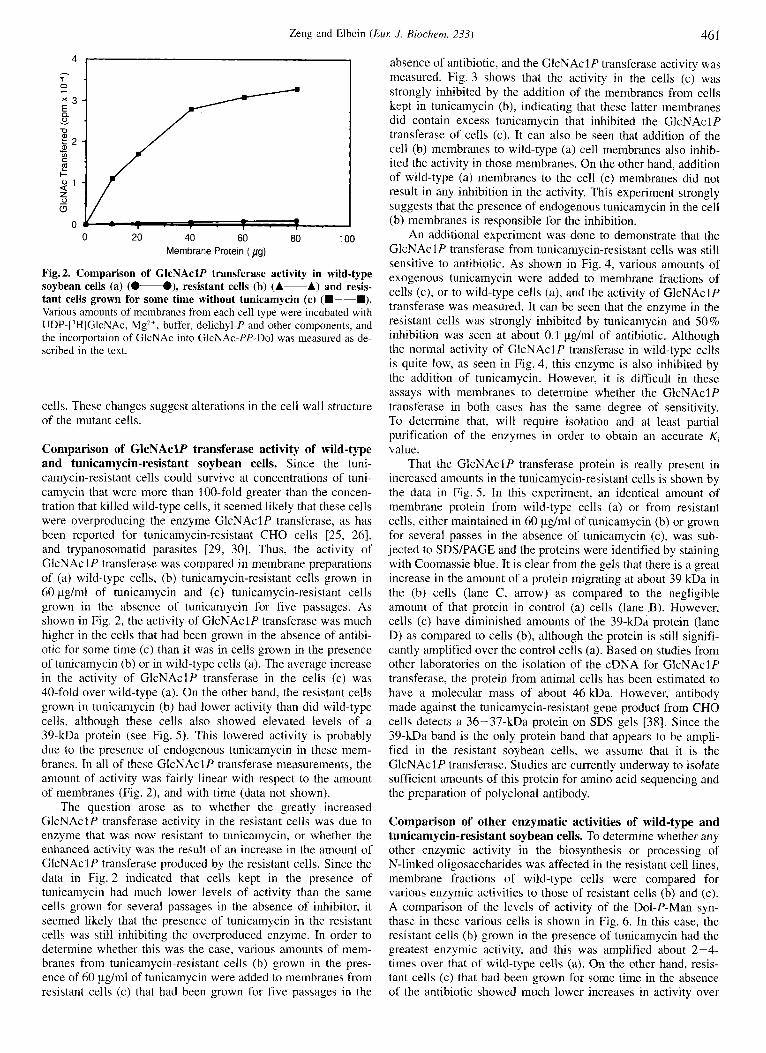
Fig. 2. Comparison of GlcNAclP transferase activity in wild-type soybean cells (a) (0-O), resistant cells (b) (A-A) and resis- tant cells grown for some time without tunicamycin (c) (m-m). Various amounts of membranes from each cell type were incubated with UDP-[’H]GlcNAc, Mgz+, buffer, dolichyl-P and other components, and the incorportaion of GlcNAc into GlcNAc-PP-Do1 was measured as de- scribed in the text.
cells. These changes suggest alterations in the cell wall structure of the mutant cells.
Comparison of GlcNAclP transferase activity of wild-type and tunicamycin-resistant soybean cells. Since the tuni- camycin-resistant cells could survive at concentrations of tuni- camycin that were more than 100-fold greater than the concen- tration that killed wild-type cells, it seemed likely that these cells were overproducing the enzyme GlcNAclP transferase, as has been reported for tunicamycin-resistant CHO cells [25, 261, and trypanosomatid parasites [29, 301. Thus, the activity of GlcNAcl P transferase was compared in membrane preparations of (a) wild-type cells, (b) tunicamycin-resistant cells grown in 60 pg/ml of tunicamycin and (c) tunicamycin-resistant cells grown in the absence of tunicamycin for five passages. As shown in Fig. 2, the activity of GlcNAclP transferase was much higher in the cells that had been grown in the absence of antibi- otic for some time (c) than it was in cells grown in the presence of tunicamycin (b) or in wild-type cells (a). The average increase in the activity of GlcNAclP transferase in the cells (c) was 40-fold over wild-type (a). On the other hand, the resistant cells grown in tunicamycin (b) had lower activity than did wild-type cells, although these cells also showed elevated levels of a 39-kDa protein (see Fig. 5) . This lowered activity is probably due to the presence of endogenous tunicamycin in these mem- branes. In all of these GlcNAclP transferase measurements, the amount of activity was fairly linear with respect to the amount of membranes (Fig. 2), and with time (data not shown).
The question arose as to whether the greatly increased GlcNAclP transferase activity in the resistant cells was due to enzyme that was now resistant to tunicamycin, or whether the enhanced activity was the result of an increase in the amount of GlcNAclP transferase produced by the resistant cells. Since the data in Fig. 2 indicated that cells kept in the presence of tunicamycin had much lower levels of activity than the same cells grown for several passages in the absence of inhibitor, it seemed likely that the presence of tunicamycin in the resistant cells was still inhibiting the overproduced enzyme. In order to determine whether this was the case, various amounts of mem- branes from tunicamycin-resistant cells (b) grown in the pres- ence of 60 pg/ml of tunicamycin were added to membranes from resistant cells (c) that had been grown for five passages in the
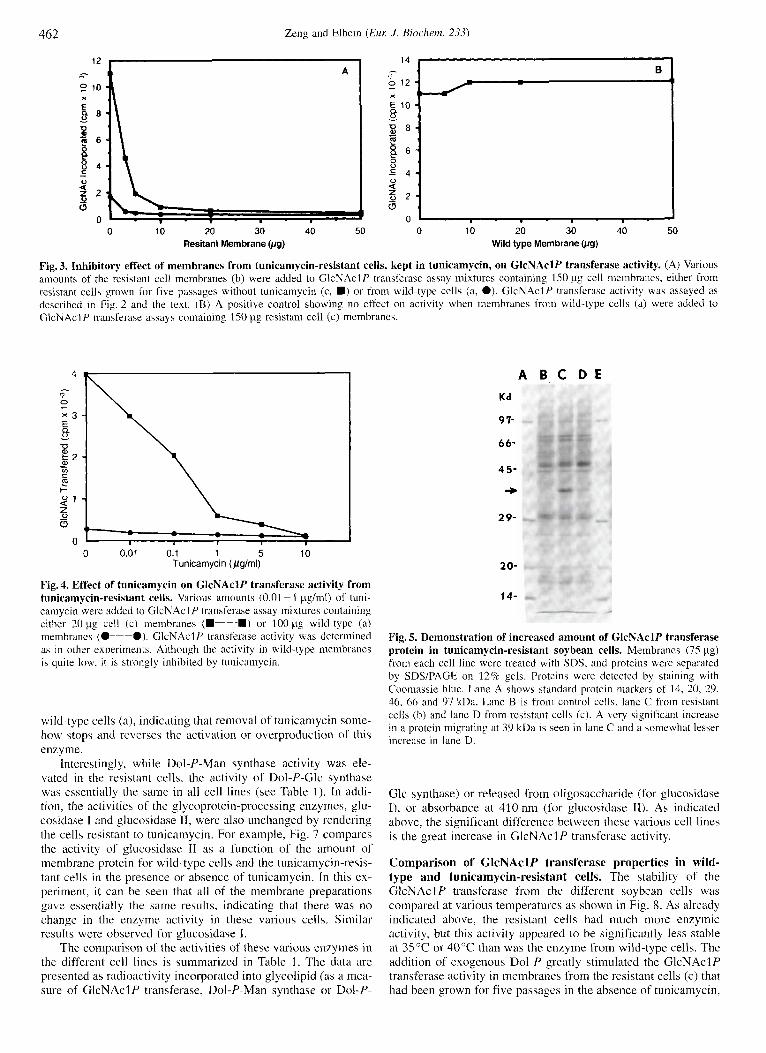
absence of antibiotic, and the GlcNAclP transferase activity was measured. Fig. 3 shows that the activity in the cells (c) was strongly inhibited by the addition of the membranes from cells kept in tunicamycin (b), indicating that these latter membranes did contain excess tunicamycin that inhibited the GlcNAclP transferase of cells (c). It can also be seen that addition of the cell (b) membranes to wild-type (a) cell membranes also inhib- ited the activity in those membranes. On the other hand, addition of wild-type (a) membranes to the cell (c) membranes did not result in any inhibition in the activity. This experiment strongly suggests that the presence of endogenous tunicamycin in the cell (b) membranes is responsible for the inhibition.
An additional experiment was done to demonstrate that the GlcNAcl P transferase from tunicamycin-resistant cells was still sensitive to antibiotic. As shown in Fig. 4, various amounts of exogenous tunicamycin were added to membrane fractions of cells (c), or to wild-type cells (a), and the activity of GlcNAclP transferase was measured. It can be seen that the enzyme in the resistant cells was strongly inhibited by tunicamycin and 50% inhibition was seen at about 0.1 pg/ml of antibiotic. Although the normal activity of GlcNAclP transferase in wild-type cells is quite low, as seen in Fig. 4, this enzyme is also inhibited by the addition of tunicamycin. However, it is difficult in these assays with membranes to determine whether the GlcNAclP transferase in both cases has the same degree of sensitivity. To determine that, will require isolation and at least partial purification of the enzymes in order to obtain an accurate K, value.
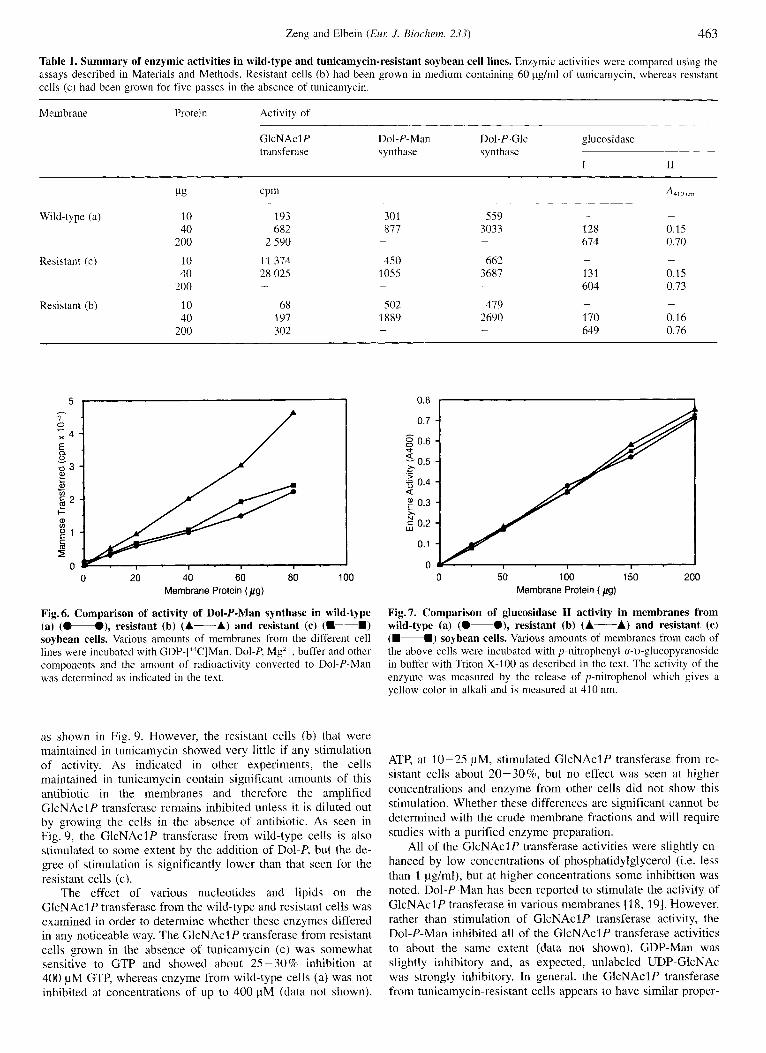
That the GlcNAclP transferase protein is really present in increased amounts in the tunicamycin-resistant cells is shown by the data in Fig. 5 . In this experiment, an identical amount of membrane protein from wild-type cells (a) or from resistant cells, either maintained in 60 pg/ml of tunicamycin (b) or grown for several passes in the absence of tunicamycin (c), was sub- jected to SDSPAGE and the proteins were identified by staining with Coomassie blue. It is clear from the gels that there is a great increase in the amount of a protein migrating at about 39 kDa in the (b) cells (lane C, arrow) as compared to the negligible amount of that protein in control (a) cells (lane B). However, cells (c) have diminished amounts of the 39-kDa protein (lane D) as compared to cells (b), although the protein is still signifi- cantly amplified over the control cells (a). Based on studies from other laboratories on the isolation of the cDNA for GlcNAcIP transferase, the protein from animal cells has been estimated to have a molecular mass of about 46kDa. However, antibody made against the tunicamycin-resistant gene product from CHO cells detects a 36-37-kDa protein on SDS gels [38]. Since the 39-kDa band is the only protein band that appears to be ampli- fied in the resistant soybean cells, we assume that it is the GlcNAclP transferase. Studies are currently underway to isolate sufficient amounts of this protein for amino acid sequencing and the preparation of polyclonal antibody.
Comparison of other enzymatic activities of wild-type and tunicamycin-resistant soybean cells. To determine whether any other enzymic activity in the biosynthesis or processing of N-linked oligosaccharides was affected in the resistant cell lines, membrane fractions of wild-type cells were compared for various enzymic activities to those of resistant cells (b) and (c). A comparison of the levels of activity of the Dol-P-Man syn- thase in these various cells is shown in Fig. 6. In this case, the resistant cells (b) grown in the presence of tunicamycin had the greatest enzymic activity, and this was amplified about 2-4- times over that of wild-type cells (a). On the other hand, resis- tant cells (c) that had been grown for some time in the absence of the antibiotic showed much lower increases in activity over
462 Zeng and Elbein (EM J . Biochern. 233)
0 10 20 30 40 50 0 10 20 30 40 50 Resitant Membrane bg) Wild type Membrane Wg)
Fig. 3. Inhibitory effect of membranes from tunicamycin-resistant cells, kept in tunicamycin, on GlcNAclP transferase activity. (A) Various amounts of the resistant cell membranes (b) were added to GlcNAclP transferase assay mixtures containing 150 pg cell membranes, either from resistant cells grown for five passages without tunicamycin (c , U) or from wild-type cells (a, e). GlcNAclP transferase activity wa? assayed as described in Fig. 2 and the text. (B) A positive control showing no effect on activity when membranes from wild-type cells (a) were added to GlcNAclP transferase assay!, containing 150 pg resistant cell (c) membranes.
4
x 3 E 8 - 7J g 2
c
C
I- 2
A B C D E Kd
9 7-
6 6-
4 5-
+ 2 9-
0 0.01 0.1 1 5 10 Tunicamycin (pglml)
Fig. 4. Effect of tunicamycin on GlcNAclP transferase activity from tunicamycin-resistant cells. Various amounts (0.01 ~ 1 pg/ml) of tuni- cainycin were added to GlcNAclP transferase assay mixtures containing either 20 pg cell ( c ) membranes (U-U) or 100 pg wild-type (a) membranes ( O p e ) . GlcNAcl P transferase activity was determined as in other experiments. Although the activity in wild-type membranes is quite low, i t is strongly inhibited by tunicamycin.
wild-type cells (a), indicating that removal of tunicamycin some- how stops and reverses the activation or overproduction of this enzyme.
Interestingly, while Dol-P -Man synthase activity was ele- vated in the resistant cells, the activity of Dol-P-Glc synthase was essentially the same in all cell lines (see Table 1 ) . In addi- tion, the activities of the glycoprotein-processing enzymes, glu- cosidase I and glucosidase 11, were also unchanged by rendering the cells resistant to tunicamycin. For example, Fig. 7 compares the activity of glucosidase I1 as a function of the amount of membrane protein for wild-type cells and the tunicamycin-resis- tant cells in the presence or absence of tunicamycin. In this ex- periment, it can be seen that all of the membrane preparations gave essentially the same results. indicating that there was no change in the enzyme activity i n these various cells. Similar results were observed for glucosidase I.
The comparison of the activities of these various enzymes in the different cell lines is summarized in Table 1 . The data are presented as radioactivity incorporated into glycolipid (as a mea- sure of GlcNAclP transferase, Dol-P-Man synthase or Dol-P-
20-
14-
Fig. 5. Demonstration of increased amount of GlcNAclP transferase protein in tunicamycin-resistant soybean cells. Membranes (75 pg) from each cell line were treated with SDS, and proteins were separated by SDS/PAGE on 12% gels. Proteins were detected by staining with Coornassie blue. Lane A shows standard protein markers of 14. 20, 29, 46, 66 and 97 kDa. Lane B is from control cells, lane C froin resistant cells (b) and lane D from resistant cells (c). A very significant increase in a protein migrating at 39 kDa is seen in lane C and a somewhat lesser increase in lane D.
Glc synthase) or released from oligosaccharide (for glucosidase I), or absorbance at 410 nm (for glucosidase TI). As indicated above, the significant difference between these various cell lines is the great increase in GlcNAcl P transferase activity.
Comparison of GlcNAclP transferase properties in wild- type and tunicamycin-resistant cells. The stability of the GlcNAclP transferase from the different soybean cells was compared at various temperatures as shown in Fig. 8. As already indicated above, the resistant cells had much more enzymic activity, but this activity appeared to be significantly less stable at 35°C or 40°C than was the enzyme from wild-type cells. The addition of exogenous Dol-P greatly stimulated the GlcNAclP transferase activity in membranes from the resistant cells (c) that had been grown for five passages in the absence of tunicamycin,
Zeng and Elbein ( E M J. Biorlzem. 233) 463
Table 1. Summary of enzymic activities in wild-type and tunicamycin-resistant soybean cell lines. Enzymic activities were compared using the assays described in Materials and Methods. Resistant cells (b) had been grown in medium containing 60 Fg/ml of tunicamycin, whereas resistant cells (c) had been grown for five passes in the absence of tunicamycin.
Membrane Protein Activity of
GlcNAclP Dol-P-Man Dol-P-Glc glucosidase transferase s ynthase s ynthase
I I1
Wild-type (a) 10 40
200
Resistant (c) 10 40
200
Resistant (b) 10 40
200
193 682
2 590
11 374 28 025 -
68 197 302
301 817 -
450 1055
502 1889
-
-
- 559 3033 128
614 -
- 662 3687 131
604 -
- 419 2690 170
649 -
-
0.15 0.70 -
0.15 0.73 -
0.16 0.76
7
x 4 -
Y - ' I . I - I
0 20 40 60 80 100 Membrane Protein ( p g )
Fig. 6. Comparison of activity of Dol-P-Man syntbase in wild-type (a) (0-O), resistant (b) (A-A) and resistant (c) (M-4) soybean cells. Various amounts of membranes from the different cell lines were incubated with GDP-['"C]Man. Dol-P, Mg", buffer and other components and the amount of radioactivity converted to Dol-P-Man was determined as indicated in the text.
as shown in Fig. 9. However, the resistant cells (b) that were maintained in tunicamycin showed very little if any stimulation of activity. As indicated in other experiments, the cells maintained in tunicamycin contain significant amounts of this antibiotic in the membranes and therefore the amplified GlcNAcI P transferase remains inhibited unless it is diluted out by growing the cells in the absence of antibiotic. As seen in Fig. 9, the GlcNAclP transferase from wild-type cells is also stimulated to some extent by the addition of Dol-P, but the de- gree of stimulation is significantly lower than that seen for the resistant cells (c).
The effect of various nucleotides and lipids on the GIcNAcl P transferase from the wild-type and resistant cells was examined in order to determine whether these enzymes differed in any noticeable way. The GlcNAcl P transferase from resistant cells grown in the absence of tunicamycin (c) was somewhat sensitive to GTP and showed about 25-30% inhibition at 400 pM GTP, whereas enzyme from wild-type cells (a) was not inhibited at concentrations of up to 400 pM (data not shown).
0.7
0.6
0.5
.$ 0.4 < 0.3
d
r * ._
x ; 0.2
0.1
0 0 50 100 150 200
Membrane Protein ( pg)
Fig. 7. Comparison of glucosidase I1 activity in membranes from wild-type (a) (0-O), resistant (b) (A-A) and resistant (c) (4-M) soybean cells. Various amounts of membranes from each of the above cells were incubated with p-nitrophenyl a-11-glucopyranoside in buffer with Triton X-100 as described in the text. The activity of the enzyme was measured by the release of p-nitrophenol which gives a yellow color in alkali and is measured at 41 0 nm.
ATP, at 10-25 pM, stimulated GlcNAclP transferase from re- sistant cells about 20-30%, but no effect was seen at higher concentrations and enzyme from other cells did not show this stimulation. Whether these differences are significant cannot be determined with the crude membrane fractions and will require studies with a purified enzyme preparation.
All of the GlcNAcl P transferase activities were slightly en- hanced by low concentrations of phosphatidylglycerol (i.e. less than 1 pg/ml), but at higher concentrations some inhibition was noted. Dol-P-Man has been reported to stimulate the activity of GlcNAcl P transferase in various membranes [18, 191. However, rather than stimulation of GlcNAclP transferase activity, the Dol-P-Man inhibited all of the GlcNAclP transferase activities to about the same extent (data not shown). GDP-Man was slightly inhibitory and, as expected, unlabeled UDP-GlcNAc was strongly inhibitory. In general, the GlcNAclP transferase from tunicamycin-resistant cells appears to have similar proper-
464 Zeng and Elbein (Eur: J . Biochern. 233)
0
10
" 20 25 30 35 40
Temperature ( O C )
Fig. 8. Effect of incubation temperature on the activity of GlcNAclP transferase in various soybean cell membranes. Incubation mixtures were prepared as indicated in the text and contained 40 pg cell (c) mem- branes (D-D) or 100 pg wild-type cell (a) inembranes (0-0). Incubation temperatures were as shown in the figure. GlcNAclP trans- ferase activity was measured as described in the text.
ties to that of wild-type cells, except for its enhanced suscep- tibility to temperataure.
Analysis of the product. To be certain that the product pro- duced from UDP-['HIGlcNAc was really GlcNAc-PP-Do1 in all of the soybean cell lines, the product that was extracted into the CHCI, phase was subjected to TLC on silica gel plates, and to column chromatography on DEAE-cellulose. The major radio- active glycolipid produced by the wild-type and resistant cells migrated with standard GlcNAc-PP-Do1 on TLC plates, and the R, value was similar to literature values for this compound. Like- wise, on DEAE-cellulose columns, the radioactive GlcNAc-lipid bound more tightly to the column than standard Dol-P-Man, and required 50 mM ammonium acetate to be completely eluted from the column. Again this is the expected behavior for a gly- colipid having a diphosphate linkage. Mild acid hydrolysis of the glycolipids from either cell line gave a radioactive product that was identified as GlcNAc by HPLC.
Structures of the lipid-linked oligosaccharides from tuni- camycin-resistant soybean cells. To determine whether resis- tance to tunicamycin resulted in any changes in the structures of
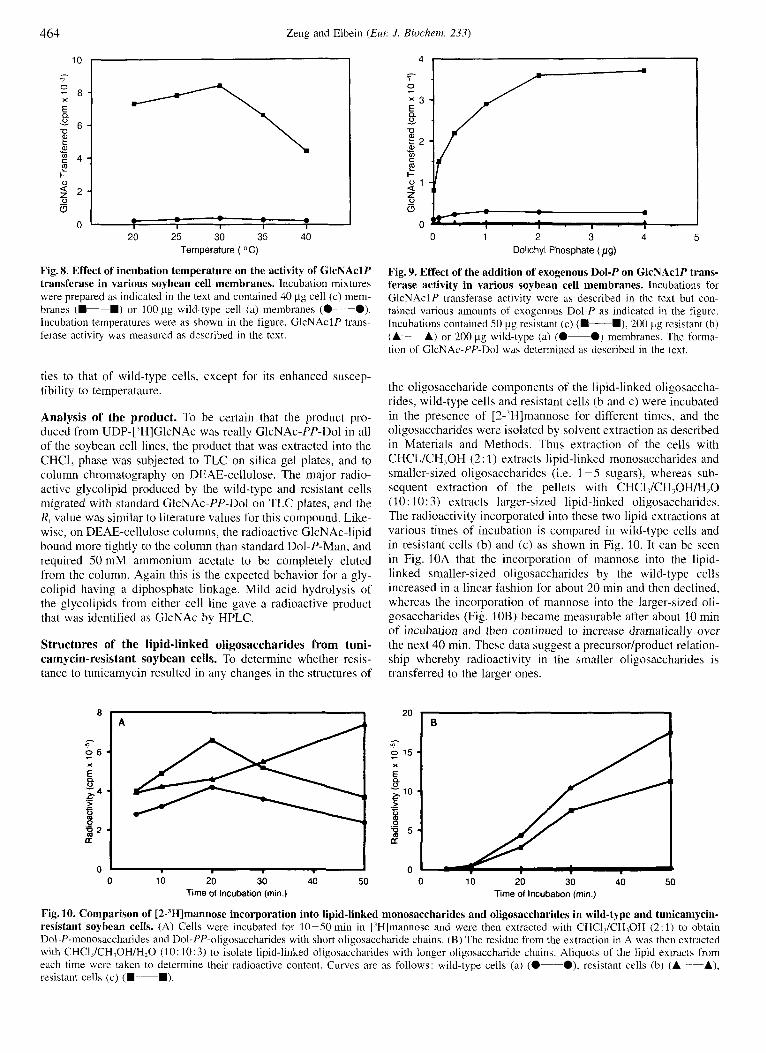
A
the oligosaccharide components of the lipid-linked oligosaccha- rides, wild-type cells and resistant cells (b and c) were incubated in the presence of [2-'H]mannose for different times, and the oligosaccharides were isolated by solvent extraction as described in Materials and Methods. Thus extraction of the cells with CHCl,/CH,OH (2: 1) extracts lipid-linked monosaccharides and smaller-sized oligosaccharides (i.e. 1 - 5 sugars), whereas sub- sequent extraction of the pellets with CHC1,/CH,0H/H20 (10: 10: 3 ) extracts larger-sized lipid-linked oligosaccharides. The radioactivity incorporated into these two lipid extractions at various times of incubation is compared in wild-type cells and in resistant cells (b) and (c) as shown in Fig. 10. It can be seen in Fig. 10A that the incorporation of mannose into the lipid- linked smaller-sized oligosaccharides by the wild-type cells increased in a linear fashion for about 20 min and then declined, whereas the incorporation of mannose into the larger-sized oli- gosaccharides (Fig. 10B) became measurable after about 10 min of incubation and then continued to increase dramatically over the next 40 min. These data suggest a precursor/product relation- ship whereby radioactivity in the smaller oligosaccharides is transferred to the larger ones.
20 B
0 10 20 30 40 50 Time of Incubation (min.)
0 10 20 30 40 50 Time of Incubation (min.)
Fig. 10. Comparison of [2-'H]mannose incorporation into lipid-linked monosaccharides and oligosaccharides in wild-type and tunicamycin- resistant soybean cells. (A) Cells were incubated for 10-50 min in ['Hlmannose and were then extracted with CHCl,/CH,OH (2: 1) to obtain Dol-P-monosaccharides and Dol-PP-oligosaccharides with short oligosaccharide chains. (B) The residue from the extraction in A was then extracted with CHCl,/CH,OH/H,O ( I 0 : 10: 3) to isolate lipid-linked oligosaccharides with longer oligosaccharide chains. Aliquots of the lipid extracts from each time were taken to determine their radioactive content. Curves are as follows: wild-type cells (a) (0-O), resistant cells (b) (A-A), resistant cells (c) (D-D).
Zeng and Elbein (Eur: J . Biochern. 233) 465
" I .I 0 1 mM 2 t + t
0 5 10 15 20 25 Fraction Number
0 5 10 15 20 25 Fraction Number
0 1 mM % I ,.- t t
C
2 50 t t
0 5 10 15 20 25 Fraction Number
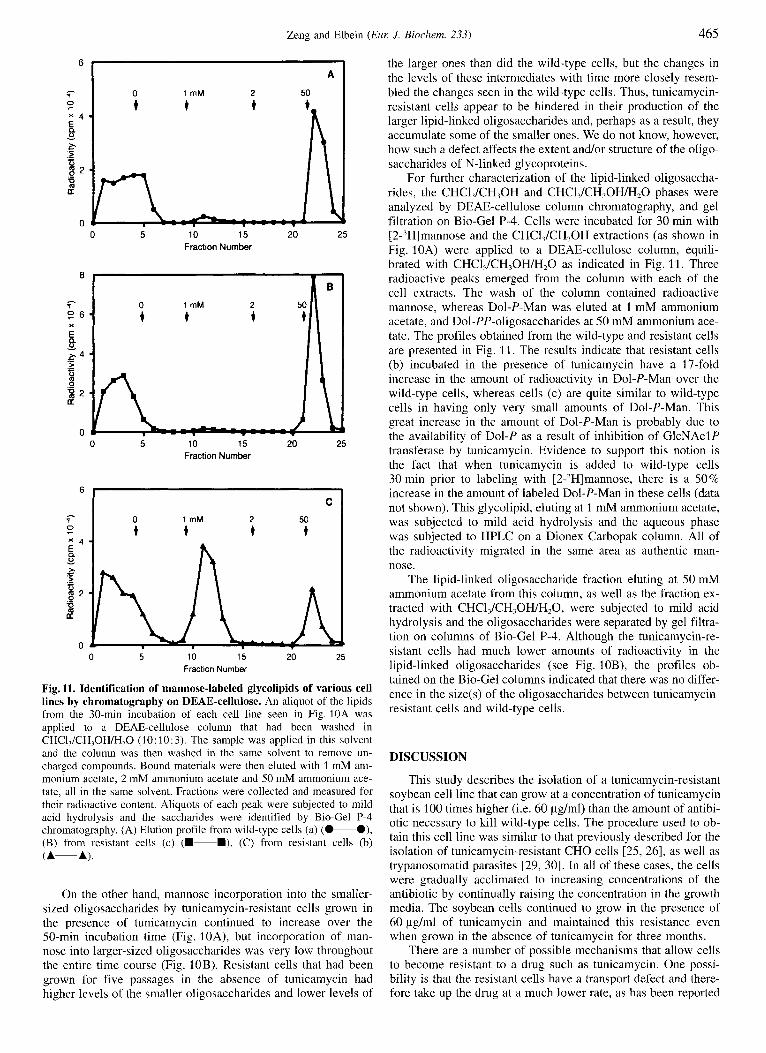
Fig. 11. Identification of mannose-labeled glycolipids of various cell lines by chromatography on DEAE-cellulose. An aliquot of the lipids from the 30-min incubation of each cell line seen in Fig. 10A was applied to a DEAE-cellulose column that had been washed in CHCI,/CH,OH/H,O (10: 10:3). The sample was applied in this solvent and the column was then washed in the same solvent to remove un- charged compounds. Bound materials were then eluted with 1 mM am- monium acetate, 2 mM ammonium acetate and 50 mM ammonium ace- tate, all in the same solvent. Fractions were collected and measured for their radioactive content. Aliquots of each peak were subjected to mild acid hydrolysis and the saccharides were identified by Bio-Gel P-4 chromatography. (A) Elution profile from wild-type cells (a) (0-O), (B) from resistant cells (c) (m-B), (C) from resistant cells (b) (A-A).
On the other hand, mannose incorporation into the smaller- sized oligosaccharides by tunicamycin-resistant cells grown in the presence of tunicamycin continued to increase over the 50-min incubation time (Fig. 1 OA), but incorporation of man- nose into larger-sized oligosaccharides was very low throughout the entire time course (Fig. 10B). Resistant cells that had been grown for five passages in the absence of tunicamycin had higher levels of the smaller oligosaccharides and lower levels of
the larger ones than did the wild-type cells, but the changes in the levels of these intermediates with time more closely resem- bled the changes seen in the wild-type cells. Thus, tunicamycin- resistant cells appear to be hindered in their production of the larger lipid-linked oligosaccharides and, perhaps as a result, they accumulate some of the smaller ones. We do not know, however, how such a defect affects the extent and/or structure of the oligo- saccharides of N-linked glycoproteins.
For further characterization of the lipid-linked oligosaccha- rides, the CHCI,/CH,OH and CHCI,/CH,OH/H,O phases were analyzed by DEAE-cellulose column chromatography, and gel filtration on Bio-Gel P-4. Cells were incubated for 30 min with [2-'Hlmannose and the CHCl,/CH,OH extractions (as shown in Fig. 10A) were applied to a DEAE-cellulose column, equili- brated with CHCl,/CH,OH/H,O as indicated in Fig. 11. Three radioactive peaks emerged from the column with each of the cell extracts. The wash of the column contained radioactive mannose, whereas Dol-P-Man was eluted at 1 mM ammonium acetate, and Dol-PP-oligosaccharides at 50 mM ammonium ace- tate. The profiles obtained from the wild-type and resistant cells are presented in Fig. 11. The results indicate that resistant cells (b) incubated in the presence of tunicamycin have a 17-fold increase in the amount of radioactivity in Dol-P-Man over the wild-type cells, whereas cells (c) are quite similar to wild-type cells in having only very small amounts of Dol-P-Man. This great increase in the amount of Dol-P-Man is probably due to the availability of Dol-P as a result of inhibition of GlcNAclP transferase by tunicamycin. Evidence to support this notion is the fact that when tunicamycin is added to wild-type cells 30 min prior to labeling with [2-,H]mannose, there is a 50% increase in the amount of labeled Dol-P-Man in these cells (data not shown). This glycolipid, eluting at 1 mM ammonium acetate, was subjected to mild acid hydrolysis and the aqueous phase was subjected to HPLC on a Dionex Carbopak column. All of the radioactivity migrated in the same area as authentic man- nose.
The lipid-linked oligosaccharide fraction eluting at 50 mM ammonium acetate from this column, as well as the fraction ex- tracted with CHCl,/CH,OH/H,O, were subjected to mild acid hydrolysis and the oligosaccharides were separated by gel filtra- tion on columns of Bio-Gel P-4. Although the tunicamycin-re- sistant cells had much lower amounts of radioactivity in the lipid-linked oligosaccharides (see Fig. 10B), the profiles ob- tained on the Bio-Gel columns indicated that there was no differ- ence in the size(s) of the oligosaccharides between tunicamycin- resistant cells and wild-type cells.
DISCUSSION
This study describes the isolation of a tunicamycin-resistant soybean cell line that can grow at a concentration of tunicamycin that is 100 times higher (i.e. 60 &ml) than the amount of antibi- otic necessary to kill wild-type cells. The procedure used to ob- tain this cell line was similar to that previously described for the isolation of tunicamycin-resistant CHO cells [25, 261, as well as trypanosomatid parasites [29, 301. In all of these cases, the cells were gradually acclimated to increasing concentrations of the antibiotic by continually raising the concentration in the growth media. The soybean cells continued to grow in the presence of 60 pg/ml of tunicamycin and maintained this resistance even when grown in the absence of tunicamycin for three months.
There are a number of possible mechanisms that allow cells to become resistant to a drug such as tunicamycin. One possi- bility is that the resistant cells have a transport defect and there- fore take up the drug at a much lower rate, as has been reported

466 Zeng and Elbein (Eur: .I. Bioclzem. 233)
for some tunicamycin-resistant CHO cells [39, 401. Another pos- sibility is that the target enzyme (i.e. GlcNAclP transferase) itself becomes resistant to the inhibitory effects of the drug. In both of these cases. the level of enzymic activity of GlcNAclP transferase would not change. However, in the soybean cells, as well as the CHO cells [25, 261 and trypanosomatid parasites [29, 301, the activity of GlcNAclP transferase in the tunicamycin- resistant cells is increased about 40-fold over the wild-type cells. This increase in enzymic activity in the soybean cells correlated with a great increase in the amount of a 39-kDa protein. Al- though the full-length cDNA from the CHO cells codes for a protein of 408 amino acids and therefore should hava a molecu- lar mass of about 46 kDa, we believe that the protein overex- pressed in soybean cells and seen on the SDS gels is the GlcNAclP transferase. In fact, antibody against the tuni- camycin-resistant gene product of CHO cells reacts with a 36- 37-kDa band on Western blots [38]. Future studies will involve preparation of polyclonal antibodies against this soybean protein to prove that it is the real GlcNAclP transferase, and then the determination of amino acid sequence in order to isolate the mRNA and cDNA.
In contrast to observations in at least one of the tunicamycin- resistant CHO cell lines where four or five different proteins were amplified [25], in soybean cells only one 39-kDa protein was increased in sufficient amounts to be detected on the SDS gels. However, the tunicamycin-resistant soybean cells also show a significant and repeatable two-fourfold increase in the Dol-P-Man synthase activity, especially in those cells grown in the presence of tunicamycin. One might expect that this increased activity was due to the increased availability of Dol-P because of the inhibition of the GlcNAclP transferase. How- ever, if this were the case, one should also expect an increase in the Dol-P-Glc synthase activity. Such an increase is not ob- served.
At this stage, we do not have an explanation for this increase in activity of the Dol-P-Man synthase. However. when the tuni- camycin-resistant soybean cells are labeled with [2-'H]mannose, there is a dramatic increase in the production or accumulation of Dol-P-Man in the cells as compared to wild-type cells. In addition, the resistant cells have significantly lower amounts of the lipid-linked oligosaccharides with longer-sized oligosaccha- ride chains. One probable explanation for these results is that the GlcNAclP transferase activity is strongly inhibited in tuni- camycin-resistant cells grown in the presence of tunicamycin even though the enzyme is greatly amplified in these cells. Thus, Dol-P is not utilized by this enzyme and is available for the formation of Dol-P-Man and Dol-P-Glc. However, lack of GlcNAc-PP-Do1 formation results in a deficiency of the accep- tor for the addition of GlcNAc, mannose and glucose to produce the Glc,Man,(GlcNAc),-PP-Dol. Thus the larger-sized lipid- linked oligosaccharides are produced at a much reduced rate as compared to control cells. and the Dol-P-monosaccharides accu- mulate since there is not enough acceptor available for these sugars
of Health (DK 21800). This research was supported by a grant from the National Institutes
REFERENCES I . Lehrman, M. A. (1991) Glycohiology I , 553-562. 2. Waechter, C. J. & Lennarz, W. J. (1976) Annu. Rev. Biochern. 45,
95-112.
3. Elbein, A. D. (1979) Annu. Rev. Plant Physiol. 30, 239-260. 4. Hubbard, S. C. & Ivatt, R. J. (1981) Annu. Rev. Bicrclzeni. 50, 555-
5 . Kornfeld, R. & Kornfeld, S. (1985) Annu. Rev. Bi<JCkem. 54, 631 -
6. Hirschberg, C. B. & Snider, M. D. (1987) Annu. Rev. Biochern. 56,
7. Chapman, A.. Fujimoto, K. & Kornfeld, S. (1980) J . B i d . Chrrn.
8. Kang, M. S., Spencer, J. P. & Elbein, A. D. (1978) J. B i d . Chenz.
9. Schutzbach, J. S.. Springfield, J. D. & Jensen, J. W. (1980) J. Biol.
10. Behrens. N. H., Parodi, A. J. & Leloir, L. E (1971) Proc. Narl A c d
21. D'Souza, C. M., Sharma, C. B. & Elbein, A. D. (1992) J . Biol.
12. Kelleher, D. J., Krebach, G. & Gilmore, R. (1992) Cell 69, 55-65, 13. Lennarz, W. J. (1987) Biocheniisrry 26, 7205-7210. 14. Keller, R. K.. Boon, D. Y. & Cruni, F. C. (1979) Biochemi.str?; 18,
15. Kaushal, G. P. & Elbein, A. D. (1985) J . B i d Chern. 260. 16303-
16. Plouhar, P. L. & Bretthauer, R. K. (1983) J . Bid . Chem. 258,
17. Shailubhai, K., Dong-Yu, B., Saxena, E. S. & Vijay, I. K. (1988) J.
18. Kean, E. L. (1982) J . Riol. Chem. 257, 7952-7954. 19. Kean, E. L. (1985) J. Biol. Chenz. 260, 12561 - 12571. 20. Elbein, A . D. (1987) Annii. Rev. Biocliern. 56, 497-534. 21. Tkacz, J. S. & Lampen, J . B. (1975) Biochem. Biophys. Res. Con/-
22. Heifetz, A,, Keenan, R. W. & Elbein, A. D. (1979) Biochemisfn 18,
23. Struck, D. K. & Lennarz, W. J . (1977) 1. Biol. Cheni. 252, 1007-
24. Kaushal, G . P. & Elbein, A. D. (1986) Plrmt Physiol. 81, 1086-
25. Waldman, B. C., Oliver, C. & Krag, S. S. (1987) J. Cell. Physiol.
26. Lehrman, M. A,, Zhu, X. & Khounlo, S. (1988) J . Biol. Chem. 263,
27. Zhu, X. & Lehrman, M. A. (1990) J. Biol. Chem. 265, 14250-
28. Scocca, J. R. & Krag, S. S. (1990) J. Biol. Cheni. 265, 20621 -
29. Kink, J. A. & Chang, K.-P. (1987) Proc. Nut1 Acad. Sci. USA 84,
30. Liu, X. & Chang, K.-P. (1992) Mol. Cell. R i d . 12, 4112-4122. 31. Rine, J., Hansen, W., Hardeman. E. & Davi5, R. (1983) Proc. Narl
32. Kaushal, G. P. & Elbein, A. D. (1987) Biochernistn 26. 7953-
33. Pan. Y. T. & Elbein, A. D. (1990) Biochemistry 29, 8077-8084. 34. Drake, R. R., Kaushal, G. P., Pastuszak, I. & Elbein, A. D. (1991)
35. Szumilo, T., Kaushal, G. P. & Elbein, A. D. (1986) Arch. Biochern.
36. Kaushal, G. P., Zeng, Y.-C. & Elbein, A. D. (1993) 1. B i d . Chern.
37. Bradford, M. M. (1976) And. Biochenz. 14, 248-254. 38. Zhu, X., Zeng, Y. & Lehrman. M. A. (1992) J. Biol. Cheni. 267.
39. Sudo, T. & Onodera, K. (1979) J. Cdl. Physiol. 101, 149-156. 40. Kuwano, M., Tabuki, T., Akiyama, S., Mifune, K., Tainura. C. &
583.
664.
63-87.
255, 4441 -4446.
253, 8860-8868.
Chem. 255, 4170-4175.
Sci. USA 68. 2857-2860.
Chem. 268, 4720-4727.
3946- 3952.
16 309.
12988- 12993.
Biol. Chem. 263, 15 964- 15 972.
niun. 65, 248-257.
21 86-2192.
1013.
1091.
131, 302 - 3 17.
19 796- 19 803.
14255.
20 626.
1253 - 1257.
Acad. Sci. USA 80, 6750-6754.
7960.
Plant Physiol. 97, 396-401.
Biophys. 247, 261 - 27 1 .
268, 14536- 14542.
8895-8902.
Tkehara, Y. (1981) Somatic Cell Genet. 7. 507-521.





























