Embed Size (px)
Citation preview

242 W. Hocker, K . Colln und E . Amtmann
Summary
To eliminate the inherent uncertainty in conveying results gained from laboratory animals onto livestock, it was attempted to determine the biological value of proteins on chicken istead of rats. I t was possible to apply a rat standard-method on chicken without additionll expenditure, thus proving the feasibility of determining the biolo- gical value of prsteins for poultry on this species itself.
Literatur
1. Arbeitskreis fur Proteinbewertung der Gesellsdlaft fur Ernahrungsphysiologie der Haus- tiere, 1964: Vorschrift zur Proteinbewertung in Versudlen an wachsenden Ratten, Z. Tier- physiol., Tierernahrg. u. Futtermittelkde. 19, 305.
Anschrift der Autoren: A 1030 Wien, Linke Bahngasse 11 , Usterreich
Aus dem Zoologisnben Institut der Universitat Koln - Direktor: Prof. Dr. Egelbaaf - und dem Anatomischen Institut der Uniuersitat Koln - Direktor: Prof . Dr. Ortmann
Ober Veranderungen der Serumproteine und Serumphosphatasen durch Hypophysektomie
und reduzierte Ernahrung bei Tauben
Von W. HOCKER, K. COLLN und E. AMTMANN I
Einleitung
Im Rahmen von Untersuchungen iiber die hornionale Kontrolle verschiedener ut- bestandteile sol1 im folgenden der EinfluB der Hypophysektomie auf die Serumpro- teine und Serumphosphatasen gepriifi werden. Wahrend zahlreiche, teilweise aller- dings widerspriichliche Befunde an Saugetieren zu diesem Fragenkomplex bereits vor- liegen, sind unsere Kenntnisse uber die hormonalen Wirkungen bei Vogeln relativ gering.
Bei Tauben wird nach Hypophysektomie die Nahrungsaufnahme stark reduziert; damit geht ein Gewichtsabfall einher, der oft 2 5 V 0 und mehr betriigt, bis im Laufe von vier bis acht Wochen p. op. zumeist ein etwa stationarer Zustand erreicht wird. Eine erste Analyse der vielschichtigen Gesamtwirkung der Hypophyseninsuffizienz bei Vogeln zielt darauf ab, den nutritiven Faktor in diesem Zusammenhang auf seine Bedeutung zu prufen.
Nach Untersuchungen an Menschen, Saugetieren und Amphibien ergeben sich bei Hypophyseninsuffizienz beziiglich der Serum- bzw. Plasmaproteine heterogene Be-
* Fiir die sorgfaltige Erledigung der technischen Arbeiten danken wir Fraulein H. KLEI.

Uber Veranderwngen der Serumproteine und Serumphosphatasen 243
funde: Keine Veranderungen des Gesamtproteins lie8en sich beim Menschen (45, 8) und bei Ratten (6) nachweisen, bei Pavianen und Hunden wurde eine geringe Sen- kung festgestellt (5, 29, 4); GOLDBERC (16) hatte beim H u n d uber der Norm liegende Werte ermittelt; derselbe Autor fand bei Kroten (Bufo arenarum) einen starken Ruckgang.
Bei Inanition ist nach einer gewissen Zeit mit einer Hypoproteinamie zu rechnen (1, 3, 15). Eine Verminderung der Nahrungsaufnahme, wie sie nach Hypophysektomie gegeben ist, fuhrt bei Ratten zu einer signifikanten Senkung des Gesamtproteins UIU
O,2 o/o (29). Sofern sich herausstellen sollte, d a 8 bei Tauben sowohl die Reduzieruiig der Nahrung als auch die Hypophysektomie ebenfalls zu einer Verminderung des Gesamtproteins fuhren, interessiert vor allem, in welchern Umfang das auftretende Defizit im GesamteiweiB auf Nahrungsfaktoren zuriickgefuhrt werden kann.
Mit Hilfe der Elektrophorese soll sodann geklart werden, wie sich die einzelnen Proteinfraktionen verhalten. Untersuchungen an Saugetieren lassen vermuten, da8 sich sowohl nach Hypophysektomie, moglicherweise aber auch bei Mangel- oder Unterernahrung das EiweiBspektrum verandert (5, 31, 58).
SchlieRlich soll der Einflu8 der Hypophysektomie und des partiellen Nahrungs- entzuges auf einzelne Proteine bzw. Proteingruppen des Serums gepruft werden. Aus den zur Wahl stehenden Fermentproteinen wurden die Serumphosphatasen ausge- wahlt, weil auf Grund der bereits vorliegenden Untersuchungen damit gerechnet werden kann, daR deren Aktivitat sowohl hormonalen als auch nutritiven Einflussen unterliegt (52, 20, 49). Allerdings sind auch hier die Befunde teilweise divergierend: So wurde einerseits bei Ratten eine Aktivitatsverminderung der alkalischen Serum- phosphatase nach Hypophysektomie festgestellt, andererseits eine Steigerung (30, 21). Befunde iiber das Verhalten der sauren Serumphosphatase von Vogeln unter ahnlichen Versuchsbedingungen liegen uns nicht vor.
Material und Methode
Als Versuchstiere dienten mannliche und weibliche adulte Brieftauben (Columba livia), die eine Woche vor Versuchsbeginn in Einzelkafigen untergebracht wurden. Die Zahl der Ver- suchstiere ist aus den Tabellen zu entnehmen. Die Gewinnung der Blutproben erfolgte durch Punktion der Flugelvene. Unmittelbar nach Gerinnung wurde das Blur zentrifugiert. Die Fermentaktivitat der Seren wurde am gleichen Tag gemessen, die Proben zur Proteinbestim- mung wurden zurneist bei -20” C eingefroren und spater aufgearbeitet.
Die Tauben wurden in drei Gruppen eingeteilt: 1. UnbeeinfluiSte Kontrolltiere: Futter und Wasser ad lib., mindestens eine Woche vor
Blutentnahme gekafigt. 2. Hungertiere: Das Futter wurde individuell so dosierc, dai3 ein weitgehend kontinuier-
licher Gewichtsverlust von ca. 4-5 g/Tag im Laufe von drei bis vier Wochen zu einer Ab- nahme des Kor ergewichtes von etwa 22 O/o fuhrte - entsprechend dem Gewichtsverlauf bei den h ypophyseitomierten Tieren der Gruppe 3. Das Gewichtsniveau wurde noch etwa zwei Wochen konstant gehalten, ehe die Blutentnahme erfolgte. - Erganzend wurde der Protein- gehalt des Serums von zwolf Tauben bei jeweils denselben Tieren vor, wahrend und nach dem Hungerversuch gemessen.
3. Hypophysektomierte Tiere: Operationsmethode: parapharyngeal in Nembutalnarkose, Prufung der Vollstandigkeit der Operation durch stereomikrosko ische Inspektion der Sella turcica und Kontrolle des Kor ergewichtsverlaufs. Im Zweifelsill wurden die Tiere aus- geschieden. Die Blutentnahme erfolgte sechs bis acht Wochen p. op.
Bestimmung der Serumproteine (Standard: Rinderserumalbumin) nach LOWRY et al. (34). Die elektrophoretische Auftrennung erfolgte auf Cellogelstreifen (Serva) mit dem Mikro- zonenelektrophoresesystem (Beckman, Model1 R-loo), Barbituratpuffer pH 8,6; Ionenstarke

W. Hdcker, K. Colln und E . Amtmann
v v u
i
- 1
I U
0,075, aufgetragene Serummenge etwa lpl, Trennzeit 60 Minuten, 0,9 rnA, konstant pro crn Streifenbreite, Farbung: Ponceau S, quantitative Auswertung rnit Ana- Iytrol (Beckman), Filter 520 mu. - Auf eine Korrektur konnte bei der Berechnung der einzelnen Proteinfrak- tionen verzichtet werden (40).
Photometrische Bestimmung der alkalischen Serum- phosphataseaktivitat nach RICK und HAUSAMEN (38) und der sauren Serumphosphataseaktivitat nach ANDERSCH und SZCYPINSKI (2); alkalische Phosphatase: mU (25' CI ml), saure Phosphatase: mU (37'Ciml) (1 IU ist die Enzymmenge, die 1 pMol Substrat in 1 Minute bei Stan- dardtemperatur umsetzt).
Fur die Bezeichnung der Serurnproteinfraktionen wurden in Obereinstimmung rnit anderen Untersuchern von Vogelseren (18, 37, 9) Nummern statt der ublichen Termini gewahlt. In diesem Zusarnrnenhang ware auch auf die Bedenken hinzuweisen, die aus der Humanmedi- zin entlehnten Bezeichnungen ohne weiteres auf Tiere zu ubertragen (53). Auflerdem sind die Angaben in der Literatur uber die Anzahl der Serumproteinfraktionen bei Tauben nicht einheitlich (53, 13, 12). Wahrscheinlich liegen folgende Entspredlungen vor: Fraktion I = Prf- albumin, Fraktion I1 = Albumin, Fraktion 111 = aG10- buline, Fraktion IV = P-Globuline, Fraktion V = 7- Globuline.
Ergebnisse
1. Serumproteine
D e r Proteingehalt des Serums unbeeinflufiter Tau- ben betragt im Mittel 4,27 g/100 ml (Tabelle 1). Bei hypophysektornierten Tieren liegt der Gesamtpro- teingehalt u m durchschnittlich 1,71 g/100 ml unter dem der Kontrolltiere. Die Hungert iere zeigen im Mittel einen u m 1,02 g/100 ml verminderten Pro- teingehalt des Serums.
Bevor diese Diff erenzen auf Bedeutsamkeit gepriift werden konnen, ist die Homogenitat der Streuungen der Versuchsgruppen nachzuweisen. Bartletts Homogenitats- test ergab ein ~ 2 ~ ~ 1 = 0,89 mit einem P > 0,05. Die drei Varianzen konnen darnit als homogen angesehen werden. Zur Prufung der Bedeutsamkeit der aufgezeigten Mittel- wertdifferenzen wurde daraufhin eine Varianzanalyse durchgefuhrt. Fur das Varianzverhaltnis der Streuungen zwisdien den Gruppen und der Reststreuung ergab sich ein F = 39,3 mit den Freiheitsgraden n1 = 2 und n, = 27 und einern P < 0,Ol.
Die Unterschiede in den mittleren Gesamtpro- teinrnengen in den drei Versuchsgruppen sind somit signifikant. Hypophysektomie fuh r t also zu einer statistisch gesicherten Verminderung des Gesamt- proteins i m Serum (Prufung durch Vergleich der Mittelwertdifferenzen mittels t-Test). Eine an-
Ober Veranderungen der Serumproteine und Serumphosphatasen 245
nahernd gleich starke Abnahme wird auch bei Hungertieren beobachtet. Es liegt daher der SchluB nahe, daB mindestens ein Teil der komplexen Wirkung der Hypophysen- insuffizienz au fdie verminderte Nahrungsaufnahme zuriidrzufiihren ist.
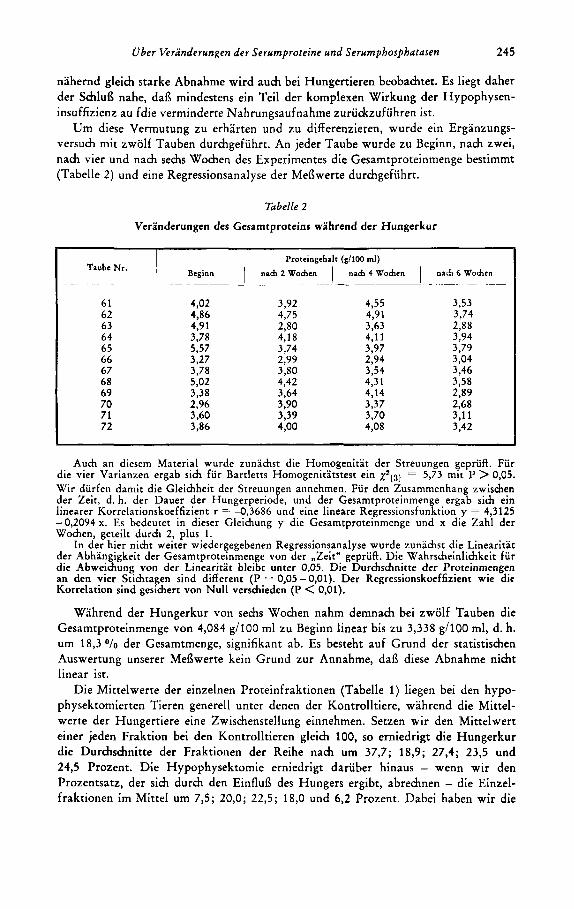
Um diese Vermutung zu erharten und zu differenzieren, wurde ein Erganzungs- versuch mit zwolf Tauben durchgefiihrt. An jeder Taube wurde zu Beginn, nach zwei, nach vier und nach sechs Wochen des Experimentes die Gesamtproteinmenge bestimmt (Tabelle 2) und eine Regressionsanalyse der MeBwerte durchgefiihrt.
Tabelle 2
Veranderungen des Gesamtproteins wahrend der Hungerkur
Taubc Nr. -
61 62 63 64 65 66 67 68 69 70 71 72
I I 1 Beginn ._
4,02 4,86 4,9 1 3,78 5,57 3,27 3,78 5,02 3,38 2,96 3,60 3,86
Proteingehrlt (g1100 ml)
na& 2 Wo&en I n a k 4 W o m e n
3,92 4,55 4,75 4,9 1 2,80 3,63 4,18 4 , l l 3,74 3,97 2,99 2,94 3,80 3,54 4,42 4,31 3,64 4,14 3,90 3,37 3,39 3,70 4,OO 4,08
I . - na& 6 Wo&en
3,53 3,74 2,88 3,94 3,79 3,04 3,46 3,58 2,89 2,68 3 , l l 3,42
Auch an diesern Material wurde zunachst die Hornogenitat der Streuungen gepruff. Fur die vier Varianzen ergab sich fur Bartletts Hornogenitatstest ein ~ ~ 1 3 1 5,73 rnit P > 0,05. Wir durfen darnit die Gleichheit der Streuungen annehrnen. Fur den Zusarnrnenhang zwischen der Zeit, d. h. der Dauer der Hungerperiode, und der Gesarntproteinrnenge ergab sich ein h e a r e r Korrelationskoeffizient r = -0,3686 und eine lineare Regressionsfunktion y = 4,3125 - 0,2094 x. Es bedeutet in dieser Gleichung y die Gesarntproteinrnenge und x die Zahl der Wochen, geteilt durch 2, plus.1.
In der hier nicht weiter wiedergegebenen Regressionsanalyse wurde zunachst die Linearitat der Abhangigkeit der Gesarntproteinrnenge von der ,Zeit" gepriifi. Die Wahrscheinlichkeit fur die Abweichung von der Linearitat bleibt unter 0.05. Die Durchschnitte der Proteinmengen an den vier Stichtagen sind different (P = 0,05 - 0,Ol). Der Regressionskoeffizient wie die Korrelation sind gesichert von Null versrhieden (P < 0,Ol).
Wahrend der Hungerkur von sechs Wochen nahm demnach bei zwolf Tauben die Gesamtproteinmenge von 4,084 g/100 ml zu Beginn linear bis zu 3,338 g/lOO ml, d. h. um 18,30/0 der Gesamtmenge, signifikant ab. Es besteht auf Grund der statistischen Auswertung unserer MeBwerte kein Grund zur Annahme, daB diese Abnahme nicht linear ist.
Die Mittelwerte der einzelnen Proteinfraktionen (Tabelle 1) liegen bei den hypo- physektomierten Tieren generell unter denen der Kontrolltiere, wahrend die Mittel- wert:e der Hungertiere eine Zwischenstellung einnehmen. Setzen wir den Mittelwert einer jeden Fraktion bei den Kontrolltieren gleich 100, so erniedrigt die Hungerkur die Durchschnitte der Fraktionen der Reihe nach um 37,7; 18,9; 27,4; 23,5 und 24,5 Prozent. Die Hypophysektomie erniedrigt dariiber hinaus - wenn wir den Prozentsatz, der s i h durch den EinfluB des Hungers ergibt, abrechnen - die Einzel- fraktionen im Mittel um 7,5; 20,O; 22,5; 18,O und 6,2 Prozent. Dabei haben wir die
246 W. Hocker, K . Colln und E. Amtmann
Werte beiden
Die
aus unserem Hungerversuch zugrunde gelegt, unter der Annahme, dafl in Gruppen die verminderte Nahrungsaufnahme zu dem gleichen Effekt fiihrt. Hypophysektomie fiihrt demnach besonders bei den Fraktionen I und V
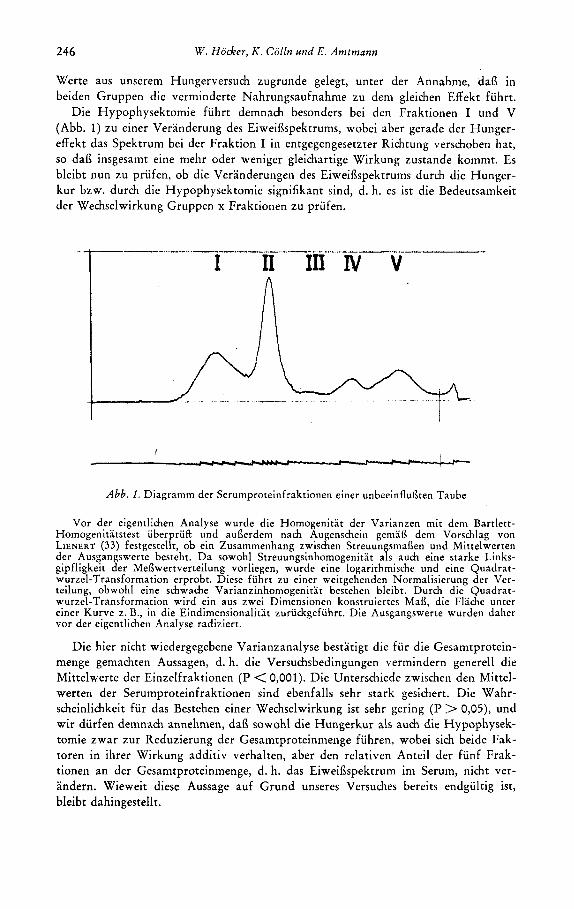
(Abb. 1) zu einer Veranderung des Eiweiflspektrums, wobei aber gerade der Hunger- effekt das Spektrum bei der Fraktion I in entgegengesetzter Richtung verschoben hat, so dai3 insgesamt eine mehr oder weniger gleichartige Wirkung zustande kommt. Es bleibt nun zu priifen, ob die Veranderungen des Eiweiflspektrums durch die Hunger- kur bzw. durch die Hypophysektomie signifikant sind, d. h. es ist die Bedeutsamkeit der Wechselwirkung Gruppen x Fraktionen zu priifen.
Abb. 1. Diagramm der Serumproteinfraktionen einer unbeeinflunten Taube
Vor der eigentlichen Analyse wurde die Homogenitat der Varianzen mit dem Bartlett- Homogenitatstest uberpruft und auBerdem nach Augenschein gemal3 dem Vorschlag von LIENERT (33) festgestellt, ob ein Zusammenhang zwischen StreuungsmaBen und Mittelwerten der Ausgangswerte besteht. D a sowohl Streuungsinhomogenitat als auch eine starke Links- gipfligkeit der MeBwertverteilung vorliegen, wurde eine logarithmische und eine Quadrat- wurzel-Transformation erprobt. Diese fiihrt zu einer weitgehenden Normalisierung der Ver- teilung, obwohl eine schwache Varianzinhomogenitat bestehen bleibt. Durch die Quadrat- wurzel-Transformation wird ein aus zwei Dimensionen konstruiertes MaB, die FlPche unter einer Kurve z. B., in die Eindimensionalitat zuriickgefuhrt. Die Ausgangswerte wurden daher vor der eigentlichen Analyse radiziert.
Die hier nicht wiedergegebene Varianzanalyse bestatigt die fur die Gesamtprotein- menge gemachten Aussagen, d. h. die Versuchsbedingungen vermindern generell die Mittelwerte der Einzelfraktionen (P < 0,001). Die Unterschiede zwischen den Mittel- werten der Serumproteinfraktionen sind ebenfalls sehr stark gesichert. Die Wahr- scheinlichkeit fur das Bestehen einer Wechselwirkung ist sehr gering (P > 0,05), und wir diirfen demnach annehmen, dai3 sowohl die Hungerkur als auch die Hypophysek- tomie zwar zur Reduzierung der Gesamtproteinmenge fiihren, wobei sich beide Fak- toren in ihrer Wirkung additiv verhalten, aber den relativen Anteil der fiinf Frak- tionen an der Gesamtproteinmenge, d. h. das Eiweiflspektrum im Serum, nicht ver- andern. Wieweit diese Aussage auf Grund unseres Versuches bereits endgiiltig ist, bleibt dahingestellt.
Uber Veranderungen der Serumproteine und Serumphosphatasen 247
! Phorphatane I Pararnrter ' Kontrolltiere
In zwei Einzelanalysen der Mef3werte der Kontrolltiere und der hypophysektomierten Tiere einerseits und der Hungertiere und der hypophysektornierten Tiere andererseits ergab sich in der ersten Analyse eine signifikante, in der zweiten eine ungesicherte Wechselwirkung Gruppen x Fraktionen. Das deutet darauf hin, da13 moglicherweise die Hypophysektomie zu einer Verschiebung des Eiweifispektrums fiihrt. Dieser Frage sol1 in weiteren Untersuchungen nachgegangen werden.
Der "Albumin-Globulin-Quotient" betragt bei unbeeinfluflten Kontrolltieren im Mittel 2.08, bei hypophysektornierten Tauben 2,04 und bei Hungertieren 2,22. Fur den Zusamrnen- hang zwischen Albuminfraktionen und Globulinfraktionen ergaben sich in der Reihenfolge der Eiweiflquotienten die folgenden Regressions- bzw. Korrelationskoeffizienten -0,61; +0,34; und +0,82 bzw. -0,48; +0,24 und 0,15. Alle diese Werte sind nicht von Null vershieden, und wie eine Covarianzanalyse uberdies zeigte, auch untereinander nicht different. Wir diirfen damit annehmen, dafi die Unterschiede in den Albumin-Globulin-Quotienten unbedeutsarn sind.
1 Hungertiere I
Hypoph yrekrornierte Ticre
2. Serumphosphatasen
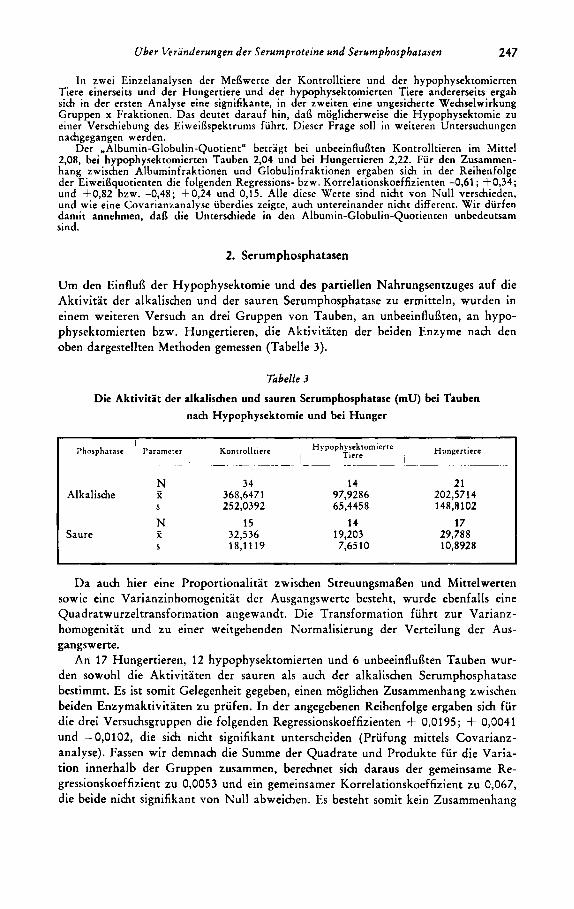
Um den Einflufl der Hypophysektomie und des partiellen Nahrungsentzuges auf die Aktivitat der alkalischen und der sauren Serumphosphatase zu ermitteln, wurden in einem weiteren Versuch an drei Gruppen von Tauben, a n unbeeinfluflten, an hypo- physektomierten bzw. Hungertieren, die Aktivitaten der beiden Enzyme nach den oben dargestellten Methoden gemessen (Tabelle 3).
Tabelle 3
Die Aktivitat der alkalischen und sauren Serumphosphatase (mu) bei Tauben nach Hypophysektomie und bei Hunger
N 34 14 - Alkalische x 368,6471 97,9286 S 252,0392 65,4458
N 15 14 Saure 2 32,536 19,203
S 18,1119 7,65 10
21 202,5714 148,8102
17 29,788 10,8928
Da auch hier eine Proportionalitat twischen Streuungsmaflen und Mittelwerten sowie eine Varianzinhomogenitat der Ausgangswerte besteht, wurde ebenfalls eine Quadratwurzeltransformation angewandt. Die Transformation fuhrt zur Varianz- homogenitat und zu einer weitgehenden Normalisierung der Verteilung der Aus- gangswerte.
An 17 Hungertieren, 12 hypophysektomierten und 6 unbeeinfluflten Tauben wur- den sowohl die Aktivitaten der sauren als auch der alkalischen Serumphosphatase bestimmt. Es ist somit Gelegenheit gegeben, einen moglichen Zusammenhang zwischen beiden Enzymaktivitaten zu prufen. In der angegebenen Reihenfolge ergaben sich fur die drei Versuchsgruppen die folgenden Regressionskoeffizienten + 0,0195 ; + 0,0041 und - 0,0102, die sich nicht signifikant unterscheiden (Priifung mittels Covarianz- analyse). Fassen wir demnach die Summe der Quadrate und Produkte fur die Varia- tion innerhalb der Gruppen tusammen, berechnet sich daraus der gemeinsame Re- gressionskoeffizient zu 0,0053 und ein gemeinsamer Korrelationskoeffizient zu 0,067, die beide nicht signifikant von Null abweichen. Es besteht somit kein Zusammenhang

248 W. Hocker, K . Colln und E. Amtmann
zwischen der Aktivitat der alkalischen und der sauren Serumphosphatase. In den folgenden Untersuchungen konnen wir damit beide Phosphatasen getrennt betrachten.
Setzen wir die mittlere Phosphataseaktivitat (untransformierte Werte) bei den Kontrolltieren gleich 100, so hat in unserem Versuch (Tabelle 3) die Hungerkur die Aktivitat im Mittel bei der alkalischen Serumphosphatase um 45, l O/o, bei der sauren um 8,5 O/o gesenkt. Die Hypophysektomie verringert die Aktivitat der alkalischen Phosphatase dariiber hinaus um weitere 28,4 O/O und die saure um 32,50/0. Eine Varianzanalyse zeigt, dafl diese Senkungen in der Enzymaktivitat sowohl bei der alkalischen als auch bei der sauren Phosphatase signifikant sind (P < 0,001 bzw. 0,05-0,Ol). Die Aktivitat der sauren Phosphatase ist aber bei den Hungertieren gegenuber der bei den Kontrolltieren nicht statistisch real erniedrigt. Wir diirfen daher annehmen, dai3 die reduzierte Nahrungsaufnahme bei den hypophysektomier- ten Tieren im Hinblick auf die Aktivitat dieses Enzyms bedeutungslos ist. - Fassen wir die transformierten Werte der Kontrolltiere und der Hungertiere zu einer Stich- probe zusammen und priifen die Differenzen der Mittelwerte zu den hypophysekto- mierten Tieren, ergibt sich eine sehr starke Sicherung fur den Unterschied.
Diskussion
Die von uns ermittelten Gesamtproteinwerte fur zehn unbeeinfluflte Tauben liegen durchschnittlich bei 4,27 g/100 nil; nach Messungen an weiteren 26 Tieren ergab sich ein Gesamtmittel von 4,15 g/100 ml. Entsprechende Angaben in der Literatur (35, 46, 42) divergieren erheblich: 2,32 g/100 ml, 5 , l bzw. 4,42-5,81 g/100 ml. - Durch die Anwendung verschiedener Methoden bei den zitierten Untersuchungen ist ein Ver- gleich der erhaltenen Gesamtproteinwerte problematisch. Dasselbe gilt fur einen Ver- gleich der einzelnen Proteinfraktionen. Wir haben bei der Wahl der elektrophore- tischen Auftrennung Wert darauf gelegt, vergleichbare Fraktionen zu gewinnen, die bei allen Versuchstieren identifizierbar waren. Dafiir muflten wir allerdings auf eine gesteigerte Differenzierung, wie sie besonders mit der Disc-Elektrophorese erreichbar gewesen ware, verzichten.
Bei der Bestimmung sowohl des Gesamtproteins als auch der Eiweiflquotienten der Vogel diirfie das Ausreiflerproblem eine gewisse Rolle spielen, weil besonders bei weiblichen Tieren mit der Eibildung erhebliche Veranderungen verbunden sind (9, 56). O b hierauf einige extreme MeBwerte auch bei unseren Befunden zuriickgefuhrt wer- den konnen, sol1 in weiteren Untersuchungen geklart werden; auch eine Beziehung zu der sben erwahnten Streuungsinhomogenitat ware in diesem Zusammenhang zu be- denken.
Hypophysektomie fuhrt bei gewissen Saugetieren in Obereinstimmung mit unseren Befunden an Tauben zu einer Verminderung des Gesamtproteins. Andererseits ergeben sich aber auch Veranderungen des Eiweiflspektrums. Bei Ratten werden durch Hypo- physektomie nur die Albumine reduziert, wogegen die Globuline sowohl relativ als auch absolut einen Anstieg erfahren (29). Dieses divergierende Verhalten der ein- zelnen Proteinfraktionen findet seinen Niederschlag in den stark erniedrigten Albu- min-Globulin-Quotienten. Ahnliche Ergebnisse wurden auch beim Pavian und Hund erzielt (5, 57). Hinsichtlich der Veranderungen der B-Globuline bei hypophysekto- mierten Ratten ergaben sich entgegengesetzte Befunde (36, 31).

Vber Veranderungen der Serumproteine und Serwmphosphatasen 249
Als Ursache fur die Senkung der Serumproteine ware u. a. an die verringerte Pro- duktion von Nebennierenrinden- und Gonadenhormonen zu denken (43, 28, 23, 44). Allerdings kann ein ACTH-Strei3 auch zur Reduzierung des Gesamtproteins fuhren (17). Vor allem aber scheint bei Tauben, wie wir auf Grund von Vorversuchen an- nehmen, das Fehlen von Prolactin von Bedeutung zu sein. Dieses Hormon stimuliert bei hypophysektomierten Tauben den Appetit, so da8 die Nahrungsaufnahme norma- lisiert wird (39, 19). Daruber hinaus erhohte Prolactin den Serumproteinspiegel um einen so hohen Betrag, dai3 vermutlich nicht nur der Unterernahrungseffekt kompen- siert wird, sondern da8 zusatzlich mit einer Steigerung der Proteinsynthese zu rech- nen ist.
Hungerversuche niit einem Nahrungsentzug, wie er der reduzierten Nahrungs- aufnahme nach Hypophysektomie entspricht, ergaben bezuglich der Serumproteine ebenso divergierende Befunde wie nach Hypophysektomie: Wahrend einerseits (29) abgeschwachte, aber entsprechende Reaktionen wie bei Hypophyseninsuffizienz beob- achtet wurden (geringe Abnahme des Gesamtproteins, starke Senkung der Albumine, Zunahme der Globuline), ergaben andererseits Untersuchungen von WEIMER (58) .- ebenfalls an Ratten - eine Reaktion, die mit unseren Befunden grundsatzlich uberein- stimmt.
Gleichzeitige Hamatokritmessungen berechtigen zu der Annahme, dad die Redu- zierung der Serumproteine nicht auf einer Hydramie beruht: Bei den Hungertieren waren die Hamatokritwerte ebenso wie nach Befunden an Ratten (58) erhoht; die Senkung des Hamatokrits nach Hypophysektomie entspricht dem Ruckgang der Erythropoiese (das zugrunde liegende Zahlenmaterial sol1 nach Abschlui3 der Ver- suche uber Veranderungen des roten Blutbildes demnachst vorgelegt werden).
Hemerkenswert ist die hohe Aktivitat der alkalischen Serumphosphatase bei Tau- ben (369 mU im Mittel), verglichen mit anderen Tieren: Huhn: 170 bzw. 200 m u , Pferd: 135 m u , Rind: 163 m u , Schaf: 267 m u , Schwein: 150 mU und Hund: 85 mU (55, 50, 22). Wenn auch keine eindeutige Erklarung fur diese starke Aktivitlt gegeben werden kann, so scheint doch ein Zusammenhang moglich zu dem ungewohn- lich hohen Gehalt und Umsatz an energiereichen Phosphorverbindungen bei Tauben- erythrocyten (14). Die groaen Streuungen in der Aktivitat der alkalischen Serum- phosphatase bei Saugetieren (51) sind auch bei Huhnern gefunden worden (7, 47) und diirften damit vermutlich auch bei Vogeln typisch fur dieses Enzym sein. Da wir gelegentlich auch bei mannlichen Tauben extrem hohe Werte (uber 800 m u ) erhielten, konnten die Befunde von BELL (7), wonach die Eischalenbildung zu einer starken Er- hohung der Enzymaktivitat fuhrt, nicht als Erklarung herangezogen werden.
Auf die unterschiedlichen Befunde uber das Verhalten der Enzymaktivitat bei hypophysektomierten Ratten wurde bereits einleitend hingewiesen. Verglichen mit den Untersuchungen von LI und Mitarbeitern (30), die ebenfalls eine Senkung der Phosphataseaktivitat feststellten, ist bei Tauben die Reaktion wesentlich starker.
Als Ursachen fur die verminderte Enzymaktivitat durflen Ausfall bzw. Verminde- rung mehrerer Hormone in Frage kommen: Untersuchungen an Huhnern ergaben. da8 Schilddrusen- und Nebennierenrindenhormone die Fermentaktivitat positiv beein- flussen (55, 48); Oestrogene haben nur bei Hennen eine stimulierende Wirkung (50, 54). Ob die fehlende Produktion von Somatotropin nicht nur bei Ratten (32), son- dern auch bei Vogeln von Bedeutung ist, bleibt zu prufen. Ferner deuten einige Vor- versuche mit Prolactin darauf hin, dad auch dieses Hormon eine Wirkung auf die

250 W. Hockey, K. Co11n und E . Amtmann
Fermentaktivitat besitzt. Beim Menschen lassen sich die erniedrigten Phosphatasewerte bei hypophysaren Zwergen durch Gaben von Hypophysenextrakten normalisieren (59). Die Befunde an Hungertieren ergeben ein grundsatzlich ubereinstimmendes Bild: Nahrungsniangel senkt die Fermentaktivitat (52, 20, 25).
Im Gegensatz zur alkalischen Serumphosphatase wird die Aktivitat der sauren Serumphosphatase durch Nahrungsentzug nicht beeinfluat. Bei Ratten wurde sogar ein Anstieg der Enzymaktivitat nach Redutierung der Nahrung beobachtet (26). Die Stabilitat dieses Fermentes gegeniiber gewissen experimentellen Einwirkungen, die bei der alkalischen Serumphosphatase zu Veranderungen fiihren, bestatigen Versuche an Kuken und Ratten (27, 11, 24). Nach diesen Befunden ist es nicht iiberraschend, dai3 die Priifung der Korrelation zwischen saurer und alkalischer Serumphosphatase --
ebenso wie bei gewissen Saugetieren (10, 41) - negativ ausfiel.
Zusammenfassung
1. Serumproteine und Serumphosphatasen sind bei hypophysektomierten Tauben signifikant erniedrigt.
2. Reduziert man die Nahrungsaufnahme und damit das Korpergewicht in dem Um- fang, wie man es nach Hypophysektomie beobachtet, wird der Proteingehalt aller Fraktionen vermindert. Die Aktivitat der alkalischen, nicht aber der sauren Serum- phosphatase wird gesenkt. Es besteht keine Korrelation zwischen alkalischer und saurer Serumphosphatase.
3. Eine Veranderung des Eiweiflspektrums bei hypophysektomierten Tieren ist wahr- scheinlich, aber nicht eindeutig gesichert.
4. Die verminderte Nahrungsaufnahme ist eine wesentliche Folge der Hypophysekto- mie; innerhalb der Gesamtwirkung der Hypophyseninsuffizienz stellt sie die Hauptvariationsursache fur die Senkung der Serumproteine und der alkalischen Serumphosphatase dar.
Summary
1. Serum proteins and serum phosphatases are significantly lowered in hypophysecto- mized pigeons.
2. The reduction of food intake and of body weight in the extend as related to hypo- physectoniy lowers the level of all protein fractions and the activity of the alkaline but not of the acid phosphatase. There is no correlation between the activity of alkaline and acid serum phosphatase.
3. The interaction between the protein fractions in hypophysectomized pigeons is evident but not significant.
4. The reduced food intake as followed from hypophysectomy represents the main source of variation in the decrease of the serum proteins and alkaline phosphatase among the combined effect of the failure of pituitary hormones.
Literatur
1 . AMBEGOAKAR, D. S . , and CHANDRAN, K., 1959: Physiological effects of low protein diets. 11. Studies on hemoglobin, erythrocytes and plasma protein formation in rat. Indian J. med.

Uber Veranderungen der Serumproteine und Serurnphosphatasen 25 1
Res. 47. 552-562. - 2. ANDERSCH, M. A., and SZCYPINSKI, A. J., 1947: vgl. Merkotest N r . 3305, Amer. J. clin. Path. 17, 751. - 3. ANDERSON, C. G., and ALTMAN, A., 1951: The electrophoretic serum-protein pattern in malignant malnutrition. Lancet, I, 203-204. - 4. BAKER, R., and MIII.ER jr., H., 1951: Janendocrinectomy" and the serum proteins in the dog. Endocrinology 49, 484-489. - 5. BEARDWOOD, C., GILBERT, C., and GILLMAN, J., 1962: Endocrine control of the plasma albumin/globulin ratio. Nature 195. 710-711. - 6. BER- NASCONI, C., 1956: Serum proteins and protein-bound carbohydrates in the rat treated with an anterior pituitary extract. Acta endocrinol. 23, 184-195. - 7. BELL, D. J., 1960: Tissue components of the domestric fowl. 4. Plasma-alkaline-phosphatase activity. Biochem. J. 75, 224-229. - 8. BRACKENRIDCE, C. J., and CSILLAG, E. R., 1962: A quantitative electrophoretic survey of serum protein fractions in health and disease. Acta med. scand. 172, Suppl. 383. - 9. BRANDT, L. W., CLI:GG, R. E., and ANDREWS, A. C., 1951: The effect of age and degree of maturity on the serum proteins of the chicken. J. biol. Chem. 191, 105-111. - 10. BUCH- WALD, K. W., and HUDSON, L., 1944: The biohemical effects of sex hormones on acid and alkaline phosphatase activity, calcium and phosphorus. Endocrinology 35. 73-82. - 11. CORDIER, D., et CHANEL, J., 1959: Action de l'histamine sur I'activitC des phosphatases acides e t alcalines d u serum chez le rat. J. Physiol. 51, 439-440. - 12. DESBOROUGH, S., and IRWIN, M. R., 1966: Additional variations in serum proteins in Columbidae. Physiol. 2001. 39, 66-69. - 13. DEUTSCH, H. F., and GOODLOE, M. B., 1945: An electrophoretic survey of various animal plasma. J. biol. Chem. 161, 1-20. - 14. GERLACH, E., FLKKENSTEIN, A., und FREIJND, K. J., 1957: Konzentration und Turnover der Adcnosin- und Guanosinphosphate sowie anderer saureloslicher Phosphorverbindungen in Taubenerythrocyten. Studien mit s*P- markiertem Orthophosphat. Pfliigers Arch. ges. Physiol. 263, 682-703. - 15. GOHR, H., HUSSONG, H. J., und LANCENBERG, H., 1950: Untersuchungen iiber die Verlnderungen des SerumeiweiRbildes bei den verschiedenen Verlaufsformen der Hungerkrankheit, zugleich ein Beitrag zur Schadigung der Leberfunktion bei Inanition. 2. inn. Med. 5, 22-31. - 16. GOLD- BERG, 1.. 1938: Action d e I'insuffisance hypophysaire sur les protCines d u plasma sanguin. C. r. SCance SOC. Biol. 128, 1135-1137. - 17. HARTENBACH, W., und RITTER, R., 1957: Untersuchungen iiber das Verhalten der Serum-EiweiR-Korper unter Einwirkung eines A C T H - Stresses. Munch. med. Wschr. 99, 215-217. - 18. HEIM, W. G., and SCHECHTMAN, A. M., 1954: Electrophoretic analysis of the serum of the chicken during development. J. biol. Chem. 209, 241-247. - 19. HOCKER, W., AMTMANN, E., und KRIESTEN, K., 1967: Ober die Wirkung der Hypophysektomie auf die Wasserstofiionenkonzentration in verschiedenen Abschnitten des Magen-Darm-Kanals der Taube. Z. vergl. Physiol. 56, 221-231. - 20. JACKSON, S. H., 1952: The effect of food ingestion on intestinal and serum alkaline phosphatase in rats. J. biol. Chern. 198, 553-559. - 21. JONES, L. M., and SHINOWARA, G. Y., 1942: Serum inorganic phosphate ,,alkaline" phosphatase activity in hypophysectomized rats. J. biol. Chern. 142, 935-937. - 22. KOLB, E., 1955: Untersuchungen iiber die AktivitHt von Serumfermenten (Diastase, Lipase, saure und alkalische Phosphatase) bei Haustieren. Naturwissenschaften 42, 418-419. - 23. KRUSKEMPER, H., 1958: Uber den EinfluR von Cortison und Schilddriisen- hormonen auf die Serumproteine. Arch. exp. Path. Pharmakol. 233, 284-295. - 24. KUSCH, T., und RINK, D., 1963: Die Veranderungen der sauren Serumphosphatase (Typ 11) durch Chlorpromazin und das Verhalten der Prostataphosphatase. Acta biol. med. german. 11, 480-184. - 25. LARON, Z., FORTUNATO, J., and KLEIN, R., 1957: Serum alkaline phosphatase response to the administration of cortisone in rat. Endocrinology 61, 483-487. - 26. LARON, Z., and KOWADLE, A., 1961: Serum acid phosphatase response to starvation and cortisone administration in rats. Metabolism 10, 1100-1104. - 27. LESHER, S., and BURMESTER, B. R., 1955: Plasma phosphatase activities of normal and lymphomatous chickens. Cancer Res. 15, 537-540. - 28. LEVCHFNKO, M. N., 1962: Role of the functional interaction of the thyroid gland and adrenals of the blood serum. Fiziol. Z. 8, 630-637. - 29. LEVIN, L., and LEATHEM, J. H., 1942: The relation of the pituitary, thyroid and adrenal glands to the maintenance of normal serum albumin and globulin levels. Am. J. Physiol. 136, 303-313. - 30. LI. C. H.. KALMAN, C., EVANS. H. M., and SIMPSON, M. E., 1946: The effect of hypophysectomy and adrenocorticotropic hormone on the alkaline phosphatase of rat plasma. J. biol. Chem. 163, 715-721. - 31. LI, <:. H., and REINHARDT, W. O., 1947: Electrophoresis of rat plasma. 11. The effect of adrenocorticotropic hormone. J. biol. Chem. 167, 487-493. - 32. LI, C. H., KALMAN, C., and EVANS, H. M., 1947: The effect of hypophyseal growth hormone on the alkaline phosphatase of rat serum. J . biol. Chem. 169, 625-629. - 33. LIENERT, G. A., 1962: Uber die Anwendung von Variablen-Transformationen in der Psychologie. Biometr. 2. 4, 145-181. - 34. LOWRY, 0. H., ROSEBROUCH, N. J., FARR, N. J., and RANDALL, R. J., 1951: Protein measurement with the Fohn phenol reagent. 1. biol. Chem. 193, 265-275. - 35. MANIIEL, P., CLAVERT, J.. et MANDEL, L., 1947: Modificationes de sirurn apr& ligature des vaisseau hipatiques chez. le pigeon soumis A I'action de la folliculine. C. r. Seance SOC. Biol. 141, 913-914. - 36. MOORE, D. H. , LEVIN, L., and LEATHEM, J. H., 1944: The n-globulin fraction of the serum of normal and hypophysectomized rats. J. biol. Chem. 153, 349-353. -

252 W . Hocker, K . Colln und E. Amtmann
37. MOORE, D. H., 1948: Effect of reciprocal steroid treatment on the electrophoretic patterns of fowl sera. Endocrinology 42, 38-45. - 38. RICK, W., und HAUSAMEN, T. V., 1965: vgl. Merkotest 3314, Z. analyt. Chem. 212, 267. - 39. RIDDLE, O., 1963: Prolactin in vertebrate function and organization. J. nat. Cancer Inst. 31, 1039-1110. - 40. RITTS jr., R. E., and OUDRICK, W’., 1964: Electrophoresis of serum proteins on cellulose acetate. Am. J. clin. Path. 41, 321-331. - 41. ROSENMUND, H., 1953: Untersuchungen iiber die saure Phosphataseaktivi- t i t mit besonderer Beriicksichtigung der Prostataphosphatase und des Blutserums. Helv. med. Acta, Ser. A, 20, Suppl. 33, Basel. - 42. SCHERMER, S., 1954: Die Blutmorphologie der Laboratoriumstiere. Leipzig: Johann Ambrosius Barth. - 43. SCHMIDT, W., 1962: Verande- rungen der SerumeiweiCkorper von Ratten nach Oestrogengaben. Zbl. Gynak. 84, 179-185. - 44. SHAPIRO, A. B., and SCHECHTMAN, A. M., 1949: Effect of adrenal cortical extract on the blood picture and serum proteins of fowl. Proc. SOC. exp. Biol. Med. 70, 440-445. - 45. SHEEHAN, H. L., and SUMMERS, V. K., 1949: The syndrome of hypopituitarisrn. Quart. J. Med. 42, (N. S. 18), 319-378. - 46. STURKIE, P. D., 1954: Avian Physiology. Ithaca, New York: Bailliire, Tindall and Cox, London and Comstock Publ. - 47. TANABE, Y., NAKAYAMA, R., SASAKI, K., and HOSODA, T., 1954: Sex and age differences in estrogen induced serum vitellin production in the chick. Tap. J. zootech. Sci. 25, 137-141. - 48. TANABE, Y., 1960: Thyroidal control of serum alkaline phosphatase level in the chicken. Endocr. jap. 7, 153-1 56. - 49. TANABE, Y., and W r ~ c o x , F. H., 1960: Effects of age, sex and line on serum alkaline phosphatase of the chicken. Proc. SOC. exp. Biol. Med. 103, 68-70. - 50. TANABE, Y., 1962: Influence of age upon the ability of thyroxine and estrogen to increase serum alkaline phos- phatase of the chicken. Gen. comp. Endocr. 2, 446-452. - 51. TUBA, J., BAKER, D. B., and CANTOR, M. H., 1949: The relationship of serum phosphatase to sex hormones. Can. 1. Res. E. 27, 202-209. - 52. TUBA, I., and MADSEN, N. B., 1952: The relationship of dietary factors to rat serum alkaline phosphatase. 11. The effect of total food consumption, methionine, choline, and vitarnine B,, in normal and alloxan diabetic rats. Can J. Med. Sci. 30, 18-25. - 53. URIEL, J., FINE, J. M., et LE BOURDELLES, F., 1957: Contribution ?I I’itude des prot6ines des sCrums d’animaux. Bull. SOC. Chim. Biol. 39, 1415-1427. - 54. URIST, M. R., 1959: The effect of calcium deprivation upon blood, adrenal cortex, ovary, and skleton in domestic fowl. Recent Prog. Horrn. Res. 15, 455-481. - 55. URIST, M. R., and DEUTSCH, N. M., 1960: Effects of cortisone upon blood, adrenal cortex, gonads and the development of osteoporosis in birds. Endocrinology 66, 805-818. - 56. VASTONE, W. E., MAW, W. A., and COMMON, R. H., 1955: Levels and partition of the fowl’s serum proteins in relation to age and egg production. Can. J. Biochem. Physiol. 33, 891-903. - 57. WARNER, R. C., WEBER, I., DE BODO, R. C., and KURTZ, M., 1957: Effect of hypophysectomy and subsequent administration of growth hormone and ACTH on serum proteins of dogs. Am. J. Physiol. 190, 121-128. - 58. WEIMER, H. E., and GODFREY, J. F., 1964: Effects of inanition, semistarvation and proteinfree diets on serum proteins. Am. J. Physiol. 206, 331-337. - 59. ZUR HORST-MEYER, H., 1954: Das Verhalten der alkalischen Phosphatase bei hypophysaren und myxodematosen Zwergen vor und wahrend der Behandlung. Arztl. Wschr. 9. 657-658.
AnschriP der Autoren: Dr. WALTER HOCKER und KLAUS COLLN, Zoologisches Institut, 5 Koln-Lindenthal, Weyertal 119, und Dr. EDUARD AMTMANN, Anatomisches Institut. 5 Koln-Lindenthal, Lindenburg





























