Embed Size (px)
Citation preview

ULTRASTRUCTURE OF MITOSIS AND
CYTOKINESIS IN THE MULTINUCLEATEGREEN ALGA ACROSIPHONIA
PEGGY R . HUDSON and J . ROBERT WAALAND
From the Department of Botany, University of Washington, Seattle, Washington 98195
ABSTRACT
The processes of mitosis and cytokinesis in the multinucleate green algaAcrosiphonia have been examined in the light and electron microscopes. The courseof events in division includes thickening of the chloroplast and migration ofnumerous nuclei and other cytoplasmic incusions to form a band in which mitosisoccurs, while other nuclei in the same cell but not in the band do not divide .Centrioles and microtubules are associated with migrated and dividing nuclei butnot with nonmigrated, nondividing nuclei . Cytokinesis is accomplished in theregion of the band, by means of an annular furrow which is preceded by a hoop ofmicrotubules . No other microtubules are associated with the furrow . Characteris-tics of nuclear and cell division in Acrosiphonia are compared with those of othermultinucleate cells and with those of other green algae .
INTRODUCTION
274
In multinucleate cells, nuclear division may occursynchronously, asynchronously, or in a wavespreading from one part of the cell to another (fora general discussion, see Agrell, 1964 ; Grell, 1964 ;Erickson, 1964). Cytokinesis may or may not beassociated with nuclear division (Grell, 1964 ; Jbns-son, 1962 ; Kornmann, 1965, 1966 ; Schussnig,1931, 1954 ; Lewis, 1909). In the multinucleategreen alga Acrosiphonia, an unusual pattern ofnuclear and cell divison occurs (Jbnsson, 1962 ;Kornmann, 1965): some of the nuclei of eachmultinucleate cell gather to form a band of nucleilocated in the region where cytokinesis will lateroccur. Next the nuclei in the band undergo syn-chronous division, forming two bands of daughternuclei that move away from one another duringingrowth of an annular septum in the region of thebands . Meanwhile, nuclei that are not part of the
band remain scattered in the cytoplasm at somedistance from the band and do not participate inmitosis . The recently divided nuclei soon scatterinto the cytoplasm. Thus, as in uninucleate cells,nuclear and cell division in Acrosiphonia areclosely coordinated spatially and temporally, butin the multinucleate Acrosiphonia, a substantialportion of the nuclei in a dividing cell do notparticipate in the synchronous nuclear divisionswhich occur in the band . This paper presents theresults of observations which shed new light on thisunusual pattern of nuclear and cell division . Wehave studied this process by means of sequentialphotomicrography of living, dividing cells andhave examined the process at the ultrastructurallevel . We have also compared this process withnuclear and cell division as it occurs in othermultinucleate cell types. In addition, we have
THE JOURNAL OF CELL BIOLOGY - VOLUME 62, 1974 - pages 274-294
on January 3, 2019jcb.rupress.org Downloaded from http://doi.org/10.1083/jcb.62.2.274Published Online: 1 August, 1974 | Supp Info:

compared our results with those reported for othergreen algae, since nuclear and cell division patternsappear to be of importance in considering thephylogeny of these organisms (Pickett-Heaps,1972 b: Pickett-Heaps and Marchant, 1972; Stew-art et al ., 1973) .
MATERIALS AND METHODS
The isolate of Acrosiphonia spinescens (Kütz.) Kjellm .which has been used in this study was originally obtainedfrom zooids released from green unicellular endophytesof the foliose red alga Schizymenia sp . (Wynne no . 1620)collected 12 May 1968 on the east side of FoulweatherBluff, Skunk Bay, Hansville, Wash., by Dr . Michael J .Wynne. Since that time it has been grown in unialgalculture at 10 ° C in Provasoli's Enriched Seawater me-dium (PES) (Provasoli, 1968) in Pyrex no . 3250 storagedishes . Illumination at about 1,200 lx is provided bycool-white fluorescent lights, on a 16-h light/8-h darkphotoregime .We have followed the recommendation of Wille
(1900) that multinucleate, branched members of theAcrosiphoniaceae be called Acrosiphonia and uninu-cleate branched ones be called Spongomorpha . Thetaxonomic and nomenclatural problems in this group arerather involved and will not be dealt with here .
The details of the growth and life history of this algacorrespond in general to those described by Kornmann(1965), JSnsson (1962), and Chihara (1969) for A .spinescens (Kütz.) Kjellm .
In obtaining material for the cytological and ultra-structural study of nuclear and cell division, we havetaken advantage of the fact that cells of Acrosiphoniaplants grown in alternating cycles of light and darkundergo most nuclear and cell divisions after entering thedark cycle (J6nsson, 1962 ; Kornmann, 1965) . Cells at thetips of filaments divide within about I h after entering thedark cycle ; the process occurs somewhat later and lessfrequently in intercalary cells . Tip cells have been used inthis study because it is easy to obtain large numbers ofdividing tip cells at one time, and because, in preparingcells for electron microscopy, we have greater successwith tip cells than with intercalary cells .
Light Microscopy
Living cells in division were studied by placing shortfilaments in PES on glass slides and making a thickmount by supporting the coverglass with cover glassfragments . The thick preparation was placed on aheating-cooling stage (Leitz model 80, E . Leitz, Inc .,Rockleigh, N . J .) maintained at 10 °C. Condensation onthe top of the cold cover slip was prevented by applying athin film of Spray-Kleen (American Optical Corp .,Research Div., Framingham Center, Mass .) (H . H .Heunert, personal communication) to the upper face of
the cover slip before placing it over the alga . With achange of medium approximately once per hour, many ofthe tip cells completed nuclear and cell division within afew hours, the time required depending mainly on thestage of the process of division in which we found thecells when we put them on the stage . It is easy to identifya cell in which the division processes have begun, sincethe presence of the band of aligned nuclei in tip cells isevident to the unaided eye . Micrographs were takenmostly with bright-field optics to reduce the problemscreated by the high refractivity of the cell walls and thenumerous pyrenoids .
A modification of Buffaloe's fixative (Buffaloe, 1958 ;Kapraun, 1969) was used before acetocarmine stainingof plants for squash preparations .
Sections of material prepared for electron microscopy(see below) were cut at I µm thickness with glass knivesand stained with Richardson's stain (Richardson et al .,1960) for subsequent light microscope examination .
Electron Microscopy
Several different techniques were used in attempting tofind a satisfactory method of preparing Acrosiphonia forultrastructural studies . The large, highly vacuolate cellswere quite susceptible to collapse and plasmolysis . Bestresults were obtained when we used phosphate-buffered2.5% glutaraldehyde followed by postfixation in 1%phosphate-buffered osmium tetroxide, with osmolalitiesof the fixing and rinsing solutions adjusted to approxi-mately that of sea water by the addition of NaCl (Cloneyand Florey, 1968) . After OSO .4 postfixation the materialwas rinsed in buffer solutions with gradual stepwisedecreases in osmolality as recommended by Burr andWest (1970) and then embedded in agar and passedthrough a series of water-methyl Cellosolve solutions toabsolute methyl Cellosolve before ethanol dehydration,as recommended by Fowke and Pickett-Heaps (1969 a) .The material was rinsed in propylene oxide and infil-trated and embedded in Epon 812 (Luft, 1961) .
Sections were cut with a diamond knife and stainedwith uranyl acetate (omitted when in block staining wasdone using 1% uranyl nitrate in 70% ethanol overnight)and lead citrate (Reynolds, 1963) . They were examinedwith a Zeiss 9S-2 electron microscope .
RESULTS
Thallus Structure and Interphase
Cell Structure
Acrosiphonia is a uniseriate, filamentousbranched green alga which is a characteristicmember of many marine algal communities incool-temperate and cold regions and is quitecommon in Puget Sound . Each plant is differenti-ated into an upright and rhizoidal portion . The
P. R . HUDSON AND J . R . WAALAND Ultrastructure of Mitosis and Cytokinesis
275

upright, branched shoot system consists of cellsabout 100 µm in diameter ; the rhizoidal system ismade up of branched filaments 30-40 µm indiameter that can terminate in small adherentdisks composed of repeatedly branched filamentsof cells 8-10 µm in diameter . In the uprightportion of the plant, the cell length is typically oneto three times the diameter but may reach amillimeter or more in rapidly growing tip cells .
Young cells of the upright part of the plant eachcontain a peripheral layer of cytoplasm averagingabout 5 µm in thickness, appressed to the cell wallby the large central vacuole . The single largereticulate chloroplast containing numerous pyre-noids is the most striking component of the cellswhen they are viewed at low magnification (Figs .5, 6) . Smaller organelles and inclusions are ob-scured by the chloroplast when it over- or underliesthem, but they are more easily seen when theyoccur in the cytoplasm filling the perforations inthe chloroplast (Fig . 1) .
Ultrastructurally, the cells of Acrosiphonia arelike those of many other green algae. The chloro-plast contains lamellae characteristically consist-ing of two thylakoids each (Fig . 10) which appearto enlarge and protrude as finger-like projectionsinto the polypyramidal pyrenoids (Chadefaud,1941) . Numerous mitochondria about 0.5 x 1-5µm in longitudinal section, dictyosomes, lengths ofendoplasmic reticulum, vesicles, and a number ofnuclei 3-5 µm in diameter containing one promi-nent nucleolus each, are scattered apparently atrandom in the cytoplasm . The number of nucleiper cell varies according to the size of the cell andthe volume of its cytoplasm, ranging from two orthree nuclei in tiny rhizoid cells, to over a thousandnuclei in large tip cells . The difference betweennuclei which are going to divide and those whichare not is not evident structurally in interphasecells. We have not observed microtubules or
276
centrioles in association with interphase nucleieither in dividing or nondividing cells . Mi-crotubules, though sparse, seem to be consistentlyoriented parallel to the longitudinal axis of theinterphase cell . The walls of young tip cells may beless than 1 µm in thickness (Fig . 3), whereas thewalls of older cells may reach 10 µm in thickness,and appear to be composed of several layers (Fig .2) . Ultrastructurally these layers are made up offibrillar material, which in section presents aherringbone effect at least in the innermost layers(Fig . 9) . A somewhat more electron-dense nonfi-brillar outer wall layer about 35 nm thick ispresent in some cells (Fig . 9) ; it sometimes is lost inpreparation . This wall thickening with advancingage appears to be a consequence of continued walldeposition combined with minimal elongationgrowth of intercalary cells .
Mitosis and Cytokinesis
BAND FORMATION : Our light micro-scope observations on nuclear and cell divisionagree with those of J6nsson (1962) and to someextent with those of Kornmann (1965) on A .spinescens . We have found that the same processtakes place in A . coalita, another species commonin cool marine waters along the Pacific coast ofNorth America .
At low magnifications in the living tip cells, thenuclei cannot be observed as they undergo divisionwithin the band because of the highly refractilechloroplast and pyrenoids nearby, and because ofthe thickness of the specimen . In the thinnerpreparations necessary for observations at highermagnifications, the refraction problems are quitesevere, and the cells do not survive for the length oftime necessary to complete division . A preliminaryattempt to bleach a "window" in the cytoplasm(Kamitsubo, 1972) in the area in which nuclear
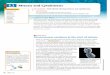
FIGURE I Phase-contrast longitudinal surface view of interphase cell . Note reticulate chloroplast (cps) withrefractile pyrenoids (py), and numerous small nuclei (n) . x 545 .
FIGURE 2 Thick cross section of old cell, showing thick, layered wall (w), nuclei (n), pyrenoids (py), andlarge central vacuole (v) . x 1,200 .
FIGURE 3 Thick longitudinal section of cell with aligned premitotic nuclei . Note elongate, teardrop-shapednuclei (n) with nucleoli, thickened chloroplast (cpl), thin wall (w) which is characteristic of young cells .x 860 .
FIGURE 4 Postdivision migration of bands of nuclei (n) in intercalary cell . Single arrows mark nuclearbands, double arrow marks septum . Pyrenoids are also stained by acetocarmine technique . x 530 .
THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974
P. R. HUDSON AND J . R. WAALAND Ultrastructure ofMitosis and Cytokinesis
277

band formation and cell division could be pre-dicted to occur bleached the chloroplast but killedthe cell . In our work we have attempted to relatethe changes in band shape which we observed inliving tip cells to the band shapes which appear tobe characteristic of the stages in nuclear divisionwhich we have observed in fixed and sectionedmaterial .
While cytoplasmic streaming is not evident inliving cells of Acrosiphonia, rearrangements ofnuclei and of local regions of the chloroplast andsome other organelles do precede nuclear and celldivision .
The first visible event in the division process is alocal thickening of the chloroplast, which indicatesthe region of the cell in which nuclear and celldivision will occur (Fig . 5 a-c) . Within the next 30min, a bright colorless band forms at the peripheryof the thickening chloroplast, which is thus ex-cluded to regions above, below, and inward fromthe bright band (Fig . 5 d) . Numerous nuclei, inassociation with a particular arrangement of someother organelles (discussed below), constitute theband (Figs . 3, 7) .
Comparison of the division process in tip cellswith that in intercalary cells reveals some differ-ences in location of the band of nuclei, number ofnuclei contained in the band, partitioning of nucleiinto daughter cells, size of daughter cells formed,and frequency of division . In tip cells, the bandforms at a distance about 1/4 to'/ the length of thecell back from its tip, and is up to six nuclei inheight and four nuclei in thickness . The band ofnuclei undergoes changes in shape during thedivision process, and three to five times as manydivided nuclei are partitioned into the smaller,newly formed tip cell as into the larger, newsubapical cell. The new tip cell then elongatesrapidly during the 12-24-h period preceding thenext division, increasing its postdivision volume bya factor of three to five, while the new subapicalcell elongates only a few micrometers, if at all,during the same period . In contrast, the band individing intercalary cells is characteristically onlyone nucleus in height and thickness, and is sym-metrical throughout division, with equal numbersof daughter nuclei partitioned into daughter cellsof equal size (Fig . 4). Although they continue todeposit cell wall material, intercalary cells do notelongate significantly, and instead add to thethickness of their walls . In addition, they divideless frequently than do tip cells .
278 THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974
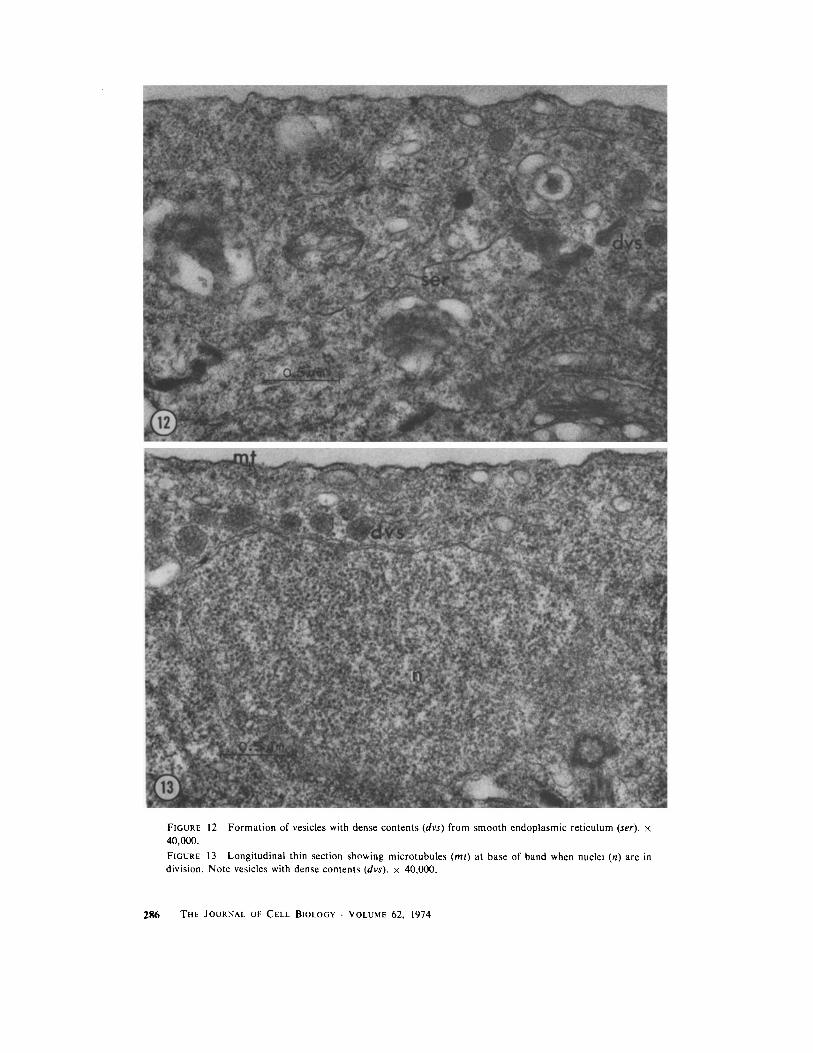
When first formed, the bright band in tip cells isonly a few micrometers in height, and appearslongitudinally symmetrical . As it increases in size,however, it extends and thickens asymmetrically,so that its thickest region is near its base, and itsupper portion tapers out somewhat toward the celltip (Figs . 5 e-g, 6 a, b) . The arrangement of thecomponents of a band of this shape is unusual . Thenuclei, which constitute the greater part of thevolume of the band, form its center . They areteardrop-shaped in longitudinal section, their nar-row ends appearing to focus on a region at the baseof the band (Figs . 3, 7) . At this stage the nucleoliare quite apparent as large, dark-staining bodiesat the larger ends of the nuclei . Centrioles arelocated at the narrow ends of the elongated nuclei(Figs . 7, 8) and a few microtubules oriented ap-proximately parallel to the longitudinal axis of thecell interpolate between nuclei or between theoutermost nuclei and the plasmalemma, near thenarrowed tips of the nuclei (Fig . 8) . Several layersof endoplasmic reticulum form the upper andlower boundaries of the band, and some endo-plasmic reticulum also is present between nucleiwithin the band (Fig . 7) . Vesicles and dictyosomesappear throughout the band but are most concen-trated toward its base, where they are located inthe area that constitutes the apparent focal pointfor the elongated nuclei . The vesicles in the bandare of two types which can be distinguished bytheir contents : the majority of them have electron-lucent contents and the others have electron-dense, granular contents (Figs . 7, 7 a, 12-16) . Wehave not observed intermediates between thesetwo types. Apparently the vesicles with electron-dense contents are formed from smooth endoplas-mic reticulum (Fig. 12) and in addition their size(0 .5-3 .0 gm diameter) and their electron-densecontents agree with those described for the generalclass of organelles identified as microbodies(Newcomb and Frederick, 1971 ; Graves et al .,1971) . We have not yet performed cytochemicaltests to determine whether they contain enzymescommonly found in microbodies . The origin of thevesicles with electron-lucent contents is not clear .The vesicles with electron-dense contents aregrouped at the focal point of the nuclei, while themore numerous vesicles with electron-lucent con-tents are scattered throughout the base of theband (Figs . 7, 7 a, 13) .MITOSis : About 1 .5-2 h after the first slight
thickening of the chloroplast is observed, the
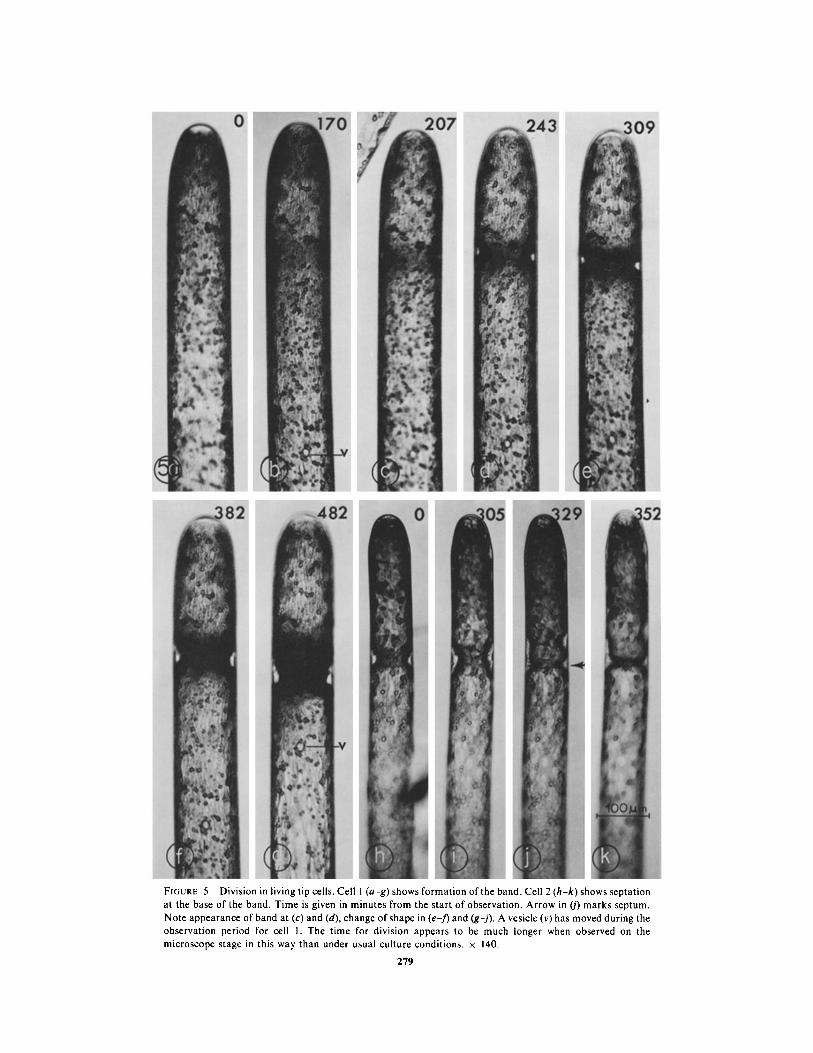
FIGURE 5 Division in living tip cells . Cell I (a-g) shows formation of the band . Cell 2 (h-k) shows septationat the base of the band . Time is given in minutes from the start of observation . Arrow in (j) marks septum .Note appearance of band at (c) and (d), change of shape in (ef) and (gj) . A vesicle (v) has moved during theobservation period for cell 1 . The time for division appears to be much longer when observed on themicroscope stage in this way than under usual culture conditions . x 140.
279
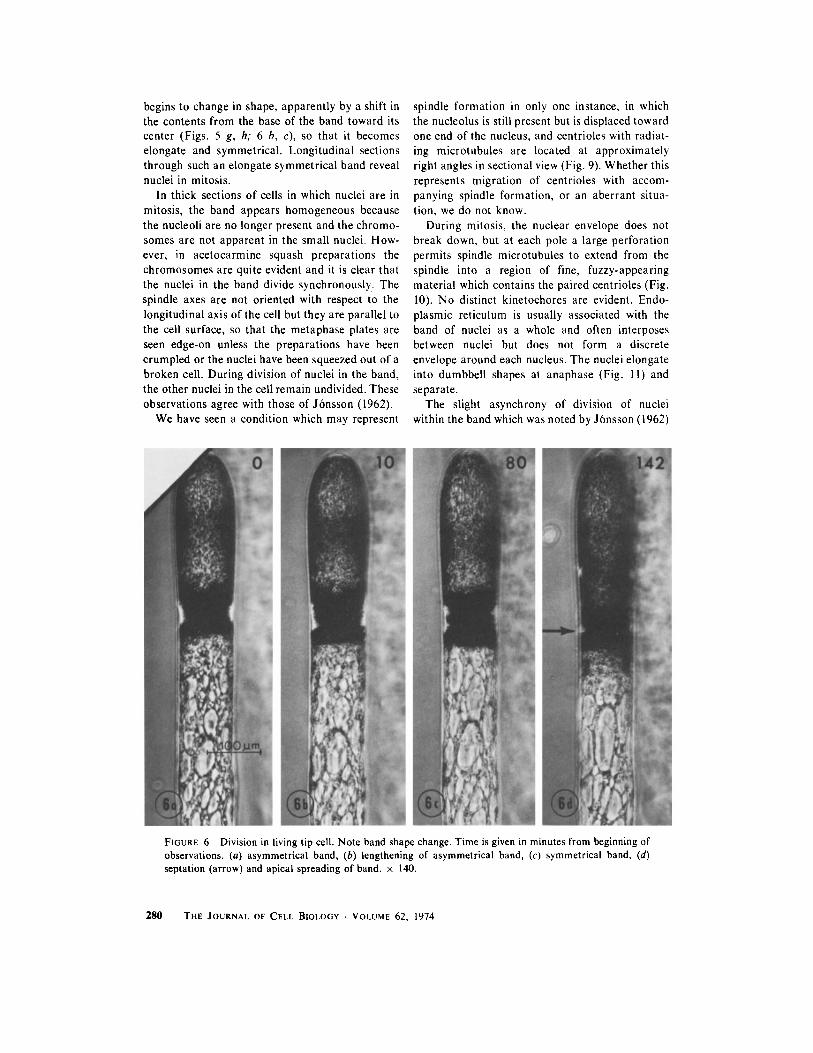
begins to change in shape, apparently by a shift inthe contents from the base of the band toward itscenter (Figs . 5 g, h; 6 b, c), so that it becomeselongate and symmetrical . Longitudinal sectionsthrough such an elongate symmetrical band revealnuclei in mitosis .
In thick sections of cells in which nuclei are inmitosis, the band appears homogeneous becausethe nucleoli are no longer present and the chromo-somes are not apparent in the small nuclei . How-ever, in acetocarmine squash preparations thechromosomes are quite evident and it is clear thatthe nuclei in the band divide synchronously . Thespindle axes are not oriented with respect to thelongitudinal axis of the cell but they are parallel tothe cell surface, so that the metaphase plates areseen edge-on unless the preparations have beencrumpled or the nuclei have been squeezed out of abroken cell . During division of nuclei in the band,the other nuclei in the cell remain undivided . Theseobservations agree with those of Jbnsson (1962) .
We have seen a condition which may represent
28 0
spindle formation in only one instance, in whichthe nucleolus is still present but is displaced towardone end of the nucleus, and centrioles with radiat-ing microtubules are located at approximatelyright angles in sectional view (Fig . 9) . Whether thisrepresents migration of centrioles with accom-panying spindle formation, or an aberrant situa-tion, we do not know .
During mitosis, the nuclear envelope does notbreak down, but at each pole a large perforationpermits spindle microtubules to extend from thespindle into a region of fine, fuzzy-appearingmaterial which contains the paired centrioles (Fig .10) . No distinct kinetochores are evident . Endo-plasmic reticulum is usually associated with theband of nuclei as a whole and often interposesbetween nuclei but does not form a discreteenvelope around each nucleus . The nuclei elongateinto dumbbell shapes at anaphase (Fig . 11) andseparate .
The slight asynchrony of division of nucleiwithin the band which was noted by Jbnsson (1962)
FIGURE 6 Division in living tip cell . Note band shape change . Time is given in minutes from beginning ofobservations . (a) asymmetrical band, (b) lengthening of asymmetrical band, (c) symmetrical band, (d)septation (arrow) and apical spreading of band . x 140 .
THE JOURNAL OF CELL BIOLOGY • VOLUME 62, 1974
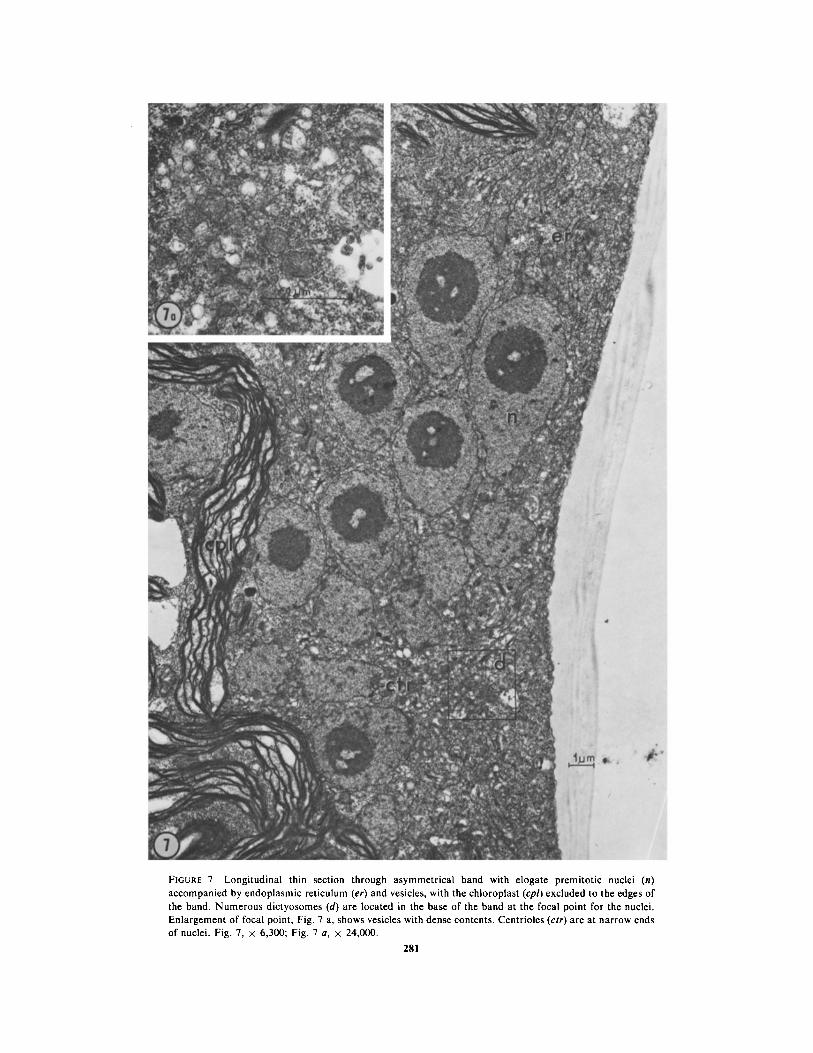
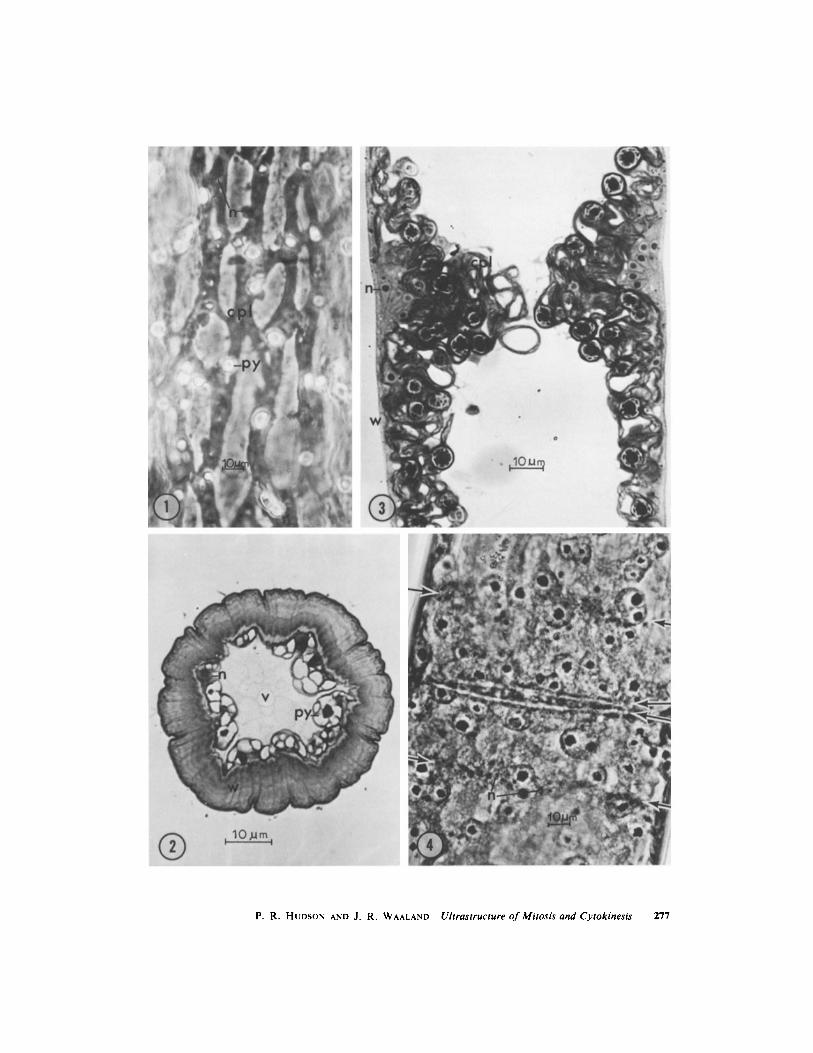
FIGURE 7 Longitudinal thin section through asymmetrical band with elogate premitotic nuclei (n)accompanied by endoplasmic reticulum (er) and vesicles, with the chloroplast (cpl) excluded to the edges ofthe band . Numerous dictyosomes (d) are located in the base of the band at the focal point for the nuclei .Enlargement of focal point, Fig . 7 a, shows vesicles with dense contents . Centrioles (ctr) are at narrow endsof nuclei . Fig . 7, x 6,300 ; Fig . 7 a, x 24,000 .
2 81
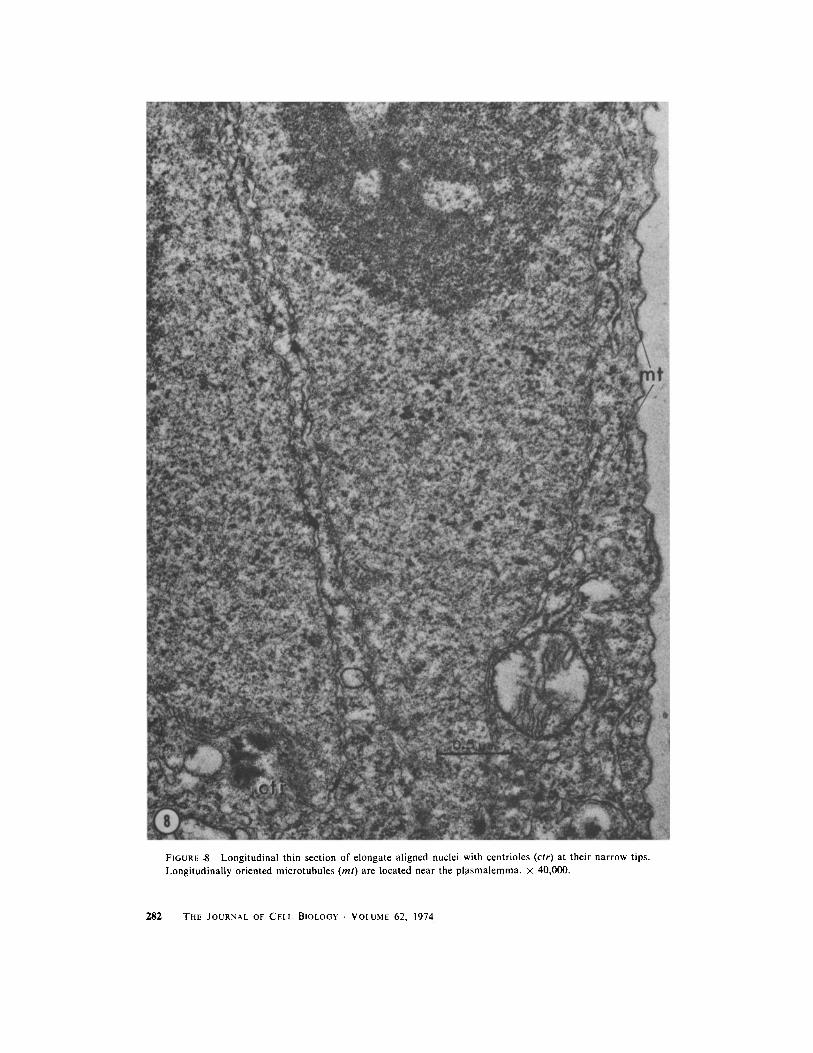
FIGURE $ Longitudinal thin section of elongate aligned nuclei with centrioles (ctr) at their narrow tips .Longitudinally oriented microtubules (ml) are located near the plasmalemma . x 40,000 .
282
THE JOURNAL OF CELL BIOLOGY • VOLUME 62, 1974
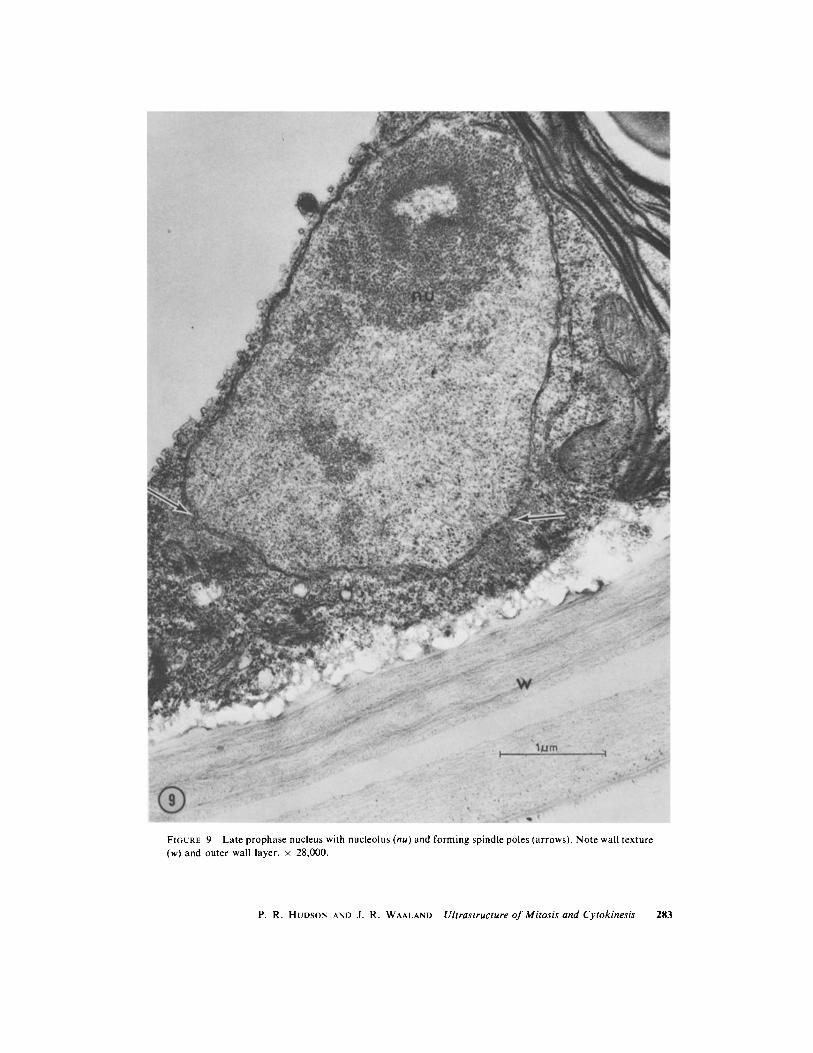
FIGURE 9 Late prophase nucleus with nucleolus (nu) and forming spindle poles (arrows) . Note wall texture(w) and outer wall layer . x 28,000 .
P . R . HUDSON AND J. R. WAALAND Ultrastructure of Mitosis and Cytokinesis
283
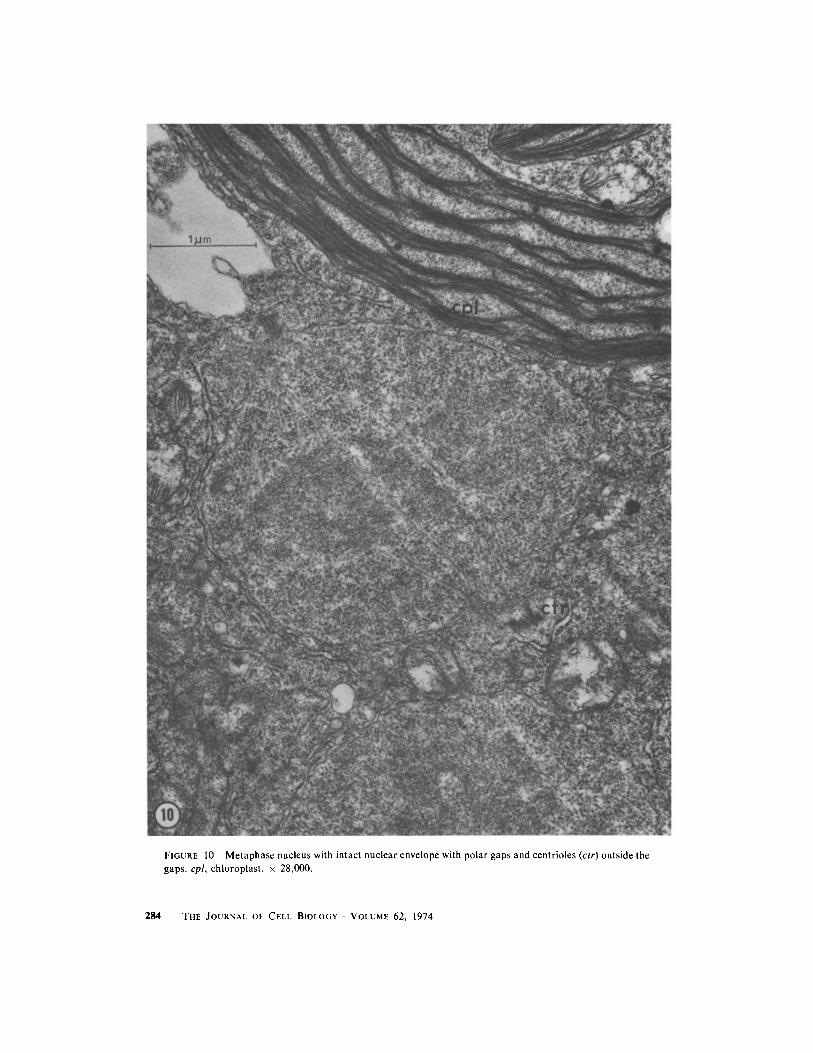
FIGURE 10 Metaphase nucleus with intact nuclear envelope with polar gaps and centrioles (ctr) outside thegaps . cpl, chloroplast . x 28,000 .
2 8 4
THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974
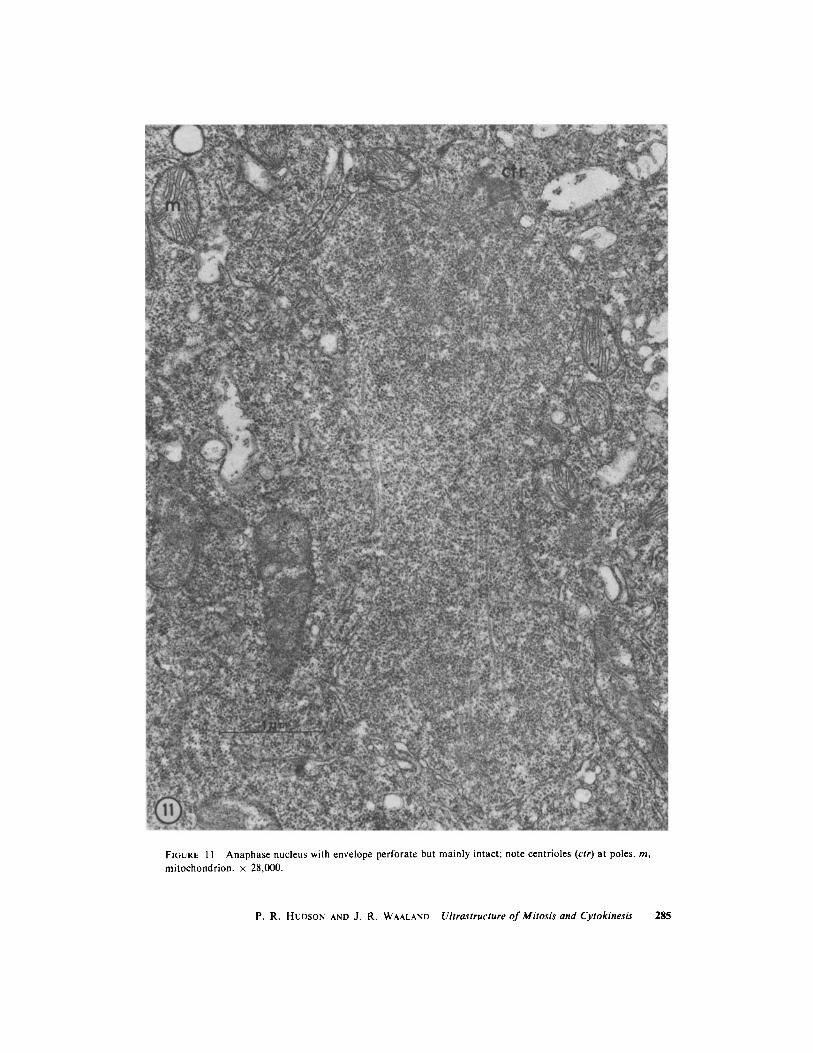
FIGURE I I Anaphase nucleus with envelope perforate but mainly intact ; note centrioles (ctr) at poles . m,mitochondrion . x 28,000 .
P. R. HUDSON AND J. R. WAALAND Ultrastructure of Mitosis and Cytokinesis
285
286
FIGURE 12 Formation of vesicles with dense contents (dvs) from smooth endoplasmic reticulum (ser) . x40,000 .
FIGURE 13 Longitudinal thin section showing microtubules (ml) at base of band when nuclei (n) are indivision . Note vesicles with dense contents (dvs) . x 40,000 .
THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974

asymmetrical broad-based, narrow-topped bandwas also encountered in this study, in the case of aparticularly large band of nuclei in a tip cell . Inthis case, nuclei at the upper and lower edges of theband were in metaphase when nuclei in the centerof the band were in late anaphase .
Although the nucleolus is still present at the timewhen chromatin is condensing (Fig . 9), it is notapparent at metaphase (Fig . 10) when the nucleo-plasm contains a slightly greater density of granu-lar material . After division the nucleolus is obviousagain (Fig . 4), but we have not observed stages inits reformation .POSTMITOTIC MIGRATION AND CYTO-
KINESIS : Within 0 .5 h of the time that the pre-mitotic, broad-based band begins to change inshape, most of the band contents are displacedapically away from the original band base loca-tion, and cytokinesis takes place near the originalbase location (Figs . 5j, k; 6 d). The site at whichthe initiated septum appears is somewhat variablewith relation to the base of the group of nuclei,since sometimes it is entirely below the group andsometimes within the basal part of the group .
In newly formed tip cells the bright band ofrecently divided nuclei remains evident for up to anhour, as it moves and disperses toward the tip ofthe cell (Fig . 6 d; Fig. 4, intercalary cell) . Thespreading and apical displacement of the bandseems too rapid to be accounted for simply by theresumption of tip growth immediately after divi-sion, since it can shift 100 µm in 30 min, and theoverall rate of elongation of the tip cell is about700 um in 24 h . Also, the distance from the tip ofthe cell to the top of the band decreases during thisspreading, while the distance from the cell tip tothe new septum remains the same .
Cytokinesis is accomplished by the infurrowingof an annular septum (Figs . 14-16) . Its locationmay be determined as early as the asymmetricband stage before mitosis, when nuclei appear tobe focused toward the base of the band . At thattime, the group of vesicles with electron-densecontents is present in a small area of cytoplasm inthe base of the band, near the plasmalemma (Figs .7, 10) . When nuclei in the band are at metaphase,this group of vesicles is in the same position, but inaddition there is a group of two to three mi-crotubules external to them (Fig . 13) . Becausethese microtubules appear in precise or slightlyoblique cross section when observed in longitu-dinal sections of cells, we believe that they areperpendicular to the longitudinal axis of the cellsand are arranged around the cytoplasmic periph-
ery just internal to the plasmalemma . When theinitiated septum is actually visible as a projectionfrom the old cell wall, a larger group of 8-20 mi-crotubules is located at the leading edge of theseptum (Figs . 14, 15) .
We have not observed other arrays of mi-crotubules to be present in the cytoplasm beyondthe advancing septum, in contrast with the reportsof such microtubules in other green algae. Wewould expect that such microtubules would bevisible to us since we can detect microtubules in thecytoplasm of interphase cells, in mitotic spindles,and in the small group closely associated with thegrowing septum .
During cytokinesis, the vesicles with electron-dense contents are located just internal to theleading edge of the septum (Figs . 13-15). Thevesicles with electron-lucent contents are concen-trated along the entire surface of the growingseptum (Fig . 16) . Numerous dictyosomes are alsopresent in the region of the growing septum . Thestainable fibrillar material present in the older cellwall is not visible either in the newest portions ofthe forming septum or in the dictyosomes orelectron-lucent vesicles near it .DISCUSSIONUnusual Features of Mitosis andCytokinesis in Acrosiphonia
SYNCHRONY : The cytology of this unusualseparation of nuclei into different dividing andnondividing roles in division has been discussedextensively by Jônsson (1962) and Kornmann(1965) . Because the plants arise from uninucleatemeiospores, or in some cases, from unfused ga-metes, they begin life as uninucleate cells . They arealso capable of vegetative propagation from elon-gate rhizoids or from rhizoidal disks of small cellscontaining only a few nuclei per cell . For cells tobecome multinucleate, karyokinesis must occur anumber of times in the absence of cytokinesis . Invery young filaments which are undergoing rapidextension and expansion growth, a band of nucleiis formed, but all nuclei in the cell undergo division(Jônsson, 1962). In somewhat older filaments inwhich cells are still actively dividing vegetatively,the nuclei in the band divide while those in the endsof the cell do not (Jônsson, 1962 ; Kornmann, 1965 ;this paper) . The basis for this transition is notknown . However, it apparently is not permanent,since in the formation of gametes, cells are cleavedinto uninucleate units, and there is no evidence toindicate that any nuclei are excluded from thisprocess (Jônsson, 1962 ; Hudson, unpublished re-
P. R. HUDSON AND J. R. WAALAND Ultrastructure of Mitosis and Cytokinesis 287
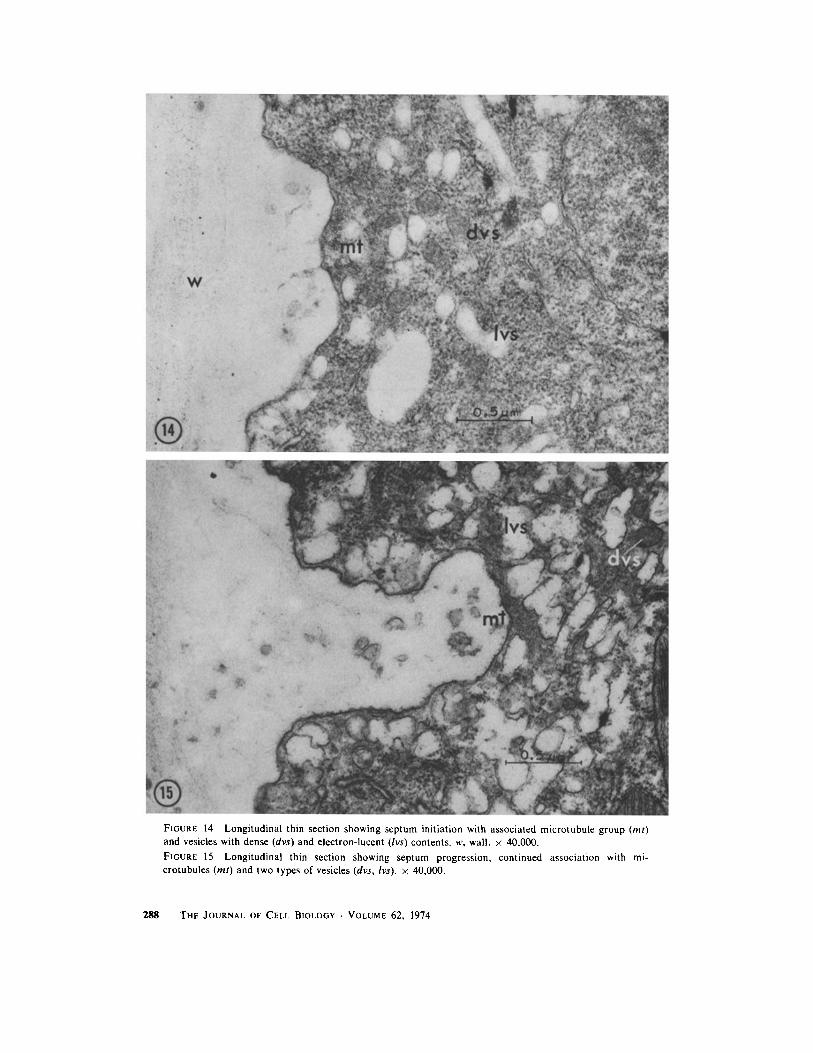
FIGURE 14 Longitudinal thin section showing septum initiation with associated microtubule group (mt)and vesicles with dense (dvs) and electron-lucent (lvs) contents. w, wall . x 40,000 .
FIGURE 15 Longitudinal thin section showing septum progression, continued association with mi-crotubules (ml) and two types of vesicles (dvs, lvs) . x 40,000 .
288
THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974
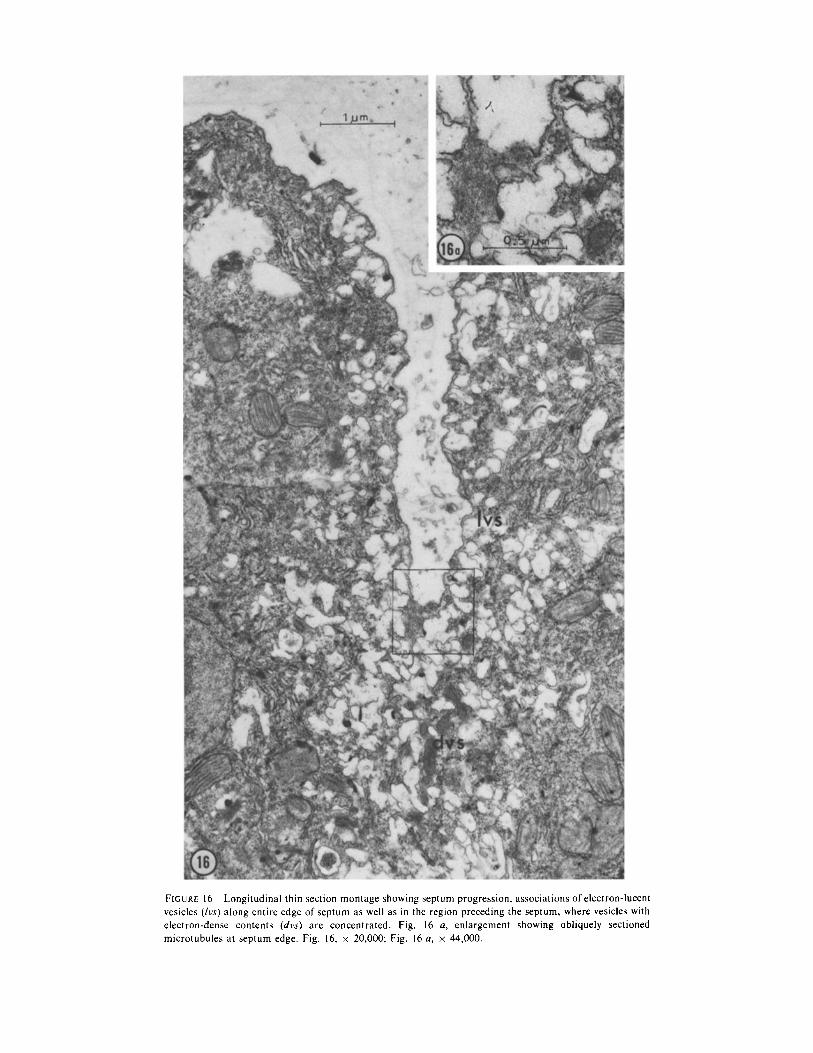
FIGURE 16 Longitudinal thin section montage showing septum progression, associations of electron-lucentvesicles (!vs) along entire edge of septum as well as in the region preceding the septum, where vesicles withelectron-dense contents (dvs) are concentrated . Fig . 16 a, enlargement showing obliquely sectionedmicrotubules at septum edge . Fig . 16, x 20,000; Fig . 16 a, x 44,000 .

suits) . The roles of the nuclei and the coordinationof nuclear and cell division thus can vary duringthe life of the cells .
C E N T R I O L E s : The presence or absence of cen-trioles at different times in the same organism isnot unique to Acrosiphonia; ephemeral centriolesare found in many plants which have flagellatecells at some time in their life histories . Centriolescommonly are found at the poles of the mitoticspindle even when there is no evidence that flagel-late cells are about to be formed (Pickett-Heaps,1969 a, 1971) . However, in most such plants,centrioles are present continuously in interphase .In Chara (Pickett-Heaps, 1967), Oedogonium(Pickett-Heaps and Fowke, 1969), and Bulbocha-ete (Pickett-Heaps, 1973 b) there are no centriolespresent at vegetative mitosis although male ga-metes with flagella are formed when sexual repro-duction occurs . In the fungus Labyrinthula (Per-kins and Amon, 1969 ; Porter, 1972) and in thegreen alga Kirchneriella (Pickett-Heaps, 1970),what seem to be very simple centrioles are pro-duced just before mitosis and are not present ininterphase . Thus the absence of centrioles inassociation with interphase nuclei in Acrosiphoniais unusual but not unprecedented .MOVEMENT OF NUCLEI : Before division in
Acrosiphonia the centrioles are located at thenarrow end of a teardrop-shaped nucleus in theband, with the nucleolus in the broad opposite end .We have not been able to detect these nuclei inmotion in living cells . However, the shape of thenucleus and the location of the centrioles is likethat reported by Girbardt (1968) and by Wilsonand Aist (1967) for moving nuclei in fungi, inwhich a kinetochore equivalent is present and cen-trioles are lacking . In the desmid Closterium lit-torale (Pickett-Heaps and Fowke, 1970), whichalso lacks centrioles, a similarly elongate nucleusis preceded by a microtubule center in relocationof the nucleus in a daughter cell after division .
Before mitosis in uninucleate cells it is notuncommon for the nucleus to take up a positiondifferent from that which it occupies at interphase .However, it is unusual for this repositioning tooccur in multinucleate cells . In the giant amoebaPelomyxa carolinenis (= Chaos chaos), Kudo(1947) reported that the entire amoeba contractsbefore mitosis and many nuclei. divide in protru-sions extending from the main body of theamoeba. Thus it is rather difficult to compare thepositions of the nuclei at mitosis with the positions
2 9 0 THE JOURNAL OF CELL BIOLOGY • VOLUME 62, 1974
in interphase because the whole organism haschanged its shape . Urospora, another green algaquite similar to Acrosiphonia, provides the onlyother example of readily identifiable movement ofnuclei to new positions in the cell before vegetativemitosis and cytokinesis in a multinucleate cell .However, in Urospora, (Kornman, 1966), all nu-clei in a cell divide, not just the ones aligned in aband at the future site of cytokinesis .
Comparison with OtherMultinucleate Organisms
In a multinucleate cell, nuclear division may besynchronous, asynchronous, or may occur as awave of division passing from one part of the cellto another (Agrell, 1964 ; Erickson, 1964 ; Grell,1964) . In studying the series of nuclear divisions inthe formation of pollen in orchids, Heslop-Harri-son (1966, 1968) found that cytoplasmic continui-ties between nuclei may be important in maintain-ing synchrony . In species in which septation occurslate in the series of mitoses, mitotic synchronypersists for more nuclear division cycles than inspecies in which septation begins after the first fewcycles of mitosis . In Drosophila embryos, nucleardivision is synchronous for the 12 mitotic cyclesthat precede the first cell division that leads to theformation of blastoderm (Fullilove and Jacobsen,1971). However, some multinucleate algae haveasynchronous division of nuclei within the samecell (Godward, 1966-Siphonocladales ; J6nsson,1962; Schussnig, 1931-Cladophora), so that al-though cytoplasmic continuity may promote orallow synchrony of nuclear division, it does notappear to impose synchrony .
In the multimicronucleate ciliate Spirostomum,it is reported that nuclear division is synchronousbut that only 20-30% of the nuclei undergo mitosisat one time (Sesachar and Padmavathi, 1956) .Whether the nuclei are assembled in groups beforedivision and whether cytokinesis is closely relatedto mitosis, apparently have not been determined .However, according to Grell (1964), mitotic syn-chrony in multinucleate protozoans is rare, so thatSpirostomum appears to be unusual among proto-zoans .
Comparison with Other Green AlgaeMitosis and cytokinesis have been studied ultra-
structurally in a number of green algae andcomparison of the characteristics observed may be

useful in phylogenetic considerations (Pickett-Heaps, 1972 b, c; Pickett-Heaps and Marchant,1972; Stewart et al ., 1973) .Among the ultrastructural studies on mitosis
and cytokinesis in green algae, only those on thevegetatively nonseptate Bryopsis (Burr and West,1970) and on the coenobial Hydrodictyon (Mar-chant and Pickett-Heaps, 1970) can provide exam-ples of these processes in multinucleate greenalgae. Both undergo septation regularly only dur-ing formation of motile uninucleate reproductivecells . Thus septation in those multinucleate-algaemay be considered to be basically multiple cleav-age of a multinucleate mass resulting in theformation of uninucleate units . This occurs inmany organisms in early embryonic developmentor in formation of reproductive cells . Gametogene-sis and zoosporogenesis in Acrosiphonia entail justsuch a cleavage pattern . In contrast, except for thepresent study on Acrosiphonia, members of theCladophorales, which are composed of filamentsof multinucleate cells which regularly undergocytokinesis vegetatively, have not been studiedultrastructurally with regard to mitosis and cyto-kinesis .MITOSis : The closed mitotic spindle with polar
perforations in Acrosiphonia is like that in manygreen algae, including Chlamydomonas reinhardtii(Johnson and Porter, 1968), Ova mutabilis (LO-vlie and BrAten, 1970), Oedogonium sp . (Pickett-Heaps and Fowke, 1969), Bulbochaete hiloensis,(Pickett-Heaps, 1973 b), Chlorella pyrenoidosa(Atkinson et al ., 1971 ; Wilson et al., 1973),Hydrodictyon reticulatum (Marchant and Pickett-Heaps, 1970), and Bryopsis hypnoides (Burr andWest, 1970) . However, the perforations in thenuclear envelope at anaphase in Acrosiphonia alsoshow some similarity to the partial breakdown ofthe nuclear envelope in Stigeoclonium and Ulo-thrix (Floyd et al ., 1972 a) and Microspora(Pickett-Heaps, 1973 a) . The complete breakdownof the nuclear envelope is reported in Closteriumlittorale (Pickett-Heaps and Fowke, 1970), Cos-marium botrytis (Pickett-Heaps, 1972 a), Spi-rogyra sp . (Fowke and Pickett-Heaps, 1969 a),Chara (Pickett-Heaps, 1967), Klebsormidium flac-cidum (Floyd et al ., 1972 b), and K. subtillissimum(Pickett-Heaps, 1972 c) .
The presence of centrioles at the poles of themitotic nucleus does not appear to be directlyrelated to the open or closed condition of themitotic nucleus : Ulothrix (Floyd et al ., 1972 a) and
Klebsormidium (Floyd et al., 1972 b; Pickett-Heaps, 1972 c) have both polar centrioles anddispersal of the nuclear envelope, while Spirogyra(Fowke and Pickett-Heaps, 1969), Closterium(Pickett-Heaps and Fowke, 1970), and Chara(Pickett-Heaps, 1967) have dispersal of the nuclearenvelope but lack centrioles . Except for Oedogo-nium (Pickett-Heaps and Fowke, 1969) and Bulbo-ehaete (Pickett-Heaps, 1973 b), species with anintact nuclear envelope have centrioles present justoutside the gaps at the spindle poles . In Stigeo-clonium (Floyd et al ., 1972 a) the centriolesactually enter the nucleus at prophase . In Chlorella(Wilson et al ., 1973) and Chlamydomonas (John-son and Porter, 1968) the centrioles appear to bemore directly related to initiation of the septum atcytokinesis than to mitosis .CYTOKINESIS : Microtubules were not ob-
served in the plane of septation in Ulva (L¢vlie andBrAten, 1970), Klebsormidium flaccidum (Floyd etal ., 1972 b), K . subtillissimum (Pickett-Heaps,1972 c), or in the present study of A . spinescens .Microtubules appear to be involved in septumformation in a number of green algae, where theyare arranged in the plane of a developing cell plate,the group of microtubules being termed a "phyco-plast" by Pickett-Heaps (1972 b) . This group ofalgae includes Oedogonium (Pickett-Heaps andFowke, 1969), Bulbochaete (Pickett-Heaps, 1973b), Chlorella pyrenoidosa (Atkinson et al ., 1971 ;Wilson et al ., 1973), Scenedesmus (Nilshammer etal ., 1972), Ulothrix fimbriata (Floyd et al ., 1972a), U. belkae (Stewart et al ., 1973), Stigeocloniumhelveticum (Floyd et al ., 1972 a), S . farctum, S .tenue, Chaetophora incrassata, Uronema confer-vicolum, Schizomeris leibleinii, and Draparnaldiaplumosa (Stewart et al ., 1973). In Microspora(Pickett-Heaps, 1973 a) a phycoplast is associatedwith a combination of an annular septum and a cellplate . The presence of microtubules in the plane ofseptation in the Drosophila zygote (Fullilove andJacobsen, 1971) indicates that this sort of involve-ment is not confined to green algae . By contrast,microtubules oriented perpendicular to the planeof septation, which along with vesicles fusing toform a cell plate constitute the phragmoplastfound in higher plants, have been observed inChara (Pickett-Heaps, 1967), which is not gener-ally considered to be very closely related to the restof the green algae . In Spirogyra (Fowke andPickett-Heaps, 1969 b), septation begins by in-growth of an annular septum and ends with a small
P. R. HUDSON AND J . R. WAALAND Ultrastructure of Mitosis and Cytokinesis 291

phragmoplast between the daughter nuclei . Thephragmoplast in Coleochaete (McBride, 1970 ;Marchant and Pickett-Heaps, 1973) is initiatedbetween the daughter nuclei during cytokinesisalong the radius of the discoid thallus, but developsfrom a more lateral position during cytokinesisparallel to the circumference of the thallus .
Microtubules which appear to mark the locationof future septation were first described in develop-ing stomatal complexes in wheat, by Pickett-Heaps and Northcote (1966) . Such microtubuleshave since been observed in some other higherplants (Burgess and Northcote, 1967-Phleum ;Cronshaw and Esau, 1968-Nicotiana) and insome green algae (Johnson and Porter, 1968-C .reinhardtii; Pickett-Heaps, 1970-Kirchneriella ;Atkinson et al ., 1971-Chlorella) . Perhaps the oneor two microtubules next to the just-initiatedseptum in C. littorale (Pickett-Heaps and Fowke,1970) are comparable . The function of thesegroups of microtubules remains unclear (Pickett-Heaps, 1969 b; Burgess and Northcote, 1967) . Themicrotubules in wheat are present at preprophaseand disappear at later stages of mitosis, which ledPickett-Heaps (1969 b) to suggest that they mightrepresent the effect of mobilization of micro-tubules from positions in the cytoplasm for usein the mitotic spindle . However, this does not seemto be likely in Acrosiphonia, in which moremicrotubules are present at the leading edge of thegrowing septum after mitosis than are present atthe septum initiation site when they are firstnoticeable, at metaphase . In Acrosiphonia, themicrotubules apparently form a band just internalto the advancing annular septum, somewhat likethe position of the group of microtubules internalto the furrow in C. reinhardtii (Johnson andPorter, 1968) .
SUMMARY AND CONCLUSIONS
Acrosiphonia is like most of the green algae thusfar investigated ultrastructurally in that kineto-chores were not observed and the nuclear envelopeis intact at mitosis, with polar gaps that open ontoa dense amorphous area containing centrioles . It islike Ulva and Klebsormidium, in that it lackseither the phycoplast or the phragmoplast type ofseptation observed in many other green algae . It isdistinct among the green algae and other plantsbecause before mitosis it forms a large band ofnuclei accompanied by a characteristic array ofendoplasmic reticulum, dictyosomes, and vesicles,
2 9 2 THE JOURNAL OF CELL BIOLOGY • VOLUME 62, 1974
bounded by the large reticulate chloroplast, acharacteristic shared only by its close relative,Urospora . It is not known whether the migratednuclei of Urospora are associated with centriolesand microtubules while nonmigrated nuclei lackthis association, as is the case in Acrosiphonia .However, nuclei in Urospora do not appear to berole-separated, since regardless of their position,all nuclei in a cell of Urospora undergo synchro-nous division . As far as we know, the only parallelto mitotic-non mitotic role separation among nu-clei occurs in a multimicron ucleate ciliate,Spirostomum (Sesachar and Padmavathi, 1956),which unfortunately does not appear to have beenstudied further. This kind of relationship amongnuclei may represent just one sort of approach tothe general problem of propagating the cells ofmultinucleate, multicellular organisms. Perhaps inAcrosiphonia the gathering of nuclei into a band isa way of insuring that at least some nuclei aredistributed into each daughter cell, and in thespecial case of tip cells, of distributing the largernumber of nuclei into the cell which will increasemost rapidly in volume . The formation of the bandof nuclei along with the accompanying cytoplasmiccomponents also in some way might define thelocation of cytokinesis .
The kind assistance and encouragement of Dr . M . J .Wynne, especially in the beginning stages of this study,have been most helpful . Dr. R. E. Norris providedinvaluable technical assistance and the heating-coolingstage used for studies of living, dividing cells . The adviceand assistance of Dr . E . F . Haskins in many aspects ofthis study are greatly appreciated .
This investigation was carried out with the aid of aNational Science Foundation pre-doctoral fellowship (toP. R . Hudson) and Public Health Service Training Grantno. HD-00266 from National Institute of Child Healthand Human Development .Received for publication 3 August 1973, and in revisedform 11 March 1974 .
REFERENCES
AGRELL, 1 . 1964. Natural division synchrony and mitoticgradients in metazoan tissues . In Synchrony in CellDivision and Growth . E, Zeuthen, editor . IntersciencePublishers, Inc ., John Wiley & Sons, Inc ., New York .
ATKINSON, A . W ., JR ., B . E . S. GUNNING, P . C . L . JOHN,
and W. MCCULLOUGH . 1971 . Centrioles and mi-crotubules in Chlorella . Nat . New Biol. 234:24-25 .
BUFFALOE, N. D . 1958 . A comparative cytological studyof four species of Chlamydomonas . Bull . Torrey Bot .Club . 85:157-178 .

BURGESS, J ., and D . H . NORTHCOTE . 1967 . A function ofthe preprophase band of microtubules in Phleumpratense . Planta (Berl .) . 75:319-326 .
BURR, F. A ., and J . A . WEST . 1970 . Light and electronmicroscope observations on the vegetative and repro-ductive structures of Bryopsis hypnoides . Phycologia.9 :17-37 .
CHADEFAUD, M . 1941 . Les pyrénoides des algues . Ann .S ci . Nat . Bot . Biol . Veg . Ser . I t . 2 :1-44 .
CHIHARA, M . 1969 . Culture study of Chlorochytriuminclusum from the northeast Pacific . Phycologia .8:127-133 .
CLONEY, R. A ., and E . FLOREY . 1968 . Ultrastructure ofcephalopod chromatophore organs . Z. Zellforsch .Mikrosk . Anat . 89:250-280 .
CRONSHAW, J ., and K . ESAU . 1968 . Cell division inleaves of Nicotiana . Protoplasma . 65:1-24 .
ERICKSON, R. O . 1964. Synchronous cell and nucleardivision in tissue of higher plants . In Synchrony in CellDvision and Growth . E . Zeuthen, editor . IntersciencePublishers, Inc ., John Wiley & Sons, Inc ., New York .
FLOYD, G . L., KD . STEWART, and K . R . MATTOX . 1972a. Comparative cytology of Ulothrix and Stigeocloni-um. J . Phycol. 8 :68-80 .
FLOYD, G. L ., K . D . STEWART, and K. R . MATTOX . 1972b . Cellular organization, mitosis and cytokinesis in theUlotrichalean alga Klebsormidium . J. Phycol .8:176-183 .
FOWKE, L. C ., and J . D . PICKETT-HEAPS . 1969 a . Celldivision in Spirogyra . I . Mitosis . J. Phycol. 5:240-
259 .FOWKE, L . C ., and J . D . PICKETT-HEAPS . 1969 b . Cell
division in Spirogyra . II . Cytokinesis . J. Physol .5:273-280 .
FULLILOVE, S. L . and A . G. JACOBSEN . 1971. Nuclearelongation and cytokinesis in Drosophila montana .Dev. Biol. 26:560-577 .
GIRBARDT, M . 1968 . Ultrastructure and dynamics of themoving nucleus . Symp . Soc. Exp. Biol. 22:249-259 .
GODWARD, M . B . E . 1966. The Chromosomes of theAlgae . St . Martin's Press Inc ., New York .
GRAVES, L . B., Jr ., L. HANZELY, and R. N . TRELEASE .1971 . The occurrence and fine-structural characteri-zation of microbodies in Euglena gracilis . Prolo-plasma . 72:141-152 .
GRELL, K . G. 1964 . The protozoan nucleus . In The Cell .J . Brachet and A. Mirsky, editors . VIII-79 .
HESLOP-HARRISON, J . 1966 . Cytoplasmic continuitiesduring spore formation in flowering plants . Endeavour(Engl . Ed.) . 25:65-72 .
HESLOP-HARRISON, J . 1968 . Synchronous pollen mitosisand the formation of the generative cell in massulateorchids . J. Cell Sci. 3 :457-466 .
JOHNSON, U. G ., and K . R . PORTER. 1968 . Finestructure of cell division in Chlamydomonas reinhardi.Basal bodies and microtubules . J. Cell Biol.38:403-425 .
JdNSSON, S. 1962 . Recherches sur des Cladophoracées
marines . (structure, reproduction, cycles comparés,conséquences systématiques) . Ann. Sci. Nat. Bot . Biol .Veg . Ser. 12 . 3 :25-230.
KAMITSUBO, E . 1972 . A "window technique" for detailedobservation of characean cytoplasmic streaming . Exp .Cell Res . 74:613-616 .
KAPRAUN, D . 1969 . Field and cultural studies of thegenera Ulva and Enteromorpha . Ph.D. Thesis. Uni-versity of Texas, Austin, Texas .
KORNMANN, P. 1965 . Zur Analyse des Wachstums unddes Aufbaus von Acrosiphonia . Helgolaender Wiss .Meeresunters . 12:219-238 .
KORNMANN, P. 1966. Wachstum und Zellteilung beiUrospora. Helgolaender Wiss . Aeeresunters . 13:73-83 .
KORNMANN, P. 1970. Phylogentische Beziehungen inder Grilnalgengattung Acrosiphonia. HelgolaenderWiss . Meeresunters . 21:292-304 .
KUDO, R. R . 1947 . Pelomyxa carolinensis Wilson, II .Nuclear division and plasmotomy . J. Morphol.80:93-125 .
LEWIS, 1 . F . 1909 . The life-history of Griffithsia bor-netiana . Ann . Bot . (Lond .) . 23:639-690 .
L¢VLIE, A., and T . BRATEN . 1970. On mitosis in themulticellular alga Ulva mutabilis F¢yn . J. Cell Sci .6:109-219 .
LUFT, J . H . 1961 . Improvements in epoxy resin embed-ding methods . J. Biophys . Biochem . Cytol . 9:409-414 .
MCBRIDE, G . E . 1970 . Cytokinesis and ultrastructure inFritschiella tuberosa Iyengar . Arch. Protistenkd.112 :365-375 .
MARCHANT, H. J ., and J. D . PICKETT-HEAPS . 1970 .Ultrastructure and differentiation of Hydrodictyonreticulatum . I . Mitosis in the coenbium . Aust . J . Biol .Sci . 23:1173-1186 .
MARCHANT, H. J ., and J . D. PICKETT-HEAPS . 1973 .Mitosis and cytokinesis in Coleochaete sculata . J.Phycol . 9:461-471 .
NEWCOMB, E . H ., and S . E . FREDERICK. 1971. Distribu-tion and structure of plant microbodies (peroxisomes) .In Photosynthesis and Photorespiration . M . O . Hatch,C. B. Osmond, and R . O . Slatyer, editors . Intersci-ence Publishers, Inc ., John Wiley & Sons, Inc ., NewYork. 442-457 .
NILSHAMMAR, M ., B . WALLIS, and A. KYLIN . 1972 . Theeffects of calcium deficiency on the ultrastructure ofthe green alga Scenedesmus. Z. Pfanzenphysiol .66 :197-205 .
PERKINS, F . O ., and J . P . AMON . 1969 . Zoosporulation inLabyrinthula sp . : an electron microscopic study . J .Protozool. 16:235-257 .
PICKETT-HEAPS, J . D . 1967 . Ultrastructure and differen-tiation in Chara sp . Aust . J . Biol . Sci . 20:883-894 .
PICKETT-HEAPS, J . D. 1969 a . The evolution of themitotic apparatus : an attempt at comparative ultra-structural cytology in dividing plant cells . Cytobios .3:257-280 .
PICKETT-HEAPS, J .D. 1969 b . Preprophase microtubules
P. R. HUDSON AND J . R . WAALAND Ultrastrueture of Mitosis and Cytokinesis 293

and stomatal differentiation : some effects of centrifu-gation on symmetrical and asymmetrical cell division .J.Ultrastruct . Res . 27:24-44 .
PICKETT-HEAPS, J . D. 1970. Mitosis and autosporeformation in the green alga Kirchneriella lunaris .Protoplasma . 70:225-247 .
PICKETT-HEAPS, J . D.1971 . The autonomy of the centri-ole : fact or fallacy? Cytobios . 3 :205-214 .
PICKETT-HEAPS, J . D. 1972 a . Cell division in Cos-marium botrytis .J . Phycol . 8 :343-360 .
PICKETT-HEAPS, J . D. 1972 b . Variation in mitosis andcytokinesis in plant cells : its significance in phylogenyand evolution of ultrastructural systems . Cytobios .5:59-77 .
PICKETT-HEAPS, J . D. 1972 c. Cell division in Klebsor-midium subtillissimum (formerly Ulothrixsubtillissima) and its possible phylogenetic signifi-cance . Cytobios . 6 :167-183 .
PICKETT-HEAPS, J . D. 1973 a . Cell division and wallstructure in Microspora. New Phytol . 72 :347-355,
PICKETT-HEAPS, J . D . 1973 b . Cell division inBulbochaete . 1 . Divisions utilizing the wall ring . J .Phycol. 9 :408-420 .
PICKETT-HEAPS, J . D., and L. C . FowKE . 1969 . Celldivision in Oedogonium . I . Mitosis, cytokinesis andcell elongation . Aust.J . Biol . Sci . 22 :857-894 .
PICKETT-HEAPS, J . D ., and L. C . FOWKE . 1970 . Mitosis,cytokinesis and cell elongation in the desmid Closter-ium littorale . J . Phycol . 6 :189-215 .
PICKETT-HEAPS, J . D ., and H . J . MARCHANT . 1972 . Thephylogeny of the green algae : a new proposal . Cy-tobios . 6 :255-264 .
PICKETT-HEAPS, J . D ., and D. H . NORTHCOTE . 1966 .Cell division in the formation of the stomatal complexof the young leaves of wheat . J. Cell Sci . 1 :121-128 .
PORTER, D . 1972 . Cell division in the marine slime mold,Labyrinthula sp ., and the role of the bothrosome in
294
THE JOURNAL OF CELL BIOLOGY . VOLUME 62, 1974
extracellular membrane production . Protoplasma .74:427-448 .
PROVASOLI, L. 1968 . Media and prospects for cultivationof marine algae . In Culture and Collections of Algae .A . Watanabe and A . Hattori, editors . Proceedings U .S.-Japan Conference Hakona, Sept . 1966. JapaneseSociety of Plant Physiology . 63-75.
REYNOLDS, E . S. 1963 . The use of lead citrate at high pHas an electron-opaque stain in electron microscopy . J .Cell Biol . 17 :208-212 .
RICHARDSON, K . C., L. JARRETT, and E. H FINKE . 1960 .Embedding in epoxy resins for ultrathin sectioning inelectron microscopy . Stain Technol. 35 :313-323 .
SCHUSSNIG, B. 1931. Die somatische and heterotypeKernteilung bei Cladophora Suhriana Kützing . Planta(Berl .) . 13 :474-528 .
SCHUSSNIG, B. 1954 . Gonidiogenese, Gametogenese andMeiose bei Cladophora glomerata (L .) Kützing . Arch .Protistenkd. 100 :287-322 .
SESACHAR, B. R ., and P. B . PADMAVATHI . 1956 . Syn-chronous micronuclear divisions in multimicronucle-ate ciliates . Curr . Sci . (India) . 25 :281-282 .
STEWART, K . D., K . R . MATTOX, and G . L . FLOYD .1973 . Mitosis, cytokinesis, the distribution of plasmo-desmata and other cytological characteristics in theUlotrichales, Ulvales and Chaetophorales : phyloge-netic and taxonomic considerations . J. Phycol .9 :128-140 .
WILLE, N . 1900 . Die Zellkerne bei Acrosiphonia (J . Ag .)Kjellm . Botanisches Centralblatt . 81 :238-239 .
WILSON, C. L ., and J . R . AIST . 1967 . Motility of fungalnuclei . Phytopathology . 57 :769 .
WILSON, H . J ., F . WANKA, and H . F . LINSKENS . 1973 .The relationship between centrioles, microtubules andcell plate initiation in Chlorella pyrenoidosa . Planta(Berl .) . 109 :259-267 .







![Practice For May: Cell Ultrastructure [114 marks]blogs.4j.lane.edu/.../2018/02/Cell-Ultrastructure-Test-1.pdfPractice For May: Cell Ultrastructure [114 marks]1. Which structure found](https://img.pdfslide.net/doc/110x75/5eda4db5b3745412b5711d9c/practice-for-may-cell-ultrastructure-114-marksblogs4jlaneedu201802cell-ultrastructure-test-1pdf.jpg)