Embed Size (px)
Citation preview

ARTICLE
Using mark–recapture information to validate and assess ageand growth of long-lived fish speciesMartin J. Hamel, Jeff D. Koch, Kirk D. Steffensen, Mark A. Pegg, Jeremy J. Hammen,and Mathew L. Rugg
Abstract: Long-lived species from marine and freshwater environments have experienced declines linked to anthropogeniceffects such as overexploitation, dam construction, and habitat modification. An understanding of the age structure and theassociated dynamics determined from these data for long-lived species is critical for both perseverance of at-risk species andmaintenance of exploited species. We used pallid sturgeon (Scaphirhynchus albus) to evaluate the efficacy of mark–recapture datafrom known-age, hatchery-reared fish (ages 1 to 7) to corroborate age and growth estimates obtained from sectioned pectoral finrays. Accuracy of age estimates from known-age fish was 13%, whereas 72% of estimates were within 2 years of the true age.Annual growth was significantly different between estimated growth (back-calculated) and actual observations of tagged pallidsturgeon. Age for pallid sturgeon of any given size was estimated with parameters derived from mark–recapture data, and thepredicted length-at-age relation was similar to observations from known individuals. In instances where age determination forall ages of interest cannot be verified, mark–recapture appears to be a viable solution for examining growth and has shownpromise as a tool for estimating ages in long-lived species with calcified structures that are difficult to read.
Résumé : Les espèces longévives de milieux marins et d’eau douce ont connu des baisses d’abondance associées a des effetsanthropiques tels que la surexploitation, la construction de barrages et la modification des habitats. La compréhension de lastructure par âge et de la dynamique associée déterminée a partir de ces données pour les espèces longévives est essentielle tantpour la persévérance des espèces en péril que pour le maintien des espèces exploitées. Nous avons utilisé le scaphirhynque blanc(Scaphirhynchus albus) pour évaluer l’efficacité de données de marquage–recapture pour des poissons d’âge connu (âgés de 1 a7 ans) élevés en alevinière pour la vérification des estimations de l’âge et du taux de croissance obtenues a partir de coupes derayon de nageoire pectorale. L’exactitude des estimations de l’âge pour les poissons d’âge connu était de 13 %, alors que 72 % desestimations différaient de moins de 2 ans de l’âge réel. Il y avait une différence significative entre la croissance annuelle estimée(par rétrocalcul) et la croissance obtenue a partir d’observations sur des scaphirhynques blancs marqués. L’âge de scaphi-rhynques blancs de toute taille a été estimé en utilisant des paramètres tirés de données de marquage–recapture, et la relationprédite de la longueur selon l’âge était semblable aux observations pour des individus d’âge connu. Dans les cas où la détermi-nation de l’âge pour tous les âges d’intérêt ne peut être vérifiée, l’approche de marquage–recapture semble être une solutionvalable pour examiner la croissance et s’annonce prometteuse comme outil pour estimer les âges chez les espèces longévivesdont les structures calcifiées sont difficiles a déchiffrer. [Traduit par la Rédaction]
IntroductionLong-lived fish species (>20 years) pose a unique set of chal-
lenges for assessment and monitoring, as population-level responsesmay take years to be realized. In marine fisheries, long-lived speciesare often targeted for commercial harvest (Trippel 1995; Hutchingsand Reynolds 2004). Overexploitation of many fish stocks has re-sulted in a collapse of the fishery. Complete fishing closures, al-though unlikely, may be the only means to restore abundance toprevious levels (Pauly et al. 2002; Hutchings and Reynolds 2004).Furthermore, many long-lived fish species such as those that livein or near coral reefs have exhibited declines due to destruction ofhabitat (Coleman et al. 2000; Hughes et al. 2003; Bellwood et al.2004). Long-lived fishes from freshwater systems are not exemptto excessive exploitation and have also been subject to intenseanthropogenic effects such as dam construction, pollution, andhabitat modifications that have further hindered population sta-bility (Boreman 1997; Dudgeon et al. 2006; Leveque et al. 2008).
Many long-lived species have adapted a periodic life-history strat-egy where longevity compensates for variation in juvenile survivaland environmental influence on reproductive success (Winemiller2005). Sporadic recruitment patterns limit these species’ ability toincrease population size quickly, making long-lived species highlysusceptible to declines in abundance (i.e., anthropogenic effects).Therefore, an understanding of the age structure and the interact-ing dynamics (i.e., growth and mortality) of long-lived species iscritical for both perseverance of at-risk species and maintenanceof exploited species.
Several long-lived species inhabit fresh waters of North Americaand most inhabit large rivers. Fishes such as sturgeon (genus:Acipenser and Scaphirhynchus), paddlefish (Polyodon spathula), alliga-tor gar (Atractosteus spatula), and several large species from familiesCatostomidae and Cyprinidae have been known to live greaterthan 30 years (Scoppettone 1988; Bemis et al. 1997; Pikitch et al.2005; Buckmeier et al. 2012). These long-lived species, particularlysturgeons and paddlefish, have received considerable attention in
Received 24 July 2013. Accepted 17 January 2014.
Paper handled by Associate Editor Josef Michael Jech.
M.J. Hamel, M.A. Pegg, J.J. Hammen, and M.L. Rugg. University of Nebraska – Lincoln, 3310 Holdrege Street, Lincoln, NE 68583, USA.J.D. Koch. Kansas Department of Wildlife, Parks, and Tourism, Pretty Prairie, KS 67570, USA.K.D. Steffensen. Nebraska Game and Parks Commission, Lincoln, NE 68503, USA.Corresponding author: Martin J. Hamel (e-mail: [email protected]).
559
Can. J. Fish. Aquat. Sci. 71: 559–566 (2014) dx.doi.org/10.1139/cjfas-2013-0393 Published at www.nrcresearchpress.com/cjfas on 21 January 2014.
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
AL
HO
USI
E U
NIV
ER
on
07/1
4/14
For
pers
onal
use
onl
y.

recent years because of range-wide declines linked to anthropo-genic effects such as overharvest, dam construction, and loss ofhabitat (Boreman 1997). Determining the age structure has been,and will continue to be, an integral component for assessment ofthese long-lived species so that demographic models used to pre-dict population viability and responses to management actionscan be assessed (Bajer and Wildhaber 2007; Koch et al. 2009; Jageret al. 2010; Steffensen et al. 2013). Age of fishes is typically deter-mined by observing periodic growth increments that are producedin calcified structures. If growth marks are formed consistently andcan be interpreted accurately, an estimate of age is produced(Campana and Neilson 1985). Similarly, the distance between growthmarks relative to the size of the calcified structure can often becorrelated to annual, somatic growth in the fish (Francis 1990).These procedures are based on assumptions of consistent deposi-tion of growth marks and that the distance between marks are indirect proportion to the somatic growth (i.e., increase in length) ofthe fish. However, validation of these techniques is an often over-looked prerequisite before using age data for analyses (Beamishand McFarlane 1983).
Validation for individual species should include a verification ofgrowth increment periodicity across the entire age range of inter-est, particularly for long-lived species (Beamish and McFarlane1983; Campana 2001). For example, age estimates from lake stur-geon (Acipenser fulvescens) fin rays were accurate to age-14, anderror increased with age thereafter (Bruch et al. 2009). Failure toproperly verify absolute age estimates can have drastic effects,such as providing overly optimistic or pessimistic growth andmortality estimates. Campana (2001) reported several examples oflong-lived species (i.e., orange roughy (Hoplostethus atlanticus), wall-eye pollock (Gadus chalcogrammus), among others) that experi-enced overexploitation due to the severe underestimation of ages.Most validation studies of long-lived species to date have verifiedthe periodicity and accuracy of growth marks for young individ-uals, but waiting for known-age fish to attain old ages is rarelyutilized and may not always be feasible given the time required(Campana 2001).
Bomb-radiocarbon, radiochemical dating, release of known-ageand marked fish, and mark–recapture techniques have been usedor proposed to attain absolute age validation for long-lived fishes,yet these techniques often require substantial financial and timecommitment from researchers to validate. The accuracy, commit-ment, and expense associated with these techniques will limittheir use in many instances. For example, bomb-radiocarbon is atechnique that examines the amount of 14C that was incorporatedinto the otolith of a fish after nuclear testing began in the 1950s. Asmall amount of 14C indicates the pre-1958 era, while elevatedlevels of 14C indicate the era after nuclear testing. Referencecurves from 14C analysis for fish age validation can be used toestimate the age of fish sampled that were hatched during thenuclear testing period (approximately 1958 to 1968). This ap-proach does not explicitly validate a certain age, only that it washatched before or after nuclear testing began. This technique willeventually be unavailable for most fish species unless archivedcollections are used.
Mark–recapture techniques are well-suited for long-lived spe-cies and can be used to validate both the periodicity of growthincrement deposition and the absolute age. When fish age isknown at marking, either directly or inferred (i.e., young fish),absolute age validation can be performed when fish are recap-tured. However, if fish age is unknown at marking, a calcium-binding chemical such as oxytetracycline, alizarin, or calcein canbe applied at the time of tagging to create a permanent mark onthe structure. Additional growth increments after marking can becompared with the time-at-large for recaptured fish to corroborateincrement periodicity. These methods are the preferred methods forage validation and corroboration outlined in Campana (2001), andthe only perceived limitations are the rigors and cost associated
with fish collections. This is particularly true with old fish, as theprobability of recapture decreases over time (Black et al. 2005).Though mark–recapture data are often difficult to collect, addedbenefits exist that warrant their use. For example, Paragamianand Beamesderfer (2003) used 23 years of white sturgeon (Acipensertransmontanus) mark–recapture data to estimate age and charac-terize growth patterns. In addition, various mark–recapturemodels can be used to determine population size, survival, andmovement patterns (Kendall and Bjorkland 2001; Pine et al.2003). Therefore, the objective of our study was threefold: to usemark–recapture data of known-age, subadult pallid sturgeon(Scaphirhynchus albus) to attempt to validate both age and growthestimates generated from a commonly used aging structure froma long-lived sturgeon species, to demonstrate the applicability ofusing mark–recapture for determining growth patterns, and topredict age given a specified length.
Materials and methodsWe used a mark–recapture data set coupled with a previous age
validation study (Koch et al. 2011) for pallid sturgeon to demon-strate the applicability of using these data to validate the accuracyof age estimates and to determine age and growth characteristics.The pallid sturgeon is a long-lived, fluvial specialist endemic tothe Mississippi River and Missouri River basins (Bailey and Cross1954; Dryer and Sandvol 1993). Many known-age and marked pal-lid sturgeon have been released as part of a Missouri River basin-wide propagation program. Stockings of pallid sturgeon began inthe late 1990s to early 2000s; therefore, most known-age pallidsturgeon are subadult. However, these known-age fish providedan opportunity to attempt to validate both age and growth esti-mates from a commonly used aging structure, as this informationwill be critical for future recovery efforts.
The leading edge of the pectoral fin ray is the most com-monly used age estimation structure for sturgeon of the genusScaphirhynchus. This structure can be removed nonlethally (Kochet al. 2008) and has the highest reported precision compared withother structures (Jackson et al. 2007). However, the accuracy ofage estimates obtained from pectoral fin rays (i.e., absolute age;Campana 2001) has not been successfully validated, and severalauthors have suggested these data be used with caution (Whitemanet al. 2004; Jackson et al. 2007; Kennedy et al. 2007).
Mark–recapture data for hatchery-reared pallid sturgeon fromthe 2001–2007 year classes were collected continuously by theNebraska Game and Parks Commission in the Missouri River, Ne-braska (river kilometres 811–1086). Mark–recapture events wereincluded from pallid sturgeon that were at large for a minimum of30 days after stocking or between subsequent recaptures. Thesedata were compared with fin ray analysis from a previous study(i.e., same year classes) that attempted to validate juvenile pallidsturgeon age estimates (Koch et al. 2011). We assumed that mark–recapture data were the closest semblance to actual annual mea-surements of growth on wild fish and provided an opportunity toexamine the bias and validity of using an aging structure (i.e., finray) to perform back-calculated growth estimates. Individual pal-lid sturgeon were distinguished with a unique tag or combinationof tags (e.g., PIT tag, elastomer, scute removal) that identified theyear class of the individual. All pallid sturgeon were measured tothe nearest millimetre (fork length, FL) at stocking or initial cap-ture and subsequent recapture. Fin ray cross-sections and back-calculated measurements were provided by Koch et al. (2011).
The apparent bias in growth estimation between procedureswas inferred from the comparison of observed growth incrementsfrom mark–recapture data to growth increments from length-at-age estimates generated from fin ray age data. Annual growthincrements of mark–recapture individuals were calculated fromthe following equation:
560 Can. J. Fish. Aquat. Sci. Vol. 71, 2014
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
AL
HO
USI
E U
NIV
ER
on
07/1
4/14
For
pers
onal
use
onl
y.

(1) Gi �(Lr � Lc)
Yi
where Gi is the growth for fish i, Lc is the fork length at firstcapture, Lr is the fork length at recapture, and Yi is the number ofyears between capture events. We standardized the data for vari-ous periods at large to facilitate comparisons by pairing annualgrowth (Gi) to the midpoint in length between capture events(Paragamian and Beamesderfer 2003). Only one growth measure-ment was recorded for fish that were recaptured multiple times,and the longest period of time between captures was chosen torepresent growth of that particular fish.
Determination of growth from fin rays was estimated usingtraditional back-calculation techniques (Dahl–Lea method; DeVriesand Frie 1996). Back-calculated length-at-age estimates provided byKoch et al. (2011) were generated with prior knowledge of age.Knowledge of age was needed to determine which mark was thefirst annulus because of disagreement between readers. Priorknowledge of age would theoretically reduce error in determiningthe number of annuli and should provide results that resembletrue growth patterns, if annuli deposition in fin rays followed con-temporary assumptions (e.g., proportional growth increments). Wealso wanted to determine how back-calculated growth estimateswould compare with other growth estimates without previousknowledge of age. Therefore, we solicited an independent agereader without knowledge of this study to age the fin ray sectionsanalyzed by Koch et al. (2011) and make the appropriate mea-surements between presumptive annuli. The independent readerhad prior experience aging shovelnose sturgeon (Scaphirhynchusplatorynchus) and was instructed to disregard the hypothesizedfalse inner annuli identified by Koch et al. (2011) so that resultswere directly comparable. Age estimates were compared withknown ages with a two-sample t test to determine accuracy.
Finally, we used an equal proportion approach to evaluate ifother growth rates differed from a simple technique of dividingthe fin ray into equal parts for each year the fish was alive. Theassumption is that growth is equally proportionate throughoutthe life of the fish. This equal proportionate approach served asthe null hypothesis that pallid sturgeon growth is linear. Linear-ized growth estimates for each method were compared with ananalysis of covariance (ANCOVA) using calculation method as thecovariate.
von Bertalanffy curves were derived from mark–recapture datawith a modification of the Fabens (1965) method. Growth incre-ment data were fitted to the von Bertalanffy growth curve refor-mulated to account for observed growth between capture periods,so that
(2) �L � (L∞ � Lt)(1 � e�kT)
where �L is the increase in length between capture events, t istime of tagging, T is the number of years between tagging andrecapture, L∞ is the von Bertalanffy length at infinity, and k is thevon Bertalanffy growth rate coefficient. Parameters for the vonBertalanffy growth curve were estimated iteratively using a non-linear regression approach. An estimate of the time at lengthzero (t0) cannot be estimated with this method; therefore, we usedthe formula provided by Pauly (1979):
(3) log(�t0) � �0.3922 � 0.2752logL∞ � 1.038logk
Age (t) for pallid sturgeon of any given size (Lt) could then beestimated by using a reformulation of the von Bertalanffy equa-tion (Kirkwood 1983):
(4) t � t0 � loge[(1 � Lt/L∞)/k]
Predicted ages were estimated with parameters derived from themark–recapture data and were compared with corresponding av-erage ages for pallid sturgeon of the same length that were deter-mined using fin ray data.
Mean FLs (mm) for each of the seven year classes that wereanalyzed in this study (age-1 to age-7) were calculated with each ofthe previously mentioned analytical procedures. Mean length-at-age was compared with a two-way ANOVA, and pairwise compar-isons of procedure-type for each age class were assessed withTukey’s studentized range test. All statistical tests used the statis-tical program R (version 3.0.0; R Development Core Team 2013),and � level for all analyses was set at P ≤ 0.05.
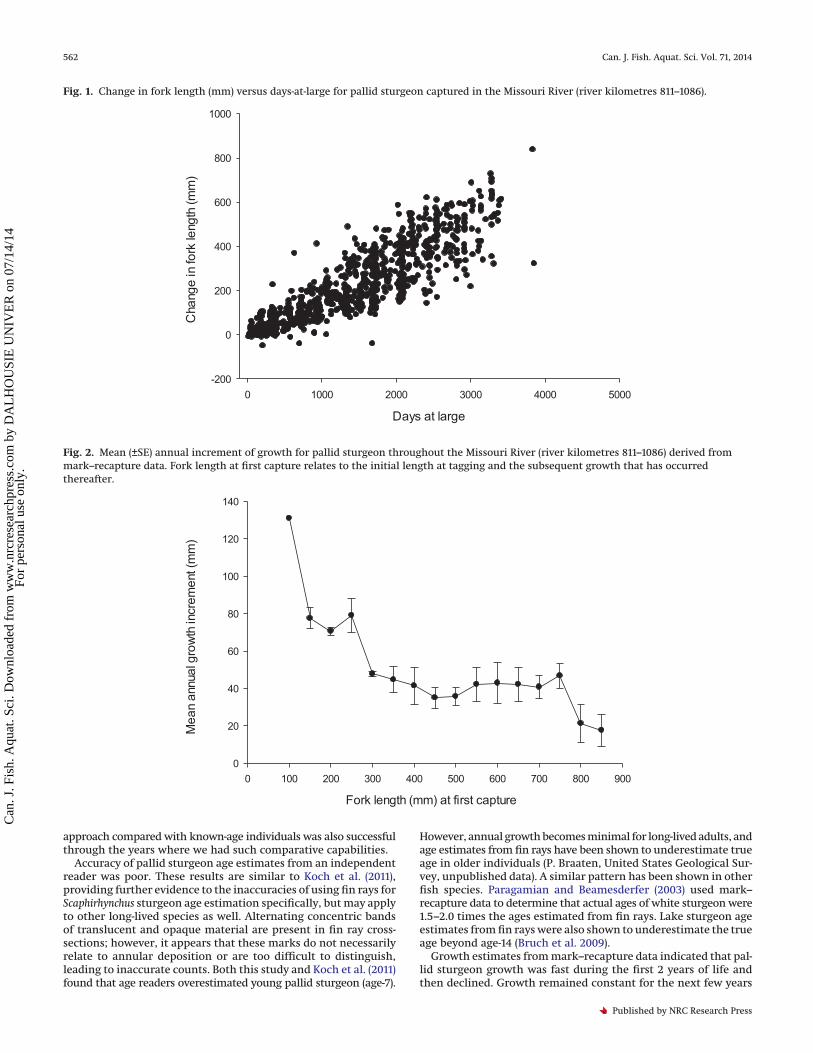
ResultsThere were 808 pallid sturgeon mark–recapture events that
were at large for 30 to 3855 days (Fig. 1). Annual growth was largestfor small pallid sturgeon (<300 mm) and declined to approxi-mately 40 mm for pallid sturgeon between 300 and 750 mm FL(Fig. 2). As pallid sturgeon approached maturity (�800 mm; Keenlyneand Jenkins 1993), annual growth declined to approximately 20 mm(Fig. 2). The estimate of mean annual growth increment varied bythe method of calculation (P < 0.001; Fig. 3). Adjusted pairwisecomparisons (Bonferroni correction; � = 0.008) indicated thatback-calculated growth estimates from the fin ray aging structurewith and without prior knowledge of age were different (P = 0.002).Back-calculated growth with knowledge of age resulted in agrowth trajectory that declined much faster at larger sizes thangrowth that was observed from mark–recapture (P = 0.003). How-ever, back-calculated growth without prior knowledge of age dis-played a similar growth trajectory as the mark–recapture data(P = 0.331), presumably because of a large degree of age overesti-mation (Table 1). Both of these procedures (i.e., mark–recaptureand back-calculations without knowing age) were not signifi-cantly different (i.e., the slopes were similar) from the null hypoth-esis of annual, equal proportionate growth (mark–recapture, P = 0.01;back-calculated without age, P = 0.354).
Accuracy of pallid sturgeon age estimates from the indepen-dent reader was lower than results reported by Koch et al. (2011)(28%–41% among three readers). The reader’s exact accuracy was13% compared with the true ages of pallid sturgeon and increasedto 43% within 1 year and 72% within 2 years of the true age. Similarto Koch et al. (2011), the reader generally overestimated the trueages because of the difficulty in discerning true annuli from pre-sumed false annuli. Age estimates varied from 1 to 14, and thelargest discrepancy in age was 8 years (Table 1).
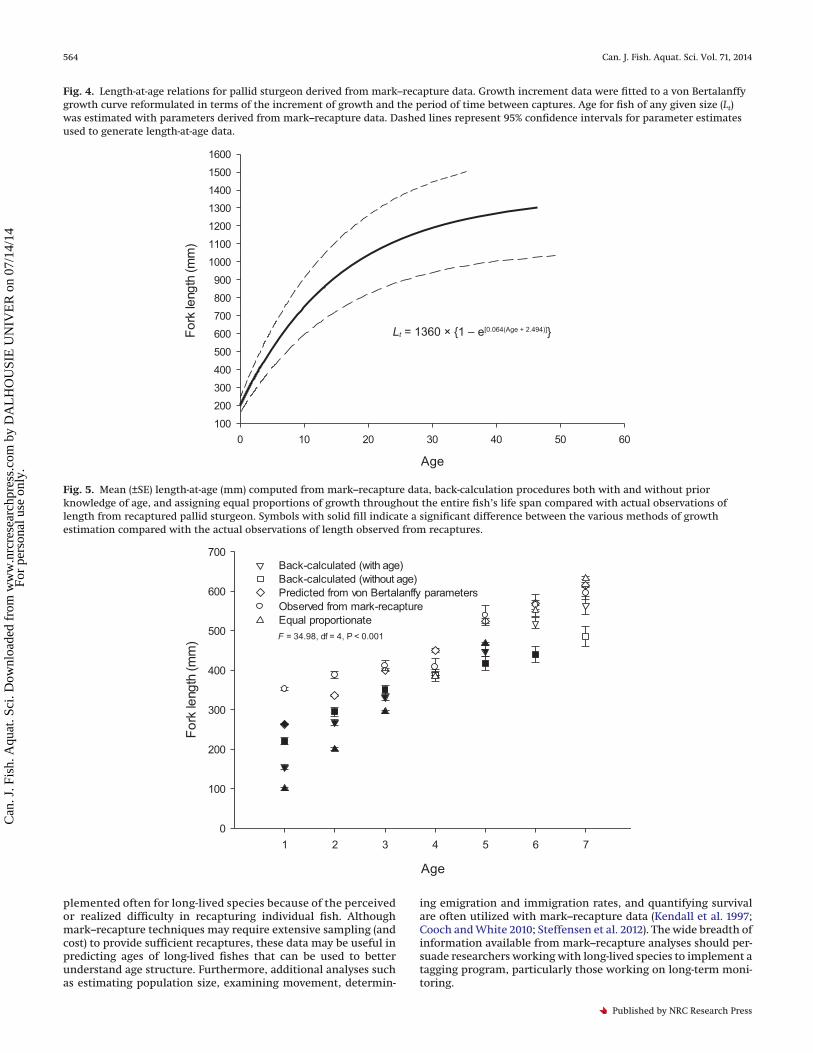
Age for pallid sturgeon of any given size was estimated withparameters derived from mark–recapture data (Fig. 4). The vonBertalanffy growth curve appeared similar to the actual lengthmeasurements observed from mark–recapture data. Mean length-at-age for all other calculation procedures was generally lowerthan the observed mark–recapture data (Fig. 5). Pairwise compar-isons of mean length observed from captured fish of all ageclasses, except age-1, were similar to the predicted length-at-agefrom the von Bertalanffy equation.
DiscussionWe used a combination of known-age pallid sturgeon (hatchery-
reared) and mark–recapture data to corroborate previous at-tempts at absolute age validation and to determine growth ratesfor pallid sturgeon in the lower Missouri River. Our results suggestthat mark–recapture data accurately portrays the growth trajec-tory of subadult, known-age pallid sturgeon and provides a meansto estimate and validate basic rate functions related to growth.Our predictions of ages of individuals using the mark–recapture
Hamel et al. 561
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
AL
HO
USI
E U
NIV
ER
on
07/1
4/14
For
pers
onal
use
onl
y.
approach compared with known-age individuals was also successfulthrough the years where we had such comparative capabilities.
Accuracy of pallid sturgeon age estimates from an independentreader was poor. These results are similar to Koch et al. (2011),providing further evidence to the inaccuracies of using fin rays forScaphirhynchus sturgeon age estimation specifically, but may applyto other long-lived species as well. Alternating concentric bandsof translucent and opaque material are present in fin ray cross-sections; however, it appears that these marks do not necessarilyrelate to annular deposition or are too difficult to distinguish,leading to inaccurate counts. Both this study and Koch et al. (2011)found that age readers overestimated young pallid sturgeon (age-7).
However, annual growth becomes minimal for long-lived adults, andage estimates from fin rays have been shown to underestimate trueage in older individuals (P. Braaten, United States Geological Sur-vey, unpublished data). A similar pattern has been shown in otherfish species. Paragamian and Beamesderfer (2003) used mark–recapture data to determine that actual ages of white sturgeon were1.5–2.0 times the ages estimated from fin rays. Lake sturgeon ageestimates from fin rays were also shown to underestimate the trueage beyond age-14 (Bruch et al. 2009).
Growth estimates from mark–recapture data indicated that pal-lid sturgeon growth was fast during the first 2 years of life andthen declined. Growth remained constant for the next few years
Fig. 1. Change in fork length (mm) versus days-at-large for pallid sturgeon captured in the Missouri River (river kilometres 811–1086).
Days at large
0 1000 2000 3000 4000 5000
Cha
nge
in fo
rk le
ngth
(mm
)
-200
0
200
400
600
800
1000
Fig. 2. Mean (±SE) annual increment of growth for pallid sturgeon throughout the Missouri River (river kilometres 811–1086) derived frommark–recapture data. Fork length at first capture relates to the initial length at tagging and the subsequent growth that has occurredthereafter.
Fork length (mm) at first capture
0 100 200 300 400 500 600 700 800 900
Mea
n an
nual
gro
wth
incr
emen
t (m
m)
0
20
40
60
80
100
120
140
562 Can. J. Fish. Aquat. Sci. Vol. 71, 2014
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
AL
HO
USI
E U
NIV
ER
on
07/1
4/14
For
pers
onal
use
onl
y.
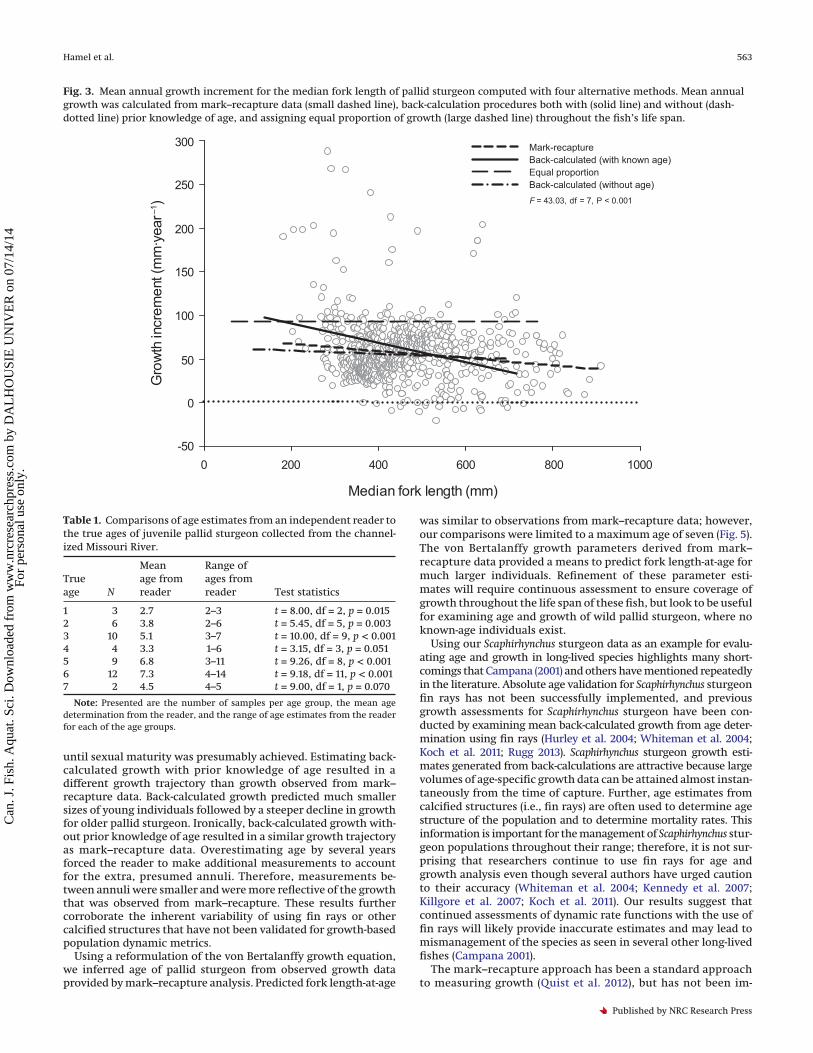
until sexual maturity was presumably achieved. Estimating back-calculated growth with prior knowledge of age resulted in adifferent growth trajectory than growth observed from mark–recapture data. Back-calculated growth predicted much smallersizes of young individuals followed by a steeper decline in growthfor older pallid sturgeon. Ironically, back-calculated growth with-out prior knowledge of age resulted in a similar growth trajectoryas mark–recapture data. Overestimating age by several yearsforced the reader to make additional measurements to accountfor the extra, presumed annuli. Therefore, measurements be-tween annuli were smaller and were more reflective of the growththat was observed from mark–recapture. These results furthercorroborate the inherent variability of using fin rays or othercalcified structures that have not been validated for growth-basedpopulation dynamic metrics.
Using a reformulation of the von Bertalanffy growth equation,we inferred age of pallid sturgeon from observed growth dataprovided by mark–recapture analysis. Predicted fork length-at-age
was similar to observations from mark–recapture data; however,our comparisons were limited to a maximum age of seven (Fig. 5).The von Bertalanffy growth parameters derived from mark–recapture data provided a means to predict fork length-at-age formuch larger individuals. Refinement of these parameter esti-mates will require continuous assessment to ensure coverage ofgrowth throughout the life span of these fish, but look to be usefulfor examining age and growth of wild pallid sturgeon, where noknown-age individuals exist.
Using our Scaphirhynchus sturgeon data as an example for evalu-ating age and growth in long-lived species highlights many short-comings that Campana (2001) and others have mentioned repeatedlyin the literature. Absolute age validation for Scaphirhynchus sturgeonfin rays has not been successfully implemented, and previousgrowth assessments for Scaphirhynchus sturgeon have been con-ducted by examining mean back-calculated growth from age deter-mination using fin rays (Hurley et al. 2004; Whiteman et al. 2004;Koch et al. 2011; Rugg 2013). Scaphirhynchus sturgeon growth esti-mates generated from back-calculations are attractive because largevolumes of age-specific growth data can be attained almost instan-taneously from the time of capture. Further, age estimates fromcalcified structures (i.e., fin rays) are often used to determine agestructure of the population and to determine mortality rates. Thisinformation is important for the management of Scaphirhynchus stur-geon populations throughout their range; therefore, it is not sur-prising that researchers continue to use fin rays for age andgrowth analysis even though several authors have urged cautionto their accuracy (Whiteman et al. 2004; Kennedy et al. 2007;Killgore et al. 2007; Koch et al. 2011). Our results suggest thatcontinued assessments of dynamic rate functions with the use offin rays will likely provide inaccurate estimates and may lead tomismanagement of the species as seen in several other long-livedfishes (Campana 2001).
The mark–recapture approach has been a standard approachto measuring growth (Quist et al. 2012), but has not been im-
Fig. 3. Mean annual growth increment for the median fork length of pallid sturgeon computed with four alternative methods. Mean annualgrowth was calculated from mark–recapture data (small dashed line), back-calculation procedures both with (solid line) and without (dash-dotted line) prior knowledge of age, and assigning equal proportion of growth (large dashed line) throughout the fish’s life span.
Gro
wth
incr
emen
t (m
m·y
ear–1
)
-50
0
50
100
150
200
250
300
Median fork length (mm)
0 200 400 600 800 1000
Mark-recapture Back-calculated (with known age)Equal proportionBack-calculated (without age)F = 43.03, df = 7, P < 0.001
Table 1. Comparisons of age estimates from an independent reader tothe true ages of juvenile pallid sturgeon collected from the channel-ized Missouri River.
Trueage N
Meanage fromreader
Range ofages fromreader Test statistics
1 3 2.7 2–3 t = 8.00, df = 2, p = 0.0152 6 3.8 2–6 t = 5.45, df = 5, p = 0.0033 10 5.1 3–7 t = 10.00, df = 9, p < 0.0014 4 3.3 1–6 t = 3.15, df = 3, p = 0.0515 9 6.8 3–11 t = 9.26, df = 8, p < 0.0016 12 7.3 4–14 t = 9.18, df = 11, p < 0.0017 2 4.5 4–5 t = 9.00, df = 1, p = 0.070
Note: Presented are the number of samples per age group, the mean agedetermination from the reader, and the range of age estimates from the readerfor each of the age groups.
Hamel et al. 563
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
AL
HO
USI
E U
NIV
ER
on
07/1
4/14
For
pers
onal
use
onl
y.
plemented often for long-lived species because of the perceivedor realized difficulty in recapturing individual fish. Althoughmark–recapture techniques may require extensive sampling (andcost) to provide sufficient recaptures, these data may be useful inpredicting ages of long-lived fishes that can be used to betterunderstand age structure. Furthermore, additional analyses suchas estimating population size, examining movement, determin-
ing emigration and immigration rates, and quantifying survivalare often utilized with mark–recapture data (Kendall et al. 1997;Cooch and White 2010; Steffensen et al. 2012). The wide breadth ofinformation available from mark–recapture analyses should per-suade researchers working with long-lived species to implement atagging program, particularly those working on long-term moni-toring.
Fig. 4. Length-at-age relations for pallid sturgeon derived from mark–recapture data. Growth increment data were fitted to a von Bertalanffygrowth curve reformulated in terms of the increment of growth and the period of time between captures. Age for fish of any given size (Lt)was estimated with parameters derived from mark–recapture data. Dashed lines represent 95% confidence intervals for parameter estimatesused to generate length-at-age data.
Lt = 1360 × {1 – e[0.064(Age + 2.494)]}
Age
0 10 20 30 40 50 60
Fork
leng
th (m
m)
100
200
300
400
500
600
700
800
900
1000
1100
1200
1300
1400
1500
1600
Fig. 5. Mean (±SE) length-at-age (mm) computed from mark–recapture data, back-calculation procedures both with and without priorknowledge of age, and assigning equal proportions of growth throughout the entire fish’s life span compared with actual observations oflength from recaptured pallid sturgeon. Symbols with solid fill indicate a significant difference between the various methods of growthestimation compared with the actual observations of length observed from recaptures.
Age
1 2 3 4 5 6 7
Fork
leng
th (m
m)
0
100
200
300
400
500
600
700Back-calculated (with age)Back-calculated (without age)Predicted from von Bertalanffy parametersObserved from mark-recaptureEqual proportionate F = 34.98, df = 4, P < 0.001
564 Can. J. Fish. Aquat. Sci. Vol. 71, 2014
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
AL
HO
USI
E U
NIV
ER
on
07/1
4/14
For
pers
onal
use
onl
y.

Our study provides an alternative approach to determininggrowth and estimating age without the use of a calcified structurethat may be subject to inaccuracies. Certainly, additional assess-ment of using the mark–recapture approach to measure age andgrowth are needed, but this approach looks to have promise com-pared with other less accurate or precise techniques. Using calci-fied structures may provide reliable results for some long-livedspecies if they can be validated; however, we recommend re-searchers understand the potential bias associated with thesestructures in long-lived species before adhering to conventionalmethods that have previously been conducted or are easier tocalculate. In instances where age determination for all ages ofinterest cannot be verified, mark–recapture appears to be a viablesolution for examining growth and has shown promise as a toolfor estimating ages in long-lived species that have calcified struc-tures too difficult to accurately and precisely read.
AcknowledgementsWe thank the pallid sturgeon population assessment crew from
the Nebraska Game and Parks Commission for their help in tag-ging and recapturing pallid sturgeon over a several-year period.We acknowledge N. Hogberg, K. Pope, R. Holland, and K. Eskridgefor reviewing earlier drafts of this manuscript. Funding wasprovided through Federal Sport Fish Restoration F-180-R andthe University of Nebraska – Lincoln School of Natural Resources.Additional resources for field collections were provided by US ArmyCorps of Engineers and the Pallid Sturgeon Population Assess-ment Project.
ReferencesBailey, R.M., and Cross, F.B. 1954. River sturgeons of the American genus
Scaphirhynchus: characters, distribution, and synonymy. Pap. Mich. Acad. Sci.Arts Lett. 39: 169–208.
Bajer, P.G., and Wildhaber, M.L. 2007. Population viability analysis of lowerMissouri River shovelnose sturgeon with initial application to the pallidsturgeon. J. Appl. Ichthyol. 23: 457–464. doi:10.1111/j.1439-0426.2007.00879.x.
Beamish, R.J., and McFarlane, G.A. 1983. The forgotten requirement for agevalidation in fisheries biology. Trans. Am. Fish. Soc. 112: 735–743. doi:10.1577/1548-8659(1983)112<735:TFRFAV>2.0.CO;2.
Bellwood, D.R., Hughes, T.P., Folke, C., and Nyström, M. 2004. Confronting thecoral reef crisis. Nature, 429: 827–833. doi:10.1038/nature02691. PMID:15215854.
Bemis, W.E., Findeis, E.K., and Grande, L. 1997. An overview of Acipenseriformes.Environ. Biol. Fishes, 48: 25–71. doi:10.1007/0-306-46854-9_4.
Black, B.A., Boehlert, G.W., and Yoklavich, M.M. 2005. Using tree-ring crossdat-ing techniques to validate annual growth increments in long-lived fishes.Can. J. Fish. Aquat. Sci. 62(10): 2277–2284. doi:10.1139/f05-142.
Boreman, J. 1997. Sensitivity of North American sturgeons and paddlefish tofishing mortality. Environ. Biol. Fishes, 48: 399–405. doi:10.1007/0-306-46854-9_28.
Bruch, R.M., Campana, S.E., Davis-Foust, S.L., Hansen, M.J., and Janssen, J. 2009.Lake sturgeon age validation using bomb radiocarbon and known-age fish.Trans. Am. Fish. Soc. 138: 361–372. doi:10.1577/T08-098.1.
Buckmeier, D.L., Smith, N.G., and Reeves, K.S. 2012. Utility of alligator gar ageestimates from otoliths, pectoral fin rays, and scales. Trans. Am. Fish. Soc.141: 1510–1519. doi:10.1080/00028487.2012.717520.
Campana, S.E. 2001. Accuracy, precision and quality control in age determina-tion, including a review of the use and abuse of age validation methods.J. Fish Biol. 59: 197–242. doi:10.1111/j.1095-8649.2001.tb00127.x.
Campana, S.E., and Neilson, J.D. 1985. Microstructure of fish otoliths. Can. J. Fish.Aquat. Sci. 42(5): 1014–1032. doi:10.1139/f85-127.
Coleman, F.C., Koenig, C.C., Huntsman, G.R., Musick, J.A., Eklund, A.M.,McGovern, J.C., Chapman, R.W., Sedberry, G.R., and Grimes, C.B. 2000. Long-lived reef fishes: the grouper–snapper complex. Fisheries, 25: 14–21. doi:10.1577/1548-8446(2000)025<0014:LRF>2.0.CO;2.
Cooch, E., and White, G. 2010. Using MARK: a gentle introduction. 8th ed. Cor-nell University, Department of Natural Resources, Ithaca, New York.
DeVries, D.R., and Frie, R.V. 1996. Determination of age and growth. In Fisheriestechniques. 2nd ed. Edited by B.R. Murphy and D.W. Willis. Am. Fish. Soc.,Bethesda, Md. pp. 483–512.
Dryer, M.P., and Sandvol, A.J. 1993. Pallid stureon recovery plan. US Fish andWildlife Service, Bismarck, North Dakota.
Dudgeon, D., Arthington, A.H., Gessner, M.O., Kawabata, Z., Knowler, D.J.,Lévêque, C., Naiman, R.J., Prieur-Richard, A.H., Soto, D., Stiassny, M.L.J., andSullivan, C.A. 2006. Freshwater biodiversity: importance, threats, status, and
conservation challenges. Biol. Rev. 81: 163–182. doi:10.1017/S1464793105006950.PMID:16336747.
Fabens, A.J. 1965. Properties and fitting of the von Bertalanffy growth curve.Growth, 29: 265–289. PMID:5865688.
Francis, R.I.C.C. 1990. Back-calculation of fish length: a critical review. J. FishBiol. 36: 883–902. doi:10.1111/j.1095-8649.1990.tb05636.x.
Hughes, T.P., Baird, A.H., Bellwood, D.R., Card, M., Connolly, S.R., Folke, C.,Grosberg, R., Hoegh-Guldberg, O., Jackson, J.B.C., Kleypas, J., Lough, J.M.,Marshall, P., Nyström, M., Palumbi, S.R., Pandolfi, J.M., Rosen, B., andRoughgarden, J. 2003. Climate change, human impacts, and the resilience ofcoral reefs. Science, 301: 929–933. doi:10.1126/science.1085046. PMID:12920289.
Hurley, K.L., Sheehan, R.J., and Heidinger, R.C. 2004. Accuracy and precision ofage estimates for pallid sturgeon from pectoral fin rays. N. Am. J. Fish. Man-age. 24: 715–718. doi:10.1577/M02-189.1.
Hutchings, J.A., and Reynolds, J.D. 2004. Marine fish population collapses: con-sequences for recovery and extinction risk. BioScience, 54: 297–309. doi:10.1641/0006-3568(2004)054[0297:MFPCCF]2.0.CO;2.
Jackson, N.D., Garvey, J.E., and Colombo, R.E. 2007. Comparing aging precisionof calcified structures in shovelnose sturgeon. J. Appl. Ichthyol. 23: 525–528.doi:10.1111/j.1439-0426.2007.00875.x.
Jager, H.I., Lepla, K.B., Van Winkle, W., James, B.W., and McAdam, S.O. 2010. Theelusive minimum viable population size for white sturgeon. Trans. Am. Fish.Soc. 139: 1551–1565. doi:10.1577/T09-069.1.
Keenlyne, K.D., and Jenkins, L.G. 1993. Age at sexual maturity of the pallidsturgeon. Trans. Am. Fish. Soc. 122: 393–396. doi:10.1577/1548-8659(1993)122<0393:AASMOT>2.3.CO;2.
Kendall, W.L., and Bjorkland, R. 2001. Using open robust design models to esti-mate temporary emigration from capture–recapture information. Biomet-rics, 57(4): 1113–1122. doi:10.1111/j.0006-341X.2001.01113.x. PMID:11764251.
Kendall, W.L., Nichols, J.D., and Hines, J.E. 1997. Estimating temporary emigra-tion using capture–recapture data with Pollock’s robust design. Ecology, 78:563–578. doi:10.1890/0012-9658(1997)078[0563:ETEUCR]2.0.CO;2.
Kennedy, A.J., Daugherty, D.J., Sutton, T.M., and Fisher, B.E. 2007. Populationcharacteristics of shovelnose sturgeon in the upper Wabash River, Indiana.N. Am. J. Fish. Manage. 27: 52–62. doi:10.1577/M06-038.1.
Killgore, K.J., Hoover, J.J., Kirk, J.P., George, S.G., Lewis, B.R., and Murphy, C.E.2007. Age and growth of pallid sturgeon in the free-flowing Mississippi River.J. Appl. Ichthyol. 23: 452–456. doi:10.1111/j.1439-0426.2007.00892.x.
Kirkwood, G.P. 1983. Estimation of von Bertalanffy growth curve parametersusing both length increment and age–length data. Can. J. Fish. Aquat. Sci.40(9): 1405–1411. doi:10.1139/f83-162.
Koch, J.D., Schreck, W.J., and Quist, M.C. 2008. Standardised removal and sec-tioning locations for shovelnose sturgeon fin rays. Fish. Manage. Ecol. 15:139–145. doi:10.1111/j.1365-2400.2008.00594.x.
Koch, J.D., Quist, M.C., Pierce, C.L., Hansen, K.A., and Steuck, M.J. 2009. Effects ofcommercial harvest on shovelnose sturgeon populations in the Upper Mis-sissippi River. N. Am. J. Fish Manage. 29: 84–100. doi:10.1577/M08-115.1.
Koch, J.D., Steffensen, K.D., and Pegg, M.A. 2011. Validation of age estimatesobtained from juvenile pallid sturgeon Scaphirhynchus albus pectoral finspines. J. Appl. Ichthyol. 27: 209–212. doi:10.1111/j.1439-0426.2010.01656.x.
Leveque, C., Oberdorff, T., Paugy, D., Stiassny, M.L.J., and Tedesco, P.A. 2008.Global diversity of fish (Pisces) in freshwater. Dev. Hydrobiol. 198: 545–567.doi:10.1007/978-1-4020-8259-7_53.
Paragamian, V.L., and Beamesderfer, R.C.P. 2003. Growth estimates from taggedwhite sturgeon suggest that ages from fin rays underestimate true age in theKootenai River, USA and Canada. Trans. Am. Fish. Soc. 132: 895–903. doi:10.1577/T02-120.
Pauly, D. 1979. Gill size and temperature as governing factors in fish growth: ageneralization of von Bertalanffy’s growth formula. Ber. Inst. f. MeereskundeUniv. Kiel. No. 63.
Pauly, D., Christensen, V., Guénette, S., Pitcher, T.J., Rashid Sumaila, U.,Walters, C.J., Watson, R., and Zeller, D. 2002. Towards sustainability in worldfisheries. Nature, 418: 689–695. doi:10.1038/nature01017. PMID:12167876.
Pikitch, E.K., Doukakis, P., Lauck, L., Chakrabarty, P., and Erickson, D.L. 2005.Status, trends and management of sturgeon and paddlefish fisheries. FishFish. 6: 233–265. doi:10.1111/j.1467-2979.2005.00190.x.
Pine, W.E., Pollock, K.H., Hightower, J.E., Kwak, T.J., and Rice, J.R. 2003. A reviewof tagging methods for estimating fish population size and components ofmortality. Fisheries, 28(10): 10–23. doi:10.1577/1548-8446(2003)28%5B10:AROTMF%5D2.0.CO;2.
Quist, M.C., Pegg, M.A., and DeVries, D.R. 2012. Age and growth. In Fisheriestechniques. 3rd ed. Edited by A.V. Alexander, D.L. Parrish, and T.M. Sutton.Am. Fish. Soc., Bethesda, Md. pp. 677–731.
R Development Core Team. 2013. R: a language and environment for statisticalcomputing [online]. R Foundation for Statistical Computing, Vienna, Austria.ISBN 3-900051-07-0. Available from http://www.R-project.org.
Rugg, M.L. 2013. Shovelnose sturgeon reproductive ecology in the lower PlatteRiver, Nebraska. Master’s thesis, University of Nebraska, Lincoln, Neb.
Scoppettone, G.G. 1988. Growth and longevity of the Cui-ui and longevity ofother catostomids and cyprinids in western North America. Trans. Am. Fish.Soc. 117: 301–307. doi:10.1577/1548-8659(1988)117<0301:GALOTC>2.3.CO;2.
Steffensen, K.D., Powell, L.A., and Pegg, M.A. 2012. Population size of hatchery-
Hamel et al. 565
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
AL
HO
USI
E U
NIV
ER
on
07/1
4/14
For
pers
onal
use
onl
y.

reared and wild pallid sturgeon in the lower Missouri River. N. Am. J. Fish.Manage. 32: 159–166. doi:10.1080/02755947.2012.661388.
Steffensen, K.D., Pegg, M.A., and Mestl, G.E. 2013. Population prediction andviability model for pallid sturgeon (Scaphirhynchus albus, Forbes and Richard-son, 1905) in the lower Missouri River. J. Appl. Ichthyol. 29: 984–989. doi:10.1111/jai.12277.
Trippel, E.A. 1995. Age at maturity as a stress indicator in fisheries. BioScience,45: 759–771. doi:10.2307/1312628.
Whiteman, K.W., Travnichek, V.H., Wildhaber, M.L., DeLonay, A., Papoulias, D.,and Tillett, D. 2004. Age estimation for shovelnose sturgeon: a cautionarynote based on annulus formation in pectoral fin rays. N. Am. J. Fish. Manage.24: 731–734. doi:10.1577/M03-090.1.
Winemiller, K.O. 2005. Life history strategies, population regulation, and impli-cations for fisheries management. Can. J. Fish. Aquat. Sci. 62: 872–885. doi:10.1139/f05-040.
566 Can. J. Fish. Aquat. Sci. Vol. 71, 2014
Published by NRC Research Press
Can
. J. F
ish.
Aqu
at. S
ci. D
ownl
oade
d fr
om w
ww
.nrc
rese
arch
pres
s.co
m b
y D
AL
HO
USI
E U
NIV
ER
on
07/1
4/14
For
pers
onal
use
onl
y.





























