Embed Size (px)
Citation preview

Publications of the University of Eastern Finland
Dissertations in Health Sciences
isbn 978-952-61-0507-9
Publications of the University of Eastern FinlandDissertations in Health Sciences
The relationship between strenuous
exercise and oxidative insults has
been implicated in muscle damage
and decreased muscle performance.
This study presents an overview
of oxidative stress-related events
and tissue protection mechanisms
in skeletal muscle after controlled
trials on a high-speed treadmill and
during the recovery in horses. The
antioxidant and tissue protective
effects of a thiol redox-modulator
α-lipoic acid are also studied.
dissertatio
ns | 070 | S
usa
nn
a Kin
nu
nen
| Oxidative S
tress in Skeletal M
uscle A
fter Acute E
xercise
Susanna Kinnunen
Oxidative Stress in Skeletal Muscle After Acute Exercise
Susanna Kinnunen
Oxidative Stress in Skeletal Muscle After Acute Exercise

SUSANNA KINNUNEN
��Oxidative�stress�in�skeletal�muscle�after�
acute�exercise��������������������
To�be�presented�by�permission�of�the�Faculty�of�Health�Sciences,�University�of�Eastern�Finland�for�public�examination�in�Mediteknia�auditorium,�Kuopio,��
on�Friday,�August�26th�2011,�at�12�noon��
Publications�of�the�University�of�Eastern�Finland�Dissertations�in�Health�Sciences�
Number�70��
Department�of�Medicine,�Institute�of�Biomedicine,�Physiology,��Faculty�of�Health�Sciences,�University�of�Eastern�Finland�
Kuopio�2011�

�������������
Kopijyvä�Oy�Kuopio,�2011�
�Series�Editors:�
Professor�Veli�Matti�Kosma,�M.D.,�Ph.D.�Institute�of�Clinical�Medicine,�Pathology�
Faculty�of�Health�Sciences��
Professor�Hannele�Turunen,�Ph.D.�Department�of�Nursing�Science�
Faculty�of�Health�Sciences��
Professor�Olli�Gröhn,�Ph.D.�A.I.�Virtanen�Institute�for�Molecular�Sciences�
Faculty�of�Health�Sciences��
Photo:�Terhi�Piispa�Helisten,�Equipose�(Equito�Ky)��
Distributor:�University�of�Eastern�Finland�
Kuopio�Campus�Library�P.O.Box�1627�
FI�70211�Kuopio,�Finland�http://www.uef.fi/kirjasto�
�ISBN�(print):�978�952�61�0507�9�ISBN�(pdf):�978�952�61�0508�6�
ISSN�(print):�1798�5706�ISSN�(pdf):�1798�5714�
ISSN�L:�1798�5706

III
Author’s�address:� Institute�of�Biomedicine,�Physiology�� � � � � � � University�of�Eastern�Finland�
KUOPIO,�FINLAND��
Supervisors:� � � Docent�Mustafa�Atalay,�M.D.,�M.P.H.,�Ph.D.�� � � � � � � Institute�of�Biomedicine,�Physiology�� � � � � � � University�of�Eastern�Finland�� � � � � � � KUOPIO,�FINLAND�� � � � � � � �
� � � � � � � Docent�Niku�Oksala,�M.D.,�Ph.D.�� � � � � � � Department�of�Vascular�Surgery�� � � � � � � Tampere�University�Hospital�
TAMPERE,�FINLAND�� � � � � � � �
� � � � � � � Professor�Emeritus�Osmo�Hänninen,�M.D.,�Ph.D.�� � � � � � � Institute�of�Biomedicine,�Physiology�� � � � � � � University�of�Eastern�Finland�� � � � � � � KUOPIO,�FINLAND��
� � � � � � � Professor�Emeritus�Pekka�Mäenpää,�M.D.,�Ph.D.�� � � � � � � Department�of�Biochemistry�� � � � � � � University�of�Eastern�Finland�� � � � � � � KUOPIO,�FINLAND��
� � � � � � � Professor�Chandan�K.�Sen,�Ph.D.�� � � � � � � Davis�Heart�and�Lung�Institute�
Department�of�Surgery�Ohio�State�University�Medical�Center�COLUMBUS,�OHIO,�USA�
�
Reviewers:�� � � Professor�Mustafa�Gül,�M.D.,�Ph.D.�� � � � � � � Department�of�Physiology�
Faculty�of�Medicine�Atatürk�University�ERZURUM,�TURKEY� ��
Professor�Emeritus�Ilkka�Alitalo,�D.V.M.�� � � � � � � Faculty�of�Veterinary�Medicine�� � � � � � � University�of�Helsinki�� � � � � � � HELSINKI,�FINLAND��
Opponent:� � � � Professor�Heikki�Kainulainen,�Ph.D.�� � � � � � � Department�of�Biology�of�Physical�Activity��� � � � � � � University�of�Jyväskylä�� � � � � � � JYVÄSKYLÄ,�FINLAND�

IV
�

V
Kinnunen Susanna
Oxidative stress in skeletal muscle after acute exercise
University of Eastern Finland, Faculty of Health Sciences, 2011
Publications of the University of Eastern Finland. Dissertations in Health Sciences 70,
2011, 73 p.
ISBN (print): 978‐952‐61‐0507‐9
ISBN (pdf): 978‐952‐61‐0508‐6
ISSN (print): 1798‐5706
ISSN (pdf): 1798‐5714
ISSN‐L: 1798‐5706
ABSTRACT:
The relationship between exercise and oxidative insults is well demonstrated and has been
implicated in muscle damage and decreased muscle performance. During heavy physical exercise
the oxygen flux to active skeletal muscle increases several folds, which enhances production of
reactive oxygen species (ROS). Studies examining racehorses in training have shown that the
most common reasons for retraction of the trotters from racecourses are musculoskeletal injuries.
This may be in part due to an exercise‐induced increase in ROS production.
This study presents an overview of ROS‐related insults and tissue protection mechanisms in
skeletal muscle after controlled trials on a high‐speed treadmill in horses. The antioxidant effects
of a natural thiol compound and redox‐modulator, α‐lipoic acid (LA), were also studied. In
addition to the acute effects of exercise, important data during the recovery period of up to 48
hours were also gained.
One bout of acute, submaximal exercise increased the concentrations of oxidised protein and
lipid products as well as the antioxidant capacity in plasma and in muscle. However,
submaximal exercise had no effect on muscle stress protein levels. LA supplementation reduced
the rate of free radical formation in skeletal muscle, activated stress protein induction and
simultaneously decreased the concentrations of oxidised protein and lipid peroxidation products.
LA appeared also to decrease ROS formation within muscle cells.
Based on this study, it can be concluded that trotters are prone to exercise‐induced oxidative
stress. However, LA supplementation may be used to improve performance by decreasing the
formation of free radicals and by increasing the rate of stress protein expression and inducing
antioxidant protection in horse skeletal muscle.
A broad‐based knowledge in exercise‐induced oxidative insults will improve our
understanding of oxygen‐related insults and their management. The results of this thesis support
previous studies that micronutrient supplementation can increase the total antioxidant status and
enhance cellular protection against exercise‐induced oxidative stress and muscle damage.
However, it should be kept in mind that prolonged mega‐dose supplementation of
micronutrients may attenuate the normal exercise‐related responses of tissues and blunt training‐
induced adaptations.
National Library of Medical Classification: QZ 180, WE 500
Medical Subject Headings: Oxidative stress; Muscle, Skeletal/physiology; Exercise; Horses

VI

VII
Kinnunen Susanna
Akuutin liikunnan aiheuttama hapetusstressi luustolihaksessa
Itä‐Suomen yliopisto, terveystieteiden tiedekunta, 2011
Publications of the University of Eastern Finland. Dissertations in Health Sciences 70 ,
2011, 73 p.
ISBN (print): 978‐952‐61‐0507‐9
ISBN (pdf): 978‐952‐61‐0508‐6
ISSN (print): 1798‐5706
ISSN (pdf): 1798‐5714
ISSN‐L: 1798‐5706
TIIVISTELMÄ:
Hapetusstressillä tarkoitetaan solujen altistumista reaktiivisille hapen muodoille, radikaaleille,
siinä määrin, että solun hapetusstressiä vastustavien puolustusjärjestelmien puskurointikyky
(antioksidatiivinen kapasiteetti) ylittyy. Liikunta lisää lihasten hapenkulutusta, jolloin myös
hapen radikaalien määrä ja hapetusstressi lisääntyvät, erityisesti silloin, kun hapenkulutus
lisääntyy nopeasti.
Tutkimuksessa selvitettiin liikunnan aiheuttaman hapetusstressin vaikutuksia luustolihaksissa.
Erityisen mielenkiinnon kohteena olivat stressiproteiinien ja antioksidatiivisten
puolustusjärjestelmien reaktiot keskiraskaaseen liikuntasuoritukseen. Tutkimuksen tavoitteena oli
selvittää pystytäänkö kudosten antioksidatiivisen puolustuksen kapasiteettia lisäämään
lämpöshokkiproteiinitasoja nostamalla ja samalla alentamaan liikunnan aiheuttamaa hapetusstressiä
α‐lipoaattilisää käyttäen.
Tutkimuksessa havaittiin, että jo yksittäinen, keskiraskas liikuntasuoritus aiheuttaa
hapetusstressiä valmennetuilla ravihevosilla. Akuutti liikunta nosti plasman ja lihasten
proteiinien ja lipidien hapettumistuotteiden määrää ja toisaalta myös kudosten antioksidatiivista
kapasiteettia, mutta ei vaikuttanut kudosten stressiproteiinitasoihin. Tutkimuksessa todettiin
rehuun lisätyn α‐lipoaatin vähentävän muodostuvien happiradikaalien määrää, nostavan
kudosten antioksidatiivista kapasiteettia ja stressiproteiinitasoja, lisäävän lihassolujen
oksidatiivista kapsiteettia sekä vähentävän lipidien ja proteiinien hapettumistuotteiden määrää
kudoksissa. α‐Lipoaatin havaittiin myös lisäävän lihaskudoksen pelkistyspotentiaalia, mikä
auttaa kudoksia suojautumaan liikunnan aiheuttamalta hapetusstressiltä.
Tutkimuksen perusteella voidaan todeta, että ravihevoset altistuvat hapetusstressille ja että
valmennuksen aikainen lipoaattilisä voi laskea hapetusstressitasoja mm. vähentämällä liikunnan
aikaisten vapaiden radikaalien muodostumisnopeutta ja jopa parantamaan suorituskykyä
lisäämällä lihasten oksidatiivista kapasiteettia ja aktivoimalla kudosten suojamekanismeja.
Antioksidanttilisiä käytettäessä on kuitenkin muistettava, että vapaat radikaalit ovat myös
maksimaalisen suorituksen edellytys. Liiallinen antioksidanttivalmisteiden käyttö voi häiritä
kudosten normaalien suojamekanismien toimintaa, jolloin happiradikaalien muodostumisen
negatiiviset seuraukset ylittävät niiden kudoksia suojaavat vaikutukset.
Yleinen suomalainen asiasanasto: rasitus; liikunta; lihakset – hapenkulutus

VIII

IX
To�your�surprise,�

X

XI
Acknowledgements��This�study�was�carried�out�at�the�Institute�of�Biomedicine,�University�of�Eastern�Finland�(former�Department�of�Physiology,�University�of�Kuopio),�the�Ylä�Savo�Vocational� Institute� in� Kiuruvesi,� Finland� (original� papers� III� and� IV)� and� at�Equine�Research�of�Agricultural�Research�Centre,�Ypäjä,�Finland�(original�papers�I�and�II).��
The� deepest� and� the� most� sincere� thanks� of� all� I� owe� to� my� principle�supervisor� docent� Mustafa� Atalay,� MD,� MPH,� Ph.D,� whose� endless� and� firm�support� and� guidance� have� dragged� me� over� the� hard� times� of� this� project.� I�could�not�have�done�this�without�you!�Thank�you!�
I�am�grateful�to�docent�Niku�Oksala�MD,�for�encouragement�and�constructive�guidance�along�the�study.�I�would�also�like�to�thank�my�retired�supervisors�Prof.�Em.�Osmo�Hänninen�and�Prof.�Em.�Pekka�Mäenpää,�as�well�as�Prof.�Chandan�K.�Sen�for�their�professional�expertise�and�support�at�all�stages�of�this�work.��
The�referees�of�this�study,�Prof.�Em.�Ilkka�Alitalo�from�University�of�Helsinki�(Finland)�and�Prof.�Mustafa�Gül�from�University�of�Atatürk�(Turkey)�are�greatly�acknowledged�for�their�constructive�criticism�and�suggestions.��
I�also�thank�Professor�Heikki�Kainulainen,�who�kindly�agreed�to�stand�as�my�honourable�opponent.�
The�contributions�of�Docent�David�Laaksonen�MD,�MPH,�Ph.D,�to�revise�the�endless�manuscripts�and�my�statistical�analyses�are�warmely�remembered.�
I�would�also�like�to�thank�the�following�persons�who�have�helped�me�during�these�years:�My�co�author�and�mentor� in� equine� science,�Seppo�Hyyppä�DVM,�from� Equine� College,� Ypäjä� (at� the� time� of� the� research:� MTT� Agricultural�Research� Centre,� Ypäjä)� for� making� these� studies� possible� and� Ms.� Marjatta�Lehtisaari�from�Ellab,�Ypäjä�for�all�the�technical�assistance�and�warm�hospitality�during�my�visits�as�well�as�all�the�help,�fun�and�encouragement.��
I� am� grateful� to� Zsolt� Radák� Ph.D,� from� Semmelweis� University,� Budapest�(Hungary)�for�expertise�and�collaboration,�and�Judit�Jakus�Ph.D,�from�Chemical�Researh� Center,� Hungarian� Academy� of� Science� (Budapest,� Hungary)� for� kind�assistance�and�for�skilled�introduction�to�EPR�method.�
I�would�like�to�thank�the�head�of�the�former�Department�of�Physiology,�Prof.�Matti�Närhi�and�the�head�of�Institute�of�Biomedicine,�Prof.�Anitta�Mahonen�for�all� the� support� and� flexibility.� Ms.� Merja� Atalay� and� my� collaborators� Marja�Leena�Hannila�MSc,�Jani�Lappalainen�MD�and�Mika�Venojärvi�Ph.D;�laboratory�technicians�Ms.�Taina�Vihavainen,�Ms.�Satu�Marttila�and�Ms.�Taija�Hukkanen�for�their�skilful� technical�assistance�and�all� the�help� they�extended��� it�would�have�taken� even� longer� to� finish� this� thesis� without� you.� And� Ms.� Eeva�Liisa�

XII
Palkispää,� Ms.� Riitta� Venäläinen,� trainer� Tero� Mäenpää� and� Purje�Ritari� for�getting�me�started�several�years�ago.�
I� am� grateful� to� Minna�Liisa� Heiskanen� Ph.D� and� her� staff� at� Equine�Information� Centre� for� everlasting� encouragement� and� support.� Matti� Notko,�Tapani� Kouvalainen� and� Leena� Rimpiläinen� for� the� opportunity� to� use� the�excellent� facilities� at� The� Ylä�Savo� Vocational� Institute� in� Kiuruvesi� for� lipoic�acid� studies;� the� students� and� the� staff� of� the� Ylä�Savo� Vocational� Institute� in�Kiuruvesi,� especially� Arja� Aalto� for� the� common� sense� and� friendship;� Timo�Vääränen,� Heli� Määttä,� Kirsi� Lustig,� Riina� Huusko,� Kirsi� Kettunen,� Satu�Riikonen,� Tuovi� Huttunen,� Katariina� Kastarinen,� Terho� Partanen� and� Alpo�Pellikka� for�all� the�help,�patience�and�everlasting� flexibility�during� these�years.�And� Kristiina� Palttala,� DVM,� from� Kiuruveden� Hevosklinikka� for� veterinary�support�and�guidance.��
I� am� thankful� to� Professor� Juhani� Syväoja� from� Department� of� Biology,� for�providing� the� facilities� in� Joensuu� Campus� and� to� Ms.� Terhi� Piispa�Helisten�(Equipose/Equito�Ky),�who�kindly�provided�the�photographs.�
I�would�like�to�thank�Juha�and�Ensio�Rissanen�for�re�introducing�me�to�horses�in�early�1990’s�and�Sari�for�making�it�such�a�pleasant�time.�I�wish�to�thank�Heli�for� sisterhood� and� recreation� as� well� as� my� dear� friends� Anneli� and� Heikki�Kosunen�for�all�the�help�and�support�during�our�daily�lives.�I�am�thankful�to�my�mother�in�law,� Sinikka� Hokkanen� and� my� late� father�in�law� Seppo� for�babysitting� during� emergencies;� and� to� my� friendly� host� in� Kuopio,� Annukka,�who�also�unexpectedly�passed�away�too�soon.�
The�warmest�thanks�I�owe�to�my�closest�family,�to�my�parents�Raija�and�Kari�Kinnunen,�my�sisters�Anna�and�Jenni�and�The�Rambœrs�for�the�fun,�support�and�togetherness�in�life.�Thank�you!�I�also�owe�a�lot�to�my�husband�Jari,�and�to�our�beautiful�daughters�Heta�Maija�and�Sanni�Kaisa,�who�have�the�unbeatable�ability�to�disturb�me�and�my�daily�routines.�I�love�you!�
This�study�has�been�financially�supported�by�National�Doctoral�Programme�of�Musculoskeletal� Disorders� and� Biomaterials,� TBDP� (former� TULES� Graduate�School� and� National� Graduate� School� of� Musculoskeletal� Disorders� and�Biomaterials,� TBGS)� and� Erkki� Rajakoski� Foundation,� Suomen� Hippos.� The�highly� specific� sampling� equipment� for� EPR� measurement� was� provided� by�Viinijärven�Metalli�–�thank�you!���
In�Kuopio,�August�2011���Susanna�Kinnunen�

XIII
List�of�the�original�publications�����This�dissertation�is�based�on�the�following�original�publications:����
I�� Kinnunen�S,�Hyyppä�S,�Lappalainen�J,�Oksala�N,�Venojärvi�M,�Nakao�C,� Hänninen� O,� Sen� C� K� and� Atalay� M.� Exercise�induced� oxidative�stress� and� muscle� stress� protein� responses� in� trotters.� Eur� J� Appl�Physiol�93:�496�501,�2005.�
�II�� Kinnunen� S,� Hyyppä� S,� Lehmuskero� A,� Oksala� N,� Mäenpää� P,�
Hänninen� O� and� Atalay� M.� Oxygen� radical� absorbance� capacity�(ORAC)� and� exercise�induced� oxidative� stress� in� trotters.�Eur� J� Appl�Physiol�95:�550�556,�2005.�
�III�� Kinnunen�S,�Hyyppä�S,�Oksala�N,�Laaksonen�D�E,�Hannila�M�L,�Sen�C�
K�and�Atalay�M.���Lipoic�acid�supplementation�enhances�heat�shock�protein� production� and� decreases� post� exercise� lactic� acid�concentrations� in�exercised�standardbred�trotters.�Res�Vet�Sci�87:�462�467,�2009.�
�IV�� Kinnunen�S,�Oksala�N,�Hyyppä�S,�Sen�C�K,�Radak�Z,�Laaksonen�D�E,�
Szabó� B,� Jakus� J� and� Atalay� M.� ��Lipoic� acid� modulates� thiol�antioxidant�defences�and�attenuates�exercise�induced�oxidative�stress�in�standardbred�trotters.�Free�Radic�Res�43(8):�697�705,�2009.�
��The�publications�were�adapted�with�the�permission�of�the�copyright�owners.��

XIV

XV
Contents 1 INTRODUCTION 1
2 REVIEW OF THE LITERATURE 3
2.1 Free radicals and reactive oxygen species 3
2.2 Oxidative stress and acute exercise 4
2.3 Markers of exercise‐induced oxidative stress 5
2.3.1 Detection of free radicals 5
2.3.2 Lipid peroxidation 6
2.3.3 Protein carbonylation 7
2.4 Antioxidant defences and other protective mechanisms
against exercise‐ induced oxidative stress 9
2.4.1 Total antioxidant capacity 9
2.4.2 Thiols as antioxidants 11
2.4.3 Other enzymatic and non‐enzymatic antioxidants 14
2.4.4 Stress protein response 15
2.5 Athletic horse and exercise‐induced oxidative stress 16
2.5.1 Exercise‐induced oxidative damage in horses 18
2.5.2 The effects of antioxidant supplementations in
sport horses 22
2.5.3 α‐Lipoic acid supplementation and horse 25
3 AIMS AND OBJECTIVES 27
4 METHODS 28
4.1 Subjects 28
4.2 Dietary assessments 28
4.2.1 α‐Lipoic acid supplementation 29
4.3 Exercise procedures 29
4.4 Sample collection 30
4.5 Biochemical methods 30
4.6 Statistical methods 32
5 RESULTS 33
5.1 Effects of acute exercise on tissue protection
mechanisms in exercising trotters (Original paper I) 33

XVI
5.2 Effects of acute exercise on biomarkers of oxidative stress
in exercising trotters (Original paper II) 34
5.3 Effects of LA supplementation on tissue protection
mechanisms after strenuous exercise in trotters (Original
paper III) 35
5.4 Effects of LA supplementation on muscle free radical
production, thiol status and certain markers of oxidative
stress after strenuous exercise in trotters (Original paper IV) 36
6 DISCUSSION 38
6.1 Free radical production during exercise 38
6.2 Thiol redox control 39
6.2.1 Glutathione redox‐status and related enzymes 40
6.2.2 Thioredoxin system 41
6.3 Lipid peroxidation 41
6.4 Protein oxidation 43
6.4.1 Thioredoxin system and protein carbonylation 44
6.5 Total antioxidant capacity 45
6.6 Stress protein response 47
6.6.1 Lipid peroxidation, protein oxidation and induction
of HSPs 48
6.6.2 Interaction between increased oxidative
capacity and HSP induction 49
7 FUTURE ASPECTS 50
8 CONCLUSIONS 51
9 REFERENCES 52
ORIGINAL PUBLICATIONS

XVII
Abbreviations�4�HNE� � � � � 4�hydroxy�noneal�ABTS� � � � � 2,2��azino�di�[3�ethylbenzthiazioline�sulphonate]�AUC�� � � � � area�under�curve�BALF� � � � � broncho�alveolar�lavage�fluid�CAT�� � � � � catalase�DCFH�� � � � � dichlorofluoroscein��DHLA� � � � � dehydrolipoic�acid�DNA� � � � � deoxiribonucleic�acid�EPR� � � � � � electron�paramagnetic�resonance�ESR� � � � � � electron�spin�resonance�FRAP� � � � � ferric�reducing�antioxidant�power�GPx� � � � � � glutathione�peroxidase�GRd�� � � � � glutathione�reductase�GRR�� � � � � glutathione�redox�ratio�GSH/GSSG� � � glutathione�(reduced/oxidised)�GST� � � � � � glutathione�S�transferase�H2O2�� � � � � hydrogen�peroxide�HO�1� � � � � hemioxygenase�1�HSF� � � � � � heath�shock�factor�HSP27� � � � � heat�shock�protein�27�HSP70� � � � � heat�shock�protein�70�HSP90� � � � � heat�shock�protein�90�HSPs� � � � � heat�shock�proteins�kDa� � � � � � kilodalton�LA� � � � � � lipoic�acid�LPO� � � � � � lipid�hydroperoxide�MDA� � � � � malondialdehyde�mRNA� � � � � messenger�RNA�NADP+/NADPH� nicotinamide�dinucleotide�phosphate�O2
��� � � � � � superoxide�anion�OBLA� � � � � onset�of�blood�lactate�accumulation�ORAC� � � � � oxygen�radical�absorbance�capacity�PCarb� � � � � proteincarbonyl�Prxs� � � � � � peroxiredoxins�RNS�� � � � � reactive�nitrogen�species�ROS� � � � � � reactive�oxygen�species�Se�� � � � � � selenium

XVIII
SET� � � � � � standardised�exercise�test�SOD�� � � � � superoxide�dismutase�TAC�� � � � � total�antioxidant�capacity�TAS� � � � � � total�antioxidant�status�TBARS� � � � � thiobarbituric�acid�reactive�substances�TEAC� � � � � trolox�equivalent�antioxidant�capacity�TGSH� � � � � total�glutathione��TRAP� � � � � total�radical�trapping�antioxidant�parameter�Trx� � � � � � thioredoxin�TrxRd� � � � � thioredoxin�reductase�VLa4� velocity�of�treadmill�resulting�in�blood�lactate�level�of�
�4�mmol/l�VO2max� � � � � maximal�oxygen�uptake�(ml�O2/kg/min)�

1�Introduction��Oxidative�metabolism�provides�an�enormous�advantage�to�succeed�in�the�great�struggle�for�existence�as�the�evolution�of�efficient�energy�production�allowed�the�development�of�complex�multicellular�organisms.�In�parallel�with�rising�oxygen�concentration�in�atmosphere,�organisms�were�obliged�to�either�escape�or�tolerate�oxygen� toxicity.� Consequently,� a� wide� variety� of� antioxidant� defence�mechanisms�evolved�to�cope�with�oxidative�insults�in�aerobic�organisms.�
It� was� first� suggested� in� early� 1950’s� (Gerschman� et� al.� 1954a)� that� the�damaging� effects� of� oxygen� could� be� attributed� to� the� formation� of� reactive�oxygen�species�(ROS).�Vigorous�production�of�ROS�is�implicated�in�aging�and�in�the�pathogenesis�of�a�number�of�diseases,� including�atherosclerosis,� cancer�and�diabetes� as� well� as� muscular� atrophy� and� chronic� inflammation� (Dröge� 2002).�Despite�all�this,�ROS�and�free�radicals�have�recently�been�found�to�be�beneficial�at�physiological�levels.�They�act�as�signalling�molecules�and�have�a�crucial�role�in�signal� transduction�within� cells� (Niess�and�Simon�2007;� Jackson�2008a;�Forman�2010).��
Excess�or�uncontrolled�production�of� reactive�oxygen�species�will�eventually�lead� to�oxidative� stress,� earlier�defined�as�a� stage�where�production�of�ROS�and�pro�oxidants� overwhelms� the� antioxidant� defence� mechanisms� of� the� cells� and�tissues�(Sies�1991).�The�more�recent,�complementary�definition�of�oxidative�stress�emphasises�the�role�of�disruption�of�thiol�redox�circuits,�resulting�in�imbalance�in�cell�signalling�and�dysfunctional�redox�control�(Jones�2008).��
Any� situation,� in�which� the� consumption�of� oxygen� is� increased,�potentially�results� in� acute� oxidative� stress� (Fisher�Wellman� and� Bloomer� 2009b).� The�relationship� between� exercise� and� oxidative� insults� is� well� demonstrated� and�implicated� in� muscle� damage� and� decreased� muscle� performance� (Reid� 2008;�Jackson�2009a;�Powers�et�al.�2010).�The�protective�mechanisms,�evolved�to�cope�with� the� reactive� oxygen� metabolites,� depend� primarily� on� the� synergism�between� several� endogenous� and� dietary� antioxidants� (Atalay� et� al.� 2006).�Epidemiological�studies�have�shown�that�adequate�daily�intake�of�micronutrients�is�associated�with� lower�cardiovascular�mortality� in�humans� (Bhupathiraju�and�Tucker�2011;�Núñez�Córdoba�and�Martínez�González�2011).��
The� series� of� studies� presented� in� this� thesis� examine� the� oxidative� stress�following� acute� exercise.� The� series� will� also� provide� an� overview� of� oxidant�related�insults�and�tissue�protection�mechanisms�in�the�exercising�horse.�Horses�possess�great� capacity� to� respond� to� repeated� stress� imposed�by� training� (Rose�and� Hodgson� 1994a);� still,� almost� thirty� years� ago,� studies� examining�

2
racehorses� in� training� showed� that� the� most� common� reason� to� impair�racehorse’s� career� and� to� cause� significant� economic� loss� for� the� industry� are�musculoskeletal� injuries� (Rose�et�al.�1983).�This� is�mainly�due� to� the�size�of� the�horse�and�the�force�generated�while�exercising�at�a� fast�pace.�Other� influencing�factors� may� include� training� methods,� length� of� the� race,� feeding� strategies� as�well�as�diverse�environmental�factors.�Studies�in�equine�exercise�physiology�and�sports� medicine� are� closely� related� to� the� animal� welfare� issues� that� are� in� the�centre� of� the� debate� these� days.� Physically� active� skeletal� muscles� produce� a�significant�amount�of�reactive�species�creating�a�mechanical�and/or�biochemical�load,� strong�enough� to� severely� injure�most�of�other� cells� (Clanton�et� al.� 1999).�The� significance� of� these� reactions� for� performance� and� athletic� capacity� is� not�fully� understood.� The� growing� awareness� and� broad�based� knowledge� in� both�oxidant�related� insults� and� related,� redox�regulated� adaptations� in� exercising�horse�will�eventually� redirect�horse� training�and�racing� industry� towards�more�sustainable�methods.�
An� innately� high� maximal� oxygen� uptake� makes� the� horse� a� good� research�subject� for� exercise�induced� oxidative� stress� studies.� It� has� also� been� reported�that� the� overtraining� syndrome� appears� to� be� similar� between� humans� and�horses� (Tyler� et� al.� 1996),� enabling� comparative� studies� (Tyler� et� al.� 1998).�However,�all�the�findings�are�not�applicable�into�humans�per�se�due�to�different�disease�predispositions�and�genetic�backgrounds�as�well�as�some�rather�unique�physiologic�adaptations�to�exercise.��

3
2�Review�of�the�literature��The� conventional� definition� of� oxidative� stress� as� a� “disturbance� in� the� pro�oxidant�antioxidant� balance� in� favour� of� the� former,� leading� to� potential�damage”�(Sies�1991)�has�recently�been�refined�(Jones�2008).�The�new�definition,�also�known�as�redox�hypothesis,�emphasises�the�role�of� the�disruption�of�cellular�reducing� circuits,� resulting� in� imbalance� in� cell� signalling� and� dysfunctional�redox�control� (Jones� 2008).� The� term� “redox”� originates� from� the� electron�transfer�from�the�reducing�agent�to�the�oxidising�agent.�
The�production�of�free�radicals�is�classically�considered�as�an�unavoidable�by�product�of�respiration,�mitochondria�being�the�major�source�for�the�production.�Radical�is�a�molecule�with�one,�unpaired�electron�on�its�valence�shell.�Molecular�oxygen�(O2)�has�two�unpaired�valence�electrons�making�it�especially�susceptible�to� radical� formation.� The� most� damaging� radicals� in� biological� systems� are�derived� from� oxygen,� and� are� known� collectively� as� reactive� oxygen� species,�ROS�(Halliwell�and�Gutteridge�2007).�It�was�first�suggested�by�Gerschman�et�al.�(1954a�and�1954b)�that�the�damaging�effects�of�oxygen�could�be�attributed�to�the�formation�ROS.�There�are�also�certain�non�radicals�classified�as�ROS,�known�as�non�radical�oxidants�(Table�1).��
The�cellular�redox�status�is�considered�as�a�balance�between�the�production�of�reactive� oxygen� and� nitrogen� species� (ROS� and� RNS� respectively),� and� their�removal� by� antioxidant� enzymes� and� small� molecular� weight� antioxidants�(Sarsour� et� al.� 2009).� The� redox�status� of� cells� and� different� tissues� is� closely�related�with�the�pathogenesis�of�several�diseases�and�appears�to�regulate�various�aspects�of�cellular�functions�in�healthy�cells.���2.1 FREE RADICALS AND REACTIVE OXYGEN SPECIES The� evolution� of� an� efficient� oxygen� delivery� system� has� placed� all� aerobic�organisms� under� trying� pro�oxidant� challenges� from� different� exo�� and�endogenous� sources.� Free� radicals� are� continuously� generated� during� normal�aerobic� life� in�mitochondria�as�oxygen� is� reduced�within� the�electron� transport�chain.�At� the�physiological� levels�ROS�regulate�many�genes� involved�with�cell�proliferation,� apoptosis� and� repair� mechanisms� for� proteins� and� nucleic� acids�(Sen�and�Packer�2000).��
The� primary� oxygen� free� radical,� superoxide�anion� (O2��)� is� dismutated� by�
superoxide�dismutases�(SODs),�generating�hydroperoxide�(H2O2).�H2O2�is�a�non�radical� able� to� diffuse� within� cell� and� across� cell� membranes� and� to� activate�

4
signalling� pathways� (Halliwell� and� Gutteridge� 2007).� H2O2� is� quickly�decomposed� by� catalase� or� peroxidases.� As� a� result,� tissue� redox� status� is�affected�by�ROS�resulting�in�oxidised�thiols�and�leading�to�activation�of�reducing�systems.� Excessive� production� of� H2O2� will� exhaust� the� peroxidative� and�reducing� systems� and� lead� to� an� imbalance� in� redox� balance.� Conversely,� a�disturbance�in�thiol�redox�balance�will�end�up�with�an�accumulation�of�H2O2.�
�Table 1�Selected free radicals and non-radical oxidants (modified from Halliwell and Gutteridge 2007). �Name Formula Comments/Examples � � �Free�radicals� � �Hydrogen�atom� H�� The�simplest�free�radical�Superoxide� O2
��� An�oxygen�centered�radical�Hydroxyl�� OH�� A�highly�reactive�oxygen�centered�radical�Thiyl� RS�� Radical�with�unpaired�electron�residing�on�sulphur�Peroxyl,�alkoxyl� RO2
�,�RO�� Oxygen�centered�radicals�formed�during�the�breakdown�of�organic�peroxides�
Non�radicals� � �Hydrogen�peroxide�
H2O2� One�of�the�most�powerful�known�oxidisers��
Hypochlorous�acid�
HOCl� Strong�oxidiser,�reacts�with�a�variety�of�biomolecules�and�is�able�to�cause�post�translational�modification�to�proteins�
Ozone� O3� Strong�oxidiser,�less�stable�than�its�diatomic�allotrope�O2�Singlet�oxygen� ½�O2� Strong�oxidiser,�very�reactive,�diamagnetic�form�of�O2�Peroxynitrite� ONOO���*)� An� oxidant� and� nitrating� agent� able� to� damage� variety� of�
biomolecules�� � �*)�Could�equally�well�be�called�a�“reactive�nitrogen�species,�RNS”��Recent� results� support� the� hypothesis� that� the� redox�cycle� within� the� cell�represents� a� regulatory� link� between� oxidative� metabolic� processes� and� cell�functions�(Burhans�and�Heintz�2009;�Sarsour�et�al.�2009).�The�production�of�ROS�is�therefore�to�be�considered�essential�for�optimal�physiological�functions.�� 2.2 OXIDATIVE STRESS AND ACUTE EXERCISE One� of� the� early� studies� provoked� a� strong� motivation� for� further� research� on�exercise�related� oxygen� toxicity.� By� using� the� electron� paramagnetic� or� spin�resonance�spectroscopy�(EPR�or�ESR�respectively)�for�the�direct�detection�of�free�radical� species� in� tissues,� (Davies� et� al.� 1982)� showed� that� exhaustive� exercise�resulted� in� a� two�� to� three�fold� increase� in� free� radical� concentrations� of� the�muscle�and�liver�of�rats�.��

5
During� heavy� physical� exercise� the� oxygen� flux� to� active� skeletal� muscle�increases� several� folds.� The� multifolded� oxidative� metabolism� is� closely�associated�with�enhanced�production�of�ROS�(Sen�et�al.�1994).�ROS�are�produced�in� multiple� sites� within� skeletal� muscle,� and� the� contractile� activity� of� fibres�exposes� them� to� higher� rates� of� oxidant� production� (Ferreira� and� Reid� 2008).�Intense�or�prolonged�muscular�exercise�may�result�in�oxidative�injury�to�muscle�lipids,� proteins� and� DNA� (Alessio� and� Goldfarb� 1988;� Reznick� et� al.� 1992a;�Lawler� et� al.� 1993b;� Lawler� et� al.� 1994;� Venditti� and� Di� Meo� 1996)� as� well� as�decreased�bioenergetic�enzyme�activities�and�muscle�fatigue�(Barclay�and�Hansel�1991;�Reid�et�al.�1992a;�Reid�et�al.�1992b;�Lawler�et�al.�1993a;�Lawler�et�al.�1993b;�Essig�and�Nosek�1997).���2.3 MARKERS OF EXERCISE-INDUCED OXIDATIVE STRESS The�direct�measurement�of�oxidative�stress� is�challenging,�due�to�the�instability�and�short�life�span�of�ROS.�However,�there�are�several�indirect�methods�to�assess�oxidative� stress.� These� techniques� measure� more� stable� products� formed� in�reactions� of� ROS� with� certain� (bio)molecules,� and� are� known� to� decrease� or�increase�as�a�result�of�oxidative�stress.��
Several�in�vitro�studies�have�revealed�that�the�oxidation�of�lipids�(Breusing�et�al.�2010)�and�proteins�(Radak�et�al.�2008)�provide�a�wide�range�of�breakdown�and�end� products� that� can� be� used� as� biomarkers� of� oxidative� stress� also� in� vivo�studies.�It�is�well�established�that�the�antioxidant�defence�systems�of�mammalian�tissues� are� capable� of� adaptation� in� response� to� chronic� exposure� to� oxidants�(Powers� and� Lennon� 1999).� The� severity� of� the� oxidative� stress� can� also� be�evaluated� indirectly� by� measuring� the� alterations� in� the� antioxidant� defence�system�of�an�organism.�These�will�be�further�discussed�in�following�chapters.���2.3.1�Detection�of�free�radicals�Free�radicals�are�continuously�produced�during�aerobic�metabolism.�Measuring�them� in� vivo� is� extremely� challenging,� and� generally� their� detection� relies� on�indirect� measurements.� However,� direct� assessment� for� the� production� is�possible� via� electron� paramagnetic� resonance� (EPR),� which� has� proved� to�provide,�with�or�without� spin� traps,� highly� sensitive�measurements� of� reactive�free� radicals� (McArdle� et� al.� 1999;� Stadler� et� al.� 2003;� Sun� et� al.� 2008;� Jackson�2008b),�also�during�exercise�and�training�(Toldy�et�al.�2005;�Traverse�et�al.�2006;�Bailey�et�al.�2007;�Davison�et�al.�2008;�Wray�et�al.�2009).�
Furthermore,� the�method,�based�on�dichlorofluoroscein� (DCFH)�fluorescence�microscopy�has�also�been�validated�to�measure�the�real�time�ROS�generation�in�single�isolated�muscle�cells�(Arbogast�and�Reid�2004).�There�are�also�two�other,�

6
less�common�techniques�introduced�in�literature:�radiolysis�and�flash�photolysis�(Knight�1998).��2.3.2�Lipid�peroxidation Membrane� lipids� are� the� major� targets� for� oxidative� insults� in� tissues.� Lipid�peroxidation�is�the�end�of�a�damaging�radical�chain�reaction�causing�substantial�damage� to� the� cell� membranes� (Sacheck� et� al.� 2003).� The� free� radical�mediated�peroxidation� of� lipids� has� received� great� deal� of� attention� in� connection� with�oxidative�stress� in�vivo� (Niki�and�Yoshida�2005).�Free�radicals�produced�during�lipid�peroxidation�have�some�very�local�effects,�because�of�their�short�life,�but�the�breakdown� products� of� lipid� hydroperoxidises� may� serve� as� “second�messengers”�of�oxidative�stress,�due�to�their�prolonged�half�life�and�their�ability�to�diffuse�from�their�site�of�formation.�
The� breakdown� products� of� lipid� peroxides,� mostly� aldehydes� such� as�malondialdehyde� (MDA)� or� 4�hydroxynonenal� (4�HNE),� have� received� much�attention�due�to�their�apparent�reactivity�and�toxicity�(Gueraud�et�al.�2010).�They�are� less� unstable� than� hydroperoxides� and� can� diffuse� from� their� site� of�formation� (Esterbauer� et� al.� 1991).� For� a� long� period� of� time,� MDA� and� HNEs�were� considered� as� toxic� end� products� of� lipid� peroxidation.� Quite� recently,�however,� it� has� been� clarified� that� they� have� a� powerful� biological� role� in� cell�signalling�under�both�pathological�and�physiological�conditions�(Gueraud�et�al.�2010).��
Since� MDA� is� generated� from� most� fatty� acids� with� more� than� two� double�bonds,�it�is�quantitatively�the�major�product�of�lipid�peroxidation�(Gueraud�et�al.�2010).� Compared� to� MDA,� HNE� is� formed� up� to� 80�fold� lower� concentrations.�However,�in�toxicological�terms,�HNE�appears�to�be�more�significant�than�MDA�due� to� its� higher� electrophilicity� and� its� implication� in� pathological� processes�(Gueraud� et� al.� 2010).� HNEs� appear� to� be� the� most� intensively� studied� lipid�peroxidation�products�so�far�(Poli�et�al.�2008).�
Additionally,� TBARS� (thiobarbituric� acid� reactive� substances)� is� a� well�established� assay� to� monitor� lipid� peroxidation� and� its� by�products,� including�lipid� hydroperoxides� and� aldehydes,� primarily� MDA� (Fisher�Wellman� et� al.�2009a).� However,� TBARS� assay� has� been� criticised� by� its� lack� of� specificity� in�complex� biological� samples� where� it� tends� to� react� with� several� other�biomolecules� (Sacheck� et� al.� 2003).� Therefore,� more� reliable� results� can� be�obtained�by�focusing�directly�on�MDA,�especially�from�tissue�homogenates.��� The� results� concerning� exercise�induced� lipid�peroxidation�are� controversial.�There� are� several� studies� reporting� no� changes� in� TBARS� (Alessio� et� al.� 2000;�Goldfarb�et�al.�2005a)�or�MDA�(Alessio�et�al.�2000;�Bloomer�et�al.�2005;�Goldfarb�et�al.�2005a;�Bloomer�et�al.�2006)�following�nearly�maximal�exercise.�This�has�been�

7
supported�with�exercising� rats,� too� (You�et�al.� 2005).�Still,� several�other� studies�have�reported�a�general� increase�in� lipid�peroxidation�in�relation�with�eccentric�exercise�(Sacheck�et�al.�2003;�Kingsley�et�al.�2006).��
The� differences� between� the� reports� are� likely� related� to� type� of� exercise�(concentric,�eccentric,�isometric�etc.)�and�its�intensity�as�well�as�duration,�alike�to�differences� in� sample� times,� training� status� of� subjects� and� to� the� chosen�analytical� methods,� differing� in� sensitivity� and� target� compounds.� The�differences�in�the�methodology�are�further�supported�by�the�study�of�Alessio�et�al.� (2000),� where� they� detect� no� change� in� levels� of� TBARS� after� exhaustive�aerobic�as�well�as�non�aerobic�isometric�exercise,�but�the�level�of�LPO�increased�significantly�after�both.��
Furthermore,�plasma�MDA�concentration�has�been�reported� to� increase�after�high�intensity�exercise�of�30�min�(Nakhostin�Roohi�et�al.�2008;�Seifi�Skishahr�et�al.�2008)� and� after� a� maximal� isometric� handgrip� test� (Rodrigues� et� al.� 2003)� in�healthy� untrained� men.� Therefore,� it� has� been� suggested� that� the� level� of� lipid�peroxidation�occurs�independently�of�exercise�intensity.�Instead�there�might�be�a�causal�link�between�hypoxia�and�levels�of�oxidative�damage�to�lipids�(Hoffman�et�al.�2007;�Møller�et�al.�2008).��
The� loss�of� fluidity� in�cell�membranes� is�closely�related�to� lipid�peroxidation�(Li�et�al.�1999;�Cazzola�et�al.�2003;�Portier�et�al.�2006;�Brzeszczynska�et�al.�2008;�Motta� et� al.� 2009).� The� overall� effects� of� lipid� peroxidation� are� to� damage�membrane� proteins� and� decrease� their� mobility,� leading� to� inactivation� of�enzymes� and� ion� channels� as� well� as� to� decreased� membrane� fluidity� and�increased� leakage� of� the� membrane� (Halliwell� and� Gutteridge� 2007).�Furthermore,� there� is� growing� evidence� that� products� generated� during� lipid�peroxidation� may� play� an� important� role� in� cellular� pathologies� and� lipid�peroxidation� is� therefore� linked� to� a�variety�of�disorders.�However,� the� role� of�lipid�peroxidation� in�signal� transduction�has�been�suggested,�mainly�expressed�by�4�HNE�(Yang�et�al.�2003).���2.3.3�Protein�carbonylation�Proteins�are�the�major�components�of�the�most�biological�systems�and�therefore�major� targets� for�ROS�attacks.�As�major�components�of�most�biological�system,�they� are� able� to� scavenge� 50�75%� of� free� radicals� (Davies� et� al.� 1999).� During�various� oxidative� stress� conditions,� protein� oxidation� results� in� inactivation� of�protein� functions� (Wong�et� al.� 2008);� furthermore,�proteins� exhibit� alteration�of�receptors,� enzymes� and� structural� proteins� (Levine� and� Stadtman� 2001).� Since�oxidative�modifications�generally�cause�loss�of�catalytic�or�structural�function�in�affected�proteins,�it�is�likely�that�the�increased�level�of�oxidised�proteins�observed�

8
during� recovery� will� have� deleterious� effects� on� cellular� and� organ� function�(Levine�and�Stadtman�2001;�Nyström�2005;�Suzuki�et�al.�2010).�
Generic� example� of� protein� modification� is� the� introduction� of� protein�carbonyl� (PCarb)� groups� into� protein� side� chains� by� a� variety� of� oxidative�pathways� including� the� oxidation� of� amino� acid� residues� or� reaction� with�primary� oxidation� products� such� as� MDA� and� 4�HNE� (Levine� and� Stadtman�2001;� Dalle�Donne� et� al.� 2006).� Carbonylation� is� an� irreversible,� non�enzymatic�modification�of�proteins�(Dalle�Donne�et�al.�2006)�and�protein�carbonyls�are�quite�stable� products� (Wong� et� al.� 2008).� However,� they� are� not� considered� as� only�damage�causing�but�can�also�serve�as�second�messengers�for�signal�transduction�(Wong�et�al.�2008;�Wong�et�al.�2010).��
Some�proteins�are�more�susceptible�to�oxidative�modifications�and�increases�in�carbonylation�than�others;�however,�the�set�of�proteins�that�become�carbonylated�differs�between�different�species�(Dalle�Donne�et�al.�2006).�Protein�carbonylation�has� become� a� widespread� indicator� of� oxidative� damage� in� cells� (Levine� and�Stadtman�2001;�Dalle�Donne�et�al.�2006;�Davies�and�Shringarpure�2006;�Wong�et�al.�2008).�
However,� it�has�recently�been�suggested�that�carbonylated�proteins�could�be�removed� from� cells� in� a� process� named� “decarbonylation”� (Wong� et� al.� 2008).�Intracellular� proteases� responsible� for� the� selective� degradation� of� oxidised�proteins� function� as� an� efficient� removal� and� repair� system� for� moderately�carbonylated� proteins� (Shringarpure� et� al.� 2003;� Dalle�Donne� et� al.� 2006).�However,� Wong� et� al.� (2008)� showed� that� decarbonylation� occurs� even� if� the�proteasomes� are� inhibited.� Nonetheless,� heavily� carbonylated� proteins� tend� to�form� high�molecular� weight� aggregates� that� are� resistant� to� degradation� and�accumulate� as� damaged� or� unfolded� proteins� possibly� inhibiting� proteasome�activity.�
Plasma�protein� carbonyls�have� been� reported� to� increase� following�different�types�of�exercise�(Bloomer�et�al.�2005;�Goldfarb�et�al.�2005a;�Bloomer�et�al.�2006;�Bloomer� et� al.� 2007;� Goldfarb� et� al.� 2007;� Lamprecht� et� al.� 2009).� There� are� a�couple� of� studies� (Bloomer� et� al.� 2005;� Morillas�Ruiz� et� al.� 2005)� showing� no�significant� changes� following� exercise� of� different� intensities.� These� differences�may� be� due� to� same� insufficiencies� in� test� protocol� and� sampling� times,� as�discussed�with�lipid�peroxidation�measures�in�chapter�2.3.2.��

9
2.4 ANTIOXIDANT DEFENCES AND OTHER PROTECTIVE MECHANISMS AGAINST EXERCISE-INDUCED OXIDATIVE STRESS Exposure�to�free�radicals�from�a�variety�of�sources�has�led�organisms�to�develop�a�series�of�defence�mechanisms�inducible�from�exposure�to�ROS�and�via�cellular�signal�molecules�(Cadenas�1997).�These�defence�mechanisms�involve�prevention�and�repair,�as�well�as�physical�and�antioxidant�defences.�A�broad�definition�of�an�antioxidant�as�any�substance,�that�when�present�at� low�concentration�compares�with�those�of�an�oxidisable�substrate�significantly�delays�or�prevents�oxidation�of�that�substrate�(Halliwell�and�Gutteridge�2007).��
Under� normal� conditions,� there� is� a� balance� between� the� activities� and� the�intracellular� levels� of� antioxidants� and� oxidants,� which� is� essential� for� the�survival� of� organisms� and� their� health� (Valko� et� al.� 2007).� However,� the�association� between� the� antioxidant� capacity� and� different� antioxidant�parameters�will�clarify�the�mechanisms�of�antioxidant�protection�and�the�effects�of�LA�on� the� tissue� thiol�antioxidant�network� in�relation�to�acute�exercise�were�not� clear� until� now.� Furthermore,� understanding� the� role� and� the� expression�patterns�of�stress�proteins�and�their�association�with�oxidative�damage�may�help�to�reduce�the�deleterious�effects�of�physical�exercise.�The�relative�importance�of�different�antioxidants�depends�also�upon�the�type�as�well�as�how�and�where�the�ROS� are� generated� (Halliwell� and� Gutteridge� 2007).� The� composition� of�antioxidant� defences� differs� between� tissues� and� cell�types� even� between� the�individual� cells� of� the� same� type.� Also� the� extracellular� fluids� have� different�protective�mechanisms�from�the�intracellular�compartments.���2.4.1�Total�antioxidant�capacity�Antioxidant� capacity� is� defined� as� the� ability� of� a� compound� to� reduce� pro�oxidants�(Prior�and�Cao�1999).�There�are�several�different�individual�antioxidants�and�antioxidant�systems�present� in�tissues�and�it� is�difficult� to�measure�each�of�them� separately.� Furthermore,� the� possible� interactions� among� different�antioxidants� in� vivo� could� also� make� the� measurement� of� any� individual�antioxidant�less�representative�of�the�overall�antioxidants�(Prior�and�Cao�1999).�
�The�increased�production�of�reactive�species�may�result�in�a�decrease�in�total�antioxidant�capacity� in�vivo;�however,� the�production�of� reactive�species�would�probably�have�to�be�very�extensive� to�disturb� the�system’s�steady�state� level�of�antioxidants� (Prior� and� Cao� 1999).� The� body’s� antioxidant� capacity� will� be�temporarily�decreased�as�its�components�are�used�to�quench�the�harmful�radicals�produced�(Fisher�Wellman�and�Bloomer�2009b).��

10
The�first�of�these�assays�becoming�popular�was�total�radical�trapping�antioxidant�parameter� (TRAP)� and� it� was� soon� followed� by� Trolox� equivalent� antioxidant�capacity� (TEAC),� ferric� reducing�antioxidant� power� (FRAP),� total� antioxidant�capacity� (TAC),� total� antioxidant� status� (TAS)� and� oxygen� radical� absorbing�capacity�(ORAC),�which�is�discussed�more�detail�later�in�this�thesis.��
There� is� a� wide� variety� of� commercial� kits� available� to� measure� total�antioxidant�capacity�of�diversity�of�body�fluids.�Although�most�of�the�assays�are�standardised� against� the� antioxidant� standard� Trolox,� these� distinct� methods�differ� largely� in� their� chemistry,� the� type� of� ROS� generated,� generation� site,�target�molecules�and�the�end�point�measured�(Pellegrini�et�al.�2003).��
The�FRAP�assay�actually�measures�the�ability�of�the�compound�to�reduce�Fe3+�to�Fe2+�and�either�of�them�is�able�to�cause�oxidative�damage�to�lipids�and�proteins�(Prior�and�Cao�1999).�It�should�be�remembered�that�a�compound�solely�reducing�Fe3+� is� not� considered� as� an� antioxidant� and� not� all� antioxidants� are� able� to�reduce� Fe3+.� For� instance,� FRAP� assay�does�not�measure�GSH,�one�of� the� most�important� antioxidants� in� vivo� (Prior� and� Cao� 1999).� However,� a� weak,� but�significant�correlation�has�been�reported�between�serum�ORAC�and�serum�FRAP�(Prior�and�Cao�1999).�
The�TAS�assay�relies�on�the�ability�of�plasma�antioxidant�substances�to�inhibit�the� oxidation� of� 2,2��azino�di�[3�ethylbenzthiazioline� sulphonate]� (ABTS).� The�capacity� of� the� antioxidants� in� the� sample� to� prevent� ABTS� oxidation� is�compared�with�that�of�trolox�(Berzosa�et�al.�2010).�
The�basic�reactions�of�TRAP�are�similar�to�those�of�ORAC�by�using�peroxyl�or�hydroxyl� radicals� as� reactive� species� and� PE� (phycoerythrin)� as� an� oxidisable�protein� substrate� (Prior� et� al.� 2005).� During� induction� period� (lag� phase)� the�oxidation� is� inhibited� by� the� plasma� antioxidants� and� compared� to� that� of� an�internal� standard� Trolox,� a� water�soluble� tocopherol� analogue,� reflecting� the�antioxidant�capacity�of�the�sample�(Prior�and�Cao�1999).�The�major�problem�with�the�original�TRAP�assay�is�that�the�oxygen�electrode�will�not�maintain�its�stability�over� the� period� of� time� required,� and� it� is� also� relatively� complex� to� perform�(Prior�and�Cao�1999).��
Taking� the� reaction� with� reactive� species� to� completion� and� using� the� AUC�(area�under� curve)� technique� for�quantification�makes� the�ORAC�assay�unique�when� compared� with� other� antioxidant� capacity� assays� (Prior� and� Cao� 1999).�ORAC� assay� represents� biologically� relevant� mechanism� measuring� both�lipophilic�and�hydrophilic�antioxidants�and�is�adapted�for�use�in�routine�quality�control�and�measurement�of�antioxidant�capacity�(Prior�et�al.�2005).�
ORAC� has� been� reported� to� increase� after� exhaustive� aerobic� and� isometric�exercise�(Alessio�et�al.�2000)�or�to�peak�at�72�hours�after�near�maximal,�eccentric�downhill�run�(Sacheck�et�al.�2003).�ORAC�levels�have�also�been�reported�not�to�

11
change� after� 30� min� submaximal� (i.e.� athlete is working below maximum effort) exercise�(Alessio�et�al.�1997)�or�four�weeks�of�cold�weather�field�training�at�high�intensity�in�trained�men�(Schmidt�et�al.�2002).���2.4.2�Thiols�as�antioxidants�The� fluctuation� in� the� redox�environment� of� a� cell� is� characteristic� to� cell� cycle�and� is� mediated� by� intracellular� changes� in� thiol� concentration.� Thiols� are�organic�sulphur�derivatives,�characterised�by�the�sufhydryl�residue�(�SH)�at�their�active� site.� Disulfide� linkage� S�S,� between� two� –SH� residues� is� an� important�determinant� of� protein� structure.� However,� conversion� of� –SH� groups� into�disulfides�is�one�of�the�earliest�markers�during�the�radical�mediated�oxidation�of�proteins�(Dean�et�al.�1997).�Another�characteristic�of�thiols�is�their�ability�to�act�as�reducing� agents� by� accepting� electrons,� leading� to� favourable� redox�milieu�within�cells.��
The�function�and�homeostasis�of�thiol�systems�contributes�to�the�most�central�feature�of�oxidative�stress.�Available�evidence� indicates� that� the�redox�sensitive�thiol�elements�function�in�cell�signalling�and�control�practically�all�aspects�of�life�(Jones� 2008).� Two� of� the� three� most� important� redox�systems� within� the�mammalian�cells�are�thiols.��Glutathione�system.�Glutathione�(��L�glutamyl�L�cysteinenyl�glycine,�GSH)�is�
the� most� abundant� thiol� in� animal� cells.� GSH� completes� various� biological�functions,� including� detoxifying� electrophilic� xenobiotics,� storing� and�transporting�cysteine,�modulating�redox�sensitive�signalling�processes,�reducing�ascorbate� and� tocopheroxyl� radicals� as� well� as� serving� as� a� cofactor� for�antioxidant� enzymes� (Sen� et� al.� 1998).� GSH� is� also� involved� in� cell� cycle�regulation�(Forman�et�al.�2009;�Zembron�Lacny�et�al.�2010).�
In�the�oxidant�thiol�reaction,�the�oxidant�is�neutralised�to�a�relatively�less�toxic�by�product� at� the� expense� of� the� reducing� power� of� GSH,� which� itself� gets�oxidised� to� a� disulfide� (GSSG).� As� a� result,� under� enhanced� oxidative� stress�conditions,� GSSG� content� increases� (Valko� et� al.� 2007).� There� is� a� specific�reductase,� glutathione� reductase� (GRd)� that� recycles� GSSG� back� to� GSH� at� the�expense� of� cellular� reducing� equivalent� NADPH.� The� glutathione� couple�(2GSH/GSSG)�represents�a�major�cellular�redox�buffer�and�can�therefore�be�used�as�an�indicator�for�the�redox�status�of�the�cell�(Dröge�2002).��
As�a� result�of�GRd�activity,� the� levels�of�GSSG�are�very� low� in�most� tissues,�and� the� ratio� of� GSH� to� GSSG� is� normally� very� high� (Niess� and� Simon� 2007;�Forman�et� al.� 2009).�While�acute�exercise� temporarily�decreases� the�GSH/GSSG�ratio�toward�an�augmented�pro�oxidant�state,�high�intensity�endurance�training�seems�to�be�capable�of�increasing�muscle�GSH�content�in�the�animal�model�(Sen�et�al.�1999a).�

12
In� response� to� oxidative� stress,� cells� try� to� maintain� glutathione� redox�state�through� diverse� mechanisms,� including� increasing� GRd� activity,� eliminating�excess�GSSG,�activating� the�de�novo�synthesis�of�glutathione�and/or�promoting�thiol� disulphide� exchange� (Filomeni� et� al.� 2002).� Level� of� oxidative� stress� is�increased,�the�GSSG�is�not�adequately�regenerated�to�GSH�and�the�excess�GSSG�is�secreted�out�of�cell�(Sen�et�al.�1993;�Forman�2010).�Prolonged�oxidative�stress,�when� the� cellular� systems� are� no� longer� able� to� counteract� the� ROS�mediated�insults,�will�cause�the�amount�of�free�GSH�to�decrease,�leading�to�irreversible�cell�degeneration�and�eventually�death�(Filomeni�et�al.�2002).�Thioredoxin� and� peroxiredoxin� systems.�The� thioredoxin� and� peroxiredoxin�
systems� represent� important�antioxidant�proteins� complementing�other� cellular�enzymic�and�non�enzymic�systems�within�cell� (Holmgren�2000).�The�thioredoxin�system�consists�of�the�two�antioxidant�enzymes�thioredoxin�(Trx)�and�NADPH�dependent�thioredoxin�reductase�(TrxRd).�As�the�intracellular�redox�homeostasis�is�ensured�primarily�by�GSH�and�Trx,�the�higher�concentrations�of�reduced�GSH�and� Trx� are� maintained� by� the� activity� of� GRd� and� TrxRd,� respectively.� In�addition� to� antioxidant� functioning� in� the� cell,� GSH�� and� Trx�systems� are�involved�in�cell�signalling�processes�(Dröge�2002).��
The� different� functions� of� Trx� are� dependent� upon� the� activity� of� TrxRd,�which� is� totally� selenium� dependent� (Arnér� 2009).� Thus,� the� effects� of�abnormalities� in� selenium�metabolism�should�not�only�affect� the� functions�and�levels�of� selenoproteins,�but�also� the�cellular� systems� linked� to�activities�of�Trx�(Arnér�2009).��
Peroxiredoxins� (Prxs)� are� a� group� of� peroxidases� reducing� intracellular�peroxides�(e.g.�H2O2)�with�the�thioredoxin�system�as�the�specific�electron�donor�(Zhang�et�al.�2009).�Prxs�are�known�to�mediate�signal�transduction�in�mammalian�cells� and� regulated� by� changes� in� redox�status� (Nordberg� and� Arner� 2001).� In�addition,� these� multifunctional� proteins� directly� reduce� some� ROS� as� well� as�refold�oxidised�proteins� and� low�molecular�weight� compounds� (Nordberg�and�Arner�2001).����Lipoic�acid.�As�an�essential�coenzyme�for�pyruvate�dehydrogenase�and���
ketoglutarate� dehydrogenase,� ��lipoic� acid� (LA)� plays� a� critical� role� in�mitochondrial� energy� metabolism,� ultimately� resulting� in� the� production� of�adenosine� triphosphate,�ATP� (Petersen�Shay�et� al.� 2008;� Satoh�et� al.� 2008).� It� is�found� in� most� prokaryotic� and� eukaryotic� microorganisms� as� well� as� in� many�plant�and�animal�tissues�(Sen�and�Packer�2000).�LA�has�an�eight–carbon�structure�that�contains�a�disulfide�bond�as�part�of�dithiolane�ring�with�five�carbon�tail.�It�is�also� known� as� thiotic� acid,� 1,2�dithiolane�3�pentanoic� acid,� 1,2�dithiolane�3�valeric�acid�and�6,8�dithioctic�acid.�At�physiologic�pH,�lipoic�acid�is�anionic�and�in�this�form�it�is�commonly�called�lipoate.��

13
LA�was�tentatively�classified�as�a�vitamin�after�its�isolation,�but�it�was�later�found�to�be�synthesised�by�animals�and�humans.�De�novo�synthesis�appears�to�supply�all� the� necessary� LA� for� the� intermediary� metabolism.� However,� digestive�enzymes�do�not�break�the�bond�between�LA�and�lysine�effectively�and�LA�can�be�absorbed� from� the� diet� (Schreier� 2005).� It� is� transiently� accumulated� in� many�tissues� and� carried� through� the� body� in� the� blood� stream� (Petersen� Shay� et� al.�2008;�Satoh�et�al.�2008).��
In� biological� systems,� LA� is� mainly� present� in� the� lysine�bound� form,�lipoyllysine�(Schreier�2005).�It�is�detected�in�various�natural�sources,�spinach�and�broccoli�being� the�richest�vegetables� (Sen�and�Packer�2000).�The�animal� tissues,�the�most�abundant�of� lipoyllysine,�are�determined�to�be�kidney,�heart�and�liver�(Sen�and�Packer�2000).�LA,�in�its�native�form,�contains�a�disulphide�bond.�LA�is�rapidly�converted�to�DHLA�inside�of�the�cell,�transported�out�of�the�cell�and�used�as�an�antioxidant.�LA�and�DHLA�do�not�only�act�directly�as�antioxidants�through�radical�quenching,�but�also�through�recycling�of�other�antioxidants�and�possibly�through� induction� of� increased� intracellular� levels� of� glutathione� (Packer� et� al.�1995;�Khanna�et�al.�1999a).��
As�both,�water��and�fat�soluble�compound,�LA�is�able�to�regenerate�water��and�fat�soluble� antioxidants,� such� as� vitamin� C� and� E� from� their� radical� forms.�Vitamin�E�is�the�major�chain�breaking�antioxidant�that�protects�membranes�from�lipid� peroxidation.� Evidence� for� vitamin� E� recycling� by� DHLA� has� come� from�number� of� studies� (Moini� et� al.� 2002;� Petersen� Shay� et� al.� 2008).� Furthermore,�DHLA� can� reduce� both� the� ascorbate� free� radical� and� dehydroascorbic� acid� to�ascorbate,�and�may�therefore�help�to�preserve�vitamin�C�either�inside�or�outside�cells,� although� this� has� not� been� demonstrated� (Jones� 2008).� It� has� been�suggested� that� DHLA� may� also� prevent� lipid� peroxidation� by� reducing�glutathione,�which� in� turn� recycles�vitamin�E� (Ghibu�et� al.� 2008).� Furthermore,�they� proposed� that� DHLA� protects� membranes� against� oxidation� by� recycling�ascorbate,�which�in�turn�recycles�vitamin�E.��
Thiolation�of�proteins�has�been�reported�to�be�mechanism�that�protects�against�oxidative�stress�and�regulates�the�function�of�some�proteins�(Packer�et�al.�1995).�It�has�also�been�suggested�that�LA�might�function�as�an�HSP�inducer�(Gupte�et�al.�2009).�The�role�of�LA�as�an�enhancer�of�HSP� induction� is� further�supported�by�previous� studies� in� which� LA� supplementation� appeared� to� up�regulate� the�mRNA� for� HSP90,� but� had� no� effect� at� the� protein� level� in� rat� kidney� tissue�(Oksala�et�al.�2007).�Furthermore,�there�is�evidence�that�LA�and�DHLA�may�have�effects� on� regulatory� proteins� and� on� genes� involved� in� normal� growth� and�metabolism.�
Glucose�transport�has�also�been�shown�to�be�enhanced�by�DHLA�in�a�number�of� systems� and� it� is� thought� that� this� stimulation� may� be� due� to� reduction� of�

14
sulfhydryl� groups� involved� in� the� regulation� of� insulin–stimulated� glucose�transport� (Packer� et� al.� 1995;� Khanna� et� al.� 1999a;� Lappalainen� et� al.� 2010).� In�addition,� LA� as� its� amide� form,� lipoamide,� is� an� essential� cofactor� in� the�multienzyme�complexes� that� catalyse� the�decarboxylation�of���keto�acids,� such�as�pyruvate� (to� acetyl� coenzyme�A)� and���ketoglutarate� (to� succinyl� coenzyme�A)�in�the�Krebs�cycle�(Halliwell�and�Gutteridge�2007).��2.4.3�Other�enzymatic�and�non�enzymatic�antioxidants�The� main� enzymatic� antioxidant� systems� are� superoxide� dismutase� (SOD),�catalase�(CAT)�and�glutathione�peroxidase�(GPx).�Metalloprotein�SOD�catalyses�the� reaction� of� superoxide� (O2��)� to� hydrogenperoxide� (H2O2).� Existing�isoenzymes� of� mammalian� SOD� can� be� characterised� by� their� metal� ions� and�cellular� locations� (Niess� and� Simon� 2007).� Acute� strenuous� exercise� has� been�shown�to�increase�SOD�activity�in�various�rodent�tissues�(Ji�et�al.�1998);�however,�it� appears� that� the� substantial� upregulation� of� SOD� system� in� trained� subjects�requires� cumulative� stimulation,� as� it� typically� occurs� during� chronic� exercise�and�training�(Niess�and�Simon�2007).��
�In�addition,�CAT�catalyses�the�decomposition�of�hydrogenperoxide�(H2O2)�to�water� and� oxygen� and� is� mainly� present� in� peroxisomes� of� most� mammalian�cells�(Niess�and�Simon�2007).�Compared�to�SOD�and�later�GPx,�activity�of�CAT�does�not� seem� to� respond� to� regular� training� (Powers�and�Hamilton�1999)� and�the� existing� reports� are� inconsistent� and� controversial.� This� controversy� may�result� from� the� lack� of� spesificity� of� the� CAT� assay� which� is� based� on� the�decomposition� of� H2O2.� On� the� other� hand,� CAT� response,� that� takes� place� in�peroxisomes,� may� also� be� acute� and� a� detectable� response� may� demand� more�strenuous�exercise.�Nevertheless�it�has�to�be�kept�in�mind�that�there�is�a�delicate�a� chain� reaction� among� antioxidant� ezymes� and� an� increase� in� SOD� activity�should� be� ensued� by� the� rise� in� the� activity� of� CAT,� GPx� or� other� peroxides�including�peroxiredoxins.���� One�specific�antioxidant�enzyme�is�hemioxygenase�(HO),�also�known�as�heat�shock� protein� 32,� exists� in� two� isoforms:� the� inducible� HO�1� (32� kDa)� and� the�constitutive� HO�2� (36� kDa).� It� protects� the� cell� against� oxidative� stress� by�reducing� intracellular� pool� of� free� iron� via� induction� of� ferritin� synthesis.� In�addition�HO�1�catalyses�the�initial�step�in�the�degradation�of�heme�to�bilirubin,�which� is� known� to� be� a� potent� water�soluble� antioxidant.� HO�1� protein� and�mRNA�are�strongly�induced�among�various�stressors�by�ROS,�and�it�functions�as�a� useful� marker� for� cellular� oxidative� stress� at� mRNA� level,� too� (Valko� et� al.�2007).�
Selenoprotein� GPx� catalyses� the� reduction� of� H2O2� and� a� wide� range� of�complex�organic�hydroperoxides�to�water�and�oxygen�by�using�GSH�as�electron�

15
donor�(Niess�and�Simon�2007;�Forman�2010).�Therefore�GPx�is�located�in�both�the�cytosol�and�mitochondrional�matrix� in�close�vicinity� to�cellular�sources�of�H2O2�formation.� Data� regarding� the� stimulating� impact� of� regular� training� in� GPx�activity� in� skeletal� muscle� are� reasonably� consistent� (Ji� et� al.� 1998).� It� has� been�shown� that� upregulation� of� GPx� acitivity� is� related� to� training� volume� and� it�appears� that� that� increased� GPx� acitivity� due� to� regular� exercise� is� limited� to�oxidative� skeletal�muscles� and� that� the� response�of�mitochondrial�GPx� is�more�pronounced�than�that�of�its�cytosolic�fraction�(Powers�and�Hamilton�1999).��
Selenium� (Se)� acts� as� a� cofactor� of� glutathione� peroxidase� and� is� therefore�necessary� to� maintain� full�strength� GSH�dependent� antioxidant� defense� (Burk�1983).� Dietary� deficiency� of� selenium� remarkably� lowers� tissue� glutathione�peroxidase� activity� and� thus� makes� the� tissue� more� susceptible� to� oxidative�damage� (Sen�and�Packer�2000).� Instead,�ebselen� (2�phenyl�1,2�benzisoselenazol�3(2H)�one)� a� synthetic,� non�toxic� seleno�organic� compound;� is� an� effective�scavenger� of� organic� hydroperoxides,� particularly� lipid� hydroperoxides�(Nakamura�et�al.� 2002).�Thus,� the�particular� interest� in�ebselen� is�of�mimicking�glutathione�peroxidase,�GPx�(Sies�1993;�Schewe�et�al.�1994;�Nakamura�et�al.�2002).�
There� are� also� several� low�molecular�mass� compounds� that� are� relevant� in�antioxidant�defence.�These�are�further�divided�into�endogenous�compounds,�like�bilirubin,� melatonin,� oestrogen,� coenzyme� Q10� and� uric� acid,� as� well� as�compounds� obtained� from� diet,� like� vitamins� E� and� C,� carotenoids� and�flavonoids� (Halliwell� and� Gutteridge� 2007).� However,� these� antioxidant�compounds�are�not�further�discussed�in�this�thesis.���2.4.4�Stress�protein�response�Heat� shock� proteins� (HSPs)� represent� cell� protective� and� antioxidant� system�against� stress� and� cell� damage� (Atalay� et� al.� 2004;� Morton� et� al.� 2009a).� HSP�expression� is� considered� as� an� adaptation�mechanism�and�a�marker� of� various�cellular�insults,�including�exercise�induced�oxidative�stress�(Powers�and�Lennon�1999;� Morton� et� al.� 2009a).� HSPs� are� a� group� of� highly� conserved� proteins,�expressed� in�both�pro�� and�eukaryotes.�Prevalent�HSPs�are�HSP27,�HSP60�and�the�family�of�HSP70�(Powers�and�Lennon�1999;�Niess�and�Simon�2007;�Morton�et�al.� 2009a).� Another� key� member� of� stress� proteins� 90�kDa� heath� shock� protein�(HSP90)�accounts� for�nearly�2%�of� total�protein� in�most�unstressed�cells� and� is�involved�in�essential�physiological�processes,�including�protein�trafficking,�signal�transduction,�and�specific�steroid�hormone�signalling�(Whitham�and�Fortes�2008;�Harkins�2009;�Michel�and�Vanhoutte�2010).�
In�response� to�environmental�stress,� the�major� function�of�HSPs� is� to�defend�cells� against� damage� by� binding� to� partially� denatured� proteins,� dissociating�

16
protein�aggregates,�regulating�the�correct�folding�and�co�ordinating�the�transport�of�newly�synthesised�polypeptides�(Fehrenbach�et�al.�2001).��
The� characteristic� feature� of� the� heat� shock� response� is� the� rapid� stress�induced�synthesis�of�HSPs.�The�transcriptional�response�is�mediated�by�a�family�of� heat� shock� transcription� factors� (HSFs)� that� are� activated� upon� specific�environmental� and� physiologic� events� resulting� in� HSP� gene� expression� and�eventually�HSP�synthesis�(Niess�and�Simon�2007;�Calderwood�et�al.�2009;�Anckar�and� Sistonen� 2010).� The� diversity� of� HSFs� expands� the� realm� of� regulatory�signals� for� the� inducible� expression� of� chaperones� and� other� stress�responsive�genes�(Morimoto�and�Santoro�1998).��
The�heat�shock�response�is�accelerated�by�a�variety�of�stressful�conditions,�like�elevated�core�temperature,�ischemia,�oxidative�stress,�and�increased�intracellular�calcium,�electro�mechanical�coupling,�stress�on�intermediate�filaments,�glycogen�and� ATP� depletion,� acidosis� and� oxidative� stress,� which� all� exists� during�exhaustive� and� prolonged� physical� exercise� (Essig� and� Nosek� 1997;� Liu� and�Steinacker�2001;�Morton�et�al.�2009b;�Anckar�and�Sistonen�2010).�Therefore� it� is�not�surprising�that�it�has�been�shown�already�in�the�late�1990s�that�exercise�up�regulates�the�expression�of�HSPs�(Kelly�et�al.�1996;�Essig�and�Nosek�1997;�Locke�1997).� Endurance� training� appears� to� induce� HSP� response� in� skeletal�(Thompson� et� al.� 2002;� Atalay� et� al.� 2004;� Moran� et� al.� 2004)� and� in� cardiac�muscle�(Powers�et�al.�2008;�Kavazis�2009;�Golbidi�and�Laher�2010).�The�induction�is�suggested�to�occur�in�an�intensity��and�duration�dependent�manner�(Ogawa�et�al.�2011).��
In� addition,� anesthesia� associated� muscle� disorder,� like� malignant�hyperthermia� (Denborough� 1998)� and� exertional� myopathies,� like�rhabdomyolysis� in� horse� (Aleman� et� al.� 2009)� and� PSE� (pale,� soft,� exudative)�meat�in�swine�and�poultry�are�stress�derived�and�therefore,�may�be�linked�also�to�stress�protein� induction�at� least� in� chicken� (Yu�et� al.� 2008;�Al�Aqil� and�Zulkifli�2009)�and�pigs�(Bao�et�al.�2009;�Yu�et�al.�2009).�Furthermore,�transport�stress�is�of�major�concern�and�predisposing�factor�for�stress�response�in�pigs�(Bao�et�al.�2009)�and�poultry�(Al�Aqil�and�Zulkifli�2009;�Dadgar�et�al.�2010).��2.5 ATHLETIC HORSE AND EXERCISE-INDUCED OXIDATIVE STRESS The� high� athletic� performance� of� the� horse� results� from� biomechanical� factors�and� integration� of� the� major� body� systems� delivering� energy� and� oxygen�throughout� the� body� (Rose� and� Hodgson� 1994a).� Selected� physiological�characteristics�of�human�athlete�and�horse�are�summarised�in�table�2.��
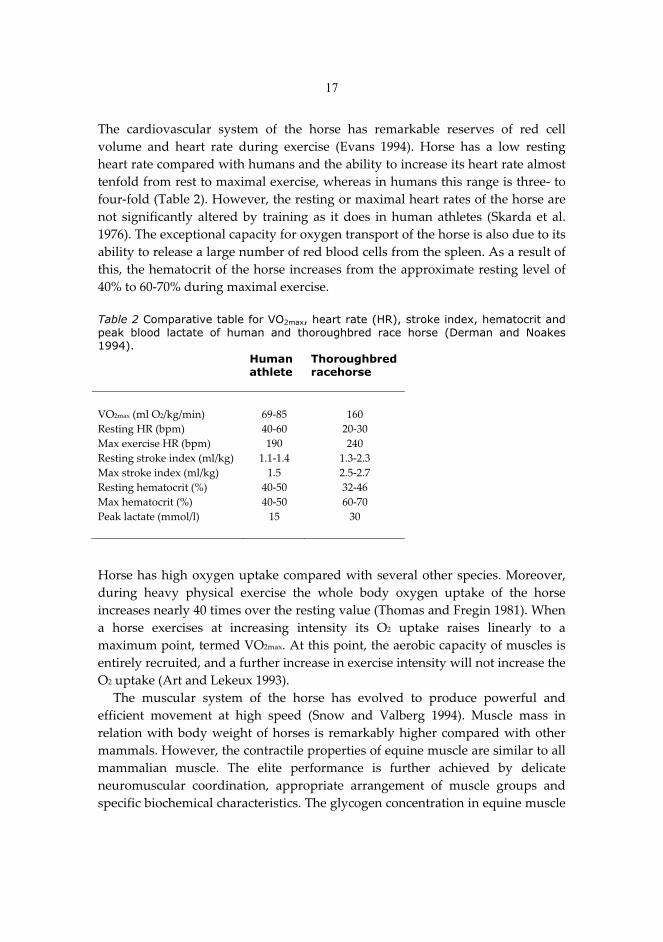
17
The� cardiovascular� system� of� the� horse� has� remarkable� reserves� of� red� cell�volume� and� heart� rate� during� exercise� (Evans� 1994).� Horse� has� a� low� resting�heart�rate�compared�with�humans�and�the�ability�to�increase�its�heart�rate�almost�tenfold�from�rest�to�maximal�exercise,�whereas�in�humans�this�range�is�three��to�four�fold�(Table�2).�However,�the�resting�or�maximal�heart�rates�of�the�horse�are�not� significantly� altered� by� training� as� it� does� in� human� athletes� (Skarda� et� al.�1976).�The�exceptional�capacity�for�oxygen�transport�of�the�horse�is�also�due�to�its�ability�to�release�a�large�number�of�red�blood�cells�from�the�spleen.�As�a�result�of�this,�the�hematocrit�of�the�horse�increases�from�the�approximate�resting�level�of�40%�to�60�70%�during�maximal�exercise.���Table 2 Comparative table for VO2max, heart rate (HR), stroke index, hematocrit and peak blood lactate of human and thoroughbred race horse (Derman and Noakes 1994). � Human
athlete Thoroughbred racehorse
� � �VO2max�(ml�O2/kg/min)� 69�85� 160�Resting�HR�(bpm)� 40�60� 20�30�Max�exercise�HR�(bpm)� 190� 240�Resting�stroke�index�(ml/kg)� 1.1�1.4� 1.3�2.3�Max�stroke�index�(ml/kg)� 1.5� 2.5�2.7� �Resting�hematocrit�(%)� 40�50� 32�46�Max�hematocrit�(%)� 40�50� 60�70�Peak�lactate�(mmol/l)� 15� 30�� � � �Horse�has�high�oxygen�uptake�compared�with�several�other�species.�Moreover,�during� heavy� physical� exercise� the� whole� body� oxygen� uptake� of� the� horse�increases�nearly�40�times�over�the�resting�value�(Thomas�and�Fregin�1981).�When�a� horse� exercises� at� increasing� intensity� its� O2� uptake� raises� linearly� to� a�maximum�point,�termed�VO2max.�At�this�point,�the�aerobic�capacity�of�muscles�is�entirely�recruited,�and�a�further�increase�in�exercise�intensity�will�not�increase�the�O2�uptake�(Art�and�Lekeux�1993).��
The� muscular� system� of� the� horse� has� evolved� to� produce� powerful� and�efficient� movement� at� high� speed� (Snow� and� Valberg� 1994).� Muscle� mass� in�relation�with�body�weight�of�horses� is� remarkably�higher�compared�with�other�mammals.�However,�the�contractile�properties�of�equine�muscle�are�similar�to�all�mammalian� muscle.� The� elite� performance� is� further� achieved� by� delicate�neuromuscular� coordination,� appropriate� arrangement� of� muscle� groups� and�specific�biochemical�characteristics.�The�glycogen�concentration�in�equine�muscle�

18
is�relatively�high,�approximately�50%�higher�than�in�human�muscle�(Lindholm�et�al.�1974a).��
Horses� possess� a� large� intramuscular� buffering� capacity� binding� and�removing� H+�ions,� hence� fighting� back� the� deleterious� effects� of� lactate�accumulation�during�intense�exercise.�This�buffering�capacity�includes�proteins,�bicarbonate,� inorganic� phosphate� and� carnosin,� all� enabling� horses� to� tolerate�higher�concentrations�of�lactate�in�muscles�(Hyyppä�and�Pösö�1998).�The�muscle�buffering�capacity�of�the�horse�is�much�greater�compared�with�humans�(Harris�et�al.�1990).��2.5.1�Exercise–induced�oxidative�damage�in�horses��In� horses� and� humans� there� is� a� linear� relationship� between� maximal� oxygen�uptake�and�speed�during�submaximal�exercise.�However,�in�horses,�the�maximal�oxygen�uptake� is�able� to� increase�up� to�30� to�40� folds�within�60�seconds�of� the�onset�of�exercise�(Derman�and�Noakes�1994).�Due�to�dense�capillary�network�of�the�regularly� trained�muscles�and�high�oxygen�uptake�of� the�horse,� the�oxygen�flux� in� the� active� peripheral� skeletal� muscle� fibres� may� increase� over� 100� fold�during� exercise.� It� has� been� estimated� that� from� 1� to� 5%� of� inhaled� oxygen� in�aerobes�will�form�reactive�oxygen�species,�ROS�(Halliwell�and�Gutteridge�2007).�Evidently,� this� can� expose� the� horse� to� oxidant�related� insults.� There� is,� to� our�knowledge,� only� one� study� using� electron� spin� resonance� (ESR)� to� detect�superoxide�scavenging�ability�in�equine�serum�after�racing�(race�length�was�not�mentioned)�and�submaximal�treadmill�exercise�as�well�as�after�5,�23�and�40�hours�of�transport�(Ishida�et�al.�1999).�Ishida�et�al.�(1999)�reported�no�change�following�exercise,�but�significant�increase�in�superoxide�scavenging�ability�after�23�and�40�hours�of�transport.�
There�is�a�wide�variety�of�studies�concerning�the�exercise�associated�oxidative�insults�in�horses�but�no�uniform�evidence�of�the�occurrence�of�exercise�induced�oxidative� stress� in� the� horse� in� general� (Mills� et� al.� 1996;� Balogh� et� al.� 2001;�Deaton�et�al.�2002;�Hoffman�et�al.�2002;�Kirschvink�et�al.�2002;�Marlin�et�al.�2002;�Hargreaves�et�al.�2002a;�2002b).�The�lack�of�unequivocal�knowledge�of�exercise�induced� oxidative� stress� is� mainly� due� to� the� variation� in� contributing� factors�(exercise� intensity,� diet,� training� history� and� level� of� fitness)� as� well� as� to� the�variable�specificity�and�sensitivity�of�different�analytical�methods.��Lipid� peroxidation� in� response� to� acute� exercise.� The� different� lipid�
peroxidation� products� have� been� measured� from� skeletal� muscle� and� from�plasma�in�horses.�The�amount�of�thiobarbituric�acid�reactive�substances�(TBARS)�and� malondialdehyde� (MDA)� have� been� reported� to� increase� post�exercise� in�plasma� (Brady� et� al.� 1978;� McMeniman� and� Hintz� 1992;� Mills� et� al.� 1996;�Chiaradia�et�al.�1998;�White�et�al.�2001;�Marlin�et�al.�2002)�as�well�as� in�muscle�

19
(Matsuki�et�al.�1991).�Furthermore,�there�are�two�reports�that�have�measured�no�change�in�plasma�MDA�or�TBARS�following�exercise�(Ono�et�al.�1990;�Siciliano�et�al.� 1996)� and� two� reporting�a�decrease� (Avellini� et� al.� 1999;�Balogh�et� al.� 2001).�Although�TBARS�and�MDA�are�well�established�methods�to�assess�the�intensity�of� lipid�peroxidation,� especially� the�TBARS�assay� is�questioned�due� to� its�non�specificity� (Sachdev� and� Davies� 2008).� Furthermore,� there� is� great� variation�between� the� individual� horses� and� horse� breeds,� training� background,� feeding�regimens�and�exercise�test�intensity�in�reported�studies.��
The� return� of� the� lipid� peroxidation� markers� back� to� the� basal� level� in� red�blood�cells�has�been�reported�to�occur�as�soon�as�after�a�one�hour�recovery�from�light�exercise�of�10�min� (Brady�et�al.�1978)�and�after�24�hours�of� recovery� from�more� strenuous� exercise� (Matsuki� et� al.� 1991;� Mills� et� al.� 1996).� Elevation� in�conjugated� dienes� suggests� increased� lipid� peroxidation� (Duthie� et� al.� 1990;�Alessio�1993;�Clarkson�1995).�There�is�one�study�reporting�increased�conjugated�dienes� after� exercise� in� horses;� however,� there� were� no� detectable� changes� in�plasma�TBARS�at�the�given�time�(Siciliano�et�al.�1996).��
Exercise�induced�rigidity�in�the�erythrocyte�membrane�has�been�suggested�to�contribute�to�the�increased�rate�of�lipid�peroxidation�in�man�(Brzeszczynska�et�al.�2008;� Berzosa� et� al.� 2010).� Exercise�induced� decrease� in� erythrocyte� membrane�fluidity� has� also� been� observed� in� horses� after� cross�training� and� standardised�exercise�test�(Avellini�et�al.�1995;�Portier�et�al.�2006;�de�Moffarts�et�al.�2007).�The�loss� of� fluidity� in� erythrocyte� cell� membranes� has� been� detected� in� vitro� by�stressing� the� isolated� red� blood� cells� with� t�butyl� hydroperoxide,� t�ButOOH�(Avellini� et� al.� 1995).� Outcome� of� this� trial� was� increased�cholesterol:phospholipid�ratio,�suggesting�loss�of�elasticity�in�the�cell�membranes�(Avellini�et�al.�1995).��Protein�oxidation� in� response� to�acute� exercise.�A�standardised� exercise� test�
for� regularly� trained� three�day�eventing�horses�has�been� reported� to� induce�an�increase�in�plasma�oxidised�proteins�after�a�24�hour�recovery�(Portier�et�al.�2006;�de� Moffarts� et� al.� 2007).� Another� paper� has� reported� the� measures� of� blood�oxidant� and� antioxidant� markers� in� healthy� competition� horses� of� different�breeds�(Kirschvink�et�al.�2006).�Show�jumpers�appeared�to�have�higher�oxidised�protein�levels�in�plasma�compared�with�thoroughbred�racehorses,�and�mares�to�have� lower� levels� of� oxidised� proteins� compared� with� geldings� or� stallions�(Kirschvink�et�al.�2006).�In�this�study�trotters�were�not�considered.��Antioxidant� capacity� in� response� to� acute� exercise.� Plasma� antioxidant�
capacity� (PAOC)�and�plasma�antioxidant� reactivity� (TAR)�has�been�reported� to�increase�in�thoroughbred�racehorse�five�minutes�after�1000�m�gallop�at�maximal�velocity� (White� et� al.� 2001).� In� addition,� the� ferric� reducing� ability� of� plasma�(FRAP)�has�been�reported�to�increase�significantly�after�24�hour�recovery�period�

20
in� pentathlon� horses� after� two� show� jumping� courses� (Balogh� et� al.� 2001).� The�total�antioxidant�status�(TAS)�changes�in�opposite�direction�compared�to�FRAP;�however� the� decrease� was� not� statistically� significant� (Balogh� et� al.� 2001).�According� to� this� study,� it� can� be� suggested� when� antioxidant� capacity� is�assessed� using� different� methods,� highly� different� results� may� be� obtained�(Balogh� et� al.� 2001).� This� may� seem� contradictory,� but� the� differences� between�the�two�methods�have�to�be�considered�(Balogh�et�al.�2001).��� The�total�peroxyl�radical�trapping�ability�of�plasma�(TRAP)�is�also�reported�to�increase� in� plasma� after� strenuous� exercise� of� four� consecutive� days� in� trained�standardbred� trotters� (Räsänen� et� al.� 1996).� The� TRAP� increased� as� a�consequence�of�the�six�bouts�of�exercise�with�increasing�intensity�on�two�separate�days� three� days� apart,� peaking� within� 1�2� hours� post�exercise� (Räsänen� et� al.�1996).��
There�were�no�difference�in�plasma�iron�binding�antioxidant�activity�(IBAA%)�due� to�different�environmental� conditions� (Mills� et�al.� 1996).�Although�exercise�resulted� in� decrease� in� IBAA%� post�exercise,� the� decrease� was� statistically�significant�only�5�min�after�the�first�exercise�test�of�moderate�intensity�in�series�of�three�exercise�tests�separated�by�fortnights�(Mills�et�al.�1996).�Glutathione�status�in�response�to�acute�exercise.�Submaximal�exercise�appears�
not� to� induce� any� increase� in� glutathione� (GSH)� concentration� in� erythrocytes�(Mills�et�al.�1996;�Balogh�et�al.�2001;�Kirschvink�et�al.�2002).�However,�significant�increase� is� reported� in� red� blood� cells� after� a� 24�hour� recovery� (Balogh� et� al.�2001).�Endurance�exercise�over�80�km,�has�been�reported�to�induce�severe�decline�in�GSH� (Marlin� et� al.� 2002;�Hargreaves� et� al.� 2002a;� 2002b)�or� total� glutathione�concentration� (TGSH� =� GSH� +� GSSG)� in� red� blood� cells� (Marlin� et� al.� 2002).�Significant�decrease�in�GSSG�levels�has�also�been�reported�following�endurance�exercise�(Marlin�et�al.�2002).�The�concentration�of�oxidised�glutathione�(GSSG)�is�further� decreased� after� a� 16�hour� recovery,� while� TGSH� and� GSH� remained�unchanged� (Marlin� et� al.� 2002).� Although,� the� cumulative� effects� of� exercise�induced� oxidative� stress� are� present� during� three�month� racing� period� of�racehorses� (de�Moffarts� et� al.� 2005)� as�well� as�during� consecutive� standardised�exercise�tests,�SETs�(Mills�et�al.�1996).�According�to�(Mills�et�al.�1996),�there�were�no�detectable�GSH�or�GSSG�in�the�plasma�at�any�stage�of�each�SET.�In�contrast,�acute�exercises�have�also�been�reported�to�increase�GSH�in�plasma�(Chiaradia�et�al.�1998).�This�is�supported�by�the�previous�report�by�(Brady�et�al.�1977),�where�exhaustive�exercise�increased�GSH�levels�in�blood,�but�not�in�erythrocytes�alone.��
Geldings� and� stallions� appear� to� have� higher� TGSH�level� compared� with�mares;�furthermore,� the�GSSG�concentration�appears�to�higher� in�geldings�than�in� stallions� and� mares� (Kirschvink� et� al.� 2006).� Equestrian� discipline� or� riding�style�appeared�to�have�no�effect�on�TGSH�or�GSSG�levels�(Kirschvink�et�al.�2006).�

21
The� glutathione� redox� ratio� (GRR)� is� considered� as� one� of� the� most� sensitive�indices� of� oxidative� stress� following� closely� the� concentration� of� GSSG,� and� is�strongly�affected�by�glutathione�reductase�(GRd)�activity.�It�has�been�previously�reported� that� the� GRd�activities� in� equine� blood� increases� in� relation� with�submaximal� exercise� (Brady� et� al.� 1977;� Brady� et� al.� 1978;� Mills� et� al.� 1996).�Plasma� GRd� activity� has� been� reported� to� remain� unaffected� by� seven�week�interval�type�conditioning�period�on�the�treadmill�(Ji�1990).�There�have�been�no�previous�studies�about�TRx�responses�to�exercise�in�horses�so�far.�Antioxidant� enzymes� in� response� to� acute� exercise.�Glutathione� peroxidase�
(GPx)� has� been� reported� to� decrease� (Ono� et� al.� 1990)� or� remain� unchanged�(Balogh�et�al.�2001)�after�acute,�submaximal�jumping�exercise�in�horses.�Training�periods� of� up� to� three� months� with� gradually� increasing� training� intensity,�appeared�not�to�have�an�effect�on�GPx�activity�in�plasma�(Ji�1995)�or�erythrocytes�(Ji� 1990;� Avellini� et� al.� 1995)� after� submaximal� exercise.� Anyway,� endurance�exercise�over�80�km�has�reported�to�increase�GPx�activity�solely�in�erythrocytes�(Hargreaves� et� al.� 2002a).� The� rate� of� GPx� activity� in� erythrocytes� is� therefore�suggested�to�correlate�positively�with�the�duration�of�exercise�(Hargreaves�et�al.�2002a)� and� with� the� environmental� conditions,� especially� the� temperature�(Hargreaves�et�al.�2002b).�Thoroughbred�racehorses�are�reported�to�have�higher�GPx� activity� compared� with� show�jumpers� or� standardbred� trotters� with� no�effect�of�gender�(Kirschvink�et�al.�2006).�
Extra�cellular�superoxide�dismutase�(SOD)�in�plasma�(McMeniman�and�Hintz�1992)�and�cytosolic�CuZn�SOD�in�red�blood�cells�(Ji�1990;�Ono�et�al.�1990;�Balogh�et�al.�2001)�have�been�reported�to�be�mainly�unchanged�due�to�exercise� .�Mares�have�the�highest�SOD�activities,�followed�by�stallion�and�geldings�and�it�appears�to�decline�with�age�(Kirschvink�et�al.�2006).�Prolonged�effects�of�exercise�are�seen�as�level�of�SOD�activity�decreases�after�three�month�racing�period;�even�though,�the� physical� exertion� was� fairly� extensive� with� daily� training� and� weekly� the�mean�of�4.5�races�per�horse�(de�Moffarts�et�al.�2005).��
Catalase� (CAT)� activity� has� not� been� widely� measured� in� horses� (Ji� 1990).�However,� there� is� a� recent� study� measuring� increased� CAT� activity� from�bronchoalveolar� lavage� fluid� (BALF� or� BAL)� of� thoroughbred� colts� in� training�compared�with�stabled�colts� (Michelotto�et�al.�2010).�BAL� is� considered� to�be�a�valuable�technique�for�evaluating�poor�performance�in�athletic�horses�(Rose�and�Hodgson�1994b).��Non�enzymatic�antioxidants.�The�vitamin�E�status�of�horse�blood�or�skeletal�
muscle�is�not�altered�by�a�single�bout�of�repeated�(Siciliano�et�al.�1996)�or�short�duration� (McMeniman� and� Hintz� 1992)� submaximal� exercise.� Even� prolonged,�endurance�exercise�of�80�160�km,�appears�not� to�affect�plasma�vitamin�E� levels�during� the� exercise� or� after� 60� min� recovery� period� (Marlin� et� al.� 2002;�

22
Hargreaves�et�al.�2002a;�2002b).�However,�the�horses�represented�different�breeds�and�were�not�fed�or�trained�consistently.�Furthermore,�some�of�the�horses�were�withdrawn� from� the� race� due� to� exhaustion� or� insufficient� recovery.� Anyhow,�the� plasma� vitamin� C� levels� have� been� reported� to� decrease� during� recovery�from�prolonged�exercise�(Marlin�et�al.�2002;�Hargreaves�et�al.�2002a;�2002b).�Stress� protein� response� to� acute� exercise.� There� is� only� minor� information�
available� concerning� stress� protein� induction� in� relation� to� exercise� in� horses.�This� is� interesting,� because� HSPs� are� known� to� be� evolutionarily� conserved�proteins� synthesised� in� response� to� physical,� chemical� or� biological� stress.�However,�in�situ�hybridisation�has�been�used�to�establish�the�presence�of�HSP72�mRNA�in�skeletal�muscle� (Pösö�et�al.�2002).� It�appears� to� increase� immediately�after� single� bout� of� exercise� and� descend� back� to� basal� level� after� 24�hour�recovery,�peak�lactate�level�correlating�positively�with�HSP72�mRNA�level�(Pösö�et�al.�2002).�In�addition,�regular�training�further�enhances�the�levels�the�level�of�HSP72�mRNA�(Pösö�et�al.�2002).���2.5.2�The�effects�of�antioxidant�supplementations�in�sport�horses�Antioxidant�supplementation�within�equestrian�sports�is�at�least�as�popular�as�it�is� in� human� sports.� There� are� several� myths� concerning� antioxidants� and�performance� in� horses,� since� the� knowledge� concerning� equine� antioxidant�supplementation�is�scanty.�� Previous�studies�have�shown�that�selenium�(Se)�supplementation�has�no�effect�on�plasma�levels�of�GPx�after�56�days�in�pregnant�mares�or�110�days�in�young,�non�exercised� horses� (Richardson� et� al.� 2006;� Karren� et� al.� 2010),� respectively.�There�were�no�changes�in�GPx�activity�in�RBC�or�muscle�either;�even�though�the�plasma�concentration�of�Se�was�significantly� increased� (Richardson�et�al.�2006).�The�geographical�sources�for�forages�were�not�available�in�either�of�the�studies.�
Antioxidant� supplementation� will� eventually� increase� the� levels� of� given�antioxidants�in�plasma�(McMeniman�and�Hintz�1992;�Siciliano�et�al.�1996;�White�et� al.� 2001;� de� Moffarts� et� al.� 2005).� However,� this� does� not� necessarily� lead� to�increased�vitamin�E�levels�in�muscle�(McMeniman�and�Hintz�1992;�Siciliano�et�al.�1996)�unless�the�supplement�concentration�is�high�enough�(Siciliano�et�al.�1996).�It� has� been� suggested� earlier� that� 150� to� 250� IU� of� vitamin� E/kg� dry� matter� is�needed� to� keep� serum� vitamin� E� concentration� from� declining� in� horses� on�regular�exercise�(Saastamoinen�and�Juusela�1993).��
Furthermore,� plasma� vitamin� E� concentrations� have� been� reported� to� be�higher� in� vitamin� E+C� supplemented� groups� compared� to� vitamin� E�supplementation� itself� (Williams� et� al.� 2004).� This� difference� progressively�diminished�during�an�endurance�race�as�ascorbic�acid�increased�in�the�vitamin�E�group�but�remained�unchanged�in�E+C�group�(Williams�et�al.�2004).�It�has�been�

23
established� that� vitamin� C� regenerates� vitamin� E� by� reducing� tocopheroxyl�radicals�that�are�generated�while�vitamin�E�scavenges�free�radicals�(Packer�et�al.�1979).��
The� increased� vitamin� E� concentration� may� also� be� due� to� an� increased�mobilisation�of�intracellular�ascorbic�acid�stores�in�vitamin�E�group,�whereas�the�E+C�group�was�able�to�maintain�ascorbic�acid�levels�using�the�exogenous�source�for�its�antioxidant�capacity�(Williams�et�al.�2004).�Furthermore,�plasma�vitamin�C�concentration� has� previously� been� reported� to� increase� after� vitamin� E�supplementation�in�exercised�horses�(McMeniman�and�Hintz�1992).��
Selenium�supplementation�together�with�vitamin�E�was�reported�to�reduce�the�severity�of�tying�up,�a�milder�form�of�exertional�rhabdomyolysis,�compared�with�healthy�horses�(El�Deeb�and�El�Bahr�2011).�At�the�same�time�with�relieve�of�the�symptoms,� there� was� a� significant� decrease� in� several� enzymatic� and� non�enzymatic� antioxidants.� This� suggests� an� association� of� antioxidant� protection�with�oxidative�stress�related�diseases.��Effects� on� lipid� peroxidation.�The� effects� of� antioxidant� supplementation� on�
rate� of� lipid� peroxidation� are� quite� controversial� in� horses.� An� exercise�related�decrease� in� blood� lipid� peroxides� during� 15� min� recovery� remains� non�significant� after� supplementation� with� (��3)�vitamin� compared� with� placebo�administrated� horses� (de� Moffarts� et� al.� 2007).� Daily� training� and� dietary�supplementation� with� vitamin� E� and� selenium� has� been� reported� to� decrease�exercise�induced�production�of�MDA� in�plasma� (McMeniman�and�Hintz� 1992).�However,� this� had� no� effect� on� plasma� or� muscle� TBARS� after� exhaustive�exercise� (McMeniman� and� Hintz� 1992).� Furthermore,� dietary� supplementation�with�vitamin�E�and�selenium�during�three�month�training�period�did�not�prevent�the� increase� in�MDA�concentration�after�submaximal�exercise�test�and�recovery�period�of�18�hours�(Chiaradia�et�al.�1998).�Still,�plasma�LPO�has�been�reported�to�increase� after� 80�km� ride� in� moderate� climate� and� remained� high� after� 30�min�recovery�in�vitamin�supplemented�horses�(Williams�et�al.�2004).��
Intramuscular� administration� of� vitamin� E� together� with� selenium� had� no�effect�on�LPO�concentration�in�erythrocytes�after�submaximal�exercise�(Ono�et�al.�1990).� Intravenous� vitamin� C� administration� blunted� the� exercise�induced�increase�in�plasma�TBARS�after�stimulated�gallop�race�(White�et�al.�2001).��
Dietary� supplementation� with� vitamin� E� had� no� effect� on� the� level� of�conjugated�dienes�in�skeletal�muscle�(Siciliano�et�al.�1996).��Effects�on�protein�oxidation.�Dietary�antioxidant�supplementation�with�(��3)�
vitamin� appeared� to� have� no� effect� on� the� plasma� concentration� of� oxidised�proteins�(Portier�et�al.�2006;�de�Moffarts�et�al.�2007).�These�are�practically�the�only�available� study� of� protein� oxidation� in� exercising� horses,� excluding� studies�presented�in�this�thesis.�

24
Effects�on�antioxidant� capacity.�The�dietary� supplementation�with� commercial�antioxidant� supplement� (containing� ascorbic� acid,� ��tocopherol� acetate,� ��carotene,� selenium� and� variety� of� trace� elements)� increased� the� antioxidant�capacity�in�horse�plasma�(de�Moffarts�et�al.�2005).��� Intravenous�administration�of�ascorbate�has�been�reported�to�increase�plasma�antioxidant� capacity� at� rest� (White� et� al.� 2001).� However,� after� five�minute�recovery� from�1000�m�race�at�maximum�speed,� there�was�no�change� in�plasma�antioxidant�capacity�in�vitamin�C�treated�horses�compared�with�the�non�treated�horses� with� lower� pre�� race� level� of� antioxidant� capacity� (White� et� al.� 2001).�Furthermore,�in�untreated�horses�the�increase�was�significant�(White�et�al.�2001).�Effects�on�glutathione�status.�Total�glutathione�concentration�in�erythrocytes�
has�been�reported�to�peak�after�a�21�km�ride�and�decline�until�end�of�an�80�km�race� and� after� 30� min� recovery,� without� an� effect� of� vitamin� supplementation�(Williams�et�al.�2004).�Furthermore,� in�white�blood�cells� the�peak�concentration�was�reached�at�the�end�of�race�(80�km)�and�it�remained�high�after�recovery�of�30�min�(Williams�et�al.�2004).��
The�whole�blood�concentrations�of�GSH,�GSSG�and�GRR(%)�were�not�affected�by� three�month� antioxidant� supplementation� during� training� period� compared�with�placeo�treated�controls�(de�Moffarts�et�al.�2005).�However,�the�rate�of�time�effect�was�stronger�in�supplemented�horses�(de�Moffarts�et�al.�2005).�� No�supplement�related�differences�was�detected�for�blood�GSH,�GSSG�or�GRR�following� (��3)�vitamin� supplementation� at� rest� (de� Moffarts� et� al.� 2007).�However,�an�exercise�related�decrease�in�GSH�during�15�min�recovery�remained�non�significant� after� supplementation� compared� with� placebo�treated� controls�(de�Moffarts�et�al.�2007).�There�were�no�treatment��or�exercise�related�differences�detected�in�GSSG�or�GRR�(de�Moffarts�et�al.�2007).�Effects� on� activities� of� antioxidant� enzymes.�Whole� blood� GPx� activity� is�
reported� not� to� alter� after� supplementation� with� vitamin� E� (McMeniman� and�Hintz� 1992)� or� combination� of� several� antioxidants� (de� Moffarts� et� al.� 2005);�where,� however,� the� GPx� activity� was� reported� to� decrease� in� placebo� control�group�(de�Moffarts�et�al.�2005).�This�blunting�effect� is� further�supported�by�the�earlier�study�by�Ono�et�al.�(1990).�It�has�also�been�reported�that�the�GPx�activity�in�erythrocytes�of�supplemented�horses,�decreases�during�the�first�21�km�of�80�km� ride� and� increases� towards� the� finish�line� at� 80� km,� peaking� after� 30� min�recovery� (Williams� et� al.� 2004).� Daily� training� combined� with� dietary�supplementation� with� solely� vitamin� E� (Ji� 1990)� or� vitamin� E� together� with� Se�(Avellini� et� al.� 1999)� augmented� the� GPx� activity� in� erythrocytes� and� in�lymphocytes,�respectively.�In�horses�receiving�vitamin�supplementation,�the�GPx�activity�in�white�blood�cells�peaked�after�30�min�recovery�(Williams�et�al.�2004).�

25
Dietary� supplementation� with� commercial� antioxidant� supplement� and/or�vitamin�E�has�been�reported�not�to�alter�the�antioxidant�enzyme�response�to�an�acute�exercise�bout�measured�as�SOD�(Ji�1990;�de�Moffarts�et�al.�2005;�de�Moffarts�et�al.�2007)�or�CAT�(Ji�1990).��Effects�on�cell�membranes.�Supplementation�with�certain�antioxidants�appears�
not�to�have�straight�effect�on�erythrocyte�membrane�fluidity�(Portier�et�al.�2006;�de�Moffarts�et�al.�2007).�However,�erythrocytes’�resistance�to�in�vitro�peroxidative�stress� induction� has� been� reported� to� strengthen� by� daily� training� and� dietary�supplementation�with�vitamin�E�and�selenium�(Avellini�et�al.�1999).��
Vitamin� E� supplementation� has� no� significant� effect� on� exercise�induced�increase� in� plasma� levels� of� creatine� kinase,� CK� (McMeniman� and� Hintz� 1992;�Siciliano� et� al.� 1996)� or� aspartate� aminotransferase,� AST� (Siciliano� et� al.� 1996).�However,� intravenous� supplementation�with�vitamin� C� tended� to�decrease� the�CK� level� at� rest� but� amplified� its� post�exercise� increase� compared� with� non�supplemented� horses� (White� et� al.� 2001).� In� vitamin� supplemented� horses� the�plasma�CK�and�AST�are� reported� to� increase�with�distance,�both�peaking�at�80�km�(Williams�et�al.�2004).��2.5.3� L �supplementation�and�horse�There�are�only�a�few�studies�available� in�the�literature�on�the�effects�of���lipoic�acid�(LA)�in�horse�and�observed�effects�of�LA�supplementation�are�summarised�in�table�3.���Table 3 The effects of LA on different biochemical parameters in horses. Type of exercise
Horses Dose of LA
Specimens Parametres Reference
No�exercise�
n�=�12�(5)�6�17�yrs�
xx�
10�mg/kg�2�weeks�
RBC�WBC�
Plasma�
TGSH�,�GPx��GPx�,�TGSH�,�
GPx�,�LPO��
Williams�et�al.�2002�
� � � � � �Endurance�(simulated)�
n�=�12�(4)�Arabian�
10�mg/kg�2�weeks�
Blood�RBC�WBC�
Plasma�
TGSH�,�GRR��GPx��GPx��
CK�,�AST�,�lactate��
LPO�,�vit�E�,�vit�C��
Williams�et�al.�2004�
xx = thoroughbred, RBC = red blood cells, WBC = white blood cells, TGSH = total glutathione concentration, GPx = glutathione peroxidise, LPO = lipid hydro peroxides, GRR = glutathione redox ratio, CK = creatine kinase, AST = aspartate aminotransferase, vit E = vitamin E, vit C = vitamin C
ipoic acid

26
��Lipoic� acid� supplemented� horses� are� reported� to� increase� their� antioxidant�status� and� to� decrease� plasma� muscle� enzyme� concentrations� and� white� blood�cell� apoptosis� during� and� after� exercise� similarly� to� vitamin� E� supplemented�horses�(Williams�et�al.�2004).�
Lipoic� acid� supplementation� increased� the� levels� of� ascorbate� and� ��tocopherol� in�plasma�and� the�GSH�concentrations� in�whole�blood�compared� to�control.� Vitamin� E� and� lipoic� acid� supplemented� groups� had� about� 40%� more�GSH,� 30%� more���tocopherol� and� 15%� more� ascorbate� than� the� control� group�(Williams�et�al.�2004).�This�is�suggested�to�represent�the�recycling�and�scavenging�of�antioxidant�radicals�using�the�exogenous�sources�of� the�vitamin�E�and� lipoic�acid�(Williams�et�al.�2004).��

27
3�Aims�and�objectives��The� general� aim� for� this� thesis� was� to� clarify� the� susceptibility� of� the� horse� to�exercise�induced� oxygen� insults� and� to� define� the� capacity� of� the� thiol�antioxidant�system�and�tissue�protection�in�the�muscle�of�the�horse.�The�studies�were� designed� to� gain� better� understanding� of� the� occurrence� of� exercise�induced�oxidative�stress�and�its�management�in�standardbred�trotters.�They�will�provide�valuable�information�on�tissue�protection�mechanisms�in�animals�with�a�remarkably� high� maximal� oxygen� uptake,� VO2max,� and� may� help� to� reduce� the�deleterious�effects�of�heavy�physical�exercise.�To�a�certain�extent,�these�outcomes�may�be�applicable�to�humans�as�well.�
�The�more�specific�objectives�of�these�series�of�studies�were:��I� To� test� the� effects� of� acute� exercise� on� stress� protein� expression� in� horse�
muscle� and� to� clarify� their� role� and� expression� patterns� in� relation� with�oxidative�insult��
�II� To� examine� the� association� of� the� plasma� antioxidant� capacity� with�
different� antioxidant� parameters� and� certain� oxidative� stress� markers�following� acute� exercise� and� to� elucidate� the� mechanisms� of� antioxidant�protection�in�trained�standardbred�trotters�
�III� To�clarify�the�effects�of���lipoic�acid,�LA,�on�exercise�induced�stress�protein�
induction�and�on�the�oxidative�capacity�of�skeletal�muscle��IV� To�clarify�the�antioxidant�effects�of�LA,�different�indices�of�oxidative�stress�
and�on�tissue�protection�mechanisms�in�trained�standardbred�trotters�

28
4�Methods��The� Ethics� committee� for� Animal� experiments� of� the� Agricultural� Research�Centre� of� Ypäjä� and� The� Ethics� Committee� for� Animal� Experiments� of� the�University� of� Kuopio� approved� all� studies.� The� design� and� interventions� are�summarised�in�table�4.��
Table 4 The design, size, intervention and length and objectives of the studies. Papers Design Size Intervention Objectives
I�II� Controlled�
trial�with�modified�SET�
8�horses�(3�mares,�5�stallions)�
None.��
To�explore�the�mechanisms�of�antioxidant�protection�in�horses�in�response�to�acute�exercise��
III�IV� Case�control�study�with�strenuous�interval�–type�exercise�
6�horses�(2�mares,�4�geldings)�
5�week�LA�supplementation�(25�mg/kg/d).��
Management�of�exercise�induced�oxidative�stress�
SET�=�standardised�exercise�test,�LA�=���lipoic�acid�
4.1 SUBJECTS �Original� papers� I� and� II.� The� study� was� carried� out� in� Agricultural� Research�Centre�in�Ypäjä,�Finland�Eight�clinically�normal�standardbred�trotters,�6�9�years�of�age�and�435�517�kg�in�weight,�were�tested�on�treadmill�for�total�time�of�53�min.�Three� of� the� horses� were� mares� and� five� stallions,� all� of� which� had� been� in�regular�training�for�years.��
Original� papers� III� and� IV.� The� study� was� carried� out� in� the� Ylä�Savo�Vocational� Institute� in� Kiuruvesi,� Finland.� Six� clinically� healthy� standardbred�trotters,�5�13�years�of�age,�and�408�500�kg�in�weight,�were�examined�in�the�study.�Two� of� the� horses� were� mares� and� four� geldings.� All� the� horses� had� been� in�regular�training�for�several�years.��4.2 DIETARY ASSESSMENTS �All�the�horses�in�these�four�papers�were�standardbred�trotters�and�housed�in�box�stalls�and�fed�haylage�(free�access)�and�oats�(2.2�±�0.24�kg)�sufficient�to�meet�the�recommended� nutrient� requirements� (Meyer� 1996)� and� to� maintain� ideal� body�

29
condition� score� (Henneke� et� al.� 1983).� The� horses� were� routinely� checked� by�experienced�veterinarian�prior�to�exercise�tests�in�all�studies.�
In� original� papers� III� and� IV� the� administration� of� ancillary� forages� and�additional�vitamins�was�discontinued�for�five�weeks�before�starting�the�series�of�tests� to� rule� out� a� previous� antioxidant� supplementation� effect.� Horses� were�monitored�regularly�by�experienced�veterinarian�during�the�trials.���4.2.1� �supplementation�The� supplementation�with���lipoic� acid� (LA)�was� conducted�only� in�papers� III�and� IV.� The� main� purpose� of� the� above� mentioned� five�week� period� without�additional� vitamins� (denominated� later� as� “control”)� was� to� reveal� the� true�antioxidant�effects�of�LA�supplementation.�After�this�five�week�period�the�horses�were� supplemented� with� LA� 25� mg/kg� body� weight/day� (from� Changshu�Fushilai� Medicine� &� Chemical� Co.,� LTD,� Changsu,� Jiangsu,� China)� for� five�consecutive� weeks.� The� purity� of� LA� was� tested� and� compared� with� a� reagent�grade� LA� by� using� high�performance� liquid� chromatography,� HPLC,� methods�(Sen�et�al.�1999a).�LA�was�weighted,�mixed�with�molasses�and�fed�to�horses�prior�to�concentrates.��4.3 EXERCISE PROCEDURES �The�physical�fitness�is�routinely�evaluated�with�a�standard�exercise�test�(SET)�in�horses� too� (Persson� 1983).� The� SET� for� horses� consists� of� a� 10�min� warm�up�period�at�1.7�m/s,�followed�by�4�exercise�intervals,�2�min�each,�at�speeds�of�6,�7,�8,�9�and�if�the�horse�is�fit�enough,�at�10�m/s�on�a�high�speed�treadmill�with�a�2.5�3o�incline.�Blood�samples�for�lactate�analysis�are�collected�before�the�test�and�during�the�last�10�s�of�each�interval.�After�the�trotting�stages,�the�horses�are�walked�on�treadmill� 20� min� for� active� recovery� and� the� blood� samples� are� collected� after�every�five�min�in�order�to�follow�the�recovery�rate�of�the�horses.��
Anaerobic� threshold� or� the� onset� of� blood� lactate� accumulation� (OBLA)� is�calculated�from�the�velocity�of�the�treadmill�and�blood�lactate�concentration�after�each�interval�during�the�SET�(Persson�1983).�The�speed�of�treadmill�leading�on�to�blood�lactate�concentration�of�4�mmol/l�(VLa4)�can�also�be�used�as�rough�estimate�of�horses’�aerobic�capcity�and�performance.�
Original� papers� I� and� II.� The� exercise� protocol� was� based� on� the� modified�SET�for�horses�(see�above).�The�animals�were�exercised�on�the�treadmill�at�the�3°�uphill�slope�for�53�min.�The�detailed�protocol�is�shown�in�detail�in�original�paper�I.�
Original�papers�III�and�IV.�The�physical�fitness�of�the�horses�was�evaluated�one�week�before�the�performance�test�with�a�SET.�In�the�subsequent�performance�
Lipoic acid

30
test� the� treadmill� speed� was� sustained� under� the� individual� VLa4� to� make� sure�that� lactic� acid� will� not� disrupt� the� functioning� of� the� skeletal� muscles.� The�performance�test�protocol�is�presented�in�detail�in�original�paper�III.��
In� original� papers� III� and� IV,� both� the� SET� and� the� performance� tests� were�rerun�and�all�the�samples�taken�again�after�the�five�week�LA�supplementation.���4.4 SAMPLE COLLECTION �The�blood�samples�in�all�series�of�tests�were�collected�from�jugular�vein�and�all�the� biopsy� specimens� obtained� from� the� middle� gluteal� muscle� under� local�anaesthesia� as� described� previously� by� Lindholm� and� Piehl� (1974b).� Sample�collection� points� are� introduced� and� the� procedure� for� sample� preparation� is�described�in�detail�in�the�original�papers.� 4.5 BIOCHEMICAL METHODS �Muscle�enzymes�and� lactate.�Analyses�of�blood� lactate,�plasma�creatine�kinase�(CK)� and� plasma� aspartate� aminotransferase� (AST)� were� carried� out� in� the�Laboratory� of� Equine� Hospital� (Ypäjä,� Finland).� Lactate� concentrations� (as� L�lactate)�were�measured�from�blood�with�an�enzymatic�lactate�analyser�(YSI�2300�STAT,�Yellow�Springs� Instrument�Co.,�Yellow�Springs,�OH,�USA)�using� lactate�oxidase�as�a�catalyst.�Analyses�of�plasma�CK�and�AST�were�carried�out�by�using�a� clinical� chemistry� analyzer� Kone�Pro� (Konelab,� Thermo� Clinical� Labsystems�Oy,�Finland)�according�to�IFCC�reference�procedures�(Schumann�et�al.�2002a�and�2002b)�respectively).�The�heart�rates�were�measured�using�Horse�Tester�PEH200�(Polar�Electro�Co,�Kempele,�Finland)�in�the�papers�I�and�II�and�Polar�S810�heart�rate�meter�(Polar�Electro�Co,�Kempele�Finland)�in�papers�III�and�IV.�
Stress� protein� induction.� Heat� shock� protein� (HSP)� expression� was�determined�using�Western�Blot�as�described�in�detail�in�original�paper�I.��
Protein� oxidation.� Protein� carbonyls� (PCarb)� were� measured� as� a� marker� of�protein� oxidation.� Oxidised� proteins� were� derived� by� 2�4�dinitrophynylhydrazine� (DNPH)� and� measured� using� Western� Blot� for� muscle�homogenates� and� enzyme�linked� immunosorbent� assay� (ELISA)� for� plasma�samples�(Gordillo�et�al.�2002;�Atalay�et�al.�2004).��
Aerobic�capacity�of�the�muscles.�Citrate�synthase�(CS)�activity�was�measured�as� described� previously� (Shepherd� and� Garland� 1969)� and� muscle� ��glucuronidase�activity�was�measured�by�using�the�method�of�Barret�(1972).�Electron�paramagnetic�resonance�(EPR).�Electron�paramagnetic�resonance�(EPR)�spectroscopy�was�used�to�measure�the�in�vivo�generated�steady�state�free�radical�concentration� of� muscle� samples.� EPR� or� electron� spin� resonance� (ESR)�

31
spectroscopy�is�known�as�a�sensitive�method�to�detect�and�identify�free�radicals�and�other�chemical�species�that�have�one�or�more�unpaired�electrons.��
The�measurements�were�carried�out�in�Chemical�Research�Center,�Hungarian�Academy�of�Science� (Budapest,�Hungary)�with�an�X�Band�computer�controlled�spectrometer� constructed� by� Magnettech� GmbH� (Berlin,� Germany).�Approximately�100�mg�of�tissue�samples�were�frozen�into�a�rod�shaped�form�and�spectra�of�the�samples�were�recorded�at�77�K�using�a�quartz�finger�dewar�filled�up�with�liquid�nitrogen�(Stadler�et�al.�2003).�For�evaluation,�a�method�of�double�integration� of� the� EPR� signals� with� Mn/MnO� as� an� internal� standard�(Rockenbauer�et�al.�2001).��
Thioredoxin�system.� Thioredoxin� (Trx)� activity� was� determined� using�endpoint�assay�(IMCO�Corporation�Ltd�AB),�the�principle�of�the�assay�being�the�rapid�reaction�between�reduced�thioredoxin�and�protein�disulphide.�Thioredoxin�reductase�(TrxRd)�activity�assay�(IMCO�Corporation�Ltd�AB)�was�based�on�using�a�relative�excess�of�thioredoxin.�
Total�antioxidant�capacity.�Oxygen�radical�absorbing�capacity�(ORAC)�assays�were� performed� using� a� multi�well� plate� reader� according� to� the� methods�previously�published�(Prior�et�al.�2005).�The�antioxidant�capacity�of�the�samples�was�measured�by�the�inhibition�of�the�decrease�of�the�fluorescence�of�fluorescein�(FL,� Na� salt,� Riedel�De� Haen� Aldrich� Milwaukee,� WI,� USA).� Trolox� (Aldrich,�Milwaukee,� WI)� was� used� as� a� control� standard.� Final� results� were� calculated�using�the�differences�of�areas�under�the�FL�decay�curves�between�the�blank�and�a�sample�and�quantified�according�to�Trolox�standards�and�expressed�as��mol.��
Lipid� peroxidation.� Lipid� hydroperoxides� (LPO)� in� whole� plasma� were�determined�as�described�by�Arab�and�Steghens�(2004)�based�on�oxidation�of�Fe2+�to� Fe3+� by� lipid� hydroperoxides� under� acidic� conditions,� followed� by�complexation� of� Fe3+� by� xylenol� orange.� Peroxidative� damage� to� cellular� lipid�constituents�was�determined�by�measuring�the�total�malondialdehyde�(MDA)�in�plasma� and� muscle,� and� was� measured� according� to� the� method� of� Gerard�Monnier�et�al.�(1998)�later�described�by�Breusing�et�al.�(2010).�
As� a� marker� of� lipid� peroxidation,� the� levels� of� 4�hydroxynoneal� (4�HNE)�protein�adducts�were�measured�from�middle�gluteal�muscle�using�Western�Blot�techniques�described�in�detail�in�original�paper�I.��
Glutathione�related�enzymes�in�muscle�and�in�erythrocytes.�The�activities�of�erytrocyte� glutathione� peroxidase� (GPx),� glutathione� reductase� (GRd)� and�glutathione�S�transferase�(GST)�were�determined�spectrophotometrically�(Sen�et�al.�1992).��
Total� and� oxidised� glutathione� concentrations� and� glutathione� related�enzymes� in� muscle.� The� muscle� tissues� for� total� glutathione� concentration�(TGSH)� and� oxidised� glutathione� (GSSG)� analysis� were� deproteinised� with�

32
perchlorous� acid� (PCA;� for� only� TGSH� measurement)� in� original� paper� I� and�with� 5%�metaphosphorous� acid� (MPA;� for� TGSH� and� GSSG� measurements)� in�original�paper�III.��
Plasma� vitamin� E� concentration.� Plasma� vitamin� E� concentration� was�determined� by� high�performance� liquid� chromatography� (De� Leenheer� et� al.�1979).�
Total� protein� concentration.�Total�protein� concentration� for�all� samples�was�detected�by�using�a�BCA�protein�assay�kit�(Pierce,�Rockford,�IL,�USA).��4.6 STATISTICAL METHODS �The�data�were�analysed�by�SPSS�(Statistical�Package�for� the�Social�Sciences)� for�Windows�11.0�(in�original�papers�I�and�II)�and�14.0�(in�original�papers�III�and�IV).�Means� and� standard� errors� of� means� (SEM)� were� calculated� in� all� studies� and�multiple�comparisons�were�performed�as�described�below.��
Original�papers� I� and� II.�Data�were�analyzed�using�analysis�of�variance� for�repeated� measures.� Paired� samples� t�test� was� used� to� analyze� the� difference�between�the�means.�Pearson’s�correlation�coefficient�was�used�to�investigate�the�relationship� between� variables.� The� Bonferroni� correction� was� used� to� control�overall�error�rate.�The�level�of�significance�was�set�at�p�<�0.05.�
Original� papers� III� and� IV.� Multivariate� linear� mixed� model� was� used� to�assess� whether� duration� of� exercise� and� use� of� LA� have� an� effect� on� physical�quantities,� as� it� takes� into� account� the� correlation� structure� of� the� data� due� to�repetitions.� Also� the� paired� samples� t�test� was� used� to� assess� whether� the� LA�supplementation�had�an�effect�on�blood�lactate�before�and�after�the�exercise�and�heart� rate� during� the� exercise.� Spearman’s� correlation� coefficient� was� used� to�assess�the�correlation�between�variables.�P�values�less�than�0.05�were�treated�as�statistically�significant.�

33
5�Results��5.1 EFFECTS OF ACUTE EXERCISE ON TISSUE PROTECTION MECHANISMS IN EXERCISING TROTTERS (ORIGINAL PAPER I) �The� exercise� test�was� a� controlled� trial�which�was�modified� from� standardised�exercise�test�(SET).�The�blood�lactate�concentration�started�to�increase�while�the�treadmill� speed� reached� 6� m/s� and� peaked� at� the� highest� speed.� The� lactate�concentration�was�significantly�higher�(p�<�0.05)�still,�after�20�minutes�of�recovery�compared�to�the�pre�exercise�level.�Stress�protein�induction.�The�submaximal�exercise�did�not�induce�constitutive�
heath� shock� protein� 70� (HSC70),� heat� shock� protein� 70� (HSP70),� heat� shock�protein� 90� (HSP90)� or� glucose�regulated� protein� 75� (GRP75)� expression� in� the�middle�gluteal�muscle�of�the�horse�(Table�3�in�original�paper)�measured�after�4�hour�recovery.�Concomitantly�the�DNA�binding�activity�and�protein�expression�of�heat�shock�factor�1�(HSF1)�were�not�activated�either.�Protein�oxidation.�A�single�bout�of�acute�exercise� increased�significantly� the�
amount� of� protein� carbonyls� (PCarb)� in� the� gluteus� medius� muscle� after� four�hour� recovery� (p� <�0.01,�Table�3� in�original�paper).�Furthermore,�plasma�PCarb�increased� immediately� after� exercise� and� was� further� elevated� after� 4�hour�recovery.� After� 24� hours,� the� level� had� started� do� decrease;� even� though� the�concentrations� were� still� significantly� higher� compared� with� the� resting� levels�(Table�4�in�original�paper).��Enzyme� activities.� Muscle� citrate� synthase� (CS)� activity,� marker� of� the�
oxidative� capacity,� showed� a� tendency� to� decrease� after� 4�hour� recovery� (p� =�0.058).�However,�the�activity�of���glucuronidase�was�not�affected�by�the�exercise.�There� were� no� changes� in� muscle� glutathione�S�transferase� (GST)� or�glutathioneperoxidase�(GPx)�activities�during�recovery.��
Plasma� aspartate� aminotransferase� (AST)� activity� increased� significantly�immediately� after� and�at� four�hours� after� exercise� (p� <� 0.05)�which� returned� to�pre�exercise�level�at�24�hours�of�recovery.��Histology.�To�support�the�exercise�induced�increase�in�AST,�light�microscopy�
sections�of�middle�gluteal�muscle�showed�a�mild�swelling�and� inflammation�of�muscle�fibres.�Classification�of�muscle�fibres�by�ATPase�showed�that�18�±�2%�of�fibres�were�type�I�and�82�±�2%�were�type�II�fibres.��

34
5.2 EFFECTS OF ACUTE EXERCISE ON BIOMARKERS OF OXIDATIVE STRESS IN EXERCISING TROTTERS (ORIGINAL PAPER II) �The� exercise� test� was� a� controlled� trial,� modified� from� SET.� Blood� lactate�concentration� started� to� increase� while� the� treadmill� speed� reached� 6� m/s� and�peaked�at�the�highest�speed.�The�lactate�concentration�was�significantly�higher�(p�<�0.05)�still,�after�20�minutes�of�recovery�compared�to�the�pre�exercise�level.�Total� antioxidant� capacity.�The� plasma� oxygen� radical� absorbance� capacity�
(ORAC)�increased�immediately�after�the�exercise�compared�to�pre�exercise�level�(p�<�0.001).�ORAC�levels�started�to�decrease�after�the�recovery�period�of�24�hours,�but�remained�significantly�higher�than�prior�to�exercise�(p�<�0.01).�Lipid� peroxidation.� The� concentration� of� lipid� hydroperoxides� (LPO)�
increased�(p�<�0.05)�immediately�after�the�exercise�compared�to�pre�exercise�level.�However,�the�levels�of�4�hydroxynoneal(4�HNE)�modified�proteins�in�the�middle�gluteal�muscle�were�at�the�pre�exercise�level�after�four�hour�recovery.��Blood� glutathione� status.�Total� glutathione� concentration� (TGSH)� remained�
unchanged� during� the� exercise� and� recovery.� There� were� no� changes� in�erythrocyte� GPx,� GST� or� glutathione� reductase� (GRd)� activities� prior� to� or�following�exercise.�Alike,�there�were�no�changes�in�GPx�or�GST�activities�in�the�gluteus�medius�muscle�of�the�horse�following�exercise.��Plasma� vitamin� E.� Plasma� vitamin� E� levels� were� not� affected� by� the� acute�
exercise�(Table�5).�� Table 5 Plasma vitamin E levels (mg/l) after acute exercise.
Pre Post 4 h recovery 24 h recovery
1.2 ± 0.09 1.1 ± 0.21 1.4 ± 0.29 1.3 ± 0.18
Values are means ± SEM, level of significance was set at p < 0.05. �Correlations.�Resting�ORAC�levels�correlated�positively�with�plasma�LPO�levels�after� 4�hour� recovery� (r� =� �0.71,� p� <� 0.05)� as� well� as� with� blood� TGSH�concentration�after�four�and�24�hours�of�recovery�(r�=�0.75,�p�<�0.5�and�r�=�0.83,�p�<�0.01,�respectively).��

35
5.3 EFFECTS OF LA SUPPLEMENTATION ON TISSUE PROTECTION MECHANISMS AFTER STRENUOUS EXERCISE IN TROTTERS (ORIGINAL PAPER III) �The� following�results�were�obtained�after�an�exercise� test�which�was�strenuous�interval� run� on� a� high�speed� treadmill� with� 2.5%� slope.� The� speed� of� the�treadmill�was�individually�defined�for�each�horse�based�on�SET�done�during�the�previous�week.�Stress� protein� induction.� Intense� aerobic� exercise� had� no� overall� significant�
effect�on�HSP�levels�in�the�horses.�However,�in���lipoic�acid�(LA)�supplemented�horses�heat�shock�protein�70�(HSP70)�increased�19%,�and�was�significantly�higher�after� the� 24�hour� recovery� compared�with�non�supplemented�horses� (p� <� 0.05).�LA�supplementation� increased� the�basal� level�of�heat�shock�protein�90� (HSP90)�by�24%�and�the�overall�levels�of�constitutive�heath�shock�protein�70�(HSC70)�by�12%�(p� <�0.05� in�both).�There�were�no�significant�changes� in�muscle�heat� shock�protein�60� (HSP60),�heat�shock�protein�25� (HSP25)�or�glucose�regulated�protein�75�(GRP75)�responses�due�to�LA�supplementation�or�exercise.�Enzyme� activities.� LA�supplementation� increased� citrate� synthase� (CS)�
activity� in�equine�muscle�at�rest� (p�<�0.05).�On�the�other�hand,�only� in�controls,�muscle�CS� activity� increased� in� response� to� exercise� (p� <� 0.05,� Fig� 2� in� original�paper).��
Plasma� creatine� kinase� (CK)� activity� increased� significantly� in� response� to�exercise� both� before� and� after� LA� supplementation� (p� <� 0.001� and� 0.05�respectively).�However,�in�controls�the�CK�level�remained�elevated�still�after�the�24�hour�recovery�compared�with�the�resting�levels�(p�<�0.05).�In�response�to�LA�supplementation,�the�increase�in�plasma�CK�concentration�from�the�baseline�to�6�hours� after� recovery� was� only� 35%,� compared� with� a� 97%� increase� in� non�supplemented� horses.� Although� this� distinction� between� CK� levels� was� not�statistically�significant,�the�difference�due�to�LA�was�significant�(p�<�0.05).�There�were� no� statistically� significant� changes� in� plasma� aspartate� aminotransferase�(AST)�due�to�exercise�or�LA�supplementation.�Blood� lactic� acid� and� heart� rate.� LA� supplementation� reduced� the� post�
exercise�concentration�of�blood�lactate�without�significant�changes�in�heart�rates�(p�<�0.05).�Correlations.�After� 24�hour� recovery�plasma�AST� correlated�negatively�with�
the� resting� levels� of� HSP25� and� HSP60� (r� =� �0.829,�p�<� 0.05� in� both)� before� LA�supplementation.� Similarly,� the� plasma� AST� level� correlated� negatively� with�muscle�HSP25�level�(r�=��0.943,�p�<�0.01)�after�the�recovery�period�of�24�hours�and�with�muscle�HSP90�level�after�48�hours�(r�=��0.886,�p�<�0.05).�

36
After� LA� supplementation� plasma� CK� level� correlated� negatively� with� muscle�HSP25�level�(r�=��1.000,�p�<�0.001)�and�further�with�muscle�HSC70�after�48�hour�recovery�(r�=��0.886,�p�<�0.05).� 5.4 EFFECTS OF LA SUPPLEMENTATION ON MUSCLE FREE RADICAL PRODUCTION, THIOL STATUS AND CERTAIN MARKERS OF OXIDATIVE STRESS AFTER STRENUOUS EXERCISE IN TROTTERS (ORIGINAL PAPER IV) The� exercise� test� was� controlled� trial,� modified� from� SET.� Blood� lactate�concentration� started� to� increase� while� the� treadmill� speed� reached� 6� m/s� and�peaked�at�the�highest�speed.�The�lactate�concentration�was�significantly�higher�(p�<�0.05)�still,�after�20�minutes�of�recovery�compared�to�the�pre�exercise�level.��Free�radical�production.�Based�on�the�electron�paramagnetic�resonance�(EPR)�
assay,� compared� to� controls� LA� supplementation� blunted� the� exercise�induced�increase� in� the� muscle� free� radical� concentration,� whereas� the� amount� of� free�radicals� in� non�supplemented� horses� increased� significantly� immediately� after�exercise�compared�with�resting�levels�(p�<�0.05,�Figure�1�in�original�paper).��Lipid� peroxidation.� The� LA� supplementation� also� decreased� the� exercise�
induced� increase� in� muscle� malondialdehyde� (MDA)� concentrations� (p� <� 0.05).�The� overall� trend� in� muscle� MDA� level� was� consistent� with� those� of� LPO.�However,�the�change�in�the�magnitude�and�the�statistical�significance�was�not�as�evident�in�LPO�as�it�was�in�MDA.��
In� plasma,� five�week� LA� administration� decreased� exercise�induced� LPO�compared� with� control� (p� <� 0.05).� The� LA� supplementation� also� decreased� the�exercise�induced�increases�in�plasma�MDA�concentrations�(p�<�0.05).�The�overall�trend� in� plasma� MDA� levels� was� consistent� with� those� of� LPO.� In� muscle,� the�change� in� the� magnitude� and� the� statistical� significance� was� not� as� evident� in�MDA�as�it�was�in�LPO.��Protein�oxidation.�In�muscle�LA�supplementation�had�no�statistically�relevant�
effect�on�the�exercise�related�protein�carbonyl�(PCarb)�levels.��Total� antioxidant� capacity.� In� non�supplemented� horses,� intense� acute�
exercise� increased� the� plasma� oxygen� radical� absorbance� capacity� (ORAC)�immediately�after�exercise�(p�<�0.01).�ORAC�also�remained�higher�after�recovery�period�of� two�hours� (p� <� 0.05).�The�LA�supplementation�attenuated� the�overall�exercise�induced�ORAC�response�(p�<�0.05).��Thiol� redox� status.� There� was� no� significant� change� in� muscle� thioredoxin�
(Trx)� activity� following� exercise� or� LA� supplementation.� At� the� same� time,�thioredoxin� reductase� (TrxRd)� activity� during� recovery� was� significantly�increased�following�LA�supplementation�(p�<�0.05).��

37
The� LA� supplementation� increased� the� overall� amount� of� muscle� TGSH� (p� <�0.05).� After� 6�hour� recovery,� the� increase� was� significantly� higher� in� LA�supplemented� horses� than� in� non�supplemented� horses� (p� <� 0.05).� Although,�there� were� no� significant� changes� in� muscle� oxidise� glutathione� (GSSG)� or�glutathione�redox�ratio�(GRR%�=�GSSG/TGSH�x�100)�in�non�supplemented�or�LA�supplemented�horses.�
Muscle� GRd� activity� was� significantly� higher� in� LA� supplemented� horses�compared�with�non�supplemented�animals�after�24�hour�recovery.�Furthermore,�the�muscle�GPx�activity�tended�to�be�higher�after�LA�supplementation�compared�with� non�supplemented� horses� after� 6�hour� recovery� (p� =� 0.08).� Despite� the�statistical�non�significance,�LA�induced�increase�in�muscle�GPx�activity�was�33%�at� rest� and� varied� from� 43%� to� 81%� depending� on� the� time� of� recovery;� the�average�increase�rate�following�LA�supplementation�was�57%.�Correlations.� In� paper� IV,� LA�induced� increase� in� pre�exercise� TrxRd�level�
appeared�to�decrease�post�exercise�EPR�as�well�as�muscle�PCarb,�LPO�and�MDA�levels� after� 24�hour� recovery� (r� =� �0.900,� p� <� 0.05� in� all).� The� rest� of� the�correlations�are�summarised�in�table�2�of�the�original�paper.�

38
6�Discussion��Exercise,� free� radicals� and� oxidative� stress� form� a� complex� triangle� with�important� influences� on� health� and� disorder.� Increased� generation� of� reactive�oxygen� species� (ROS)� is� linked� with� aging� and� in� the� pathogenesis� of� several�diseases.�In�the�studies�of�this�thesis,�we�detected�the�free�radical�and�non�radical�mechanisms� of� exercise�induced� oxidative� stress� and� formed� an� extensive�overview�of�oxidant�related�insults�and�tissue�protection�mechanisms�in�models�with�remarkably�high�oxidative�capacity.�Previous�reports�have�studied�the�acute�effects�of� exercise�on�horse,�and� therefore�we�aimed� to�pay�special�attention� to�recovery.�According�to�our�hypothesis,�exercise�induced�ROS�generation�and�the�length�of�recovery�period�may�play�a�significant�role�in�connection�with�exercise�induced�oxidative�damage.�
Athletic�capacity�and�potential� to�respond�to� the�repeated�stress� imposed�by�strenuous,� long�term� training� makes� horse� an� excellent� animal� for� exercise�studies.�As�far�as�the�biochemistry�is�concerned,�the�skeletal�muscle�of�the�horse�adapts�to�endurance�exercise�training�similarly�to�the�human�muscle�(Tyler�et�al.�1996;�1998).�Furthermore,�it�has�been�proved�that�the�changes�in�muscle�fibers�of�adolescent�thoroughbreds�are�caused�by�training�and�not�by�growth�(Yamano�et�al.� 2002).�The�physiological� similarities�of�mammals�and� the�uniform�muscular�structure�make� it�possible� to�apply� the� findings� from�human�studies� to�horses.�However,�there�are�certain�differences�that�have�to�be�taken�into�account.�These�discrepancies�are�discussed�in�detail�in�chapter�2.5.�
The� findings� of� the� present� thesis� show� that� the� induction� of� antioxidant�defences� following� acute� physical� stress� can� be� interpreted� as� a� protective�response� and� the� extent� of� induction� may� reflect� the� intensity� or� level� of�oxidative�stress�(Sen�and�Packer�2000;�Atalay�et�al.�2006)�also�in�horses.��
It� is� of� great� importance� for� the� field� of� exercise� science� to� gain� more�information� of� the� tissue� protection� mechanisms,� their� induction,� recovery�pattern� and� management.� Furthermore,� there� is� an� ever�increasing� debate�concerning�the�welfare�of�domestic�animals�used�for�sports.�Studies�like�this�give�important�information�for�the�horse�industry�on�how�to�take�better�care�of�horses�and�how�to�improve�their�well�being.���6.1 FREE RADICAL PRODUCTION DURING EXERCISE �Our�study�(original�paper� IV)�on�electron�paramagnetic� resonance� (EPR)�signal�induced�by�interval�type�exercise�appears�to�be�the�first�ever�published�in�horses.�

39
There� are,� to� my� knowledge,� no� previous� studies� measuring� the� rate� of� free�radical� production� in� equine� muscle,� even� though� exercise� is� a� well� known�source� for� reactive�oxygen� species� (ROS)� and� free� radicals� leading� to� oxidative�stress.��
At� the� biochemical� level� it� is� clear� that� ROS� modify� muscle� components�during�strenuous�exercise.�Cellular�structures�that�are�located�nearest�the�sites�of�ROS�production�expose�to�the�highest�local�concentrations�and�are�most�likely�to�be�affected� (Ferreira�and�Reid�2008).�The�observed�exercise�induced� increase�of�free� radical� production� in� horse� skeletal� muscle� is� in� line� with� the� previous�reports�of�diverse�tissues�of�humans�(Davies�et�al.�1982;�Ashton�et�al.�1998;�Bailey�et�al.�2007)�and�in�other�mammals�(Tanabe�et�al.�2006;�Traverse�et�al.�2006).�The�five�week���lipoic�acid� (LA)� supplementation�decreased� free� radical� formation.�The�perceived�decrease�in�the�amount�of�free�radicals�is�supported�by�the�earlier�reports� on� both� LA� and� its� reduced� form� dehydrolipoicacid� (DHLA)� directly�scavenging�ROS�(Petersen�Shay�et�al.�2008).�
It�is�already�known�that�for�maximal�performance�an�optimal�level�of�ROS�is�needed� (Reid� et� al.� 1992b).� Yet,� our� results� suggest� that� a� decrease� in� exercise�induced� ROS� production� following� LA� supplementation� does� not� cause� any�decrease� in� the� performance.� The� overdose� of� nutritional� antioxidants� may,�however,� decrease� the� maximal� performance� by� attenuating� the� essential� ROS�production.� LA� supplementation� may� reduce� the� indices� of� exercise�induced�oxidative� stress� directly� by� augmenting� intracellular� protective� mechanisms�against�oxidative�insult�and�indirectly�by�decreasing�the�ROS�production�during�exercise.��6.2 THIOL REDOX CONTROL �Since�all�biological�systems�contain�redox�elements�(e.g.�redox�sensitive�cysteine�residues),� whose� organisation� and� activity� occurs� through� thiol� redox� circuits�(e.g.� thioredoxin,� TRx� and� glutathione,� GSH),� thiol� oxidation� is� considered� the�most� sensitive� marker� of� oxidative� stress� (Jones� 2008).� These� small� molecular�weight� thiols� are� evolutionarily� conserved� and� have� versatile� functions� within�cells�and�serve�to�maintain�an�overall�reducing�environment�within�cells�(Grune�et� al.� 1997;� Demasi� et� al.� 2001;� Jones� 2008).� In� the� oxidant�thiol� interaction� the�oxidant� is�neutralised�to�a�relatively� less� toxic�by�product�at� the�expense�of� the�reducing�power�of�thiols.��
In�our�study� (original�paper� IV),� there�was�a� strong�association�between�Trx�and� glutathione� throughout� the� strenuous� exercise� protocol,� even� though�exercise� itself� had� no� significant� effect� on� either� of� the� thiols� separately.� In�

40
contrast,�five�week�lipoic�acid�(LA)�supplementation�up�regulated�muscles�total�glutathione�(TGSH)�levels.���6.2.1�Glutathione�redox�status�and�related�enzymes�The�glutathione�redox�ratio�(GSSG/TGSH)�is�considered�to�be�a�reliable�indicator�of� tissue� redox� status� (Schafer� and� Buettner� 2001;� Ferreira� and� Reid� 2008)� and�oxidative� stress� (Jones� 2006;� Jones� and� Liang� 2009;� Adimora� et� al.).� However,�there�were�no�significant�changes�in�oxidised�glutathione�(GSSG)�or�GSSG/TGSH�ratio�in�response�to�exercise�or�during�recovery�in�these�series�of�studies.��
The�exercise�induced�changes�in�redox�status�have�not�always�been�consistent�in�humans�(Goldfarb�et�al.�2005a;�Goldfarb�et�al.�2005b;�Goldfarb�et�al.�2008)�or�in�horses�(Balogh�et�al.�2001;�Marlin�et�al.�2002;�Hargreaves�et�al.�2002a;�2002b;�de�Moffarts� et� al.� 2004;� de� Moffarts� et� al.� 2005;� 2006;� Williams� and� Carlucci� 2006;�Marañón�et� al.� 2008).�Neverthless,� the� findings�of� the�present� thesis� are� in� line�with� the� earlier� studies� with� erythrocytes� in� pentathlon� horses� (Balogh� et� al.�2001)�and�in�human�marathon�runners�(Kaikkonen�et�al.�2002)�with�no�exercise�induced�change�in�glutathione�concentration�immediately�after�exercise.��
The� changes� in� glutathione� redox� status� are� short�lived,� partly� due� to� rapid�conversion� of� GSSG� back� to� GSH� by� glutathione� reductase� (GRd),� which� is� an�important� determinant� of� GRR� and� cellular� protection� against� oxidative� stress�(Hayes� and� McLellan� 1999;� Goldfarb� et� al.� 2005b;� Forman� et� al.� 2009;� Forman�2010).� LA� supplementation� increased� GRd� activity� after� 24�hour� recovery,�suggesting�increased�protection�against�exercise�induced�oxidative�damage.�The�increase� in�TGSH�concentration� in�muscle�during�recovery�can�be�attributed� to�the� enhanced� regeneration� of� GSH� through� increased� GRd� activity� and� to� the�GSH�sparing�effect�of�LA�(Packer�et�al.�1995;�Khanna�et�al.�1999a).��
Along�with�its�direct�antioxidant�properties,�glutathione�serves�as�substrate�for�glutathione� peroxidase� (GPx)� in� the� enzymatic� breakdown� of� peroxides,� e.g.�hydrogen� peroxide� and� lipid� hydroperoxides� and� for� other� enzymes� like�glutathione� reductase� (GRd)� and� glutathione�S�transferase� (GST).� Parallel� to�unchanged�TGSH�levels,�there�were�no�changes�in�GPx,�GRd�or�GST�activities�in�red�blood�cells�and�or�in�gluteus�medius�muscle�post�exercise�or�during�recovery�in� either� of� the� studies.� However,� there� was� significantly� increased� overall�activity� of� muscle� GPx� after� LA� supplementation,� which� is� consistent� with� the�previous�studies�(Kumaran�et�al.�2004;�Chae�et�al.�2008).�The�LA�related�increase�in�the�activity�of�GPx�is�supported�by�earlier�reports�on�horse�white�blood�cells�and� on� equine� erythrocytes� following� supplementation� with� an� antioxidant�cocktail� containing� daily� quantities� of� 11500� mg� ascorbic� acid,� 7000� mg� ��tocopherol�acetate�and�500�mg���carotene�(Williams�et�al.�2002).�

41
The� increase� in�GPx�activity� following�LA�supplementation�may�partly�explain�the� slight� increase� of� GSSG� following� supplementation� as� GSH� is� used� up�substrate� for� GPx.� The� increased� GPx� activity� may� therefore� lead� to� decreased�exercise�induced� lipid� peroxidation� (LPO)� since� LPO� can� be� inhibited�enzymatically� or� non�enzymatically.� Moreover,LPO� products� are� better�substrates�for�GPx�than�for�GSH�itself�(Niki�and�Yoshida�2005).�Furthermore,�LA�has� free� radical� scavenging�properties�and� is�potentially�able� to� inhibit� the� free�radical�mechanisms�of�LPO�.��
The� strong� positive� correlation� between� plasma� aspartate� aminotransferase�(AST)�and�erythrocyte�GPx�activity�immediately�after�exercise�suggests�incipient�muscle�damage,�possibly�caused�by�ROS.�However,�it�should�also�be�considered�that�increased�ROS�may�cause�inactivation�of�these�enzymes�(Atalay�et�al.�2000).�
The� overall� decrease� in� GPx� activity� during� a� long� training� period� has� also�been�previously�reported�in�the�horse�(Avellini�et�al.�1995).�It�has�been�suggested�to�be�of�a�different�origin�(i.e.�selenium�deficiency�or�prolonged�physical�stress)�than� that� in� the� acute� exercise� model� (Balogh� et� al.� 2001).GSTs� are� usually�involved�in�the�metabolism�of�xenobiotics,�but�some�of�them�may�be�important�protectors�against�lipid�peroxidation�(Halliwell�and�Gutteridge�2007).���6.2.2�Thioredoxin�system�Both�thioredoxin�(Trx)�and�thioredoxin�reductase�(TrxRd)�play�an�essential�role�in� cell� function� and� protection� by� limiting� oxidative� stress� directly� via� their�antioxidant� effects,� and� indirectly� by� protein–protein� interactions� with� key�signalling� molecules� (Lillig� and� Holmgren� 2007).� The� five�week� LA�supplementation� increased�the�reductive�potential�of� the�middle�gluteal�muscle�by�increasing�the�recovery�activities�of�TRxRd�and�GRd�within�the�muscle.�Even�so,�supplementation�had�no�effect�on�muscle�TRx�activity�itself.�Protective�role�of�TRx� system� against� oxidative� insults� is� supported� in� the� present� study� by� the�negative� correlations� between� TRxRd� activity� at� rest� and� post�exercise� free�radical�formation.�Furthermore,�in�LA�supplemented�horses�higher�TRx�activity�at�rest�was�associated�with�lower�post�exercise�glutathione�redox�ratio�(GRR%�=�GSSG/TGSH*100%),� supporting� the� hypothesis� of� TRx� protection� against�oxidative�stress.�This�may,�at�least�in�part,�explain�the�TGSH�sparing�effect�of�LA�(see�chapter�6.2).��6.3 LIPID PEROXIDATION �As�a�well�established�mechanism�of�cellular�damage,�lipid�peroxidation�is�widely�accepted�as�an�indicator�of�oxidative�stress.� In�these�series�of�studies,�moderate�(original�paper�II)�as�well�as�more�strenuous�(original�paper�IV)�exercise�appears�

42
to� multiply� the� amount� of� plasma� lipid� peroxidation� products� (LPO)�immediately� post�exercise� in� exercising� horses,� which� is� in� agreement� with�previous� studies� (McMeniman� and� Hintz� 1992;� Avellini� et� al.� 1995;� Mills� et� al.�1996;� Chiaradia� et� al.� 1998;� White� et� al.� 2001;� Marlin� et� al.� 2002).� After� more�moderate� exercise,� the� level�of�LPO� in�plasma� tended� to� remain�higher�at� 24�h�recovery�compared�with�the�resting�levels�(original�paper�II).��
Lipid� hydroperoxides� are� not� stable� end� products� of� lipid� peroxidation� and�serve� as� substrates� for� several� enzymes,� such� as� glutathione� peroxidase� (see�chapter� 6.2.1.)� and� undergo� secondary� reactions� (Niki� and� Yoshida� 2005).�Therefore� the� level� of� lipid� hydroperoxides� does� not� necessarily� represent� the�extent� of� lipid� peroxidation� in� vivo� (Niki� and� Yoshida� 2005).� The� primary�products� of�hydroperoxides�decompose�by�various�mechanisms� to�more� stable�secondary�products,�like�aldehydes,�with�potential�toxicity�(Gueraud�et�al.�2010).�
After�more�strenuous�interval�type�exercise�(original�paper�IV),�we�found�the�level� of� lipid� peroxidation� products� to� return� back� to� pre�exercise� level� faster�after� more� strenuous� exercise� compared� with� less� intense� exercise.� In� the�previous� studies,�plasma�concentrations�of�malondialdehyde� (MDA)�have�been�reported�to�recover�to�pre�exercise�levels�by�24�h�after�prolonged�exercise�(Mills�et�al.�1996)�and�in�red�blood�cells�as�soon�as�after�one�hour�recovery�(Brady�et�al.�1978)� and� after� 24�hour� recovery� (Matsuki� et� al.� 1991).� The� exercise�induced�increase� in� lipid�peroxidation�products� is�also�reported�to� increase�significantly�after� several� hours� (>� 6� h)� or� days� (24�72� h)� following� eccentric� stimulus� in�humans�(Sacheck�et�al.�2003;�Kingsley�et�al.�2006).�However,�in�humans�eccentric�exercise�has� increased�plasma�LPO�up� to� four�days� after� exercise� (Childs� et� al.�2001).� Some� exercise�related� pathologic� conditions� are� also� associated� with�inflammatory� processes.� The� late� increase� in� lipid� peroxidation� markers� after�damaging� exercise� may� be� due� to� inflammation.� Even� though� neutrophils� are�important�in�initiating�tissue�repair,�they�may�cause�further�damage�and�impair�functions� in� skeletal� muscle� by� producing� reactive� species� (Pizza� et� al.� 2005;�Tiidus�and�Enns�2009).�Further�research�is�needed�to�establish�whether�delayed�increase� in� lipid� peroxidation� markers� is� relevant� to� athletic� horses.� Five�week�LA�supplementation�appeared�to�decrease�plasma�LPO�concentration�before�and�after� exercise.� This� may� be� due� to� the� preventive,� scavenging,� quenching� or�removing�actions�of�LA�towards�active�oxidants�(Niki�and�Yoshida�2005).���
Lipid�peroxidation�in�tissues�is�closely�related�to�loss�of�cell�membrane�fluidity�and� exercise�induced� oxidative� stress� negatively� affects� the� fluidity� of�erythrocyte� membranes� by� increasing� lipid� peroxidation� (Cazzola� et� al.� 2003;�Sentürk�et�al.�2005;�Gueraud�et�al.�2010).�We�found�the�concentrations�of�muscle�LPO� and� plasma� MDA� being� strongly� associated� with� the� plasma� activities� of�creatine�kinase�(CK)�and�aspartate�aminotransferase�(AST),�illustrating�the�strong�

43
connection�between� lipid�peroxidation�and�muscle�damage.�This� is� in� line�with�the�earlier�studies�in�horses�(White�et�al.�2001)�as�well�as�in�young�and�older�men�(Sacheck�et�al.�2003).�The�increased�plasma�level�of�muscle�enzymes�is�considered�as�a�marker�of� increased�membrane�permeability�due� to�exercise.�The�extent�of�exercise�induced� damage� in� muscles� is� evaluated� by� measuring� the� levels� of�these�enzymes�from�plasma�or�serum.�The�enzymes�are�present�in�plasma�all�the�time�at�low�levels�but�released�more�extensively�into�blood�flow�as�a�result�of�cell�damage.� The� scale� of� the� damage� is� evaluated� according� to� the� magnitude� of�concentration,�also�in�horses�(Anderson�1976).��
The�observed�tendency�towards�an�increase�in�plasma�CK�levels�following�LA�supplementation� (original� paper� IV)� is� supported� by� the� earlier� study� with�vitamin� E� supplementation� and� eccentric� exercise� in� older� men� (Sacheck� et� al.�2003).�However,�there�are�also�other�findings�inconsistent�with�this�(Williams�et�al.� 2004),� demonstrating� lower� CK� levels� with� LA� supplementation� following�endurance�exercise.�Nonetheless,�the�duration�of�exercise�(Child�et�al.�1998),�age,�gender�and�exercise�training�(Williams�et�al.�2004)�may�also�influence�the�release�of�CK�during�exercise.�As�LA�supplementation�decreased�the�rate�of�CK�increase�during�recovery�after�constant�exercise�duration�(original�paper�IV),�we�suggest�that� LA� might� decrease� acute,� exercise�induced� muscle� damage.� One� probable�explanation� is� the� detected� decrease� in� lipid� peroxidation� resulting� in� retained�fluidity�of�cell�membranes.�The�protective�effects�of�LA�supplementation� in� the�lipid� phase� are� represented� by� decreased� oxidative� lipid� damage� and� further�supported�by�a�previous�study�by�our�group�(Khanna�et�al.�1999a)�and�by�more�recent�findings�in�horses�(Williams�et�al.�2002;�Williams�et�al.�2004),�rats�(Skibska�et�al.�2006)�and�humans�(Zembron�Lacny�et�al.�2007).��
The� increased� unsaturation� index� of� fatty� acids� has� been� reported� in� horses�during� training,� suggesting� potential� training�induced� protection� against�oxidative� stress� (Avellini� et� al.� 1995)� and� is� supported� by� the� several� latter�findings�in�humans�(Andersson�et�al.�1998;�Cazzola�et�al.�2003)�and�rats�(Li�et�al.�1999;�Quiles�et�al.�1999).�Adequate�training�appears�to�improve�performance�by�strengthening� muscles’� ability� to� resist� and� even� to� repair� exercise�induced�injury.���6.4 PROTEIN OXIDATION �Consistent� with� the� increased� LPO� levels,� we� observed� increased� protein�carbonylation� in� plasma� and� middle� gluteal� muscle� within� the� same�experimental� setting.� Our� studies� evidenced� increased� amounts� of� protein�carbonyls�in�plasma�and�muscle�induced�by�a�single�bout�of�acute,�submaximal�exercise�(original�paper�I).�The�concentration�appeared�to�peak�after�a�few�hours�

44
of� recovery� and� started� to� decline� before� the� recovery� point� of� 24� hours.�However,� the� plasma� concentration� of� protein� carbonyls� (PCarb)� was� still�significantly�higher�after�24�hour�recovery�compared�with�resting�levels�(original�paper�I).��
LA� supplementation� appeared� to� decrease� the� extent� of� protein� carbonyl�formation�during�recovery�from�strenuous�exercise�(original�paper�IV).�However,�the�decrease�did�not� reach�statistical� significance.�As� far�as�we�know,� there�are�only�two�papers�apart�from�our�own�(Portier�et�al.�2006;�de�Moffarts�et�al.�2007)�that� have� reported� oxidised� proteins� in� horses.� These� papers� report� a� slight�increase� (Portier� et� al.� 2006)� and� slight� decrease� (de� Moffarts� et� al.� 2007)� in�plasma�oxidised�proteins�with� exercise.�There� are� only� a� few� studies� reporting�decreased� protein� carbonyls� levels� after� LA� supplementation� at� rest� and� after�exercise� in� human� (Saengsirisuwan� et� al.� 2001;� Saengsirisuwan� et� al.� 2002;�Zembron�Lacny�et�al.�2009).�
Protein� carbonyl� content� is� the� most� common� biomarker� of� the� oxidative�damage�of�proteins�(Suzuki�et�al.�2010).�Nonetheless,�the�effects�of�LA�on�protein�oxidation�are�not�as�widely�studied�as�it�is�on�lipid�peroxidation.�From�scientific�literature�one� is� able� to� find� studies� to� support� all� these�opposite�observations.�Despite� the� fact� that� increased� protein� oxidation� is� well� characterised� in� active�cells� and� correlated� with� aging� (Stadtman� 2001),� published� information� on� the�exercise�induced�protein�carbonylation� is�conflicting.�These�discrepancies�could�be�explained�partly�by� the�heterogeneity�and� the�methodological� limitations�of�thestudies,�especially�in�spectrophotometric�studies.��
�6.4.1�Thioredoxin�system�and�protein�carbonylation�Intense,�graded�exercise�induced�a�strong�interaction�between�protein�carbonyls�and� muscle� thioredoxin� (Trx)� system.� This� interaction� is� recently� reported� to�increase�during�the�decarbonylation�phase,�i.e.�the�process�within�cells�in�which�carbonylation� can� be� eliminated� (Wong� et� al.� 2008).� Trx� interactions� with�carbonylated�proteins�have�been�reported�to�increase�during�the�decarbonylation�(Wong�et�al.�2008;�Suzuki�et�al.�2010;�Wong�et�al.�2010).��
Protein� carbonyls� are� derived� from� oxidative� cleavage� of� the� peptide� main�chain� and� the� oxidation� of� the� side� chains� of� certain� amino� acids.� Generally,�oxidation�reactions�will�affect�the�biochemical�properties�and�the�conformation�of�the�protein,�leading�to�different�functional�properties�and�loss�of�enzyme�activity�(Grune�et�al.�2005;�Grimm�et�al.�2010).�Oxidative�modifications�are�also�known�to�induce�cell�proteasomes�(Radak�et�al.�2000;�Grimm�et�al.�2010).��
The�results�from�Wong�et�al.�(2008)�indicate�that�Trx,�which�has�been�shown�to�play�integral�roles�in�redox�signaling�(Kondo�et�al.�2006)�may�also�regulate�signal�transduction� mechanism� involving� protein� carbonylation� and� decarbonylation.�

45
Protein� carbonylation� is� not� chemically� reversible,� neither� the� nature� of�decarbonylation�is�yet�known�(Dalle�Donne�et�al.�2006).��
In�addition,�positive�correlations�between�muscle�protein�carbonyl�levels�and�TRx� and� thioredoxin� reductase� (TRxRd)� activities� after� 24�hour� recovery� were�observed�in�LA�supplemented�horses.�Induction�of�antioxidant�defences�to�acute�stress�can�be�interpreted�as�a�protective�response,�and�the�extent�of�this�induction�may�reflect�the�levels�of�oxidative�stress�(Sen�and�Packer�2000;�Atalay�et�al.�2004;�Atalay�et�al.�2006;�Tanskanen�et�al.�2010).�Other�studies�have�reported�decreases�in� the� levels� of� protein� carbonyls� after� LA� supplementation� in� response� to�exercise�training,�but�these�studies�did�not�examine�acute�responses�to�exercise�induced�oxidative�stress�(Saengsirisuwan�et�al.�2001;�Saengsirisuwan�et�al.�2002).�
�6.5 TOTAL ANTIOXIDANT CAPACITY �The�immediate�increase�in�the�total�antioxidant�capacity�indicator,�oxygen�radical�absorbance� capacity� (ORAC),� after� acute� exercise� is� in� accordance� with� our�previous�study�with�standardbred�trotters�(Kinnunen�et�al.�2005b)�and�with�other�studies� in� horses� where� other� methods� of� total� antioxidant� capacity� including�plasma� antioxidant� capacity� (PAOC)� and� total� antioxidant� reactivity� (TAR)�methods�were�used�(White�et�al.�2001).�However,�LA�supplementation�appeared�to�blunt�the�exercise�induced�increase�in�ORAC.�This�is�supported�by�White�et�al.�(2001),� who� showed� that� intravenous� ascorbate� administration� to� horses�increased� the� PAOC� and� TAR� under� basal� conditions,� with� no� increase� in�response�to�exercise.�
Plasma� antioxidant� capacity� has� been� reported� to� increase� after� exercise� in�horses� (Räsänen�et�al.�1996;�Balogh�et�al.� 2001;�White�et�al.�2001).�An� increased�antioxidant� capacity� in� plasma� or� serum� may� not� necessarily� be� a� desirable�condition� if� it� reflects� a� response� to� increased� oxidative� stress.� It� has� been�reported�that�micronutrient�supplementations�per�se�up�regulate�total�antioxidant�capacity�of�rats�(Skibska�et�al.�2006)�and�horses�(Avellini�et�al.�1999).�Therefore�it�is� possible� that� the� antioxidant� properties� of� LA� preserve� the� antioxidant�capacity�of� the�horse�during�exercise�attenuating�the�amount�of�acute�oxidative�stress� and�decreasing� the�need� for� an� exercise�induced� increase�of� endogenous�antioxidant�levels�(Atalay�et�al.�2006).�Vice�versa,�a�decrease�in�plasma�or�serum�antioxidant� capacity� may� not� necessarily� be� an� undesirable� condition,� if� the�measurement�reflects�decreased�production�of�reactive�species.��
There� are� several� analytical� methods� developed� to� measure� the� total�antioxidant� capacity� in� biological� samples,� since� the� amount� of� different�antioxidants� makes� it� challenging� to� measure� each� of� them� separately.� The�oxygen� radical� absorbance� capacity,� ORAC,� assay� is� used� by� different�

46
laboratories� and� provides� significant� information� regarding� the� antioxidant�capacity�of�various�biological�samples�(Prior�and�Cao�1999).�We�chose�to�use�the�ORAC�assay,�as�it� is�known�as�a�reliable�and�efficient�as�well�as�simple�tool�for�evaluating� the� overall� antioxidant� capacity� in� vivo.� The� use� of� peroxyl� or�hydroxyl� radicals� as� pro�oxidants� in� the� ORAC� assay� makes� it� different� and�unique� from� the� assays� that� involve� oxidants� that� are� not� necessarily� pro�oxidants.� There� are� also� some� improvements� in� quantitation� compared� with�other�assays;�these�are�discussed�in�detail�previously�(Prior�and�Cao�1999).��
The�major� contributors�of�ORAC� in�plasma�are�uric� acid,� ascorbate� (vitamin�C),�glutathione�and���tocopherol� (vitamin�E).�Therefore,� it�should�be�taken�into�account� that�ORAC�may�better�reflect�water�soluble�antioxidant�status� (Prior�et�al.�2003)�than�lipid�phase�antioxidant�status.�The�explanation�depends�also�upon�the�conditions�under�which�the�antioxidant�capacity� is�determined,�because�the�measurement�reflects�outcomes� in�a�dynamic�system�(Prior�and�Cao�1999).�The�interpretation�of� the�changes� in�plasma�or� serum�antioxidant� capacity�becomes�complicated� by� the� diversity� of� the� analytical� methods� and� it� depends� on� the�biochemical�basis�of�the�assay;�e.g.�ferric�reducing�antioxidant�power�(FRAP)�is�a�method� that�does�not� include� thiol� antioxidants� and� therefore� it� should�not� be�used� alone� (Prior� and� Cao� 1999).� Furthermore,� based� on� existing� studies,� no�single�measurement�of�antioxidant�status�will�be�sufficient,�but�the�combination�of�different�measurements�will�be�necessary�to�adequately�assess�oxidative�stress�in�biological�systems�(Prior�and�Cao�1999).��
The� increase� in� plasma� ORAC� levels� following� exercise� could� also� be�attributed� to� increased� capillary� permeability� leading� to� redistribution� of�antioxidants�between�tissues�(Cao�et�al.�1999b).�Increased�production�of�ROS�has�been�postulated�to�decrease�the�antioxidant�capacity�of�tissues�(White�et�al.�2001)�even�after�72�h�after�exercise�(Sacheck�et�al.�2003).�It�has�been�also�speculated�that�increases� in� total� antioxidant� capacity� after� acute� exercise� might� be� due� to�hemoconcentration�(Sürmen�Gür�et�al.�1999;�White�et�al.�2001).�However,� in�the�case� of� original� paper� IV,� the� assumption� is� not� plausible� because� LA�supplementation� blunted� the� exercise�induced� increase� in� plasma� ORAC.�Furthermore,�it�can�be�suggested�that�plasma�ORAC�might�lower�plasma�levels�of� lipid� peroxidation� and� protein� oxidation� products� at� rest.� To� support� this,�higher�post�exercise�MDA�as�well�as�PCarb�levels�in�muscle�correlated�positively�with� increased�plasma�ORAC�during�recovery.�However,� it� is�suggested�earlier�that�the�increase�in�total�antioxidant�capacity�does�not�necessarily�prevent� lipid�peroxidation�in�man�(Alessio�et�al.�2000)�or�in�horse�(White�et�al.�2001)�and�this�is�also�the�case�with�protein�oxidation�(Alessio�et�al.�2000).���

47
6.6 STRESS PROTEIN RESPONSE �Although�it�is�well�documented�that�both�acute�and�chronic�exercise�can�increase�HSP�expression�(Harkins�2009;�Morton�et�al.�2009a),�we�did�not�observe�any�HSP�induction� following� acute� exercise� in� horses� (original� papers� I� and� III).� The�information� available� on� stress� protein� induction� patterns� in� horses� is� limited�and� there� is�only�one� study� reporting�HSP72�mRNA� levels;� and�as� the�authors�emphasise,� it� is� not� certain� if� the� increase� in� HSP72� mRNA� levels� led� to� an�increase�in�protein�levels�(Pösö�et�al.�2002).��
In� the� present� thesis,� the� cytoprotective� role� of� HSPs� in� skeletal� muscle� in�connection� with� strenuous� exercise� is� supported� by� the� negative� correlation�between� the� resting� levels� of� muscle� HSP60� and� HSP25� in� non�supplemented�horses� and� the� recovery� level� of� plasma� aspartate� aminotransferase� (AST)�(original� paper� III).� This� is� in� line� with� the� negative� correlations� between� the�levels� of� plasma� AST�and� HSP25� and� HSP90� and� between� the� plasma� creatine�kinase�(CK)�and�muscle�HSP25�and�HSC70�during�recovery�of�24�and�48�hours�respectively�(original�paper�III).��
The� five�week� ��lipoic� acid� (LA)� supplementation� enhanced� the� skeletal�muscle�HSP�response�(original�paper�III).�The�role�of�LA�as�an�enhancer�of�HSP�induction� is� supported� by� the� previous� study� of� our� group,� where� LA�supplementation� up�regulated� the� mRNA� for� HSP90,� but� had� no� effect� on� the�protein� levels� (Oksala� et� al.� 2007).� Additionally,� LA� supplementation� up�regulated�heat�shock�factor�1�(HSF�1)�at�both�mRNA�and�protein�levels�(Oksala�et�al.�2007).�The�mechanism�by�which�LA�increases�the�basal�levels�of�HSP90�and�the�levels�of�HSP70�during�recovery�remains�to�be�elucidated.�It�is�suggested�that�LA�might�activate�certain,�yet�unidentified,�signalling�intermediates�by�inducing�intramolecular� disulphide� bond� formation,� a� signal� for� oxidant� exposure� and�function�as�an�HSP�inducer�(McCarty�2001;�2006).�Nonetheless,�vitamin�E�and�C�supplementation� have� been� reported� to� decrease� exercise�induced� HSP�expression�in�human�skeletal�muscle�(Khassaf�et�al.�2003;�Fischer�et�al.�2006).�
Although� oxidative� stress� is� one� of� the� physiological� inducers� of� HSP�expression,�these�studies�show�that�exercise�of�this�duration�and�intensity�did�not�significantly� influence� the� levels� of� HSPs� in� the� muscle� of� regularly� trained�trotters� (original� papers� I� and� III).� The� dependence� of� the� HSP70� induction� on�exercise�intensity�has�been�demonstrated�in�humans�(Liu�and�Steinacker�2001).�In�the�studies�reporting�acute�exercise�induced�HSP�expression�in�different�species,�prolonged�and�exhaustive�exercise�protocols�have�been�used�(Salo�et�al.�1991).�
There�are�several� factors� influencing� the�HSP�machinery,�and�one�of� these� is�assumed�to�be�exercise�induced�increase�in�muscular�temperature�(Madden�et�al.�2008).� However,� it� has� recently� been� suggested� that� the� stress� response� might�

48
after�all�be�mediated�by�redox�signaling,�i.e.�transient�and�reversible�oxidation�of�muscle�proteins,�instead�of�the�increases�in�contracting�muscle�temperature�per�se�(Morton�et�al.�2009b).�However,�the�temperature�increase�presumably�retain�the�impact� on� tissue� protection� mechanism� in� exercising� horses,� as� their� skeletal�muscle� mass� is� remarkably� higher� compared� with� other� mammals.� In� general�any� event� that� results� in� protein� unfolding� or� denaturation� �� increased�temperature,� increased�oxidative�stress�itself�or�metabolic�disturbances�–�is�able�to�activate�the�heat�shock�response�(Noble�et�al.�2008).��
We� used� a� relatively� moderate� exercise� protocol,� since� our� aim� was� to�demonstrate� induction� of� HSP,� oxidative� stress� responses� and� muscle� damage�after� a� bout� of� exercise� at� intensity� typical� for� the� daily� activity� of� horses� in�training.�Furthermore,�the�horses�in�these�series�of�studies�had�all�been�in�regular�training�for�several�years,�and�were�well�adapted�to�chronic�training.�The�effect�of� a� single� bout� of� acute� exercise� on� HSP� induction� has� been� reported� to� be�smaller�in�trained�individuals�compared�to�the�untrained�individuals�(Thompson�et�al.�2002).��
Strenuous�training�will�induce�delayed�HSP�responses�in�humans�(Paulsen�et�al.� 2007)� as� well� as� in� rats� (Ogata� et� al.� 2009).� Even� though,� increased� HSP�transcription�occurs�during,�immediately�post�exercise,�or�after�several�hours�of�recovery,� the� time�course� studies� using� western� blotting� techniques� have�demonstrated� a� significant� increase� in� protein� content� not� detectable� until� 1�2�days� following� the� exercise� stress� (Morton� et� al.� 2009b).This� is� particularly�important� when� considering� athletic� horses� and� the� cumulative� effects� of�inadequate�recovery.���6.6.1�Lipid�peroxidation,�protein�oxidation�and�induction�of�HSPs�Oxidation�reactions�are�easily�transferred�from�lipids�to�proteins�(Viljanen�et�al.�2005).� Consistent� with� this� report,� we� detected� a� strong� relationship� between�post�exercise�malondialdehyde�(MDA)�levels�and�protein�carbonyl�(PCarb)�levels�during� recovery� (original� paper� IV).� More� amino� acids� are� damaged� by�secondary� lipid� oxidation� products� such� as� aldehydes� than� by� primary� lipid�oxidation� products� such� as� hydroperoxides� (Matsushita� 1975).� It� has� been�emphasised�that�the�lipid�peroxidation�end�products�are�potent�inhibitors�of�the�chaperone’s�refolding�ability�(Gueraud�et�al.�2010).�Lipid�peroxidation�products�are�able�to�diffuse�across�membranes,�allowing�the�reactive�aldehyde�containing�lipids�to�covalently�modify�proteins� localised�throughout�the�cell�and�relatively�far�away� from� the� initial� site�of�ROS� formation� (Gueraud�et�al.�2010).�This�will�eventually� lead� to� formation� of� carbonyl� groups� in� the� proteins� (Zamora� et� al.�1999).� Mildly� cross� linked,� 4�hydroxynoneal� (HNE)� modified� proteins� are�preferentially�degraded�by�the�proteasome,�but�extensive�modification�with�this�

49
cross�linking�aldehyde�leads�to�the�formation�of�protein�aggregates�(Gueraud�et�al.�2010).�The�carbonyl�groups�in�the�active�centre�of�the�incorrectly�folded�and/or�oxidatively�modified�proteins�may�trigger�the�initial�steps�in�the�degradation�of�the� protein� preventing� the� large� aggregates� of� faulty� proteins� in� cells� under�oxidative� stress� (Shringarpure� et� al.� 2001).� HSP72� is� the� major� inducible� HSP�found�in�the�nucleus�and�cytosol.�Stress�induced�HSP72�protects�proteins�against�aggregation�of�denaturation�(Gueraud�et�al.�2010).��
The�LA�induced�increase�in�the�HSP90�levels�before�the�exercise�may�further�minimise� disruption� of� muscle� homeostasis� during� acute� exercise.� However,� it�can� be� speculated� whether� the� increase� in� HSP90� levels� following� LA�supplementation� is� a� direct� cause� of� supplementation� or� consequence� of�decreased�oxidative�stress�and�LPO�levels.��
�6.6.2�Interaction�between�increased�oxidative�capacity�and�HSP�induction�Our�observation�of� the�decreased� rate� of� lactate� formation�during� exercise� and�simultaneously� increased�skeletal�muscle�CS�activity�as�a�result� from�five�week�LA� supplementation� may� be� explained� by� the� role� of� LA� on� the� oxidative�metabolism�in�mitochondria�(Sen�and�Packer�2000).�According�to�this�hypothesis,�LA� enhances� the� activities� of� pyruvate� dehydrogenase� and� ��ketoglutarate�dehydrogenase� possibly� by� improving� their� co�factor� availability� (Hagen� et� al.�2002).�This,� in� turn,�may� increase�oxidative� capacity�of� the� skeletal�muscle�and�lead�to�improved�performance�in�endurance�events.��
However,� our� group� has� previously� reported� that� intragastrically�supplemented�LA�did�not� increase� the� levels�of� lipoyl� lysine� in�skeletal�muscle�(Khanna� et� al.� 1998).� Lipoyl� lysine� is� the� bound� form� of� LA� and� a� co�factor� of�pyruvate�dehydrogenase�and���ketoglutarate�dehydrogenase� in�skeletal�muscle�(Khanna� et� al.� 1998).� On� the� other� hand,� LA� enhances� glucose� uptake� by� the�muscle�cells�by�increasing�glucose�transporters�in�the�plasma�membrane�(Packer�et� al.� 1997).� Enhanced� glucose� metabolism,� in� turn,� may� increase� pyruvate�availability� for� TCA� cycle� and� result� in� increased� oxidative� capacity� of� the�muscles�(Savitha�et�al.�2005).��
The� LA�induced� increase� in� CS� activity� may� suggest� enhancement� of� the�oxidative� capacity�of� skeletal�muscles�and� increased�performance� in� endurance�events.� We� did� not� observe� any� significant� correlation� between� the� CS� activity�and�either�HSP70�or�HSP90�at�the�protein�level,�it�has�been�reported�earlier�that�HSP72�mRNA� is�markedly�associated�with�muscle�oxidative� capacity� (Bruce�et�al.� 2003).� It� is� unclear� whether� the� increased� recovery� levels� of� muscle� HSP70�after� LA� supplementation� (original� paper� III)� is� a� direct� effect� of� LA� or� a�consequence�of�increased�oxidative�capacity�of�the�skeletal�muscle�following�LA�administration.�

50
7�Future�aspects��Different� muscle� disorders� are� a� common� cause� of� impaired� performance� in�horses.� In� the�near�future,� it�would�be� interesting�to�study�how�the�antioxidant�system�develops�in�horses�and�to�what�extent�it�adapts�to�training.�Furthermore,�the� induction� pattern� of� stress� proteins� and� its� relations� with� equine� muscle�disorders� and� transport� stress� are� an� attractive� research� subject.� This� would�certainly�be�in�the�interest�of�the�horse�industry,�since�sport�horses�are�routinely�transported�across�the�world�and�has�considerable�economical�importance.�

51
8�Conclusions�� �The� present� thesis� provided� the� most� detailed� information� to� date� on� the�response�of� antioxidant� and�HSP�defences� to� exercise�induced�oxidative� stress.�The� previous� studies� have� reported� mainly� the� acute� effects� of� exercise� of�varying� intensity.� Based� on� these� results,� it� can� be� concluded� that� horses� are�exposed� to� exercise�induced�oxidative� stress� and�oral� supplementation�with� ��lipoic�acid�(LA)�may�be�useful�for�trotters�by�decreasing�oxidative�stress�markers�and�boosting�oxidative�metabolism�and�tissue�protection�mechanisms.��
LA� appears� to� act� as� a� redox� regulator� for� thioredoxin�� and� glutathione�systems�as�well�as�a�pro�glutathione�dietary� supplement�diminishing�oxidative�insults� and� fortifying� antioxidant� protection.� However,� it� has� not� yet� been�clarified� whether� the� protein�bound� LA� itself� has� an� antioxidant� activity� or�whether� it� acts� as� a� source� of� free� LA.� The� constant� reduction� of� LA� to�dehydrolipoic� acid� (DHLA)� may� stress� the� energy� generation� capacity� and� the�reducing�power�of�cells,�especially�at�doses�higher�than�in�this�study.�
It� should� nonetheless� be� taken� into� account� that� prolonged� mega�dose�supplementation� of� micronutrients� may� have� several� adverse� health� effects.�Enhancing� tissues� antioxidant� defense� mechanisms� may� serve� as� a� potent�therapeutic� tool,�even� though�currently� tested�natural� compounds�have�not�yet�achieved�any�major�therapeutic�action�yet.���

52
9�References��Adimora�NJ,�Jones�DP,�Kemp�ML.�A�model�of�redox�kinetics�implicates�the�thiol�
proteome�in�cellular�hydrogen�peroxide�responses.�Antioxidants�&�redox�signaling�2011;13(6):731�743.�
Al�Aqil� A,� Zulkifli� I.� Changes� in� heat� shock� protein� 70� expression� and� blood�characteristics� in�transported�broiler�chickens�as�affected�by�housing�and�early�age�feed�restriction.�Poult�Sci�2009;88(7):1358�1364.�
Aleman� M,� Nieto� JE,� Magdesian� KG.� Malignant� hyperthermia� associated� with�ryanodine� receptor� 1� (C7360G)�mutation� in�Quarter�Horses.� J�Vet� Intern�Med�2009;23(2):329�334.�
Alessio� HM.� Exercise�induced� oxidative� stress.� Med� Sci� Sports� Exerc�1993;25(2):218�224.�
Alessio� HM,� Goldfarb� AH.� Lipid� peroxidation� and� scavenger� enzymes� during�exercise:� adaptive� response� to� training.� J� Appl� Physiol� 1988;64(4):1333�1336.�
Alessio�HM,�Goldfarb�AH,�Cao�G.�Exercise�induced�oxidative�stress�before�and�after�vitamin�C�supplementation.�Int�J�Sport�Nutr�1997;7(1):1�9.�
Alessio� HM,� Hagerman� AE,� Fulkerson� BK,� Ambrose� J,� Rice� RE,� Wiley� RL.�Generation� of� reactive� oxygen� species� after� exhaustive� aerobic� and�isometric�exercise.�Med�Sci�Sports�Exerc�2000;32(9):1576�1581.�
Anckar�J,�Sistonen�L.�Regulation�of�HSF1�Function�in�the�Heat�Stress�Response:�Implications�in�Aging�and�Disease.�Annu�Rev�Biochem�2010.�
Anderson� MG.� The� effect� of� exercise� on� the� lactic� dehydrogenase� and� creatine�kinase�isoenzyme�composition�of�horse�serum.�Res�Vet�Sci�1976;20(2):191�196.�
Andersson� A,� Sjödin� A,� Olsson� R,� Vessby� B.� Effects� of� physical� exercise� on�phospholipid� fatty� acid� composition� in� skeletal� muscle.� Am� J� Physiol�1998;274(3�Pt�1):E432�438.�
Arab� K,� Steghens� JP.� Plasma� lipid� hydroperoxides� measurement� by� an�automated�xylenol�orange�method.�Anal�Biochem�2004;325(1):158�163.�
Arbogast�S,�Reid�MB.�Oxidant�activity�in�skeletal�muscle�fibers�is�influenced�by�temperature,� CO2� level,� and� muscle�derived� nitric� oxide.� Am� J� Physiol�Regul�Integr�Comp�Physiol�2004;287(4):R698�705.�
Arnér� ES.� Focus� on� mammalian� thioredoxin� reductases��important�selenoproteins� with� versatile� functions.� Biochim� Biophys� Acta�2009;1790(6):495�526.�

53
Art� T,� Lekeux� P.� Training�induced� modifications� in� cardiorespiratory� and�ventilatory� measurements� in� thoroughbred� horses.� Equine� Vet� J�1993;25(6):532�536.�
Ashton� T,� Rowlands� CC,� Jones� E,� Young� IS,� Jackson� SK,� Davies� B,� Peters� JR.�Electron� spin� resonance� spectroscopic� detection� of� oxygen�centred�radicals� in� human� serum� following� exhaustive� exercise.� Eur� J� Appl�Physiol�Occup�Physiol�1998;77(6):498�502.�
Atalay�M,�Laaksonen�DE,�Khanna�S,�Kaliste�Korhonen�E,�Hanninen�O,�Sen�CK.�Vitamin�E�regulates�changes�in�tissue�antioxidants�induced�by�fish�oil�and�acute�exercise.�Med�Sci�Sports�Exerc�2000;32(3):601�607.�
Atalay�M,� Lappalainen� J,� Sen�CK.�Dietary� antioxidants� for� the� athlete.�Current�sports�medicine�reports�2006;5:182�186.�
Atalay�M,�Oksala�NK,�Laaksonen�DE,�Khanna�S,�Nakao�C,�Lappalainen�J,�Roy�S,�Hanninen� O,� Sen� CK.� Exercise� training� modulates� heat� shock� protein�response�in�diabetic�rats.�J�Appl�Physiol�2004;97(2):605�611.�
Avellini�L,�Chiaradia�E,�Gaiti�A.�Effect�of�exercise�training,�selenium�and�vitamin�E� on� some� free� radical� scavengers� in� horses� (Equus� caballus).� Comp�Biochem�Physiol�B�Biochem�Mol�Biol�1999;123(2):147�154.�
Avellini� L,� Silvestrelli� M,� Gaiti� A.� Training�induced� modifications� in� some�biochemical�defences�against�free�radicals�in�equine�erythrocytes.�Vet�Res�Commun�1995;19(3):179�184.�
Bailey� DM,� Lawrenson� L,� McEneny� J,� Young� IS,� James� PE,� Jackson� SK,� Henry�RR,� Mathieu�Costello� O,� McCord� JM,� Richardson� RS.� Electron�paramagnetic� spectroscopic� evidence� of� exercise�induced� free� radical�accumulation�in�human�skeletal�muscle.�Free�Radic�Res�2007;41(2):182�190.�
Balogh�N,�Gaal�T,�Ribiczeyne�PS,�Petri�A.�Biochemical�and�antioxidant�changes�in�plasma� and� erythrocytes� of� pentathlon� horses� before� and� after� exercise.�Vet�Clin�Pathol�2001;30(4):214�218.�
Bao�E,�Sultan�KR,�Bernhard�N,�Hartung� J.�Expression�of�heat�shock�proteins� in�tissues� from� young� pigs� exposed� to� transport� stress.� Dtsch� Tierarztl�Wochenschr�2009;116(9):321�325.�
Barclay�JK,�Hansel�M.�Free�radicals�may�contribute�to�oxidative�skeletal�muscle�fatigue.�Can�J�Physiol�Pharmacol�1991;69(2):279�284.�
Barret� AJ.� Lysosomal� enzymes.� In:� JT� Dingle� (ed.)� Lysosomes,� a� laboratory�handbook.�Amsterdam.�North�Holland.�Netherlands.�
Berzosa� C,� Gomez�Trullen� EM,� Piedrafita� E,� Cebrian� I,� Martinez�Ballarin� E,�Miana�Mena�FJ,�Fuentes�Broto�L,�Garcia�JJ.�Erythrocyte�membrane�fluidity�and�indices�of�plasmatic�oxidative�damage�after�acute�physical�exercise�in�humans.�Eur�J�Appl�Physiol�2010.�

54
Bhupathiraju� SN,� Tucker� KL.� Coronary� heart� disease� prevention:� Nutrients,�foods,�and�dietary�patterns.�Clin�Chim�Acta�2011;412(17�18):1493�1514.�
Bloomer� RJ,� Davis� PG,� Consitt� LA,� Wideman� L.� Plasma� protein� carbonyl�response� to� increasing� exercise� duration� in� aerobically� trained� men� and�women.�Int�J�Sports�Med�2007;28(1):21�25.�
Bloomer� RJ,� Goldfarb� AH,� McKenzie� MJ.� Oxidative� stress� response� to� aerobic�exercise:� comparison� of� antioxidant� supplements.� Med� Sci� Sports� Exerc�2006;38(6):1098�1105.�
Bloomer� RJ,� Goldfarb� AH,� Wideman� L,� McKenzie� MJ,� Consitt� LA.� Effects� of�acute�aerobic�and�anaerobic�exercise�on�blood�markers�of�oxidative�stress.�J�Strength�Cond�Res�2005;19(2):276�285.�
Brady�PS,�Ku�PK,�Ullrey�DE.�Lack�of�effect�of�selenium�supplementation�on�the�response� of� the� equine� erythrocyte� glutathione� system� and� plasma�enzymes�to�exercise.�J�Anim�Sci�1978;47(2):492�496.�
Brady�PS,�Shelle�JE,�Ullrey�DE.�Rapid�changes�in�equine�erythrocyte�glutathione�reductase�with�exercise.�Am�J�Vet�Res�1977;38(7):1045�1047.�
Breusing�N,�Grune�T,�Andrisic�L,�Atalay�M,�Bartosz�G,�Biasi�F,�Borovic�S,�Bravo�L,�Casals� I,�Casillas�R,�Dinischiotu�A,�Drzewinska�J,�Faber�H,�Fauzi�NM,�Gajewska� A,� Gambini� J,� Gradinaru� D,� Kokkola� T,� Lojek� A,� Luczaj� W,�Margina�D,�Mascia�C,�Mateos�R,�Meinitzer�A,�Mitjavila�MT,�Mrakovcic�L,�Munteanu�MC,�Podborska�M,�Poli�G,�Sicinska�P,�Skrzydlewska�E,�Vina�J,�Wiswedel� I,� Zarkovic� N,� Zelzer� S,� Spickett� CM.� An� inter�laboratory�validation�of�methods�of�lipid�peroxidation�measurement�in�UVA�treated�human�plasma�samples.�Free�Radic�Res�2010;44(10):1203�1215.�
Bruce�CR,�Carey�AL,�Hawley�JA,�Febbraio�MA.�Intramuscular�heat�shock�protein�72� and� heme� oxygenase�1� mRNA� are� reduced� in� patients� with� type� 2�diabetes:� evidence� that� insulin� resistance� is� associated� with� a� disturbed�antioxidant�defense�mechanism.�Diabetes�2003;52(9):2338�2345.�
Brzeszczynska� J,� Pieniazek� A,� Gwozdzinski� L,� Gwozdzinski� K,� Jegier� A.�Structural� alterations� of� erythrocyte� membrane� components� induced� by�exhaustive�exercise.�Appl�Physiol�Nutr�Metab�2008;33(6):1223�1231.�
Burhans�WC,�Heintz�NH.�The�cell� cycle� is�a� redox�cycle:� linking�phase�specific�targets�to�cell�fate.�Free�Radic�Biol�Med�2009;47(9):1282�1293.�
Burk�RF.�Biological�activity�of�selenium.�Annu�Rev�Nutr�1983;3:53�70.�Cadenas� E.� Basic� mechanisms� of� antioxidant� activity.� Biofactors� 1997;6(4):391�
397.�Calderwood�SK,�Murshid�A,�Prince�T.�The�shock�of�aging:�molecular�chaperones�
and� the� heat� shock� response� in� longevity� and� aging��a� mini�review.�Gerontology�2009;55(5):550�558.�

55
Cao�G,�Shukitt�Hale�B,�Bickford�PC,�Joseph�JA,�McEwen�J,�Prior�RL.�Hyperoxia�induced� changes� in� antioxidant� capacity� and� the� effect� of� dietary�antioxidants.�J�Appl�Physiol�1999b;86(6):1817�1822.�
Carbone�DL,�Doorn� JA,�Kiebler�Z,� Ickes�BR,�Petersen�DR.�Modification�of�heat�shock�protein�90�by�4�hydroxynonenal�in�a�rat�model�of�chronic�alcoholic�liver�disease.�J�Pharmacol�Exp�Ther�2005;315(1):8�15.�
Carbone�DL,�Doorn�JA,�Kiebler�Z,�Sampey�BP,�Petersen�DR.�Inhibition�of�Hsp72�mediated� protein� refolding� by� 4�hydroxy�2�nonenal.� Chem� Res� Toxicol�2004;17(11):1459�1467.�
Cazzola� R,� Russo�Volpe� S,� Cervato� G,� Cestaro� B.� Biochemical� assessments� of�oxidative�stress,�erythrocyte�membrane�fluidity�and�antioxidant�status�in�professional� soccer� players� and� sedentary� controls.� Eur� J� Clin� Invest�2003;33(10):924�930.�
Chae� CH,� Shin� CH,� Kim� HT.� The� combination� of� alpha�lipoic� acid�supplementation�and�aerobic�exercise�inhibits�peroxidation�in�rat�skeletal�muscles.�Nutr�Res�2008;28(6):399�405.�
Chiaradia�E,�Avellini�L,�Rueca�F,�Spaterna�A,�Porciello�F,�Antonioni�MT,�Gaiti�A.�Physical� exercise,� oxidative� stress� and� muscle� damage� in� racehorses.�Comp�Biochem�Physiol�B�Biochem�Mol�Biol�1998;119(4):833�836.�
Child� RB,� Wilkinson� DM,� Fallowfield� JL,� Donnelly� AE.� Elevated� serum�antioxidant� capacity� and� plasma� malondialdehyde� concentration� in�response� to� a� simulated� half�marathon� run.� Med� Sci� Sports� Exerc�1998;30(11):1603�1607.�
Childs�A,�Jacobs�C,�Kaminski�T,�Halliwell�B,�Leeuwenburgh�C.�Supplementation�with�vitamin�C�and�N�acetyl�cysteine�increases�oxidative�stress�in�humans�after�an�acute�muscle�injury�induced�by�eccentric�exercise.�Free�Radic�Biol�Med�2001;31(6):745�753.�
Clanton� TL,� Zuo� L,� Klawitter� P.� Oxidants� and� skeletal� muscle� function:�physiologic� and� pathophysiologic� implications.� Proc� Soc� Exp� Biol� Med�1999;222(3):253�262.�
Clarkson� PM.� Antioxidants� and� physical� performance.� Crit� Rev� Food� Sci� Nutr�1995;35(1�2):131�141.�
Dadgar� S,� Lee� ES,� Leer� TL,� Burlinguette� N,� Classen� HL,� Crowe� TG,� Shand� PJ.�Effect� of� microclimate� temperature� during� transportation� of� broiler�chickens� on� quality� of� the� pectoralis� major� muscle.� Poult� Sci�2010;89(5):1033�1041.�
Dalle�Donne� I,� Aldini� G,� Carini� M,� Colombo� R,� Rossi� R,� Milzani� A.� Protein�carbonylation,� cellular� dysfunction,� and� disease� progression.� J� Cell� Mol�Med�2006;10(2):389�406.�

56
Davies� KJ,� Quintanilha� AT,� Brooks� GA,� Packer� L.� Free� radicals� and� tissue�damage� produced� by� exercise.� Biochem� Biophys� Res� Commun�1982;107(4):1198�1205.�
Davies�KJ,�Shringarpure�R.�Preferential�degradation�of�oxidized�proteins�by�the�20S� proteasome� may� be� inhibited� in� aging� and� in� inflammatory�neuromuscular�diseases.�Neurology�2006;66(2�Suppl�1):S93�96.�
Davies� MJ,� Fu� S,� Wang� H,� Dean� RT.� Stable� markers� of� oxidant� damage� to�proteins�and�their�application� in� the�study�of�human�disease.�Free�Radic�Biol�Med�1999;27(11�12):1151�1163.�
Davison� GW,� Ashton� T,� Davies� B,� Bailey� DM.� In� vitro� electron� paramagnetic�resonance� characterization�of� free� radicals:� relevance� to� exercise�induced�lipid�peroxidation�and� implications�of� ascorbate�prophylaxis.� Free�Radic�Res�2008;42(4):379�386.�
De� Leenheer� AP,� De� Bevere� VO,� De� Ruyter� MG,� Claeys� AE.� Simultaneous�determination�of� retinol� and�alpha�tocopherol� in�human� serum�by�high�performance�liquid�chromatography.�J�Chromatogr�1979;162(3):408�413.�
de� Moffarts� B,� Kirschvink� N,� Art� T,� Pincemail� J,� Lekeux� P.� Effect� of� oral�antioxidant� supplementation� on� blood� antioxidant� status� in� trained�thoroughbred�horses.�Vet�J�2005;169(1):65�74.�
de�Moffarts�B,�Kirschvink�N,�Art�T,�Pincemail�J,�Lekeux�P.�Effect�of�exercise�on�blood� oxidant/antioxidant� markers� in� standardbred� horses:� comparison�between� treadmill� and� race� track� tests.� Equine� Vet� J� Suppl� 2006(36):254�257.�
de�Moffarts�B,�Kirschvink�N,�Art�T,�Pincemail�J,�Michaux�C,�Cayeux�K,�Defraigne�JO,� Lekeux� P.� Impact� of� training� and� exercise� intensity� on� blood�antioxidant�markers�in�healthy�standardbred�horses.�Equine�Comp�Exerc�Physiol�2004;1:1�11.�
de� Moffarts� B,� Portier� K,� Kirschvink� N,� Coudert� J,� Fellmann� N,� van� Erck� E,�Letellier�C,�Motta�C,�Pincemail�J,�Art�T,�Lekeux�P.�Effects�of�exercise�and�oral� antioxidant� supplementation� enriched� in� (n�3)� fatty� acids� on� blood�oxidant� markers� and� erythrocyte� membrane� fluidity� in� horses.� Vet� J�2007;174(1):113�121.�
Dean� RT,� Fu� S,� Stocker� R,� Davies� MJ.� Biochemistry� and� pathology� of� radical�mediated�protein�oxidation.�Biochem�J�1997;324�(�Pt�1):1�18.�
Deaton�CM,�Marlin�DJ,�Roberts�CA,�Smith�N,�Harris�PA,�Kelly�FJ,�Schroter�RC.�Antioxidant� supplementation� and� pulmonary� function� at� rest� and�exercise.�Equine�Vet�J�Suppl�2002(34):58�65.�
Demasi� M,� Shringarpure� R,� Davies� KJ.� Glutathiolation� of� the� proteasome� is�enhanced� by� proteolytic� inhibitors.� Arch� Biochem� Biophys�2001;389(2):254�263.�

57
Denborough�M.�Malignant�hyperthermia.�Lancet�1998;352(9134):1131�1136.�Derman� KD,� Noakes� TD.� Comparative� aspects� of� Exercise� Physiology.� In:� DR�
Hodgson,�JR�Rose�(eds.)�The�Athletic�Horse.�Philadelphia.�W.B.�Saunders�Company.�p.�497,�pp.�13�26.�
Doorn�JA,�Petersen�DR.�Covalent�modification�of�amino�acid�nucleophiles�by�the�lipid� peroxidation� products� 4�hydroxy�2�nonenal� and� 4�oxo�2�nonenal.�Chem�Res�Toxicol�2002;15(11):1445�1450.�
Dröge�W.�Free�radicals�in�the�physiological�control�of�cell�function.�Physiol�Rev�2002;82(1):47�95.�
Duthie� GG,� Robertson� JD,� Maughan� RJ,� Morrice� PC.� Blood� antioxidant� status�and� erythrocyte� lipid� peroxidation� following� distance� running.� Arch�Biochem�Biophys�1990;282(1):78�83.�
El�Deeb� WM,� El�Bahr� SM.� Investigation� of� selected� biochemical� indicators� of�Equine� Rhabdomyolysis� in� Arabian� horses:� pro�inflammatory� cytokines�and�oxidative�stress�markers.�Vet�Res�Commun�2011;34(8):677�689.�
Essig� DA,� Nosek� TM.� Muscle� fatigue� and� induction� of� stress� protein� genes:� a�dual� function� of� reactive� oxygen� species?� Can� J� Appl� Physiol�1997;22(5):409�428.�
Esterbauer� H,� Schaur� RJ,� Zollner� H.� Chemistry� and� biochemistry� of� 4�hydroxynonenal,� malonaldehyde� and� related� aldehydes.� Free� Radic� Biol�Med�1991;11(1):81�128.�
Evans�DL.�The�Cardiovascular�System:�Anatomy,�Physiology,� and�Adaptations�to� Exercise� and� Training.� In:� DR� Hodgson,� JR� Rose� (eds.)� The� Athletic�Horse.�Philadelphia.�W.B.�Saunders�Company.�p.�497,�pp.�129�144.�
Fehrenbach�E,�Niess�AM,�Veith�R,�Dickhuth�HH,�Northoff�H.�Changes�of�HSP72�expression�in�leukocytes�are�associated�with�adaptation�to�exercise�under�conditions� of� high� environmental� temperature.� J� Leukoc� Biol�2001;69(5):747�754.�
Ferreira� LF,� Reid� MB.� Muscle�derived� ROS� and� thiol� regulation� in� muscle�fatigue.�J�Appl�Physiol�2008;104(3):853�860.�
Filomeni� G,� Rotilio� G,� Ciriolo� MR.� Cell� signalling� and� the� glutathione� redox�system.�Biochem�Pharmacol�2002;64(5�6):1057�1064.�
Fischer�CP,�Hiscock�NJ,�Basu�S,�Vessby�B,�Kallner�A,�Sjoberg�LB,�Febbraio�MA,�Pedersen�BK.�Vitamin�E�isoform�specific�inhibition�of�the�exercise�induced�heat� shock� protein� 72� expression� in� humans.� J� Appl� Physiol�2006;100(5):1679�1687.�
Fisher�Wellman� K,� Bell� HK,� Bloomer� RJ.� Oxidative� stress� and� antioxidant�defense� mechanisms� linked� to� exercise� during� cardiopulmonary� and�metabolic�disorders.�Oxid�Med�Cell�Longev�2009a;2(1):43�51.�

58
Fisher�Wellman� K,� Bloomer� RJ.� Acute� exercise� and� oxidative� stress:� a� 30� year�history.�Dyn�Med�2009b;8:1.�
Forman� HJ.� Reactive� oxygen� species� and� alpha,beta�unsaturated� aldehydes� as�second�messengers�in�signal�transduction.�Ann�N�Y�Acad�Sci�2010;1203:35�44.�
Forman� HJ,� Zhang� H,� Rinna� A.� Glutathione:� overview� of� its� protective� roles,�measurement,�and�biosynthesis.�Mol�Aspects�Med�2009;30(1�2):1�12.�
Gerard�Monnier� D,� Erdelmeier� I,� Regnard� K,� Moze�Henry� N,� Yadan� JC,�Chaudiere�J.�Reactions�of�1�methyl�2�phenylindole�with�malondialdehyde�and�4�hydroxyalkenals.�Analytical�applications� to�a�colorimetric�assay�of�lipid�peroxidation.�Chem�Res�Toxicol�1998;11(10):1176�1183.�
Gerschman�R,�Gilbert�DL,�Nye�SW,�Dwyer�P,�Fenn�WO.�Oxygen�poisoning�and�x�irradiation:�a�mechanism�in�common.�Science�1954a;119(3097):623�626.�
Gerschman� R,� Gilbert� DL,� Nye� SW,� Fenn� WO.� Influence� of� x�irradiation� on�oxygen�poisoning�in�mice.�Proc�Soc�Exp�Biol�Med�1954b;86(1):27�29.�
Ghibu� S,� Richard� C,� Delemasure� S,� Vergely� C,� Mogosan� C,� Muresan� A.� [An�endogenous� dithiol� with� antioxidant� properties:� alpha�lipoic� acid,�potential� uses� in� cardiovascular� diseases].� Ann� Cardiol� Angeiol� (Paris)�2008;57(3):161�165.�
Golbidi�S,�Laher� I.�Molecular�mechanisms�in�exercise�induced�cardioprotection.�Cardiol�Res�Pract�2010;2011:972807.�
Goldfarb�AH,�Bloomer�RJ,�McKenzie�MJ.�Combined�antioxidant�treatment�effects�on� blood� oxidative� stress� after� eccentric� exercise.� Med� Sci� Sports� Exerc�2005a;37(2):234�239.�
Goldfarb�AH,�Garten�RS,�Chee�PD,�Cho�C,�Reeves�GV,�Hollander�DB,�Thomas�C,�Aboudehen� KS,� Francois� M,� Kraemer� RR.� Resistance� exercise� effects� on�blood� glutathione� status� and� plasma� protein� carbonyls:� Influence� of�partial�vascular�occlusion.�Eur�J�Appl�Physiol�2008;104(5):813�819.�
Goldfarb� AH,� McKenzie� MJ,� Bloomer� RJ.� Gender� comparisons� of� exercise�induced�oxidative�stress:� influence�of�antioxidant�supplementation.�Appl�Physiol�Nutr�Metab�2007;32(6):1124�1131.�
Goldfarb� AH,� Patrick� SW,� Bryer� S,� You� T.� Vitamin� C� supplementation� affects�oxidative�stress�markers�in�response�to�a�30�minute�run�at�75%�VO2max.�Int�J�Sport�Nutr�Exerc�Metab�2005b;15(3):279�290.�
Gordillo� GM,� Atalay� M,� Roy� S,� Sen� CK.� Hemangioma� model� for� in� vivo�angiogenesis:� inducible�oxidative�stress�and�MCP�1�expression�in�EOMA�cells.�Methods�Enzymol�2002;352:422�432.�
Grimm� S,� Hohn� A,� Grune� T.� Oxidative� protein� damage� and� the� proteasome.�Amino�Acids�2010.�

59
Grune�T,�Merker�K,�Jung�T,�Sitte�N,�Davies�KJ.�Protein�oxidation�and�degradation�during�postmitotic�senescence.�Free�Radic�Biol�Med�2005;39(9):1208�1215.�
Grune�T,�Siems�WG,�Petras�T.� Identification�of�metabolic�pathways�of� the� lipid�peroxidation�product� 4�hydroxynonenal� in� in� situ�perfused� rat�kidney.� J�Lipid�Res�1997;38(8):1660�1665.�
Gueraud�F,�Atalay�M,�Bresgen�N,�Cipak�A,�Eckl�PM,�Huc�L,�Jouanin�I,�Siems�W,�Uchida� K.� Chemistry� and� biochemistry� of� lipid� peroxidation� products.�Free�Radic�Res�2010;44(10):1098�1124.�
Gupte�AA,�Bomhoff�GL,�Morris�JK,�Gorres�BK,�Geiger�PC.�Lipoic�acid�increases�heat� shock� protein� expression� and� inhibits� stress� kinase� activation� to�improve�insulin�signaling�in�skeletal�muscle�from�high�fat�fed�rats.�J�Appl�Physiol�2009;106(4):1425�1434.�
Hagen� TM,� Liu� J,� Lykkesfeldt� J,� Wehr� CM,� Ingersoll� RT,� Vinarsky� V,�Bartholomew�JC,�Ames�BN.�Feeding�acetyl�L�carnitine�and� lipoic�acid� to�old� rats� significantly� improves� metabolic� function� while� decreasing�oxidative�stress.�Proc�Natl�Acad�Sci�U�S�A�2002;99(4):1870�1875.�
Halliwell� B,� Gutteridge� JMC.� Free� radicals� in� biology� and� medicine:� Oxford�University�Press,�2007.�
Hargreaves�BJ,�Kronfeld�DS,�Waldron�JN,�Lopes�MA,�Gay�LS,�Saker�KE,�Cooper�WL,� Sklan� DJ,� Harris� PA.� Antioxidant� status� and� muscle� cell� leakage�during�endurance�exercise.�Equine�Vet�J�Suppl�2002a(34):116�121.�
Hargreaves�BJ,�Kronfeld�DS,�Waldron�JN,�Lopes�MA,�Gay�LS,�Saker�KE,�Cooper�WL,�Sklan�DJ,�Harris�PA.�Antioxidant�status�of�horses�during�two�80�km�endurance�races.�J�Nutr�2002b;132(6�Suppl�2):1781S�1783S.�
Harkins�MS.�Exercise�regulates�heat�shock�proteins�and�nitric�oxide.�Exerc�Sport�Sci�Rev�2009;37(2):73�77.�
Harris� RC,� Marlin� DJ,� Dunnett� M,� Snow� DH,� Hultman� E.� Muscle� buffering�capacity�and�dipeptide�content�in�the�thoroughbred�horse,�greyhound�dog�and�man.�Comp�Biochem�Physiol�A�Comp�Physiol�1990;97(2):249�251.�
Hayes� JD,� McLellan� LI.� Glutathione� and� glutathione�dependent� enzymes�represent�a�co�ordinately�regulated�defence�against�oxidative�stress.�Free�Radic�Res�1999;31:273�300.�
Henneke�DR,�Potter�GD,�Kreider�JL,�Yeates�BF.�Relationship�between�condition�score,�physical�measurements� and�body� fat�percentage� in�mares.�Equine�Vet�J�1983;15(4):371�372.�
Hoffman�JR,�Im�J,�Kang�J,�Maresh�CM,�Kraemer�WJ,�French�D,�Nioka�S,�Kime�R,�Rundell� KW,� Ratamess� NA,� Faigenbaum� AD,� Chance� B.� Comparison� of�low�� and� high�intensity� resistance� exercise� on� lipid� peroxidation:� role� of�muscle�oxygenation.�J�Strength�Cond�Res�2007;21(1):118�122.�

60
Hoffman� RM,� Hess� TM,� Williams� CA,� Kronfeld� DS,� Griewe�Crandell� KM,�Waldron�JE,�Graham�Thiers�PM,�Gay�LS,�Splan�RK,�Saker�KE,�Harris�PA.�Speed�associated�with�plasma�pH,�oxygen�content,�total�protein�and�urea�in�an�80�km�race.�Equine�Vet�J�Suppl�2002(34):39�43.�
Holmgren� A.� Antioxidant� function� of� thioredoxin� and� glutaredoxin� systems.�Antioxidants�&�redox�signaling�2000;2(4):811�820.�
Hyyppä� S,� Pösö� AR.� Fluid,� electrolyte,� and� acid�base� responses� to� exercise� in�racehorses.�Vet�Clin�North�Am�Equine�Pract�1998;14(1):121�136.�
Ishida� N,� Hobo� S,� Takahashi� T,� Nanbo� Y,� Sato� F,� Hasegawa� T,� Mukoyama� H.�Chronological� changes� in� superoxide�scavenging� ability� and� lipid�peroxide� concentration� of� equine� serum� due� to� stress� from� exercise� and�transport.�Equine�Vet�J�Suppl�1999;30:430�433.�
Jackson�MJ.�Redox�regulation�of�skeletal�muscle.�IUBMB�Life�2008a;60(8):497�501.�Jackson� MJ.� Free� radicals� generated� by� contracting� muscle:� By�products� of�
metabolism� or� key� regulators� of� muscle� function?� Free� Rad� Biol� Med�2008b;44:132�141.�
Jackson�MJ.�Skeletal�muscle�aging:�role�of�reactive�oxygen�species.�Crit�Care�Med�2009a;37(10�Suppl):S368�371.�
Ji� LL.� Antioxidant� enzyme� response� to� exercise� in� equine� erythrocytes.� Equine�Veterinary�Science�1990;10(5):380�383.�
Ji�LL.�Exercise�and�oxidative�stress:�role�of�the�cellular�antioxidant�systems.�Exerc�Sport�Sci�Rev�1995;23:135�166.�
Ji� LL,� Leeuwenburgh� C,� Leichtweis� S,� Gore� M,� Fiebig� R,� Hollander� J,� Bejma� J.�Oxidative� stress� and� aging.� Role� of� exercise� and� its� influences� on�antioxidant�systems.�Ann�N�Y�Acad�Sci�1998;854:102�117.�
Jones�D.�Redefining�oxidative�stress.�Antioxidants�&�redox�signaling�2006;8:1865�1879.�
Jones� DP.� Radical�free� biology� of� oxidative� stress.� Am� J� Physiol� Cell� Physiol�2008;295(4):C849�868.�
Jones�DP,�Liang�Y.�Measuring� the�poise�of� thiol/disulfide� couples� in�vivo.�Free�Radic�Biol�Med�2009;47(10):1329�1338.�
Kaikkonen� J,� Porkkala�Sarataho� E,� Tuomainen� TP,� Nyyssonen� K,� Kosonen� L,�Ristonmaa�U,�Lakka� HM,�Salonen�R,�Korpela�H,� Salonen� JT.�Exhaustive�exercise� increases�plasma/serum� total� oxidation� resistance� in�moderately�trained�men�and�women,�whereas�their�VLDL�+�LDL�lipoprotein�fraction�is� more� susceptible� to� oxidation.� Scand� J� Clin� Lab� Invest� 2002;62(8):599�607.�
Karren� BJ,� Thorson� JF,� Cavinder� CA,� Hammer� CJ,� Coverdale� JA.� Effect� of�selenium�supplementation�and�plane�of�nutrition�on�mares�and�their�foals:�

61
selenium� concentrations� and� glutathione� peroxidase.� J� Anim� Sci�2010;88(3):991�997.�
Kavazis� AN.� Exercise� preconditioning� of� the� myocardium.� Sports� Med�2009;39(11):923�935.�
Kelly�DA,�Tiidus�PM,�Houston�ME,�Noble�EG.�Effect� of� vitamin�E�deprivation�and�exercise�training�on�induction�of�HSP70.�J�Appl�Physiol�1996;81:2379�2385.�
Khanna� S,� Atalay� M,� Laaksonen� DE,� Gul� M,� Roy� S,� Sen� CK.� Alpha�lipoic� acid�supplementation:�tissue�glutathione�homeostasis�at�rest�and�after�exercise.�J�Appl�Physiol�1999a;86(4):1191�1196.�
Khanna�S,�Atalay�M,�Lodge�JK,�Laaksonen�DE,�Roy�S,�Hänninen�O,�Packer�L,�Sen�CK.�Skeletal�muscle�and�liver�lipoyllysine�content�in�response�to�exercise,�training�and�dietary�alpha�lipoic�acid�supplementation.�Biochem�Mol�Biol�Int�1998;46(2):297�306.�
Khassaf� M,� McArdle� A,� Esanu� C,� Vasilaki� A,� McArdle� F,� Griffiths� RD,� Brodie�DA,�Jackson�MJ.�Effect�of�vitamin�C�supplements�on�antioxidant�defence�and�stress�proteins�in�human�lymphocytes�and�skeletal�muscle.�J�Physiol�2003;549(Pt�2):645�652.�
Kingsley�MI,�Kilduff�LP,�McEneny� J,�Dietzig�RE,�Benton�D.�Phosphatidylserine�supplementation� and� recovery� following� downhill� running.� Med� Sci�Sports�Exerc�2006;38(9):1617�1625.�
Kinnunen� S,� Hyyppa� S,� Lehmuskero� A,� Oksala� N,� Maenpaa� P,� Hanninen� O,�Atalay� M.� Oxygen� radical� absorbance� capacity� (ORAC)� and� exercise�induced�oxidative�stress�in�trotters.�Eur�J�Appl�Physiol�2005b;95(5�6):550�556.�
Kirschvink� N,� de� Moffarts� B,� Farnir� F,� Pincemail� J,� Lekeux� P.� Investigation� of�blood� oxidant/antioxidant� markers� in� healthy� competition� horses� of�different�breeds.�Equine�Vet�J�Suppl�2006(36):239�244.�
Kirschvink�N,�Smith�N,�Fievez�L,�Bougnet�V,�Art�T,�Degand�G,�Marlin�D,�Roberts�C,�Genicot�B,�Lindsey�P,�Lekeux�P.�Effect�of�chronic�airway�inflammation�and�exercise�on�pulmonary�and�systemic�antioxidant�status�of�healthy�and�heaves�affected�horses.�Equine�Vet�J�2002;34(6):563�571.�
Knight� JA.� Free� radicals:� their� history� and� current� status� in� aging� and� disease.�Ann�Clin�Lab�Sci�1998;28(6):331�346.�
Kondo� N,� Nakamura� H,� Masutani� H,� Yodoi� J.� Redox� regulation� of� human�thioredoxin� network.� Antioxidants� &� redox� signaling� 2006;8(9�10):1881�1890.�
Kumaran�S,�Savitha�S,�Anusuya�Devi�M,�Panneerselvam�C.�L�Carnitine�and�DL�alpha�lipoic�acid�reverse�the�age�related�deficit�in�glutathione�redox�state�in�skeletal�muscle�and�heart�tissues.�Mech�Ageing�Dev�2004;125:507�512.�

62
Lamprecht� M,� Oettl� K,� Schwaberger� G,� Hofmann� P,� Greilberger� JF.� Protein�modification� responds� to� exercise� intensity� and� antioxidant�supplementation.�Med�Sci�Sports�Exerc�2009;41(1):155�163.�
Lappalainen� Z,� Lappalainen� J,� Oksala� NK,� Laaksonen� DE,� Khanna� S,� Sen� CK,�Atalay� M.� Exercise� training� and� experimental� diabetes� modulate� heat�shock�protein�response�in�brain.�Scand�J�Med�Sci�Sports�2010;20(1):83�89.�
Lawler� JM,� Powers� SK,� Criswell� DS.� Inducibility� of� NADP�specific� isocitrate�dehydrogenase�with� endurance� training� in� skeletal�muscle.�Acta�Physiol�Scand�1993a;149(2):177�181.�
Lawler� JM,�Powers�SK,�Van�Dijk�H,�Visser�T,�Kordus�MJ,� Ji�LL.�Metabolic� and�antioxidant�enzyme�activities� in� the�diaphragm:�effects�of�acute�exercise.�Respir�Physiol�1994;96(2�3):139�149.�
Lawler� JM,�Powers�SK,�Visser�T,�Van�Dijk�H,�Kordus�MJ,� Ji�LL.�Acute�exercise�and� skeletal� muscle� antioxidant� and� metabolic� enzymes:� effects� of� fiber�type�and�age.�Am�J�Physiol�1993b;265(6�Pt�2):R1344�1350.�
Levine�RL,�Stadtman�ER.�Oxidative�modification�of�proteins�during�aging.�Exp�Gerontol�2001;36(9):1495�1502.�
Li� JX,� Tong� CW,� Xu� DQ,� Chan� KM.� Changes� in� membrane� fluidity� and� lipid�peroxidation�of�skeletal�muscle�mitochondria�after�exhausting�exercise�in�rats.�Eur�J�Appl�Physiol�Occup�Physiol�1999;80(2):113�117.�
Lillig� CH,� Holmgren� A.� Thioredoxin� and� related� molecules� �� from� biology� to�health�and�disease.�Antioxidants�&�redox�signaling�2007;9(1):25�47.�
Lindholm�A,�Bjerneld�H,�Saltin�B.�Glycogen�depletion�pattern�in�muscle�fibres�of�trotting�horses.�Acta�Physiol�Scand�1974a;90(2):475�484.�
Lindholm�A,�Piehl�K.�Fibre�composition,�enzyme�activity�and�concentrations�of�metabolites�and�electrolytes� in�muscles�of�standardbred�horses.�Acta�Vet�Scand�1974b;15(3):287�309.�
Liu� Y,� Steinacker� JM.� Changes� in� skeletal� muscle� heat� shock� proteins:�pathological�significance.�Front�Biosci�2001;6:D12�25.�
Locke�M.�The�cellular�response�to�exercise:�role�of�stress�proteins.�Exerc�Sport�Sci�Rev�1997;25:105�136.�
Madden� LA,� Sandstrom� ME,� Lovell� RJ,� McNaughton� L.� Inducible� heat� shock�protein� 70� and� its� role� in� preconditioning� and� exercise.� Amino� Acids�2008;34(4):511�516.�
Marañón� G,� Muñoz�Escassi� B,� Manley� W,� García� C,� Cayado� P,� Sánchez� de� la�Muela� M,� Olábarri� B,� León� R,� Vara� E.� The� effect� of� methyl� sulphonyl�methane� supplementation� on� biomarkers� of� oxidative� stress� in� sport�horses� following� jumping� exercise.� Acta� Veterinaria� Scandinavica�2008;50:45.�

63
Marlin� DJ,� Fenn� K,� Smith� N,� Deaton� CD,� Roberts� CA,� Harris� PA,� Dunster� C,�Kelly� FJ.� Changes� in� circulatory� antioxidant� status� in� horses� during�prolonged�exercise.�J�Nutr�2002;132(6�Suppl�2):1622S�1627S.�
Matsuki� N,� Tamura� S,� Ono� K,� Watari� T,� Goitsuka� R,� Yamanobe� A,� Hiraga� A,�Kubo� K,� Takagi� S,� Hasegawa� A,� et� al.� Exercise�induced� phospholipid�degradation�in�the�equine�skeletal�muscle�and�erythrocytes.�J�Vet�Med�Sci�1991;53(6):1001�1007.�
Matsushita� S.� Specific� interactions� of� linoleic� acid� hydroperoxides� and� their�secondary�degraded�products�with�enzyme�proteins.� J�Agric�Food�Chem�1975;23(2):150�154.�
McArdle� A,� van� der� Meulen� JH,� Catapano� M,� Symons� MCR,� Faulkner� JA,�Jackson� MJ.� Free� radical� activity� following� contraction�induced� injury� to�the� extensor� digitorum� longus� muscles� of� rats.� Free� Rad� Biol� Med�1999;26(9/10):1085�1091.�
McCarty�MF.�Versatile�cytoprotective�activity�of�lipoic�acid�may�reflect�its�ability�to�activate�signalling�intermediates�that�trigger�the�heat�shock�and�phase�II�reactions.�Med�Hypotheses�2001;57(3):313�317.�
McCarty� MF.� Induction� of� heat� shock� proteins� may� combat� insulin� resistance.�Med�Hypotheses�2006;66:527�534.�
McMeniman� NP,� Hintz� HF.� Effect� of� vitamin� E� status� on� lipid� peroxidation� in�exercised�horses.�Equine�Vet�J�1992;24(6):482�484.�
Meyer�H.�Pferdefütterung:�Blackwell�Wissenschafts�Verlag,�Berlin,�1996.�Michel�T,�Vanhoutte�PM.�Cellular�signaling�and�NO�production.�Pflugers�Arch�
2010;459(6):807�816.�Michelotto�PV,�Jr.,�Muehlmann�LA,�Zanatta�AL,�Bieberbach�EW,�Fernandes�LC,�
Nishiyama�A.�Platelet�activating�factor�and�evidence�of�oxidative�stress�in�the� bronchoalveolar� fluid� of� Thoroughbred� colts� during� race� training.� J�Vet�Intern�Med�2010;24(2):414�419.�
Mills�PC,�Smith�NC,�Casas�I,�Harris�P,�Harris�RC,�Marlin�DJ.�Effects�of�exercise�intensity�and�environmental�stress�on�indices�of�oxidative�stress�and�iron�homeostasis� during� exercise� in� the� horse.� Eur� J� Appl� Physiol� Occup�Physiol�1996;74(1�2):60�66.�
Moini� H,� Packer� L,� Saris� NE.� Antioxidant� and� prooxidant� activities� of� alpha�lipoic�acid�and�dihydrolipoic�acid.�Toxicol�Appl�Pharmacol�2002;182(1):84�90.�
Moran� M,� Delgado� J,� Gonzalez� B,� Manso� R,� Megias� A.� Responses� of� rat�myocardial� antioxidant� defences� and� heat� shock� protein� HSP72� induced�by�12�and�24�week�treadmill�training.�Acta�Physiol�Scand�2004;180(2):157�166.�

64
Morillas�Ruiz� J,�Zafrilla�P,�Almar�M,�Cuevas�MJ,�Lopez�FJ,�Abellan�P,�Villegas�JA,� Gonzalez�Gallego� J.� The� effects� of� an� antioxidant�supplemented�beverage� on� exercise�induced� oxidative� stress:� results� from� a� placebo�controlled� double�blind� study� in� cyclists.� Eur� J� Appl� Physiol� 2005;95(5�6):543�549.�
Morimoto� RI,� Santoro�MG.� Stress�inducible� responses� and�heat� shock�proteins:�new� pharmacologic� targets� for� cytoprotection.� Nat� Biotechnol�1998;16(9):833�838.�
Morton�JP,�Holloway�K,�Woods�P,�Cable�NT,�Burniston�J,�Evans�L,�Kayani�AC,�McArdle�A.�Exercise� training�induced�gender�specific�heat�shock�protein�adaptations�in�human�skeletal�muscle.�Muscle�Nerve�2009a;39(2):230�233.�
Morton� JP,� Kayani� AC,� McArdle� A,� Drust� B.� The� exercise�induced� stress�response� of� skeletal� muscle,� with� specific� emphasis� on� humans.� Sports�Med�2009b;39(8):643�662.�
Motta�S,�Letellier�C,�Ropert�M,�Motta�C,�Thiebault�JJ.�Protecting�effect�of�vitamin�E� supplementation� on� submaximal� exercise�induced� oxidative� stress� in�sedentary� dogs� as� assessed� by� erythrocyte� membrane� fluidity� and�paraoxonase�1�activity.�Vet�J�2009;181(3):288�295.�
Møller�P,�Risom�L,�Lundby�C,�Mikkelsen�L,�Loft�S.�Hypoxia�and�oxidation�levels�of�DNA�and� lipids� in�humans�and�animal� experimental�models.� IUBMB�Life�2008;60(11):707�723.�
Nakamura�Y,�Feng�Q,�Kumagai�T,�Torikai�K,�Ohigashi�H,�Osawa�T,�Noguchi�N,�Niki� E,� Uchida� K.� Ebselen,� a� glutathione� peroxidase� mimetic� seleno�organic� compound,� as� a� multifunctional� antioxidant.� Implication� for�inflammation�associated� carcinogenesis.� J� Biol� Chem� 2002;277(4):2687�2694.�
Nakhostin�Roohi� B,� Babaei� P,� Rahmani�Nia� F,� Bohlooli� S.� Effect� of� vitamin� C�supplementation�on�lipid�peroxidation,�muscle�damage�and�inflammation�after� 30�min� exercise� at� 75%� VO2max.� J� Sports� Med� Phys� Fitness�2008;48(2):217�224.�
Niess�AM,�Simon�P.�Response�and�adaptation�of�skeletal�muscle�to�exercise��the�role�of�reactive�oxygen�species.�Front�Biosci�2007;12:4826�4838.�
Niki�E,�Yoshida�Y.�Biomarkers�for�oxidative�stress:�measurement,�validation,�and�application.�J�Med�Invest�2005;52�Suppl:228�230.�
Noble� EG,� Milne� KJ,� Melling� CW.� Heat� shock� proteins� and� exercise:� a� primer.�Appl�Physiol�Nutr�Metab�2008;33(5):1050�1065.�
Nordberg�J,�Arner�ES.�Reactive�oxygen�species,�antioxidants,�and�the�mammalian�thioredoxin�system.�Free�Radic�Biol�Med�2001;31(11):1287�1312.�
Núñez�Córdoba� JM,� Martínez�González� MA.� Antioxidant� vitamins� and�cardiovascular�disease.�Curr�Top�Med�Chem�2011;11(14):1861�1869.�

65
Nyström� T.� Role� of� oxidative� carbonylation� in� protein� quality� control� and�senescence.�Embo�J�2005;24(7):1311�1317.�
Ogata� T,� Oishi� Y,� Higashida� K,� Higuchi� M,� Muraoka� I.� Prolonged� exercise�training� induces� long�term� enhancement� of� HSP70� expression� in� rat�plantaris� muscle.� Am� J� Physiol� Regul� Integr� Comp� Physiol�2009;296(5):R1557�1563.�
Ogawa�K,�Seta�R,�Shimizu�T,�Shinkai�S,�Calderwood�SK,�Nakazato�K,�Takahashi�K.� Plasma� adenosine� triphosphate� and� heat� shock� protein� 72�concentrations� after� aerobic� and� eccentric� exercise.� Exerc� Immunol� Rev�2011;17:136�149.�
Oksala�NK,�Laaksonen�DE,�Lappalainen�J,�Khanna�S,�Nakao�C,�Hanninen�O,�Sen�CK,� Atalay� M.� Heat� shock� protein� 60� response� to� exercise� in� diabetes:�effects� of� alpha�lipoic� acid� supplementation.� Journal� of� diabetes� and� its�complications�2006;20(4):257�261.�
Oksala� NK,� Lappalainen� J,� Laaksonen� DE,� Khanna� S,� Kaarniranta� K,� Sen� CK,�Atalay�M.�Alpha�lipoic�Acid�modulates�heat�shock�factor�1�expression�in�streptozotocin�induced� diabetic� rat� kidney.� Antioxidants� &� redox�signaling�2007;9(4):497�506.�
Ono� K,� Inui� K,� Hasegawa� T,� Matsuki� N,� Watanabe� H,� Takagi� S,� Hasegawa� A,�Tomoda� I.� The� changes� of� antioxidative� enzyme� activities� in� equine�erythrocytes� following� exercise.� Nippon� Juigaku� Zasshi� 1990;52(4):759�765.�
Packer�JE,�Slater�TF,�Willson�RL.�Direct�observation�of�a�free�radical� interaction�between�vitamin�E�and�vitamin�C.�Nature�1979;278(5706):737�738.�
Packer�L,�Tritschler�HJ,�Wessel�K.�Neuroprotection�by�the�metabolic�antioxidant�alpha�lipoic�acid.�Free�Radic�Biol�Med�1997;22(1�2):359�378.�
Packer� L,� Witt� EH,� Tritschler� HJ.� alpha�Lipoic� acid� as� a� biological� antioxidant.�Free�Radic�Biol�Med�1995;19(2):227�250.�
Paulsen�G,�Vissing�K,�Kalhovde�JM,�Ugelstad�I,�Bayer�ML,�Kadi�F,�Schjerling�P,�Hallen� J,� Raastad� T.� Maximal� eccentric� exercise� induces� a� rapid�accumulation� of� small� heat� shock� proteins� on� myofibrils� and� a� delayed�HSP70� response� in� humans.� Am� J� Physiol� Regul� Integr� Comp� Physiol�2007;293(2):R844�853.�
Pellegrini�N,�Serafini�M,�Colombi�B,�Del�Rio�D,�Salvatore�S,�Bianchi�M,�Brighenti�F.�Total�antioxidant�capacity�of�plant�foods,�beverages�and�oils�consumed�in�Italy�assessed�by�three�different�in�vitro�assays.�J�Nutr�2003;133(9):2812�2819.�
Persson� SGB.� Evaluation� of� exercise� tolerance� and� fitness� in� the� performance�horse.� In:� DH� Snow,� SGB� Persson,� JR� Rose� (eds.)� Equine� Exercise�Physiology.�Cambridge.�Burlington�Press.�United�Kingdom.��

66
Petersen� Shay� K,� Moreau� RF,� Smith� EJ,� Hagen� TM.� Is� alpha�lipoic� acid� a�scavenger�of�reactive�oxygen�species�in�vivo?�Evidence�for�its�initiation�of�stress�signaling�pathways�that�promote�endogenous�antioxidant�capacity.�IUBMB�Life�2008;60(6):362�367.�
Pizza�FX,�Peterson�JM,�Baas�JH,�Koh�TJ.�Neutrophils�contribute�to�muscle�injury�and�impair�its�resolution�after�lengthening�contractions�in�mice.�J�Physiol�2005;562(Pt�3):899�913.�
Poli� G,� Biasi� F,� Leonarduzzi� G.� 4�Hydroxynonenal�protein� adducts:� A� reliable�biomarker�of�lipid�oxidation�in�liver�diseases.�Mol�Aspects�Med�2008;29(1�2):67�71.�
Portier� K,� de� Moffarts� B,� Fellman� N,� Kirschvink� N,� Motta� C,� Letellierw� C,�Ruelland�A,�van�Erck�E,�Lekeux�P,�Couder�J.�The�effects�of�dietary�N�3�and�antioxidant� supplementation� on� erythrocyte� membrane� fatty� acid�composition� and� fluidity� in� exercising� horses.� Equine� Vet� J� Suppl�2006(36):279�284.�
Powers� SK,� Duarte� J,� Kavazis� AN,� Talbert� EE.� Reactive� oxygen� species� are�signalling� molecules� for� skeletal� muscle� adaptation.� Exp� Physiol�2010;95(1):1�9.�
Powers� SK,� Hamilton� K.� Antioxidants� and� exercise.� Clin� Sports� Med�1999;18(3):525�536.�
Powers�SK,�Lennon�SL.�Analysis�of�cellular� responses� to� free�radicals:� focus�on�exercise�and�skeletal�muscle.�Proc�Nutr�Soc�1999;58(4):1025�1033.�
Powers�SK,�Quindry�JC,�Kavazis�AN.�Exercise�induced�cardioprotection�against�myocardial� ischemia�reperfusion� injury.� Free� Radic� Biol� Med�2008;44(2):193�201.�
Prior� RL,� Cao� G.� In� vivo� total� antioxidant� capacity:� comparison� of� different�analytical�methods.�Free�Radic�Biol�Med�1999;27(11�12):1173�1181.�
Prior�RL,�Hoang�H,�Gu�L,�Wu�X,�Bacchiocca�M,�Howard�L,�Hampsch�Woodill�M,�Huang�D,�Ou�B,�Jacob�R.�Assays�for�hydrophilic�and�lipophilic�antioxidant�capacity�(oxygen�radical�absorbance�capacity�(ORAC(FL)))�of�plasma�and�other� biological� and� food� samples.� J� Agric� Food� Chem� 2003;51(11):3273�3279.�
Prior� RL,� Wu� X,� Schaich� K.� Standardized� methods� for� the� determination� of�antioxidant� capacity� and� phenolics� in� foods� and� dietary� supplements.� J�Agric�Food�Chem�2005;53(10):4290�4302.�
Pösö�AR,�Eklund�Uusitalo�S,�Hyyppä�S,�Pirilä�E.�Induction�of�heat�shock�protein�72�mRNA�in�skeletal�muscle�by�exercise�and�training.�Equine�Vet�J�Suppl�2002(34):214�218.�
Quiles� JL,� Ramirez�Tortosa� MC,� Huertas� JR,� Ibanez� S,� Gomez� JA,� Battino� M,�Mataix� J.� Olive� oil� supplemented� with� vitamin� E� affects� mitochondrial�

67
coenzyme� Q� levels� in� liver� of� rats� after� an� oxidative� stress� induced� by�adriamycin.�Biofactors�1999;9(2�4):331�336.�
Radak�Z,�Chung�HY,�Goto�S.�Systemic�adaptation�to�oxidative�challenge�induced�by�regular�exercise.�Free�Radic�Biol�Med�2008;44(2):153�159.�
Radak� Z,� Sasvari� M,� Nyakas� C,� Taylor� AW,� Ohno� H,� Nakamoto� H,� Goto� S.�Regular� training� modulates� the� accumulation� of� reactive� carbonyl�derivatives�in�mitochondrial�and�cytosolic�fractions�of�rat�skeletal�muscle.�Arch�Biochem�Biophys�2000;383(1):114�118.�
Reid�MB.�Free�radicals�and�muscle�fatigue:�Of�ROS,�canaries,�and�the�IOC.�Free�Radic�Biol�Med�2008;44(2):169�179.�
Reid� MB,� Haack� KE,� Franchek� KM,� Valberg� PA,� Kobzik� L,� West� MS.� Reactive�oxygen� in� skeletal�muscle.� I.� Intracellular�oxidant�kinetics� and� fatigue� in�vitro.�J�Appl�Physiol�1992a;73(5):1797�1804.�
Reid�MB,�Shoji�T,�Moody�MR,�Entman�ML.�Reactive�oxygen�in�skeletal�muscle.�II.� Extracellular� release� of� free� radicals.� J� Appl� Physiol� 1992b;73(5):1805�1809.�
Reznick� AZ,� Witt� E,� Matsumoto� M,� Packer� L.� Vitamin� E� inhibits� protein�oxidation� in� skeletal� muscle� of� resting� and� exercised� rats.� Biochem�Biophys�Res�Commun�1992a;189(2):801�806.�
Richardson�SM,�Siciliano�PD,�Engle�TE,�Larson�CK,�Ward�TL.�Effect�of�selenium�supplementation�and�source�on�the�selenium�status�of�horses.�J�Anim�Sci�2006;84(7):1742�1748.�
Rodrigues�JA,�Bull�ML,�Dias�GA,�Goncalves�M,�Guazzelli�JF.�Electromyographic�analysis� of� the� pectoralis� major� and� deltoideus� anterior� muscles� in�horizontal� flyer� exercises� with� loads.� Electromyogr� Clin� Neurophysiol�2003;43(7):413�419.�
Rose�RJ,�Hodgson�DR.�An�Overview�of�Performance�and�Sports�Medicine.�In:�DR�Hodgson,�JR�Rose�(eds.)�The�Athletic�Horse.�Philadelphia.�W.B.�Saunders�Company.�p.�497,�pp.�3�12.�
Rose�RJ,�Hodgson�DR.�Hematology�and�Biochemistry.�In:�DR�Hodgson,�JR�Rose�(eds.)�The�Athletic�Horse.�Philadelphia.�W.B.�Saunders�Company.�p.�497,�pp.�63�78.�
Rose�RJ,�Hodgson�DR,�Sampson�D,�Chan�W.�Changes�in�plasma�biochemistry�in�horses�competing� in�a�160�km�endurance�ride.�Aust�Vet� J�1983;60(4):101�105.�
Räsänen�LA,�Wiitanen�PA,�Lilius�EM,�Hyyppä�S,�Pösö�AR.�Accumulation�of�uric�acid� in� plasma� after� repeated� bouts� of� exercise� in� the� horse.� Comp�Biochem�Physiol�B�Biochem�Mol�Biol�1996;114(2):139�144.�

68
Saastamoinen�M,�Juusela�J.�Serum�vitamin�E�concentration�of�horses�on�different�vitamin� E� supplementation� levels.� Acta� Agriculturae� Scandinavica�1993;43(1):52�56,�56.�
Sachdev� S,� Davies� KJ.� Production,� detection,� and� adaptive� responses� to� free�radicals�in�exercise.�Free�Radic�Biol�Med�2008;44(2):215�223.�
Sacheck�JM,�Milbury�PE,�Cannon�JG,�Roubenoff�R,�Blumberg�JB.�Effect�of�vitamin�E� and� eccentric� exercise� on� selected� biomarkers� of� oxidative� stress� in�young�and�elderly�men.�Free�Radic�Biol�Med�2003;34(12):1575�1588.�
Saengsirisuwan�V,�Kinnick�TR,�Schmit�MB,�Henriksen�EJ.�Interactions�of�exercise�and�lipoic�acid�on�skeletal�muscle�glucose�transport�in�obese�Zucker�rats.�J�Appl�Physiol�2001;91:145�153.�
Saengsirisuwan� V,� Perez� FR,� Kinnick� TR,� Henriksen� EJ.� Effects� of� exercise�training�and�antioxidant�R�ALA�on�glucose� transport� in� insulin�sensitive�rat�skeletal�muscle.�J�Appl�Physiol�2002;92(1):50�58.�
Salo� DC,� Donovan� CM,� Davies� KJ.� HSP70� and� other� possible� heat� shock� or�oxidative� stress� proteins� are� induced� in� skeletal� muscle,� heart,� and� liver�during�exercise.�Free�Radic�Biol�Med�1991;11(3):239�246.�
Sarsour�EH,�Kumar�MG,�Chaudhuri�L,�Kalen�AL,�Goswami�PC.�Redox�control�of�the� cell� cycle� in� health� and� disease.� Antioxidants� &� redox� signaling�2009;11(12):2985�3011.�
Satoh�S,�Shindoh�M,�Min�JZ,�Toyooka�T,�Fukushima�T,�Inagaki�S.�Selective�and�sensitive� determination� of� lipoyllysine� (protein�bound� alpha�lipoic� acid)�in�biological�specimens�by�high�performance�liquid�chromatography�with�fluorescence�detection.�Anal�Chim�Acta�2008;618(2):210�217.�
Savitha�S,�Sivarajan�K,�Haripriya�D,�Kokilavani�V,�Panneerselvam�C.�Efficacy�of�levo� carnitine� and� alpha� lipoic� acid� in� ameliorating� the� decline� in�mitochondrial�enzymes�during�aging.�Clin�Nutr�2005;24(5):794�800.�
Schafer�FQ,�Buettner�GR.�Redox�environment�of� the�cell�as�viewed�through�the�redox�state�of�the�glutathione�disulfide/glutathione�couple.�Free�Rad�Biol�Med�2001;30(11):1191�1212.�
Schewe�C,�Schewe�T,�Wendel�A.�Strong�inhibition�of�mammalian�lipoxygenases�by�the�antiinflammatory�seleno�organic�compound�ebselen�in�the�absence�of�glutathione.�Biochem�Pharmacol�1994;48(1):65�74.�
Schmidt�MC,�Askew�EW,�Roberts�DE,�Prior�RL,�Ensign�WY,�Jr.,�Hesslink�RE,�Jr.�Oxidative� stress� in� humans� training� in� a� cold,� moderate� altitude�environment� and� their� response� to� a� phytochemical� antioxidant�supplement.�Wilderness�Environ�Med�2002;13(2):94�105.�
Schreier�P.�Chemopreventive�compounds�in�the�diet.�Dev�Ophthalmol�2005;38:1�58.�

69
Schumann�G,�Bonora�R,�Ceriotti�F,�Clerc�Renaud�P,�Ferrero�CA,�Ferard�G,�Franck�PF,� Gella� FJ,� Hoelzel� W,� Jorgensen� PJ,� Kanno� T,� Kessne� A,� Klauker� R,�Kristiansen� N,� Lessinger� JM,� Linsinger� TP,� Misaki� H,� Panteghini� M,�Pauwels� J,� Schimmel� HG,� Vialle� A,� Weidemann� G,� Siekmann� L.� IFCC�primary� reference� procedures� for� the� measurement� of� catalytic� activity�concentrations�of�enzymes�at�37�degrees�C.�Part�2.�Reference�procedure�for�the�measurement�of�catalytic�concentration�of�creatine�kinase.�Clin�Chem�Lab�Med�2002a;40(6):635�642.�
Schumann� G,� Bonora� R,� Ceriotti� F,� Ferard� G,� Ferrero� CA,� Franck� PF,� Gella� FJ,�Hoelzel�W,� Jorgensen�PJ,�Kanno�T,�Kessner�A,�Klauke�R,�Kristiansen�N,�Lessinger�JM,�Linsinger�TP,�Misaki�H,�Panteghini�M,�Pauwels�J,�Schiele�F,�Schimmel� HG,� Weidemann� G,� Siekmann� L.� IFCC� primary� reference�procedures� for� the� measurement� of� catalytic� activity� concentrations� of�enzymes� at� 37�degrees�C.� International� Federation�of� Clinical�Chemistry�and� Laboratory� Medicine.� Part� 5.� Reference� procedure� for� the�measurement� of� catalytic� concentration� of� aspartate� aminotransferase.�Clin�Chem�Lab�Med�2002b;40(7):725�733.�
Seifi�Skishahr�F,�Siahkohian�M,�Nakhostin�Roohi�B.�Influence�of�aerobic�exercise�at�high�and�moderate�intensities�on�lipid�peroxidation�in�untrained�men.�J�Sports�Med�Phys�Fitness�2008;48(4):515�521.�
Sen� CK,� Atalay� M,� Hanninen� O.� Exercise�induced� oxidative� stress:� glutathione�supplementation�and�deficiency.�J�Appl�Physiol�1994;77(5):2177�2187.�
Sen� CK,� Marin� E,� Kretzschmar� M,� Hanninen� O.� Skeletal� muscle� and� liver�glutathione� homeostasis� in� response� to� training,� exercise,� and�immobilization.�J�Appl�Physiol�1992;73(4):1265�1272.�
Sen�CK,�Packer�L.�Thiol�homeostasis�and�supplements�in�physical�exercise.�Am�J�Clin�Nutr�2000;72(2�Suppl):653S�669S.�
Sen� CK,� Rahkila� P,� Hanninen� O.� Glutathione� metabolism� in� skeletal� muscle�derived�cells�of�the�L6�line.�Acta�Physiol�Scand�1993;148(1):21�26.�
Sen� CK,� Roy� S,� Khanna� S,� Packer� L.� Determination� of� oxidized� and� reduced�lipoic� acid� using� high�performance� liquid� chromatography� and�coulometric�detection.�Methods�Enzymol�1999a;299:239�246.�
Sen�CK,�Tirosh�O,�Roy�S,�Kobayashi�MS,�Packer�L.�A�positively�charged�alpha�lipoic� acid� analogue� with� increased� cellular� uptake� and� more� potent�immunomodulatory� activity.� Biochem� Biophys� Res� Commun�1998;247(2):223�228.�
Sentürk�UK,�Yalcin�O,�Gunduz�F,�Kuru�O,�Meiselman�HJ,�Baskurt�OK.�Effect�of�antioxidant� vitamin� treatment� on� the� time� course� of� hematological� and�hemorheological� alterations� after� an� exhausting� exercise� episode� in�human�subjects.�J�Appl�Physiol�2005;98(4):1272�1279.�

70
Shepherd�D,�Garland�PB.�The�kinetic�properties�of�citrate�synthase�from�rat�liver�mitochondria.�Biochem�J�1969;114(3):597�610.�
Shringarpure� R,� Grune� T,� Davies� KJ.� Protein� oxidation� and� 20S� proteasome�dependent� proteolysis� in� mammalian� cells.� Cell� Mol� Life� Sci�2001;58(10):1442�1450.�
Shringarpure� R,� Grune� T,� Mehlhase� J,� Davies� KJ.� Ubiquitin� conjugation� is� not�required� for� the� degradation� of� oxidized� proteins� by� proteasome.� J� Biol�Chem�2003;278(1):311�318.�
Siciliano� PD,� Parker� AL,� Lawrence� LM.� Effect� of� dietary� vitamin� E�supplementation�on�the�integrity�of�skeletal�muscle�in�exercised�horses.�J�Anim�Sci�1996;75:1553�1560.�
Sies�H.�Role�of�reactive�oxygen�species�in�biological�processes.�Klin�Wochenschr�1991;69(21�23):965�968.�
Sies� H.� Ebselen,� a� selenoorganic� compound� as� glutathione� peroxidase� mimic.�Free�Radic�Biol�Med�1993;14(3):313�323.�
Skarda�RT,�Muir�WW,�Milne�DW,�Gabel�AA.�Effects�of� training�on�resting�and�postexercise� ECG� in� standardbred� horses,� using� a� standardized� exercise�test.�Am�J�Vet�Res�1976;37(12):1485�1488.�
Skibska� B,� Jozefowicz�Okonkwo� G,� Goraca� A.� Protective� effects� of� early�administration� of� alpha�lipoic� acid� against� lipopolysaccharide�induced�plasma�lipid�peroxidation.�Pharmacol�Rep�2006;58(3):399�404.�
Snow�DH,�Valberg�SJ.�Muscle�Anatomy,�Physiology,�and�Adaptations�to�Exercise�and� Training.� In:� DR� Hodgson,� JR� Rose� (eds.)� The� Athletic� Horse.�Philadelphia.�W.B.�Saunders�Company.�p.�497,�pp.�145�180.�
Stadler�K,�Jenei�V,�Bölcsházy�G,�Somogyi�A,�Jakus�J.�Increased�nitric�oxide�levels�as� an� early� sign� of� premature� aging� in� diabetes.� Free� Rad� Biol� Med�2003;35(10):1240�1251.�
Stadtman�ER.�Protein�oxidation�in�aging�and�age�related�diseases.�Ann�N�Y�Acad�Sci�2001;928:22�38.�
Sun�Y,�Guo�H,�Yu�H,�Wang�X,�Wu�J,�Xue�Y.�Bioaccumulation�and�physiological�effects�of�tetrabromobisphenol�A�in�coontail�Ceratophyllum�demersum�L.�Chemosphere�2008;70:1787�1795.�
Suzuki� YJ,� Carini� M,� Butterfield� DA.� Protein� carbonylation.� Antioxidants� &�redox�signaling�2010;12(3):323�325.�
Sürmen�Gür� E,� Oztürk� E,� Gür� H,� Pündük� Z,� Tuncel� P.� Effect� of� vitamin� E�supplementation� on� post�exercise� plasma� lipid� peroxidation� and� blood�antioxidant� status� in� smokers:� with� special� reference� to�haemoconcentration� effect.� Eur� J� Appl� Physiol� Occup� Physiol�1999;79(6):472�478.�

71
Tanabe� K,� Masuda� K,� Hirayama� A,� Nagase� S,� Kono� I,� Kuno� S.� Effect� of�spontaneous� exercise� on� antioxidant� capacity� in� rat� muscles� determined�by�electron�spin�resonance.�Acta�Physiol�(Oxf)�2006;186(2):119�125.�
Tanskanen� M,� Atalay� M,� Uusitalo� A.� Altered� oxidative� stress� in� overtrained�athletes.�J�Sports�Sci�2010;28(3):309�317.�
Thomas�DP,�Fregin�GF.�Cardiorespiratory�and�metabolic�responses�to�treadmill�exercise�in�the�horse.�J�Appl�Physiol�1981;50(4):864�868.�
Thompson� HS,� Clarkson� PM,� Scordilis� SP.� The� repeated� bout� effect� and� heat�shock� proteins:� intramuscular� HSP27� and� HSP70� expression� following�two� bouts� of� eccentric� exercise� in� humans.� Acta� Physiol� Scand�2002;174(1):47�56.�
Tiidus�PM,�Enns�DL.�Point:Counterpoint:�Estrogen�and�sex�do/do�not� influence�post�exercise�indexes�of�muscle�damage,�inflammation,�and�repair.�J�Appl�Physiol�2009;106(3):1010�1012;�discussion�1014�1015,�1021.�
Toldy�A,�Stadler�K,�Sasvari�M,�Jakus�J,�Jung�KJ,�Chung�HY,�Berkes�I,�Nyakas�C,�Radak�Z.�The� effect� of� exercise� and�nettle� supplementation�on� oxidative�stress�markers�in�the�rat�brain.�Brain�Res�Bull�2005;65(6):487�493.�
Traverse� JH,�Nesmelov�YE,�Crampton�M,� Lindstrom� P,�Thomas�DD,�Bache�RJ.�Measurement�of�myocardial�free�radical�production�during�exercise�using�EPR� spectroscopy.� Am� J� Physiol� Heart� Circ� Physiol� 2006;290(6):H2453�2458.�
Tyler�CM,�Golland�LC,�Evans�DL,�Hodgson�DR,�Rose�RJ.�Changes�in�maximum�oxygen�uptake�during�prolonged�training,�overtraining,�and�detraining�in�horses.�J�Appl�Physiol�1996;81(5):2244�2249.�
Tyler� CM,� Golland� LC,� Evans� DL,� Hodgson� DR,� Rose� RJ.� Skeletal� muscle�adaptations�to�prolonged�training,�overtraining�and�detraining�in�horses.�Pflugers�Arch�1998;436(3):391�397.�
Valko�M,�Leibfritz�D,�Moncol�J,�Cronin�MT,�Mazur�M,�Telser�J.�Free�radicals�and�antioxidants� in� normal� physiological� functions� and� human� disease.� Int� J�Biochem�Cell�Biol�2007;39(1):44�84.�
Venditti�P,�Di�Meo�S.�Antioxidants,�tissue�damage,�and�endurance�in�trained�and�untrained�young�male�rats.�Arch�Biochem�Biophys�1996;331(1):63�68.�
White�A,�Estrada�M,�Walker�K,�Wisnia�P,�Filgueira�G,�Valdes�F,�Araneda�O,�Behn�C,� Martinez� R.� Role� of� exercise� and� ascorbate� on� plasma� antioxidant�capacity� in� thoroughbred� race� horses.� Comp� Biochem� Physiol� A� Mol�Integr�Physiol�2001;128(1):99�104.�
Whitham� M,� Fortes� MB.� Heat� shock� protein� 72:� release� and� biological�significance�during�exercise.�Front�Biosci�2008;13:1328�1339.�

72
Viljanen� K,� Kylli� P,� Hubbermann� EM,� Schwarz� K,� Heinonen� M.� Anthocyanin�antioxidant� activity� and� partition� behavior� in� whey� protein� emulsion.� J�Agric�Food�Chem�2005;53(6):2022�2027.�
Williams�CA,�Carlucci�SA.�Oral�vitamin�E�supplementation�on�oxidative�stress,�vitamin�and�antioxidant�status�in�intensely�exercised�horses.�Equine�Vet�J�Suppl�2006(36):617�621.�
Williams�CA,�Hoffman�RM,�Kronfeld�DS,�Hess�TM,�Saker�KE,�Harris�PA.�Lipoic�acid� as� an� antioxidant� in� mature� thoroughbred� geldings:� A� preliminary�study.�J�Nutr�2002;132:1628S�1631S.�
Williams� CA,� Kronfeldt� DS,� Hess� TM,� Saker� KE,� Waldron� JN,� Crandell� KM,�Hoffman� RM,� Harris� PA.� Antioxidant� supplementation� and� subsequent�oxidative� stress� of� horses� during� an� 80�km� endurance� race.� J� Anim� Sci�2004;82(2):588�594.�
Wong� CM,� Cheema� AK,� Zhang� L,� Suzuki� YJ.� Protein� carbonylation� as� a� novel�mechanism�in�redox�signaling.�Circ�Res�2008;102(3):310�318.�
Wong�CM,�Marcocci�L,�Liu�L,�Suzuki�YJ.�Cell�signaling�by�protein�carbonylation�and�decarbonylation.�Antioxidants�&�redox�signaling�2010;12(3):393�404.�
Wray�DW,�Uberoi�A,�Lawrenson�L,�Bailey�DM,�Richardson�RS.�Oral�antioxidants�and�cardiovascular�health�in�the�exercise�trained�and�untrained�elderly:�a�radically�different�outcome.�Clin�Sci�(Lond)�2009;116(5):433�441.�
Yamano�S,�Eto�D,�Sugiura�T,�Kai�M,�Hiraga�A,�Tokuriki�M,�Miyata�H.�Effect�of�growth�and�training�on�muscle�adaptation�in�Thoroughbred�horses.�Am�J�Vet�Res�2002;63(10):1408�1412.�
Yang�Y,�Sharma�R,�Sharma�A,�Awasthi�S,�Awasthi�YC.�Lipid�peroxidation�and�cell�cycle�signaling:�4�hydroxynonenal,�a�key�molecule�in�stress�mediated�signaling.�Acta�Biochim�Pol�2003;50(2):319�336.�
You� T,� Goldfarb� AH,� Bloomer� RJ,� Nguyen� L,� Sha� X,� McKenzie� MJ.� Oxidative�stress� response� in� normal� and� antioxidant� supplemented� rats� to� a�downhill�run:�changes�in�blood�and�skeletal�muscles.�Can�J�Appl�Physiol�2005;30(6):677�689.�
Yu� J,�Bao�E,�Yan� J,�Lei�L.�Expression�and� localization�of�Hsps� in� the�heart� and�blood� vessel� of� heat�stressed� broilers.� Cell� Stress� Chaperones�2008;13(3):327�335.�
Yu�J,�Tang�S,�Bao�E,�Zhang�M,�Hao�Q,�Yue�Z.�The�effect�of�transportation�on�the�expression� of� heat� shock� proteins� and� meat� quality� of� M.� longissimus�dorsi�in�pigs.�Meat�Sci�2009.�
Zamora�R,�Alaiz�M,�Hidalgo�FJ.�Modification�of�histidine�residues�by�4,5�epoxy�2�alkenals.�Chem�Res�Toxicol�1999;12(7):654�660.�
Zembron�Lacny�A,�Slowinska�Lisowska�M,�Szygula�Z,�Witkowski�K,�Szyszka�K.�The� comparison� of� antioxidant� and� hematological� properties� of� N�

73
acetylcysteine�and�alpha�lipoic�acid�in�physically�active�males.�Physiol�Res�2009;58(6):855�861.�
Zembron�Lacny� A,� Slowinska�Lisowska� M,� Ziemba� A.� Integration� of� the� thiol�redox� status� with� cytokine� response� to� physical� training� in� professional�basketball�players.�Physiol�Res�2010;59(2):239�245.�
Zembron�Lacny� A,� Szyszka� K,� Szygula� Z.� Effect� of� cysteine� derivatives�administration� in� healthy� men� exposed� to� intense� resistance� exercise� by�evaluation�of�pro�antioxidant�ratio.�J�Physiol�Sci�2007;57(6):343�348.�
Zhang�B,�Wang�Y,�Su�Y.�Peroxiredoxins,�a�novel� target� in� cancer� radiotherapy.�Cancer�Lett�2009;286(2):154�160.

������������

Publications of the University of Eastern Finland
Dissertations in Health Sciences
isbn 978-952-61-0507-9
Publications of the University of Eastern FinlandDissertations in Health Sciences
The relationship between strenuous
exercise and oxidative insults has
been implicated in muscle damage
and decreased muscle performance.
This study presents an overview
of oxidative stress-related events
and tissue protection mechanisms
in skeletal muscle after controlled
trials on a high-speed treadmill and
during the recovery in horses. The
antioxidant and tissue protective
effects of a thiol redox-modulator
α-lipoic acid are also studied.
dissertatio
ns | 070 | S
usa
nn
a Kin
nu
nen
| Oxidative S
tress in Skeletal M
uscle A
fter Acute E
xercise
Susanna Kinnunen
Oxidative Stress in Skeletal Muscle After Acute Exercise
Susanna Kinnunen
Oxidative Stress in Skeletal Muscle After Acute Exercise





























