Embed Size (px)
Citation preview

Variable Expressivity in Fundus Albipunctatus SHEILA MARGOLIS, MD,* IRWIN M. SIEGEL, PhD,* HARRIS RIPPS, PhDt
Abstract: A healthy, 14-year-old girl presented with nyctalopia, good vision, and multiple, irregular, yellowish lesions of the fundus. Dark adaptometry showed prolonged cone and rod branches, elevated thresholds, and the cone-rod transition occurring after 50 minutes in darkness. Her scotopic electroretinogram (ERG) b-wave attained normal amplitudes after 45 minutes of dark adaptation. The half-time for regeneration of rhodopsin after an extensive bleach was 16 minutes, four times longer than normal, and the maximum density difference measured by fundus reflectometry was at the lower limit of the normal range. Although photopigment kinetics were significantly faster than observed in other reported cases of fundus albipunctatus, it appears likely that there is a wide spectrum of functional and funduscopic abnormalities in this disorder. However, fundus appearance, adaptometric findings, and rhodopsin determinations serve to distinguish fundus albipunctatus from other flecked retina diseases. [Key words: dark adaptometry, electroretinography, fundus albipunctatus, genetic variability, night blindness, rhodopsin kinetics.] Ophthalmology 94:1416-1422, 1987
There are a variety of retinal disorders characterized by distinctive fundus changes that do not appear to bear an obvious relation to the functional abnormalities associated with the condition. 1•2 This is particularly evident in some of the stationary forms of congenital nightblindness.2·3 In fundus albipunctatus, for example, the fundus typically contains a large number of small, punctate, yellowish-white dots, organized into a broad annular band encircling the posterior pole. 1 The functional hallmark of the disorder is the markedly prolonged cone and rod branches of the dark adaptation curve, but there is apparently no correlation between these findings and the distribution of the punctate deposits. However, unlike other forms of hereditary nightblindness, there is a clear relation between the very
From the New York Eye and Ear Infirmary,* New York University Medical Center, New York, the Lions of Illinois Eye Research lnstitute,t University of Illinois College of Medicine, Chicago.
Supported by a grant from the RP Foundation and Allied Diseases to the New York University Retinal Clinic, a research grant (EY 06516) from the National Eye Institute, unrestricted grants to the Departments of Ophthalmology at New York University Medical Center and University of Illinois Eye and Ear Infirmary from Research to Prevent Blindness, and an unrestricted grant to the Kirby Eye Institute.
Reprint requests to Sheila Margolis, MD, 16 Park Avenue, New York, NY 10016.
gradual return of sensitivity in the dark and an abnormally slow regeneration rate of the visual pigments. 2
Electrical activity, such as the electroretinogram (ERG) amplitudes and the electro-oculogram light:dark ratio, can also be shown to follow the slow return of visual pigment, a process that may require 3 hours or more in darkness after an intense bleaching exposure. 2
The patient we report here has extensive fundus lesions which appear to involve more of the retina than is usually the case in fundus albipunctatus. Nevertheless, her functional abnormalities, particularly with regard to the rod system are less severe, and she manifests a retinal dysfunction which seems to be intermediate between the typical form of fundus albipunctatus and the normal subject. This circumstance presents an opportunity to add relevant data to the presumed relationship between photopigment kinetics and the time course of dark adaptation.4 In addition, the ophthalmoscopic and functional findings exhibited by this patient pose an interesting question for the clinician: are we dealing with a unique genetic entity or simply with a variant of fundus albipunctatus?
PATIENT
A 14-year-old Puerto Rican girl was seen for a routine eye examination and complained about poor vision at
1416

MARGOLIS et al • FUNDUS ALBIPUNCTATUS
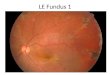
Fig 1. Photomontage of the right eye of the patient showing distribution of yellowish lesions.
night. She was in good health and there was no history of parental consanguinity or ocular disease in her family. Both parents, her maternal grandmother, a maternal uncle, and one first cousin were given eye examinations and were found to be normal.
METHODS
In addition to a complete ophthalmologic examination, color vision was evaluated with the AO-HRR test plates (American Optical Co, Southbridge, MA) and the Farnsworth D-15 panel (Psychological Corp, New York, NY), and visual fields were measured on a Goldmann perimeter with a 0.25-mm white test object under standard conditions of background luminance.
Fundus photographs including fluorescein angiograms were obtained at the time of the initial examination.
The course of dark adaptation was determined monocularly with a modified Goldmann-Weekers adaptometer; all runs were made after pupil dilation with tropicamide 1% (Mydriacyl). The retinal area tested was located in the horizontal meridian, 15 o temporal to the fovea. However, stimulus size and preadapting exposure conditions varied in accordance with the objectives of the experiment and are given in the description of the results. After at least 90 minutes ofdark adaptation, rod thresholds were measured on the same apparatus for retinal loci extending 30° temporal and 30° nasal to the fovea. Data were obtained for 3° white stimulus, using a test flash duration of 0. 7 second.
Electroretinography was performed using a Life-Tech
Model 100 recorder (Life-Tech, Houston, TX) with the amplifiers set at a bandwidth of 1 to 300 Hz. The active lead was a Burian-Allen monopolar contact lens electrode (Hansen Ophthalmic Co, Iowa City, lA), and the forehead and ear served as reference and ground, respectively. A Ganzfeld bowl provided diffuse flash stimuli. Photopic (cone-mediated) potentials were recorded under conditions of moderate (50 foot-candles) room illumination; scotopic (rod mediated) ERGs were obtained after 15 and 45 minutes of dark adaptation.
Fundus reflectometry was used to determine the bleaching and regeneration kinetics of rhodopsin within the same test region studied by dark adaptometry (i.e., 15° temporal to the fovea); a detailed description ofthe rapid scan apparatus has been published elsewhere.5 In brief, spectral scans (400-600 nm) were obtained (1) with the eye fully dark adapted for more than 1.5 hours, (2) after a 30-second exposure to an intense yellow light (Wratten # 15, 6.2 log scot td) that bleached about 99% of the available rhodopsin, and (3) at various times after the bleaching light was extinguished. Difference spectra computed from these measurements were used in determining the density loss due to the bleaching exposure (1, 2), and the kinetics of regeneration (2, 3) throughout more than 90 minutes of dark adaptation.
RESULTS
OCULAR FINDINGS
At the time of the initial examination, cycloplegic refraction was: right eye, +1.00 = -2.00 X 150; left eye,
1417

OPHTHALMOLOGY • NOVEMBER 1987 • VOLUME 94 • NUMBER 11
Fig 2. Lesions in the midperipheral retina of the right eye showing the manner in which they aggregate and become confluent (original magnification, X2).
-0.25 = -1.00 X 5; and visual acuity correctable to 20/25 in each eye. Four years later, her acuity was 20/20 in both eyes with slight changes in the astigmatic error. Except as noted below, all other phases of the examination were normal, including intraocular pressure, slitlamp study of the anterior segment, lens and vitreous, color vision, and visual fields.
Results ofophthalmoscopy showed normal-appearing optic discs, normal retinal vasculature, and well-defined foveal reflexes. However, beginning in the perimacular region and extending to the periphery, there were great numbers of yellowish deposits which varied in size and shape lying deep to the retinal vessels (probably at the level of the retinal pigment epithelium [RPE]). The fundus photomontage (Fig 1) shows the distribution of the lesions from the posterior pole to the midperiphery. The shapes ranged from dots, rods, and the comma-like flecks seen centrally, to more amorphous patterns in the far periphery where the flecks appeared to clump together and become confluent (Fig 2). The fundus appearance around the lesions was normal; there was no sign of "tapetal sheen," or changes in pigmentation. Subsequent fundus examinations performed 2 and 4 years after the photographs were taken showed no detectable changes.
Fundus fluorescein angiograms (Fig 3) showed transmission defects in the RPE scattered throughout the fundus. The location of the RPE defects were independent of the distribution of the yellow flecks,6 and included areas within the macula that were free of flecks. In addition, there was no sign of the so-called "dark choroid," a hypofluorescent phenomenon associated with large amounts oflipofuscin-like material located in the RPE; these deposits have been shown to occlude fluorescence from the choriocapillaris in diseases as diverse as fundus flavimaculatus7 and cone-rod dystrophy.8 However, close inspection of the fluorescein
Fig 3. Fundus fluorescein angiograms of the patient show that the lesions in various portions of the retina are not coincident with the small atrophic areas of the RPE which hyperfluoresce. Top, disc, right eye. Bottom, infero-nasal to disc.
photographs indicated that the individual flecks did block choroidal flush, and gave the early angiographic photographs a more patchy appearance than usual.
Figure 4 compares the course of dark adaptation for our patient with that obtained on normal subjects (dashed line); the data were obtained after similar preadapting bleaches of 3.11 log foot-lamberts (7.8 log td sec). Clearly, the two segments of the patient's dark adaptation curve are grossly abnormal; for both the coneand rod-mediated branches, the rates ofdark adaptation are slower and the threshold plateaus are higher than normal. Notice, for example, that the normal cone plateau is reached in about 8 minutes, the cone-rod transition occurs at 12 minutes, and that the rod thresholds are attained after about 25 minutes of dark adaptation. The patient data, on the other hand, show that the early cone-mediated dark adaptation process is much slower,
1418

--
MARGOLIS et al • FUNDUS ALBIPUNCT A TUS
Fig 4. Course of dark adaptation of the patient after an intense preadapting exposure. The 7-minute exposure to 3.11 log foot-lamberts (pupil diameter, 7 mm) provided a retinal illuminance of 7.8 log troland seconds.
6
~5 1/)
c:.e4 c:
Fig 5. Decreasing the preadapting luminance (to 7.2 Q)
log troland seconds) and > 3 using a larger test stimulus +
0 reduces the cone-rod break Q) 2time and allows each branch a:: ofthe dark adaptation curve to attain maximal sensitiv 0' ity in a shorter period of 0 time. _j
0
5 ?'pre-adapt to 3.11 log ft.L.
Vl
c 4 •
\ stimulus= 1.25~white Q)
c \
\ located l5°temporal retina
Q) >
3 ', --,
\ ~ 2 \
\ Q) \
'cr
] 1Eo~[--~~--~'~',_,__
.........~~--~~--~--~~--~~--~~--~~--~___ I I I I I I I I
10 20 30
the cone-rod break is at 50 minutes, and it takes more than 75 minutes for the absolute rod threshold to be reached. In addition, it should be noted that the patient could not make threshold judgments during the first few minutes of dark adaptation because of dazzle from the preadapting field, and that the rod and cone plateaus are raised 0.4 and 0.6 log units, respectively, above the corresponding levels for normal subjects. Evidence that the loss of rod sensitivity was a widespread phenomenon was seen in the retinal profile data (not shown). Although there was some variability across the horizontal meridian, threshold was elevated throughout by about 0.6 to 1.2 log units above the normal.
Dark adaptometry was performed also under less-demanding conditions of preadaptation and testing. In an attempt to reduce the dazzle effect produced by the intense bleaching exposure, and to replicate more closely the experimental conditions used in an earlier study,9
the test was repeated with a dimmer ( -0.6 log unit)
40 50 60 70 80 90 100 M1nutes in Dark
preadapting light and a larger (5.5 deg) test stimulus. As shown in Figure 5, both the normal function and that of our patient were affected by the change in test conditions, particularly as regards the time of the cone-rod transition. In both cases, the break occurred in about one-half the time seen in Figure 4 (i.e., after 5 minutes of dark adaptation in the normal and after about 22 minutes for the patient). It is apparent, therefore, that preadapting conditions exert a profound effect on psychophysical dark adaptation, and must be carefully considered when comparing results from different studies.
Figure 6 shows several photopic and scotopic ERG tracings recorded when the patient was 14 years old. The cone-mediated responses to a single flash or to 30/second high intensity flashes appeared to be lower than normal. However, it was observed that during flicker stimulation this young patient became agitated and would attempt to close her eyes. The cone implicit time (flash-to-b-wave peak) was normal. After exposure to
7' pre-adapt to 2.51og ft.L. stimulus= 5.5~ white
•.. I o c a ted 15o temporo I retina•......................-......\ ..... ,' ......\ ....\
\ ............... NORMAL
10 20 30 40 50 60 Minutes in Dark
70
1419

OPHTHALMOLOGY • NOVEMBER 1987 • VOLUME 94 • NUMBER 11
the bright flicker stimulus, the patient remained in darkness and responses were recorded after 15 and 45 minutes of dark adaptation. As shown by the lower tracings of Figure 6, normal subjects recover full b-wave amplitudes in 15 minutes, whereas the patient's ERG b-wave had grown to only half that amplitude. Notice that the amplitude reduction applies both to the a- and b-waves, indicating that the photoreceptors also require a longer time to recover from the prior light adaptation. At the end of45 minutes in the dark, however, all components ofthe ERG waveform have attained normal amplitudes.
Figure 7 A shows density difference spectra (the changes in absorbance) across the visible spectrum obtained by using fundus reflectometry at various times after exposure to an intense bleaching light. For the normal subject, rhodopsin regeneration requires about 25 minutes of darkness10; for our patient, the process takes nearly twice as long. Moreover, the maximum absorbance change for the patient (LlD (2) = 0.11) is at the lower limit of normal. 11 Notice that there is a small progressive shift in A.max from about 490 nm after 2 minutes in darkness to 510 nm after 30 minutes, a finding also reported for normal subjects and attributable to the formation and subsequent decay of an intermediate photoproduct, metarhodopsin III. 10
Figure 7B illustrates graphically the measured time course of rhodopsin regeneration. The data points were obtained from curves similar to those in Figure 7 A, but refer here to absorbance changes at a single wavelength (A. = 510 nm) relative to the maximal absorbance change for that wavelength (i.e., the percent rhodopsin regeneration as a function of time in the dark). The curve in Figure 7B shows that it takes about 16 minutes
Patient Normal
Photopic
~~d.a.
• Scotopic
J ~'M •
Fig 6. Electroretinogram recordings. The patient shows a nearly normal photopic b-wave amplitude, but after 15 minutes in the dark, a scotopic b-wave amplitude only one-half that of the normal subject. After 45 minutes in the dark, however, the b-wave amplitude increases to normal size. The vertical and horizontal calibration lines represent 200 11V and 25 msec, respectively.
• 4 s' 60 1 w .10 0
(.!) z < J: Subject' D N u
15°T w .05 u A z < "' "' 0 0 "' "' <
400 500 600 700 WAVELENGTH (nm)
100 • • ~so
~ • •
z 60 •;;; 0.. ------.
I 8~ 40 I
0 : t 1/2 =16'J:
I"'20 I
10 20 30 40 50 60 TIME IN DARK (min)
Fig 7. Fundus reflectometry. A, difference spectra recorded at times indicated after an intense bleaching exposure (7.7log troland seconds). The maximum absorbance change at the end of60 minutes is near the lower limit of normal. B, rhodopsin regeneration measured at "A = 51 0 nm; half-time of regeneration was 16 minutes.
to recover 50% ofthe total amount of rhodopsin, a value roughly four times slower than normal. 10
•12 Notice also
that nearly 1 hour is required to recover all the bleached rhodopsin, as compared to the normal value of 20 minutes.
DISCUSSION
Of all the forms of stationary nightblindness, fundus albipunctatus is unique, in that it has been possible to identify a specific retinal abnormality that accounts for functional deficits associated with the disease.2 Clearly, it is the extremely slow regeneration rate of the visual pigments that governs, to a large extent, the prolongation ofboth the rod and cone branches of the dark adaptation curve, as well as the slow recovery of the ERG . Since there are a number of complex biochemical processes involved in the resynthesis of visual pigments after bleaching (few ofwhich can be studied in vivo), the precise nature of the anomaly remains unknown. There is good evidence, however, that serum vitamin A levels are normal in fundus albipunctatus patients,6•13 and that the underlying defect probably affects a local (i.e., retinal) process involving the exchange of retinoids between the visual cells and the RPE.
Although in most circumstances the regeneration of human rhodopsin can be approximated by an exponen
1420

MARGOLIS et al • FUNDUS ALBIPUNCTATUS
Fig 8. The effect of stray light on the measurement of rhodopsin kinetics. The continuous line represents the absorbance data(~ DM) measured by fundus reflectometry in the course of dark adaptation (from Fig 7). The dashed line shows
100
80
/ /
/ /
/ / , ,, ....... -r----
the corresponding changes occurring in the rods(~ DR) as calculated from the equation relating to the two processes when f (the fractional area occupied by photoreceptor outer segments) is
c ., a. 0 ~ 0
40
-7(/
I /I I / IJ/
1 I
/ /
tY1 16'
21.5'
0.3 .20 Notice that in situ events occur more slowly than the reflectometric data suggest. As a result, the half-time of regeneration in situ occurs at 21.5 minutes, and the point at which 99% of rhodopsin is regenerated
...r::.
"' 20 /
// /
'/ V
//I / I
I I I I I
I I I I I I I
is extended from 38 to 49 minutes (arrows). 0 10 20 30 40 50 60 70
Time (min)
10 12 14tial function, - • first-order reaction kinetics do not measurements that, due to stray light, do not a priori describe the process in some of the other mammalian provide an accurate index of receptor photochemistry. species that have been tested. 15- 17 The departure from a In fact, a consideration of the stray light factor in fundus simple exponential is evident also in the reflectometric reflectometry19 enables us to derive an equation that data obtained in fundus albipunctatus2 (Fig 7). In fact, transforms the measured density differences (ADM) to both here and in the earlier study of fundus albipunc receptoral equivalents (ADR). Figure 8 illustrates the aptatus, the initial stage of the regenerative process appears plication of this equation to the experimental data of nearly linear in form, a finding that is more akin to the Figure 7B; the measured absorbance changes are plotted results obtained in animal studies than to the data on the along with the results derived from the equation that regeneration of human rhodopsin. relates these data to those occurring in situ. Notice that
In addition to this unusual photochemical anomaly, after compensating for stray light effects, the in situ there is an apparent discrepancy between the reflecto events occur more slowly than the reflectometric data metric data on rhodopsin regeneration and the subjec suggest. As a result, the half-time of regeneration is distive measurements of dark adaptation obtained under placed from 16.0 to 21.5 minutes, and the point at similar bleaching conditions (i.e., Figs 4 and 7B respec which 92% of the rhodopsin is regenerated (the pretively). Rushton and Powell 18 have shown that the sumed location of the cone-rod break) is extended to 49 cone-rod break of the normal dark adaptation curve minutes, a value clearly in line with the subjective reoccurs at a time when about 92% of the bleached rho sults cited earlier. dopsin has regenerated. Thus, the full extent of the We now return to a consideration of the disease entity measured range of rod dark adaptation, extending over manifested by this patient. Does she, by virtue of her more than 2.5 log units, corresponds to the resynthesis singular fundus appearance, the moderate prolongation of the final 8% of the available rhodopsin. Assuming of visual pigment regeneration, and dark adaptation, that a similar situation exists in patients with fundus represent a distinct type of fundus albipunctatus? The albipunctatus, a comparison of the results shown in Fig cases reported by Smith et al9 and by Krill and Folk21
ures 4 and 7 indicates a large temporal disparity; appear, in fact, to be very similar in fundus appearance whereas the cone-rod kink in the subjective function and functional findings to those ofthe patient. They also occurs at about 50 minutes, rhodopsin regeneration as noted that, like the patient in this study, the scotopic measured by fundus reflectometry is 92% complete in ERG b-wave amplitudes recovered slower than normal only 38 minutes. However, it is important to recall that after light exposure. On the other hand, there are at least subjective dark adaptation is governed primarily by the two reported cases22 with fundus lesions having the same rhodopsin changes occurring within the photoreceptors, general appearance and distribution as our patient, but whereas the data of Figure 7 are based on reflectometric who manifest the very prolonged dark adaptation (~ 3
1421

c
OPHTHALMOLOGY • NOVEMBER 1987 • VOLUME 94 • NUMBER 11
normal
.~100-0 I... Q) c 80 Q)
0'1 Q)
a:: 60 c en 40' a. 0
"'0 0 ~
a::
oguchi c.s.n.b. patient fundus albi punctatus
/ .. / ..........----~·············-··-·······················-·····
I I I I ...... I I ....·· ..I I ....········· I_ -----:.··- I .. • 50% recovery..
normal: ..•··•···••··•········•··••···••··•• patient:
fundus albi.:
Fig 9. Rhodopsin regeneration of the patient compared with congenital stationary nightblindness and Oguchi's disease (both of which are normal), and the more typical form of fundus albipunctatus (markedly4' prolonged).16'
60' .··
0 ~ ..··· o~~~~~~~~~~~~~~~~~~--
60 120 180 Time (minutes)
hours) seen usually in the more typical form of fundus albipunctatus (i.e., those having small discrete white dots arranged in an annulus around the disc and macula). To emphasize further the profound disparity between fundus changes and functional abnormalities, there is the descriptive study of Aish and Dajani23 who reported that seven siblings from the same family had a nearly identical fundus picture to that of the patient being studied, but showed no functional deficits. Unfortunately, this claim was not supported by any corroborative data.
With regard to the underlying defect in fundus albipunctatus, fundus reflectometry probably provides the most significant insight into the nature of the disorder. In the few cases studied, representing the extremes ofthe dark adaptation abnormality, there is a close correlation between the rates of subjective dark adaptation and the kinetics of photopigment regeneration. Thus, even at this level of observation, the results (Fig 9) demonstrate the variability that characterizes this form ofnightblindness.
REFERENCES
1. Franceschetti A, Chome-Bercioux N. Fundus albipunctatus cum hemeralopia. Ophthalmologica 1951; 121:185-93.
2. Carr RE, Ripps H, Siegel 1M. Visual pigment kinetics and adaptation in fundus albipunctatus. Doc Ophthalmol Proc Ser 1974; 4:193-204.
3. Carr RE, Ripps H. Rhodopsin kinetics and rod adaptation in Oguchi's disease. Invest Ophthalmol1967; 6:426-36.
4. Rushton WAH. Rhodopsin measurement and dark-adaptation in a subject deficient in cone vision. J Physiol1961; 156:193-205.
5. Carr RE, Ripps H, Siegel 1M, Weale RA. Rhodopsin and the electrical activity of the retina in congenital night blindness. Invest Ophthalmol 1966; 5:497-507.
6. Marmor MF. Fundus albipunctatus: a clinical study of the fundus
lesions, the physiologic deficit, and the vitamin A metabolism. Doc Ophthalmol1977; 43:277-302.
7. Eagle RC Jr, Lucier AC, Bernardino VB Jr, Yanoff M. Retinal pigment epithelial abnormalities in fundus flavimaculatus. Ophthalmology 1980; 87:1189-200.
8. Rabb MF, Tso MOM, Fishman G. Cone-rod dystrophy: a clinical and histopathologic report. Ophthalmology 1986; 93:1443-51.
9. Smith BF, Ripps H, Goodman GA. Retinitis punctata albescens: a functional and diagnostic evaluation. Arch Ophthalrnol 1959; 61 :93101.
10. Ripps H, Weale RA. Rhodopsin regeneration in man. Nature 1969; 222:775-7.
11. Ripps H, Brin KP, Weale RA. Rhodopsin and visual threshold in retinitis pigmentosa. Invest Ophthalrnol Vis Sci 1978; 17:735-45.
12. Alpern M. Rhodopsin kinetics in the human eye. J Physiol 1971; 217:447-71.
13. Carr RE, Margolis S, Siegel 1M. Fluorescein angiography and vitamin A and oxalate in fundus albipunctatus. Am J Ophthalmol 1976; 82:549-58.
14. Pugh EN. Rushton's paradox: rod dark adaptation after flash photolysis. J Physiol 1975; 248:413-31.
15. Weale RA. Bleaching experiments on eyes of living guinea-pigs. J Physiol1955; 127:572-86.
16. Perlman I. Kinetics of bleaching and regeneration of rhodopsin in abnormal (RCS) and normal albino rats in vivo. J Physiol 1978; 278:141-59.
17. Ripps H, Mehaffey L Ill, Siegel 1M. Rhodopsin kinetics in the cat retina. J Gen Physiol1981; 77:317-34.
18. Rushton WAH, Powell DS, The rhodopsin content and the visual threshold of human rods. Vis Res 1972; 12:1073-81.
19. Ripps H, Mehaffey L Ill, Siegel 1M, et al. Flash photolysis of rhodopsin in the cat retina. J Gen Physiol1981; 77:295-315.
20. 0sterberg G. Topography of the layer of rods and cones in the human retina. Acta Ophthalmol Suppl 1935; 6:6-102.
21. Krill AE, Folk MR. Retinitis punctata albescens: a functional evaluation of an unusual carp. Arn J Ophthalmol 1962; 53:450-5.
22. Miyake Y, Harada K. Familial fleck retina with nightblindness. Ann Ophthalmol1982; 14:836-41.
23. Sabei-Aish SF, Dajani B. Benign familial fleck retina. Br J Ophthalmol 1980; 64:652-9.
1422