Embed Size (px)
Citation preview

t~OHRLICtt, ADLER und Ka~)I~: Diff¢renzierung der Eiweil~stoffe des Weizens und Roggens. I. 85
Versuche zur chemischen Differenzierung der Eiweifistoffe des Weizens und Roggens.
I . Mi t t e i lung .
Untersuchungen der Eiwei~sto~fe mit Natriumhypochlorit. Von
M. R O H ~ C H ~ G. ADLE~ u n d 0 . K ~ M M .
Mitteilung aus der Versuehsanstalt liar Getreideverwertung, Berlin.
Mit 10 Textabbildungen.
(Eingegangen am 18. M~irz 1955.)
Ein/i~hrung.
Die Chemic der G e t r e i d e p r o t e i n e h a t in l e t z t e r Ze i t d u r c h e ine R e i h e w i c h t i g e r E r k e n n t n i s s e n e u e n A u f t r i e b e rha l t en . I n s b e s o n d e r e b e d i n g e n die A r b e i t e n y o n H E s s 1 d u r c h die B e o b a c h t u n g zweie r m e e h a n i s c h t r e n n b a r e r E i w e i l ~ k o m p o n e n t e n im W e i z e n m e h l u n d d u r c h die r S n t g e n o g r ~ p h i s c h e C h a r a k t e r i s i e r u n g der K l e b e r - b i l dung des n a t i v e n We izene iwe i~ eine R e v i s i o n der b i she r igen V o r s t e l l u n g e n f iber das Ge t r e idee iwe i~ u n d seine R o l l e bei de r Te igb i l dung .
Zweifel an der Charakterisierung des Getreideeiweil3 und an der Einheitlichkeit der isolierten Kleberkomponenten G]iadin und Glutenin bestehen ja seit langen. Obwohl diese beiden Eiwei•- kSrper nach den Befunden yon OSbOrnE 2 sich dutch ihren Aminos:~uregehalt unterscheiden sollen, hat schon O'BnI~N a angenonnen , dal~ sich eine Klcberkonponente aus der anderen her- leitet, und auch KOSUT~_~Y 4 hat das Gliadin als Hydrat des Glutenins und Glutenin als oxydiertes Anhydrid des Gliadins angesehen. Die Gfiltigkeit der OseoRN~schen Befunde ist abet his heute im Grunde genomnen noch nicht erschiittert worden.
Bedingt dutch die modernen physikalischen Arbeitsnethoden sind die Untersuchungen tiber die Uneinheitlichkeit des ~¥eizenklebers oder Zuf~illigkeit der getroffenen Feststellungen wieder in Flul~ gekonmen. So haben KREJCI und SVEDBE~G ~ bei Arbeiten n i t der Ultrazentrifuge in einer GliaclinlSsung eine Mischung yon ,,Ganz- und Halbnolekfilen" nachweisen kSnnen. Die ,,Ganzmolekfile" zeigen ein Molekulargewicht yon 34500, die ,,Halbmolekfile" yon 17250, und die ersten dissoziieren mit steigender Tempera~ur und Acidit~t in die letzteren. Die Inhonogenit~t des Gliadins durch Messung unterschiedlichcr ])iffusionskonstanten mehrerer Fraktionen ist auch yon LAMM und POLSO~ 6 besti~tigt worden, die nur die neistlSsliehe Fraktion als honogen bezeichnen. Bum~ 7 ist auf Grund seiner Molekulargewichtsbestimmungen ni t te ls osmotischer Drucknessung in verschiedenen organischen LSsungsmittein zu d e n gleichen Schlul~ gekonmen. Seine ffir Gliadin erni t te l ten Molekulargewichte variieren zwischen 40000 und 75000 je nach L5sungsmittcl.
SC~IWERT, PUTNAM und BRIGGS s haben mittels der elektrophoretischen Methode yon TISELIUS gefunden, dab Gliadin auch elektrophoretisch inhomogen ist. Das aach Extraktion des Klebers mit w~t~rigem Alkohol - - zur Entfernung des Gliadins - - zurfickbleibende Glutenin, das nach OSBOR~ nur in verdfinnten Alkali 15slich is~, muB nach BLISH und SA~DST~DT 9 gleichfalls als uneinheitlich und als ver~ndert angesehen werden.
Die Molekulareigenschaften des Klebers sind yon McCALL~ und GR~L]~ ~° in Nat r iun- salicylatlSsung durch Anwendung der Ultrazentrifuge und Diffusionsmethode untersucht worden.
HEss, K. : Kolloid-Z. 136, 84 (1954). OSBORNE, T. B. : The proteins of the wheat kernel. Carnegie Inst., Pub]. No. 84. Washington:
1907. O ' B R I ~ , M.: Ann. Bot. 9, 543 (1895).
a KOSUT~:f, T.: J. Landw. 51, 139 (1903). KRE5C~, L., u. T. SVED~EI~G: J. Amer. Chem. Soc. 57, 946 (1935). L A ~ , 0., u. A. POLSO~: Biochemic. J. 30, 528 (1936).
: BU~K, ~N. F.: J . of Biol. Chem. 124, 49 (1938). 8 SC/-IWERT, G. W., U. W. PUTNA~¢~ 11. D. ]:~. BRIGGS; Arch. Biochemistry 4, 371 (1944). 9 BLISS-I, M. J., u. ~. M. SANDSTEDT: J. of Biol. Chem. 85, 195 (1929).
~0 MeC~LL~, A. G., u. N. G n ~ L ~ : Canad. J. Res., Sect. C 20, 130 (1942).

86 M. R o m ~ c H , G. ADLER und O. K ~ A ~ :
Sic lassen vermuten, dab der Kleber ein aus sehr vielen Komponenten bestehendes Protein- system ist. Diese Komponenten variieren in fortsehreitendem MaBe hinsichtlich L6slichkeit und anderer physikaliseher Eigensehaften. Aus diesem Grunde ist es nach MCCALLA unkorrekt, den Kleber als aus nur zwei oder drei oder vier einzelnen Komponenten zusammengesetzt zu be- zeiehnen. Das yon ihm bestimmte niedrigste Molekulargewieht der am meisten 15slichen Fraktion hat 35000 betragen.
KVaL~ANN 1 nimmt an, dab der Kleber einen Eiweil3komplex darstellt, der aus Micellen versehiedener L~nge gebildet wird und deren l~ngste und stabilste die des Glutenins ist. McCALLA und Rose 2 haben bei fraktionierten F~llungen aus Kleberdispersionen in 8--10 %iger Natrinm- salieylatlSsung Eiwei$fraktionen erhalten, die sich in ihrer Aminos~iurezusammensetzung ein- deutig unterscheiden. Diese Autoren schliel3en daraus, dab der Kleber ein einziger EiweiB- komplex ist, der in eine groBe Zahl yon Fraktionen zerlegt werden kann, die sich chemiseh und physikalisch unterseheiden.
Doch nicht nur fiber die Homogenitat der Kleberkomponenten Gliadin und Glutenin ist keine endgfiltige Klarung erzielt worden, sondern auch hinsichtlieh der Einheitliehkeit der salz- und wasserlSslichen Eiweil~stoffe des Weizens, der Globuline und Albumine.
Die "Physiological and Biochemical Committees on Protein Nomenclature ''3 bezeichnen die Globuline als ,,einfache Proteine, unlSslich in reinem Wasser, aber 15s]ich in neu~ralen Salz- 15sungen starker Basen mit starken S~uren (die Grenzen der Fallbarkeit durch Ammoninmsulfat sollten nicht als Unterseheidungsmerkmal zwisehen Albuminen nnd Globulinen dienen)".
GOnTNEX und Mitarbeiter 4 wenden gegen die Anwendung dieser Bezeiehnung ffir die lViehi- proteine ein, dab durch die verschiedenen Arten yon Salzl5sungen und bei verschiedenen Konzen- trationen unterschiedliche Mengen an Proteinen aus dem Mehl gewonnen werden. STAKER und GO~TNER s lehnen aueh den Ausdruck ,,LSslichkeit" ffir die Getreideproteine ab und halten den Begrfff , ,Peptisation" ffir angebrachter. Bei der Peptisation der Getreideproteine entstehen in Abh~ngigkeit yon dem LOsungsmittel (Wasser, Salz-L5sung) Sole, die Mieellen versehiedener I)ispersionsgrade enthalten. GORTNER sehreibt: ,,Es ist nicht ausgeschlossen, dab eine groI]e Anzahl yon Gedanken in der Chemie der Proteine, an welehen man tells aus Tr~igheit, tells aus Tradition festh~ilt, ftir die weitere Entwicklung eher schiidlich als nfitzlich sind. Jedenfalls ist es im l~alle des Globulins erforderlieh, die gegenw~rtigen Ansehauungen yon dessen chemiseher Einheitliehkeit aufzugeben."
I)iese Feststellung ist aueh yon anderen Bearbeitern experimentell erhartet worden. So hat DANIELSSO~ 6 bei Behandlung eines Salzextraktes des Weizenmehles in der Ultrazentrifuge zwei untersehiedliehe Globulinkomponenten gefunden. Mehrere Globuline sind auch in den Samen anderer Ar tender Gramineen gefunden worden. QVENSEL ~ bezeiehnet vier Globuline der Gerste gem~B ihrer steigenden Molekulargewiehte Ms ct-, fl-, y- und 6-Komponenten. Im Keimling des Weizens ist nur die y-Komponente gefunden worden, w~hrend das Endosperm haupts~chlich die ~-Komponente und kleinere Betr~tge des ~-Globulins enthi~lt.
Aueh die Existenz echter Pflanzenalbumine als definierte ehemische Einheiten wird yon STAXER und GOXT~ER in Frage gestellt, und die Untersehiede gegenfiber anderen Eiwefl]fraktionen werden in der L~nge der Protein-Mieellen gesucht.
In neuerer Zeit haben nun LAws und FRANCE s bei der Elektrophorese eines w~l~rigen Extraktes des Mehles die Uneinheitliehkeit der Albumine nachgewiesen, und PENCE, ELDER und M E c I ~ ~ ist der elektrophoretisehe Naehweis yon sechs Albuminen m5glich gewesen,
Was ffir die Eiwei$stoffe des Weizens gesagt worden ist, grit naturgem~l~ auch fiir die des Roggens, die jedoeh bei weitem bisher nicht so intensiv untersucht worden sind. Die Unterschiede im Verhalten des Roggeneiwei$ und des Weizeneiweil~ sind zwar schon mehrfaeh Gegenstand yon ErSrterungen gewesen; dabei ist aber immer yon der grundsiitzliehen Identit~it der Eiweil~- komplexe, besonders aber der Gliadine, ausgegangen worden. Diese Identit~t hat eine gewisse
1 KmZL~AN~, A. G.: Nature (London) 140, 119 (1937). 1V[cCALLA, A. G., u. R. C. ROSE: Canad. J. Res. 12, 346 (1935).
3 Physiological and Biochemical Committees on Protein Nomenclature, Physiol. and Bioehem. Committees on Protein: J. of Biol. Chem. 4, 48, 51 (1908); Amer. J . Physics 21, 27, 30 (1908).
4 GORTNER, R. A., W. F. HOFF~A~ U. W. B. SINCLAIr: Kolloid-Z. 44, 97 (1928). 5 STAKER, E. V., u. R. A. GO~TNER: J . Phys. Chem. 25, 1565 (1931). 6 DANIELSSON, C. E.: Biochemie. J. 44, 387 (1949). 7 QUENSEL, O.: Diss. Univ. Upsala 1942. s LAws, W. D., u. W. G. ~ANCE: Cereal Chem. 25, 231 (1948). 9 PENCE, J. W., A. H. ELDE~ U. D. K. ~EC]=IAM: Cereal Chem. 28, 94 (1951).

Chemische Differenzierung der EiweiBstoffe des Weizens und Roggens. I. 87
Stfitze dureh I-IAGnE~ ~ erfahren, der fiber die Isolierung eines Kleber-/ihnlichen Eiweil~kSrpers aus Roggenmehl berichtet hat. Die yon HESS ~ bei der rfntgenographischen Untersuchung ermittelten Interferenzen lassen jedoch einen tiefer gehenden Unterschied vermnten.
Eigene Untersuchungen. I n der vor l iegenden Arbe i t i s t der Versuch u n t e r n o m m e n worden, die EiweiB-
stoffe des Weizens und Roggens mi t I-Iilfe der N a t r i u m h y p o c h l o r i t - R e a k t i o n zu differenzieren und festzustel len, inwieweit ihre A b t r e n n u n g au f der Basis ihrer Lfs l i chke i t in verschiedenen L5su/~gsmitteln zu ehemisch ~hnliehen oder anders- ar~igen E iwe iBkf rpe rn fiihr~. Es is t d a v o n ausgegangen worden, dab die Reak t ions - f i thigkeit der EiweiBmolekii le aueh wei tgehend mi t der r~umlichen A n o r d n u n g und F a l t u n g der P e p t i d k e t t e n zusammenh~ng t und diese durch den AblSsungsprozeB yon einem Eiwe i6komplex ver~tndert werden kann. Die Mfgl ichke i t der Beeinf lussung in t r amoleku la re r Wassers to f fb indungen , wie sic PALTLI~G und NI]~MA_~ a ffir EiweiB- molekii le postul ieren, du tch geringfiigige Einflf isse wie p~-Versehiebungen, Neu t ra l - s~lze, d i i r f te im Hinb l ick au f die Gewinnungsmethoden der GetreideeiweiBstoffe ebenfal ls eine l~olle spielen, und es is t v e r m u t e t worden, dab dies in einem abweichen- den R e a k t i o n s v e r l a u f bei ihrer Umse~zung mi t Na~r iumhypoch lor i t s ich tbar wird.
Auf den Verlauf der Umsetzung der EiweiBstoffe mit Natriumhypochlorit, bei der Kohlen- s~iure und Stickstoff entstehen, soll nicht n~her eingegangen werden. Er ist Gegenstand der Arbeiten yon RASCI-[IG4~ L&~GHELD 5 WOHL 6, ENGFELD 7, ABDEB~I~ALDE~S, ~]-OR~-~[AN ~ und GOLD- SCtIMIDT ~.
Die quantitati.ce Verfolgung der Natriumhypochlorit-Reaktion durch Messung der ent- wiekelten Gase (Kohlens~ture und Stickstoff) im W~R~tmG-Manometer ist "con ]:[ERKEN und Mitarbeitern ~0 zur Kennzeiehnung yon Serumproteinen angewendet worden. Auch SC~O~.~i~LLEa ~ hat die manometrische Messung der entwickelten Kohlens~ure aufdie Untersuchung "con Nahrungs- eiweil] angewendet, um Ver~nderungen bei weehselnden Lagerbedingungen zu .cerfo]gen. SCHOa- ~i)LLE~ hat bei seinen Untersuchungen auch die alkohollSsliche Fraktion des Weizeneiwei]~, das Gliadin, in seine Untersuchungen mit einbezogen, es aber lediglieh im Vergleich zu anderen Eiweil~kSrpern betraehtet.
W e n n die N a t r i u m h y p o c h ] o r i t - R e a k t i o n auch ke inen u n m i t t e l b a r e n E inb l i ck in die S t r u k t u r der Pro te ine zul~f~t, so g ib t doch ein unterschiedl iehes Verha l t en der Eiweif~stoffe gegenfiber N a t r i u m h y p o e h l o r i t , das in den Differenzen der gemessenen Gasmengen zum Ausd ruck kommt , Mfg l ichke i t en zu ihrer Charakter i s ie rung.
Methodik. a) Apparatur.
Ffir die Messung der bei der Umsetzung mit Natriumhypochlorit entwickelten Gasmengen ist ein WA~BU~G-Ger~t mit 6 Manometern verwendet worden. Die mit dem Manometer .cer- bundenen kegelfSrmigen Reaktionskflbehen sind mit kleinen Eins~tzen ffir Kalilauge und seit- lichen Eins~tzen zur Aufnahme der Natriumhypoeh]oritl5sung .cerschen. Die Kflbchen werden
1 ttAGBE~G, S.: Bull. i~cole Meun. belge 14, 1 (1952). 2 HEss, K.: Mfillerei, 5, Sonder-H. 28, 32 (1952). 3 PAVLIX~, L., u. C. NIE~A~: J. Amer. Chem. Soc. 61, 1860 (1939).
RASC~IG, F.: Ber. dtsch, chem. Ges. 40, 4580 (1907). LA-~GI-IELD, K.: Hoppe-Seylers Z. 121, 18 (1922). WooL, A.: Bet. dtsch, chem. Ges. 33, 2763 (1900).
7 E~'GFELD, N. ]:).: Hoppe-Seylers Z. 121, 18 (1922). s ABnER~ALD~, E., u. H. SICK~L: Hoppe-Seylers Z. 170, 134 (t927). 9 NORMAn-, M. F.: Bioehemic. J. 30, 484 (1936).
10 GOLDSCHYlIDT, ST., U. K. STRAUSS: Liebigs Ann. 471, 1 (1929). ~ HEnXE~, ill., u. J. Sc~u~: : Naunyn-Schmiedebergs Arch. exper. Path. 206,
102 (1949). - - tIERKv.~, I-I., u. tt.-O. SILEERSI]~PF: Naunyn-Schmiedebergs Arch. exper. Path. 212, 205 (1951).
~2 SCEoRMi~LL~n, Ji: Dtsch. Lebensmittel-gdseh. 48, 113, 136 (1952).

88 M. ROttl~LICH, G. AI)LE:~ und O. KRA~IM:
wie folgt gefiillt: 0,5 ml EiweiGlSsung (bis 180 ~ Stickstoff), 1,5 ml Phosphatpuffer nachS6R~WSE~ (PH 6,4) und 5 ml NatriumhypochloritlSsung (6 mg NaOC1). In zwei K51bchen werden als Thermo- barometer start der Eiwei$lSsung 0,5 ml des verwendeten LSsungsmittels und die gleiche NaOC1- Menge einpipettiert. Die hier abgelesenen Volumina werden yon den Mel3werten abgezogen, da sie Temperatur- und Luftdruckschwankungen und den Grad der Natrinmhypochloritzersetzung w/~hrend der Messung erfassen. Ffir die Stiekstoffmessungen werden zus~tzlieh 0,5 ml 10%ige K O t t in den Inneneinsatz des KSlbchens geffillt, um das entwickelte COs zu absorbieren.
Da in einigen Vorversuchen die gemessenen Stickstoffmengen nur sehr gering waren, ist auf die Ermittlung des Stickstoffwertes verzichtet und die gesamte Gasmenge gemessen und angegeben worden.
b) Herstellung des NatriumhypochIorits.
Das benStigte t typoehlorit ist stets kurz vor dem Versuch dureh Einleiten yon Chlor in n-Natronlauge hergestellt worden. Da die Na0Cl-LSsung fiir die Versuche weder zu stark alkalisch, noch zu sauer sein daft, mug sie unter Verwendung einer Glaselektrode mit n-Essigsi~ure oder n-Natronlauge auf einen p . -Wert yon 7,7 eingestellt werden. Anschliel~end wird der Hypochlorit- gehalt durch Titration mit Natriumthiosulfat ermittelt und durch Verdiinnung mit H20 auf die erforderliche Konzentration yon 6,0 rag/0,5 ml gebracht. Diese Menge reieht fiir EiweiGmengen bis zu 180 y 1N aus.
c) Die manometrische Messung.
Die gefiillten KSlbchen werden an dem Manometer befestigt und in den Thermostaten yon 38 ° C eingesetzt. Nach 10 min ffir den Temperaturausg]eich des l~eaktionsgemisches werden die Manometer gesehlossen, das Natriumhypochlorit dureh Kippen in den Reaktionsraum des KSlbehens gebraeht und 2 Std. gleiehm/~Gig gesehiittelt (125 Vor- und Rfiekw~rtsbewegungen pro min). Die Ablesungen werden in Abstanden yon 10, 20, 30, 60, 90 und 120 rain vorgenommen.
Um einen gesicherten Durchschnittswert zu erhalten, sind im allgemeinen bei jeder Messung jeweils mindestens 16 Werte ermittelt - - auf 100 y gelSsten Stiekstoff berechnet - - und die Mittelwerte graphiseh aufgetragen worden.
d) Untersuchungsmaterial.
Als Untersuehungsmaterial sind Weizenmehle und Roggenmehle gleieher Kornpartien, a b e t verschiedener Ausmahlungsgrade verwendet worden. Zur tterstellung der Eiweil~lSsungen ist
das Mehl in einer Glas-
Mehlmenge
g
' 10
LSsungsmittel
Ar~
dest. Wasser
6 10
2,0 WM ÷ 3,5 RM -[- 1,0 Kleber
10 10
SalzlSsungen w~Griger Alkohol
n-Essigs~ure ~
2 % ige HC1 2%ige X-I~PO~
~ienge ml
100 300
3 500 X 50
250
100
5O0 5OO
Ex- stopfenflasehe mit dem je- traktions- weiligen LSsungsmittel eine
dauer bestimmte Zeit lang bei S t d . Raumtemperatur geschfit-
telt, vom Ungel~isten dureh 1 Zentrifugieren abgetrennt
und nStigenfalls filtriert worden.
0,5 1 e) Die statistlsche
G~dtiglceit der Messungen. 24
Da bei den Messungen 1 zeitweise grSGere Schwan- 1 kungen auftreten, sind bei
der alkohollSsliehenFraktion des Weizens und des Roggens 40 Messungen und bei den besonders stark schwankenden Ablesungen ftir die wasserlSsliehe Fraktion des Roggens 68 Messungen vorgenommen worden. Die statistische Auswertnng der Messungen ist dann nach der yon DAEVES und BECKEL ~ entwiekelten tti~ufigkeitsan~lyse und Grol~zahlmethodik erfolgt und die sog. T~0-Spanne erreehnet worden. Sie hat bei der alkohollSsliehen Fr~ktion des Weizens 17,2 (T00~ = ~= 8,6 ram3), bei der fiir Roggen 30 (Tg0 , 2 = ~= 15 mma), bei der wasserlSslichen Fraktion des Roggens 70 (T90 , ~ = ~ 35 mm ~) betragen.
Bei Essigsaure uls L6sungsmittel sind for Roggenmehl [I~M], Weizenmehl [WM] und Kleber untersehiedliche Mengen angewendet worden.
I)AEVES, K., u..h_. BECKEL: GroGzahlforsehung uad H~ufigkeits-Analyse. Weinheim: ¥erl . Chemie 1948.
Chemische Differenzierung der EiweiBstoffe des Weizens und Roggens. I. 89
Ergebnisse. 1. Der EinJlul3 des L6sungsmittels.
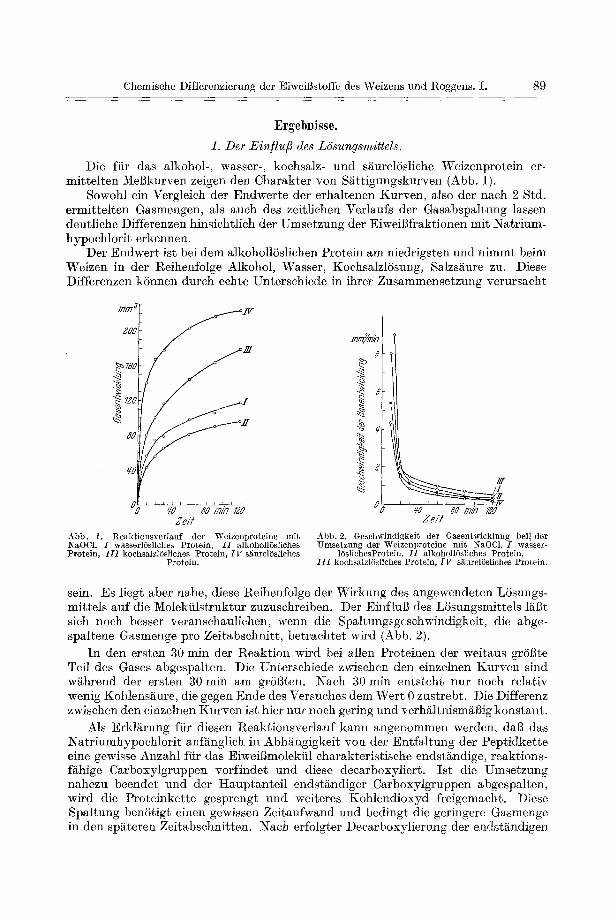
Die fiir das alkohol-, wasser-, koohsalz- und s~urelSsliehe Weizenprotein er- mittelten Mel3kurven zeigen den Charakter yon S/~ttigungskurven (Abb. 1).
Sowohl ein Vergleich der Endwerte der erhaltenen Kurven, also der naeh 2 Std. ermittelten Gasmengen, als auch des zeitliehen Verlaufs der Gasabspaltung lassen deutliehe Differenzen hinsiehtlich der Umsetzung der EiweiBfraktionen mit Natrium- hypochlorit erkennen.
Der Endwert ist bei dem alkohollSsliehen Protein am niedrigsten und nimmt beim Weizen in der Reihenfolge Alkohol, Wasser, Koehsalzl6sung, Salzsi~ure zu. Diese Differenzen kSnnen durch echte Untersehiede in ihrer Zusammensetzung verursacht
zz', L t _..........:s
k ~ d 0
0 ) t ' t L/kr i i80 1/77))7':~0
Ze/t Abb. 1. Reaktionsverlauf der Weizenproteine mit NaOCL I wasserlUsliehes Protein, I I alkohollUsliehes Protein, I I I koehsMzlUsliehes Protein, I F siiurelUsliehes
Protein.
:nmymi:7 8
>..
p i
gO 80 re~J7 IZO Zeit
Abb. 2. Gesehwindigkeit der Gasentwieklung bei] der Umsetzung der Weizenproteine mit NaOC1. I wasser-
15slichesProtein, I f alkohollSsliehes Protein, I I I kochsalzlUsliches Protein, I V sSourel6sliches Protein.
sein. Es liegt aber nahe, diese Reihenfolge der Wirkung des angewendeten Lbsungs- mittels auf die Molekiilstruktur zuzusehreiben. Der Einflul3 des LSsungsmittels 1/~13t sieh noch besser veranschauliehen, wenn die Spaltungsgesehwindigkeit, die abge- spaltene Gasmenge pro Zeitabsehnitt, betraehtet wird (Abb. 2).
In den ersten 30 min der Reaktion wh'd bei allen Proteinen der weitaus gr6Bte Teil des Gases abgespalten. Die Untersehiede zwischen den einzelnen Kurven sind wikhrend der ersten 30 rain am grSl3ten. Naeh 30 min entsteht nut noeh relativ wenig Kohlens~ure, die gegen Ende des Versuehes dem Wert 0 zustrebt. Die Differenz zwisehen den einzelnen Kurven ist bier nur noeh gering und verhikltnism~13ig konstant.
Als Erklkkrung fiir diesen Reaktionsverlauf kann angenommen werden, dab das Natriumhypoehlorit anf/knglieh in Abhangigkeit yon der Entfaltung der Peptidkette eine gewisse Anzahl fiir das Eiweigmolekiil eharakteristisehe endstandige, reaktions- fikhige Carboxylgruppen vorfindet und diese deearboxyliert. Ist die Umsetzung nahezu beendet und der Hauptanteil endstikndiger Carboxylgruppen abgespalten, wird die Proteinkette gesprengt und weiteres Kohlendioxyd freigemaeht. Diese Spaltung benStigt einen gewissen Zeitaufwand und bedingt die geringere Gasmenge in den spkkteren Zeitabschnitten. Naeh erfolgter Decarboxylierung der endst/mdigen

90 M. ROHRLICtt, G. ADLER und O. KRAMM:
Carboxylgruppen verlauft die weitere Spaltung der Ket ten dureh das Natrium- hypoehlorit bei allen Fraktionen mit ann~hernd gleieher Gesehwindigkeit.
Nur die salzsaure Fraktion (Abb. 2, Kurve IV) zeigt hier eine Ausnahmestellung. Naehdem in den ersten 30 rain ein sehr groger Betrag an C02 freigemaeht worden ist, ist die weitere Gasentwicklung, genau wie bei den anderen EiweiBstoffen, relativ gering. Die Kurve der Gasentwicklung nimmt aber in einem welt schnelleren Mage
ab und erreicht in einem frfiheren Zeitpunkt ihren ~m ~ [ / ' ~ Endwert als die Kurven der fibrigen Fraktionen. Es ist
/6of . / ~ ~ N denkbar, dab sehon unter der Einwirkung der Salzs~ure eine Aufspaltung der Peptidkette erfolgt ist und dag
120~ / / J " ~ / " ~ ] das Hypoehlori t auf eine grSBere Anzahl freiliegender ~ g Endgruppen trifft als bei den anderen Fraktionen.
o z7 Einen ~hnliehen Verlauf, wenn auch mit anderen 8 o ~ _ o ° Werten, zeigen die Eiweigstoffe des Roggens (Abb. 3).
Vergleicht man die nach 2 Std. Reaktionszeit gemesse- gOlt]/ nen Endwerte, so ergibt sich eine gewisse Ubereinstim-
mung der Werte ffir die alkohol- und wasserl6slichen EiweiBfraktionen des Weizens und l~oggens. Die s~ure-
°c ' ~)' ' '8) rz/;7 '30 15sliehen Fraktionen unterscheiden sieh jedoch, selbst Zell
unter Berfieksichtigung der relativ grogen Sehwankungs- Abb. a. l~eaktionsverlauf der breite der MeBwerte, reeht erheblieh (Tab. 1). Dag aueh
Roggenproteine mi t NaOC1. r wasserlSsliehes Protein, II alko- die Art der S~ure ffir die Struktur der herausgelSsten hollUsliehes Protein, I f I koehsalz- Eiweigfraktion eine t~olle spielt, kommt im VerhMten des 16sliehes Protein, I V salzs~iure- 15sliches Protein, V phosphor- Phosphors~ure-15slichen Eiweig zum Ausdruek. I)urch
s~iurelSsliches Protein. diesen Befund erseheint die yon HAGBEaG angegebene MSglichkeit, mit Hilfe von Si~uren aus Roggenmehl eine
dem Weizenkleber iihnliche Substanz zu isolieren, in einem anderen Licht. Dar- fiber und ebenfalls fiber das Verhalten der salzlSs]iehen Proteinfraktion wird sp~ter zu spreehen sein.
Tabelle
Mehl Yore
Weizen t%oggen Roggen Weizen Weizen Weizen Weizen Roggen Roggen Weizen Roggen Roggen Weizen Roggen Roggen Roggen
1. Gemessene Gasmenge bei der Reaktion der Proteine des Weizens und Roggens mit NaOC1.
Asche N/10 g i~Iehl
% mg
0,75 133 0,51 66 1,15 168 0,75 133
66 168 133 66
168 133 66
168 168
LSsungsmittel
Wasser
/15%ig ]35% ig J55%ig
Alkohol |72 %ig ]72%ig t72 % ig
NaC1-Lsg. 10%ig
HC1 2 % ig
HAP04 2%ig
0,51 1,15 0,75 0,51 1,15 0,75 0,51 1,15 1,15
N herausgelSst aus I0 g Mehl
rag %
24,4 18,3 21,0 31,8 34,0 20,0 21,7 16,3 28,4 21,3 56,7 42,6 58,7 44,1 28,0 42,1 47,0 28,2 15,2 11,4 21,0 32,3 34,0 20,4 19,0 14,3 28,0 42,5 38,0 22,8
101,0 64,5
N[it NaOC1 Gasmenge umgesetzte nach 2 Std.
M.enge N pro 100 ~ N nlm 3
122,6 27,0 48,0 43,4 56,7
113,4 117,3 46,0 79,0 38,0 36,0 57,0 19,0 28,0 39,0
I 101,0
120 126 126
99 97
100 96 89 90
180 176 151 225 164 174 112

Chemische Differenzierung der Eiweigstoffe des Weizens und Roggens. [. 91
Die 1Jbereinstimmung der mit wgl3rigem Alkohol weehse]nder Kon- zentrat ion gewonnenen Eiweigex- t rak te ha t dazn angeregt, die alko- holischen Ex t r ak t e (Prolamine) yon Getreideart Hafer, Gerste und Mats auf ihre geakt ionsfghigkei t gegeniiber Na- t r iumhypochlor i t zu untersuehen. Gerste . . . . .
Wie die ermit tel ten Megwerte Haler . . . . . (Tab. 2) zeigen, ist die Uberein- Mats . . . . . . s t immung unter Ber~eksiehtigung Weizen . . . . der Sehwankungsbrei te der Mes- Roggen . . . . sungen eine recht gate. Die Reak- tionsf~higkeit der MkohollSsliehen der Menge der vorbiandenen Fettkomp.onenten. Die vor des t tafer- und Maissehrotes mit Ather ermittel ten Reakt ionskurven (Abb. 4) verlaufen versehieden.
Wenn aueh der Fet tgehal t des Weizens und Roggens nieht so hoeh ist wie der yon I-Iafer und Mats, so mug doeh aueh hier angenommen werden, dag die Reaktions- fghigkeit des MkohollSsliehen Proteins dureh den Fe t t - gehalt beeinfluBt wird. Es seheint eine Analogie zu den Befunden yon T ~ a v , BVF~'E und Bn~TON ~ vorzuliegen, die eine gr6Bere Resistenz gegen fermentat ive Angriffe bet 8erumproteinen feststellen konnten, die dureh Lipoid- bindnng gesehiitzt waren.
Tabelle 2. Gemeasene Gasmenge bet der Reaktion der al]coholl6slichen Proteine verschledener Cerealien mit
Natriumhypochlorit.
~I n de IMi tNaOOl I e ge s ~ . _ ~ e s _ a _ a - - i I:~aSlilenge l{Sslichm~ u ~ U~a~e~ I ~aaeh 2 Std. Proteins J b~r°~a~2l ~ pro 100 y .X
°/o i ' ~ ~ rl] Ill ~
24,0 87 90 16,4 98 (51) lO0 17,2 i 85 (57) 100 44,0 117 96 28,2 79 90
Eiwei0fraktion wird jedoeh beeinflul3t yon und naeh der En t fe t tung
2. Der Ein/lufi der Salz~onzentration, des Kations und dea Anions au[ die gelgste Eiweififralction.
Die Wirkung verschiedener Salze auf die physikalischen Eigenschaften der EiweiBk6rper hat t to~msmEn 2 durch Au£ stellung sog. lyotroper Reihen charakterisiert, und HOFF~V und Go~m~E~ a vermochten such ffir die PeptJsierung der Pro-
aJ I
q~
0 ¢0 ,70 mli7 120 Ze/7
Abb. 4. Einflul3 der Lipoidkompo- nente auf den l~eaktionsverlauf des Haferprolamins mit R'aOC1.
I entfettet, I I nicht, entfettet.
teine des Weizens solche lyotropen Reihen aufzust.ellen. Bei Salzen mit Kalium Ms Kation haben HOJr:P~AS; and GO~T~R die Reihenfolge F < SO~ < C1 < Tartrat < CrQ < Brund bet Salzen mit Chlorals Anion die tl.eihenfolge Na < K < Ba < Li < Sr < Mg < Ca aufgefunden. Asl]erdem konnten sie beobachten, dab mit steigender Konzentration das Peptisierungsvermggen der Alkalihalogenide sinkt and dag Untersehiede in der Wasserstoffionkonzentration ffir die lyotropen Reihen nicht verantwortlich sind.
Es ha t nun interessiert, wie sich die dutch LSsungen verschiedener Salze gelSsten Eiweigfrakt ionen bet der Behandlung mit Nat r iumhypochlor i t verhalten. Werden mit clan verschiedenen SMzlSsungen nur verschieden groBe Mengen des gleiehen Proteins herausgelSst oder werden Proteine herausgelSst, die sieh strukturell unterseheiden .~
Die Gewinnung der einzelnen Ex t rak te ist genau nach der yon GORT~R ange- ftihrten Vorsehrift erfolgt, und zur Ex t rak t ion sind folgende Salz]6snngen verwendet worden: n-K2SO4-L5sung , n-KBr-L6snng, n - N a 2 H P Q . L S s u n g ' n . K 2 H P O c L 6 s u n g ' 10% ige KC1-LSsung, n-NaC1-LSsung, n-KC1-LSsung, n-Na2804-LSsnng ' ]0%ige NaC/-L6sung.
1 TAYAI2, t~., G . M. :BUFFE U. J. BRETOR-: B u l l . Soe . Chim. biol. 29, 671, 677, 780 (1947). HOF~ISTER, F. : Naunyn-Sehmiedebergs Arch. expel P~th. 24, 247 (1888).
a t Io~IA~, W, F., u. R. A. GOI~T~En: Cereal Chem. 4, 221 (1927).
92 M. ROHRLIOH, G. ADLEI¢ und O. KI~A~:
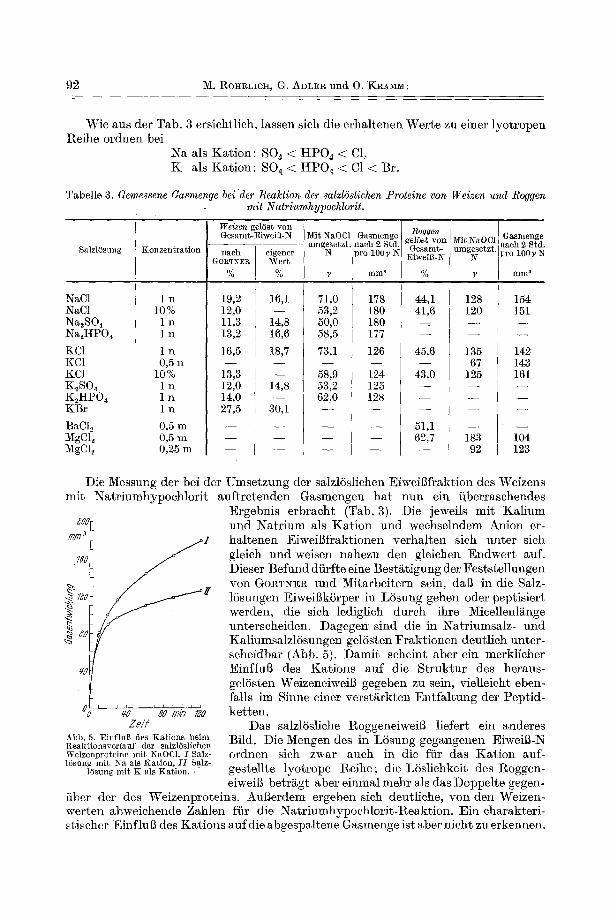
Wie aus der Tab. 3 ersichtl ich, lassen sich die e rha l tenen W e r t e zu einer lyo t ropen Reihe o rdnen bei
N a als K a t i o n : SO~ < HPO~ < CI, K als K a t i o n : SO~ < HPO~ < C1 < Br.
Tabelle 3. Gemessene Gasmenge bei "der Reaktion der salzlSslichen Proteine von Weizen und Roggen mit Natriumhypochlorit.
Weizen gel6st yon [ Roggen [ Gesamt-EiweiB-N Mit NaOCI ] Gasmenge Mit NaOC1 Gasmenge
umgesetzt, nach 2 Std. gelUs~ yon nach 2 Std. Gesamt- umg~etzt. Salzl6sung Konzen~ra~ion nach eigener N pro 100 y N :Eiwei~-N ~ro 100 ~ N GoRTNEI~ Wert
% % ~ mm 3 % mm 3
NaC1 NaC1 Na2SO4 Na~HPO,
KC1 KC1 KC1 K~S04 K2HPO4 KBr
BaC12 MgC12 MgO12
1 n 19,2 10% 12,0
1 n 11,3 1 n 13,2
1 n 16,5 0,5 n
10% 13,3 1 n 12,0 1 n 14,0 1 n 27,5
0,5 m 0,5 m 0,25 m
16,1
1 ,8 !6,6
18,7
1 ,8 3 ,1
71,0 53,2 50,0 58,5
73,1
57, 53,2 62,0
178 180 180 177
126
125 128
44,1 41,6
45,6
4;,0
51,1 62,7
128
67 125
183 92
154 151
142 143 161
104 123
Die Messung der bei der Umse tzung der salzl6slichen Eiweil~fraktion des Weizens mi t N a t r i u m h y p o c h l o r i t au f t r e t enden Gasmengen h a t nun ein i iberrasehendes
.:oo rn/n~ I
7#0
z
¢5
Zeit Abb. 5. Einflug des Kations beim Reaktionsverlauf der salzlUsliehea Weizenproteine mit NaOCl. I Salz- 16sung mit Na als Kation, II Salz-
]6sling mit K als :Kation.
Ergebnis e rb rach t (Tab. 3). Die jeweils mi t K a l i u m u n d N a t r i u m als K a t i o n und wechselndem Anion er- ha l t enen Eiweil3fraktionen verha! ten sich un te r sich gleich und weisen nahezu den gleichen E n d w e r t auf. Dieser Befund di i r f te eine Besti~tigung der Fes t s te l lungen yon GO~TNE~ und Mi ta rbe i t e rn sein, dag in die Salz- ]6sungen Eiweigk6rper in L6sung gehen oder pep t i s ie r t werden, die sich lediglich du tch ihre Micellenli~nge unterscheiden. Dagegen sind die in Na t r iumsa lz - und Kal iumsa lz lSsungen gelSsten F r a k t i o n e n deut l ieh unter - scheidbar (Abb. 5). D a m i t scheint aber ein merkl icher E in f lug des K a t i o n s auf die S t r u k t u r des heraus- gelSsten WeizeneiweiB gegeben zu sein, vie]leicht eben- falls im Sinne e iner versti~rkten En~fal tung der Pep t id - ke t ten .
Das salzlSsliehe Roggeneiweig l iefert ein anderes Bild. Die Mengen des in L6sung gegangenen Eiweig-N ordnen sieh zwar aueh in die fiir das K a t i o n auf- ges te l l te lyo t rope Reihe ; die L6sl iehkei t des Roggen- eiweig be t r~g t aber e inmal mehr als das Doppe l t e gegen-
fiber der des Weizenpro te ins . A u g e r d e m ergeben sieh deutl iehe, yon den Weizen- wer ten abweiehende Zahlen ffir die Na t r i umhypoe h lo r i t -Re a k t i on . E in eharakter i - st iseher EinfluB des Ka t i ons au fd i e abgespa l t ene Gasmenge ist aber n ieht zu erkennen.
Chemische Differenzierung der Eiweiflstoffe des Weizens und Roggens. I. 93
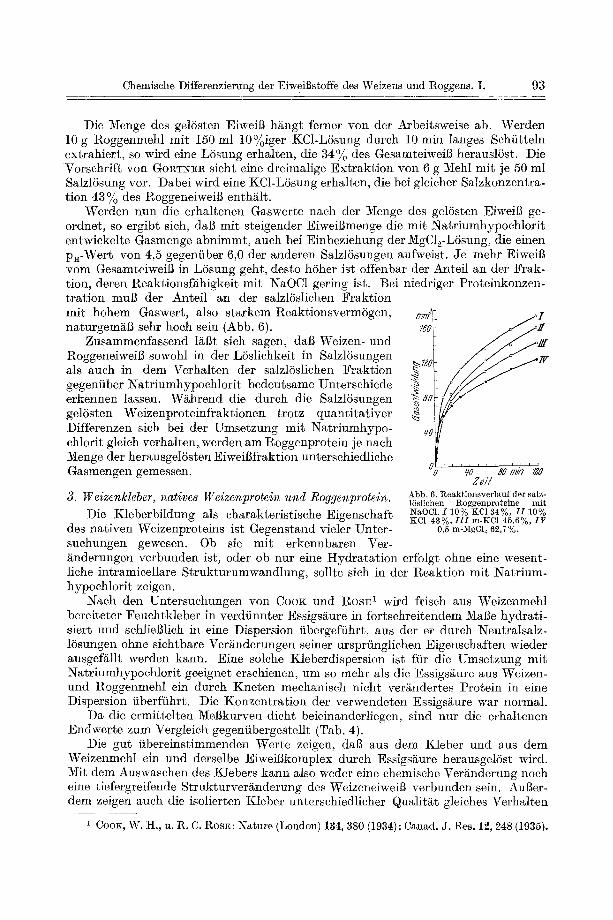
Die Menge des gelSsten EiweiB h/~ngt ferner v o n d e r Arbeitsweise ab. Werden 10 g Roggenmehl mit 150 ml 10%iger KC1-LSsung dureh 10 min langes Sehiitteln extrahiert, so wird eine L6sung erhalten, die 34% des GesamteiweiI~ herauslSst. Die Vorsehrift yon GORT~R sieht eine dreimalige Extrakt ion yon 6 g Mehl mit je 50 ml Salzl6sung vor. Dabei wird eine KC1-LSsung erhalten, die bei gleicher Salzkonzentra- tion 43 % des RoggeneiweiB enthg~lt.
Werden nun die erhaltenen Gaswerte naeh der Menge des gelSsten Eiweig ge- ordnet, so ergibt sieh, daB mit steigender Eiweigmenge die mit Natriumhypochlorit entwiekelte Gasmenge abnimmt, auch bei Einbeziehung der 3/igC12-L6sung , die einen p~-Wert yon 4,5 gegenfiber 6,0 der anderen SalzlSsungen aufweist. Je mehr EiweiB veto Gesamteiweig in LSsung geht, desto hSher ist offenbar der Anteil an der Frak- tion, deren geaktionsf/~higkeit mit NaOC1 gering ist. Bei niedriger Proteinkonzen- tration mug der Anteil an der salzlSsliehen Fraktion mit hohem Gaswert, also starkem ReaktionsvermSgen, naturgemgg sehr hoeh sein (Abb. 6).
Zusammenfassend l~Bt sieh sagen, ' dab Weizen- und I~oggeneiweiB sowohl in der LSsliehkeit in SalzlSsungen als such in dem Verhalten der salzl6sliehen Fraktion gegenfiber Natriumhypoehlorit bedeutsame Untersehiede erkennen lassen. W~hrend die dutch die SalzlSsungen gelSsten Weizenproteinfraktionen trotz quantitativer Differenzen sieh bei der Umsetzung mit Natriumhypo- ehlorit gleieh verhalten, werden am Roggenprotein je naeh Menge der herausgelSsten EiweiBfraktion untersehiedliehe Gasmengen gemessen.
3. Weizenkleber, natives Weizenprotein und Roggenprotein. Die Kleberbildung als charakteristische Eigenschaft
des nativen Weizenproteins ist Gegenstand vieler Unter- suchungen gewesen. Ob sie mit erkennbaren Ver- i~nderungen verbunden ist, oder ob nur eine Hydratat ion erfolgt ohne eine wesent- liche intramicellare Strukturumwandlung, sollte sich in der t~eaktion mit Natrium- hypochlorit zeigen.
Nach den Untersuchungen yon Cook und l~osn 1 wird frisch aus Weizenmehl bereiteter Feuehtkleber in verdfinnter Essigs~ure in fortschreitendem MaBe hydrati- siert und sehlieBlich in eine Dispersion iibergeffihrt, aus der er durch Neutralsalz- 16sungen ohne sichtbare Ver~nderungen seiner ursprfinglichen Eigenschaften wieder ausgef£11t werden kann. Eine solehe Kleberdispersion ist ffir die Umsetzung mit Natriumhypochlorit geeignet erschienen, um so mehr als die Essigsi~ure aus Weizen- und Roggenmehl ein durch Kneten mechanisch nieht veri~ndertes Protein in eine Dispersion iiberfiihrt. Die Konzentration der verwendeten Essigsiiure war normal.
Da die ermittelten ?¢[eBkurven dicht beieinanderliegen, sind nur die erhaltenen Endwerte zum Vergleich gegenfibergestellt (Tab. 4).
Die gut fibereinstimmenden Werte zeigen, dab aus dem Kleber und aus dem Weizenmehl ein und derselbe Eiweigkomplex durch Essigsgure herausgelSst wird. Mit dem Auswaschen des Klebers kann also weder eine chemische Ver~nderung noch eine tiefergreifende StrukturverSmderung des WeizeneiweiB verbunden sein. AuGer- dem zeigen auch die isolierten Kleber unterschiedlieher Qualit/~t gleiches Verhalten
CooK, W. H., u. R. C. Rosin: Nature (London) 134, 380 (1934); Canad. J. Res. 12, 248 (1935).
H /80
N
%
qo
, , , r , i , i i , 1
O0 ¢0 80 fn/b 7#0 Zeit
Abb. 6. ~,eak~ionsverl~uf der salz- 16sliohen l~oggenproteine mit NaOC1. I 10% KC1 34%, I I 10% KCI 43%, I I I m-KCl 45>6%, fp
0,5 m-MgCl~ 62,7%.
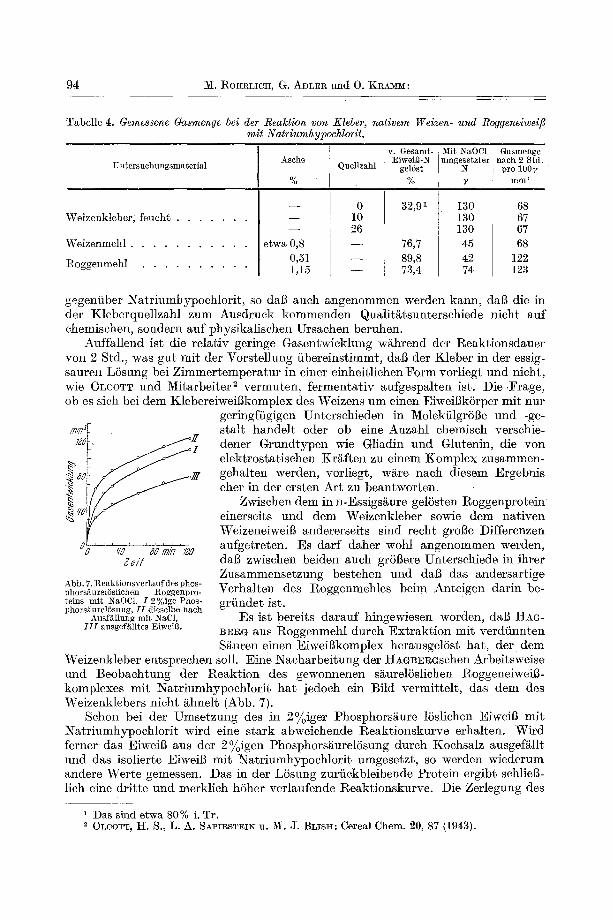
94 M. ROItRLICIC[, G. ADLER und O. KRAMM:
Tabelle 4. Gemessene Gasmenge bei der Realction yon Kleber, nativem Weizen- und Roggeneiweifi mit Natriumhypochlorit.
Un~ersuchungsmateriM
Weizenkleber; feueht . . . . . . .
Weizenmehl . . . . . . . . . . .
Roggenmehl . . . . . . . . . .
Asche
% Quellzahl
e t w a 0,8 0,51 1,I5
v. Gesamt- Mi~ NaOC1 I Gasmenge Eiweifi-N umgesetzter nach 2 Std.
gel6st N ! pro 100y
100 32, 91
26 - - 76,7 - - I 89,8 - - ', 73,4
130 68 130 67 130 67 45 68 42 122 74 123
gegeniiber Natriumhypochlorit, so dag aueh angenommen werden kann, dab die in der Kleberquellzahl zum Ausdruek kommenden QuMit/~tsuntersehiede nieht auf ehemisehen, sondern auf physikalisehen Ursaehen beruhen.
Auffallend ist die relativ geringe Gasentwieklung wahrend der I~eaktionsdauer yon 2 Std., was gut mit der Vorstellung iibereinstimmt, dal3 der Kleber in der essig- sauren LSsung bei Zimmertemperatur in einer einheitliehen Form vorliegt und nieht, wie OLeOTT und Mitarbeiter 2 vermuten, fermentativ aufgespalten ist. Die .Frage, ob es sieh bei dem KlebereiweiBkomplex des Weizens um einen EiweiBk6rper mit nur
geringfiigigen Unterschieden in Mo]ekiilgrSBe und -ge- mm 3 stalt handelt oder ob eine Anzahl ehemiseh versehie-
:zo ~ ~ B ' dener Grundtypen wie Gliadin und Glutenin, die yon ~ I elektrostatisehen Kr~ften zu einem Komp]ex zusammen- ~8~ ~r gehalten werden, vorliegt, w~re naeh diesem Ergebnis ~ eher in der ersten Art zu beantworten.
Zwisehen dem in n-Essigs~ure gelSsten Roggenprotein einerseits und dem Weizenkleber sowie dem nativen WeizeneiweiB andererseits sind reeht groBe Differenzen
' '~) . . . . . . . . aufgetreten. Es darf daher wohl angenommen werden, 0 00 fnl)7 720
Ze/7 d~13 zwisehen beiden aueh grSgere Untersehiede in ihrer Zusammensetzung bestehen und dab das andersartige
Abb. 7.Reaktionsverlaufdesphos- Verhalten des Roggenmehles beim Anteigen darin be- phorsi~urelSsliehen ttoggenpro- reins mit NaOC1. I 2%ige Phos- griindet ist. phors~iurelSsung, II dieselbe nach
Ansf~llung mit NaC1, ]~S ist bereits darauf hingewiesen worden, dab HAG- I I I ausgefglltes EiweiB. BErG aUS Roggenmehl dutch Extraktion mit verdiinnterL
S~uren einen EiweiBkomplex herausgel6st hat, der dem Weizenkleber entspreehen soll. Eine Naeharbeitung der HaGB~RGsehen Arbeitsweise und Beobaehtung der Reaktion dos gewonnenen s~urelSsliehen RoggeneiweiB- komplexes mit Natriumhypoehlorit hat jedoeh ein Bild vermittelt, das dem des Weizenklebers nieht/ihnelt (Abb. 7).
Sehon bei der Umsetzung des in 2% iger Phosphors~ure 15sliehen Eiweig mit Natriumhypoehlorit wird eine stark abweiehende Reaktionskurve erhalten. Wird ferner das Eiweig aus der 2% igen Phosphors~urel6sung dutch Koehsalz ausgef~llt und das isolierte EiweiB mit Natriumhypoehlorit umgesetzt, so werden wiederum andere Werte gemessen. Das in der L6sung zuriickbleibende Protein ergibt sehlieg- lieh eine dritte und merklieh h6her verlaufende l~eaktionskurve. Die Zerlegung des
Das sind etwa 80% i. Tr. 20LCOT% I-I. S., L. A. SAPIt'STEIN U. M. J. BLIStt: Cereal Chem. 20, 87 (1943).

Chemische Differenzierung der Eiweil~stoffc des Weizens und Roggens. I. 95
in 2~oiger Phosphors/~ure gel6sten EiweiBkomplexes in eine f~l lbare F rak t i on , die nur wenig mi t N a t r i u m h y p o e h l o r i t reagier t und eine zweite in LSsung verble ibende und merkl ieh reakt ionsf~higere F r a k t i o n m a e h t eine ~ n d e r u n g der E iweiBs t ruk tur durch die Ausf/~llung wahrseheinl ich. Sie zeigt auBerdem deut l ieh die Abh~ingigkeit der I~eakt ionsf~higkei t des gewonnenen Pro te ins yon der Arbei tsweise. DaB die ausgef/i l l te E iwe ig f rak t ion nahezu vollsti~ndig in 72 ~oigem Alkohol 15slich ist, weist wel ter d a r a u f hin, dab zwisehen dem naeh HAGB~RG aus Roggenmehl gewonnenen E iwe igkomplex und dem K l e b e r k o m p l e x des Weizens keine Ahnl iehke i t bes tehen kann.
4. Der Ein/lu/3 der Temperatur au] das Klebereiwei/3.
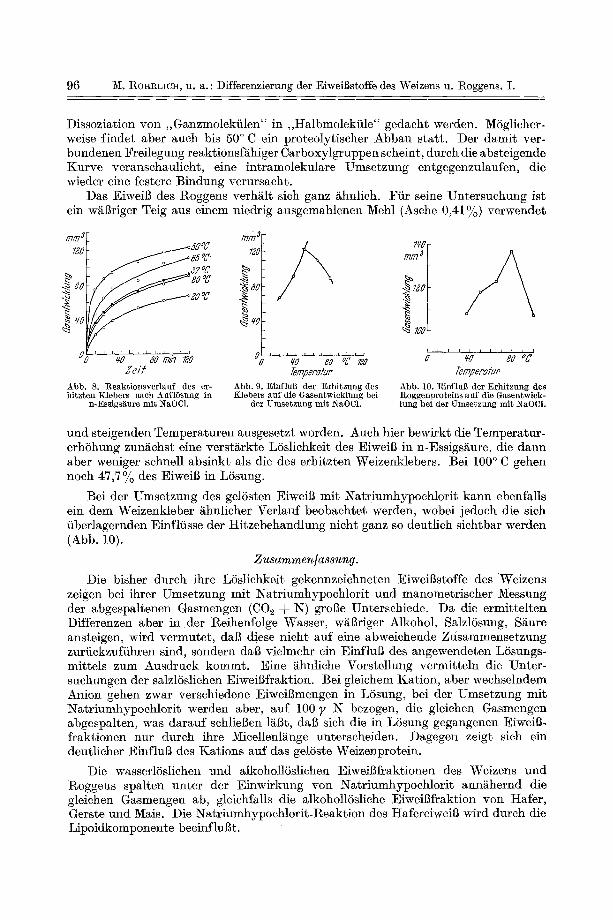
Nach OLCOTT und BLIsg 1 erfolgt beim Trocknen des Weizenklebers unter Atmosph/irendruck, insbesondere abet beim Kochen mit Wasser, eine vonder Dauer der Erhitzung abh/ingige Denatu- rierung. DaB dcr Feuchtigkeitsgehalt des Klebers bei der Hitzedenaturierung yon Bedeutung ist, haben P ~ c ~ und Mitarbeiter ~ beobachtet. Wiihrend bei niedrigem Feuchtigkeitsgehalt die I)enaturierung relativ gering ist, steigt sie bei einem Wassergehalt yon 35--40 % stark an. PENOn hat die Veri~nderungen der Eigenschaften des erhitzten Klebers am Abfall der LSslichkcit in verdfinnter Essigs~ure und an der Abnahme des Volumens yon Geb~icken, die unter Zusatz des erhitzten Klebers hergestellt worden waren, beobachtet. Nach S~ARP und GORT~ER a ist die alkohollSsliche EiweiBfraktion, das Gliadin, wenigcr temperaturempfind]ieh als die Glutenin- komponente. Doch land ROI-IRLICtI a, dal~ beim Erhitzen des Feuchtkleberkomplexes unter Atmosph~irendruck bei 100 ° C auch die Menge des alkohollSslichen Anteils stark verringcrt ist gegeniiber der des im Vakuum getrockneten Klebers.
Beim Trocknen des Klebers mfissen also in t r amoleku la re Umse tzungen verlaufen, die eine S t r u k t u r v e r ~ n d e r u n g hervorrufen, eine Vermutung , die sieh aueh durch die N a t r i u m h y p o c h l o r i t - R e a k t i o n yon F e u c h t k l e b e r (etwa 60% H~O-Gehalt) , der s te igenden Tempera tu r en ausgese tz t wurde, ha t bes t~t igen lassen. Die Tab. 5 gibt ein Bi ld fiber die in n-Essigs~iure in L6sung gegangene KlebereiweiBmenge in Ab- h~tngigkeit yon der Tem- peratur. Bis zu einer Temperatur yon 50°C s te ig t die LSsl ichkei t des Klebers in n-Essig- s~ture an, dann abe t f~ll t
s i c sehroff ab. Bei einer einstf indigen Erh i t zung des Klebers au f 100 ° C sind nur noch e twa 7~0 des Gesamte iweig 15slieh.
Tabclle 5. Einflufl der Temperatur au/ die Menge des aus Weizen- kleber bzw. _Roggeneiweifl mit n-Essigsiiure herausgelSsten Proteins.
3ffaterial
Weizenkleber . . . RoggeneiweiB . .
20o°c yo
78,5 73,5
Oel6stes Protein, bet. als N bei
STo°C 50£C 65°C go %
82,1 85,6 64,7 79,0 82,2 82,4
80 ° C 100 ° C % %
30,9 6,7 66,0 47,7
Die durch N a t r i u m h y p o e h l o r i t ve ru r sach te Gasentwick- lung h a t bei e twa 50 ° C ein ~ [ax imum (Abb. 8).
\ ¥ e r d e n die e rha l tenen E n d w e r t e in Abh~tngigkeit yore Tempera tu rans t i eg auf- ge t ragen und die e rha l tenen K u r v e n verlitngert., so erh~It m a n das in Abb. 9 gezeigte Bild.
Der Ver lauf der K u r v e n ve rmi t t e l t die Vorstel lung, dal3 das M a x imum der Gas- abspa l tung dureh (3ber lagerung zweier en tgegengese tz t ger ichte ter Reak t ionen bed ing t ist. Bei dem Anst ieg der Kurve , der eine Aufspa l tung der Eiweil?ket te ver- m u t e n l~tl3t, kann an die eingangs erwfihnte, yon KI~EJcI und SV E D ~a G angenommene
10LCOTT, H. S., u. M. J. BLISI~: Trans. Amer. Assoc. Cereal Chemists 2, 20 (1944). 2 PIercE, J. W., A. IVIoHAg~AD U. D. K. I~{EOJ&4_lW: Cereal Chem. 30, 2, 115 (1953). 3 Sm~m~, p. F., u. R. A. Got~T~m~: Univ. Minnesota, Agric. Exper. Stat., Techn. Bull. No. 19
(1923). 4 RomcnIcg, M.: Jahresber. Vcrsuchsanst. Getreideverwert. 19g0/~l, S. 81.
96 ~ . I~OHRLIClt, U. 8,. : Differenzierung der Eiweil~stoffe des Weizens u. Roggens. I.
Dissoziation yon ,,Ganzmolekiilen" in ,,Halbmolekiile" gedacht werden. MSglicher- weise finder aber auch bis 50 ° C ein proteolytischer Abbau start. Der damit ver- bundenen Freilegung reaktionsfghiger Carboxylgruppen scheint, durch die absteigende Knrve veranschaulich~, eine intramolekulare Umsetzung entgegenzulaufen, die wieder eine festere Bindung verursacht.
Das Eiweil~ des Roggens verhglt sieh ganz ghnlich. Fiir seine Untersuchung ist ein wgl~riger Teig aus einem niedrig ausgemahlenen Mehl (Asehe 0,41%) verwendet
rnn73 /50°E
//~ 7 .eev ~ 80
o 0 80 rn/n ' ~0 Zeif
Abb. 8. Reaktionsverlauf des er- hitzten Klebers nach Aufl6snng in
n-Essigsgure mit ~¢aOC1.
m/77 3
I¢OF A /17/1"13 t
,f Ig a f ~
/erzosr~/ur Abb. 9. Einflu2 der Erhitzung des Klebers auf die Gasentwicklung bei
der Umsetzung mit NaOC1.
) r i I p I I I I | ~
o ~0 80 ~g TenvperoluP
Abb. 10. Einflul~ der Erhi tzung des Roggenproteins auf die Gasentwick- lung bei der Umsetzuug mit ~aOCl.
und steigenden Temperaturen ausgesetzt worden. Aueh hier bewirkt die Temperatur- erhShung zungchst eine verstgrkte LSslichkeit des Eiweil~ in n-Essigsgure, die dann aber weniger schnell absinkt als die des erhitzten Weizenklebers. Bei 100 ° C gehen noch 47,7 °/o des Eiweil~ in LSsung.
Bei der Umsetzung des gdSsten Eiweil~ mit Natriumhypoehlorit kann ebenfalls ein dem Weizenkleber i~hnlicher Verlauf beobaehtet werden, wobei jedoch die sich iiberlagernden Einflfisse der Hitzebehandlung nicht ganz so deutlich sichtbar werden (Abb. 10).
Zusammen/assung.
Die bisher durch ihre LSslichkeit gekennzeichneten Eiweil~stoffe des "Weizens zeigen bei ihrer Umsetzung mit Natriumhypochlorit und m~nometriseher Messung der abgespaltenen Gasmengen (C02 d- N) grebe Unterschiede. Da die ermittelten Differenzen aber in der l~eihenfolge Wasser, wgBriger Alkoho], SalzlSsung, Sgure ansteigen, wird vermutet, dal~ diese nieht auf eine abweichende Zusammensetzung zurfickzufiihren sind, sondern dal~ vielmehr ein Einflul~ des angewendeten LSsungs- mitte]s zum Ausdi'uck kommt. Eine ghnliche Vorstellung vermitteln die Unter- suchungen der salzlSslichen Eiweil~fraktion. Bei gleichem Kation, aber weehselndem Anion gehen zwar verschiedene Eiweil~mengen in LSsung, bei der Umsetzung mit Natriumhypochlorit werden aber, auf 100 y N bezogen, die gleiehen Gasmengen abgespalten, was darauf sehliel~en lgl3t, dal~ sich die in LSsung gegangenen EiweiB- fraktionen nut durch ihre Micellenlgnge unterscheiden. Dagegen zeigt sieh ein deutlieher Einflul~ des Kations auf das gelSste Weizenprotein.
Die wasserlSslichen und alkohollSslichen Eiweil~fraktionen des Weizens und ]~oggens spalten unter der Einwirkung von Natriumhypochlorit ann~hernd die gleiehen Gasmengen ab, gleichfalls die alkohollSsliche Eiweil~fraktion von Hafer, Gerste und Mais. Die Natriumhypoehlorit-Reaktion des Hafereiweil~ wird dureh die Lipoidkomponente beeinflul~t.

J, SCltORMI~LLER und L. LEICtITEI~: Beitr/ige zur Bioehemie der K/~sereifung. XIV. 97
Bei der Umse tzung der essigsauren Dispers ion versehiedener sich durch ihre Quel lbarke i t un te r sehe idender Weizenkleber und des na t iven Weizenmehle iweig mi t N a t r i u m h y p o e h l o r i t en t s tehen nahezu gleiche Gasmengen, so dal3 angenommen werden kann, dab dureh das Auswasehen des Klebers aus dem Weizenmehl m i t Wasse r nnd bei der Dispergierung in n-Essigs£ure kein A b b a u oder keine wesent l iehe U m w a n d l u n g ve rursaeh t wird. Die in der Quellzahl naeh B ~ I ~ S n und KOOr~WAh'~ ausgedrf iekten Qual i tg t sunte rseh iede der Weizenkleber haben phys ika l i sehe Ur- saehen.
Roggenpro te in in ve rd i inn te r Ess igg iure dispergier t , e rg ib t bei der Umse tzung mi t N a t r i u m h y p o e h l o r i t andere W e r t e als na t ives ~Veizenmehleiwefi3 und Kleber . Es muB demnaeh aueh eine andere S t r u k t n r aufweisen.
Beitr/ige zur Biochemie der K/isereifung. X I V . Mitteilung.
Das V o r k o m m e n y o n L y s i n d e c a r b o x y l a s e in r e i f e n d e m S a u e r m i l c h M i s e .
Von
J . SCHORMULLER u n d L. LEICHTER.
Mitteilung aus dem Institut /i~r Lebensmittdchemie und Lebensmitteltechnologie der Technischen Univemitiit Berlln-Charlottenburg*.
Mit 7 Textabbildungen.
(Eingegangen am 11. Januar 1955.)
I n der vorhergehenden Arbe i t 1 h a t t e n wir das Vorkommen yon Glu tamins~ure- deea rboxy lase in re i fendem Sauermilehk~se naehgewiesen und das in Rede s tehende F e r m e n t nigher gekennzeichnet . Gleiehzeit ig wurde au f Grund e ingehender Er- 5 r te rung der diesbezi igl ichen L i t e r a t u r fiber Anwesenhe i t verschiedener Amine in K~se die Vermutung gei~ui3ert, dal~ in de ra r t igem re i fendem Mater ia l mi t dem Vor- k o m m e n auch andere r Aminosi~uredeearboxylasen zu rechnen ist. Von solchen F e r m e n t e n h a b e n wir in der hier vor l iegenden Unte r suchung Auf t r e t en und Verha l ten der Lys indeca rboxy lase i iberprfif t . I h r Vorkommen in re i fendem K~se war r ech t wahrscheinl ich, da verschiedene Angaben der L i t e r a tu r Cadaver in als korrespondie- rendes A m i n der Lys indeca rboxy l i e rung in K~se erw~hnen ~.
Lysindecarboxylase ill Mikroorganismen. Es ist schon lange bekannL dai3 Coli-Stamme, Pseudomonas fluoreseens und verschiedene
Baeillus-Arten l(+)-Lysill decarboxylierend abzubauen verm6gen 3. GALE und seine Schule ~ haben als besonders aktiven Tr~ger dieses Ferments Bacterium cadaveris (Stature ~r. 6578 der
* Die vorliegende Arbeit wurde durch eine Beihilfe der Deutschen Forschungsgemeinschafg gefSrdert. Ihr sei hierftir auch an dieser Stelle verbindlichst gedankt.
Sc~oa~iffLL~, J., u. L. LnlC~TEn: Diese Z. 102, 13 (1955). 2 WIXTE~ST~I~, E., u. I. T~5~-I: Hoppe-Seylers Z. 86, 28 (1902). - - vA~- SLY~:E, D. D., u.
E. B. HA~T: Amer. Chem. J. 30, 8 (1903). - - SI~O~A~T, P., u. J. M~¥~vgo~: Netherl. Milk and Dairy J. 6, 1 (1952). - - BLoo~, 1~. J. : J. Dairy Sci. 34, 1 (1951). - - SILVE~X.¢~, G. J., u. F. V. KOSlKOWSKI: J. Dairy Sci. 37, 645 (1954).
_~ltere Literatur bei A. J ~ K E : Arch. Mierobioh 15, 472 (1951). - - PO~E~, J. R. : Bacterial Chemistry and Physiology. S. 870. ~ew York: J. Wiley a. Sons Inc. 1947.
G~L~, E. F., u. H. M. R. Eels : Nature (London) 1~2, 327 (1943); Biochemic. J. 38, 232 (1944). - - G~L~, E. F. : Biochemic. J. 89, 46 (19¢5); Adw in Enzymol. 6, 1 (1946).-- B ~ s c ~ o , H . : Biochim. et Biophysica Aeta 4, 130 (1950). - -L~nD~, H. : Respiratory Enzymes. Minneapolis: Burgess 1949.
Z. Lebensmi~tel-Unters. u. -Forsch., :Band 102. 7