
A Bayesian Approach to Assess the Tanscriptome of Bone Aging and the Role of the Brd2
Gene in the Regulation of Sex Linked Bone Loss
Amira I. Hussein, Ph.D.1, Joseph Wu, Ph.D.2, Mayetri Gupta, Ph.D.3, Louis Gerstenfeld, Ph.D.1. 1Boston University Medical Center, Boston, MA, USA, 2Boston University, Boston, MA, USA, 3University
of Glasgow, Glasgow, United Kingdom.
Disclosures: A.I. Hussein: None. J. Wu: None. M. Gupta: None. L. Gerstenfeld: None.
Introduction: Aging and sex are among the major risk factors of osteoporosis. Aging involves numerous
biological processes. Longitudinal microarray studies have contributed immensely to the understanding
of aging and the associated pathogenesis. However, one of the main challenges in such studies lies in
the lack of multi-factorial (for example sex and genetic mutation) computation approaches to assess
temporal microarray profiles. An aging study was carried out in male and female mice expressing
reduced expression of the transcriptional regulator bromoDomain2 (Brd2lo). Mice with reduced
expression of BrD2 become obese but do not develop insulin resistance [1] hence eliminating the
confounding effects caused by Type II diabetes. Analyses of the bone phenotype showed that female
mice with Brd2 mutation had an age-dependent bone loss, while no differences were found in male
mice [2]. The goals of this study were to: (1) cluster and then characterize the gene expression profiles
of aging male and female mice with and without the Brd2 mutation using a new Bayesian statistical
algorithm; and (2) assess the biological functions of the genes in each cluster.
Methods: Specimens. Male and female wild type (WT) and Brd2lo (Het) mice on a C57BL/6 background
were used in a longitudinal aging study (3, 6, 9, and 12 months; 4 mice/time point; approved by IACUC).
Microarray Analysis. Total RNAs were isolated separately from the both right and left whole humeri of
mice tissues using procedures developed for murine bones [3]. One microgram of RNA was labeled and
used for hybridization. The GeneChip Mouse Gene 1.0ST Arrays were used for our studies (Affymetrix,
Santa Clara, CA). Out of a total of 21,225 probes, only genes expressing 3-fold change between the
maximum and the minimum values across all samples were used in this study (3,950 genes). Gene
Cluster Discovery Algorithm. A Bayesian modeling approach that captures information on three different
levels (sex, genotype and time) simultaneously was developed in R. The algorithm relies on detecting
groups of genes that behave in a similar way over time, under a particular combination of factors, and
differ from other set(s) of genes in their pattern of behavior. Each group of genes is defined as a cluster.
Biological Functions. The biologic functions of the gene in each cluster were assessed using Ingenuity
Pathway Analysis tool (IPA, QIAGEN Redwood City). All genes expressing 3-fold change as well as genes
from each cluster identified by the gene cluster discovery algorithm were imported into IPA. Gene
functions were grouped into general categories such as skeletal and muscular, immune, and
inflammatory related functions. Only biological functions with p < 0.05 were considered.
Results: Eighteen clusters were identified using the gene cluster discovery algorithm (4 subsets are
shown in Figure 1). Notable differences among the clusters were observed in the patterns of the
differential gene expression. For example, the gene profiles in cluster 4 indicate that the set of genes in
this cluster have similar expression levels within the male mice. While the expression levels for the
female heterozygous mice are different from the wild-type female mice but resemble that of the male

mice. Figure 2 represents a bar plot of all biological functions for each cluster (with 15 genes or more) as
well as the set of all genes used in the gene cluster discovery algorithm (3-fold: 3,950 gene set). Clusters
4, 14, and 18 had the highest percentage of skeletal and muscular related genes. The majority of the
skeletal and muscular related genes in Cluster 4 had skeletal (bone)-related functions, while the genes in
Cluster 18 had muscle-related functions only. Cluster 14 had a small number of genes (15 genes).
Therefore, Cluster 4 was chosen for further assessment of skeletal-related biological functions. Many of
the skeletal related biological processes indicated that over time, the Brd2 female mice resemble the
gene expression levels in male mice. For example, for metabolic bone disease, all genes in this cluster
were down-regulated in male (wild type and Brd2lo) and Brd2lo female mice relative to wild type female
mice (figure 3).
Discussion: Consistent with the known and/or predicted phenotypes ~30% and ~10% respectively of the
genes overall that were differentially expressed showed a relationship to the
hematological/immunological and skeletal and muscular tissue functions (figure 3; 3-fold bar). A
qualitative assessment of the profiles from the gene cluster discovery algorithm facilitated the
identification of gene groups that show commonality by sex or genetic mutation as well as the
interaction between the two factors. The results indicate that over time, the Brd2 female mice resemble
the gene expression levels in male mice. These results are in agreement with quantitative data on bone
loss in the trabecular bone compartment of the tibia of these same groups. Brd2 has no effect on bone
loss in male mice over time, whereas in female mice, the Brd2 mutation resulted in more bone loss over
time compared to wild type in a manner similar to male mice. Taken together, the outcomes indicate
that even though females have decreased bone volume fraction compared to males, their bone
metabolism is increased.
Significance: Clustering of genes based on the differential gene expression among male, female, with
and without Brd2 mutation was achieved through the use of a novel multi-factorial Bayesian statistical
approach. The expression of the skeletal related gene show that female mice with the Brd2 mutation
have similar gene expression profiles to male mice, while no differences were found between wild type
and Brd2 male mice. These data identify a gene (BrD2) that belongs to a gene family that imparts
epigenetic regulation and may be part of the mechanisms that control sex regulated differences in male
and female bone loss with aging.
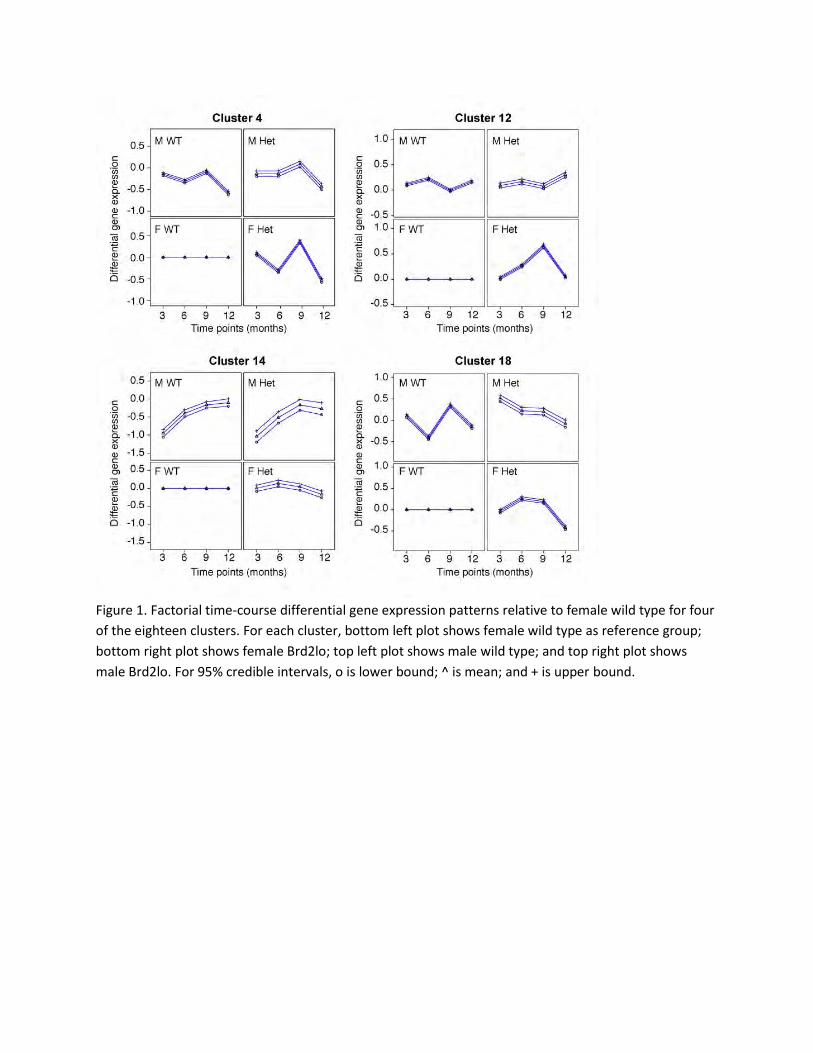
Figure 1. Factorial time-course differential gene expression patterns relative to female wild type for four
of the eighteen clusters. For each cluster, bottom left plot shows female wild type as reference group;
bottom right plot shows female Brd2lo; top left plot shows male wild type; and top right plot shows
male Brd2lo. For 95% credible intervals, o is lower bound; ^ is mean; and + is upper bound.
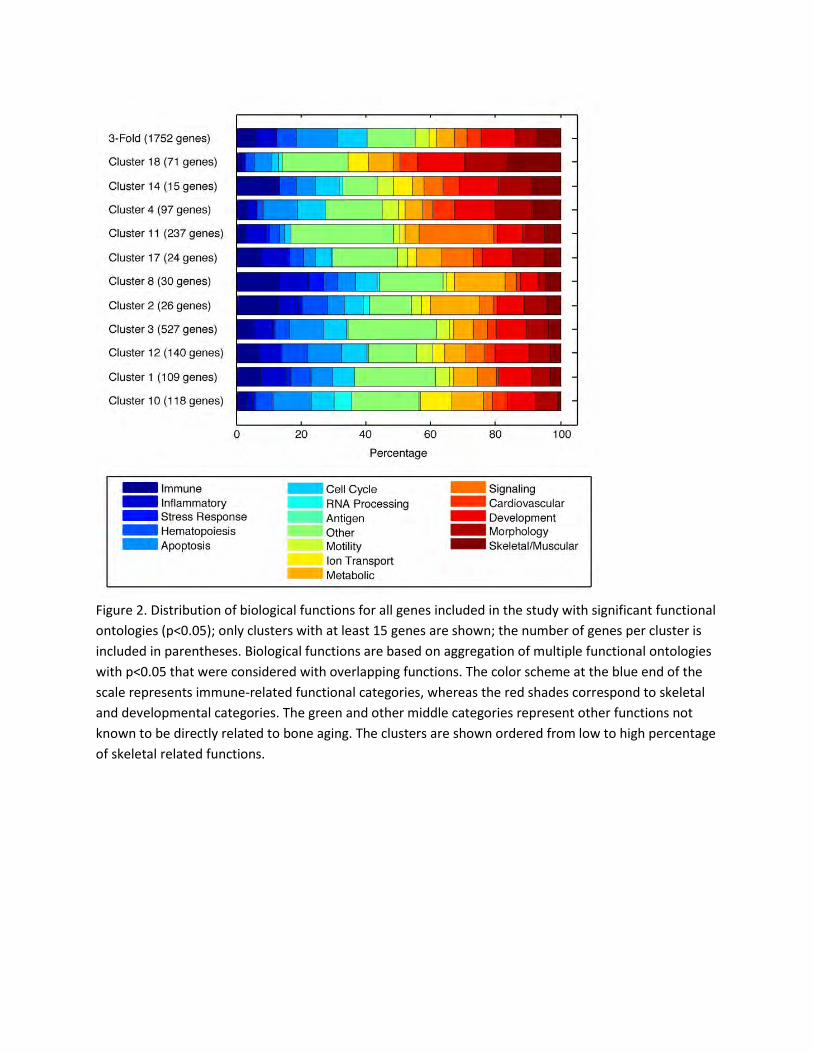
Figure 2. Distribution of biological functions for all genes included in the study with significant functional
ontologies (p<0.05); only clusters with at least 15 genes are shown; the number of genes per cluster is
included in parentheses. Biological functions are based on aggregation of multiple functional ontologies
with p<0.05 that were considered with overlapping functions. The color scheme at the blue end of the
scale represents immune-related functional categories, whereas the red shades correspond to skeletal
and developmental categories. The green and other middle categories represent other functions not
known to be directly related to bone aging. The clusters are shown ordered from low to high percentage
of skeletal related functions.

Figure 3. Network Relationship of Expressed Gene Group for the Highest Ranked Disease Associated
Function in Clusters 4. The similarity of the networks in the three groups indicate that similar genetic
mechanisms are turned on in the Brd2lo female mice, as in the males.
ORS 2015 Annual Meeting
Poster No: 1408
Recommended

















