
]REVIEWS
T h e interphase nucleus of p!a_n.t_~ .~s usually imagined, albeit unconsciously, as a bowl of spaghetti - strands of interphase chromosomes running at random through the volume of the nucleus. During cell div- ision, these strands are organized into discrete meta- phase chromosomes, but most preparations still give the impression of little or no suprachromosomal or- ganizatic, n. Recent work, both in plants and animals, is questioning some of these hidden assumptions, and suggests that it may be possible to relate nuclear archi- tecture to aspects of gone expression and chromosome behaviour.
Such assumptions were not made when chromo- somes were first described. Early drawings show sym- metrical arrangements of chromosomes and their arms throughout the cell cycle, and the physical disposition of chromosomes within the nucleus was regarded as important. For example, Fig. 1 shows a drawing from sections of nuclei in the desert plant Yucca, in which similarly sized chromosomes are together, and large chromosomes lie in a peripheral domain surrounding the smaller chromosome#.
Nuclear architecture describes the structure and pattern of the nucleus. To understand the architecture, we must know about the three-dimensional organ- ization of the nucleus, including both the position and identity of each chromosome. These two simple ideals have rarely been achieved because the nucleus is dynamic (moving through the cell cycle, transcribing RNA and replicating DNA), small (typically 10 I.tm diameter) and its chromosomes thread-like (0.2 om diameter at interphase and 1 ~tm at metaphase) and often with similar morphologies. The techniques of chromosome spreading pioneered in plants by Darlington and colleague# largely changed the way chromosomes were examined. Spread and squashed preparations of metaphases are ideal for most cyto- genetics: they enable the counting of chromosomes, and the examination of chromosome morphology. Chromosomes can be easily ident~ed in banded, two-dimensional spreads of metaphases made for the light micro- scope, and three-dimensional position can be reconstructed from sections, but combining the two techniques is diffi- cult.
Many attempts have been made to analyse the disposition of chromosomes at metaphase in spread preparations3. Although lalge numbers of dividing cells can be obtained easily, and almost any plant can be used, analysis of archi- tecture is difficult because the three- dimensional nucleus has been reduced to two dimensions. Large sample sizes may only increase the chance of assess- ing artefacts of spreading. In order to overcome the difficulty of identifying chromosomes, chromosomes with par- ticular morphologies, including hetero- chromatic blocks (large tandemly repeated DNA sequences, which may relate to the C-banding patterns seen on
Nuclear architecture in plants J.S. HESLOP-HARRISON AND M.D. BENNETt
Structure within the nucleus of plants is becoming imcreasiagIF clear in both metapbase and interphase nucleg Mthough there are conflicting data about the relative positions of individual and pairs of chromosomes. At interphase, imdltvidual chromosomes may gefteral~ occupy discrete domains that are wet Intermixed with other chromosomes. Aspects of mechanical chromosome behatvlour and even of gene expression may correlate with imterpbase chromosome positiot~ aml imply that a better understanding of nuclear architecture is required
chromosomes), have often been examined. Avivi et al. 4 analysed the positions of mitotic wheat chromo- somes that were missing a whole arm (telocentric chromosomes), and so could be easily identified. However, we reported that large systematic errors were introduced when positions of telocentric chromosomes paired with norrnai chromosomes ('marked' bivalents) were analysed in squashes of wheat meiotic prep- arationsS. At metaphase I, we found that there was a strong tendency for any marked bivalent to lie near the edge of the metaphase spread preparation, regard- less of which particular chromosome type gave rise to the marked bivalent.
Within the past ten years, interest has returned to e~amination of whole nuclei, from both plants and animals. In plants, extensive work has been carried out using serial section reconstructions at meiosis 6 and at mitosisT, s, particularly in the cereals. The newer techniques of confocal microscope reconstructions and computer processing of h~lages of sections9,10 are also being increasingly used. The problem of chromosome identification, at both interphase and metaphase, is
~ 0 A drawing by Miiller from 19091, showing sections u¢ nuclei in the dc~,:rt plant Yucca, where similarly sized chromosomes are together through the division. Large chromosomes lie in a peripheral domain, surrounding the smaller chromosomes.
~G D~Cr~BER 1990 VOL. 6 NO. 12
01990 Elsevier Science Publis~e~ Lid (UK) 0 l ( ~ • 9479/~0/$02.00
~ E V I E W S
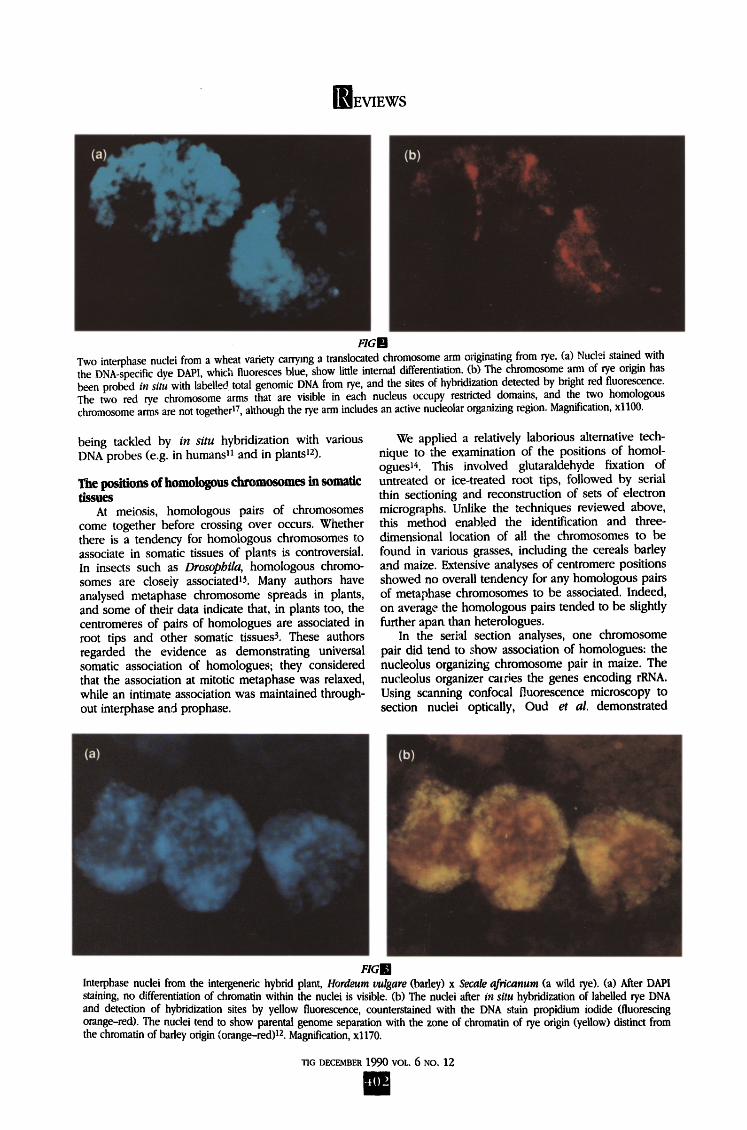
Two interphase nuclei from a wheat variety carrying a translocated chromosome arm originating from rye. (a) Nuclei stained with the DNA-specific dye DAPI, which fluoresces blue, show little internal differentiation. (b) The chromosome arm of rye origin has been probed in situ with labelled total genomic DNA from rye, and the sites of hybridization detected by bright red fluorescence. The two red rye chromosome arms that are visible in each nucleus occupy restricted domains, and the two homologous chromosome arms are not together 17, although the rye arm includes an active nucleolar organizing region. Magnification, x1100.
being tackled by in situ hybridization with various DNA probes (e.g. in humans n and in plantsl2).
The positions of homologous chromosomes in somatic tissues
At meiosis, homologous pairs of chromosomes come together before crossing over occurs. Whether there is a tendency for homologous chromosomes to associate in somatic tissues of plants is controversial. In insects such as Drosophila, homologous chromo- somes are closely associatedl3. Many authors have analysed metaphase chromosome spreads in plants, and some of their data indicate that, in plants too, the centromeres of pairs of homologues are associated in root tips and other somatic tissue#. These authors regarded the evidence as demonstrating universal somatic association of homologues; they considered that the association at mitotic metaphase was relaxed, while an intimate association was maintained through- out interphase and prophase.
We applied a relatively laborious alternative tech- nique to the examination of the positions of homol- ogue# 4. This involved glutaraldehyde fixation of untreated or ice-treated root tips, followed by serial thin sectioning and reconstruction of sets of electron micrographs. Unlike the techniques reviewed above, this method enabled the identification and three- dimensional location of all the chromosomes to be found in various grasses, including the cereals barley and maize. Extensive analyses of centromere positions showed no overall tendency for any homologous pairs of metaphase chromosomes to be associated. Indeed, on average the homologous pairs tended to be slightly further apart than heterologues.
In the serial section analyses, one chromosome pair did tend to show association of homologues: the nucleolus organizing chromosome pair in maize. The nucleolus organizer carries the genes encoding rRNA. Using scanning confocal fluorescence microscopy to section nuclei optically, Oud et al. demonstrated
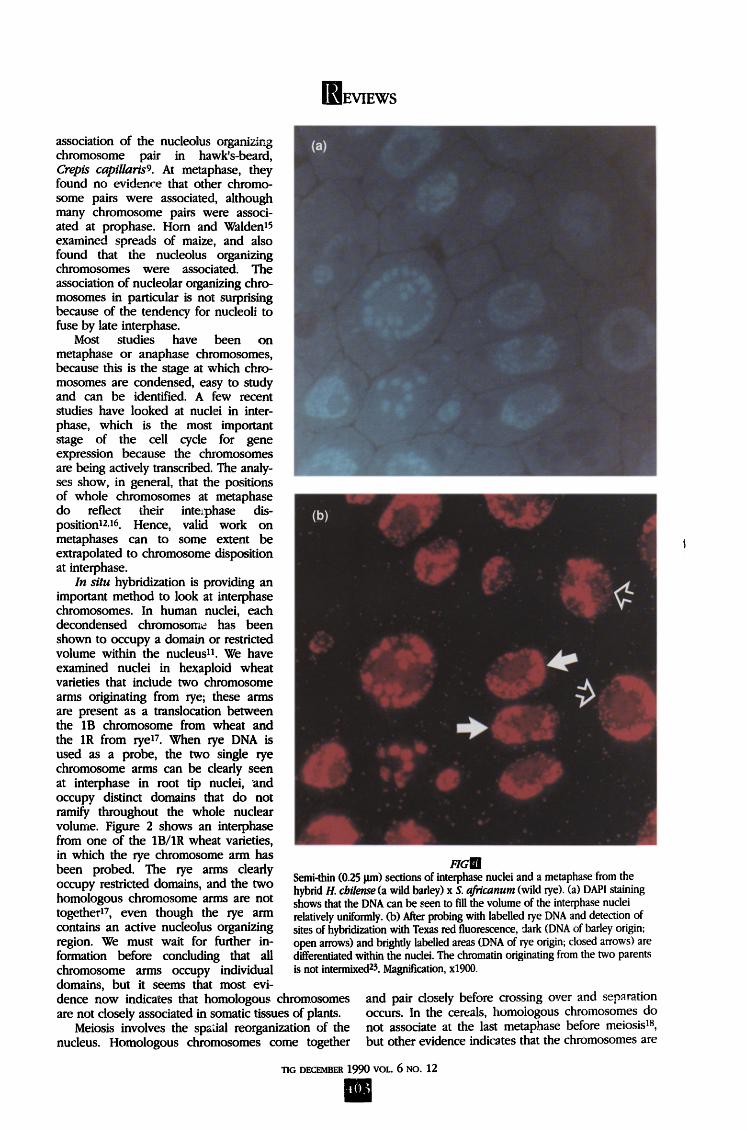
FIGIR Interphase nuclei from the intergeneric hybrid plant, Hordeum vulgare (barley) x Secale africanum (a wild rye). (a) After DAPl staining, no differentiation of chromatin within the nuclei is visible. (b) The nuclei after fn situ hybridization of labelled rye DNA and detection of hybridization sites by yellow fluorescence, counterstained with the DNA stain propidium iodide (fluorescing orange-red). The nuclei tend to show parental genuine separation with the zone of chromatin of rye origin (yellow) distinct from the chromatin of barley origin (orange-red)12. Magnification, x1170.
TIG DECEMBER 1990 VOL. 6 NO. 12
t()2
~ E V I E W S
association of the nucleolus organizing ' chromosome pair in hawk's-beard, Crepis capillaris9. At metaphase, they found no evidence that other chromo- some pairs were associated, although many chromosome pairs were associ- ated at prophase. Horn and Walden 1~ examined spreads of maize, and also found that the nucleolus organizing chromosomes were associated. The association of nucleolar organizing chro- mosomes in particular is not surprising because of the tendency for nucleoli to fuse by late interphase.
Most studies have been on metaphase or anaphase chromosomes, because this is the stage at which chro- mosomes are condensed, easy to study and can be identified. A few recent studies have looked at nuclei in inter- phase, which is the most important stage of the cell cycle for gene expression because the chromosomes are being actively transcribed. The analy- ses show, in general, that the positions of whole chromosomes at metaphase do reflect their inte~phase dis- positionlZ, 16. Hence, valid work on metaphases can to some extent be extrapolated to chromosome disposition at interphase.
In situ hybridization is providing an important method to look at interphase chromosomes. In human nuclei, each decondensed chromosome has been shown to occupy a domain or restricted volume within the nucleus n. We have examined nuclei in hexaploid wheat varieties that include two chromosome amls originating from rye; these arms are present as a translocation between the 1B chromosome from wheat and the 1R from rye ~7. When rye DNA is used as a probe, the two single rye chromosome arms can be clearly seen at interphase in root tip nuclei, "and occupy distinct domains that do not ramify throughout the whole nuclear volume. Figure 2 shows an interphase from one of the 1B/1R wheat varieties, in which the rye chromosome arm has been probed. The rye arms clearly occupy restricted domains, and the two homologous chromosome arms are not togetheri7, even though the rye arm contains an active nucleolus o~anizing region. We must wait for further in- formation before concluding that all chromosome arms occupy individual domains, but it seems that most evi- dence now indicates that homologous chromosomes are not closely associated in somatic tissues of plants.
Meiosis involves the spatial reorganization of the nucleus. Homologous chromosomes come together
F1al l Semi-thin (0.25 lira) sections of interphase nuclei and a metaphase from the hybrid H. cbilense (a wild barley) x S. africanum (wild rye). (a) DAPI staining shows that the DNA can be seen to fill the volume of the interphase nuclei relatively uniformly. (b) After probing with labelled rye DNA and detection of sites of hybridization with Texas red fluorescence, dark (DNA of barley origin; open arrows) and brighdy labelled areas (DNA of rye origin; closed arrows) are differentiated within the nuclei. The chromatin originating from the two parents is not intermixed z3. Magnification, x1900.
and pair closely before crossing over and separation occurs. In the cereals, homologous chromosomes do not associate at the last metaphase before meiosis TM, but other evidence indicates that the chromosomes are
TIG DECEMBER 1990 VOL. 6 NO. 12

EVIEWS
at ',east partially aligned by early leptotene6. Therefore, the last premeiotic interphase must be critical to the events of recombination, and the major spatial re- organization might explain, at least in part, its long duration.
The positions of genomes in hybrids Examination of metaphases from various species
often shows that chromosomes are grouped; Fig. 1 shows an example from Yucca. In spread preparations of various intergeneric hybrids from both cereal plants~9.20 and Nicotiana cell culture#], genomes orig- inating from the two parental chromosome sets are spatially separated.
Reconstructions of electron micrographs of serially sectioned cells confirmed the light microscope observations of spread metaphases]9,2a. However, reconstructions of conventionally stained thin sections do not allow ider~tification of individual chromo- somes at interphase. In situ hybridization enabled Schwarzacher et aDZ to show that the chromosomes of the two parental genomes tended to lie in distinct and separate domains throughout the cell cycle in spread preparations of an intergeneric cereal hybrid, Hordeum chilense x Secale africanum. Figure 3 shows micrographs from interphases of another hybrid, H. vulgare x S. africanum, which also tends to show genome separation at interphase. However, there is the possibility that artefacts from the spreading method, or the differential penetration of probe or detection reagents influence the observation. Leitch et al. 23 made semi-thin sections of the H. chilense hybrid, and probed the Secale genome, which was shown to occupy an independent domain from the Hordeum genome in the individual sections (Fig. 4). Thus, genome separation occurs throughout the cell cycle, and is a feature of the architecture of the interphase nucleus in these hybrids between widely diverged species.
How are such orders maintained? They are presum- ably under genetic control, and not merely a packing phenomenon, because smaller chromosomes may be central or peripheral in different hybrids2a. Even in a single hybrid, different parental genomes may lie in different domains in different tissuesa4. The work of Gleba et al.Z' showed that a spindle poison disrupted genome separation, which was restored after removal of the poison. That result implies self-maintenance of chromosome position, and perhaps that microtubule proteins are involved, but nuclear envelope attach- ments or the nuclear skeleton may also play a part in maintaining nuclear architecture.
The rehtive positions of individual chromosomes Whether chromosomes lie in nonrandom positions
with respect to each other and the nuclear envelope is a controversial issue in the study of nuclear architec- ture. Patterns of associations between nonhomologous chromosomes have been noted within metaphase spread# and between telomeres of chromosomes at prophase2~. Various authors have also noted regular patterns in the sizes and shapes of groups of chromo- somes, or in the distribution of repetitive sequences. Most plants have many hundreds of times the
minimum amount of DNA they require for encoding their genes. If there were no constraints on how the extra DNA were carried, the chromosomes would be free to alter in size and shape. However, most species have patterns in their karyotypes, both with respect to the DNA contents of individual chromosomes 26 and chromosome morphology. Shchapova 2~ found that, in various species, every long chromosome arm could be paired with a short chromosome arm such that the sum of the lengths of each pair in the species was a constant. Bennett 7 examined various cereal karyo- types, and noted another pattern: there were pairs of long and pairs of short chromosome arms, each of which tended to be similar in size. Schweizer and Loid128 observed that pairs of chromosome arms had similar C-band distribution. The data suggested to them that C-band patterns might have evolved via interchromosomal transfer of heterochromatic sequences between chromosome segments that are in close proximity.
These models enable the relative positions of chro- mosomes to be predicted, and preparations can be tested to see if the predictions are confirmed. Tests have been made using both metaphase spread prep- arations 4 and reconstructions of metaphasesa9, but statistical methods for analysing the results are ex- tremely difficult to devise.
The statistical analysis of order If order is strongly expressed and clear, then no
analysis is required; for example, Lin30 has shown that the 12 chromosomes of Rhoeo spathacea are arranged in a definite sequence in a ring at meiosis because of a particular pattern of reciprocal translocations between different chromosome arms. Beyond this, the methods of analysis are, almost without exception, contro- versial31, and usually developed to suit a particular situation.
Many factors make analysis difficult. The variability of chromosome position, at least at metaphase, is extremely high, both in spread and reconstructed cells. Cells have different shapes and sizes, and hence must be standardized. Chromosomes are physically con- strained within the nucleus, so their positions fit no standard distribution. And finally, the nonrandom polarity of chromatin within the nucleus of many cell type# z, with centromeres lying at one end and telo- meres at the opposite, affects chromosomes differen- tially.
Many analyses - for somatic association of homol- ogues, and association of heterologues, for example - involve the comparison of intercentromere distances. Without careful statistical evaluation, the distances can become dependent on each other because groups of separation distances are used 14. Tests for order within sets, particularly on small numbers of nuclei, tend to be extremely insensitive. Novel, particularly non- parametric and multivariate statistical approaches are likely to be required for the analysis and prediction of nuclear architecture, and some interesting approaches are now being developed33. These will minimize the assumptions being made about chromosome dis- position, and enable specific tests of models to be made.
TIG DECEMBER 1990 VOL. 6 NO. 12
t_{)t

[~qEVIEWS
Consequences of ordering in the nucleus Chromosome el imination
The loss of single chromosomes, or whole genomes, is a clear consequence of their positioning at a critical time. Their position within the cell at division excludes them from the daughter nuclei. In six differ- ent hybrids between various cereals that we have investigated 8, the chromosomes of the genome that is peripheral tend to be lost. While peripheral position- ing does not always lead to elimination of a genome's chromosomes, at least in cereal hybrids it does pre- dispose to this mitotic instability.
Gene expression The consequences of nuclear order for gene
expression are as yet unclear. Transformation exper- iments suggest that, in general, chromosomal position may have little or no effect on gene expression, but other observations have revealed at least some situations in which the nuclear position of a gene can influence its expression. In plants, there is only one set of genes that can be easily visualized by microscopy when they ace being expressed - the nucleolar organizing genes that are transcribed within, and give rise to, the nucleolus, in root tips, the nu- cleolus is normally central within the interphase nucleus. In the cereal hybrids that show strong parental genome separation, with one genome around the other, the nucleolar organizing genes in the peripheral genome (for example the rye nucleolar organizing genes in Figs 3 and 4) are not expressed, but the nucleolar organizing genes from the central genome, which surrounds the nucleolus, are active8, 29. In ad- dition, many hybrid plants show a strong tendency to resemble one parent much more than the other in their gross appearance and behaviour. Where the hybrid shows strong concentric genome separation, the hybrid resembles the parent contributing the outer genome s. Thus the intranuclear position of a parental genome at interphase may affect the extent to which its genes are expressed.
Conclusions Interest in the architecture and organization of
interphase nuclei and metaphases is increasing. Whether ordering has implications for plants beyond those discussed above (including, perhaps, genomic imprinting) is not yet known. However, in human nuclei, Borden and Manuelidis34 have shown that the relative position of the X chromosome alters in patients suffering from epilepsy - an important dis- covery indicating that order may directly correlate with cell and organism behaviour. The potential importance o f chromosome position, because of its effect on gene expression, is being recognized in plants, although further studies are required.
Technical advances in fluorescent light microscopy, confocal microscopy, and in situ hybridization that have been made within the past three years are now enabling us to attack the problems of nuclear architec- ture directly. For the first time, we can study the dis- position of whole genomes, chromosomes, repetitive DNA sequences and genes within active, and even dif- ferentiated, interphase nuclei.
Acknowledgements We thank BP Venture Research Unit for enabling our
work on the architecture of the nucleus. We also thank our collaborators, and in particular Drs Andrew Leitch and Trude Schwarzacher, for their contributions to our research.
References 1 MOiler, C. (1909)Jahrb. Wiss. Bot. 47, 99-117 2 Darlington, C.D. (1937) Recent Advances in Cytology
(2nd edn) J.&A. Churchill 3 Avivi, L. and Feldman, M. (1980) Hum. Genet. 55,
281-295 4 Avivi, L., Feldman, M. and Brown, M. (1982)
Chromosoma 86, 1-16 5 Heslop-Harrison, J.S., Chapm~n, V. and Bennett, M.D.
(1985) Heredity 55, 93-103 6; von Wettstein, D., Rasmussen, S.W. and Holm, P.B.
(1984) Annu. Rev. Genet. 18, 331-413 7 Bennett, M.D. (1982) in Genome Evolution (Dover, G.A.
and Flavell, R.B., eds), pp, _239--261, Academic Press 8 Bennett, M.D. (1984) in Gene Manipulation in Plant
Improvement (16th Stadler Genetics Symposium) (Gustafson, J.P., ed.), pp. 469--502, Plenum Press
9 0 u d , J.L. et al. (1989) J. Cell Sci. 92, 329-339 10 Rawlins, D.J. and Shaw, P.J. (1990) Chromosoma 99,
143-155 11 Lichter, P. et al. (1988) Hum. Genet. 80, 224-234 12 Schwarzacher, T., Leitch, A.R., Bennett, M.D. and Heslop-
Harrison, J.S. (1989) Ann. Bot. 64, 315-324 13 Metz, C.W. (1916)J. Exp. Zool. 21,213-279 14 Heslop-Harrison, J.S., Smith, J.B. and Bennett, M.D.
(1988) Chromosoma96, 119-131 15 Hom, J.D. and Walden, D.B. (1978) Genetics88, 181-200 16 Cremer, T. etal. (1982) Hum. Genet. 62, 201-209 17 Heslop-Harrison, J.S., Schwarzacher, T. and Leitch, A.R.
(1990) Heredity65, 385-392 18 Bennett, M.D. (1984) Symp. Soc. Exp. Biol. 38, 87-121 19 Finch, R.A., Smith, J.B. and Bennett, M.D. (1981)J. Cell
Sci. 52, 391-403 20 Linde-Laursen, I. and von Bothmer, R. (1988) Theor
Avpl. Genet. 76, 897-908 21 Gleba, Y.Y. et al. (1987) Proc. NatlAcad. Sci USA 84,
3709-3713 22 Schwarzacher-Robinson, T., Finch, R.A., Smiih, J.B. and
Bennett, M.D. (1987)J. (,'ell Sci. 87, 291-304 23 Leitch, A.R. et al. (1990) J. Cell Sci. 95, 335-341 24 Finch, R.A. (1983) Chromosoma 88, 386-393 25 Ashley, T. (i979)./. Cell Sci. 38, 357-367 26 Narayan, R.K.J. and Durrant, A. (1983) Genetica 61, 47-53 27 Shchapova, A.I. (1969) Tsitologia 13, 1157-1164 [with
English summary; NB for 'different' read 'equal'] 28 Schweizer, D. and Loidl, J. (1987) Chromosomes Today 9,
61-74 29 Heslop-Harrison, J.S. and Bennett, M.D. (1984)J.
Embryol. Exp. morphol. 83 (Suppl.), 51-73 30 l~,n, Y.J. (1979) Chromosoma 71, 109-127 3 / Callow, R.S. (i985) Heredity 54, 171-177 32 Rabl, C. (1885) Morphol.Jahrb. 10, 214-330 33 Dorninger, D. and Timischl, W. (1987) Heredity 58,
321-325 34 Borden, J. and Manuelidis, L. (1988) Science 242,
1687-1691
~.£ HESLoP-HaRRlSON IS IN THE KARYOBIOLOGY GROUP, J l I ]CENTRE FOR PLANT SC~NC~ RESF~.RCH, COIaEY LANai [ N o - - c a NR4 7UJ, UK a~vD M.D. BENNEIT IS IN THE] IJcoRF~ LamgaoRg, R o z ~ BOT~VlC GatO~N~ K~v, I ~ R~CH~ON~ TW9 3 ~ V~ I
"rig O~CEMBER 1990 VOW.. 6 NO. 12
Recommended

















