
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS
Vol. 348, No. 1, December 1, pp. 107–115, 1997Article No. BB970378
Reconstitution Premixes for Assays Using PurifiedRecombinant Human Cytochrome P450, NADPH-Cytochrome P450 Reductase, and Cytochrome b5
Peter M. Shaw,*,1 Natilie A. Hosea,† David V. Thompson,*Janean M. Lenius,* and F. Peter Guengerich†,1
*PanVera Corporation, 545 Science Drive, Madison, Wisconsin 53711; and †Department of Biochemistryand Center in Molecular Toxicology, Vanderbilt University School of Medicine, Nashville, Tennesee 37232-0146
Received June 17, 1997, and in revised form August 18, 1997
zymes and the development of the P450 premixes thatremain active after being stored frozen should allowThe development of enzyme and buffer premixes forfor rapid identification of novel P450 substrates andin vitro biotransformation assays is described. Theinhibitors and the development of large-scale screen-protein premixes contain a mixture of three recombi-ing assays. q 1997 Academic Pressnant human proteins, cytochrome P450 (P450) 3A4,
Key Words: cytochrome P450; reconstitution; recom-NADPH-P450 reductase, cytochrome b5 , and lipo-binant; premixes; oxidation.somes. The buffer premix contains reagents which,
when diluted, provide for optimal metabolic activitywith selected P450 3A4 substrates. P450 3A4 premixeswere competent in the oxidation of known substrates
Cytochrome P450 (P450 and/or CYP)2 enzymes (ECincluding testosterone, midazolam, nifedipine, eryth-1.14.14.1) comprise a large family of enzymes involvedromycin, benzphetamine, and amitriptyline. Premixesin the oxidative biotransformation of a wide variety ofstored at0807C for 2 months and those that underwentorganic molecules which include endogenous as well asan additional five freeze/thaw cycles were able to hy-
droxylate testosterone at turnover rates similar to xenobiotic compounds e.g., steroids, ecosanoids, aro-freshly prepared reconstitution mixes. In addition, matic hydrocarbons, pesticides, and drugs (1, 2). In thepremixes stored unfrozen at 47C for 2 weeks showed liver, P450 enzymes are located in the endoplasmic re-no significant loss in the rate of testosterone 6b-hy- ticulum of hepatocytes along with NADPH-P450 reduc-droxylation by P450 3A4. Premixes prepared with and tase (EC 1.6.2.4), which donates electrons for catalysis,without reduced glutathione, a component which had and cytochrome b5 (EC 4.4.2 group), which can modu-previously been found to be important for P450 3A4 late P450 catalysis in vitro, apparantly with or withoutreactions, were equally efficient at carrying out testos- electron transfer in various situations (3).terone hydroxylation under these conditions. Kinetic Currently, P450 enzymes representing five gene fam-parameters determined for the metabolism of testos- ilies (4) are known to be expressed to some extent interone, amitriptyline, nifedipine, and benzphetamine human adult liver: CYP1A2, CYP2A6, CYP2B6, CYP-using P450 3A4 premixes were compared with human 2C8, CYP2C9, CYP2C18. CYP2C19, CYP2D6, CYP-pooled microsomes and insect microsomes prepared 2E1, CYP3A4, CYP3A5, CYP4A9, CYP4A11, andfrom cells infected with a baculovirus containing two
CYP7. Both genetic and environmental factors play acDNA inserts coding for P450 3A4 and NADPH-P450role in the types of allelic variants and levels of individ-reductase. Each format gave different Vmax and Km val-ual enzymes observed in different human (ethnic) pop-ues indicating different catalytic efficiencies. Analysisulations and individuals within these groups. It hasof P450 1A2 premixes which contained different lipid
concentrations indicated that Vmax and Km could be al-tered. The availability of human P450 recombinant en- 2 Abbreviations used: CYP and P450, cytochrome P450; HPLC,
high-pressure liquid chromatography; GSH, reduced glutathione;Hepes; 4-(2-hydroxy)-1-piperazineethanesulfonic acid; Chaps, 3-[(3-
1 To whom correspondence should be addressed. Fax: (608) 233 cholamidopropyl)dimethylammonio]-1-propanesulfonate; BCA, bi-cinchoninic acid.3007. E-mail: [email protected].
1070003-9861/97 $25.00Copyright q 1997 by Academic PressAll rights of reproduction in any form reserved.
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca

108 SHAW ET AL.
been well documented that the type and levels of these ment of novel P450 3A4 premixes that can be storedand used conveniently when they are thawed. The ad-hepatic isozymes expressed by an individual can have
a major impact on the detoxification of xenobiotics, the vantage of these premixes is that they overcome boththe perception and the technical barriers associatedactivation of carcinogens, and the metabolism of a wide
variety of pharmacologically important drugs (1, 2). with performing large numbers of P450 assays usingpurified components in a reliable fashion.Members of the CYP3A subfamily, which represent ap-
proximately 30% and sometimes as much as 60% of thetotal expressed P450 in adult human liver, are known MATERIALS AND METHODSto be involved in the metabolism of numerous com-
Protein Expression and Purificationpounds including erythromycin, nifedipine, testoster-Plasmids containing modified cDNAs encoding P450 3A4 (8) orone, cyclosporine, lidocaine, imipramine, quinidine,
human cytochrome b5 (18) were transformed into Escherichia colimidazolam, verapamil, troleandomycin, and terfe-and grown in liquid expression medium. Cells were harvested andnadine (1, 2). recombinant proteins were purified with minor modification of estab-
One approach that we have used to better under- lished procedures for P450 3A4 (8, 9) and human cytochrome b5 (18).stand and characterize the molecular interactions and Human NADPH-P450 reductase was purified from insect micro-
somes infected with a baculovirus that contained a human cDNAsubstrate specificity of CYP3A members is by purifica-clone (19), as previously described (6). Rat reductase was purifiedtion of the native (5–7) and, more recently, the bacteri-from E. coli with minor modification of a method previously outlinedally expressed recombinant isozymes (8, 9). Substrate (20).
specificity of the purified enzymes is determined byreconstitution of enzyme activity. In vitro reconstitu- Reconstitution and Biotransformation Assaystion assays require combining multiple components in-
Simplified premix system. A batch of the 51 P450 3A4 proteincluding purified P450, NADPH-P450 reductase, lipidspremix (0.5 mM P450 3A4, 1.0 mM NADPH-P450 reductase, 0.5 mMthat have been sonicated to make liposomes, detergent, cytochrome b5 , 0.5 mg Chaps/ml, 0.1 mg/ml liposomes [L-a-dilauroyl-
and buffer additives. Many techniques have been devel- sn-glycero-3-phosphocholine, L-a-diloleoyl-sn-glycero-3-phosphocho-oped that use different methods to combine the compo- line, L-a-dilauroyl-sn-glycero-3-phosphoserine (1:1:1, w/w/w per mil-
liliter)], 3.0 mM GSH, and 50 mM potassium Hepes, pH 7.4) and anents to form a functional enzyme complex, which is51 buffer mix (200 mM potassium Hepes, pH 7.4, 12 mM GSH, andthen used for biotransformation assays (10–14). Recon-150 mM MgCl2) were prepared and frozen at 080 and 0207C in 1.0-stitution of P450 3A4 appears to be more complex than and 1.5-ml aliquots. The 51 protein premixes were made as follows
for other P450s. For example, reasonable turnovers for and are based on optimized conditions previously published (8, 9,a variety of P450 2E1 and CP450 1A2 substrates can be 12). Liposomes were prepared as previously described, in the absence
of Ar (9). The purified proteins and liposomes were mixed togetherachieved in reconstitution systems containing a singleon ice; then Hepes, GSH, and CHAPS were added, with water usedlipid, L-a-dilauroyl-sn-glycero-3-phosphocholine (15,to adjust the components to the final concentration as described16). However, optimal activity for P450 3A4 substrates above. P450 1A2 mixes were made as described above with 0.5 mM
such as testosterone and nifedipine, but not others such P450 1A2 and 0.25 mM NADPH-P450 reductase, and without cyto-chrome b5 .as erythromycin and benzphetamine, is affected by the
A typical 300-ml biotransformation reaction was prepared by mix-composition of the liposomes, the concentration of diva-ing (on ice) 60 ml of 51 P450 3A4 protein premix and an equal volumelent cations, the presence of GSH, the buffer type, andof 51 buffer mix. H2O and substrate were then added to bring the
the ratio of the other protein components (8, 9, 12, 13). reaction almost to the working 11 concentration. The subsequentFor many researchers, the reconstitution process is addition of 6 ml of 100 mM NADPH yielded the exact 11 concentra-
tion. Some experiments, indicated in the legends, used an NADPH-considered tedious, time consuming, and prone to error.generating system composed of 0.5 mM NADP/, 2 units of glucoseThis is due, in part, to the multiple pipetting required6-phosphate dehydrogenase/ml, and 10 mM glucose 6-phosphate. Un-for each of the various stages (combining all the compo- less otherwise stated (in other reactions where the final reaction
nents for reconstitution, preincubations for complex volume was different), the concentration of the premixes remainedformation, and the subsequent biotransformation reac- as described above.tions) and because classically all these stages needed Fresh standard mix. The individual protein and buffer compo-
nents were aliquoted from stock solutions into 1.5-ml microcentrifugeto be performed on the same day due to lack of informa-tubes to achieve approximately 2.51 solutions and incubated on icetion on stability of the protein components. Histori-for 10 min. H2O and substrate were added and the mixtures werecally, microsomal P450s were never frozen in the ab- preincubated at 377C for 3 min, followed by addition of NADPH to
sence of glycerol or other polyols because of early evi- start the biotransformation reactions. The addition of substrate anddence showing the protective action of these agents NADPH adjusted the components to the same concentrations as for
the simplified mixtures.(17). In early work, a protective effect of GSH was re-ported (17), and thiols are still used in purification andstorage of P450s. Analytical Assays
In an effort to make the whole process simpler and The assay conditions varied depending on the substrate tested.more amenable for others, we have abandoned the tra- Most of the assays were modified so that reactions could be performed
in 1.5-ml microfuge tubes.ditional methodology and describe here the develop-
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca

109CYTOCHROME P450 RECONSTITUTED PREMIXES
In the testosterone 6b-hydroxylation assay, the final concentrationof testosterone was 200 mM unless otherwise indicated. The reactionswere stopped after 10 min at 377C either by addition of 90 ml of 0.66mM 11b-hydroxytestosterone, dissolved in a 2:1 mixture of CH3CNand CH3OH, or with 0.2 vol of a mixture of 1 M Na2CO3 and 2 MNaCl (pH 10.5). HPLC analysis of the products after extraction with1 ml of CH2Cl2, was performed as previously described (5).
Midazolam 1- and 4-hydroxylation assays were performed as de-scribed previously (21). Nifedipine assays were performed as de-scribed previously (5).
Benzphetamine N-demethylation was measured following the re-action in the presence of 1.0 mM d-benzphetamine. Reactions (0.5ml) were quenched by the addition of 0.2 vol of a mixture of 1 MNa2CO3 and 2 M NaCl (pH 10.5) and extracted with 2 vol of CH2Cl2.The contents of the tubes were mixed using a vortex device and thelayers were separated by centrifugation (450g for 10 min). An aliquotof the organic (lower) layer was transferred to a glass tube and driedunder N2. The residue was dissolved in 100 ml of column mobilephase and the substrate and product (N-benzylamphetamine) wereseparated on a Develosil ODS-HG-5 HPLC column using a mixture of10 mM potassium phosphate (pH 4.0):CH3CN (35:65, v/v) containing0.02% (v/v) (C2H5)3N. The flow rate was 1.5 ml per minute and detec-tion was at 254 nm.
Amitriptyline N-demethylation assays were done as described else- FIG. 1. Sodium dodecyl sulfate–polyacrylamide gel electrophoresiswhere (22). Erythromycin metabolism was determined by measuring analysis of purified proteins. Each protein (2 mg) was loaded onto aformaldehyde production, using the Nash procedure as previously 4–20% acrylamide preprepared gradient Novex gel (San Diego, CA)described (23, 24). Methoxyresorufin O-dealkylation was measured and electrophoresed under denaturing conditions according to theas previously described (5) with slight modification. Reactions were manufacturer’s instructions. Lanes 1 and 6, Mr markers (150, 100,perfomed with 2.5 pmol of P450 1A2 premix, in 120 mM sodium/ 75, 50, 35, 25, and 15 kDa); Lane 2, cytochrome b5 , Lane 3, P450potassium phosphate buffer, pH 7.7. NADPH (final concentration 3A4; Lane 4, rat NADPH-P450 reductase, Lane 5, human NADPH-5 mM) was used to start the reaction. Fluorescence increase was P450 reductase.measured on a Beacon fluoresence polarization instrument (PanVeraCorp., Madison WI) in intensity mode with excitation and emissionfilters at 530 and 580 nm, respectively. Microsomal reactions wereperformed, as above, with 2 mg of protein. The purification of the components used for reconsti-
Protein concentration was estimated using the bicinchoninic acid tution experiments followed standard, well-docu-(BCA) assay (Pierce, Rockford, IL). P450 and cytochrome b5 concen-mented procedures and the purity of the proteins wastrations and NADPH-P450 reduction activity were measured as pre-
viously described (6) using extinction coefficients of 91 mM01 ú90% as demonstrated using sodium dodecyl sulfate–cm01(De450–490 for Fe2/CO vs Fe2/), 100 mM01 cm01 (De425 for Fe2/ polyacrylamide gel electrophoresis as a criterion (Fig.vs Fe3/), and 21 mM01 cm01(De550 for reduced cytochrome c), respec- 1). The specific content or activity of each of the threetively.
proteins, shown in Table I, was in good agreement withpreviously published results (8, 18, 20, 27).
ReagentsHuman pooled microsomes were purchased from XenoTech L.L.C. Stability
(Kansas City, KS). Midazolam and the 1- and 4-hydroxy metaboliteswere a kind gift from Hoffman–La Roche (Nutley, NJ). P450 3A4 Our rationale for deciding how to design the proteinBaculosomes were prepared at PanVera Corporation. Briefly, Tri- premixes was defined by several criteria that had to bechoplusia ni cells were infected with a baculovirus containing cDNA met to allow flexibility for P450 3A4 biotransformationinserts for human CYP3A4 and rabbit NADPH-P450 reductase as
reactions: (i) Different final concentrations of P450previously descibed (25). The infected cells were harvested by centrif-needed to be achieved in the reaction mix, typicallyugation and microsomes prepared using a procedure described else-
where (26). All other reagents were obtained from commercial between 10 and 200 pmol P450 per milliliter, to accom-sources and were analytical grade or better. modate the expected variation in turnover rates for
different substrates. (ii) Most P450 substrates are hy-drophobic and have to be dissolved in either an organicRESULTS AND DISCUSSIONsolvent (which should make up õ2% of the total reac-Purification tion volume to prevent interference in oxidation reac-tions) or a relatively large amount of aqueous solution.We focused our initial studies on P450 3A4 because
of its importance in drug metabolism and because re- Therefore, the buffer and protein premixes wouldpreferably make up 50% or less of the total reactionconstitution of this P450 is more complex than others.
We reasoned that if we succeeded in developing a sim- volume. (iii) As in (ii), sufficient volume should be left toallow for addition of NADPH or an NADPH-generatingplified system for P450 3A4, then other P450s could
easily be substituted into a similar system. system.
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca
110 SHAW ET AL.
TABLE IITABLE I
Stability of Premixes after Storage at 0807CaPurified Proteins Used in Reconstitution Assaysa
Specific content or Rate of 6b-hydroxytestosteroneformationPurified protein specific activity
(nmol/min/nmol of CYP3A4)P450 3A4 11 nmol/mg proteinP450 1A2 16 nmol/mg protein Format Expt 1 Expt 2Cytochrome b5 60 nmol/mg proteinNADPH-P450 reductase (rat) 55 mmol/min/mg protein Prepared on the same day 26.0 { 1.6 (n Å 5) 26.8 { 1.7 (n Å 5)NADPH-P450 reductase (human) 24 mmol/min/mg protein Two months storage 30.1 { 0.4 (n Å 4) 38.0 { 2.5 (n Å 5)
Two months storage /five freeze/thaws 27.6 { 3.3 (n Å 5) 37.3 { 4.3 (n Å 5)a Protein concentration, hemoprotein concentration, and NADPH-
cytochrome c reduction activity were measured as described underMaterials and Methods. a The mean turnover number, standard deviation, and number of
independent testosterone reactions performed from a standard re-constitution mix made on the day, and from a single set of 5X P4503A4 protein and buffer premixes that had undergone zero and fivePrevious studies had shown that optimal P450 3A4further successive freeze/thaw cycles are shown. Reconstitution con-biotransformation reactions required the inclusion of 3 ditions were performed as described under Materials and Methods.
mM GSH and 30 mM MgCl2 (8, 13). We therefore optedto make two premixes at a 51 concentration that wouldoccupy 40% of the total reaction volume, leaving 60%
least 2 weeks, as shown in Figs. 2 and 3. In addition,for the addition of substrate and electron donor. Thethe absence of GSH in the premixes had an insignifi-buffer premix contained high concentrations of MgCl2,cant effect on testosterone hydroxylation activity forGSH, and Hepes buffer that, when diluted fivefold,the time period tested, 1 week. The results from thewould give the previously defined optimal componentvarious stability tests at different temperatures dem-concentrations (9). Similarly, the 51 protein premixonstrated the feasibility of using premixes of P450 andcontained 0.5 mM P450 3A4 at a 1:2:1 molar ratio ofassociated electron transfer proteins for biotransforma-P450 3A4 to NADPH-P450 reductase to cytochrome b5 .tion reactions. These results also demonstrated that itThe ratio of the proteins was chosen based on previouswas possible to make up large batches of premixes withexperiments that had proven optimal for several P450recombinant proteins to generate a single batch of pre-3A4 biotransformation reactions and the concentrationmix that could be used for multiple reactions, thusof the P450 after dilution to 11 would be 100 nM.
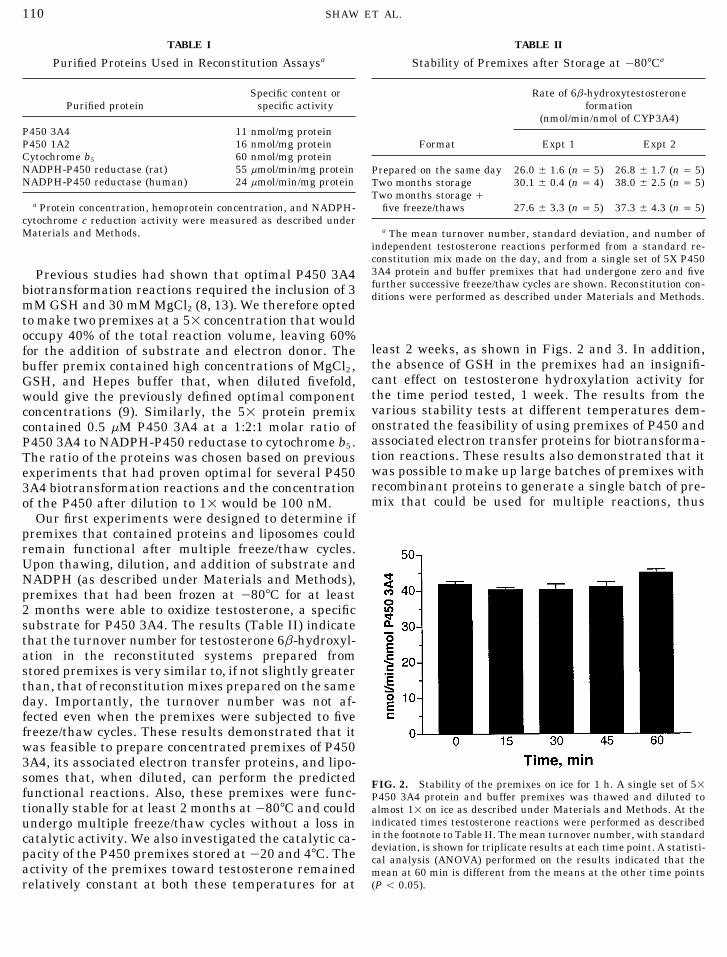
Our first experiments were designed to determine ifpremixes that contained proteins and liposomes couldremain functional after multiple freeze/thaw cycles.Upon thawing, dilution, and addition of substrate andNADPH (as described under Materials and Methods),premixes that had been frozen at 0807C for at least2 months were able to oxidize testosterone, a specificsubstrate for P450 3A4. The results (Table II) indicatethat the turnover number for testosterone 6b-hydroxyl-ation in the reconstituted systems prepared fromstored premixes is very similar to, if not slightly greaterthan, that of reconstitution mixes prepared on the sameday. Importantly, the turnover number was not af-fected even when the premixes were subjected to fivefreeze/thaw cycles. These results demonstrated that itwas feasible to prepare concentrated premixes of P4503A4, its associated electron transfer proteins, and lipo-somes that, when diluted, can perform the predicted FIG. 2. Stability of the premixes on ice for 1 h. A single set of 51functional reactions. Also, these premixes were func- P450 3A4 protein and buffer premixes was thawed and diluted totionally stable for at least 2 months at0807C and could almost 11 on ice as described under Materials and Methods. At the
indicated times testosterone reactions were performed as describedundergo multiple freeze/thaw cycles without a loss inin the footnote to Table II. The mean turnover number, with standardcatalytic activity. We also investigated the catalytic ca-deviation, is shown for triplicate results at each time point. A statisti-pacity of the P450 premixes stored at 020 and 47C. The cal analysis (ANOVA) performed on the results indicated that the
activity of the premixes toward testosterone remained mean at 60 min is different from the means at the other time points(P õ 0.05).relatively constant at both these temperatures for at
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca
111CYTOCHROME P450 RECONSTITUTED PREMIXES
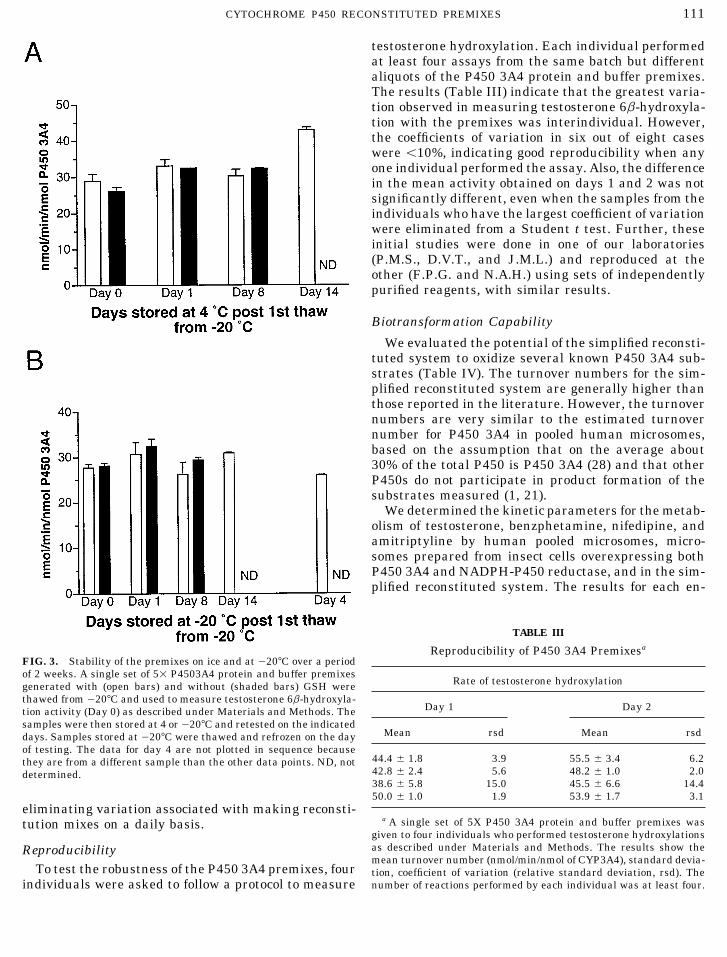
testosterone hydroxylation. Each individual performedat least four assays from the same batch but differentaliquots of the P450 3A4 protein and buffer premixes.The results (Table III) indicate that the greatest varia-tion observed in measuring testosterone 6b-hydroxyla-tion with the premixes was interindividual. However,the coefficients of variation in six out of eight caseswere õ10%, indicating good reproducibility when anyone individual performed the assay. Also, the differencein the mean activity obtained on days 1 and 2 was notsignificantly different, even when the samples from theindividuals who have the largest coefficient of variationwere eliminated from a Student t test. Further, theseinitial studies were done in one of our laboratories(P.M.S., D.V.T., and J.M.L.) and reproduced at theother (F.P.G. and N.A.H.) using sets of independentlypurified reagents, with similar results.
Biotransformation Capability
We evaluated the potential of the simplified reconsti-tuted system to oxidize several known P450 3A4 sub-strates (Table IV). The turnover numbers for the sim-plified reconstituted system are generally higher thanthose reported in the literature. However, the turnovernumbers are very similar to the estimated turnovernumber for P450 3A4 in pooled human microsomes,based on the assumption that on the average about30% of the total P450 is P450 3A4 (28) and that otherP450s do not participate in product formation of thesubstrates measured (1, 21).
We determined the kinetic parameters for the metab-olism of testosterone, benzphetamine, nifedipine, andamitriptyline by human pooled microsomes, micro-somes prepared from insect cells overexpressing bothP450 3A4 and NADPH-P450 reductase, and in the sim-plified reconstituted system. The results for each en-
TABLE III
Reproducibility of P450 3A4 Premixesa
FIG. 3. Stability of the premixes on ice and at 0207C over a periodof 2 weeks. A single set of 51 P4503A4 protein and buffer premixes
Rate of testosterone hydroxylationgenerated with (open bars) and without (shaded bars) GSH werethawed from 0207C and used to measure testosterone 6b-hydroxyla-
Day 1 Day 2tion activity (Day 0) as described under Materials and Methods. Thesamples were then stored at 4 or0207C and retested on the indicated
Mean rsd Mean rsddays. Samples stored at 0207C were thawed and refrozen on the dayof testing. The data for day 4 are not plotted in sequence because
44.4 { 1.8 3.9 55.5 { 3.4 6.2they are from a different sample than the other data points. ND, not42.8 { 2.4 5.6 48.2 { 1.0 2.0determined.38.6 { 5.8 15.0 45.5 { 6.6 14.450.0 { 1.0 1.9 53.9 { 1.7 3.1
eliminating variation associated with making reconsti-a A single set of 5X P450 3A4 protein and buffer premixes wastution mixes on a daily basis.
given to four individuals who performed testosterone hydroxylationsas described under Materials and Methods. The results show theReproducibilitymean turnover number (nmol/min/nmol of CYP3A4), standard devia-
To test the robustness of the P450 3A4 premixes, four tion, coefficient of variation (relative standard deviation, rsd). Thenumber of reactions performed by each individual was at least four.individuals were asked to follow a protocol to measure
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca

112 SHAW ET AL.
TABLE IV
Turnover Numbers of Various Probe SubstratesUsing the P450 3A4 Premixesa
RateActivity measured (nmol/min/nmol of CYP3A4)
Testosterone 6b-hydroxylation 43.9Midazolam 1-hydroxylation 9.0Midazolam 4-hydroxylation 9.0Erythromycin N-demethylation 8.8Benzphetamine N-demethylation 4.8Nifedipine oxidation 33.0
a Reconstitution reactions were performed as described under Ma-terials and Methods, except for N-demethylations, which were per-formed using a regenerating system and premixes that contained a2X P450 3A4 protein premix. The final concentration of each sub-strate was 200 mM testosterone, 200 mM nifedipine, 300 mM midazo-lam, 10 mM erythromycin, and 2 mM benzphetamine. The resultsshown are the average of at least duplicate reactions.
zyme system were analyzed using nonlinear regression(Fig. 4) which yielded estimates of Vmax
3 and Km values(Table V). The kinetic parameters for any one substratewere different in each of the P450 systems. The insectcell microsomes were observed to have the highest Vmax
values in all cases. However, comparison of the threesystems indicated that the differences in Vmax valueswere not as great for amitriptyline and nifedipine asthey were for benzphetamine and testosterone; the Vmax
values for pooled human microsomes and the premixeswere within a factor of two. The apparent Km valuesfor the reconstituted system were higher than those forthe pooled microsomes although the differences in theKm values were dependent on the substrate examined.The insect microsomes were observed to have the low-est apparent Km values for testosterone, nifedipine, andamitriptyline but not benzphetamine. Interestingly,sigmoidal profiles, suggestive of cooperativity, were ob-served in some instances (indicated with Hill coeffi-cients in Table V), as seen previously (22). The differ-ences in the kinetic parameters in each system for anyone substrate may result, in part, from differences inelectron transfer capacity to the P450 (13). This maynot be surprising when one considers the differentphysical environments in which the P450 is located andalso the estimated ratio of NADPH-P450 reductase toP450 3A4 in each of the systems, approximatly 2:1,0.2:1 and 8:1 for the reconstituted, human pooled, andinsect microsomes, respectively; the higher levels ofNADPH-P450 reductase probably explain the largerVmax values in baculosomes. In addition, another factor
3 We report Vmax values in nanomoles of substrate converted perminute per nanomole of P450, because it makes comparison betweenthe purified system and both insect and human microsomes more
TA
BL
EV
Kin
etic
Par
amet
ers
for
CY
P3A
4M
etab
olis
ma
Tes
tost
eron
eA
mit
ript
ylin
eB
enzp
het
amin
eN
ifed
ipin
e
P45
0sy
stem
Vm
axK
mV
max
/Km
nV
max
Km
Vm
ax/K
mn
Vm
axK
mV
max
/Km
nV
max
Km
Vm
ax/K
mn
Hu
man
mic
roso
mes
46{
246{
81
1.6{
0.2
7.7{
0.3
100{
100.
077
1.9{
0.2
19{
135{
170.
54n
/a32{
223{
61.
4n
/aR
econ
stit
ute
d47{
212
0{
200.
4n
/a9.
3{
1.0
490{
800.
019
1.5{
0.1
12{
244
00{
1300
0.00
27n
/a35{
137{
30.
951.
4{
3B
acu
loso
mes
150{
1025{
66
1.7{
0.2
8.5{
0.1
56{
30.
15n
/a84{
1321
00{
700
0.04
0n
/a45{
214{
43.
22.
0{
0.4
aT
he
kin
etic
para
met
ers
wer
eca
lcu
late
dfr
omth
efi
tted
curv
esin
Fig
.4
usi
ng
the
com
pute
rpr
ogra
mG
raph
Pad
PR
ISM
(San
Die
go,
CA
).V
max
valu
esar
ein
nm
ol/m
in/
nm
olof
CY
P3A
4;K
mva
lues
are
inm
M.
Th
eda
tare
flec
tsth
ebe
stfi
t(j
udg
edby
the
Pva
lue)
acco
rdin
gto
aco
mpa
riso
nof
aM
ich
aeli
s–M
ente
nan
dsi
gmoi
dal
equ
atio
ns.
Wh
ere
n/a
isdi
spla
yed,
the
best
fit
was
toa
Mic
hae
lis–
Men
ten
curv
e.In
the
case
sof
abe
stfi
tto
asi
gmoi
dal
curv
eth
eH
ill
coef
fici
ent
(n)
was
indi
cate
d.T
he
equ
atio
ns
use
dw
ere:
Mic
hae
lis-
Men
ten
,vÅ
(Vm
axS
)/(K
m/
S);
sigm
oida
l,vÅ
(Vm
axS
n)/
(Kn m/
Sn).
relevant.
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca
113CYTOCHROME P450 RECONSTITUTED PREMIXES
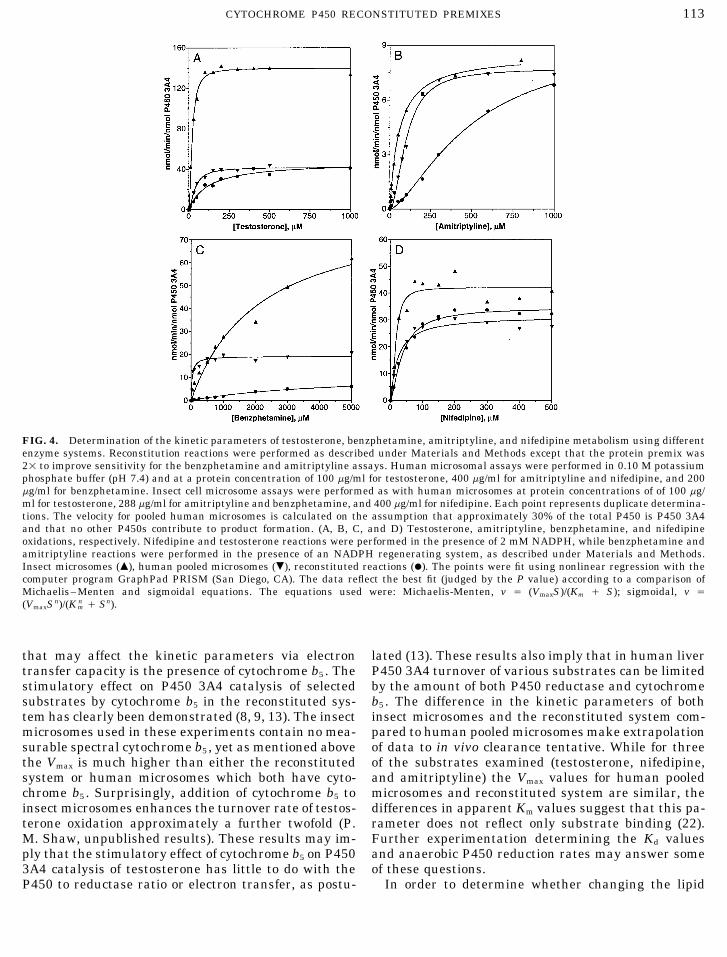
FIG. 4. Determination of the kinetic parameters of testosterone, benzphetamine, amitriptyline, and nifedipine metabolism using differentenzyme systems. Reconstitution reactions were performed as described under Materials and Methods except that the protein premix was21 to improve sensitivity for the benzphetamine and amitriptyline assays. Human microsomal assays were performed in 0.10 M potassiumphosphate buffer (pH 7.4) and at a protein concentration of 100 mg/ml for testosterone, 400 mg/ml for amitriptyline and nifedipine, and 200mg/ml for benzphetamine. Insect cell microsome assays were performed as with human microsomes at protein concentrations of of 100 mg/ml for testosterone, 288 mg/ml for amitriptyline and benzphetamine, and 400 mg/ml for nifedipine. Each point represents duplicate determina-tions. The velocity for pooled human microsomes is calculated on the assumption that approximately 30% of the total P450 is P450 3A4and that no other P450s contribute to product formation. (A, B, C, and D) Testosterone, amitriptyline, benzphetamine, and nifedipineoxidations, respectively. Nifedipine and testosterone reactions were performed in the presence of 2 mM NADPH, while benzphetamine andamitriptyline reactions were performed in the presence of an NADPH regenerating system, as described under Materials and Methods.Insect microsomes (m), human pooled microsomes (.), reconstituted reactions (l). The points were fit using nonlinear regression with thecomputer program GraphPad PRISM (San Diego, CA). The data reflect the best fit (judged by the P value) according to a comparison ofMichaelis–Menten and sigmoidal equations. The equations used were: Michaelis-Menten, v Å (VmaxS)/(Km / S); sigmoidal, v Å(VmaxSn)/(K n
m / Sn).
that may affect the kinetic parameters via electron lated (13). These results also imply that in human liverP450 3A4 turnover of various substrates can be limitedtransfer capacity is the presence of cytochrome b5 . The
stimulatory effect on P450 3A4 catalysis of selected by the amount of both P450 reductase and cytochromeb5 . The difference in the kinetic parameters of bothsubstrates by cytochrome b5 in the reconstituted sys-
tem has clearly been demonstrated (8, 9, 13). The insect insect microsomes and the reconstituted system com-pared to human pooled microsomes make extrapolationmicrosomes used in these experiments contain no mea-
surable spectral cytochrome b5 , yet as mentioned above of data to in vivo clearance tentative. While for threeof the substrates examined (testosterone, nifedipine,the Vmax is much higher than either the reconstituted
system or human microsomes which both have cyto- and amitriptyline) the Vmax values for human pooledmicrosomes and reconstituted system are similar, thechrome b5 . Surprisingly, addition of cytochrome b5 to
insect microsomes enhances the turnover rate of testos- differences in apparent Km values suggest that this pa-rameter does not reflect only substrate binding (22).terone oxidation approximately a further twofold (P.
M. Shaw, unpublished results). These results may im- Further experimentation determining the Kd valuesand anaerobic P450 reduction rates may answer someply that the stimulatory effect of cytochrome b5 on P450
3A4 catalysis of testosterone has little to do with the of these questions.In order to determine whether changing the lipidP450 to reductase ratio or electron transfer, as postu-
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca
114 SHAW ET AL.
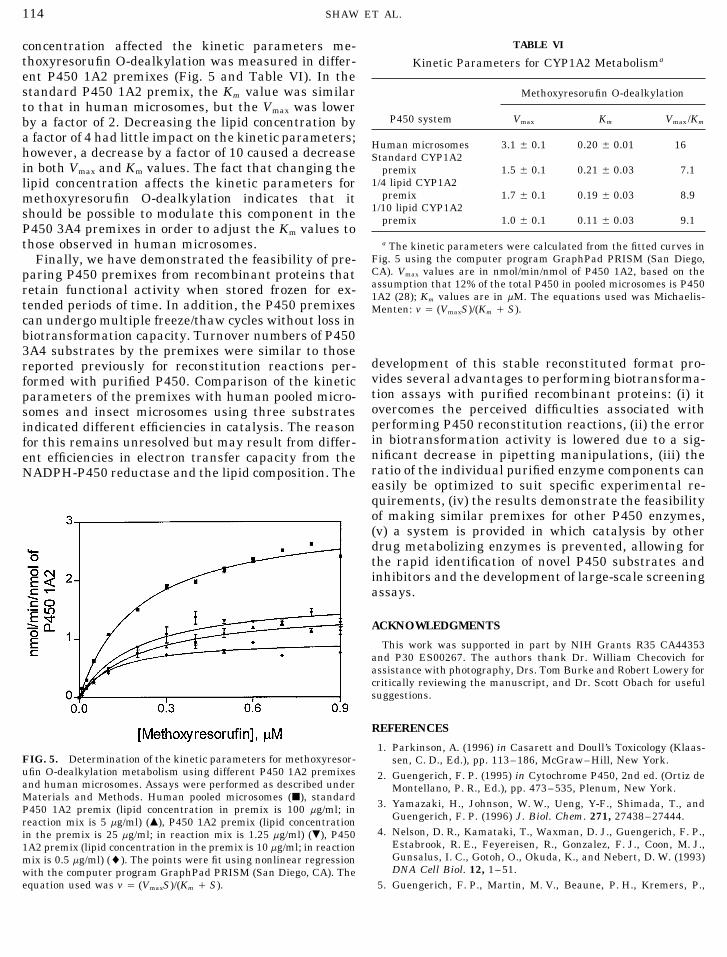
TABLE VIconcentration affected the kinetic parameters me-thoxyresorufin O-dealkylation was measured in differ- Kinetic Parameters for CYP1A2 Metabolisma
ent P450 1A2 premixes (Fig. 5 and Table VI). In thestandard P450 1A2 premix, the Km value was similar Methoxyresorufin O-dealkylationto that in human microsomes, but the Vmax was lower
P450 system Vmax Km Vmax/Kmby a factor of 2. Decreasing the lipid concentration bya factor of 4 had little impact on the kinetic parameters;
Human microsomes 3.1 { 0.1 0.20 { 0.01 16however, a decrease by a factor of 10 caused a decrease Standard CYP1A2in both Vmax and Km values. The fact that changing the premix 1.5 { 0.1 0.21 { 0.03 7.1
1/4 lipid CYP1A2lipid concentration affects the kinetic parameters forpremix 1.7 { 0.1 0.19 { 0.03 8.9methoxyresorufin O-dealkylation indicates that it
1/10 lipid CYP1A2should be possible to modulate this component in the premix 1.0 { 0.1 0.11 { 0.03 9.1P450 3A4 premixes in order to adjust the Km values tothose observed in human microsomes. a The kinetic parameters were calculated from the fitted curves in
Fig. 5 using the computer program GraphPad PRISM (San Diego,Finally, we have demonstrated the feasibility of pre-CA). Vmax values are in nmol/min/nmol of P450 1A2, based on theparing P450 premixes from recombinant proteins thatassumption that 12% of the total P450 in pooled microsomes is P450retain functional activity when stored frozen for ex- 1A2 (28); Km values are in mM. The equations used was Michaelis-
tended periods of time. In addition, the P450 premixes Menten: v Å (VmaxS)/(Km / S).can undergo multiple freeze/thaw cycles without loss inbiotransformation capacity. Turnover numbers of P4503A4 substrates by the premixes were similar to those
development of this stable reconstituted format pro-reported previously for reconstitution reactions per-vides several advantages to performing biotransforma-formed with purified P450. Comparison of the kinetiction assays with purified recombinant proteins: (i) itparameters of the premixes with human pooled micro-overcomes the perceived difficulties associated withsomes and insect microsomes using three substratesperforming P450 reconstitution reactions, (ii) the errorindicated different efficiencies in catalysis. The reasonin biotransformation activity is lowered due to a sig-for this remains unresolved but may result from differ-nificant decrease in pipetting manipulations, (iii) theent efficiencies in electron transfer capacity from theratio of the individual purified enzyme components canNADPH-P450 reductase and the lipid composition. Theeasily be optimized to suit specific experimental re-quirements, (iv) the results demonstrate the feasibilityof making similar premixes for other P450 enzymes,(v) a system is provided in which catalysis by otherdrug metabolizing enzymes is prevented, allowing forthe rapid identification of novel P450 substrates andinhibitors and the development of large-scale screeningassays.
ACKNOWLEDGMENTS
This work was supported in part by NIH Grants R35 CA44353and P30 ES00267. The authors thank Dr. William Checovich forassistance with photography, Drs. Tom Burke and Robert Lowery forcritically reviewing the manuscript, and Dr. Scott Obach for usefulsuggestions.
REFERENCES
1. Parkinson, A. (1996) in Casarett and Doull’s Toxicology (Klaas-FIG. 5. Determination of the kinetic parameters for methoxyresor- sen, C. D., Ed.), pp. 113–186, McGraw–Hill, New York.ufin O-dealkylation metabolism using different P450 1A2 premixes 2. Guengerich, F. P. (1995) in Cytochrome P450, 2nd ed. (Ortiz deand human microsomes. Assays were performed as described under Montellano, P. R., Ed.), pp. 473–535, Plenum, New York.Materials and Methods. Human pooled microsomes (j), standard
3. Yamazaki, H., Johnson, W. W., Ueng, Y-F., Shimada, T., andP450 1A2 premix (lipid concentration in premix is 100 mg/ml; inGuengerich, F. P. (1996) J. Biol. Chem. 271, 27438–27444.reaction mix is 5 mg/ml) (m), P450 1A2 premix (lipid concentration
4. Nelson, D. R., Kamataki, T., Waxman, D. J., Guengerich, F. P.,in the premix is 25 mg/ml; in reaction mix is 1.25 mg/ml) (.), P450Estabrook, R. E., Feyereisen, R., Gonzalez, F. J., Coon, M. J.,1A2 premix (lipid concentration in the premix is 10 mg/ml; in reactionGunsalus, I. C., Gotoh, O., Okuda, K., and Nebert, D. W. (1993)mix is 0.5 mg/ml) (l). The points were fit using nonlinear regressionDNA Cell Biol. 12, 1–51.with the computer program GraphPad PRISM (San Diego, CA). The
equation used was v Å (VmaxS)/(Km / S). 5. Guengerich, F. P., Martin, M. V., Beaune, P. H., Kremers, P.,
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca

115CYTOCHROME P450 RECONSTITUTED PREMIXES
Wolff, T., and Waxman, D. J. (1986) J. Biol. Chem. 261, 5051– 16. Sandhu, P., Guo, Z., Baba, T., Martin, M. V., Tukey, R., andGuengerich, F. P. (1994) Arch. Biochem. Biophys. 309, 168–177.5060.
17. Ichikawa, Y., and Yamano, T. (1967) Biochem. Biophys. Acta6. Shaw, P. M., Barnes, T. S., Cameron, D., Engeset, J., Melvin,131, 490–497.W. T., Omar, G., Petrie, J. C., Rush, W. R., Snyder, C. P., Whit-
ing, P. H., Wolf, C. R., and Burke, M. D. (1989) Biochem. J. 263, 18. Holmans, P. L., Shet, M. S., Martin-Wixtrom, C. A., Fisher,653–663. C. W., and Estabrook, R. E. (1994) Arch. Biochem. Biophys. 312,
554–565.7. Wrighton, S. A., and VandenBranden, M. (1989) Arch. Biochem.19. Shephard, E. A., Palmer, C. N. A., Segall, H. J., and Phillips, I.Biophys. 268, 144–151.
(1992) Arch. Biochem. Biophys. 294, 168–172.8. Gillam, E. M. J., Baba, T., Kim, B.-R., Ohmori, S., and Guenge-20. Shen, A. L., Porter, T. D., Wilson, T. E., and Kasper, C. B. (1989)rich, F. P. (1993) Arch. Biochem. Biophys. 305, 123–131.
J. Biol. Chem. 264, 7584–7589.9. Gillam, E. M. J., Guo, Z., Ueng, Y.-F., Hiroshi, H., Cock, I., Reilly,21. Gorski, J. L., Hall, S. D., Jones, D. R., VandenBranden, M., andP. E. B., Hooper, W. D., and Guengerich, F. P. (1995) Arch. Bio-
Wrighton, S. A. (1994) Biochem. Pharmacol. 47, 1643–1653.chem. Biophys. 317, 374–384.22. Ueng, Y-F., Kuwabara, T., Chun, Y-J., and Guengrich, F. P.10. Muller-Enoch, D., Church, P., Fleischer, S., and Guengerich,
(1997) Biochemistry 31, 370–381.F. P. (1984) J. Biol. Chem. 259, 8174–8182.23. Nash, T. (1951) Biochem. J. 55, 416–421
11. Eberhart, D. C., and Parkinson, A. (1991) Arch. Biochem. Bio-24. Werringloer, J. (1978) Methods Enzymol. 52, 297–302.phys. 291, 231–240.25. Lee, C. A., Kadwell, S. H., Kost, T. A., and Serabjit-Singh, C. J.
12. Imaoka, S., Imai, Y., Shimada, T., and Funae, Y. (1992) Biochem- (1995) Arch. Biochem. Biophys. 319, 157–167.istry 31, 6063–6069.
26. Penman, B. W., Reece, J., Smith, T., Yang, C. S., Gelboin, H. V.13. Yamazaki, H., Ueng, Y-F., Shimada, T., and Guengerich, F. P. Gonzalez, F. J., and Crespi, C. L. (1993) Pharmacogenetics 3, 28–
(1995) Biochemistry 34, 8380–8389. 39.14. Ingelman-Sundberg, M., Hagbjork, A.-L., Ueng, Y.-F., Yamazaki, 27. Yamano, S., Aoyama, T., McBride, O. W., Hardwick, J. P., Gel-
H., and Guengerich, F. P. (1996) Biochem. Biophys. Res. Com- boin, H. V., and Gonzalez, F. J. (1989) Mol. Pharmacol. 35, 83–mun. 221, 318–322. 88.
28. Shimada, T. Yamazaki, H., Mimura, M., Inui, Y., and Guenge-15. Gillam, E. M. J., Guo, Z., and Guengerich, F. P. (1994) Arch. Bio-chem. Biophys. 312, 59–66. rich, F. P. (1994) J. Pharmacol. Exp. Ther. 270, 414–423.
AID ABB 0378 / 6b44$$$301 10-28-97 08:33:47 arca
Recommended

















