Embed Size (px)
Citation preview

Molecular & Cell Biology
S. Rahgozar,PhD
University of Isfahan
Faculty of Science
5. Bioenergetics and Metabolism
5.1. Mitochondria and Oxidative Phosphorylation
93-1392
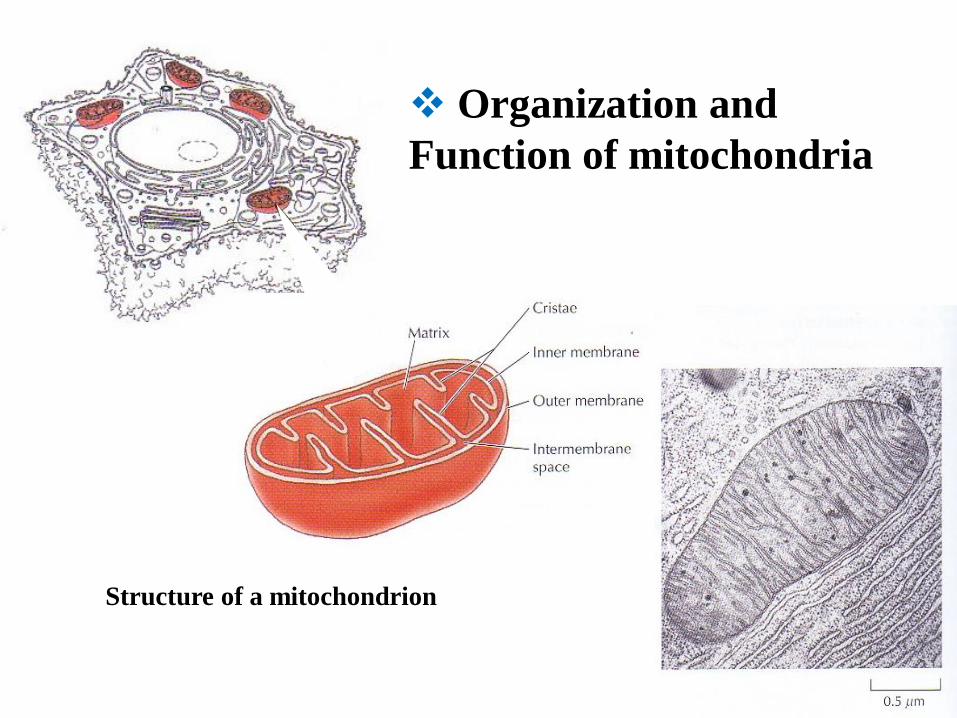
Structure of a mitochondrion
Organization and
Function of mitochondria
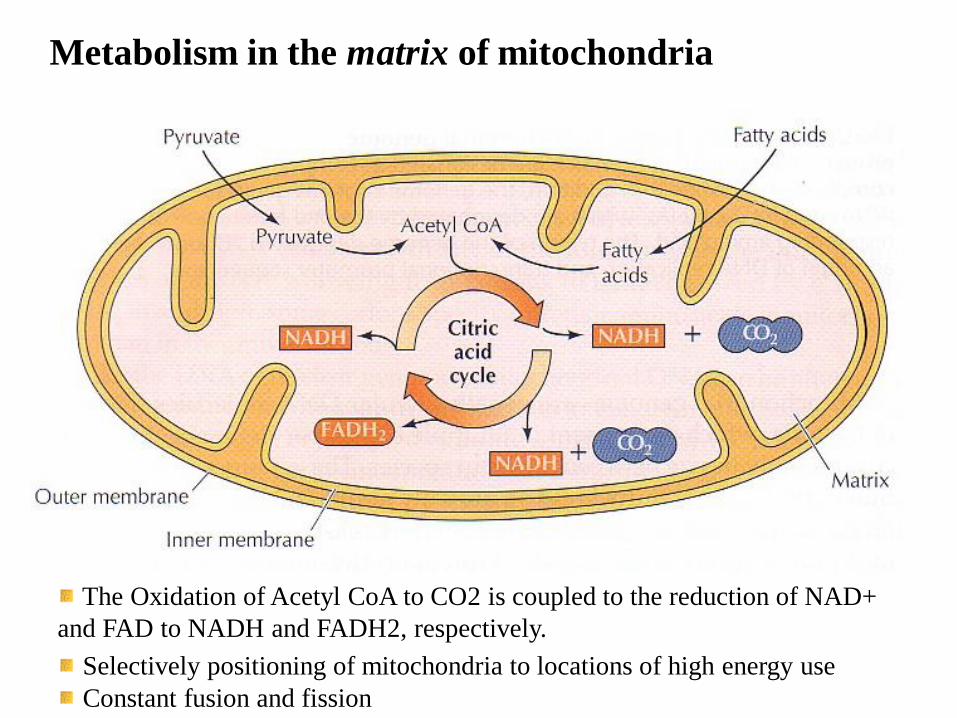
Metabolism in the matrix of mitochondria
The Oxidation of Acetyl CoA to CO2 is coupled to the reduction of NAD+
and FAD to NADH and FADH2, respectively.
Selectively positioning of mitochondria to locations of high energy use
Constant fusion and fission
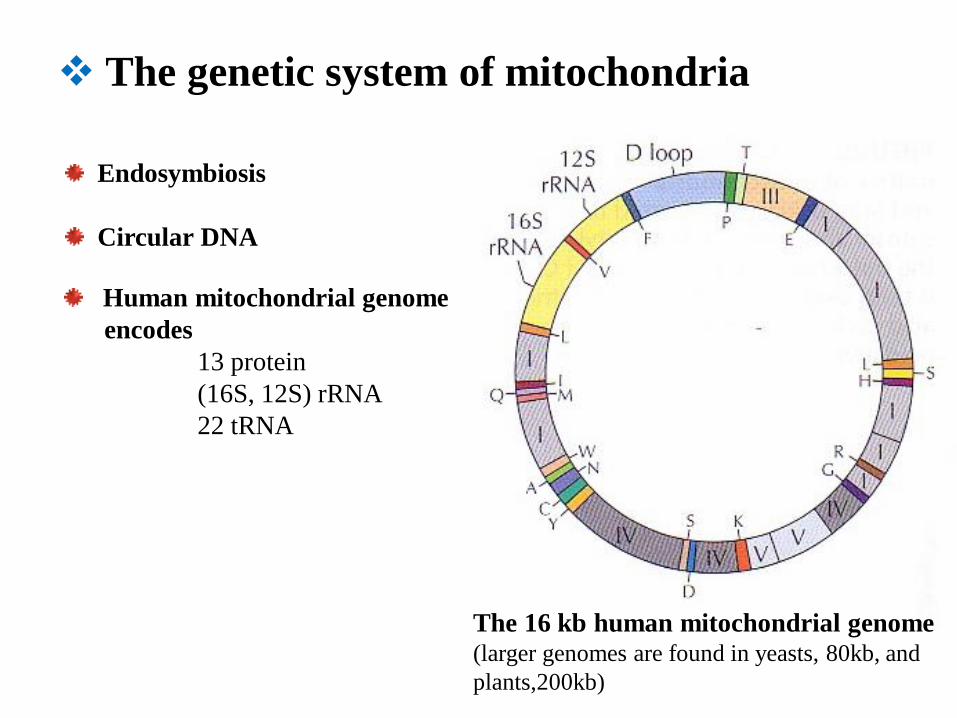
The genetic system of mitochondria
The 16 kb human mitochondrial genome (larger genomes are found in yeasts, 80kb, and
plants,200kb)
Endosymbiosis
Circular DNA
Human mitochondrial genome
encodes
13 protein
(16S, 12S) rRNA
22 tRNA
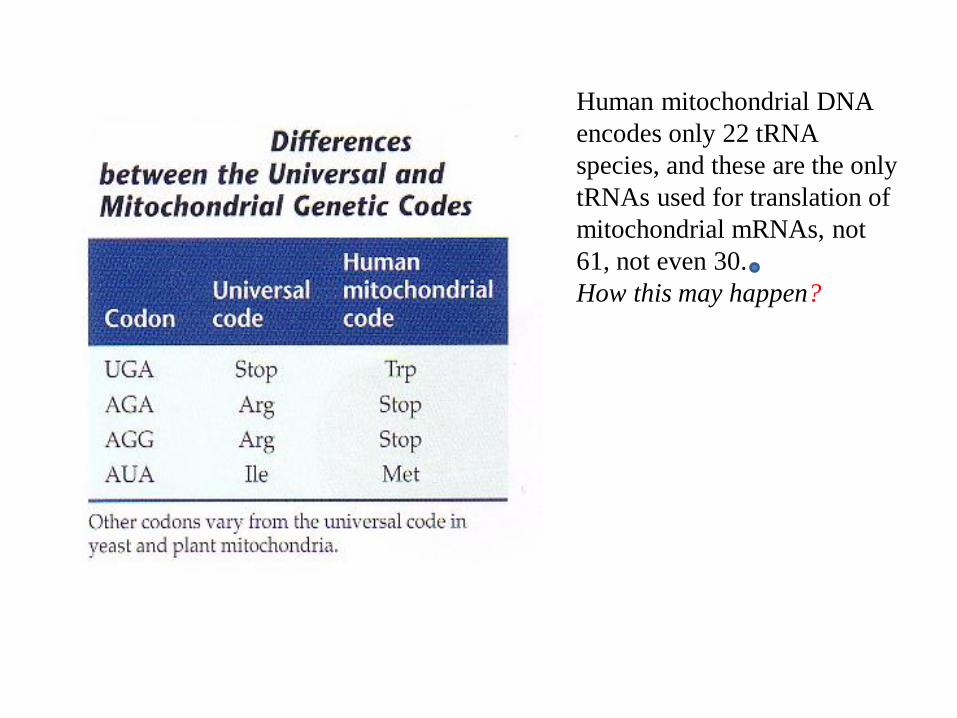
Human mitochondrial DNA
encodes only 22 tRNA
species, and these are the only
tRNAs used for translation of
mitochondrial mRNAs, not
61, not even 30.
How this may happen?
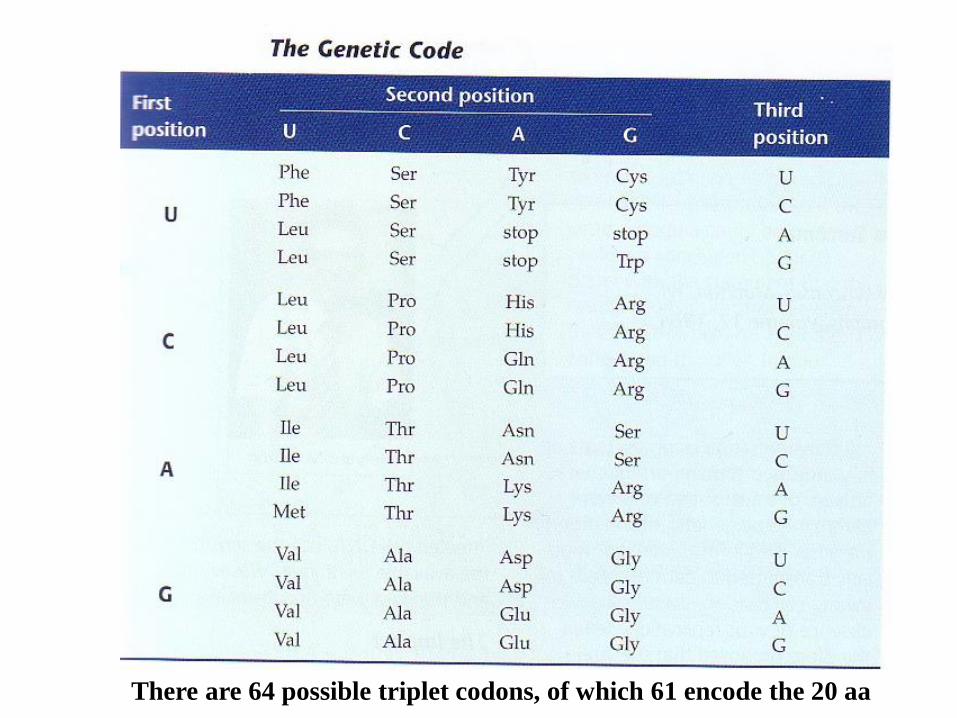
There are 64 possible triplet codons, of which 61 encode the 20 aa
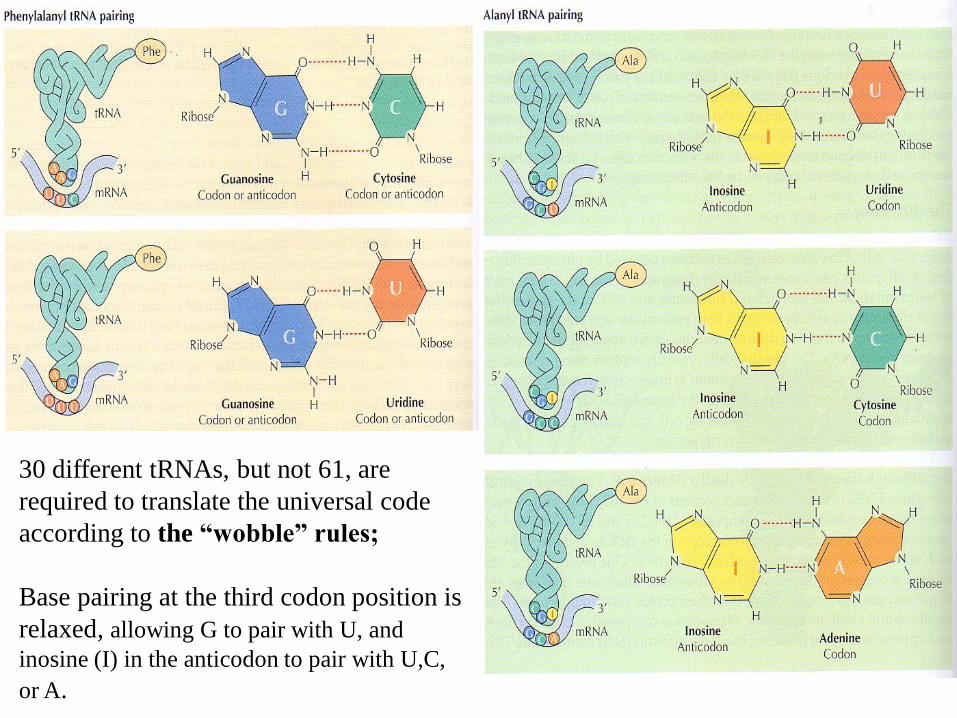
30 different tRNAs, but not 61, are
required to translate the universal code
according to the “wobble” rules;
Base pairing at the third codon position is
relaxed, allowing G to pair with U, and
inosine (I) in the anticodon to pair with U,C,
or A.

Mitochondrial Mutations are
associated with a number of diseases
Mutations in one mitochondrial
tRNA gene are associated with
metabolic syndrome, the condition
associated with obesity and diabetes.
Mutations in mitochondrial genes that encode components of the electron
transport chain are associated with Leber’s hereditary optic neuropathy, a
disease that leads to blindness.
The progress of accumulation of mutations in mitochondrial DNA during the
lifetime of individuals has been suggested to contribute to the process of aging.
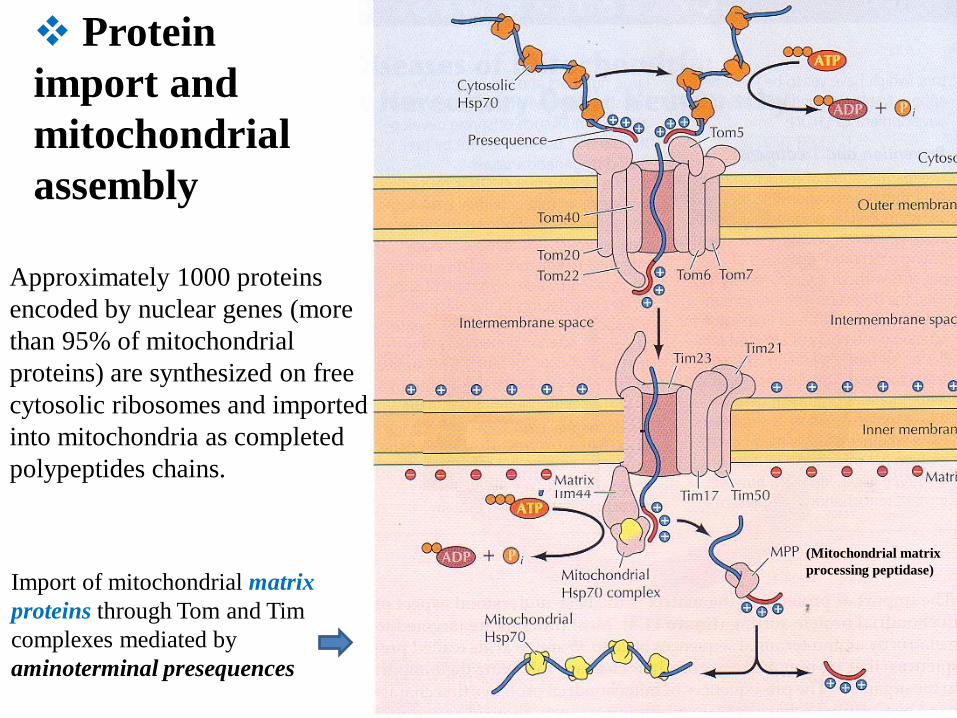
Protein
import and
mitochondrial
assembly
Approximately 1000 proteins
encoded by nuclear genes (more
than 95% of mitochondrial
proteins) are synthesized on free
cytosolic ribosomes and imported
into mitochondria as completed
polypeptides chains.
Import of mitochondrial matrix
proteins through Tom and Tim
complexes mediated by
aminoterminal presequences
(Mitochondrial matrix
processing peptidase)
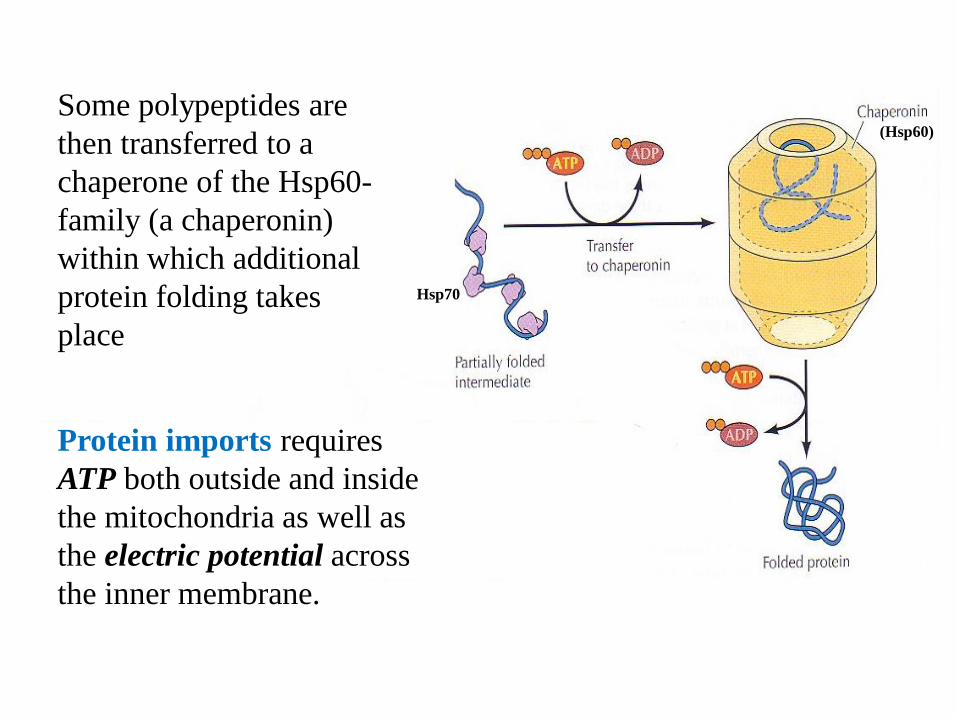
Hsp70
(Hsp60)
Some polypeptides are
then transferred to a
chaperone of the Hsp60-
family (a chaperonin)
within which additional
protein folding takes
place
Protein imports requires
ATP both outside and inside
the mitochondria as well as
the electric potential across
the inner membrane.
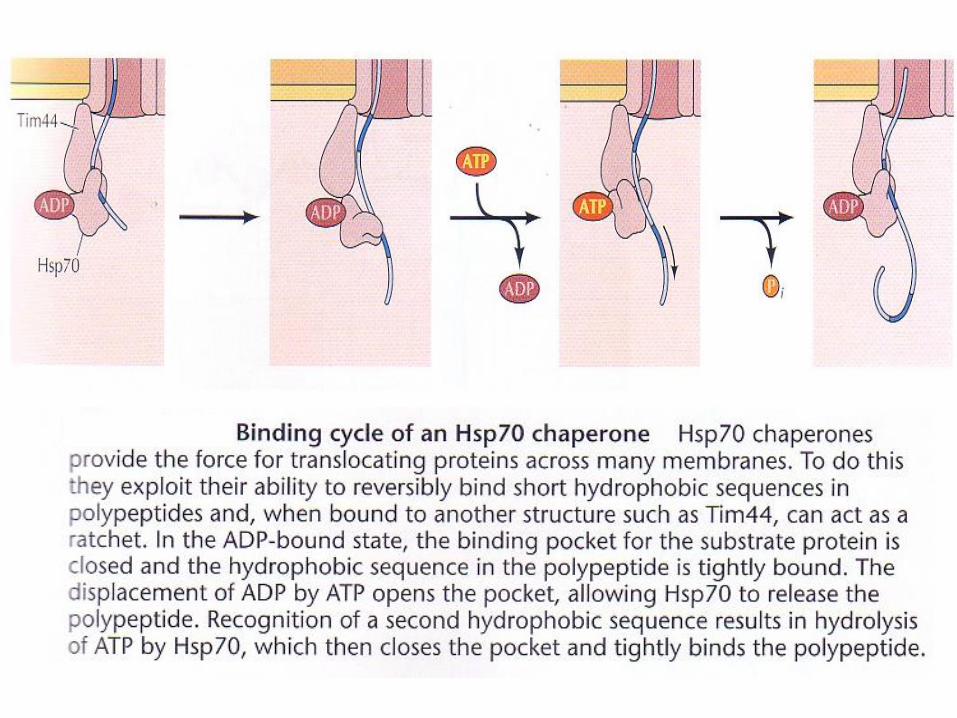
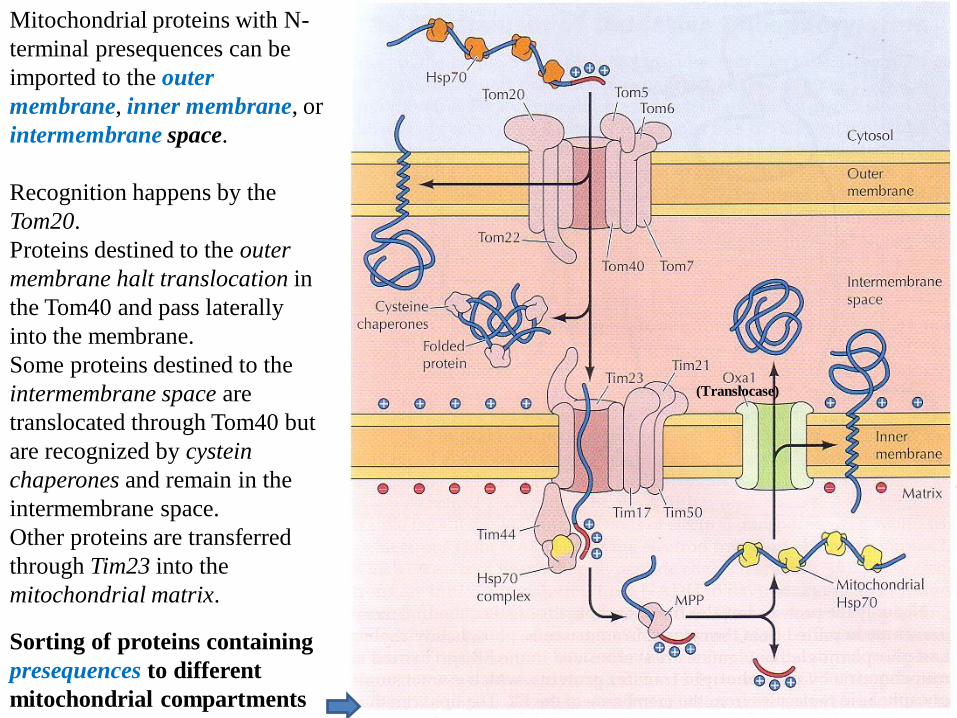
Sorting of proteins containing
presequences to different
mitochondrial compartments
Mitochondrial proteins with N-
terminal presequences can be
imported to the outer
membrane, inner membrane, or
intermembrane space.
Recognition happens by the
Tom20.
Proteins destined to the outer
membrane halt translocation in
the Tom40 and pass laterally
into the membrane.
Some proteins destined to the
intermembrane space are
translocated through Tom40 but
are recognized by cystein
chaperones and remain in the
intermembrane space.
Other proteins are transferred
through Tim23 into the
mitochondrial matrix.
(Translocase)
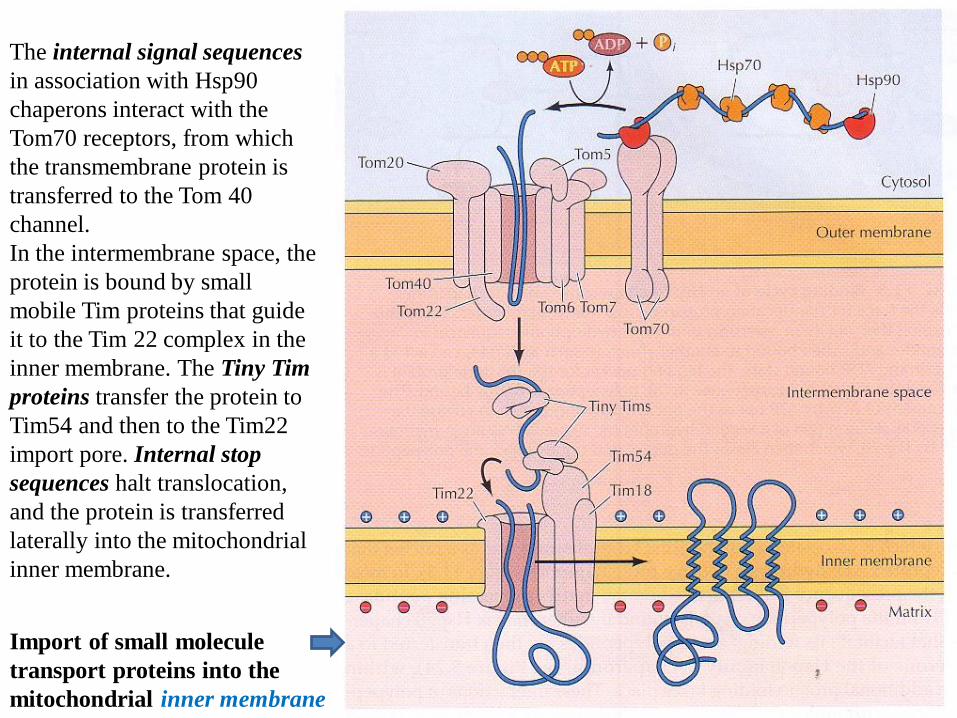
Import of small molecule
transport proteins into the
mitochondrial inner membrane
The internal signal sequences
in association with Hsp90
chaperons interact with the
Tom70 receptors, from which
the transmembrane protein is
transferred to the Tom 40
channel.
In the intermembrane space, the
protein is bound by small
mobile Tim proteins that guide
it to the Tim 22 complex in the
inner membrane. The Tiny Tim
proteins transfer the protein to
Tim54 and then to the Tim22
import pore. Internal stop
sequences halt translocation,
and the protein is transferred
laterally into the mitochondrial
inner membrane.
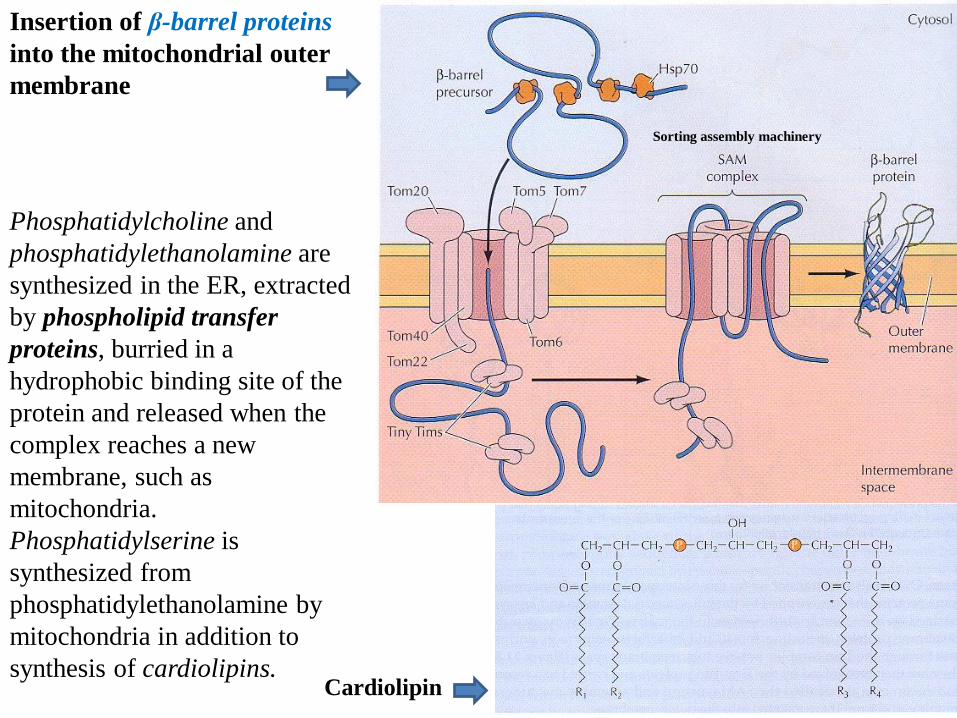
Insertion of β-barrel proteins
into the mitochondrial outer
membrane
Phosphatidylcholine and
phosphatidylethanolamine are
synthesized in the ER, extracted
by phospholipid transfer
proteins, burried in a
hydrophobic binding site of the
protein and released when the
complex reaches a new
membrane, such as
mitochondria.
Phosphatidylserine is
synthesized from
phosphatidylethanolamine by
mitochondria in addition to
synthesis of cardiolipins.
Sorting assembly machinery
Cardiolipin

The mechanism of oxidative phosphorylation
During oxidative phosphorylation, electrons derived from NADH and
FADH2 combine with O2, and the energy released from these
oxidation/reduction reactions is used to drive the synthesis of ATP from ADP.
The transfer of energy from NADH/FADH2 to O2 is a very energy-yielding
reaction with ΔG=-52.5 kcal/mol from each pair of electrons transferred.
To be harvested in usable form, this energy must be produced gradually by the
passage of electrons through a series of carriers, which constitute the electron
transport chain.
These carriers are organized into four complexes in the inner mitochondrial
membrane called: complex I (consisting of 40 polypeptide chains), coenzyme
Q (ubiquinone), complex III, complex IV (cytochrome oxidase). complex
V(ATP synthase) then serves to couple the energy-yielding reactions of
electron transport to ATP synthesis.
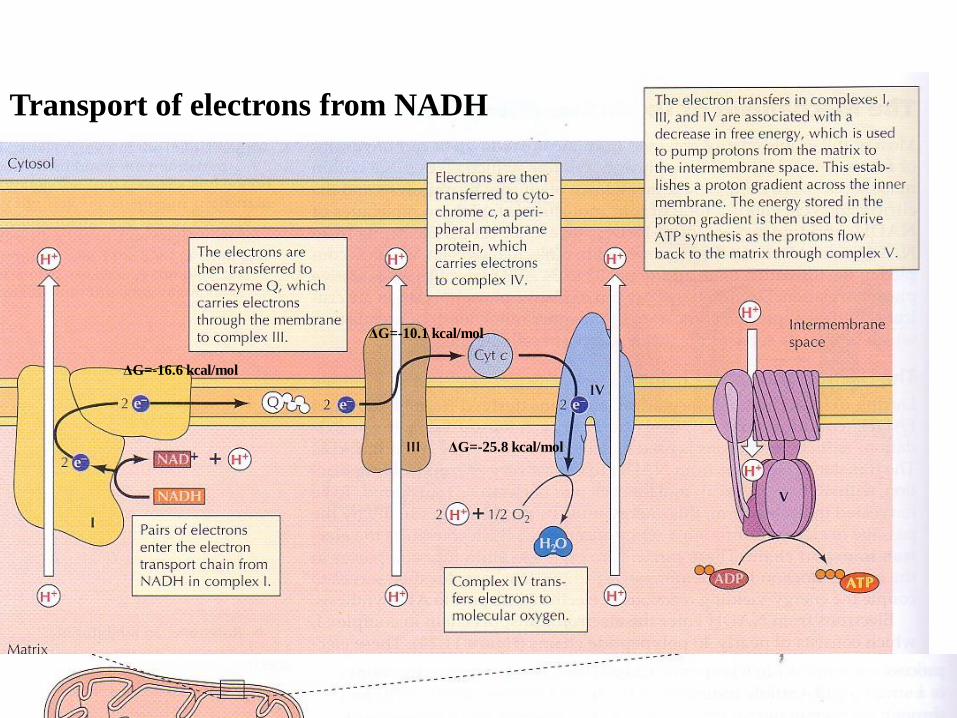
Transport of electrons from NADH
ΔG=-16.6 kcal/mol
ΔG=-10.1 kcal/mol
ΔG=-25.8 kcal/mol
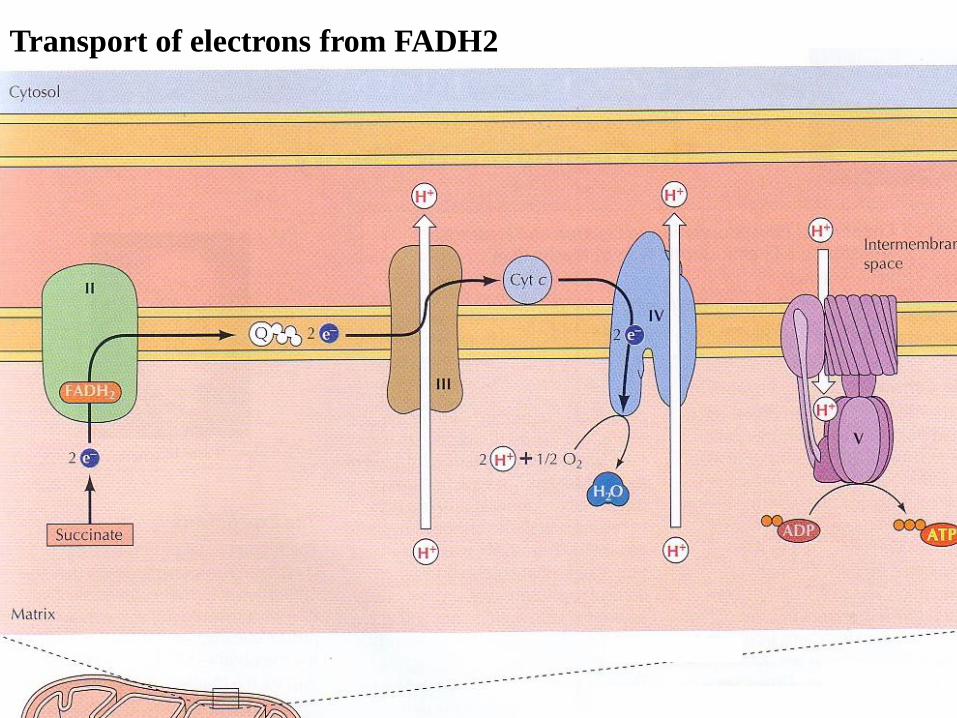
Transport of electrons from FADH2
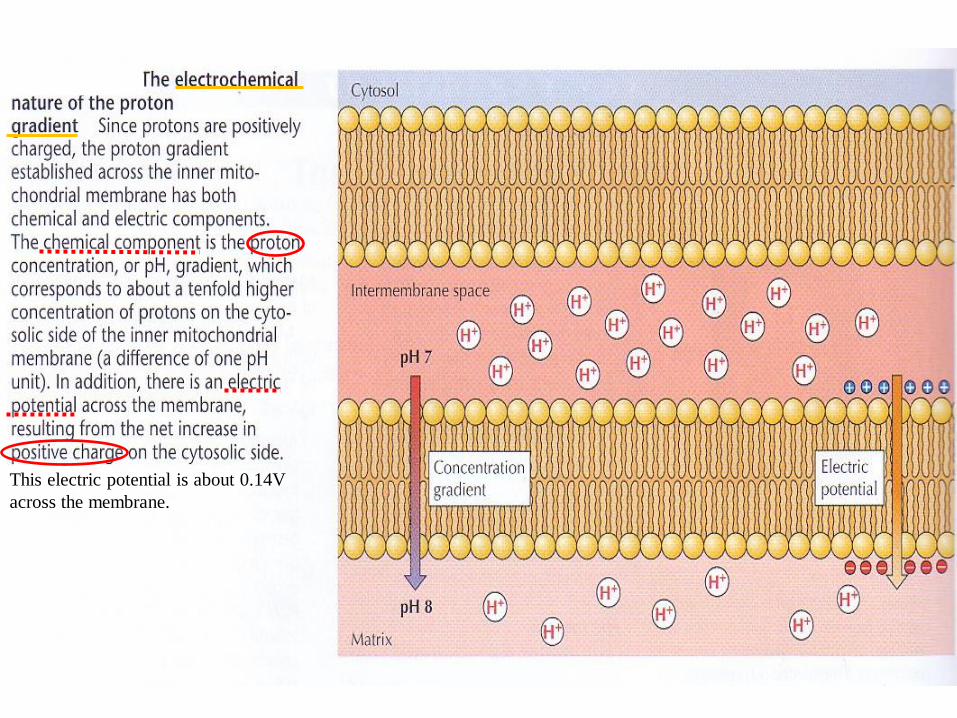
This electric potential is about 0.14V
across the membrane.
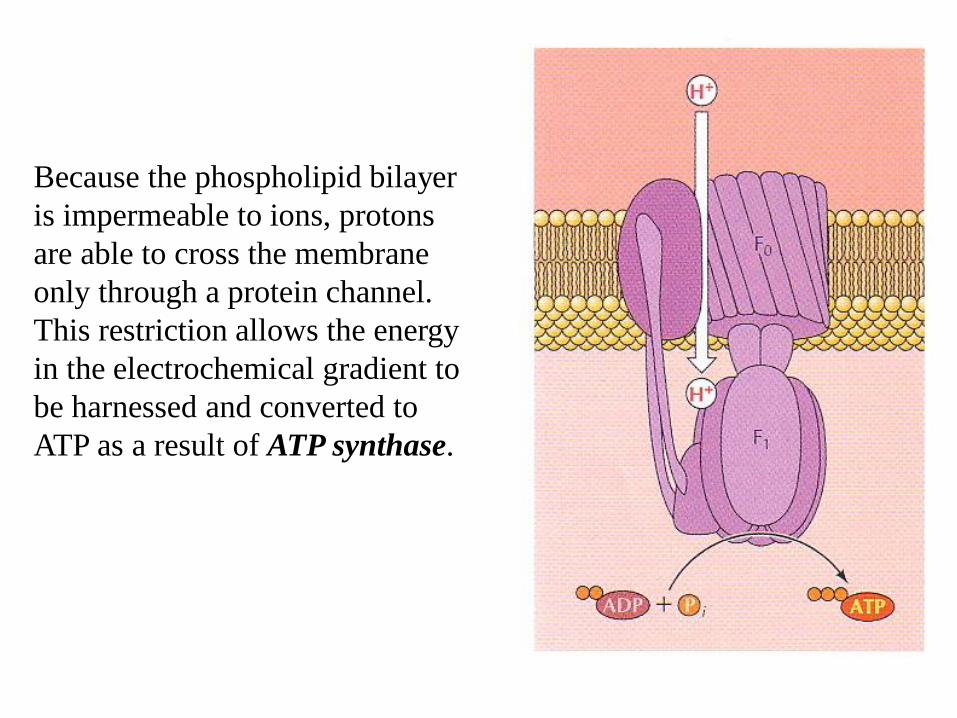
Because the phospholipid bilayer
is impermeable to ions, protons
are able to cross the membrane
only through a protein channel.
This restriction allows the energy
in the electrochemical gradient to
be harnessed and converted to
ATP as a result of ATP synthase.
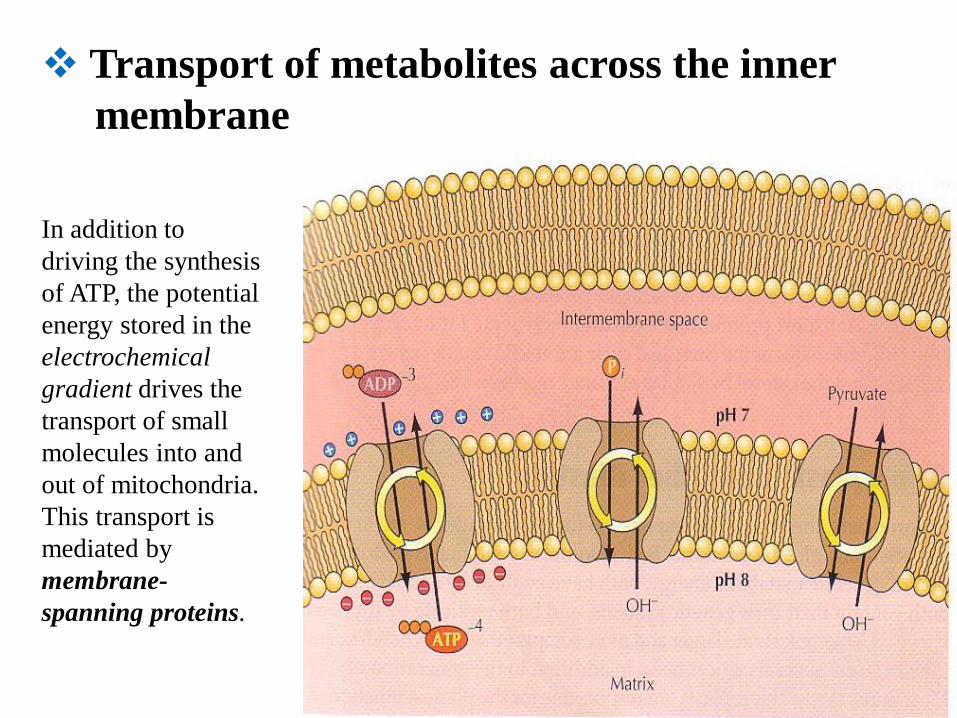
Transport of metabolites across the inner
membrane
In addition to
driving the synthesis
of ATP, the potential
energy stored in the
electrochemical
gradient drives the
transport of small
molecules into and
out of mitochondria.
This transport is
mediated by
membrane-
spanning proteins.





![11 30 201 9tÎ My 30 years with mitochondria- then, now ... · My 30 years with mitochondria- then, now, tomorrow.] Professor of Pathology & Cell Biology and Neurology Director, Neuromuscular](https://img.pdfslide.net/doc/110x75/5f5b60309fc7c66724528f5c/11-30-201-9t-my-30-years-with-mitochondria-then-now-my-30-years-with-mitochondria-.jpg)