Embed Size (px)
DESCRIPTION
Citation preview

ATP SYNTHESIS
Centre for Nano science and TechnologyCourse: Biology for Nanotechnology.Code: NST 623Course instructor: Dr. S.Kannan.
PRESENTED BY
ROOPAVATH UDAY KIRANM.Tech 1st year

Outline
• Introduction
• Electron-Transfer Reactions in Mitochondria
• ATP Synthesis
• Regulation of Oxidative Phosphorylation
• General Features of Photophosphorylation
• Light Absorption
• The Central Photochemical Event: Light-
Driven Electron Flow
• ATP Synthesis by Photophosphorylation

Adenosine Triphosphate
Energy sourcephotosynthesis and cellular
respiration
Signal
transductionsecond messenger cAMP
DNA replicationAMP

Structure
Purine base
1’C5’C
Pentos sugar
Three phosphate groups

• Substrate-level phosphorylation
direct transfer of a phospate group to ADP

In mitochondrion


• Chemiosmotic Phosphorylation
Electrochemical gradient + Osmosis
1.Oxidative Phosphorylation
2. Photophosphorylation

ATP is synthesized using the same strategy in oxidative phosphorylation and
photophosphorylation
• Oxidative phosphorylation is the process in which ATP isgenerated as a result of electron flow from NADH orFADH2 to O2 via a series of membrane-bound electroncarriers, called the respiratory chain (reducing O2 to H2Oat the end).
• Photophosphorylation is the process in which ATP (andNADPH) is synthesized as a result of electron flow fromH2O to NADP+ via a series of membrane-bound electroncarriers (oxidizing H2O to O2 at the beginning).

• Oxidative phosphorylation and photophosphorylation are
mechanistically similar in three respects.
(1) Both processes involve the flow of electrons through a
chain of membrane-bound carriers.
(2) The free energy made available by this ―downhill‖
(exergonic) electron flow is coupled to the ―uphill‖
transport of protons across a proton-impermeable
membrane, conserving the free energy of fuel oxidation
as a. transmembrane electrochemical potential
(3) The transmembrane flow of protons down their
concentration gradient through specific protein channels
provides the free energy for synthesis of ATP,
catalyzed by a membrane protein complex (ATP
synthase) that couples proton flow to phosphorylation of
ADP.

ATP GenerationGlycolysis
• Conversion of glucose to pyruvate
• Net synthesis of 2 ATP by substrate level
phosphorylation
Krebs Cycle
• Converts pyruvate to acetyl CoA & carbon dioxide
• 10 molecules of coenzymes NADH and 2 of FADH2 are
produced. Results in synthesis of 30 ATP and 4 ATP molecules,
respectively in the respiratory chain.
Electron Transport (Respiratory) Chain
• The reduced coenzymes enter into the respiratory
chain of the inner mitochondrial membrane
• Electron transport along the chain generates a proton
electrochemical gradient and this is used to produce ATP


Chemiosmotic theory:
• Introduced by Peter Mitchell in 1961
• Transmembrane differences in proton
concentration are the reservoir for the energy
extracted from biological oxidation reactions.
• It provides insight into the processes of
oxidative phosphorylation and
photophosphorylation, and into such
apparently disparate energy transductions as
active transport across membranes and the
motion of bacterial flagella.

Proton Gradient Across the Membrane:
“Chemiosmosis”
• It is the universal mechanism of ATP productionwhich involves the production of a proton motiveforce (pmf) based on a proton gradient acrossthe membrane.
• Energy to establish this electrochemical protongradient is provided by the energy released aselectrons move to lower energy levels down theelectron transport chain and the coupling of thisfree energy to the movement of protons across theIMM against the proton gradient [from matrix toIMS]
• ATP is synthesized by the ATP synthase FoF1
complex : protons move with the proton gradientthrough FoF1 to generate ATP [from IMS to matrix]

The chemiosmoticmodel of Mitchell

OXIDATIVE PHOSPHORYLATION
• The discovery in 1948 by Eugene Kennedy and
Albert Lehninger that mitochondria are the site of
oxidative phosphorylation in eukaryotes marked the
beginning of the modern phase of studies in
biological energy transductions.
• Oxidative phosphorylation begins with the entry of
electrons into the respiratory chain.
• Most of these electrons arise from the action of
dehydrogenases that collect electrons from
catabolic pathways and funnel them into universal
electron acceptors—nicotinamide nucleotides
(NAD+ or NADP+) or flavin nucleotides (FMN or
FAD).


• The mitochondrial respiratory chain consists of a series of
sequentially acting electron carriers, most of which are
integral proteins with prosthetic groups capable of
accepting and donating either one or two electrons.
• Three types of electron transfers occur in oxidative
phosphorylation:
(1) Direct transfer of electrons, as in the reduction of Fe+3
to Fe+2;
(2) Transfer as a hydrogen atom (H+ +e); and
(3) Transfer as a hydride ion (:H), which bears two
electrons.
• The term reducing equivalent is used to designate a
single electron equivalent transferred in an oxidation-
reduction reaction.

Electrons collected in NADH and FADH2 are
released and transported to O2 via the respiratory
chain
• The chain is located on the convoluted inner
membrane (cristae) of mitochondria in
eukaryotic cells (revealed by Eugene
Kennedy and Albert Lehninger in 1948) or
on the plasma membrane in prokaryotic cells.
• A 1.14-volt potential difference (E`0)
between NADH (-0.320 V) and O2 (0.816 V)
drives electron flow through the chain.

• The respiratory chain consists of four large multi-
protein complexes (I, II, III, and IV; three being
proton pumps) and two mobile electron carriers,
ubiquinone (Q or coenzyme Q, and cytochrome c.
• Prosthetic groups acting in the proteins of
respiratory chain include flavins (FMN, FAD),
hemes (heme A, iron protoporphyrin IX, heme C),
iron-sulfur clusters (2Fe-2S, 4Fe-4S), and copper.

Four multi-protein Complexes (I, II, III, and IV)
Two mobileElectron carriers
III
III
IV

• Ubiquinone (also called coenzyme Q, or simply
Q) is a lipid-soluble benzoquinone with a long
isoprenoid side chain
• Because ubiquinone is both small and
hydrophobic, it is freely diffusible within the lipid
bilayer of the inner mitochondrial membrane and
can shuttle reducing equivalents between other,
less mobile electron carriers in the membrane. And
because it carries both electrons and protons, it
plays a central role in coupling electron flow to
proton movement.

Complete reduction
of ubiquinone
requires two
electrons and two
protons, and occurs
in two steps through
the semiquinone
radical
intermediate.

Heme groups of cytochrome proteins
Heme groupsOf cytochromes

Different types of iron-sulfur centers•Iron atoms cycle between Fe2+
(reduced) and Fe3+(oxidized).
•At least eight Fe-S proteinsact in the respiratory chain.
4Fe-4S2Fe-2S
A ferredoxin

NADH:Ubiquinone
Oxidoreductase
a.k.a. Complex I
• One of the largest macro-
molecular assemblies in the
mammalian cell
• Over 40 different polypeptide
chains, encoded by both nuclear
and mitochondrial genes
• NADH binding site in the matrix
side
• Non-covalently bound flavin
mononucleotide (FMN) accepts
two electrons from NADH
• Several iron-sulfur centers pass
one electron at the time toward
the ubiquinone binding site

NADH:Ubiquinone Oxidoreducase is a
Proton Pump
• Transfer of two electrons from NADH to ubiquinone is
accompanied by a transfer of protons from the matrix (N)
to the inter-membrane space (P)
• Experiments suggest that about four protons are
transported per one NADH
NADH + Q + 5H+N = NAD+ + QH2 + 4 H+
P
• Reduced coenzyme Q picks up two protons
• Despite 50 years of study, it is still unknown how the four
other protons are transported across the membrane

Iron-Sulfur Centers
• Found in several proteins of
electron transport chain,
including NADH:ubiquinone
oxidoreductase
• Transfers one electron at a
time

Succinate Dehydrogenase
a.k.a. Complex II
• FAD accepts two
electrons from succinate
• Electrons are passed, one
at a time, via iron-sulfur
centers to ubiquinone
that becomes reduced
QH2

• The cytochromes are proteins with
characteristic strong absorption of visible light,
due to their iron-containing heme prosthetic
groups. Mitochondria contain three classes of
cytochromes, designated a, b, and c, which are
distinguished by differences in their light-
absorption spectra.
• Each type of cytochrome in its reduced (Fe2)
state has three absorption bands in the visible
range

Cytochrome bc1 Complex a.k.a. Complex III
• Uses two electrons from QH2 to reduce two
molecules of cytochrome c

The Q Cycle
• 4 H+ / 2 e-that reach CytC
• 2 H+ from QH2
• 2 H+ from the matrix

Cytochrome c
• Cytochrome c is a soluble
heme-containing protein
in the intermembrane
space
• Heme iron can be either
ferrous(Fe3+, oxidized) or
ferric(Fe2+, reduced)
• Cytochrome c carries a
single electron from the
cytochrome bc1 complex
to cytochrome oxidase

Cytochrome c Absorbs Visible
Light
• Intense Soret band near
400 nm absorbs blue light
and gives cytochrome c
an intense red color
• Cytochromes are
sometimes named by the
position of their longest-
wavelength peak

Cytochrome Oxidase
a.k.a. Complex IV
• Mammalian cytochrome oxidase is a membrane
protein with 13 subunits
• Contains two heme groups
• Contains copper ions
– Two ions (CuA) form a binuclear center
– Another ion (CuB) bonded to heme forms Fe-Cu
center

Cytochrome C Oxidase (complex IV) Transport

Structure of the Cytochrome C Oxidase Monomer
• The heme groups are
shown in blue and red
and copper sites in
green
• The catalytic core
consists of I yellow, II
blue, III pink
• The entire complex
consists of 13 subunits

A proposed reaction cycle for the four-electronreduction of O2 by cytochrome oxidase (at theHeme a3-CuB center)

Structure of Beef Heart Cytochrome Oxidase
The protein is a dimer of two 13 monomers
3 dimensional structure of beef heart cytochrome
oxidase at 2.8 angstrom resolution

The order of the many electron carriers on the
respiratory chain have been elucidated via various
studies• Measurement of the standard reduction potential
(E`0)): Electrons tend to transfer from low E`0
carriers to high E`0 carriers (but may deviate from
this in real cells).
• Oxidation kinetics studies: Full reduction followed
by sudden O2 introduction; earlier oxidation, closer
to the end of the respiratory chain; using rapid and
sensitive spectrophotometric techniques to follow
the oxidation of the cytochromes, which have
different wavelength of maximal absorption).

Electron carriers may have an order of increasing E`0

• the standard reduction potentials of theindividual electron carriers have beendetermined experimentally . We would expectthe carriers to function in order of increasingreduction potential, because electrons tend toflow spontaneously from carriers of lower Eto carriers of higher E.
• The order of carriers deduced by this method isNADH → Q → cytochrome b →cytochrome c1 → cytochrome c →cytochrome a → cytochrome a3 → O2.

• Effects of various specific inhibitors: those
before the blocked step should be reduced and
those after be oxidized.
• Isolation and characterization of each of the
multiprotein complexes: specific electron
donors and acceptors can be determined for
portions of the chain.

Various inhibitors generate various patterns of
reduced/oxidized carriers
Reduced Oxidized
Reduced Oxidized
Reduced

Oxidative
Phosphorylation
(0n inner membrane
of mitochondria)

Electron transfer to O2 was found to be coupled to ATP synthesis from ADP + Pi in isolated mitochondria
• ATP would not be synthesized when only ADPand Pi are added in isolated mitochondriasuspensions.
• O2 consumption, an indication of electron flow,was detected when a reductant (e.g., succinate) isadded, accompanied by an increase of ATPsynthesis.
• Both O2 consumption and ATP synthesis weresuppressed when inhibitors of respiratory chain(e.g., cyanide, CO, or antimycin A) was added.
• ATP synthesis depends on the occurrence ofelectron flow in mitochondria.

• O2 consumption (thus electron flow) wasneither observed if ADP was not added tothe suspension, although a reductant isprovided!
• The O2 consumption was also not observed in thepresence of inhibitors of ATP synthase (e.g.,oligomycin or venturicidin).
• Electron flow also depends on ATP synthesis!

Electron transfer was found to be obligatorily
coupled to ATP Synthesis in isolated
mitochondria suspensions:
neither occurs without the other.

3 D Model of ATP Synthase:
An Electrical Mechano-Chemical
Molecular Complex
• The Fo portion is composed of
integral transmembranous
proteins a, b and 9-14 copies of c
which forms a ring-like structure
in the plane of the membrane.
• The F1 head piece is composed of
a hexagonal array of alternating
and subunits, a central protein
with a helical coil that associates
with and proteins and extends
into the c protein ring in the Fo.

Atomic Force Microscopy of C-subunit Ring Structures
Isolated from Chloroplast ATP Synthase and Inserted
Into Liposomes

c ring & a subunit structure
•each c subunit has 2 membrane-spanning a helices– midway along 1 helix: asp– COOH↔COO–
•a subunit has 2 half-channels
H+ path
•H+ from cytosol diffuses via half-channel to asp on c ring subunit (c1)
•this subunit can now move to interface membrane, allowingc ring to rotate
•c9 now interfaces matrix half-channel, allowing H+ to diffuse into matrix
c ring
subunit a
H+ path through membrane
c1 c9
matrixhalf-channel
cytosolichalf-channel
asp
subunit ac subunit

cannot rotate ineither direction
can rotate clockwise
matrix
H+ flow drives rotation of c ring

Binding-change mechanism of ATP synthesis
• Rotation of gama subunit drives release of tightlybound ATP
• 3 active sites cycle through 3 structural states:O, open; L, loose-binding; T, tight-binding
• At T site, ADP + Pi ATP, but ATP can’t dissociate• G rotation causes T O, L T, O L• As a result of the TO structural change,
ATP can now dissociate from what is now an O site.
T O
ATP
ADP + Pi
ATP
120° rotation of (counterclockwise)
T
TO
O
L L
1 1
2 2
3 3

Synthesis of ATP: Rotary Catalysis• ATP is synthesized by coupling the energy liberated during
proton translocation through the FoF1 to a motive force thatrotates
the C ring structure and the attached subunit.
• -subunits contain the catalytic sites of ATP synthesis. 120degree
units of rotation of the protein around the stationary /
hexagonal array results in altered associations of the protein
with the protein forming the L, T and O states for the 3 β-subunits.
ATP is produced in the T state where the ∆G = ~ 0.
• Each rotation of 360 degrees of the γ subunit results in 3 ATP,one
for each β-subunit.
The model shows the rotation as arbitrarily clockwise.
∆G = ~ 0

Nature 386, 299 - 302 (20 March 1997); doi:10.1038/386299a0
Direct observation of the rotation of F1-ATPaseHIROYUKI NOJI*, RYOHEI YASUDA†, MASASUKE YOSHIDA* & KAZUHIKO KINOSITA JR†
†Department of Physics, Faculty of Science and Technology, Keio University, Hiyoshi 3-14-1, Kohoku-ku, Yokohama 223, Japan

Transport across inner mitochondrial membrane• p also drives flow of substances across inner membrane• Transported by specific carrier proteins
• Cotransport: coupled transport of 2 substances–Symport:
both move in same direction
CH3CCOO– + H+
O
HPO4= + H+
–Antiport:each movesin oppositedirection
ADP-ATPexchange

Active Transport of ATP, ADP & Pi
• Adenine Nucleotide Translocase
– Antiporter
– (ATP4-matrixADP3-
inter membrane)
• Phosphate trans locase
– Symporter
– {Pi- , H+} inter membrane => {Pi- , H+} matrix

Summary of ATP synthesis & translocation of ATP,ADP & Pi

Energy of Light is Used to
Synthesize ATP in
Photosynthetic Organisms
• Light causes charge separation between
a pair chlorophyll molecules
• Energy of the oxidized and reduced
chlorophyll molecules is used drive
synthesis of ATP
• Water is the source of electrons that are
passed via a chain of transporters to the
ultimate electron acceptor, NADP+
• Oxygen is the byproduct of water
oxidation


Various Pigments Harvest the
Light Energy
The energy is transferred to the photosynthetic reaction center





Light-Induced Redox Reactions and Electron
Transfer Cause Acidification of Lumen
The proton-motive force across the thylakoidmembrane drives the synthesis of ATP

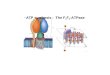
Flow of Protons: Mitochondria,
Chloroplasts, Bacteria
• Mitochondria and chloroplasts arose endosymbionts - entrapped bacteria
• Bacterial cytosol became mitochondrial matrix and chloroplast stroma

Photophosphorylation(on thylakoid of chloroplasts)

References
• Lehninger Principles of Biochemistry, 5th
Edition- © 2008 W.H Freeman and company.
• Fundamentals of Biochemistry- a text book ,
H.P. Gajera, S.V. Patel, B.A. Golakiya.
• Fundamentals of Biochemistry- J.L. Jain,
Sunjay Jain, Nitin Jain.