Embed Size (px)
Citation preview

SCHIZOPHRENIA - GENETICS
PRESENTER - Dr. Sriram.R, 2nd year MD PG
CHAIRPERSON – Dr. Thenmozhi, AP
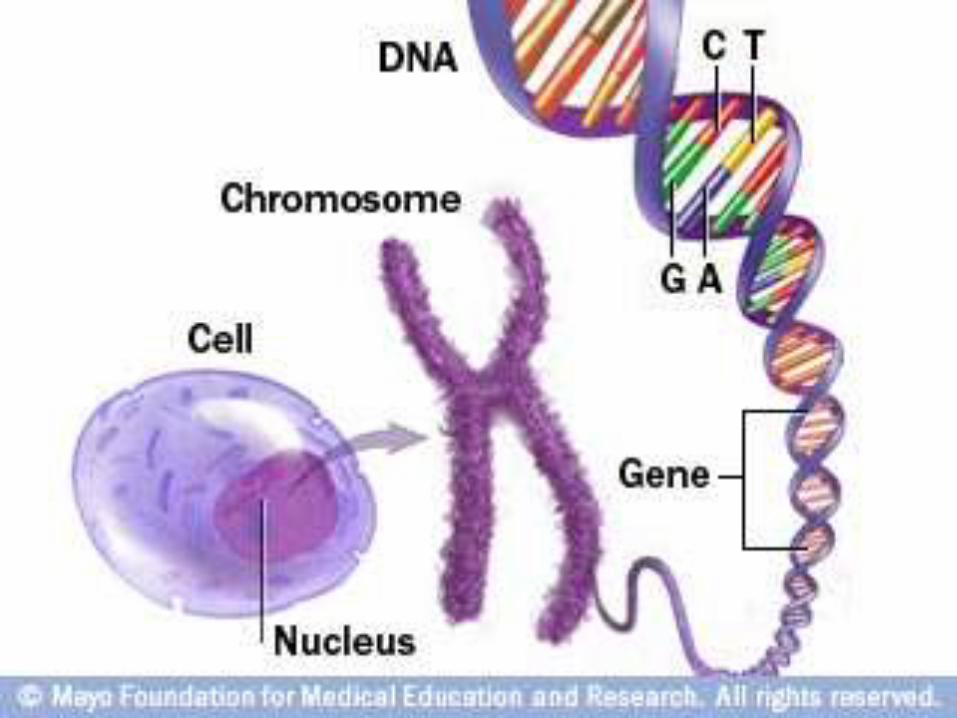
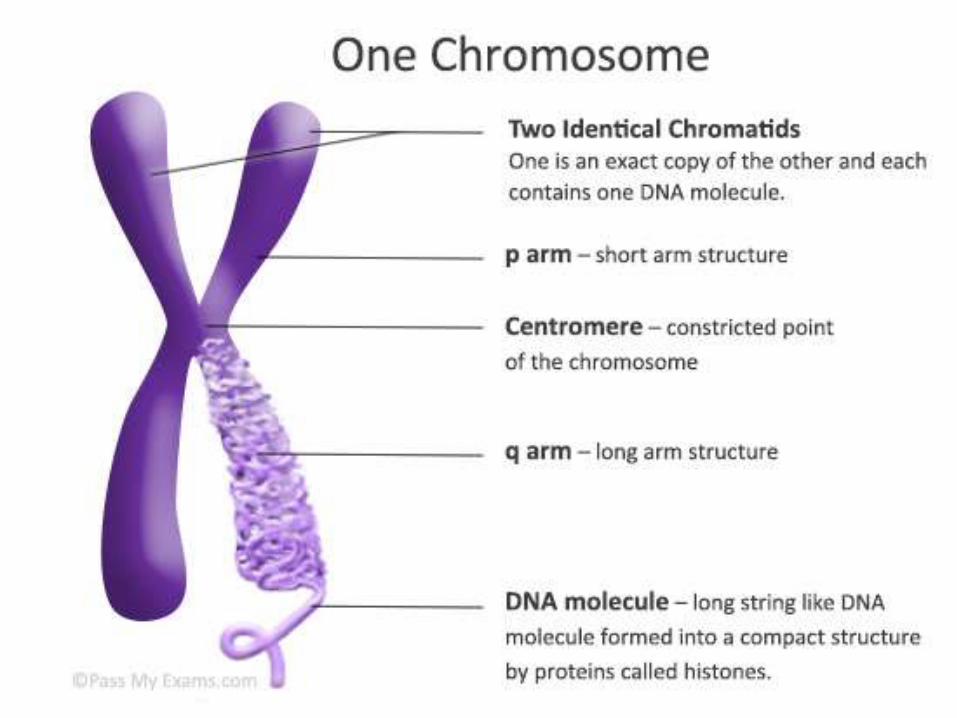
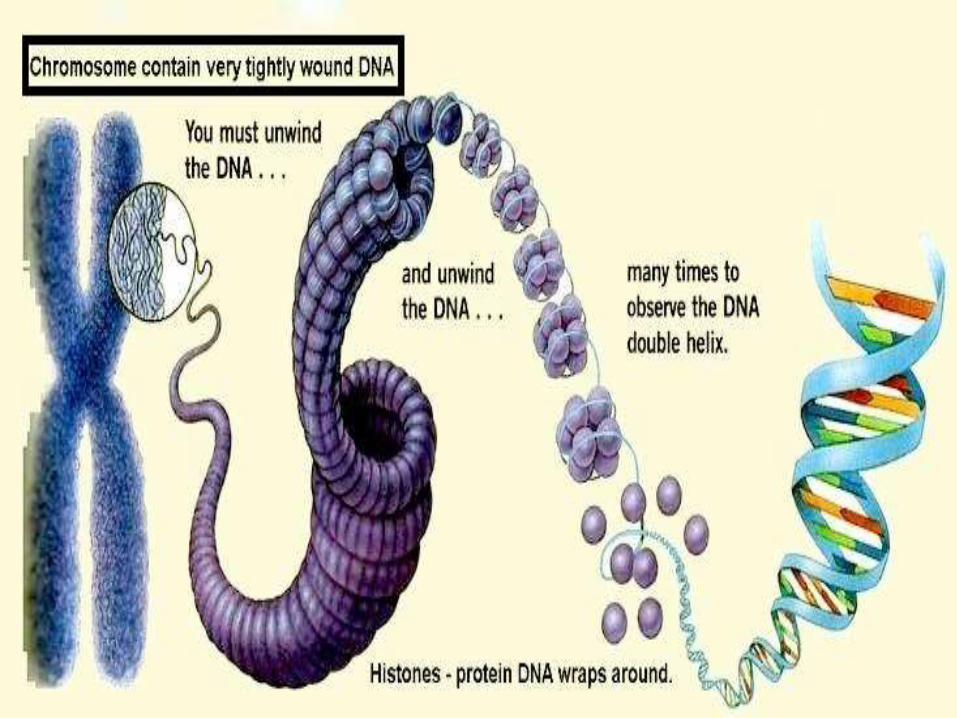

CONTENTS
• Abbreviations used
• Why study genetics of schizophrenia?
• Genetic terms used in SCZ
• Sub-groups of genetics in SCZ
• Family studies
• Twin studies
• Adoption studies
• Linkage studies
• Mode of inheritance
• Association studies and GWAS
• Chromosomal aberrations and CNVs
• Pleiotropy and overlap with BPAD/Autism
• Future directions

ABBREVIATIONS USED
• SCZ – Schizophrenia
• MZ- monozygotic
• DZ- dizygotic
• GWAS- Genome-wide Association Studies
• LD- Linkage Disequilibrium
• ISC- International Schizophrenia Consortium
• MGS- Molecular Genetics of Schizophrenia
• SGENE- Schizophrenia Genetics Consortium

Why study genetics of schizophrenia?
• Overall, psychiatric diseases are
– First-rank public health problems
– Cause enormous morbidity, mortality and personal/societal cost
– Mostly idiopathic
– Despite considerable research, little known for certain about the disease etiology
• Genetic knowledge of SCZ
– Can give a definite biological basis for distinguishing affected from non-affected (Sullivan 2010)
– Can guide for newer treatments

GENETIC TERMS USED IN SCZ
• Concordance: the probability that a pair of individuals will both have a certain characteristic, given that one of the pair has the characteristic. For example, twins are concordant when both have or both lack a given trait. (Lewontin,1982)
• Heritability: proportion of the variance of a phenotype (disease,trait) that is due to genes, estimated from risks to twins and other relatives
• Mendelian disease: caused by a (usually rare) change(mutation) in DNA sequence on one(dominant) or both(recessive) of an individual’s pair of chromosomes
• Complex disease: caused by an interaction of multiple genetic and/or environmental factors
• Allele: A variant of the similar DNA sequence located at a given locus is called an allele.
• Haplotype: collection of specific alleles in a cluster of tightly-linked genes on a chromosome that are likely to be inherited together
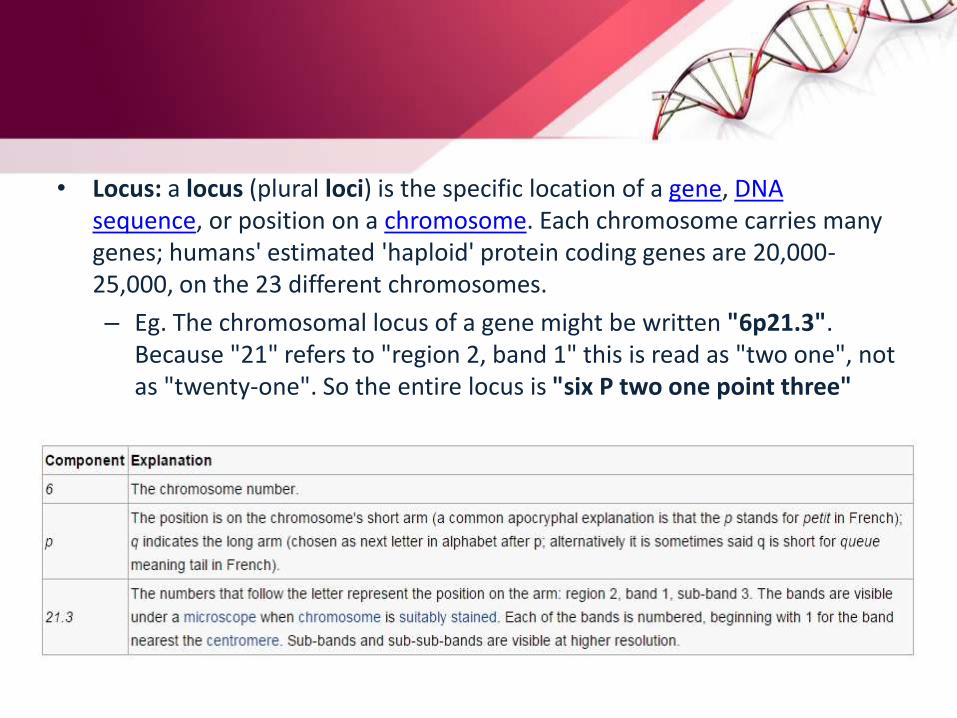
• Locus: a locus (plural loci) is the specific location of a gene, DNA sequence, or position on a chromosome. Each chromosome carries many genes; humans' estimated 'haploid' protein coding genes are 20,000-25,000, on the 23 different chromosomes.
– Eg. The chromosomal locus of a gene might be written "6p21.3". Because "21" refers to "region 2, band 1" this is read as "two one", not as "twenty-one". So the entire locus is "six P two one point three"

• Genetic map: The ordered list of loci known for a particular genome is called a genetic map.
• Gene mapping: Gene mapping is the process of determining the locus for a particular biological trait.
• Genetic linkage is the tendency of alleles that are located close together on a chromosome to be inherited together during meiosis. Genes whose loci are nearer to each other are less likely to be separated onto different chromatids during chromosomal crossover, and are therefore said to be genetically linked. In other words, the nearer two genes are on a chromosome, the lower is the chance of a swap occurring between them, and the more likely they are to be inherited together.
• Linkage disequilibrium(LD): non-random association of alleles at two or more loci that descend from single, ancestral chromosomes (Reich, 2001)
– A variant that is highly correlated with a truly causal variant will show a similar statistical association to phenotype.
– If the LD is widespread, many fewer markers will need to be assayed (Psychiatric GWAS consortium coordinating committee 2009)
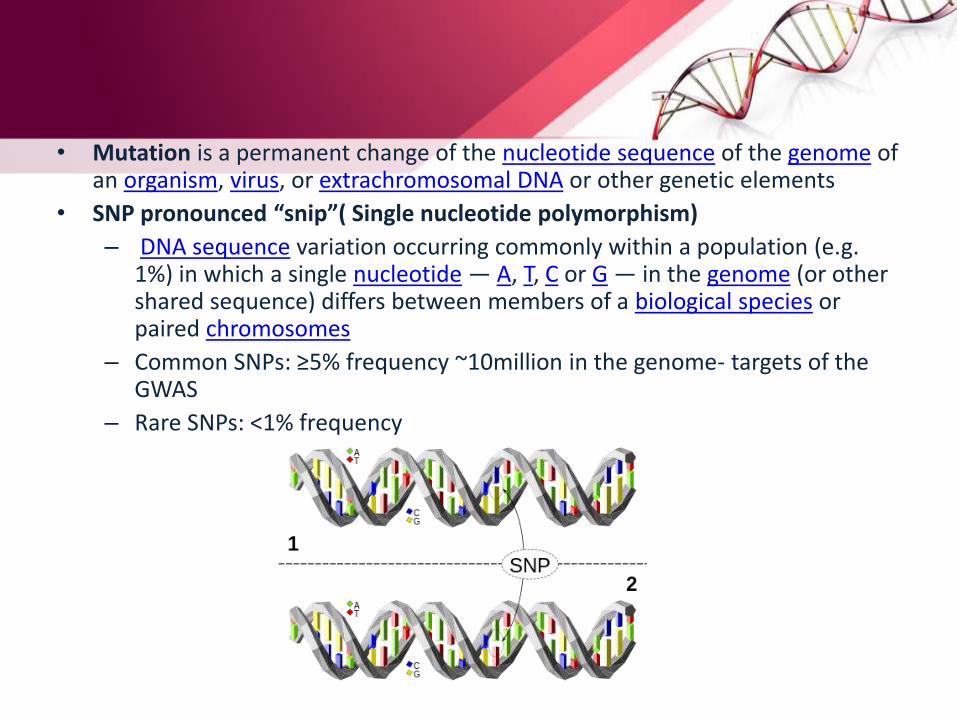
• Mutation is a permanent change of the nucleotide sequence of the genome of an organism, virus, or extrachromosomal DNA or other genetic elements
• SNP pronounced “snip”( Single nucleotide polymorphism)
– DNA sequence variation occurring commonly within a population (e.g. 1%) in which a single nucleotide — A, T, C or G — in the genome (or other shared sequence) differs between members of a biological species or paired chromosomes
– Common SNPs: ≥5% frequency ~10million in the genome- targets of the GWAS
– Rare SNPs: <1% frequency

• Copy-number variations (CNVs)—a form of structural variation—are alterations of the DNA of a genome that results in the cell having an abnormal or, for certain genes, a normal variation in the number of copies of one or more sections of the DNA.
– CNVs correspond to relatively large regions of the genome that have been deleted (fewer than the normal number) or duplicated (more than the normal number) on certain chromosomes.
– For example, the chromosome that normally has sections in order as A-B-C-D might instead have sections A-B-C-C-D (a duplication of "C") or A-B-D (a deletion of "C").
• This variation accounts for roughly 13% of human genomic DNA and each variation may range from about one kilobase (1,000 nucleotide bases) to several megabases in size (Pawel et al, 2010)

• Genome-wide association study (GWA study, or GWAS), also known as whole genome association study (WGA study, or WGAS) or common-variant association study (CVAS), is an examination of many common genetic variants in different individuals to see if any variant is associated with a trait.
– GWAS typically focus on associations between single-nucleotide polymorphisms (SNPs) and traits like major diseases.
– compare the DNA of two groups of participants: people with the disease (cases) and similar people without (controls). Each person gives a sample of DNA, from which millions of genetic variants are read using SNP arrays.
– If one type of the variant (one allele) is more frequent in people with the disease, the SNP is said to be "associated" with the disease.
• A polygene, multiple factor, multiple gene inheritance, or quantitative gene is a group of non-allelic genes that together influence a phenotypic trait. The precise loci or identities of the non-allelic genes are often unknown to biologists.

• The common disease-common variant (often abbreviated CD-CV) hypothesis predicts that common disease-causing alleles, or variants, will be found in all human populations which manifest a given disease.
– Common variants (not necessarily disease-causing) are known to exist in coding and regulatory sequences of genes.
– According to the CD-CV hypothesis, some of those variants lead to susceptibility to complex polygenic diseases.
– Each variant at each gene influencing a complex disease will have a small additive or multiplicative effect on the disease phenotype
• Pleiotropy occurs when one gene influences multiple, seemingly unrelated phenotypic traits, an example being phenylketonuria, which is a human disease that affects multiple systems but is caused by one gene defect.

SUB-GROUPS OF GENETICS IN SCZ
• Biochemical genetics: biochemical reactions by which genetic determinants are replicated and produce their effects
• Developmental genetics: how the expression of normal genes controls growth and developmental processes
• Molecular genetics: structure and functioning of genes at molecular level.
• Cytogenetics: chromosomes
• Population genetics: mathematical properties of genetic transmission in families and populations- evolutionary genetics, genetic demography, quantitative genetics, genetic epidemiology
• Quantitative genetics: goal is to partition the observed variation of phenotypes into genetic and environmental components
• Genetic epidemiology: understanding the causes, distribution and control of disease in groups of relatives and the multifactorial causes of disease in populations

GENETIC BASIS OF SCHIZOPHRENIA

FAMILY STUDIES
• Study of families of probands to see if the relatives of the probandshave increased risk of developing the disease
• Ernst Rudin(1916): first systematic family study
• Other prominent researchers: Edith Zerbin-Rudin, Irving I. Gottesman, Franz Kallmann, Manfred Bleuler
• Data showed the familial basis of schizophrenia with increased risk of developing schizophrenia in the relatives of schizophrenic patients
• Bezugsziffer: an age-adjusted size of the sample which takes into account the fact that younger persons have not passed through the full period of risk- traditionally 15-39yrs
• Average lifetime prevalence risk of 10% in siblings and children
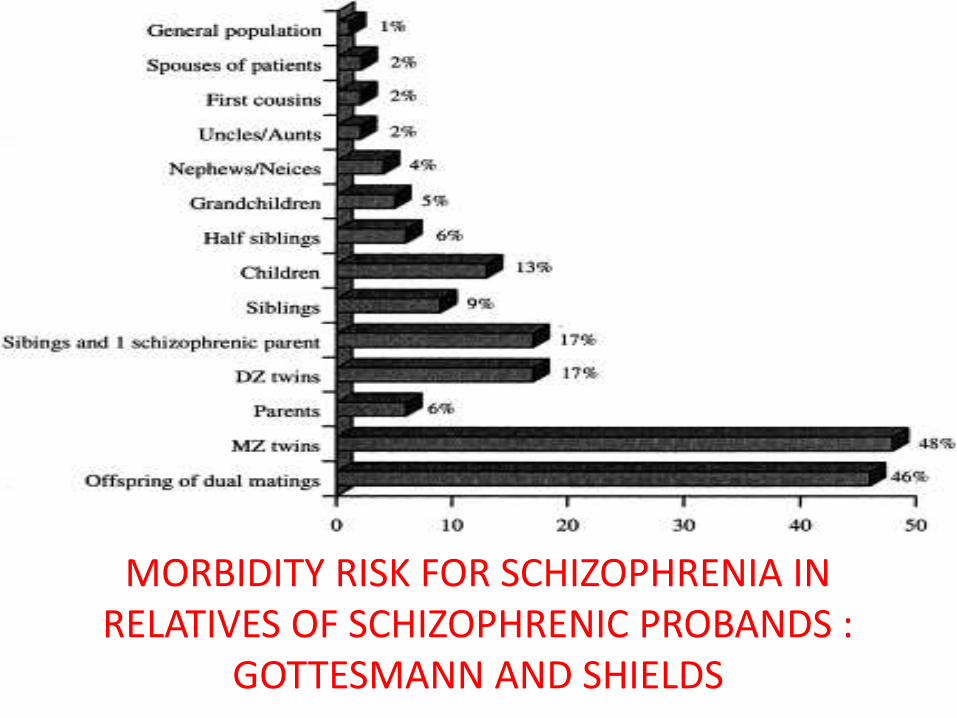
MORBIDITY RISK FOR SCHIZOPHRENIA IN RELATIVES OF SCHIZOPHRENIC PROBANDS :
GOTTESMANN AND SHIELDS

• Lower risk among parents explained by the reduced reproductive fitness associated with schizophrenia and the possibility of de novo mutations causing the illness in the offspring*
• Earlier studies questioned on the methodological grounds: no control groups, diagnoses not made blind, no structured interviews or operationalized diagnostic criteria used
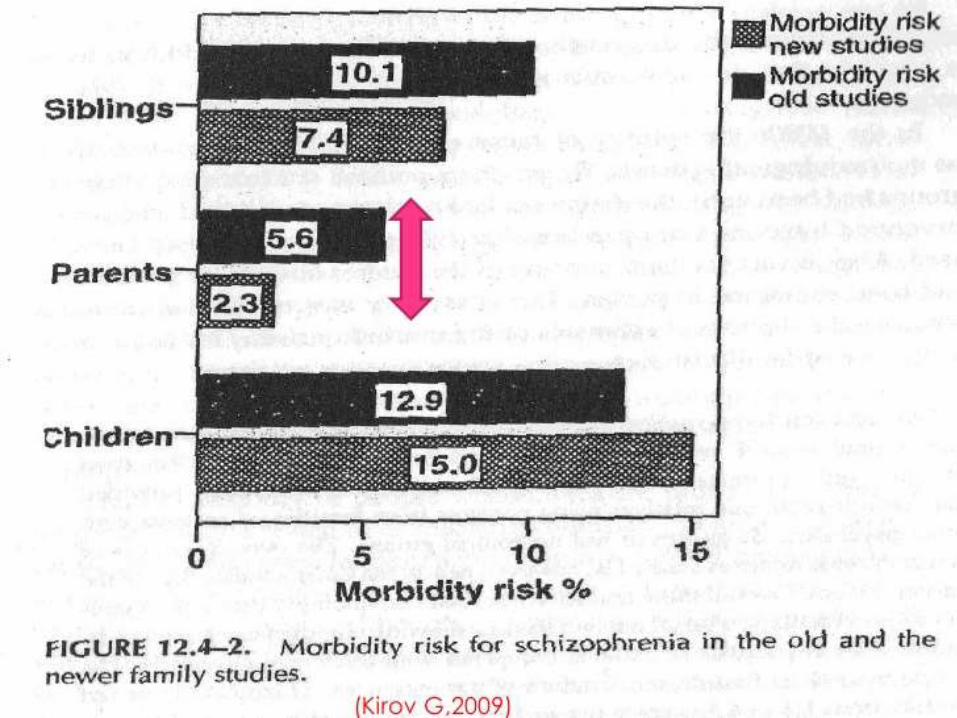

• No major difference in the findings of newer studies as compared to that of older ones
• Significant difference for parents(5.6% in older ones vs. 2.3% in newer) could be due to application of more stringent diagnostic criteria*
• Confirmed the higher risk of SCZ in the relatives of the probands but did not delineate the role of shared genetic or environmental factors in this difference
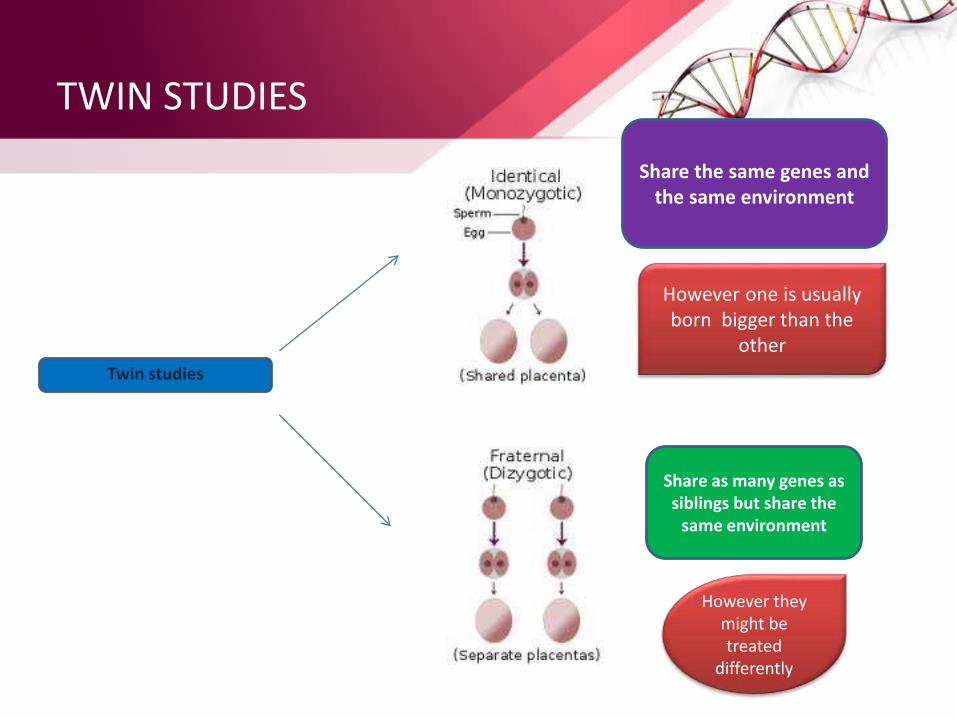
Twin studies
Share the same genes and the same environment
However one is usually born bigger than the
other
Share as many genes as siblings but share the
same environment
However they might be treated
differently
TWIN STUDIES

• Compare the concordance rates in Monozygotic(MZ) and Dizygotic(DZ) twins
• MZ twins share all their genes while DZ twins share on average 50% of their genes
• Assuming twins share common environment,
– higher concordance in MZ twins than in DZ implies genetic origin
– Concordance of less than 100% in MZ twins indicate the role of environment

•Gottesman and Shields reviewed the results of 5 twin studies looking for concordance rates for schizophrenia. These studies looked at 210 MZ twins and 319 DZ twins.It was found that in MZ twins there was a concordance rate of 35-58% compared with dizygotic (DZ) twin rates that ranged from 9-26%. They also found a concordance rate in MZ twins of 75-91% when the sample was restricted to the most severe form of schizophrenia.
•Cardno (2002) – showed concordance rate of 26.5% MZ and 0% for DZ

• Pairwise concordance: simply the number of concordant pairs divided by the total number of pairs
• Probandwise concordance: each of the concordant twin is counted i.e. the pair is counted twice and is ascertained by – number of affected co-twins/ the number of probands– Gives the risk for the twin of a person suffering from SCZ to
become ill him/her-self – Preferred by geneticists- technically more correct and directly
comparable to population risks reported in family studies• Probandwise concordance in MZ twins: 25%(n=8)(Essen-
Moller,1970) 78%(n=245)(Kallmann,1946)• Probandwise concordance in same-sex DZ twins:
0%(n=50)(Cardno,1998)- 28% (n=25)(Franzek and Beckmann,1998)• Heritability: 41%- 90%- very similar estimates with both the
methodologically superior and inferior studies (Sullivan)

• Meta-analyses of heritability: – Sullivan- 81% (Sullivan, 2007) – Cardno and Gottesmann- 88% (Cardno, 2000)
• High agreement between studies conducted in different countries over nearly a century
• Calculation based on assumptions: – Polygenic multifactorial threshold model– Similar risk of SCZ in twins as in general population – DZ and MZ twins share similar environment
• Identical twins reared apart: theoretically nullifies the effects of shared environment (Gottesmann and Shields) = 64%
• Possible explanations for the discordance in MZ twins:– Affected co-twin suffers from an environmentally determined form of
the disorder – Both twins inherited the same genetic liability but only expressed in
the affected twin• Explained by the study of risk in the offspring of discordant SCZ twins

• First study – 11 SCZ twins, 47 offspring with 6 developing SCZ = 16.8% – 6 unaffected twins, 24 offspring with 4 developing SCZ = 17.4%
• Second study – 28 offspring of SCZ twins with 3 developing SCZ = 10.7% – 45 offspring of unaffected twins with 1 developing SCZ = 2.2%
• Combined: – risk among offspring of SCZ twins, 9/75 = 12%– Risk among offspring of unaffected, 5/69 = 7.2%
• (P=0.38) No significant difference, similar morbidity risk consistent with unexpressed risk
(Kirov , 2009)

ADOPTION STUDIES
• Allow dissection of genetic from environmental contributions in ways that twin studies cannot.
• If there is a genetic component to the disorder, the similarity between the adopted children and their biological parents should be higher than the similarity between adopted children and their adoptive parents.
• Adoption itself does not increase the risk for developing SCZ among adopted children.
• Leonard Heston, 1966: – 47 adopted children of mothers suffering from SCZ and other
psychoses, separated within 3days of birth- by age 36, 5 developed SCZ (10.6%)
– None out of 50 children in the control group had SCZ• David Rosenthal, 1971:
– Studied 5500 adoptees 14/52 (26.9%) children of SCZ parents had SCZ-spectrum disorder
– 12/67 control (17.9%) had illness of similar spectrum

• M. W. Higgins, 1976:
– 50 children of SCZ mothers
– 4/23 (17.9%) children reared by SCZ mothers had SCZ
– Of 25 children adopted away, 4(16%) SCZ
• Seymour S. Kety et al, 1994
– 14/279 (5%) biological relatives of SCZ adoptees had chronic SCZ
– None out of 111 adoptive relatives had SCZ
– 1/351 (0.3%) biological + adoptive relatives of unaffected adoptees had SCZ
• Pekka Tienari, 2000:
– 164 adopted children of mothers suffering from SCZ
• SCZ- 11(6.7%)
• Schizoaffective- 1(0.6%)
• Schizotypal personality disorder- 4(2.4%)
• Overall narrow spectrum SCZ disorder- 10.4%
– 197 control adoptees
• SCZ - 4(2%)

• CONCLUSION:
– Overall at least 10% risk of developing SCZ and other narrow-spectrum SCZ disorders in adopted away children of SCZ parents
– Risk similar to that in offspring in family studies -> genetic basis of transmission of SCZ

LINKAGE STUDIES
• Use of large number of small families containing individuals who are definitely affected rather than large, multigenerational pedigrees
• At least 27 whole genome studies that analyzed between 1 to 294 pedigree containing between 32 to 669 patients of SCZ
• J. A. Badner and E.S. Gershon - susceptibility genes on chromosomes -› 8p, 13q and 22q
• Cathryn M. Lewis et al ->
– Strong evidence for 2q
– 1q, 3p, 5p, 6p, 8p, 11q, 14q, 20q and 22q
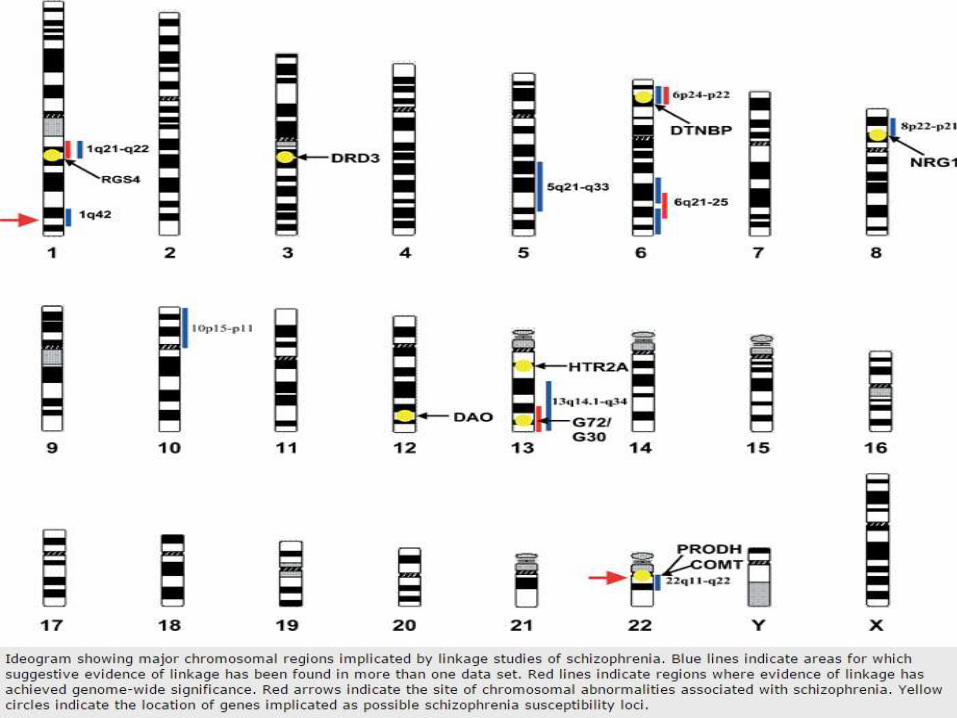

• Meta-analysis by Sullivan- › Only 42%, 35%, 14%, 6% and 3% of all known genes were implicated by zero, one, two, three and four linkage studies
• It is an imprecise tool
• Possible explanations for the varied linkage findings:
– Different genes operate in different populations
– SCZ is caused by the effect of many genes of small effect, so studies had no power to detect the loci

MODE OF INHERITANCE
• Hypotheses regarding genetic background of common diseases including SCZ:– Common disease/common variant hypothesis:
• Common diseases caused by common variants • Joint action of several common genetic variants, each has a small
effect on disease susceptibility, together with environmental factors • Could range into thousands
– Multiple rare variants in different genes, which have low population frequencies, operate in different individuals: • lack of families with clear cut Mendelian Inheritance • Inability of linkage studies to find any causative mutation • Mathematical modeling is inconsistent with single gene of large effect • A small number of cases of SCZ could be due to rare chromosomal
aberrations with high penetrance
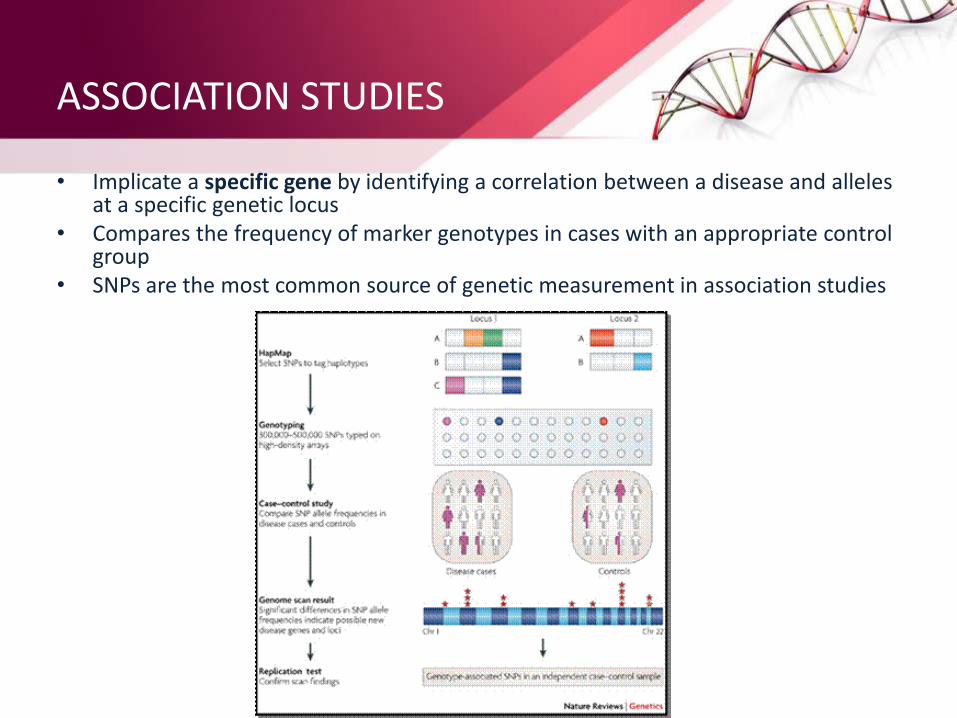
ASSOCIATION STUDIES
• Implicate a specific gene by identifying a correlation between a disease and alleles at a specific genetic locus
• Compares the frequency of marker genotypes in cases with an appropriate control group
• SNPs are the most common source of genetic measurement in association studies

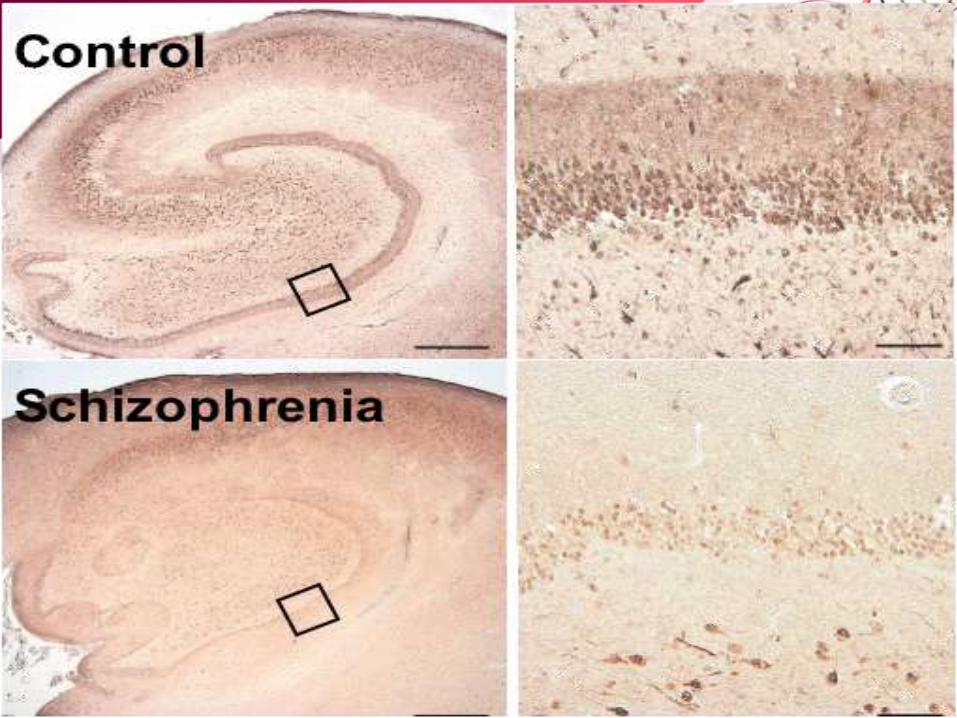
• Dystrobrevin-binding protein1(DTNBP1)/dysbindin– First reported by Richard E. Straub and colleagues in 2002 – On chromosome 6p22.3 – Konrad Talbot(2004)- presynaptic dystrobrevin-independent fraction
reduced in SCZ brain within certain glutamatergic neurons in the hippocampus-> Associated with increased expression of vesicular glutamate transporter type 1 ->Alteration in presynaptic glutamate function
– The expression of dysbindin proteins is decreased in the brains of schizophrenia patients, and neurons in mice carrying a deletion in the dysbindin gene have fewer dendritic spines (Jie-Min Jia et al, 2014)
– Significant associations found between SCZ and several SNPs and multimarker haplotypes spanning DTNBP1
– Support from other large studies as well - at least 10 studies – Some studies showing no association– Inconsistencies indicative of presence of multiple susceptibility and
protective alleles

• Neuregulin 1(NRG1) – Encodes multiple proteins with diverse range of functions in the
brain • Cell-cell signaling , ErbB receptor interactions ,Axon guidance,
Synaptogenesis, Glial differentiation, Myelination, Neurotransmission • Located on 8p21-22
– Plays a role in synaptic plasticity. It has been shown that a loss of Neuregulin 1 within cortical projection neurons results in increased inhibitory connections and reduced synaptic plasticity
– First implicated from linkage study in Icelandic sample– Further positive findings supported by studies from UK, Irish,
Chinese, Bulgarian and South African samples– Only 3 other studies replicating the specific haplotype ->
differences in linkage disequilibrium

• Other genes:
– Catechol-O-Methyltransferase (COMT)
– Proline Dehydrogenase (PRODH)
– Regulator of G-protein signaling4 (RGS4)
– D-Amino-acid oxidase (DAO)
– D-Amino-acid oxidase activator (DAOA)
– G72/G30
– CAPON
– AKT1

GWAS
• Common variant SNPs: – minor allele frequency of SNPs>0.05– The effect sizes of the associations likely to be very small, so
hundreds and even thousands of genes might contribute small effects to the pathogenesis of SCZ
• RELN gene: – › Relin protein- a serine protease important in corticogenesis
(Hong et al, 2000) – › Implicated in neurotransmitter-related GSK3β signaling and
regulation of NMDA receptor activation (Herz, 2006) – › Polymorphism in RELN associated with neurocognitive
endophenotypes of SCZ (working memory and executive functioning) (Wedenoja, 2008)

• Genome-pooling based study – rs11064768 in intron 1 of CCDC60 on 12q24.23 – rs11782269 on 8p23.1 – RBP1 on 3q23- implicated in SCZ pathogenesis – Not replicated in other studies and no genome-wide significance– Suggestive of neurodevelopmental hypothesis of SCZ (Keshavan et al
2004)
• Zinc finger protein 804A(ZNF804A)– › Located on 2q32.1 – › Putative transcription factor – › Shown to be associated with disturbed connectivity between the
dorsolateral prefrontal cortex (DLPFC) & the hippocampus; between rt. and lt. hemisphere (Esslinger et al 2009)
– › Strong association with SCZ and BPAD – › Supported by replication in other large studies as well
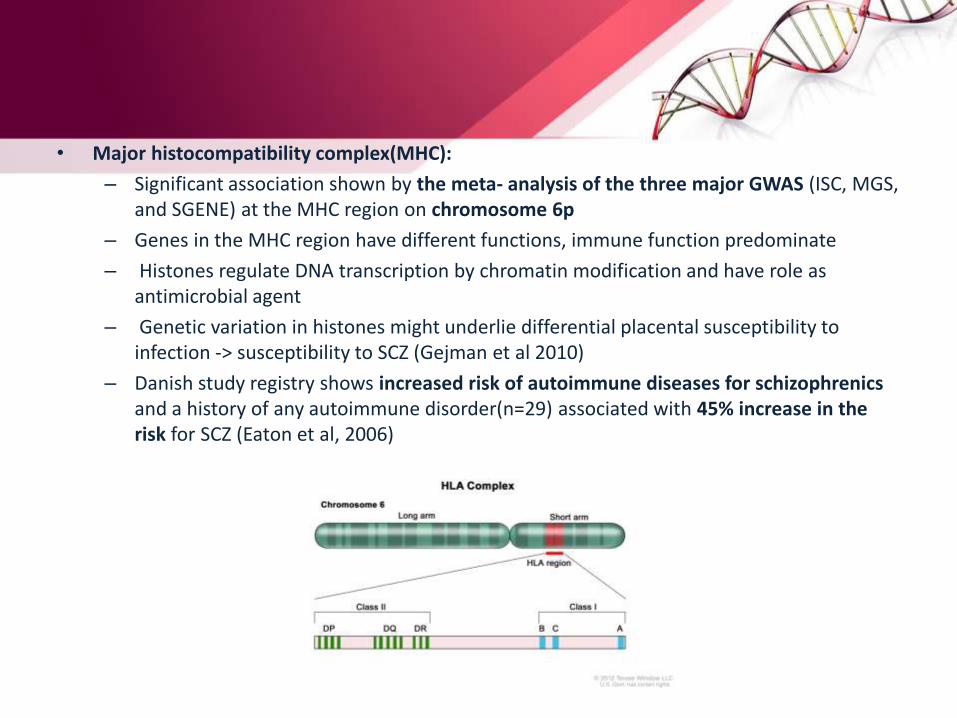
• Major histocompatibility complex(MHC):
– Significant association shown by the meta- analysis of the three major GWAS (ISC, MGS, and SGENE) at the MHC region on chromosome 6p
– Genes in the MHC region have different functions, immune function predominate
– Histones regulate DNA transcription by chromatin modification and have role as antimicrobial agent
– Genetic variation in histones might underlie differential placental susceptibility to infection -> susceptibility to SCZ (Gejman et al 2010)
– Danish study registry shows increased risk of autoimmune diseases for schizophrenicsand a history of any autoimmune disorder(n=29) associated with 45% increase in the risk for SCZ (Eaton et al, 2006)

• Neurogranin(NRGN):
– On chromosome 11
– Encodes a postsynaptic protein kinase substrate that binds calmodulin, mediating NMDA receptor signaling
– important for learning & memory, relevant to proposed glutamate pathophysiology of SCZ (Wang et al, 2008)
• Transcription factor 4 (TCF4):
– › On chromosome 18
– › Neuronal transcriptional factor essential for brain development, esp. neurogenesis
– › Mutations cause Pitt-Hopkins syndrome, a neuro-developmental disorder

• International Schizophrenia Consortium ( ISC) conclusion: – › Thousands of common polygenic variants with very small individual effects
explain about 1/3rd of the total variation in genetic liability to SCZ (Purcell et al, 2009)
– › The remaining heritability is still missing even after the well powered GWAS studies

Chromosomal aberrations and CNVs
• CNVs: – › are stretches of genomic deletions & duplications ranging
from 1kb to several Mb – › Likely to have larger phenotypic effects than SNPs – › Only rare(<1%) and large(>100kb) CNVs have been
implicated in SCZ
• Chromosome 22q11.21 deletion syndrome • Velocardiofacial syndrome • Increased risk for SCZ , >30% carriers develop
psychosis,80% of which as SCZ • Largest known individual risk factor for SCZ second only
to having an identical twin with SCZ

• 2p16.3 Neurexin1 (NRXN1) deletion:– NRXN presynaptic cell adhesion molecule– Interacts with post synaptic cell adhesion molecules including neuroligins– Believed to play important role in release of neurotransmitter from
presynaptic vesicles and together with neuroligins involved in synapse formation and use-dependent validation of neural circuits
– Partial overlapping observed in mentally retarded and autistic patients
• Disrupted In Schizophrenia (DISC1): – Balanced chromosomal translocation in(1,11) (q42;q14.3) – Disrupt two genes in chromosome1 : DISC1 & DISC2 – Strong evidence of linkage to- SCZ, BPAD and recurrent depression– May contribute to SCZ by affecting neuronal functions dependent on intact
cytoskeletal regulation such as • Neuronal migration• Neurite architecture • Intracellular transport

OTHER CNVs:
• 15q13.1 duplication
– Duplicated interval in the proband contains 3 genes of which APBA2 appears most likely
– APBA2 interacts with NRXN1
• 1q21.1 deletion
• 15q13.2 deletion
• 15q13.3 deletion
– Also found in pts. With mental retardation and seizures
• 16p11.2 duplication

Pleiotropy and overlap with BPAD/Autism
• Many of the genes positive for SCZ also positive for BPAD and vice versa
– With BPAD • DISC1
• NRG1
• RELN
• ANK3
– With autism - Neurexin 1
• Could have implication for diagnostis of SCZ and also lead to newer etiological and pathophysiologicalexplanations of the psychiatric disorders
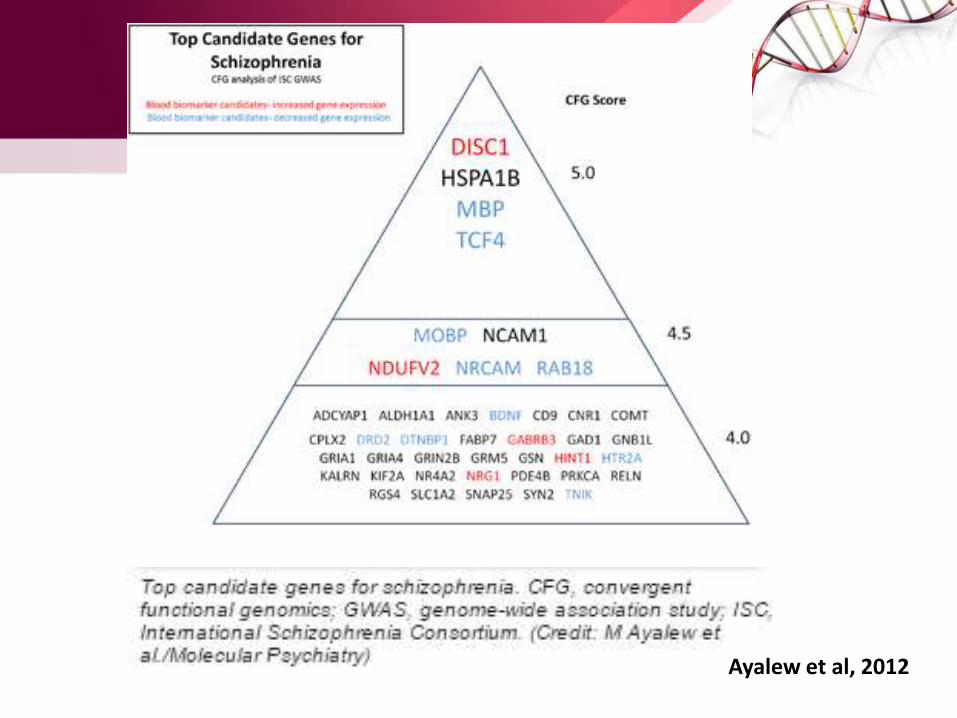
Ayalew et al, 2012

FUTURE DIRECTIONS
• Standardization of the phenotypes: – › Syndromal diagnosis – › Broad variations
• The effect size associated with common variant is very low and the number of total susceptibility variants may be in the order of thousands requiring upto 100,000 cases and controls for replicating the findings – › To achieve such sample sizes with detailed and consistent
phenotype measurement is a challenge• Combining all diseases of a spectrum e.g. psychosis, broadly
can give large sample size and also detect genes that overlap• Narrow the phenotype ->more homogenous sub-group ->
smaller number of genes of greater effect sizes

• Use of endophenotypes (Gould 2006): – Disease associated phenotypes that are heritable, state independent,
cosegregate with families and also found in unaffected family members • E.g. Abnormal eye movement while tracking a moving object in screen • Neurocognitive deficits- COMT & RELN• Structural imaging phenotypes
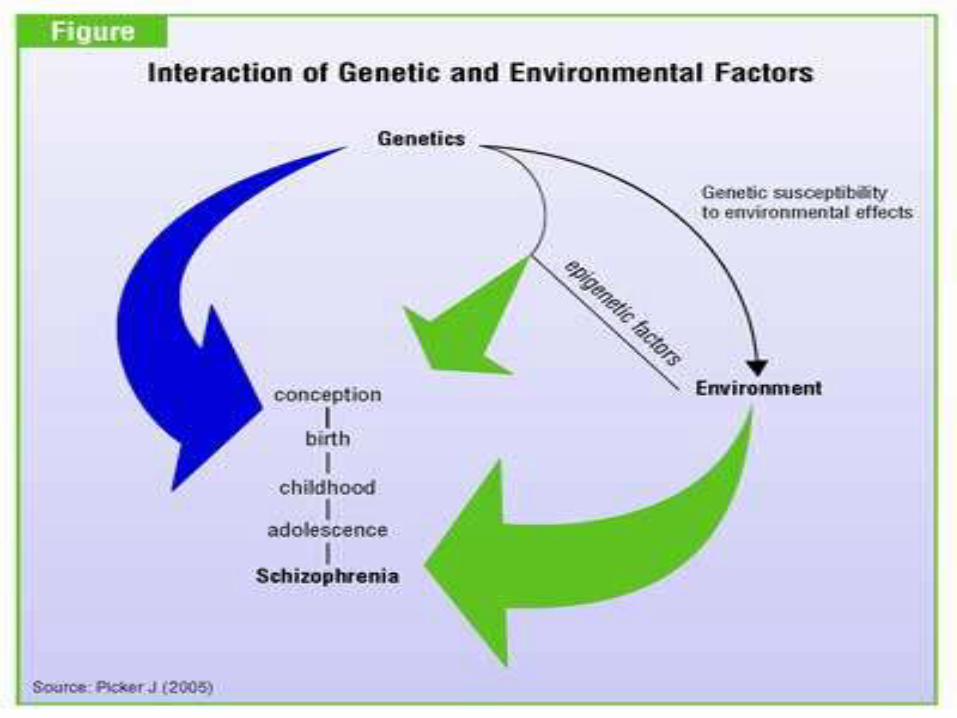
– Creates phenotypically more homogenous group• Consideration of the effect of environmental factors such as maternal
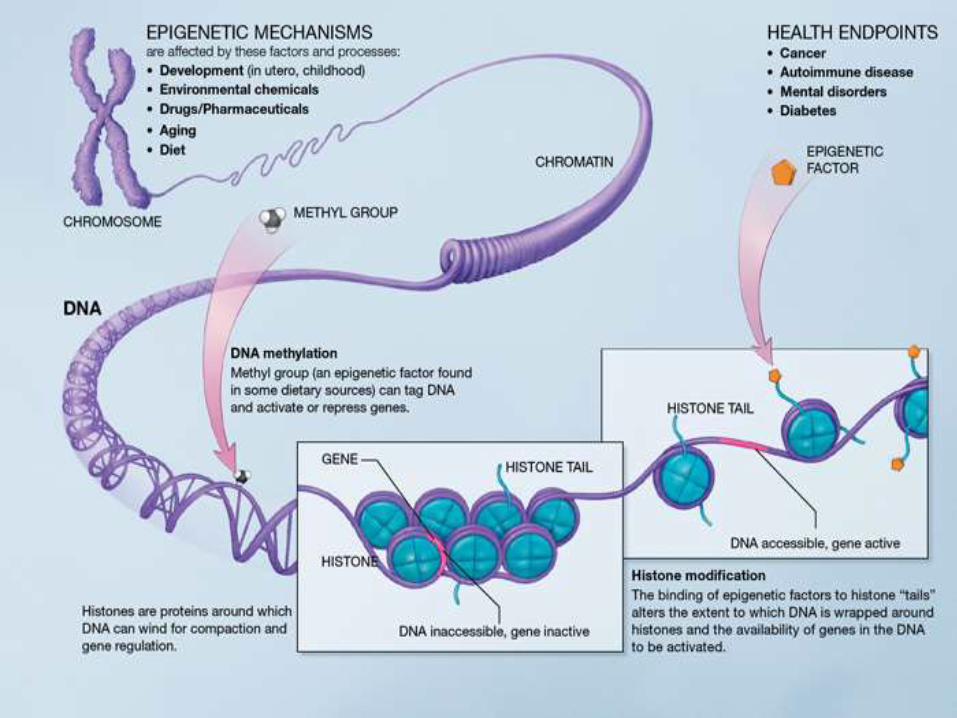
infections, and drug use• Consideration of epigenetic mechanisms• Use of high-throughput whole-genome sequencing: Has potential to
detect virtually all SNPs, CNVs and epigenetic modifications*, Will provide comprehensive information of an individual at the DNA level. Cost is a concern.
• More accurate study of the target organ in SCZ research (brain) can lead to more objective and reliable associations with the genetic variants

REFERENCES• Sullivan PF. The psychiatric GWAS consortium: Big science comes to psychiatry. Neuron
2010;68(2): 182-186
• Psychiatric GWAS Consortium Coordinating Committee. Genomewide association studies: history, rationale, and prospects for psychiatric disorders. Am J Psychiatry 2009;166:540-556
• Kirov G, Owen MJ: Genetics of schizophrenia. In: Sadock BJ, Sadock VA, Ruiz P, editors.Kaplanand Saddock’s Comprehensive Textbook of Psychiatry.9th edition. Lippincot Williams and Wilkins; Philadelphia;2009.p 1462-1475
• Sullivan PF, Kendler KS, Neale MC: Schizophrenia as a complex trait. Evidence from a meta-analysis of twin studies. Arch Gen Psychiatry. 2007;60:1187. Cited in: Kirov G, Owen MJ: Genetics of schizophrenia. In: Sadock BJ, Sadock VA, Ruiz P, editors.Kaplan and Saddock’sComprehensive Textbook of Psychiatry.9th edition. Lippincot Williams and Wilkins; Philadelphia;2009.p 1462-1475.
• Cardno AG, Gottesman II: Twin studies of schizophrenia: from bow- and-arrow concordances to star war MMx and functional genomics. Am J Med Genet. 2000;97:12. Cited in: Kirov G, Owen MJ: Genetics of schizophrenia. In: Sadock BJ, Sadock VA, Ruiz P, editors.Kaplan and Saddock’s Comprehensive Textbook of Psychiatry.9th edition. Lippincot Williams and Wilkins; Philadelphia;2009.p 1462-1475.
• Pawel Stankiewicz, James R. Lupski (2010). "Structural Variation in the Human Genome and its Role in Disease". Annual Review of Medicine 61: 437–455.
• Stearns, F. W. (2010). One Hundred Years of Pleiotropy: A Retrospective. Genetics 186(3):767-773. http://www.genetics.org/content/186/3/767.abstract

• Hong SE, Shugart YY, Huang DT, et al. Autosomal recessive lissencephaly with cerebellarhypoplasia is associated with human RELN mutations. Nat Genet. 2000;26:93-96. Cited in: Tiwari AK, Zai CC, Muller DJ, Kennedy JL. Genetics in schizophrenia: where are we and what next? Dialogues in clinical neuroscience 2010;3(12):289-303.
• Herz J, Chen Y. Reelin, lipoprotein receptors and synaptic plasticity. Nat Rev Neurosci. 2006;7:850-859. Cited in: Tiwari AK, Zai CC, Muller DJ, Kennedy JL. Genetics in schizophrenia: where are we and what next? Dialogues in clinical neuroscience 2010;3(12):289-303.
• Wedenoja J, Loukola A, Tuulio-Henriksson A, et al. Replication of link-age on chromosome 7q22 and association of the regional Reelin gene with working memory in schizophrenia families. Mol Psychiatry. 2008;13:673-684. Cited in: Tiwari AK, Zai CC, Muller DJ, Kennedy JL. Genetics in schizophrenia: where are we and what next? Dialogues in clinical neuroscience 2010;3(12):289-303.
• Keshavan MS, Kennedy JL, Murray R, eds. Neurodevelopment and Schizophrenia. Cambridge, UK: Cambridge University Press; 2004. Cited in: Tiwari AK, Zai CC, Muller DJ, Kennedy JL. Genetics in schizophrenia: where are we and what next? Dialogues in clinical neuroscience 2010;3(12):289-303
• Agarwal A, Zhang M, Trembak-Duff I, Unterbarnscheidt T, Radyushkin K, Dibaj P et al. (2014). "Dysregulated expression of neuregulin-1 by cortical pyramidal neurons disrupts synaptic plasticity". Cell Reports 8 (4): 1130–45.
• M Ayalew et al., Convergent functional genomics of schizophrenia: from comprehensive understanding to genetic risk prediction, Molecular Psychiatry, 2012

• Esslinger C, Walter H, Kirsch P, et al. Neural mechanisms of a genome-wide supported psychosis variant. Science. 2009;324:605. Cited in : Tiwari AK, Zai CC, Muller DJ, Kennedy JL. Genetics in schizophrenia: where are we and what next? Dialogues in clinical neuroscience 2010;3(12):289-303
• Gejman VP, Sanders AR, Duan J. the role of genetics in the etiology of Schizophrenia. Psychiatr Clin North Am. 2010; 33(1): 35–66
• Eaton WW, Byrne M, Ewald H, et al. Association of schizophrenia and autoimmune diseases: linkage of Danish national registers. Am J Psychiatry 2006;163:521. Cited in: Gejman VP, Sanders AR, Duan J. the role of genetics in the etiology of Schizophrenia. Psychiatr Clin North Am. 2010; 33(1): 35–66
• Purcell SM, Wray NR, Stone JL, et al. Common polygenic variation contributes to risk of schizophrenia and bipolar disorder. Nature 2009;460:748
• Gould TD, Gottesman, II. Psychiatric endophenotypes and the development of valid animal models. Genes Brain Behav. 2006;5:113- 119. cited in : Tiwari AK, ZaiCC, Muller DJ, Kennedy JL. Genetics in schizophrenia: where we are and what next. Dialogues in clinical neuroscience 2010;3(12): 289-303.
• Stahl SM, editor. Stahl’s Essential Psychopharmacology. 3rd edition. Cambridge University Press; New Delhi;2008. p. 318.

THANK YOU





























