Embed Size (px)
Citation preview

Lars Juhl JensenEMBL Heidelberg
Dynamic complex formationduring the yeast cell cycle

© Chen et al., Mol. Biol. Cell, 2004
A qualitative model of the yeast cell cycle
• Should be accurate even at the level individual interactions
• Provides a global overview of temporal complex formation
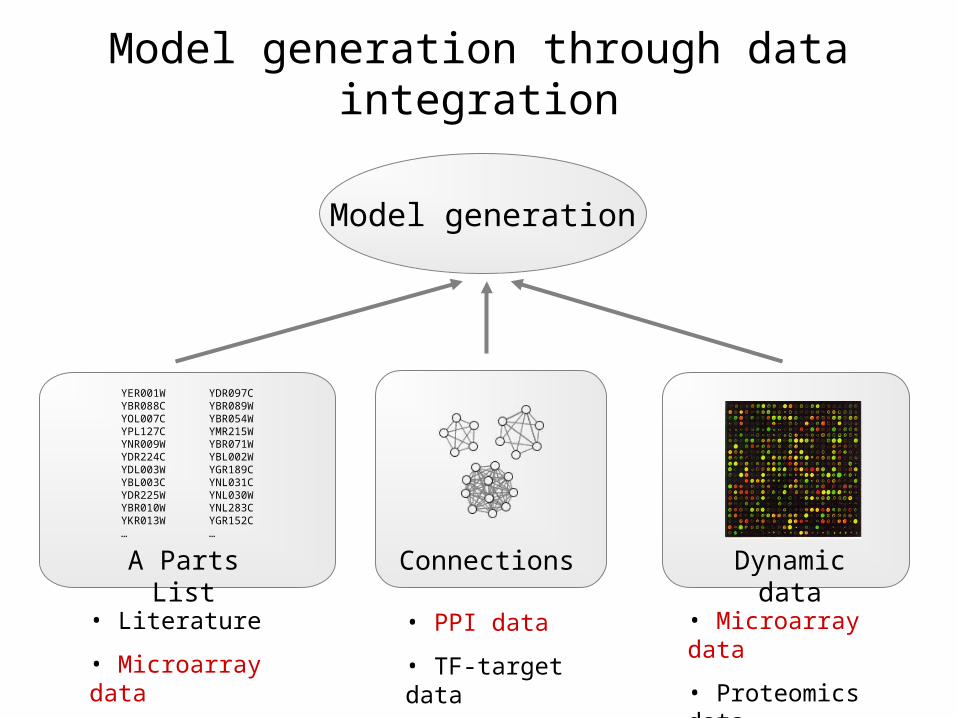
Model generation
A Parts List
• Literature
• Microarray data
Dynamic data
• Microarray data
• Proteomics data
• PPI data
• TF-target data
Connections
YER001WYBR088CYOL007CYPL127CYNR009WYDR224CYDL003WYBL003CYDR225WYBR010WYKR013W…
YDR097CYBR089WYBR054WYMR215WYBR071WYBL002WYGR189CYNL031CYNL030WYNL283CYGR152C…
Model generation through data integration
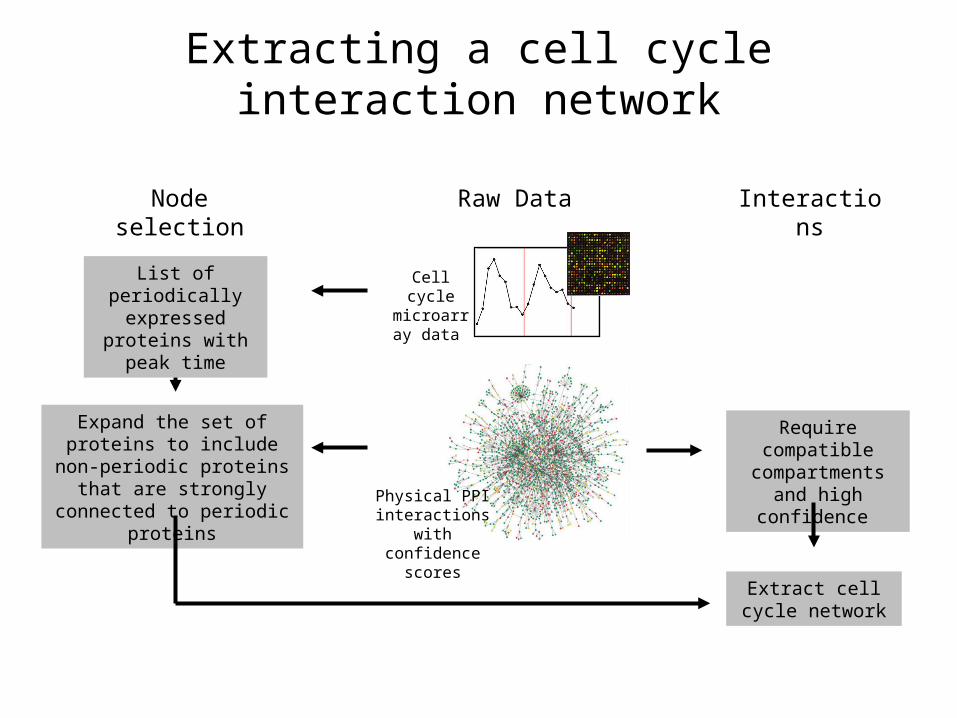
Cell cycle microarray
data
Physical PPI interactions with
confidence scores
Expand the set of proteins to include non-periodic proteins that are strongly connected
to periodic proteins
Raw DataNode selection
List of periodically expressed proteins
with peak time
Interactions
Require compatible compartments and high confidence
Extract cell cycle network
Extracting a cell cycle interaction network
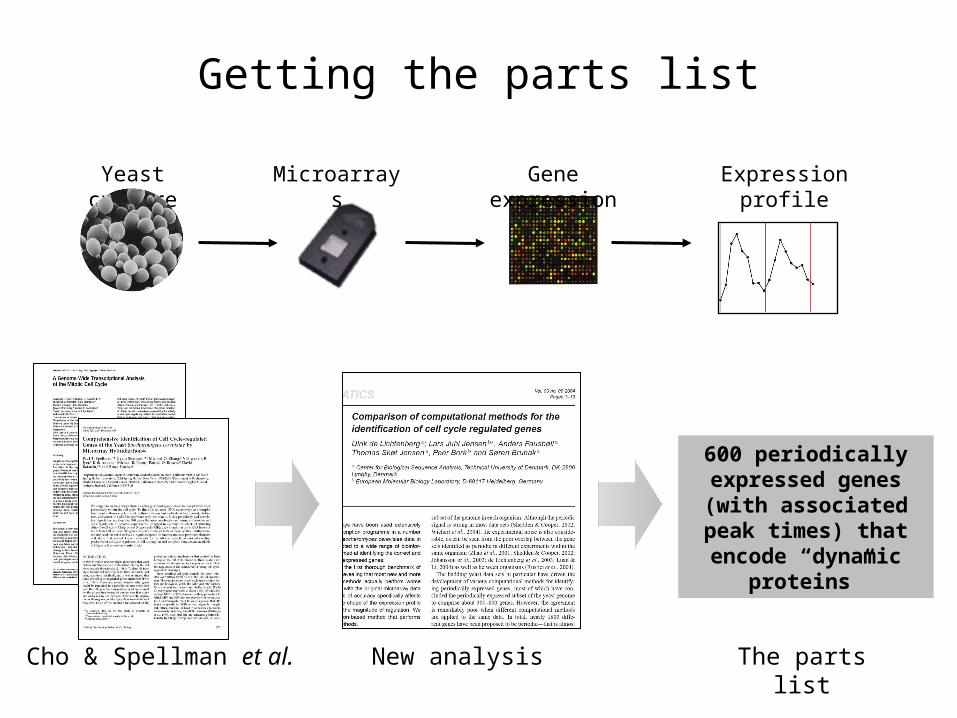
Cho & Spellman et al.
Yeast culture Microarrays Gene expression Expression profile
600 periodically expressed genes (with associated peak times) that encode “dynamic
proteins”
The parts listNew analysis
Getting the parts list

Topology based quality scores
• We developed scoring schemes for identifying sticky interactions
• Yeast two-hybrid data: S1 = -log((N1+1)·(N2+1))
• Scoring scheme for complex pull-down data: S2 = log[N12·N/((N1+1)·(N2+1))]
• Score calibration against KEGG A different raw quality score is
used for each evidence type Quality differences exist among
data sets of the same type

Filtering by subcellular localization
• Proteins cannot interact if they are not in the same place Large-scale subcellular localization screens have been made in yeast A matrix can be constructed that described the compartments between
which interactions should be allowed Two proteins cannot interact if no combination of observed subcellular
compartments allow for interaction
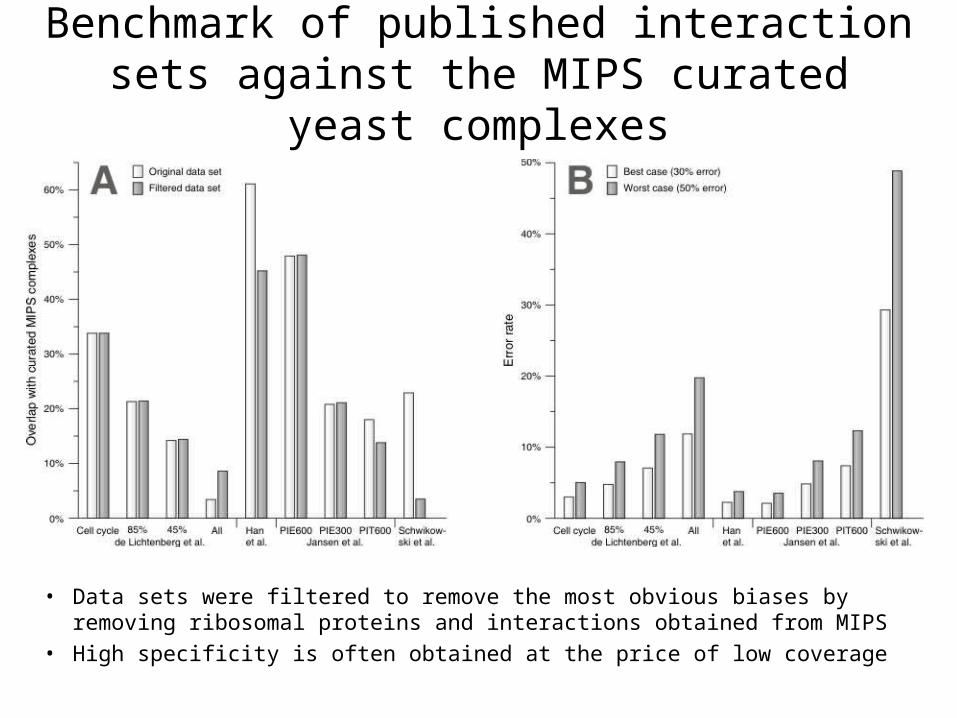
Benchmark of published interaction sets against the MIPS curated yeast complexes
• Data sets were filtered to remove the most obvious biases by removing ribosomal proteins and interactions obtained from MIPS
• High specificity is often obtained at the price of low coverage
G1
S
M
G2
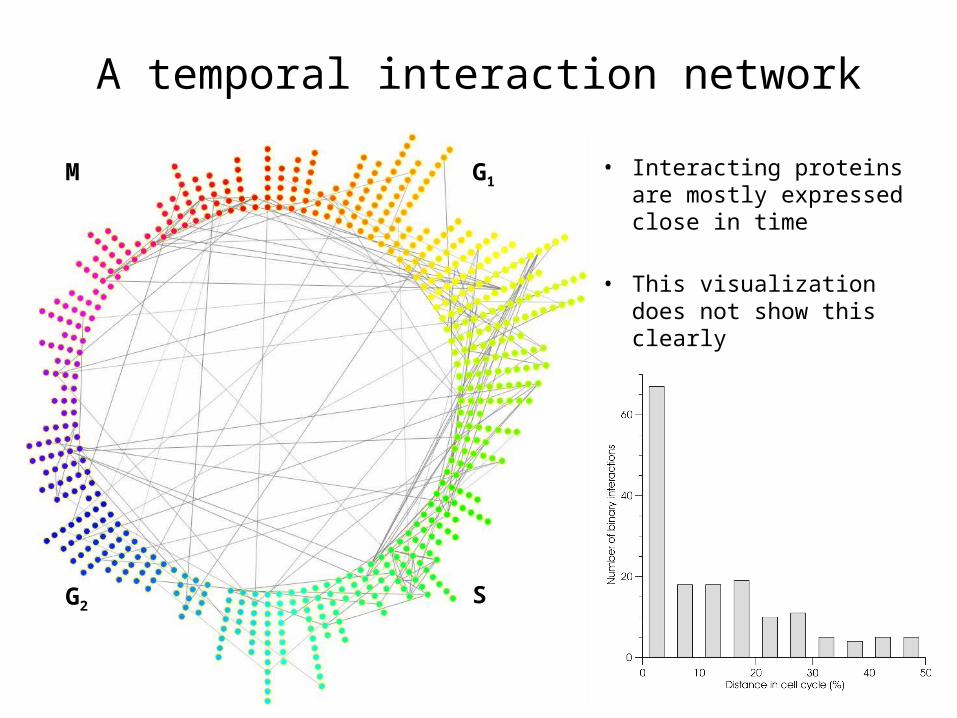
A temporal interaction network
• Interacting proteins are mostly expressed close in time
• This visualization does not show this clearly
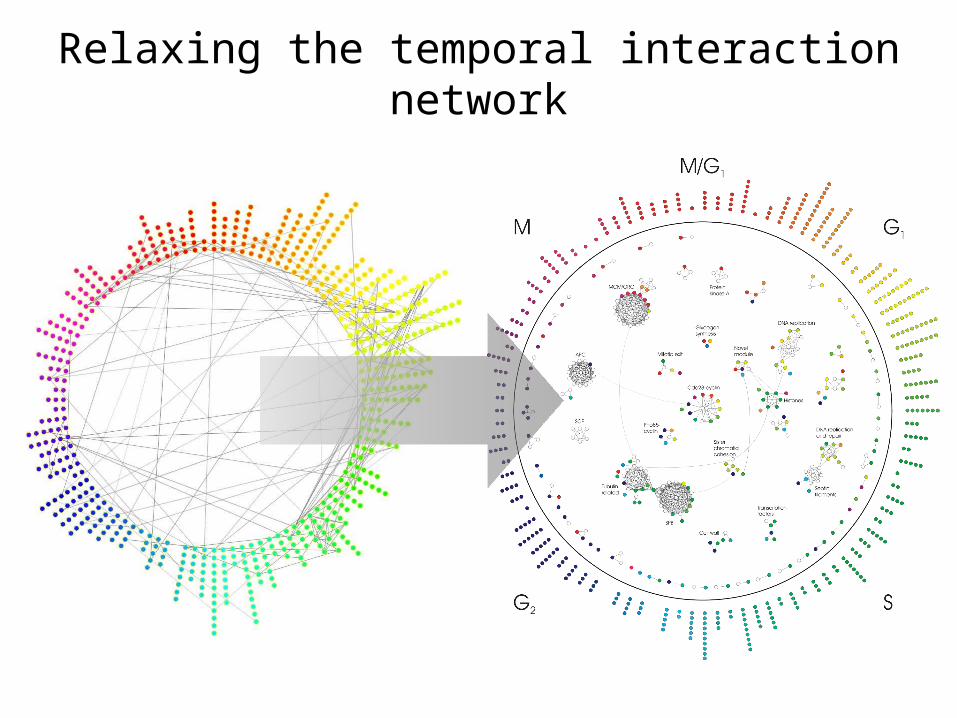
Relaxing the temporal interaction network
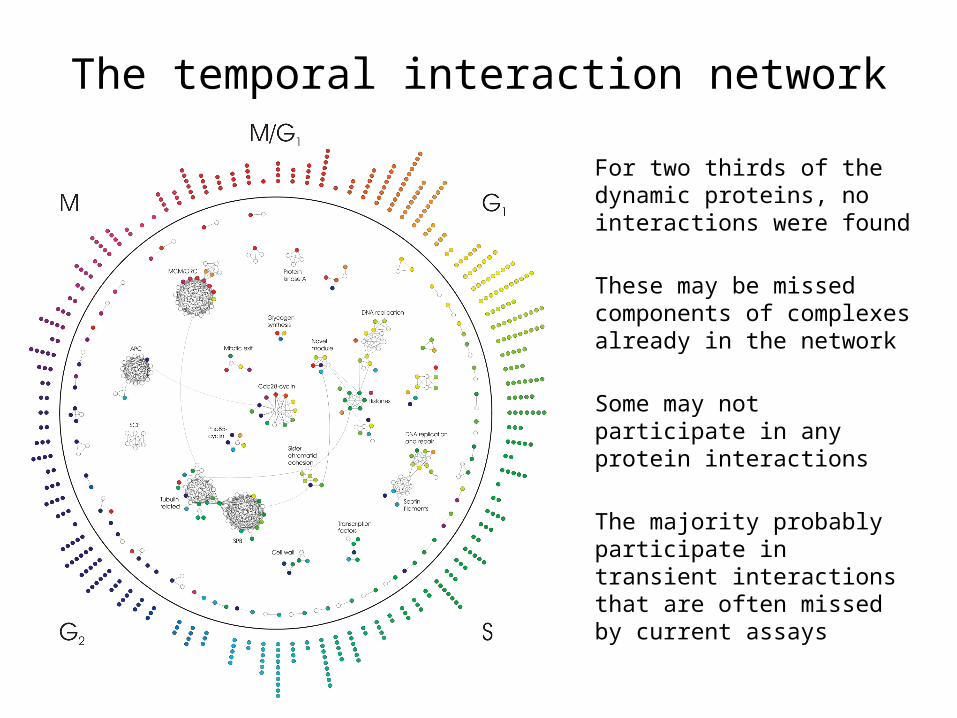
The temporal interaction network
For two thirds of the dynamic proteins, no interactions were found
These may be missed components of complexes already in the network
Some may not participate in any protein interactions
The majority probably participate in transient interactions that are often missed by current assays
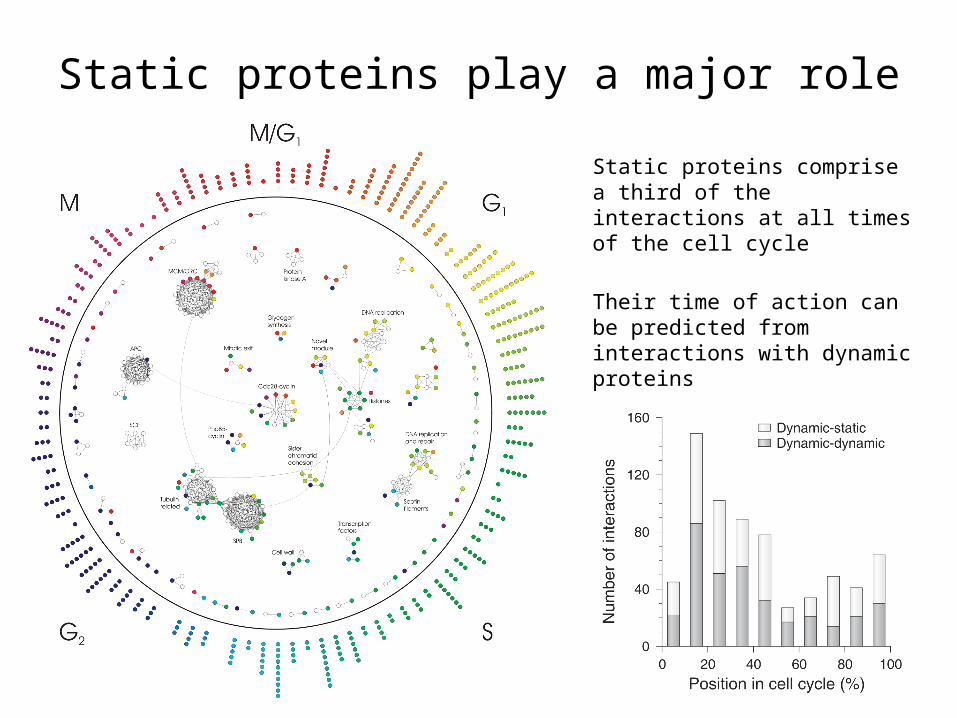
Static proteins play a major role
Static proteins comprise a third of the interactions at all times of the cell cycle
Their time of action can be predicted from interactions with dynamic proteins
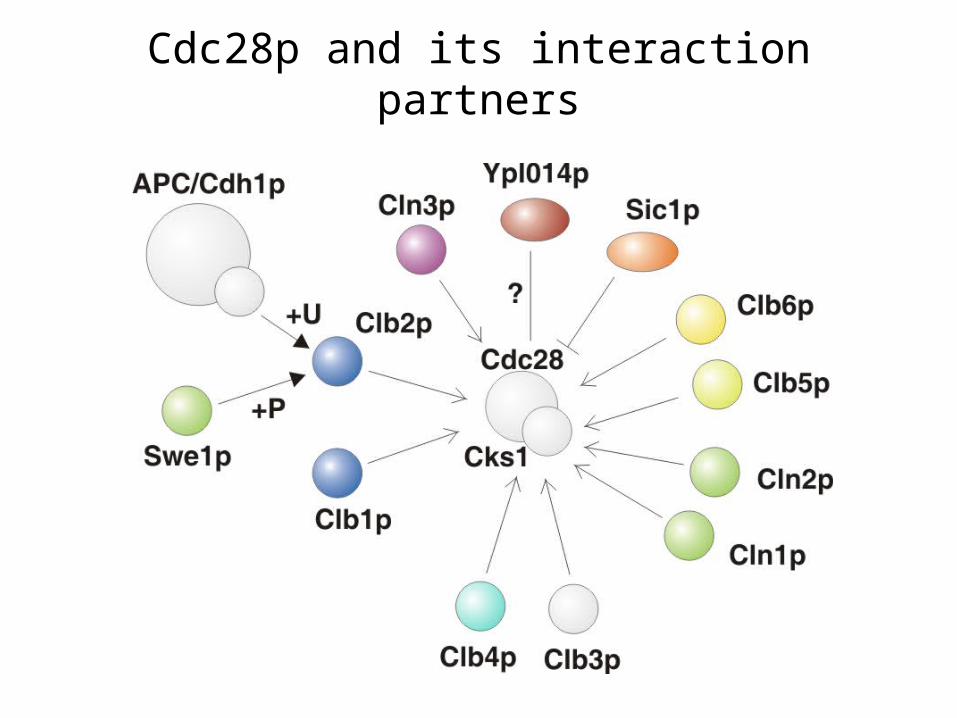
Cdc28p and its interaction partners

Pho85p and associated cyclins
Pcl7p has so far not been considered a cell cycle related cyclin, but we see it periodically expressed
We suggest the phosphoprotein Mmr1p to be a Pho85p-Pcl7p substrate
The phosphorylation of Swi5p by Pho85p-Pcl2p is correctly reproduced
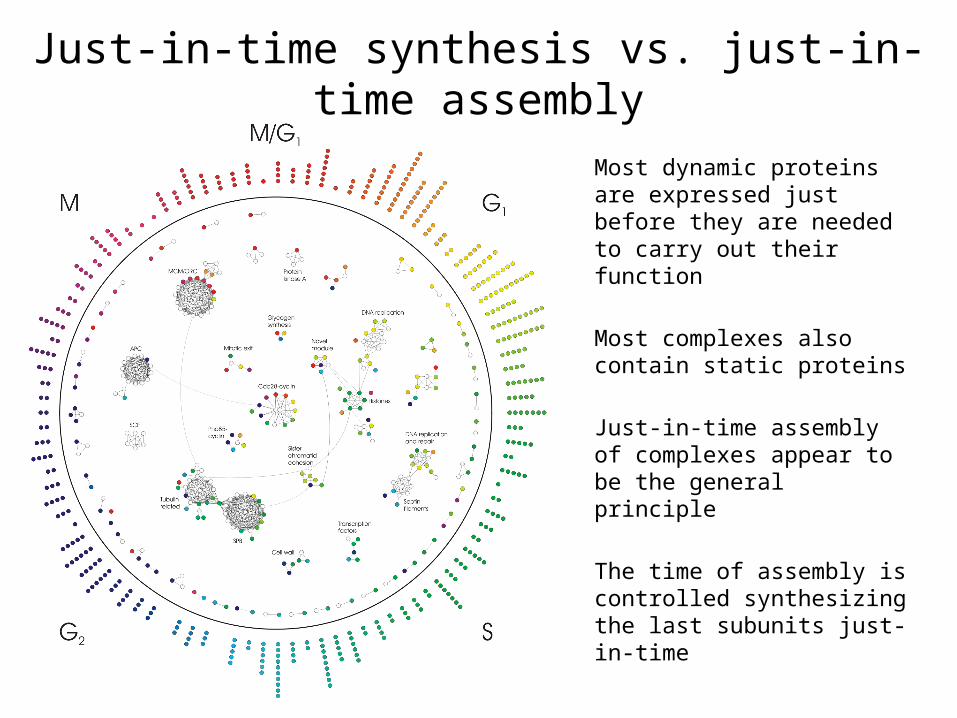
Just-in-time synthesis vs. just-in-time assembly
Most dynamic proteins are expressed just before they are needed to carry out their function
Most complexes also contain static proteins
Just-in-time assembly of complexes appear to be the general principle
The time of assembly is controlled synthesizing the last subunits just-in-time
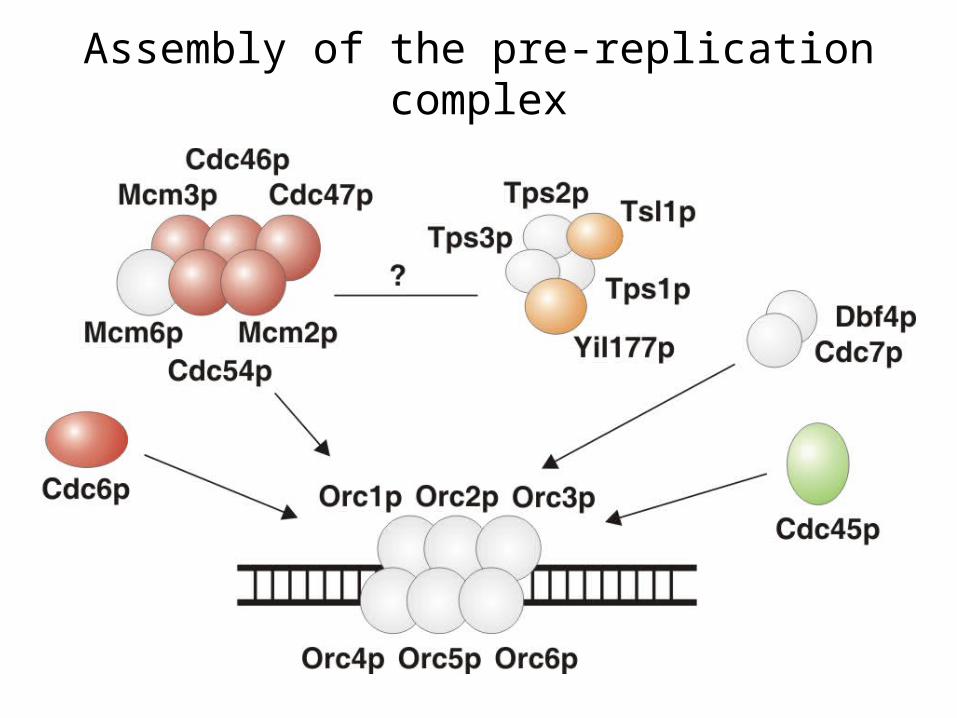
Assembly of the pre-replication complex
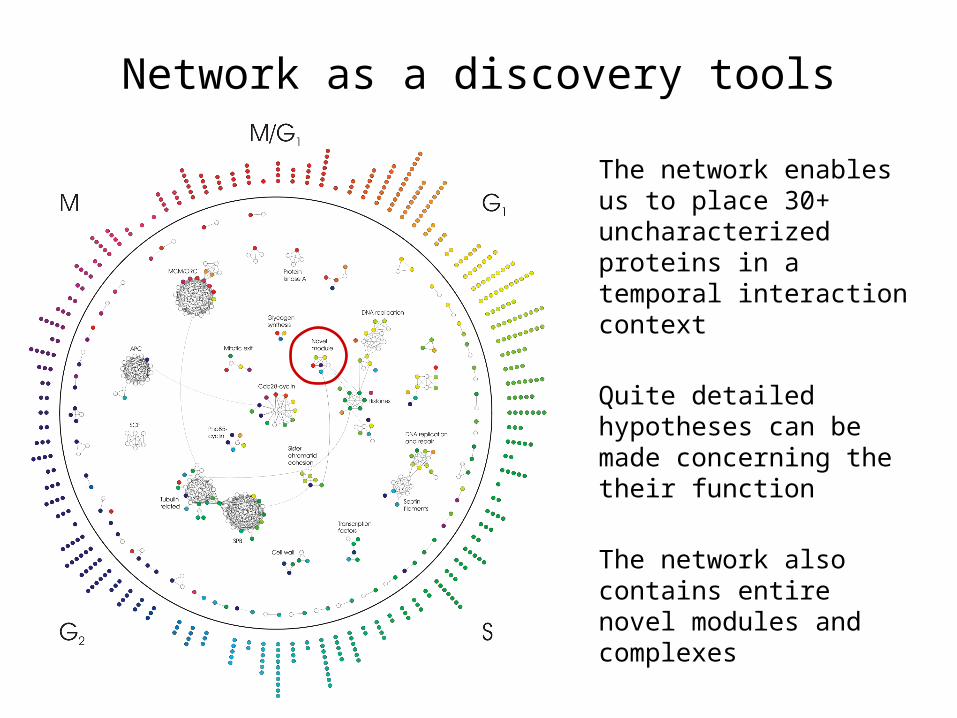
Network as a discovery tools
The network enables us to place 30+ uncharacterized proteins in a temporal interaction context
Quite detailed hypotheses can be made concerning the their function
The network also contains entire novel modules and complexes
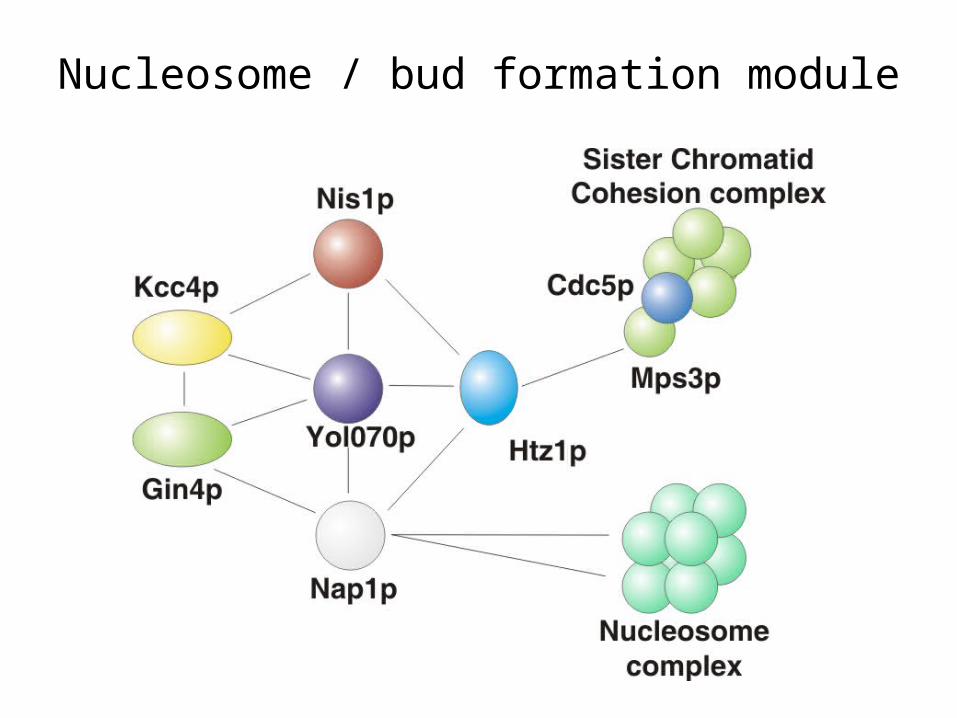
Nucleosome / bud formation module

Rediscovering the “party” hubs and “date” hubs
“Date” hubs: the hub protein interacts with different proteins at different times.
“Party” hubs: the hub protein and its interactors are expressed close in time.
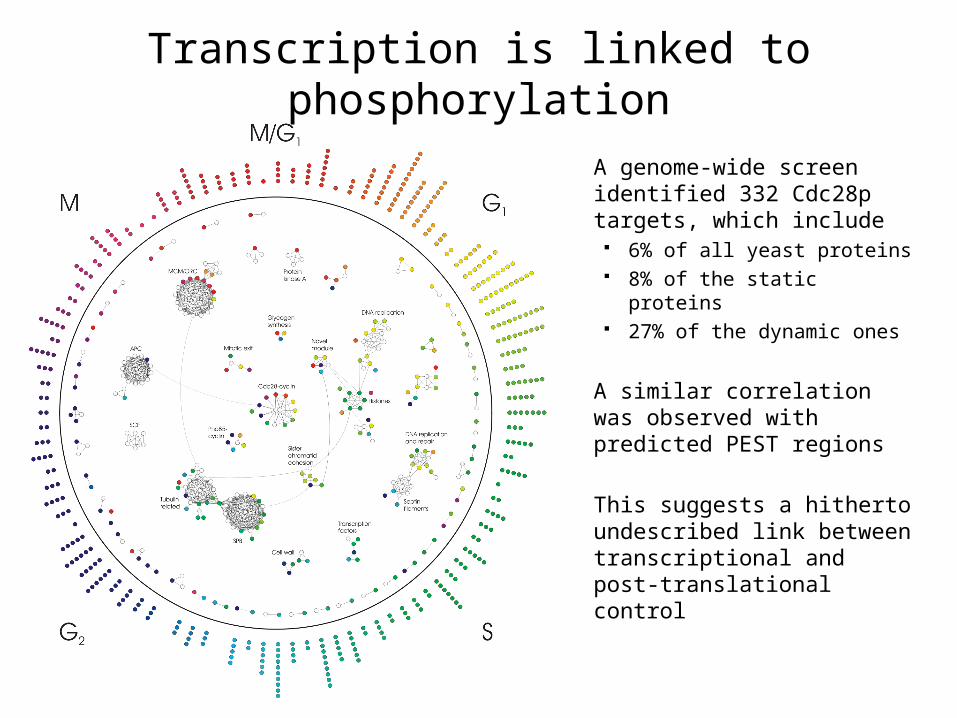
Transcription is linked to phosphorylation
A genome-wide screen identified 332 Cdc28p targets, which include 6% of all yeast proteins 8% of the static proteins 27% of the dynamic ones
A similar correlation was observed with predicted PEST regions
This suggests a hitherto undescribed link between transcriptional and post-translational control

Conclusions
• It is possible to construct highly reliable models from microarray data and high-throughput interaction screen
• Temporal interaction networks can provide an overview of how and when protein complexes are assembled
• Different mechanism for regulating protein activity appear to be tightly linked to each other

Acknowledgments
• The yeast cell cycle interaction network Ulrik de Lichtenberg Søren Brunak Peer Bork
• Re-analysis of cell cycle microarray expression data Thomas Skøt Jensen Anders Fausbøll
• Also thanks to Sean Hooper Christian von Mering

Thank you!





























