Embed Size (px)
Citation preview

1 23
Applied Microbiology andBiotechnology ISSN 0175-7598 Appl Microbiol BiotechnolDOI 10.1007/s00253-012-3907-5
Phanerochaete chrysosporium produces adiverse array of extracellular enzymes whengrown on sorghum
Anamika Ray, Sayali Saykhedkar,Patricia Ayoubi-Canaan, StevenD. Hartson, Rolf Prade & AndrewJ. Mort

1 23
Your article is protected by copyright and
all rights are held exclusively by Springer-
Verlag. This e-offprint is for personal use only
and shall not be self-archived in electronic
repositories. If you wish to self-archive your
work, please use the accepted author’s
version for posting to your own website or
your institution’s repository. You may further
deposit the accepted author’s version on a
funder’s repository at a funder’s request,
provided it is not made publicly available until
12 months after publication.

GENOMICS, TRANSCRIPTOMICS, PROTEOMICS
Phanerochaete chrysosporium produces a diverse arrayof extracellular enzymes when grown on sorghum
Anamika Ray & Sayali Saykhedkar &
Patricia Ayoubi-Canaan & Steven D. Hartson &
Rolf Prade & Andrew J. Mort
Received: 25 October 2011 /Revised: 11 January 2012 /Accepted: 14 January 2012# Springer-Verlag 2012
Abstract In an effort to understand how fungi degrade bio-mass, we grew Phanerochaete chrysosporium on sorghumstover and chronicled the growth of the fungus over the courseof 14 days. The fungal mass grew steadily until the fifth day,reaching 0.06 mg of cells per milligram of dry mass, which fellby the seventh day and stayed at nearly the same level untilday 14. After 1 day, hemicellulases, cellulases, and polygalac-turonases were detected in the extracellular fluid at 1.06, 0.34,and 0.20 U/ml, respectively. Proteomic studies performed withthe extracellular fluid using liquid chromatography–tandemmass spectrometry identified 57, 116, and 102 degradativeenzymes targeting cellulose, hemicellulose, pectin, lignin, pro-teins, and lipids on days 1, 7, and 14, respectively. Significantconcentrations of breakdown products of the sorghum poly-saccharides were detected in the extracellular fluid indicatingthat the enzymes were breaking the polysaccharides, and after14 days, almost 39% of the sorghum sugars had been used bythe fungus. Our results suggest that P. chrysosporium producesa set of enzymes to degrade the components of lignocellulosefrom the beginning of its growth, but modifies the complementof enzymes it secretes over time to adapt to the particularsubstrate available.
Keywords Phanerochaete chrysosporium . Proteomics .
Sorghum . Lignocellulose
Introduction
Sorghum is an attractive bioenergy crop in the mid-USA forproduction of fermentable sugars because it is a high yield cropwith sparse water requirements and robust growth character-istics (Lacewell et al. 1986; Hallam et al. 2001). Forage sor-ghum is grown onmore than 6million acres of land in the USAeach year producing approximately 58 million tons of biomass(stover) (http://www.afdc.energy.gov/afdc/data/biomass_resources.html, http://www.brdisolutions.com/default.aspx;Corredor et al. 2009). The primary constituent of sorghum,the lignocellulose, is comprised of cellulose, hemicellulose,and lignin, which are interlinked by a combination ofcovalent bonds and noncovalent linkages (Pérez et al.2002; Sanchez 2009). The natural resistance of sorghumstover to enzymatic attack is attributed to the complexityof the lignin structure and the intricate architecture of thelignocellulose.
Many efforts have been made to develop efficient processesfor breakdown and conversion of lignocellulosic biomass tofermentable sugars. These processes include a pretreatment todisrupt the lignin encrusting the polysaccharides and to reducethe crystallinity of the cellulose to make it more accessible tocellulases. The prevalent pretreatments include acid hydrolysis(Saha et al. 2005), alkali extraction (Gassan andBledzki 1999),steam explosion (Brownell et al. 2004), carbon dioxide explo-sion (Zheng et al. 1998), ammonia fiber explosion (Teymouriet al. 2005), ozonolysis (Vidal and Molinier 1988), ionizingradiation, microwave, and hydrothermolysis (Yu et al. 2009).These pretreatment methods suffer from several drawbacks,which include lack of selectivity, need for corrosion-resistant
Electronic supplementary material The online version of this article(doi:10.1007/s00253-012-3907-5) contains supplementary material,which is available to authorized users.
A. Ray (*) : S. Saykhedkar : P. Ayoubi-Canaan : S. D. Hartson :A. J. MortDepartment of Biochemistry and Molecular Biology,Oklahoma State University,Stillwater, OK 74078, USAe-mail: [email protected]
R. PradeDepartment of Microbiology and Molecular Genetics,Oklahoma State University,Stillwater, OK 74078, USA
Appl Microbiol BiotechnolDOI 10.1007/s00253-012-3907-5
Author's personal copy

high-pressure reactors, accumulation of compounds inhibitoryto subsequent fermentation, and the disposal of harmful chem-icals in an environmental-friendly way (Shi et al. 2008; Zhenget al. 2011).
Fungi are the only microbes that can turn over biomassalmost completely under mild conditions without any harshchemical or physical treatment (Crawford and Crawford1980). The white-rot fungus, Phanerochaete chrysosporium,is a widely studied basidiomycete, whose well-annotated ge-nome (Martinez et al. 2004) provides insight into the genesand proteins that play an important role in lignocellulosedegradation. This fungus employs an extensive collection ofextracellular hydrolytic and oxidative enzymes (Kirk andFarrell 1987; Kersten and Cullen 2007). Hydrolytic enzymesystems include cellulases and hemicellulases to degrade pol-ysaccharides, and oxidative enzymes include lignin peroxi-dases (EC 1.11.1.14) (LiP) and manganese peroxidases (EC1.11.1.13) (MnP) to depolymerize lignin and open phenylrings (Cullen and Kersten 1996; Cullen 1997).
The efficiency of biodegradation of lignocelluloses is de-pendent on environmental conditions and cultivation methods(Reddy and D’Souza 1994). Solid-state cultivation is a methodwhere the environmental conditions are akin to the naturalhabitat of this fungus (Datta et al. 1991; Shi et al. 2008). Somestudies have been reported on the growth of P. chrysosporiumon wood chips as submerged cultures (Abbas et al. 2005; Satoet al. 2007), on cellulose and on defined media (VandenWymelenberg et al. 2005, 2006) with subsequent analysis ofextracellular enzymes. No studies have been reported to dateon the growth of P. chrysosporium on sorghum for studyingthe enzyme system utilized to deconstruct lignocelluloses.
The objective of this study was to identify, in a temporalfashion, the complete set of enzymes that P. chrysosporiumproduces for growth on sorghum stover under solid-statecultivation conditions, very close to the natural conditions.Results illuminate a working enzymatic array, whose designand function should facilitate the development of an efficientin vitro saccharification system.
Materials and methods
Strains and growth conditions
P. chrysosporium strain RP78 (FGSC strain 9002) wasobtained from the Fungal Genetics Stock Center (http://www.fgsc.net/) and used throughout the study. Sorghumstover (variety mix of AtX2752/RtX2783 and AtX2752/RtX430) was ground in a Thomas Wiley® Mini-Mill(Thomas Scientific, Swedesboro, NJ, USA) by passingthrough a 60-mesh screen. Three grams of powdered sor-ghum was moistened with 6 ml of water in a Petri plate,mixed well, and autoclaved for an hour at 121 °C. Fungal
spores were counted using a hemacytometer (SpectrumScientifics, Philadelphia, PA, USA) and adjusted to amillion spores per milliliter in water. Fifteen millilitersof minimal media (5 ml of 20× nitrate salts, 0.1 ml of1,000× trace elements, pH 6.5 adjusted to a final volumeof 100 ml with water) (Pontecorvo 1969) and 1 ml ofspore suspension (∼106 spores) were added to each Petriplate, which was then incubated at 37 °C, under 70%relative humidity for 1, 2, 3, 5, 7, and 14 days. Sorghumplates with everything but fungal inoculation, incubatedalong with inoculated plates, served as controls. All thechemicals used in this study were purchased from Sigma-Aldrich (St. Louis, MO, USA), unless otherwise stated.
Microscopy and sample preparations
For scanning electron microscopy (SEM), fungal mycelialmats grown under solid-state cultivation conditions were cutinto 5-mm squares and fixed in 2.5% glutaraldehyde in 0.2 Msodium phosphate, pH 7.2 for 2 h. The glutaraldehyde solu-tion was removed and replaced with 0.2 M sodium phosphate,pH 7.2, and the samples were washed thrice for 20 min. Thesamples were postfixed in 1% osmium tetraoxide in H2O for1 h. The samples were then dehydrated in a series of ethanolsolutions (50%, 70%, 90%, 95%, 100%, 100%, and 100%) for15 min each, critical point dried, and mounted on aluminumstubs using silver paint adhesive. They were then coated withgold/palladium and observed using an FEI quanta 600 elec-tron microscope (FEI, Hillsboro, OR, USA).
For transmission electron microscopy (TEM), the fixationwas done as for the SEM samples. Postfixation was done with1% osmium tetraoxide/1.5% potassium ferricyanide in 0.1%sodium cacodylate buffer for 1 h. The dehydration steps werethe same as those conducted for SEM samples. The sampleswere further washed twice with 100% propylene oxide andincubated overnight in propylene oxide/polyresin/bed 812(Polysciences Inc., Warrington, PA, USA) followed by em-bedding in polyresin/bed 812 and polymerization at 60 °C.Sectioning was done using a Leica EMCU 6 ultramicrotome(Midwest Lab Equipment, Iowa City, IA, USA), and sec-tioned samples were placed on carbon/formavar-coated nickelgrids, stained with uranyl acetate and lead citrate, and ob-served using a JEOL 2100 transmission electron microscope(JEOL, Austin, TX, USA).
Chitin estimation
The total biomass from 1, 2, 3, 5, 7, and 14 days of growthwas freeze dried, powdered, mixed thoroughly, and then5 mg samples were transferred to glass vials. The sampleswere vortexed in 3 ml of 6 N HCl and heated at 100 °C for6 h. The HCl was evaporated using a speed-vac overnight.Ten microliters of 1 M ammonium hydroxide was added to
Appl Microbiol Biotechnol
Author's personal copy

neutralize any remaining acid followed by 100 μl of 20 mg/mlof potassium borohydride in dimethyl sulfoxide (DMSO) andincubated for 90 min at 40 °C (Penman et al. 2000) to reducethe sugars to alditols. The reaction was stopped by adding10 μl of glacial acetic acid. Twenty microliters of methylimidazole and 200 μl of acetic anhydride were incubated withthe samples for 10 min at room temperature to acetylate thealditols (Mort et al. 1983). Five hundred microliters of waterwas added to the final reaction mix, and the acetylated alditolswere adsorbed to a C18 Sep-Pac column (Waters Co., Milford,MA, USA). The reagents were washed from the column withwater and the acetylated alditols eluted in dichloromethane,while the polyphenols remained adsorbed to the Sep-Pak. Thedichloromethane was dried over sodium sulfate and then evap-orated to dryness. Twenty-five microliters of ethyl acetate wasadded to the dried sample, out of which, 1 μl was injected intothe gas chromatograph (Agilent, Santa Clara, CA, USA) forestimation of chitin. One hundred nanomoles of methyl gluc-amine was added to each sample as an internal standard. Astandard curve was prepared using known amounts of glucos-amine ranging from 10 to 250 nmol. The glucosamine contentof 1 mg aliquots of dried P. chrysosporium grown in liquidmedia (191.25 nmol) was used as a conversion factor toestimate milligrams of fungus per milligram of dry mass.
Liquid chromatography–tandem mass spectrometry
The fungal cultures grown on sorghum for the aforementioneddays were washed with 5 ml autoclaved water and the filtratescollected and frozen in small aliquots. These aliquots repre-senting the extracellular extracts were used for proteomicsstudies and enzyme assays. The extracellular extracts (days 1,7, and 14) were dried and then dissolved in 8 M urea, 100 mMTris-HCl, 5 mM Tris(2-carboxy ethyl)phosphine (TCEP-HCl),pH 8.5, and incubated at room temperature for 20 min. Sub-sequently, samples were alkylated using 10 mM idoacetamidefor 15 min in the dark at room temperature. The samples werediluted with four volumes of 100 mM Tris, pH 8.5, anddigested with 4 μg/ml trypsin at 37 °C overnight. Digestedsamples were adjusted to 1% formic acid and analyzed byliquid chromatography–tandem mass spectrometry (LC–MS/MS) using an LTQ-Orbitrap XL hybrid mass spectrometer(Thermo Fisher Scientific, San Jose, CA, USA). An Eksigentspilt less LC pump (Eksigent, Dublin, CA, USA) was used toseparate peptide populations on analytical C18 nano columns(Bruker-Michrom Inc., Auburn, CA, USA), with the columneffluent sprayed directly into a New Objective pico view ionsource. The MS/MS method consisted of the orbitrap massspectrometer collecting a high-resolution scan of intact pepti-des, while the LTQ ion trap simultaneously performedMS/MSfragmentation analysis of each of the three most abundantpeptides. The LC–MS/MS raw files were submitted for localdatabase searching using Mascot (v2.2.2 fromMatrix Science,
Boston, MA, USA) against sequences from P. chrysosporium(http://pedant.gsf.de/), as well as against a sorghum database(http://www.phytozome.net/sorghum). Detailed informationfor each protein along with its accession numbers wereobtained from the Pedant database (http://pedant.gsf.de/). MSmass accuracies of 10 ppm and stated MS/MSmass accuraciesof 0.80 Da were used. Searches were validated using Scaf-fold_2_06_02 (Proteome Software Inc., Portland, OR, USA)and the Peptide Prophet algorithm. A stringent criterion forprotein identification was used, where only protein probabilitythresholds greater than 99% were accepted and at least twopeptides needed to be identified, each with 95% certainty.Protein candidates containing similar peptides were groupedto fulfill the principles of parsimony. Search results were alsochecked for false discoveries using reversed decoy sequencedatabases, and no decoy sequences were detected, thus bring-ing our false discovery rate to zero. Changes in protein expres-sion between samples from different time points wereexamined using the spectral count method (Old et al. 2005).SignalP was used to predict secretion signals in the identifiedproteins (http://www.cbs.dtu.dk/services/SignalP/) (Bendtsenet al. 2004). Additional information for the presence of a signalpeptide was obtained by accessing the following URL with themodel number, e.g., http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db0Phchr1&id03651. Domain searching was doneusing the SMART program (http://smart.embl-heidelberg.de/smart/set_mode.cgi?NORMAL01) (Schultz et al. 2000).
Capillary zone electrophoresis (CZE) and dinitrosalicylicacid (DNS) assay
Enzyme activities were measured semi-quantitatively in theextracellular extracts by capillary zone electrophoresis (CZE)(Naran et al. 2007). Two microliters of 8-aminopyrene-1,3,6-trisulfonate (APTS)-labeled substrates (xylohexaose, cello-pentaose, arabinoheptaose, and mannohexaose), 1 μl of extra-cellular extract, along with 22 μl of 50 mM ammoniumacetate buffer of optimum pH for respective enzymes wasincubated for 1 h at 37 °C. The enzymes were heat inactivatedat 100 °C for 5 min followed by analysis of the fluorescentproducts using CZE (Bio-Rad, Hercules, CA, USA). Thedinitrosalicylic acid (DNS) assay, developed by Sumner andGraham (1921) for determination of reducing sugar, was usedfor quantitative determination of enzyme activity. The DNSreagent consisted of 0.75% DNS, 0.5% phenol, 0.5% sodiummetabisulfite, 1.4% sodium hydroxide, 21% sodium potassi-um tartrate (Miller 1959; Squina et al. 2009). For the assay, weused 10 μl of extracellular extract, 20 μl of 50 mM ammoni-um acetate buffer of optimum pH, and 20 μl of appropriatesubstrate (1% larchwood xylan for xylanase, 1% carboxy-methyl cellulose for cellulase, 1% pectic acid for pectinase,and 0.5% locust bean gum for mannanase). The reactionswere incubated for 30min at 37 °C and terminated by addition
Appl Microbiol Biotechnol
Author's personal copy

of 40 μl of DNS reagent followed by heating at 100 °C for5 min. The assays were conducted in 96-well flat bottompolystyrene plates (Corning, NY, USA), and absorbance read-ings at 550 nm were obtained using the Infinite® 200 seriesplate reader (Tecan, San Jose, CA, USA).
Estimation of oligosaccharides in extracellular extractsby gas liquid chromatography (GLC)
Twenty-five microliters of extracellular extracts were mixedwith 100 nmol of inositol (internal standard) and dried. Twohundred microliters of 1.5 M HCl in methanol was added toeach sample followed by 100 μl of methyl acetate and heatedat 80 °C for a minimum of 3 h, which converted the sugars tomethyl glycosides (Komalavilas and Mort 1989). The vialswere cooled, followed by addition of a few drops of t-butanolfollowed by the evaporation of the solvents under a stream ofnitrogen at room temperature. Twenty-five microliters of a1:1:5 mixture of hexamethyldisilazane:trimethylchlrosilane:pyridine was added, and the samples were incubated for atleast 15 min. The samples were evaporated under nitrogen gasand dissolved in 50 μl isooctane, out of which, 1 μl wasinjected in the gas chromatograph (Agilent, Santa Clara, CA,USA). The amount of each sugar in the sample was calculatedby using the formula: area of sugar peak in sample/area ofinositol peak in the sample/area of sugar peak in the standards/area of inositol in standards×1000number of nanomoles inthe sample.
Estimation of sugars in the residual sorghum by Saemanhydrolysis
Saeman hydrolysis was done on the total biomass on theplates after 1, 7, and 14 days of fungal growth to estimatetotal sugars in a single procedure (Saeman et al. 1954).Three hundred micrograms of dried biomass was mixedwith 3 ml of 72% sulfuric acid and incubated at 30 °C for60 min. The samples were diluted to 4% sulfuric acid withwater and autoclaved for 60 min at 121 °C. After neutrali-zation to pH 5–6 with calcium carbonate, 25 μl of samplewas processed for methanolysis, trimethylsilylation, and gasliquid chromatography (GLC) analysis as described above.
Results
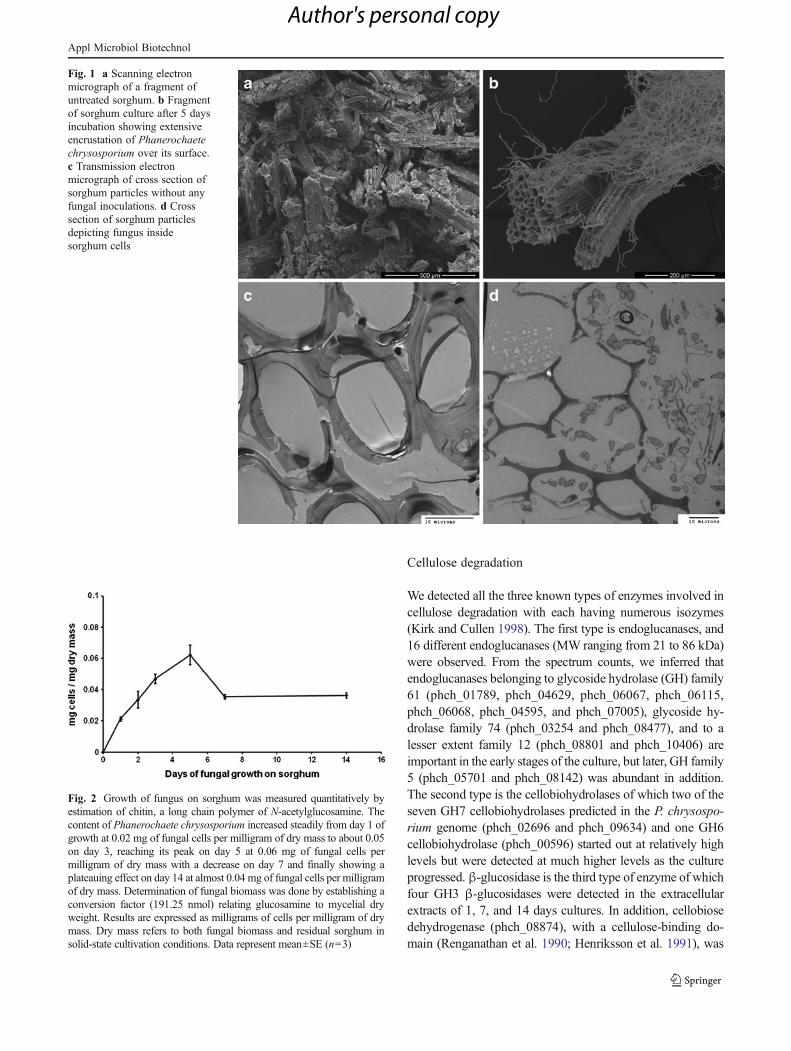
The growth of fungus on sorghum was monitored usingscanning and transmission electron microscopy (SEM andTEM). Figure 1a shows a SEM picture of the sorghumparticles without any fungus, while Fig. 1b shows thegrowth of fungus on sorghum on the fifth day after inocu-lation. Sorghum particles are shown with fungal myceliaintertwined around them. Figure 1c shows a transmission
electron microscope picture of sorghum particles withoutfungus, while Fig. 1d depicts the TEM picture of sorghumwith fungus growing inside the sorghum cells.
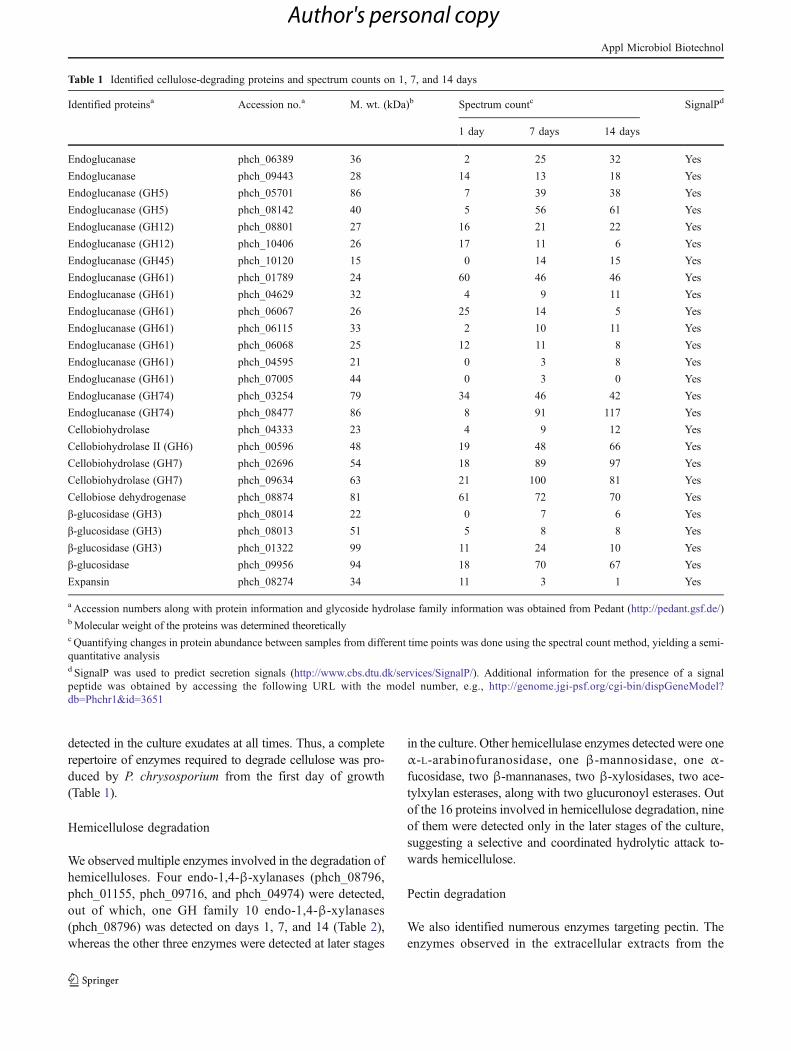
In order to quantitate the growth of fungus on sor-ghum, we estimated the content of chitin by GLC. Chitinis a crystalline polymer of N-acetyl-β-D-glucosamine, akey component of fungal cell walls which is absent insorghum and thus serves as a measure of fungal growthin mixed fungal/plant culture systems. The growth of P.chrysosporium expressed as the dry weight of fungalcells per milligram of dry biomass showed a distinctivepattern with a steady increase from the first day ofgrowth at 0.02 mg of fungal cells per milligram of drymass to almost 0.05 on day 3 and reaching its peak onday 5 at 0.06 mg of fungal cells per milligram of drymass. Fungal dry mass decreased by day 7 to 0.04 mg offungal cells per milligram of dry mass and remainedconstant until day 14 (Fig. 2). No chitin was detectedin the control samples, i.e., sorghum without any fungalinoculation.
Keeping in mind the diversity of enzymes required forcomplete degradation of sorghum, combined with previousknowledge of the recalcitrance of plant cell walls to enzy-matic breakdown, we decided to study the secretome of P.chrysosporium grown on sorghum for 1, 7, and 14 daysusing LC–MS/MS. The concentrations of total protein in the15-ml extracellular fluid rinsed from the sorghum slurry ondays 1, 7, and 14 were 0.220, 0.787, and 0.938 mg/ml,respectively. When the peptides generated from these frac-tions were analyzed by LC–MS/MS and identified by data-base searching, we found evidence for the presence ofenzymes from each of the major degradative families in-cluding cellulases, hemicellulases, pectinases, peroxidases,proteases, and lipases. We also found proteins involved infungal growth and turnover along with other miscellaneousproteins including hypothetical proteins (Tables 1, 2, 3, 4, 5,and 6). Proteomic studies identified 57, 116, and 102 pro-teins on days 1, 7, and 14, respectively. Fifty-one proteinswere found to be present at all three time points, and another51 were found on days 7 and 14, but not on day 1. Fourproteins were found on day 1 only, 12 on day 7 only, andtwo on days 1 and 7 only. Almost all of the enzymesdirected toward cellulose degradation were present on alldays. On the other hand, a little less than half of the hemi-cellulases were found from the first day on with the restbeing produced later. Most of the pectin-degrading enzymeswere found only after the first day, as were the proteases andhypothetical proteins. Detailed information for all proteinsand the complete dataset are presented in the SupplementaryMaterial in Table S1. To analyze changes in the secretion ofindividual enzymes as the cultures proceeded, we comparedtheir spectrum counts from the LC–MS/MS analysis of eachtime point.
Appl Microbiol Biotechnol
Author's personal copy
Cellulose degradation
We detected all the three known types of enzymes involved incellulose degradation with each having numerous isozymes(Kirk and Cullen 1998). The first type is endoglucanases, and16 different endoglucanases (MW ranging from 21 to 86 kDa)were observed. From the spectrum counts, we inferred thatendoglucanases belonging to glycoside hydrolase (GH) family61 (phch_01789, phch_04629, phch_06067, phch_06115,phch_06068, phch_04595, and phch_07005), glycoside hy-drolase family 74 (phch_03254 and phch_08477), and to alesser extent family 12 (phch_08801 and phch_10406) areimportant in the early stages of the culture, but later, GH family5 (phch_05701 and phch_08142) was abundant in addition.The second type is the cellobiohydrolases of which two of theseven GH7 cellobiohydrolases predicted in the P. chrysospo-rium genome (phch_02696 and phch_09634) and one GH6cellobiohydrolase (phch_00596) started out at relatively highlevels but were detected at much higher levels as the cultureprogressed. β-glucosidase is the third type of enzyme of whichfour GH3 β-glucosidases were detected in the extracellularextracts of 1, 7, and 14 days cultures. In addition, cellobiosedehydrogenase (phch_08874), with a cellulose-binding do-main (Renganathan et al. 1990; Henriksson et al. 1991), was
Fig. 2 Growth of fungus on sorghum was measured quantitatively byestimation of chitin, a long chain polymer of N-acetylglucosamine. Thecontent of Phanerochaete chrysosporium increased steadily from day 1 ofgrowth at 0.02 mg of fungal cells per milligram of dry mass to about 0.05on day 3, reaching its peak on day 5 at 0.06 mg of fungal cells permilligram of dry mass with a decrease on day 7 and finally showing aplateauing effect on day 14 at almost 0.04 mg of fungal cells per milligramof dry mass. Determination of fungal biomass was done by establishing aconversion factor (191.25 nmol) relating glucosamine to mycelial dryweight. Results are expressed as milligrams of cells per milligram of drymass. Dry mass refers to both fungal biomass and residual sorghum insolid-state cultivation conditions. Data represent mean±SE (n03)
Fig. 1 a Scanning electronmicrograph of a fragment ofuntreated sorghum. b Fragmentof sorghum culture after 5 daysincubation showing extensiveencrustation of Phanerochaetechrysosporium over its surface.c Transmission electronmicrograph of cross section ofsorghum particles without anyfungal inoculations. d Crosssection of sorghum particlesdepicting fungus insidesorghum cells
Appl Microbiol Biotechnol
Author's personal copy
detected in the culture exudates at all times. Thus, a completerepertoire of enzymes required to degrade cellulose was pro-duced by P. chrysosporium from the first day of growth(Table 1).
Hemicellulose degradation
We observed multiple enzymes involved in the degradation ofhemicelluloses. Four endo-1,4-β-xylanases (phch_08796,phch_01155, phch_09716, and phch_04974) were detected,out of which, one GH family 10 endo-1,4-β-xylanases(phch_08796) was detected on days 1, 7, and 14 (Table 2),whereas the other three enzymes were detected at later stages
in the culture. Other hemicellulase enzymes detected were oneα-L-arabinofuranosidase, one β-mannosidase, one α-fucosidase, two β-mannanases, two β-xylosidases, two ace-tylxylan esterases, along with two glucuronoyl esterases. Outof the 16 proteins involved in hemicellulose degradation, nineof them were detected only in the later stages of the culture,suggesting a selective and coordinated hydrolytic attack to-wards hemicellulose.
Pectin degradation
We also identified numerous enzymes targeting pectin. Theenzymes observed in the extracellular extracts from the
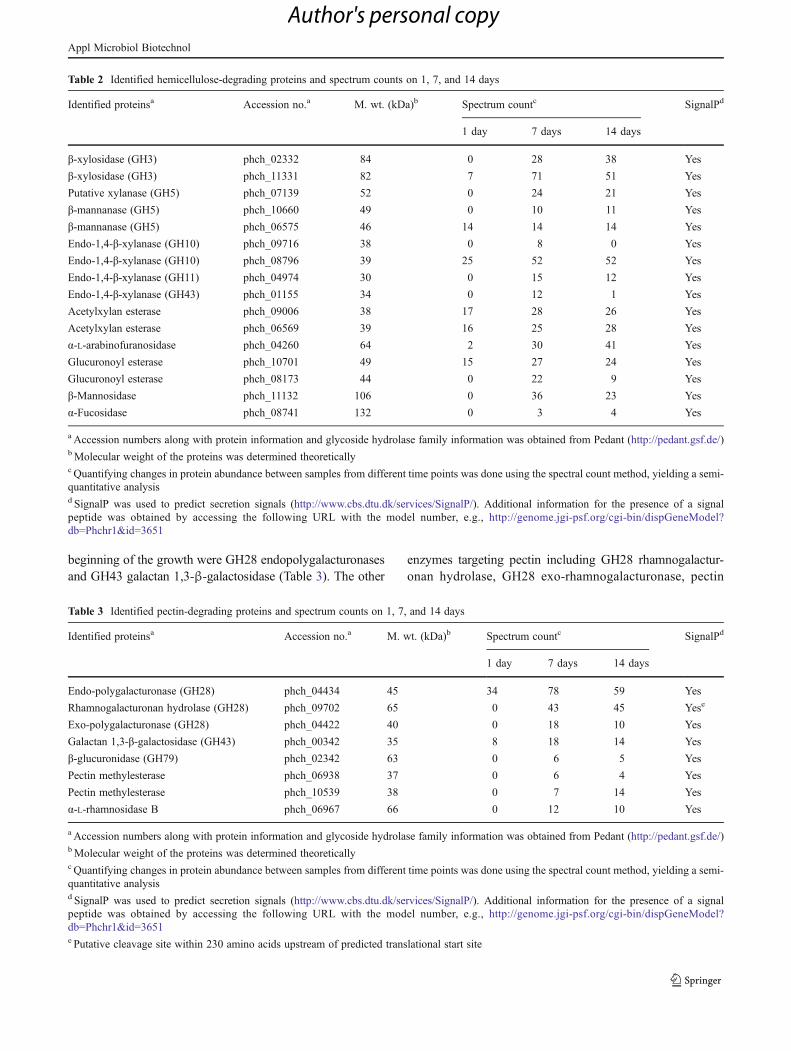
Table 1 Identified cellulose-degrading proteins and spectrum counts on 1, 7, and 14 days
Identified proteinsa Accession no.a M. wt. (kDa)b Spectrum countc SignalPd
1 day 7 days 14 days
Endoglucanase phch_06389 36 2 25 32 Yes
Endoglucanase phch_09443 28 14 13 18 Yes
Endoglucanase (GH5) phch_05701 86 7 39 38 Yes
Endoglucanase (GH5) phch_08142 40 5 56 61 Yes
Endoglucanase (GH12) phch_08801 27 16 21 22 Yes
Endoglucanase (GH12) phch_10406 26 17 11 6 Yes
Endoglucanase (GH45) phch_10120 15 0 14 15 Yes
Endoglucanase (GH61) phch_01789 24 60 46 46 Yes
Endoglucanase (GH61) phch_04629 32 4 9 11 Yes
Endoglucanase (GH61) phch_06067 26 25 14 5 Yes
Endoglucanase (GH61) phch_06115 33 2 10 11 Yes
Endoglucanase (GH61) phch_06068 25 12 11 8 Yes
Endoglucanase (GH61) phch_04595 21 0 3 8 Yes
Endoglucanase (GH61) phch_07005 44 0 3 0 Yes
Endoglucanase (GH74) phch_03254 79 34 46 42 Yes
Endoglucanase (GH74) phch_08477 86 8 91 117 Yes
Cellobiohydrolase phch_04333 23 4 9 12 Yes
Cellobiohydrolase II (GH6) phch_00596 48 19 48 66 Yes
Cellobiohydrolase (GH7) phch_02696 54 18 89 97 Yes
Cellobiohydrolase (GH7) phch_09634 63 21 100 81 Yes
Cellobiose dehydrogenase phch_08874 81 61 72 70 Yes
β-glucosidase (GH3) phch_08014 22 0 7 6 Yes
β-glucosidase (GH3) phch_08013 51 5 8 8 Yes
β-glucosidase (GH3) phch_01322 99 11 24 10 Yes
β-glucosidase phch_09956 94 18 70 67 Yes
Expansin phch_08274 34 11 3 1 Yes
a Accession numbers along with protein information and glycoside hydrolase family information was obtained from Pedant (http://pedant.gsf.de/)bMolecular weight of the proteins was determined theoreticallyc Quantifying changes in protein abundance between samples from different time points was done using the spectral count method, yielding a semi-quantitative analysisd SignalP was used to predict secretion signals (http://www.cbs.dtu.dk/services/SignalP/). Additional information for the presence of a signalpeptide was obtained by accessing the following URL with the model number, e.g., http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db0Phchr1&id03651
Appl Microbiol Biotechnol
Author's personal copy
beginning of the growth were GH28 endopolygalacturonasesand GH43 galactan 1,3-β-galactosidase (Table 3). The other
enzymes targeting pectin including GH28 rhamnogalactur-onan hydrolase, GH28 exo-rhamnogalacturonase, pectin
Table 2 Identified hemicellulose-degrading proteins and spectrum counts on 1, 7, and 14 days
Identified proteinsa Accession no.a M. wt. (kDa)b Spectrum countc SignalPd
1 day 7 days 14 days
β-xylosidase (GH3) phch_02332 84 0 28 38 Yes
β-xylosidase (GH3) phch_11331 82 7 71 51 Yes
Putative xylanase (GH5) phch_07139 52 0 24 21 Yes
β-mannanase (GH5) phch_10660 49 0 10 11 Yes
β-mannanase (GH5) phch_06575 46 14 14 14 Yes
Endo-1,4-β-xylanase (GH10) phch_09716 38 0 8 0 Yes
Endo-1,4-β-xylanase (GH10) phch_08796 39 25 52 52 Yes
Endo-1,4-β-xylanase (GH11) phch_04974 30 0 15 12 Yes
Endo-1,4-β-xylanase (GH43) phch_01155 34 0 12 1 Yes
Acetylxylan esterase phch_09006 38 17 28 26 Yes
Acetylxylan esterase phch_06569 39 16 25 28 Yes
α-L-arabinofuranosidase phch_04260 64 2 30 41 Yes
Glucuronoyl esterase phch_10701 49 15 27 24 Yes
Glucuronoyl esterase phch_08173 44 0 22 9 Yes
β-Mannosidase phch_11132 106 0 36 23 Yes
α-Fucosidase phch_08741 132 0 3 4 Yes
a Accession numbers along with protein information and glycoside hydrolase family information was obtained from Pedant (http://pedant.gsf.de/)bMolecular weight of the proteins was determined theoreticallyc Quantifying changes in protein abundance between samples from different time points was done using the spectral count method, yielding a semi-quantitative analysisd SignalP was used to predict secretion signals (http://www.cbs.dtu.dk/services/SignalP/). Additional information for the presence of a signalpeptide was obtained by accessing the following URL with the model number, e.g., http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db0Phchr1&id03651
Table 3 Identified pectin-degrading proteins and spectrum counts on 1, 7, and 14 days
Identified proteinsa Accession no.a M. wt. (kDa)b Spectrum countc SignalPd
1 day 7 days 14 days
Endo-polygalacturonase (GH28) phch_04434 45 34 78 59 Yes
Rhamnogalacturonan hydrolase (GH28) phch_09702 65 0 43 45 Yese
Exo-polygalacturonase (GH28) phch_04422 40 0 18 10 Yes
Galactan 1,3-β-galactosidase (GH43) phch_00342 35 8 18 14 Yes
β-glucuronidase (GH79) phch_02342 63 0 6 5 Yes
Pectin methylesterase phch_06938 37 0 6 4 Yes
Pectin methylesterase phch_10539 38 0 7 14 Yes
α-L-rhamnosidase B phch_06967 66 0 12 10 Yes
a Accession numbers along with protein information and glycoside hydrolase family information was obtained from Pedant (http://pedant.gsf.de/)bMolecular weight of the proteins was determined theoreticallyc Quantifying changes in protein abundance between samples from different time points was done using the spectral count method, yielding a semi-quantitative analysisd SignalP was used to predict secretion signals (http://www.cbs.dtu.dk/services/SignalP/). Additional information for the presence of a signalpeptide was obtained by accessing the following URL with the model number, e.g., http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db0Phchr1&id03651e Putative cleavage site within 230 amino acids upstream of predicted translational start site
Appl Microbiol Biotechnol
Author's personal copy
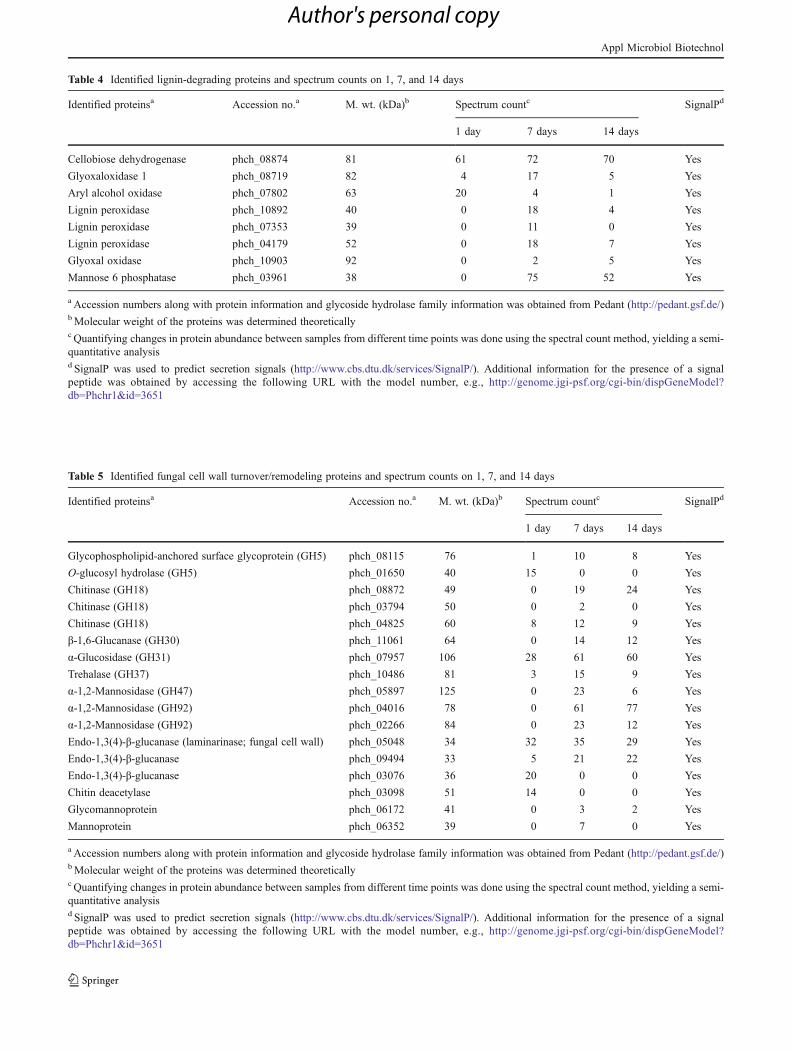
Table 4 Identified lignin-degrading proteins and spectrum counts on 1, 7, and 14 days
Identified proteinsa Accession no.a M. wt. (kDa)b Spectrum countc SignalPd
1 day 7 days 14 days
Cellobiose dehydrogenase phch_08874 81 61 72 70 Yes
Glyoxaloxidase 1 phch_08719 82 4 17 5 Yes
Aryl alcohol oxidase phch_07802 63 20 4 1 Yes
Lignin peroxidase phch_10892 40 0 18 4 Yes
Lignin peroxidase phch_07353 39 0 11 0 Yes
Lignin peroxidase phch_04179 52 0 18 7 Yes
Glyoxal oxidase phch_10903 92 0 2 5 Yes
Mannose 6 phosphatase phch_03961 38 0 75 52 Yes
a Accession numbers along with protein information and glycoside hydrolase family information was obtained from Pedant (http://pedant.gsf.de/)bMolecular weight of the proteins was determined theoreticallyc Quantifying changes in protein abundance between samples from different time points was done using the spectral count method, yielding a semi-quantitative analysisd SignalP was used to predict secretion signals (http://www.cbs.dtu.dk/services/SignalP/). Additional information for the presence of a signalpeptide was obtained by accessing the following URL with the model number, e.g., http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db0Phchr1&id03651
Table 5 Identified fungal cell wall turnover/remodeling proteins and spectrum counts on 1, 7, and 14 days
Identified proteinsa Accession no.a M. wt. (kDa)b Spectrum countc SignalPd
1 day 7 days 14 days
Glycophospholipid-anchored surface glycoprotein (GH5) phch_08115 76 1 10 8 Yes
O-glucosyl hydrolase (GH5) phch_01650 40 15 0 0 Yes
Chitinase (GH18) phch_08872 49 0 19 24 Yes
Chitinase (GH18) phch_03794 50 0 2 0 Yes
Chitinase (GH18) phch_04825 60 8 12 9 Yes
β-1,6-Glucanase (GH30) phch_11061 64 0 14 12 Yes
α-Glucosidase (GH31) phch_07957 106 28 61 60 Yes
Trehalase (GH37) phch_10486 81 3 15 9 Yes
α-1,2-Mannosidase (GH47) phch_05897 125 0 23 6 Yes
α-1,2-Mannosidase (GH92) phch_04016 78 0 61 77 Yes
α-1,2-Mannosidase (GH92) phch_02266 84 0 23 12 Yes
Endo-1,3(4)-β-glucanase (laminarinase; fungal cell wall) phch_05048 34 32 35 29 Yes
Endo-1,3(4)-β-glucanase phch_09494 33 5 21 22 Yes
Endo-1,3(4)-β-glucanase phch_03076 36 20 0 0 Yes
Chitin deacetylase phch_03098 51 14 0 0 Yes
Glycomannoprotein phch_06172 41 0 3 2 Yes
Mannoprotein phch_06352 39 0 7 0 Yes
a Accession numbers along with protein information and glycoside hydrolase family information was obtained from Pedant (http://pedant.gsf.de/)bMolecular weight of the proteins was determined theoreticallyc Quantifying changes in protein abundance between samples from different time points was done using the spectral count method, yielding a semi-quantitative analysisd SignalP was used to predict secretion signals (http://www.cbs.dtu.dk/services/SignalP/). Additional information for the presence of a signalpeptide was obtained by accessing the following URL with the model number, e.g., http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db0Phchr1&id03651
Appl Microbiol Biotechnol
Author's personal copy
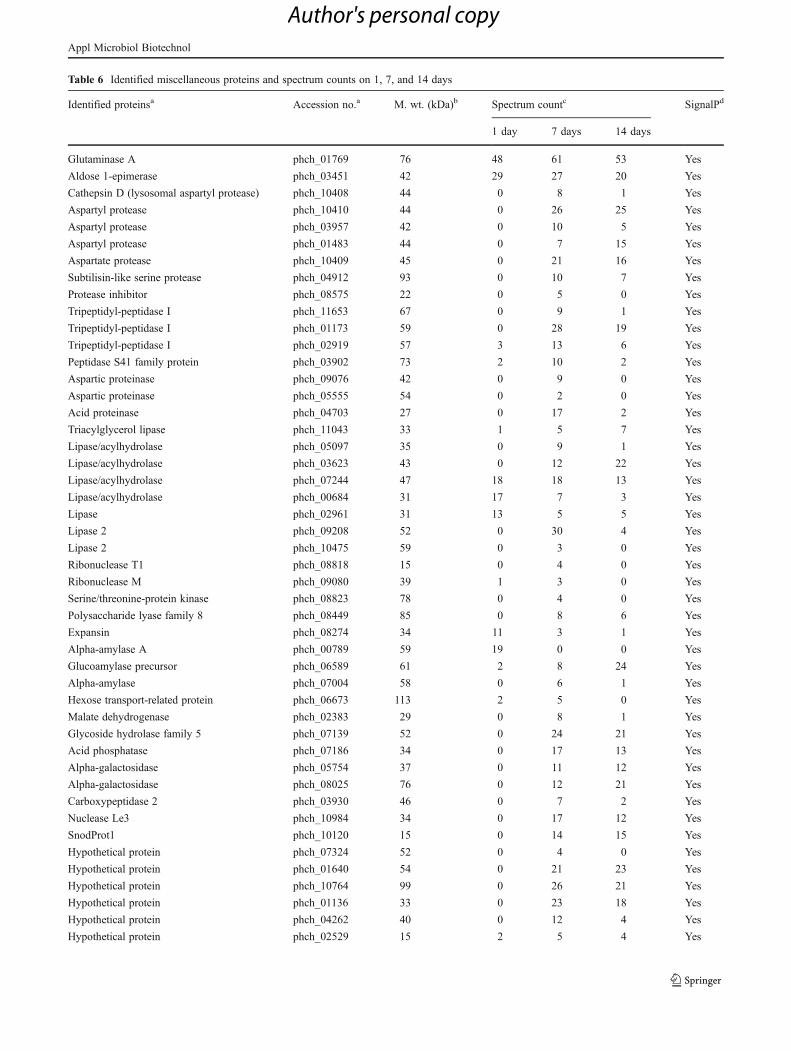
Table 6 Identified miscellaneous proteins and spectrum counts on 1, 7, and 14 days
Identified proteinsa Accession no.a M. wt. (kDa)b Spectrum countc SignalPd
1 day 7 days 14 days
Glutaminase A phch_01769 76 48 61 53 Yes
Aldose 1-epimerase phch_03451 42 29 27 20 Yes
Cathepsin D (lysosomal aspartyl protease) phch_10408 44 0 8 1 Yes
Aspartyl protease phch_10410 44 0 26 25 Yes
Aspartyl protease phch_03957 42 0 10 5 Yes
Aspartyl protease phch_01483 44 0 7 15 Yes
Aspartate protease phch_10409 45 0 21 16 Yes
Subtilisin-like serine protease phch_04912 93 0 10 7 Yes
Protease inhibitor phch_08575 22 0 5 0 Yes
Tripeptidyl-peptidase I phch_11653 67 0 9 1 Yes
Tripeptidyl-peptidase I phch_01173 59 0 28 19 Yes
Tripeptidyl-peptidase I phch_02919 57 3 13 6 Yes
Peptidase S41 family protein phch_03902 73 2 10 2 Yes
Aspartic proteinase phch_09076 42 0 9 0 Yes
Aspartic proteinase phch_05555 54 0 2 0 Yes
Acid proteinase phch_04703 27 0 17 2 Yes
Triacylglycerol lipase phch_11043 33 1 5 7 Yes
Lipase/acylhydrolase phch_05097 35 0 9 1 Yes
Lipase/acylhydrolase phch_03623 43 0 12 22 Yes
Lipase/acylhydrolase phch_07244 47 18 18 13 Yes
Lipase/acylhydrolase phch_00684 31 17 7 3 Yes
Lipase phch_02961 31 13 5 5 Yes
Lipase 2 phch_09208 52 0 30 4 Yes
Lipase 2 phch_10475 59 0 3 0 Yes
Ribonuclease T1 phch_08818 15 0 4 0 Yes
Ribonuclease M phch_09080 39 1 3 0 Yes
Serine/threonine-protein kinase phch_08823 78 0 4 0 Yes
Polysaccharide lyase family 8 phch_08449 85 0 8 6 Yes
Expansin phch_08274 34 11 3 1 Yes
Alpha-amylase A phch_00789 59 19 0 0 Yes
Glucoamylase precursor phch_06589 61 2 8 24 Yes
Alpha-amylase phch_07004 58 0 6 1 Yes
Hexose transport-related protein phch_06673 113 2 5 0 Yes
Malate dehydrogenase phch_02383 29 0 8 1 Yes
Glycoside hydrolase family 5 phch_07139 52 0 24 21 Yes
Acid phosphatase phch_07186 34 0 17 13 Yes
Alpha-galactosidase phch_05754 37 0 11 12 Yes
Alpha-galactosidase phch_08025 76 0 12 21 Yes
Carboxypeptidase 2 phch_03930 46 0 7 2 Yes
Nuclease Le3 phch_10984 34 0 17 12 Yes
SnodProt1 phch_10120 15 0 14 15 Yes
Hypothetical protein phch_07324 52 0 4 0 Yes
Hypothetical protein phch_01640 54 0 21 23 Yes
Hypothetical protein phch_10764 99 0 26 21 Yes
Hypothetical protein phch_01136 33 0 23 18 Yes
Hypothetical protein phch_04262 40 0 12 4 Yes
Hypothetical protein phch_02529 15 2 5 4 Yes
Appl Microbiol Biotechnol
Author's personal copy

methylesterases, α-L-rhamnosidase, and GH79 β-glucuronidase were detected on the seventh and 14th day(Table 3).
Lignin degradation
We identified key enzymes involved in lignin degradation.Three lignin peroxidases (LiPs) were detected, of which,phch_07353 was detected on day 7 only, while the othertwo (phch_10892 and phch_04179) were detected on days 7and 14, whereas manganese peroxidase (MnP) was notdetected in our study. Mannose-6-phosphatase implicatedin post-translation modification of LiPs (Rothschild et al.1999) was detected on days 7 and 14 (Table 4). One glyoxaloxidase (GLX) was observed on day 1 and an additional onedetected on days 7 and 14 along with aryl alcohol oxidase.These oxidases may be involved in the generation of hydro-gen peroxide as substrate for the peroxidases (Kersten 1990;Kersten and Kirk 1987). Cellobiose dehydrogenase(phch_08874), another key enzyme observed in abundancein all the days from the beginning of growth, has beenshown not only to degrade cellulose but also xylan andlignin (Henriksson et al. 1995).
Fungal cell turnover/cell wall remodeling
Various proteins involved in fungal cell wall turn over andcell wall remodeling were identified. Three α-1,2-mannosi-dases were detected in the exudates on days 7 and 14. Twoendo-1,3(4)-β-glucanases (phch_05048 and phch_09494)were detected on days 1, 7, and 14 with an additional one(phch_03076) being detected on day 1 only. Three chiti-nases (phch_08872, phch_03794, and phch_04825) weredetected, of which, phch_04825 was detected on all the3 days studied, and a chitin deacyetylase (phch_03098)was identified on day 1 only. Several other proteins involvedin cell wall remodeling were also identified (Table 5).
Miscellaneous proteins
Several proteins identified in this category included fiveaspartyl proteases, three tripeptidyl peptidases, one aldose-1-epimerase, glutaminase A, aspartic proteinase and acidproteinases, eight lipases, two ribonucleases, two glycosylhydrolases, two α-amylases, one glucoamylase, one car-boxypeptidase, eight hypothetical proteins, and severalothers (Table 6).
In order to check the presence of enzyme activity in theculture exudates, we used CZE. The four fluorescent sub-strates, aminopyrene trisulfonate (APTS)-labeled cellopen-taose, xylohexaose, mannohexaose, and GalA9, werehydrolyzed via a combination of endo- and exo-activities(results not shown). For quantitative measurement of enzymeactivity against the major classes of cell wall polymers, weused the DNS-reducing group assay. Of the four enzymeswhose activities were assessed, xylanase showed the highestactivity from day 1 onwards at 1.06 U/ml and peaked on thefifth day at 1.5 U/ml followed by a slight decrease to 1.37U/mlby the 14th day of growth. Polygalacturonase also showed anupward trend in activity from day 1 at 0.20 U/ml to 1.23 U/mlon day 7 where the activity was the highest followed by adecrease on 14th day to 0.49 U/ml. Compared to xylanase andpolygalacturonase activities, cellulase activity was lower, andthe general pattern was a steady increase from day 1 to day 5followed by a decrease on day 7 and then leveling off. Cellu-lase, like xylanase, showed peak activity at day 5 at 0.67 U/ml,but the activity of xylanase was almost 2.24-fold higher thancellulase. Activity against cellulose and pectic acid increasedgradually over the first 5 days of culture, whereas activityagainst xylan appeared on the first day and remained relativelyconstant during the rest of the culture period. Even though β-mannanases and a β-mannosidase were detected in the LC–MS/MS studies, activity against galactomannanwas too low tobe measured by the DNS assay but could be easily detected bythe more sensitive CZE assay. To estimate the productivity of
Table 6 (continued)
Identified proteinsa Accession no.a M. wt. (kDa)b Spectrum countc SignalPd
1day 7days 14days
Hypothetical protein phch_07380 108 3 31 15 Yes
Hypothetical protein phch_04700 14 28 40 27 Yes
a Accession numbers along with protein information and glycoside hydrolase family information was obtained from Pedant (http://pedant.gsf.de/)bMolecular weight of the proteins was determined theoreticallyc Quantifying changes in protein abundance between samples from different time points was done using the spectral count method, yielding a semi-quantitative analysisd SignalP was used to predict secretion signals (http://www.cbs.dtu.dk/services/SignalP/). Additional information for the presence of a signalpeptide was obtained by accessing the following URL with the model number, e.g., http://genome.jgi-psf.org/cgi-bin/dispGeneModel?db0Phchr1&id03651
Appl Microbiol Biotechnol
Author's personal copy
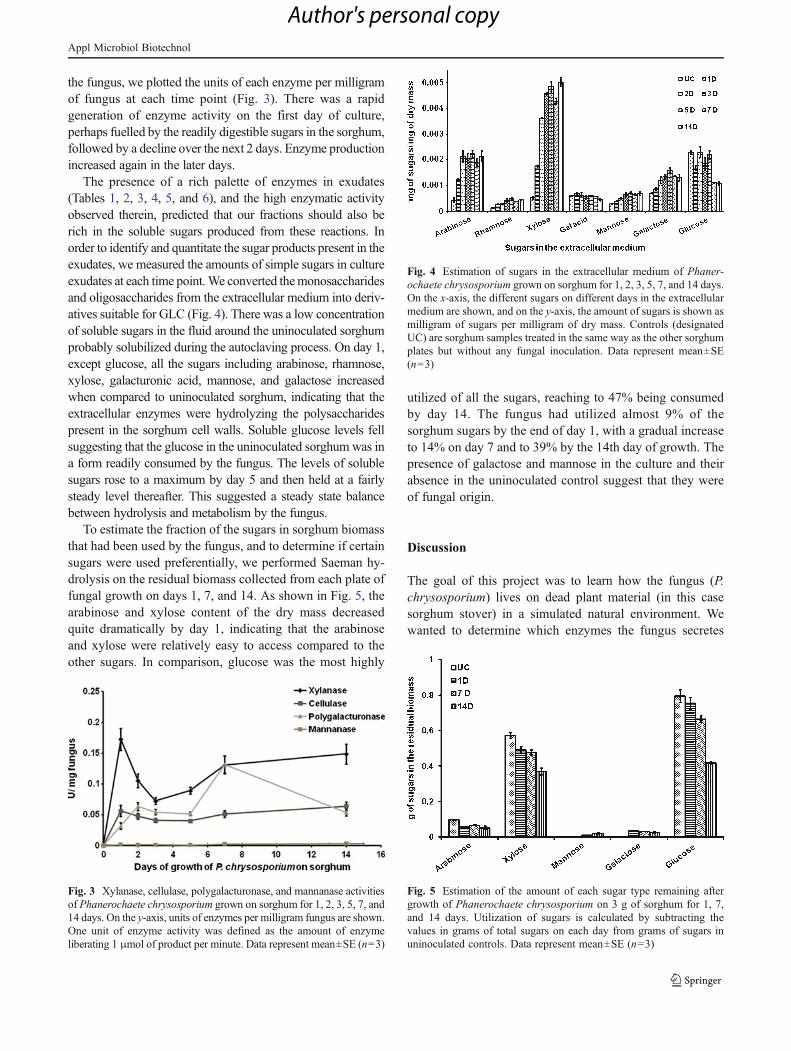
the fungus, we plotted the units of each enzyme per milligramof fungus at each time point (Fig. 3). There was a rapidgeneration of enzyme activity on the first day of culture,perhaps fuelled by the readily digestible sugars in the sorghum,followed by a decline over the next 2 days. Enzyme productionincreased again in the later days.
The presence of a rich palette of enzymes in exudates(Tables 1, 2, 3, 4, 5, and 6), and the high enzymatic activityobserved therein, predicted that our fractions should also berich in the soluble sugars produced from these reactions. Inorder to identify and quantitate the sugar products present in theexudates, we measured the amounts of simple sugars in cultureexudates at each time point.We converted themonosaccharidesand oligosaccharides from the extracellular medium into deriv-atives suitable for GLC (Fig. 4). There was a low concentrationof soluble sugars in the fluid around the uninoculated sorghumprobably solubilized during the autoclaving process. On day 1,except glucose, all the sugars including arabinose, rhamnose,xylose, galacturonic acid, mannose, and galactose increasedwhen compared to uninoculated sorghum, indicating that theextracellular enzymes were hydrolyzing the polysaccharidespresent in the sorghum cell walls. Soluble glucose levels fellsuggesting that the glucose in the uninoculated sorghum was ina form readily consumed by the fungus. The levels of solublesugars rose to a maximum by day 5 and then held at a fairlysteady level thereafter. This suggested a steady state balancebetween hydrolysis and metabolism by the fungus.
To estimate the fraction of the sugars in sorghum biomassthat had been used by the fungus, and to determine if certainsugars were used preferentially, we performed Saeman hy-drolysis on the residual biomass collected from each plate offungal growth on days 1, 7, and 14. As shown in Fig. 5, thearabinose and xylose content of the dry mass decreasedquite dramatically by day 1, indicating that the arabinoseand xylose were relatively easy to access compared to theother sugars. In comparison, glucose was the most highly
utilized of all the sugars, reaching to 47% being consumedby day 14. The fungus had utilized almost 9% of thesorghum sugars by the end of day 1, with a gradual increaseto 14% on day 7 and to 39% by the 14th day of growth. Thepresence of galactose and mannose in the culture and theirabsence in the uninoculated control suggest that they wereof fungal origin.
Discussion
The goal of this project was to learn how the fungus (P.chrysosporium) lives on dead plant material (in this casesorghum stover) in a simulated natural environment. Wewanted to determine which enzymes the fungus secretes
Fig. 3 Xylanase, cellulase, polygalacturonase, and mannanase activitiesof Phanerochaete chrysosporium grown on sorghum for 1, 2, 3, 5, 7, and14 days. On the y-axis, units of enzymes per milligram fungus are shown.One unit of enzyme activity was defined as the amount of enzymeliberating 1 μmol of product per minute. Data represent mean±SE (n03)
Fig. 4 Estimation of sugars in the extracellular medium of Phaner-ochaete chrysosporium grown on sorghum for 1, 2, 3, 5, 7, and 14 days.On the x-axis, the different sugars on different days in the extracellularmedium are shown, and on the y-axis, the amount of sugars is shown asmilligram of sugars per milligram of dry mass. Controls (designatedUC) are sorghum samples treated in the same way as the other sorghumplates but without any fungal inoculation. Data represent mean±SE(n03)
Fig. 5 Estimation of the amount of each sugar type remaining aftergrowth of Phanerochaete chrysosporium on 3 g of sorghum for 1, 7,and 14 days. Utilization of sugars is calculated by subtracting thevalues in grams of total sugars on each day from grams of sugars inuninoculated controls. Data represent mean±SE (n03)
Appl Microbiol Biotechnol
Author's personal copy

and the order of secretion of enzymes as it consumes thevarious components of sorghum biomass. After identifyingthe enzymes secreted by the fungus, we were interested indevising a minimal set of enzymes, which could be usedeconomically in vitro to convert the sorghum to fermentablesugars.
From Figs. 1 and 2, it is evident that the fungus growsrapidly for the first few days after germination on the sorghum,and our proteomics results revealed a comprehensive set ofenzymes to degrade sorghum (Tables 1, 2, 3, 4, 5, and 6).Although the hyphae continued to penetrate further in themoist sorghum until the 14th day of incubation, the dry weightof fungus decreased after 5 days and reached a steady statebetween seventh and 14th day. We hypothesize this was aresult of autolysis of the older hyphae. The chitinases, gluca-nases, proteases, and mannosidases identified in our studycould be involved in autolysis at the seventh and 14th day(Emri et al. 2008; Yamazaki et al. 2007). It is probably advan-tageous for the fungus to recycle the nitrogen needed for chitinsynthesis.
From the concentration of soluble sugars that we found inthe culture fluid, it is clear that the fungus was carbonlimited compared to normal laboratory culture conditionswhere the fungus is grown in 1% glucose. Despite thepresence of more than 7 units of cellulase activity and15 units of xylanase activity per plate, only 50% of thecellulose and 33% of the xylose had been consumed by14 days. This much enzyme activity could digest all of thecellulose and xylan in less than 12 h under ideal conditions.So, the recalcitrance of the sorghum to enzyme digestionwas evident.
Our proteomics results showed that the fungus produceda limited set of enzymes to attack all of the major compo-nents of the sorghum immediately after spore germination,but as the culture progressed, probably guided by the natureof the fragments generated by the initial enzyme set, a morecomprehensive array of enzymes was induced, suited tobreakdown of all the different linkages present.
Previous studies (Vanden Wymelenberg et al. 2005,2006, 2009, 2010) have reported on the proteins in theextracellular fluid around P. chrysosporium growing undervarious nutrient conditions in liquid medium. Their majoremphasis was upon the ligninolytic systems; however,when ball milled aspen was present, sets of glycosidehydrolases were identified that were similar, but not iden-tical to those that we found in sorghum-grown cultures.We compared the proteins found in cultures grown onsorghum to cultures grown on ball milled aspen where34 proteins were found in both, 86 were detected exclu-sively in the sorghum system that we studied, and 55were found only in the ball milled aspen cultures. Theidentities of the various proteins are reported in the Sup-plementary Material in Table S2.
As one might expect, almost half of the cellulose-directedenzymes were common to both the sorghum and aspensystems. Only one cellobiohydrolase was unique to theaspen system. One third of the xylanases were common toboth with none being unique to aspen. About one third ofthe ten pectinases were common between sorghum andaspen with two being unique to aspen. The proteins thatcould be assigned to being a part of the lignin degradationsystem were totally distinct between the two systems. Onlyenzymes that could be presumed to be involved in genera-tion of hydrogen peroxide were found in the aspen system,whereas lignin peroxidases and cellobiose dehydrogenasewere found in the sorghum system. Only a single enzymefor remodeling fungal cell walls was found in the aspensystem, but 17 were found in the sorghum system. Wedesignated a large group of proteins in the miscellaneouscategory where most of the lipases and proteases were seenonly in the sorghum system. A large number of hypotheticalproteins and proteins which one would expect to be intra-cellular were unique to the aspen system. From the ratherlarge differences in the secretome of P. chrysosporiumgrowing on sorghum vs. aspen, we suggest that the funguscaters its enzyme secretion according to the particularhabitat.
We were particularly interested in finding out what sys-tems the fungus used to overcome the recalcitrance of thebiomass to enzyme hydrolysis and if it could be possible tospeed up the hydrolysis by augmenting these systems. Wefound ample evidence of several systems used to dissociatethe lignin from hemicelluloses as well as to degrade thelignin directly and systems to attack crystalline cellulose.
We detected two glucuronoyl esterases, which hydrolyzethe ester linkage between hemicellulose and lignin, and somake the xylan more accessible (Li et al. 2007) (Table 2).One of them (phch_10701) was detected from the beginningof growth, while the other one (phch_08173) was expressedon days 7 and 14. The enzyme form (phch_10701) containscarbohydrate-binding module family-1 domain (CBD).CBDs are known to increase the concentration of enzymeson their substrates (Linder et al. 1995; Shoseyov et al.2006). The expression of phch_10701 from the beginningagrees with the hypothesis that this enzyme having the CBDis useful in the initial stage of polysaccharide degradationwhen most of the substrate is still insoluble. It has beensuggested that the enzyme without a CBD (phch_08173)takes over when most of the substrate is solubilized (Duranováet al. 2009). However, after 7 days, there is still much insolublexylan, and the CBD-containing enzyme persists. Previousstudies (Vanden Wymelenberg et al. 2009) reported only theCBD version of glucuronoyl esterase in some of the mediacontaining cellulose, but not in wood-containing media. Sur-prisingly, glucuronoyl esterase was also reported to be presentin the medium of the fungus in stationary phase growing on B3
Appl Microbiol Biotechnol
Author's personal copy

medium with adequate glucose and nitrogen (VandenWymelenberg et al. 2009).
Cellobiose dehydrogenase (phch_08874) was detectedwith a high spectrum count throughout the culture period.This enzyme oxidizes cellobiose and cello-oligomers andcan use the electrons from the reaction to reduce a broadrange of electron acceptors including ferric ions. The result-ing ferrous ions can then undergo Fenton chemistry produc-ing hydroxyl radicals from hydrogen peroxide. Thehydroxyl radicals can attack lignin-causing depolymeriza-tion. The hydroxyl radicals have also been proposed tocause chain breaks in polysaccharides, including crystallinecellulose (Henriksson et al. 2000). This enzyme was notdetected in culture media when P. chrysosporium was grownin ball milled aspen (Vanden Wymelenberg et al. 2010) or inbasal media supplemented with Avicel but was found inNorkran’s medium and Wood’s medium supplemented withAvicel (Vanden Wymelenberg et al. 2009).
We saw lignin peroxidases which use hydrogen peroxideto become strong enough oxidants to oxidize the phenolicrings of lignin and other aromatics (Wong 2009). The hy-drogen peroxide needed for these reactions could be gener-ated from glyoxal by glyoxal oxidases (phch_08874 andphch_10903) (Ander and Marzullo 1997) which were alsofound in our study. Previous studies reported multiple ligninperoxidases in nitrogen or carbon-limited basal media, whilefewer were detected in the replete medium and none in thecultures supplemented with wood (Vanden Wymelenberg etal. 2009).
The crystalline nature of cellulose contributes to its inac-cessibility to cellulases. Recently, family 61 endoglucanaseswere found to enhance the digestibility of cellulose bycellulases in lignocellulose, but not pure cellulose (Harriset al. 2010). Seven of the detected endoglucanases in ourstudy belongs to the “enigmatic” GH61 family. The mech-anism by which they do this is probably explained by thework of Vaaje-Kolstad et al. (2005), who found that chitin-binding protein 21 (CBP 21) from Serratia marcescenscauses chain scissions in crystalline chitin via a mechanisminvolving water, molecular oxygen, an electron donor, and adivalent metal coordinated to surface residues of the protein(Vaaje-Kolstad et al. 2010). There is cleavage of glycosidiclinkages and oxidation of the sugar whose bond was brokento an acid. Family 61 endoglucanases have a very similarstructure to the CBP 21, and in the crystal structure of thefamily 61 endoglucanase from Hypocrea jecorina, there is acluster of hydrophilic amino acids surrounding a divalentmetal ion on the surface of the protein just as there is in CBP21 (Karkehabadi et al. 2008). It is likely that they act in asimilar way on crystalline cellulose as CBP 21 does onchitin. Very recent work explained the lack of cellulaseenhancement on pure cellulose by family 61 endogluca-nases. It was reported by Quinlan et al. (2011) that the
family 61 endoglucanases need a reductant (present as gal-late in the lignocellulose samples but missing in pure cellu-lose samples) for activity. Presumably, the reductant reducesa cupric ion in the active site, which can then react withmolecular oxygen with subsequent oxidation of the cellu-lose on which the enzymes are bound. The oxidation of thecellulose leads to chain breaks, and so, new ends becomeaccessible to cellulases. Thus, our finding of seven differentfamily 61 endoglucanases, some with very high spectrumcounts, suggests their great importance for the degradationof cellulose by P. chrysosporium.
A search for conserved domains within the family 61 endo-glucanases shows that two of them (phch_04629 andphch_06115) contain cellulose-binding domains at theirC-termini, one phch_07005 has an extension before and afterthe glycoside hydrolase family 61 (Glyco_hydro_61) domain,and one is shorter than the others and contains a domainhomologous to tryptophan synthase beta subunit-like PLP-dependent enzymes. The other three (phch_06067,phch_06068, and phch_01789), including the one with thehighest spectrum count (phch_01789), appear to contain onlya glyco_hydro_61 domain. There are eight additional openreading frames (ORFs) in the genome containing a glyco_hy-dro_61 domain, of which, three of them have a carbohydrate-binding module (CBM), and the others do not. With theexception of phch_01789 detected in most of the culture con-ditions (except in aspen), and phch_04629 in some cellulose-containing media, the family 61 endoglucanases identified inthe various liquid cultures by Vanden Wymelenberg et al.(2005, 2009) were different from the ones that we detectedfrom the solid state sorghum cultures. The regulation of family61 endoglucanases and functioning of a CBM domain on aprotein that supposedly binds cellulose at its active site remainto be explained.
It is puzzling to see that pectin-degrading enzymes werequite abundant in our study even though sorghum containsvery little pectin. Vanden Wymelenberg et al. (2009, 2010)also detected endo- and exo-polygalacturonases along withrhamnogalacturonan hydrolase in their cultures containingcellulose or wood. Perhaps the suggestion that pectinasesopen up the wall for subsequent attack by other enzymes(Karr and Albersheim 1970) is correct, despite the smallproportion of this polymer in sorghum. We did see rham-nose and galacturonic acid in the extracellular fluid, whichprobably served as inducers for the production of the otherpectin-degrading enzymes detected in our study (Table 3).Our results, thereby, hint that the attack of pectin seemsstrategic with the backbone being attacked first followedby the side chains.
In summary, we found that P. chrysosporium secretes awide variety of enzymes for its successful growth on sor-ghum. It is not yet possible to decide on a small subset of theenzymes we detected to make an efficient cell-free mix to
Appl Microbiol Biotechnol
Author's personal copy

degrade sorghum. Further studies are required to test theeffectiveness of individual cloned glucuronoyl esterases,cellobiose dehydrogenases, lignin peroxidases, and family61 endoglucanases in making sorghum more digestible toxylanases and cellulases. Nonetheless, our results probe intothe proteome of this white-rot fungus, and monitoring theproteome over a period of time gave us useful informationregarding the secretion pattern of different protein familiesand their individual members.
Acknowledgments This workwas supported byUSDACSREES grant2007-35504-18244, a grant from The Oklahoma Bioenergy Center, andthe Oklahoma Agricultural Experiment Station. Microscopy work wasperformed in the OSUMicroscopy Laboratory using electron microscopespurchased with grants from the National Science Foundation. We appre-ciate the technical assistance offered by the staff of the Laboratory. Massspectrometry analyses were performed in the DNA/Protein ResourceFacility at Oklahoma State University, using resources supported by theNSF MRI and EPSCoR programs (award No. 0722494). We appreciatethe help and technical assistance offered by Janet Rogers in the DNA/Protein Resource Facility.
References
Abbas A, Koc H, Liu F, Tien M (2005) Fungal degradation of wood:initial proteomic analysis of extracellular proteins of Phanero-chaete chrysosporium grown on oak substrate. Curr Genet47:49–56
Ander P, Marzullo L (1997) Sugar oxidoreductases and veratryl alcoholoxidase as related to lignin degradation. J Biotechnol 53:115–131
Bendtsen JD, Nielsen H, von Heijne G, Brunak S (2004) Improvedprediction of signal peptides: SignalP 3.0. J Mol Biol 340:783–795
Brownell HH, Yu EKC, Saddler JN (2004) Steam-explosion pretreat-ment of wood: effect of chip size, acid, moisture content andpressure drop. Biotechnol Bioeng 28:792–801
Corredor DY, Salazar JM, Hohn KL, Bean S, Bean B, Wang D (2009)Evaluation and characterization of forage sorghum as feedstockfor fermentable sugar production. Appl Biochem Biotechnol158:164–179
Crawford DL, Crawford RL (1980) Microbial degradation of lignin.Enzym Microb Technol 2:11–22
Cullen D (1997) Recent advances on the molecular genetics of ligni-lolytic fungi. J Biotechnol 53:273–289
Cullen D, Kersten PJ (1996) Enzymology and molecular biology oflignin degradation. In: Bramble R, Marzluf G (eds) The mycotaIII. Springer, Berlin, pp 297–314
Datta A, Bettermann A, Kirk TK (1991) Identification of a specificmanganese peroxidase among ligninolytic enzymes secreted byPhanerochaete chrysosporium during wood decay. Appl EnvironMicrobiol 57:1453–1460
Duranová M, Spániková S, Wösten HA, Biely P, de Vries RP (2009)Two glucuronoyl esterases of Phanerochaete chrysosporium.Arch Microbiol 191:133–140
Emri T, Molnár Z, Szilágyi M, Pócsi I (2008) Regulation of autolysisin Aspergillus nidulans. Appl Biochem Biotechnol 151:211–220
Gassan J, Bledzki AK (1999) Alkali treatment of jute fibers: relation-ship between structure and mechanical properties. J Appl PolymSci 71:623–629
Hallam A, Anderson IC, Buxton DR (2001) Comparative economicanalysis of perennial, annual, and intercrops for biomass produc-tion. Biomass Bioenergy 21:407–424
Harris PV, Welner D, McFarland KC, Re E, Navarro Poulsen JC, BrownK, Salbo R, Ding H, Vlasenko E, Merino S, Xu F, Cherry J, LarsenS, Lo Leggio L (2010) Stimulation of lignocellulosic biomass hy-drolysis by proteins of glycoside hydrolase family 61: structure andfunction of a large, enigmatic family. Biochemistry 49:3305–3316
Henriksson G, Pettersson G, Johansson G, Ruiz A, Uzcategui E (1991)Cellobiose oxidase from Phanerochaete chrysosporium can becleaved by papain into two domains. Eur J Biochem 196:101–106
Henriksson G, Ander P, Pettersson B, Pettersson G (1995) Cellobiosedehydrogenase (cellobiose oxidase) as a wood degrading enzyme.Studies on cellulose, xylan and synthetic lignin. Appl MicrobiolBiotechnol 42:790–796
Henriksson G, Johansson G, Pettersson G (2000) A critical review ofcellobiose dehydrogenases. J Biotechnol 78:93–113
Karkehabadi S, Hansson H, Kim S, Piens K, Mitchinson C, SandgrenM (2008) The first structure of a glycoside hydrolase family 61member, Cel61B from Hypocrea jecorina, at 1.6 A resolution. JMol Biol 383:144–154
Karr AL, Albersheim P (1970) Polysaccharide-degrading enzymes areunable to attack plant cell walls without prior action by a “wall-modifying enzyme”. Plant Physiol 46:69–80
Kersten PJ (1990) Glyoxal oxidase of Phanerochaete chrysosporium:its characterization and activation by lignin peroxidase. Proc NatlAcad Sci U S A 87:2936–2940
Kersten P, Cullen D (2007) Extracellular oxidative systems of thelignin-degrading basidiomycete Phanerochaete chrysosporium.Fungal Genet Biol 44:77–87
Kersten PJ, Kirk TK (1987) Involvement of a new enzyme, glyoxaloxidase, in extracellular H2O2 production by Phanerochaetechrysosporium. J Bacteriol 169:2195–2201
Kirk TK, Cullen D (1998) Enzymology and molecular genetics ofwood degradation by white-rot fungi. In: Young RA, Akhtar M(eds) Environmentally friendly technologies for the pulp andpaper industry. Wiley, New York, pp 273–308
Kirk TK, Farrell RL (1987) Enzymatic “combustion” the microbialdegradation of lignin. Annu Rev Microbiol 41:465–505
Komalavilas P, Mort AJ (1989) The acetylation of O-3 of galacturonicacid in the rhamnose-rich portion of pectins. Carbohydr Res189:261–272
Lacewell RD, Teague PW, Masud SM (1986) Economics and financialfeasibility of high energy sorghum. In: Hiller EA (ed) Sorghumfor methane production: annual report. Gas Research Institute,Chicago, pp 7.1–7.25
Li XL, Spániková S, de Vries RP, Biely P (2007) Identification ofgenes encoding microbial glucuronoyl esterases. FEBS Lett581:4029–4035
Linder M, Lindeberg G, Reinikainen T, Teeri TT, Pettersson G (1995)The difference in affinity between two fungal cellulose-bindingdomains is dominated by a single amino acid substitution. FEBSLett 372:96–98
Martinez D, Larrondo LF, Putnam N, Gelpke MD, Huang K, ChapmanJ, Helfenbein KG, Ramaiya P, Detter JC, Larimer F, CoutinhoPM, Henrissat B, Berka R, Cullen D, Rokhsar D (2004) Genomesequence of the lignocellulose degrading fungus Phanerochaetechrysosporium strain RP78. Nat Biotechnol 22:695–700
Miller GL (1959) Use of dinitrosalicylic acid reagent for determinationof reducing sugar. Anal Chem 31:426–428
Mort AJ, Parker S, Kuo M (1983) Recovery of methylated saccharidesfrom methylation reaction mixtures using Sep-pak1 C18 car-tridges. Anal Biochem 133:380–384
Naran R, Pierce ML, Mort AJ (2007) Detection and identification ofrhamnogalacturonan lyase in intercellular spaces of expandingcotton cotyledons. Plant J 50:95–107
Appl Microbiol Biotechnol
Author's personal copy

Old WM, Meyer-Arendt K, Aveline-Wolf L, Pierce KG, Mendoza A,Sevinsky JR, Resing KA, Ahn NG (2005) Comparison of label-free methods for quantifying human proteins by shotgun proteo-mics. Mol Cell Proteomics 10:1487–1502
Penman D, Britton G, Hardwick K, Collin HA, Isaac S (2000) Chitinas a measure of biomass of Crinipellis perniciosa causal agent ofwitches broom disease of Theobroma cacao. Mycol Res 104:671–675
Pérez J, Muñoz-Dorado J, De-la-Rubia T, Martínez J (2002) Biodeg-radation and biological treatments of cellulose, hemicellulose andlignin: an overview. Int Microbiol 5:53–63
Pontecorvo G (1969) Genetic analysis of “somatic” cells in filamen-tous fungi. Wistar Inst Symp Monogr 9:1–8
Quinlan RJ, Sweeney MD, Lo Leggio L, Otten H, Poulsen JC, JohansenKS, Krogh KB, Jørgensen CI, TovborgM, Anthonsen A, Tryfona T,Walter CP, Dupree P, Xu F, Davies GJ, Walton PH (2011) Insightsinto the oxidative degradation of cellulose by a copper metalloen-zyme that exploits biomass components. Proc Natl Acad Sci U S A108:15079–15084
Reddy CA, D’Souza TM (1994) Physiology and molecular biology ofthe lignin peroxidases of Phanerochaete chrysosporium. FEMSMicrobiol Rev 13:137–152
Renganathan V, Usha SN, Lindenburg F (1990) Cellobiose oxidizingenzymes from lignocellulose degrading basidiomycete Phanero-chaete chrysosporium: interaction with microcrystalline cellulose.Appl Microbiol Biotechnol 32:609–613
Rothschild N, Lefkowitz A, Hadar Y, Dosoretz C (1999) Extracellularmannose-6-phosphatase of Phanerochaete chrysosporium: a lig-nin peroxidase-modifying enzyme. Arch Biochem Biophys372:107–111
Saeman JF, Moore WE, Mitchell RL, Millet MA (1954) Techniques forthe determination of pulp constituents by quantitative paper chro-matography. TAPPI 37:336–343
Saha BC, Iten LB, Cotta MA, Wu YV (2005) Dilute acid pretreatment,enzymatic saccharification and fermentation of wheat straw toethanol. Process Biochem 40:3693–3700
Sanchez C (2009) Lignocellulosic residues: biodegradation and bio-conversion by fungi. Biotechnol Adv 27:185–194
Sato S, Liu F, Koc H, Tien M (2007) Expression analysis of extracel-lular proteins from Phanerochaete chrysosporium grown on dif-ferent liquid and solid substrates. Microbiology 153:3023–3033
Schultz J, Copley RR, Doerks T, Ponting CP, Bork P (2000) SMART: aweb-based tool for the study of genetically mobile domains.Nucleic Acids Res 28:231–234
Shi J, Chinn MS, Sharma-Shivappa RR (2008) Microbial pretreatmentof cotton stalks by solid state cultivation of Phanerochaete chrys-osporium. Bioresour Technol 99:6556–6564
Shoseyov O, Shani Z, Levy I (2006) Carbohydrate binding modules:biochemical properties and novel applications. Microbiol MolBiol Rev 70:283–295
Squina FM, Mort AJ, Decker SR, Prade RA (2009) Xylan decompo-sition by Aspergillus clavatus endoxylanase. Protein Expr Purif68:65–71
Sumner JB, Graham VA (1921) Dinitrosalicylic aid: a reagent for theestimation of sugar in normal and diabetic urine. J Biol Chem47:5–9
Teymouri F, Laureano-Perez L, Alizadeh H, Dale BE (2005) Optimi-zation of the ammonia fiber explosion (AFEX) treatment param-eters for enzymatic hydrolysis of corn stover. Bioresour Technol96:2014–2018
Vaaje-Kolstad G, Horn SJ, van Aalten DM, Synstad B, Eijsink VG(2005) The non-catalytic chitin-binding protein CBP21 from Ser-ratia marcescens is essential for chitin degradation. J Biol Chem280:28492–28497
Vaaje-Kolstad G, Westereng B, Horn SJ, Liu Z, Zhai H, Sørlie M,Eijsink VG (2010) An oxidative enzyme boosting the enzymaticconversion of recalcitrant polysaccharides. Science 330:219–222
Vanden Wymelenberg A, Sabat G, Martinez D, Rajangam AS, TeeriTT, Gaskell J, Kersten PJ, Cullen D (2005) The Phanerochaetechrysosporium secretome: database predictions and initial massspectrometry peptide identifications in cellulose-grown medium. JBiotechnol 118:17–34
Vanden Wymelenberg A, Minges P, Sabat G, Martinez D, Aerts A,Salamov A, Grigoriev I, Shapiro H, Putnam N, Belinky P, DosoretzC, Gaskel J, Kersten P, Cullen D (2006) Computational analysis ofthe Phanerochaete chrysosporium v2.0 genome database and massspectrometry identification of peptides in ligninolytic cultures revealcomplex mixtures of secreted proteins. Fungal Genet Biol 43:343–356
Vanden Wymelenberg A, Gaskell J, Mozuch M, Kersten P, Sabat G,Martinez D, Cullen D (2009) Transcriptome and secretome anal-yses of Phanerochaete chrysosporium reveal complex patterns ofgene expression. Appl Environ Microbiol 75:4058–4068
Vanden Wymelenberg A, Gaskell J, Mozuch M, Sabat G, Ralph J,Skyba O, Mansfield SD, Blanchette RA, Martinez D, Grigoriev I,Kersten P, Cullen D (2010) Comparative transcriptome and secre-tome analysis of wood decay fungi Postia placenta and Phaner-ochaete chrysosporium. Appl Environ Microbiol 76:3599–3610
Vidal PF, Molinier J (1988) Ozonolysis of lignin—improvement of invitro digestibility of poplar sawdust. Biomass 16:1–17
Wong DW (2009) Structure and action mechanism of ligninolyticenzymes. Appl Biochem Biotechnol 157:174–209
Yamazaki H, Yamazaki D, Takaya N, Takagi M, Ohta A, Horiuchi H(2007) A chitinase gene, chiB, involved in the autolytic process ofAspergillus nidulans. Curr Genet 51:89–98
Yu J, Zhang JF, He J, Liu Z, Yu Z (2009) Combinations of mildphysical or chemical pretreatment with biological pretreatmentfor enzymatic hydrolysis of rice hull. Bioresour Technol100:903–908
Zheng Y, Lin HM, Tsao GT (1998) Pretreatment for cellulose hydro-lysis by carbondioxide explosion. Biotechnol Prog 14:890–896
Zheng R, Zhang H, Zhao J, Lei M, Huang H (2011) Direct andsimultaneous determination of representative byproducts in alignocellulosic hydrolysate of corn stover via gas chromatogra-phy–mass spectrometry with a Deans switch. J Chromatogr A1218:5319–5327
Appl Microbiol Biotechnol
Author's personal copy





























