Embed Size (px)
Citation preview

2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD)-Induced Accumulationof Biliverdin and Hepatic Peliosis in Rats
Marjo Niittynen,*,1 Jouni T. Tuomisto,* Seppo Auriola,† Raimo Pohjanvirta,*,‡,§ Paula Syrjala,‡Ulla Simanainen,* Matti Viluksela,* and Jouko Tuomisto*
*Department of Environmental Health, Laboratory of Toxicology, National Public Health Institute, Box 95, FIN-70701 Kuopio, Finland;†Department of Pharmaceutical Chemistry, University of Kuopio, Box 1627, FIN-70211 Kuopio, Finland; ‡National Food and
Veterinary Research Institute, Kuopio Department, Box 92, FIN-70701 Kuopio, Finland; and §Department of Food andEnvironmental Hygiene, Faculty of Veterinary Medicine, Box 57, FIN-00014 University of Helsinki, Helsinki, Finland
Received July 9, 2002; accepted September 16, 2002
2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) is a widespread,persistent, and highly toxic environmental pollutant. The mostTCDD-sensitive and the most TCDD-resistant rat strains (Long-Evans [Turku/AB] and Han/Wistar [Kuopio], respectively) werecrossbred to separate the alleles of two genes (Ahr and an uniden-tified gene “B”) mediating resistance against TCDD toxicity. Dur-ing crossbreeding, a new type of toxicity in livers of both sexes wasdetected, characterized macroscopically by intense dark green toblack color and swelling that appeared most frequently after alarge dose (300 �g/kg or more as a single intragastric dose) and afollow-up period of more than three weeks. Therefore, studieswere undertaken to identify the causative pigment chemically andto examine the hepatotoxicity histologically. The pigment frac-tions were separated by thin layer chromatography and thenanalyzed by HPLC and electrospray mass spectrometry. The pig-ment was found to consist of biliverdin and several biliverdin-related compounds. In liver histopathology carried out on malerats, progressive sinusoidal distension and hepatic peliosis withmembrane-bound cysts were seen. The clinical manifestations ofpigment accumulation were recorded most often in intermediatelyresistant rat lines such as line B (homozygous for the gene B), butnever occurred in rats expressing only the Han/Wistar (Kuopio)-type Ah receptor with an altered transactivation domain structure.
Key Words: TCDD; biliverdin; bilirubin; hepatotoxicity; accu-mulation; porphyrin metabolism; peliosis.
Polychlorinated dibenzo-p-dioxins (PCDDs) are wide-spread, persistent, and highly toxic environmental pollutants.2,3,7,8-Tetrachlorodibenzo-p-dioxin (TCDD) is the most po-tent congener among PCDDs and the most thoroughly inves-tigated model compound of this class of chemicals. Thesecompounds typically elicit a variety of biological and toxicresponses ranging from induction of cytochrome P450 (CYP)isoforms 1A1, 1A2, and 1B1 to reproductive and developmen-tal defects, immunotoxicity, thymus atrophy, endocrine imbal-ance, altered intermediary metabolism, liver toxicity, cancer,
and wasting syndrome (reviewed by Pohjanvirta and Tuomisto,1994).
Sensitivity to TCDD varies greatly among mammalian spe-cies and also among strains of the same species. The largestinterstrain difference reported in rats occurs between the Han/Wistar (Kuopio; H/W) and Long-Evans (Turku/AB; L-E) sub-strains with LD50 values � 9600 and 10–20 �g/kg TCDD,respectively (Pohjanvirta and Tuomisto, 1994). Recent studiesrevealed that the exceptional TCDD resistance of H/W ratsmainly derives from a point mutation in intron 10 of its arylhydrocarbon receptor (AhR) gene and a consequent alterationin the transactivation domain of AhR protein (Pohjanvirta etal., 1998; Tuomisto et al., 1999). It was also previously dem-onstrated that there are probably two separate genes affordingresistance to TCDD (Pohjanvirta, 1990), the other one beingstill unidentified and therefore called “gene B.” Subsequently,classical crossbreeding methods were used to segregate thesetwo genes from H/W and L-E parent strains into novel rat lines(Tuomisto et al., 1999). Three lines resulted from the breedingexperiments: line A homozygous for the mutated AhR(Ahrhw/hw, hw denoting an allele originating from the H/Wstrain) and normal gene B (Bwt/wt, wt denoting a wild-type allelefrom the L-E strain); line B homozygous for the auxiliaryresistance allele (Bhw/hw) and normal AhR (Ahrwt/wt); and line Chomozygous for wild-type alleles of both resistance genes(Tuomisto et al., 1999). LD50 values for lines A, B, and Cwere � 10000, 400–800, and 20–40 �g/kg TCDD, respec-tively. Thus, line A was as resistant as H/W, line C almost assensitive as L-E, and line B was intermediately resistant.
In necropsies carried out on TCDD-treated rats during theH/W � L-E crossing, several rats in F1, F2, and F2 � L-Egenerations were surprisingly discovered to exhibit a previ-ously unseen liver toxicity syndrome. The most severe caseshad large (more than double the expected size) swollen liversthat were mottled and dark green or black in color. When thetissue was cut with a knife, several milliliters of dark green toblack fluid leaked out. These findings prompted us to chemi-cally and histologically characterize the responsible pigment
1 To whom correspondence should be addressed. Fax: �358 17 201265.E-mail: [email protected].
TOXICOLOGICAL SCIENCES 71, 112–123 (2002)Copyright © 2002 by the Society of Toxicology
112
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

and thus to gain further insight into the possible metabolicdisturbances in these rats.
MATERIALS AND METHODS
Chemicals. TCDD (CAS# 1746-01-6; mw 321.9; purity � 99% as ana-lyzed by gas chromatography-mass spectrometry) was purchased from UFAOil Institute (Ufa, Russia). It was dissolved in corn oil (Sigma, St. Louis, MO).The composition of the buffer used in liver perfusions was 25 mM N-(2-hydroxyethyl)piperazine-N�-(2-ethanesulfonic acid) (HEPES)/1.5 mM ethyl-enediaminetetraacetic acid (EDTA)/10% glycerol/1 mM dithiothreitol, pH 7.4(abbreviated as HEGD). HEGD reagents were purchased from Sigma. Biliv-erdin dihydrochloride (80% purity), hemin chloride, and Tris-Cl (Trizma,reagent grade) were obtained from Sigma. In preliminary studies standardsolutions of biliverdin and hemin were made according to Bonkovsky et al.(1986). In mass spectrometric analysis 0.1 mM solution of biliverdin inmethanol was used as a standard. Sucrose (analytical grade) was purchasedfrom BDH Laboratory Supplies (Poole, England). Analytical grade acetoneand chloroform were purchased from Labscan Ltd. (Dublin, Ireland). Concen-trated hydrochloric acid and trichloroacetic acid were obtained from Merck(Darmstadt, Germany). Water was purified with a Milli-Q Water purificationsystem (Millipore, Bedford, MA). Methanol (HPLC-grade) and formic acid(98%) were purchased from J. T. Baker (Deventer, Netherlands).
Animal husbandry. All rats were obtained from the breeding colony of theNational Public Health Institute, Kuopio, Finland. They were housed in groupsin stainless-steel wire-mesh cages with pelleted R3 or R36 feed (Ewos,Sodertalje, Sweden) and tap water available ad libitum. The temperature in theanimal room was 21 � 1°C, relative humidity 50 � 10%, and lighting cycle12/12 h light/dark. The study plans were approved by the Animal ExperimentCommittee of the University of Kuopio and the Kuopio Provincial Govern-ment (permits 29.1.96/5Zd, 29.1.96/8Zd, 19.6.97/47Zd, and STO90/5.2.98).
Macroscopic examinations. A large number of rats (323) were tested withTCDD during the production, selection, and characterization of the new linesA, B, and C (for details, see Tuomisto et al., 1999). These rats included thethree new lines, their hybrid offspring with L-E rats, and H/W � L-E F2generation rats. They represented both genders and various age groups (4–57weeks at the time of exposure) and were treated intragastrically with a widerange of single TCDD doses (10–2000 �g/kg). The rats were euthanized a daybefore the expected death or at six weeks postexposure. However, several ratswere found dead although effort was made to identify moribund rats. Mostnonsurvivors lived two to four weeks after treatment. All rats were examinedmacroscopically at necropsy.
Instrumentation. Two different liquid chromatographic systems wereused. First, we used Waters HPLC (Waters, Milford, MA) equipped withWaters autosampler 717 and Waters diode array detector 996. In liquidchromatographic separations before mass spectrometric detection the Rheos4000 HPLC (Flux Instruments, Danderyd, Sweden) equipped with LaChromautosampler L-7200 Merck Hitachi (Hitachi, Tokyo, Japan) and Spectroflow757 UV detector was used. In both HPLC-systems the same Symmetry C18
column (5 �m; 4.6 � 150 mm) with a guard column (Waters) was used. Massspectrometric analysis was performed using a Finnigan LCQ ion trap massspectrometer fitted with an electrospray ionization source (San Jose, CA).
Pigment extraction. The livers for the preliminary HPLC-analyses werefrom an experiment designed to verify the genotype of line B by determiningthe sensitivity of male and female hybrid offspring following line B � L-Ematings. The rats were dosed intragastrically with 50 or 100 �g/kg TCDD atthe age of 8–10 weeks and monitored for 42 days. They were then killed bydecapitation. However, a few rats were ether-anesthetized and the livers werefurther perfused with HEGD to reduce the amount of blood heme in thesamples.
Later, when an experiment primarily for pigment extraction was planned,line B female rats were chosen for studies because the incidence of thesyndrome was high and the rats were homozygous in respect of the resistancegenes (Ahrwt/wt, Bhw/hw). Rats were dosed intragastrically with 300 �g/kg TCDD
at the age of 9–11 weeks. This dose was chosen because it is large enough tocause the syndrome with somewhat high incidence but it is usually not lethalto these rats. After 3 to 5 weeks, all rats were ether-anesthetized and liverswere perfused with HEGD.
In preliminary studies the pigment was extracted according to Bonkovsky etal. (1986) with minor modifications (samples were centrifuged and filteredusing 0.45 �m Spartan 30/B filter [Schleicher & Schuell, Dassel, Germany]before HPLC-analysis). Later and in mass spectrometric analysis the extractionprocedure was as follows. Liver homogenates (10%, w/v) were prepared in0.25 M sucrose/20 mM Tris-Cl (pH � 7.4) using a teflon-pestled Potter-Elvehjem glass homogenizer in a Heidolph homogenizer device. Proteins wereprecipitated by adding 0.8 ml 15% trichloroacetic acid to 0.5 ml of homoge-nate. The resultant mixture was vortex-mixed and centrifuged (10,800 � g, 4min, room temperature). Uncolored supernatant was discarded. Precipitate wassuspended in 0.8 ml of ice-cold acetone/concentrated HCl (97.5:2.5, v/v),vortex-mixed, and centrifuged (10,800 � g, 4 min, room temperature). Super-natants of 10 parallel samples were combined. The resulting solution, whichnow included the pigment, was concentrated under nitrogen to a final volumeof about 500 �l. At this stage, the mixture appeared in two phases. The lowerone contained the pigment and its volume was 50–80 �l. The upper phase wasdiscarded. Samples were kept on ice during extraction.
Preparative thin-layer chromatography. Concentrated pigment was sep-arated to its components by applying it on silica gel plates (SIL G-25,Macherey-Nagel, Duren, Germany). Development was done with chloroform:methanol:water (65:25:3). Bands were scraped off the plates and suspended inmethanol (300 �l). The resultant mixture was vortex-mixed and silica wasremoved by centrifugation. Methanol extracts were filtered using 0.45 �mSpartan 30/B filter (Schleicher & Schuell, Dassel, Germany) and stored at–20°C for further analysis.
Liquid chromatography. In preliminary studies, liquid chromatography(solvents, gradient, and flow rate) was performed according to Bonkovsky etal. (1986) using UV-VIS–detection at wavelength 377 nm. Spectral databetween 250 and 750 nm was collected. In mass spectrometric analysis thefollowing liquid chromatography conditions were used. Methanol containing0.2% formic acid and water with 0.2% formic acid were used as eluents. A 20min gradient was employed to increase methanol from 50 to 100%. Methanolwas maintained at 100% for 5 min, after which the column was restored toinitial conditions in 2 min. An automatic injection volume of 20 �l with aflow-rate of 1 ml/min was used. The eluate from HPLC column was divided ina 1:9 ratio. The smaller part was directed to the mass spectrometer and thelarger to the UV detector. UV detection was carried out at 377 nm.
Electrospray mass spectrometry. All extracts were analyzed using posi-tive ion mode. The capillary temperature was 225°C, source voltage 4 kV,sheath gas flow 100 (arbitrary units, scale 0–100 units), capillary voltage 20 V,and tube lens offset 10 V. Maximum ion time was 200 ms.
Histopathology. In the histological examination, ten-week-old male rats ofline B were used. The rats were dosed 300 �g/kg intragastrically and decap-itated 2, 7, 14, 28, or 35 days later. Liver samples were preserved in 10%neutral buffered formalin, dehydrated, embedded in paraffin wax and cut to thethickness of 5 �m. The tissue slices were mounted on glass slides, stained withMayer’s hematoxylin and eosin or sudan IV, and examined using light micro-scope independently by two pathologists (P.S., R.P.).
RESULTS
Necropsy Observations
In the course of the crossbreeding to separate the resistancegenes, a large number of rats of both genders and of variousage groups (4–57 weeks) were treated with a wide range ofTCDD doses (10–2000 �g/kg) and subjected to macroscopicexamination at necropsy. When the first observations of theblack liver syndrome were made, these findings were recorded
113TCDD AND BILIVERDIN ACCUMULATION IN RAT LIVER
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from
from all rats during subsequent necropsies. Due to the hetero-geneous rat population, all age groups were combined and twodose groups were formed, namely “low” (10–100 �g/kg) and“high” (300–2000 �g/kg).
The majority of severe black liver syndrome cases wererecorded in line B, line B � L-E, or line A � L-E rats, i.e., inrats with at least one resistance allele. In these lines, theincidence of the syndrome increased with dose. In line B malesthe incidence was 4/27 (15%) with a low dose and 9/20 (45%)with a high dose. In line B females the incidences were 2/22(9%) and 25/48 (52%), respectively. In line A � L-E malesthere were no cases (0/5) with the low dose, and the high doseresulted in 3/6 (50%) incidence. In line A � L-E females theincidences were 1/13 (8%) and 4/9 (44%), respectively.
Intriguingly, however, no cases appeared in line A rats orother rats homozygous for the resistance allele of the AhR gene(Ahrhw/hw) although 14 line A males and 22 females weretreated with doses of 700–2000 �g/kg. No black livers wereseen even in three line A males that were given extremely high
total doses of 12,000, 18,000, and 30,000 �g/kg TCDD (un-published data). In addition, 6 males and 9 females fromH/W � L-E F2 generation with verified Ahrhw/hw phenotypeswere treated with 1000 �g/kg and no black livers were seen.
Sensitive lines (C and C � L-E), which due to their sensi-tivity were only given low doses (10–100 �g/kg), exhibitedonly a few mild cases. It is not clear at this point whether thisis a true line difference or whether it merely reflects the lowerdoses used in these lines. In line C males and females theincidences were 2/40 (5%) and 0/35, respectively. In line C �L-E males and females there were 5/9 (56%) and 0/4 cases,respectively.
In all these cases the incidences were at least that mentionedabove; they may have been slightly higher, because postmor-tem changes in liver had made classification unreliable in a fewrats that were found dead (these were classified as “no syn-drome”). The proportion of these rats was highest in line Cfemales (23%) and in the low dose group of line A � L-Emales (20%), and it was 10% or less in all other groups.
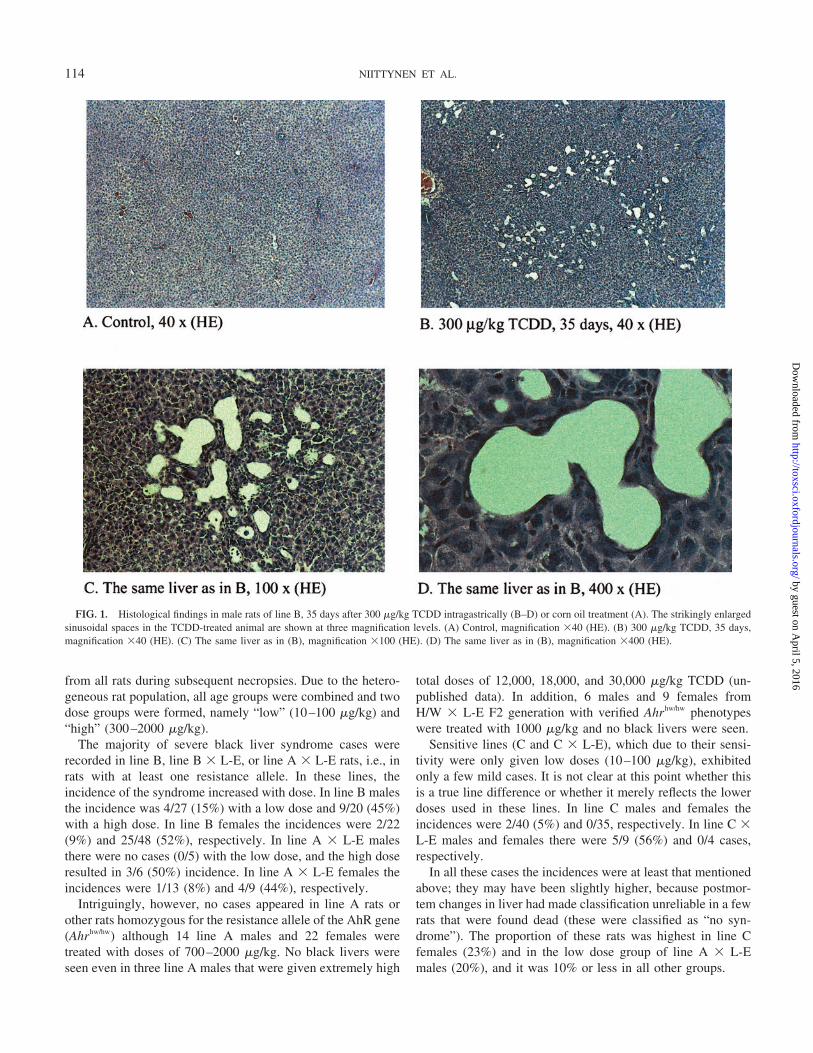
FIG. 1. Histological findings in male rats of line B, 35 days after 300 �g/kg TCDD intragastrically (B–D) or corn oil treatment (A). The strikingly enlargedsinusoidal spaces in the TCDD-treated animal are shown at three magnification levels. (A) Control, magnification �40 (HE). (B) 300 �g/kg TCDD, 35 days,magnification �40 (HE). (C) The same liver as in (B), magnification �100 (HE). (D) The same liver as in (B), magnification �400 (HE).
114 NIITTYNEN ET AL.
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

Although the incidence increased with dose, the proportionof severe and mild black liver syndromes did not show a doseresponse according to the data from lines that were treated bothwith the high and the low dose. In the groups with at least 3cases of the syndrome, the proportion of severe cases was25–66% of all cases irrespective of the dose.
The age of rat at exposure does not seem to be a criticalfactor in the development of the syndrome. The syndrome wasobserved both in rats exposed at the age of 5 weeks as well asin rats exposed at the age of 50 weeks. Usually it took 3–5weeks for syndrome to develop. Of all observations, only 6 outof 99 were made before day 21 postexposure.
Histopathology
In the TCDD-treated rats, a characteristic liver response(Pohjanvirta et al., 1989; Pohjanvirta and Tuomisto, 1994) wasdiscernible from day 7 on. This consisted of swollen hepato-cytes with hydropic degeneration, formation of giant hepato-cytes with multiple nuclei, necrosis of single cells or of smallfoci, increased number of mitotic figures, vacuolization andinfiltration of inflammatory cells in sinusoids. There was in-terindividual variation in severity of these manifestations ofTCDD toxicity. Additionally, however, in the present studypeculiar histopathological changes were found in line B ratsthat have never been recorded in L-E or H/W rats in responseto TCDD or other dioxin congeners. From day 28 on, hepaticsinusoids displayed a clear tendency towards distension. Thisdilatation was progressive and reached a hepatic peliosis-likestage with membrane-bound cysts by day 35 in 4 out of 5 rats;one of these showed a mild black liver syndrome at necropsy.Some of the cysts appeared empty while others containedproteinaceous material, erythrocytes, or leukocytes (Figs. 1B–1D). At later phases, sinusoidal distension was accompanied bymild to moderate hepatic fibrosis and modest bile duct prolif-eration.
One of the TCDD-treated rats of line B, which had to beeuthanized on day 22 in a moribund state, exhibited excessivehepatic steatosis.
Preliminary HPLC Analyses
The dark green color of the pigment suggested that it couldcontain biliverdin, a heme breakdown product. We first ana-lyzed the pigment with mere extraction followed by HPLC-analysis (Bonkovsky et al., 1986). However, the resultingchromatograms were not unambiguous. In the chromatogramsof black liver samples there were a number of poorly separatedpeaks near the retention time of biliverdin. Many of thesepeaks had UV-VIS spectrum similar to that of the biliverdinstandard. We concluded that the peak that had identical reten-tion time and UV-VIS spectrum with the standard was likely tobe biliverdin. The next task was the identification of the peaksthat had similar UV-VIS spectra but different retention times.The largest peak in the chromatograms was assumed to beheme on the basis of similar UV-VIS spectrum and identicalretention time with an authentic standard.
Based on these observations the procedure was further mod-ified as follows. Livers were perfused in order to diminish theamount of blood heme. The perfusion changed the color of thelivers from red with black spots to pale with green spots. Thisindicates that the pigment of interest was indeed green. Formore efficient extraction of pigment, the extraction procedurewas optimized (see Materials and Methods). In addition, thepigment was purified by preparative thin-layer chromatographyprior to further analysis. These modifications improved thereliability of identification of the compounds, which was fi-nally completed by mass spectrometry.
Thin-Layer Chromatography
The green liver pigment from a TCDD-treated rat separatedinto three main bands by preparative thin-layer chromatogra-phy (TLC; Fig. 2). The approximate Rf values were 0.67 (band1, most polar), 0.71 (band 2), and 0.87 (band 3, least polar).Bands 1 and 2 had a bluish-green color. Band 3 was green,having some brown pigment mixed in it. In control samplesfrom untreated rats bands 1 and 2 were missing. Instead, therewas a band having about the same Rf value as band 3 in a
FIG. 2. Thin-layer chromatogram of hepatic pigments from female rats of line B, 35 days after 300 �g/kg TCDD intragastrically or vehicle. BV, biliverdinstandard. The three TLC bands analyzed further are numbered.
115TCDD AND BILIVERDIN ACCUMULATION IN RAT LIVER
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from
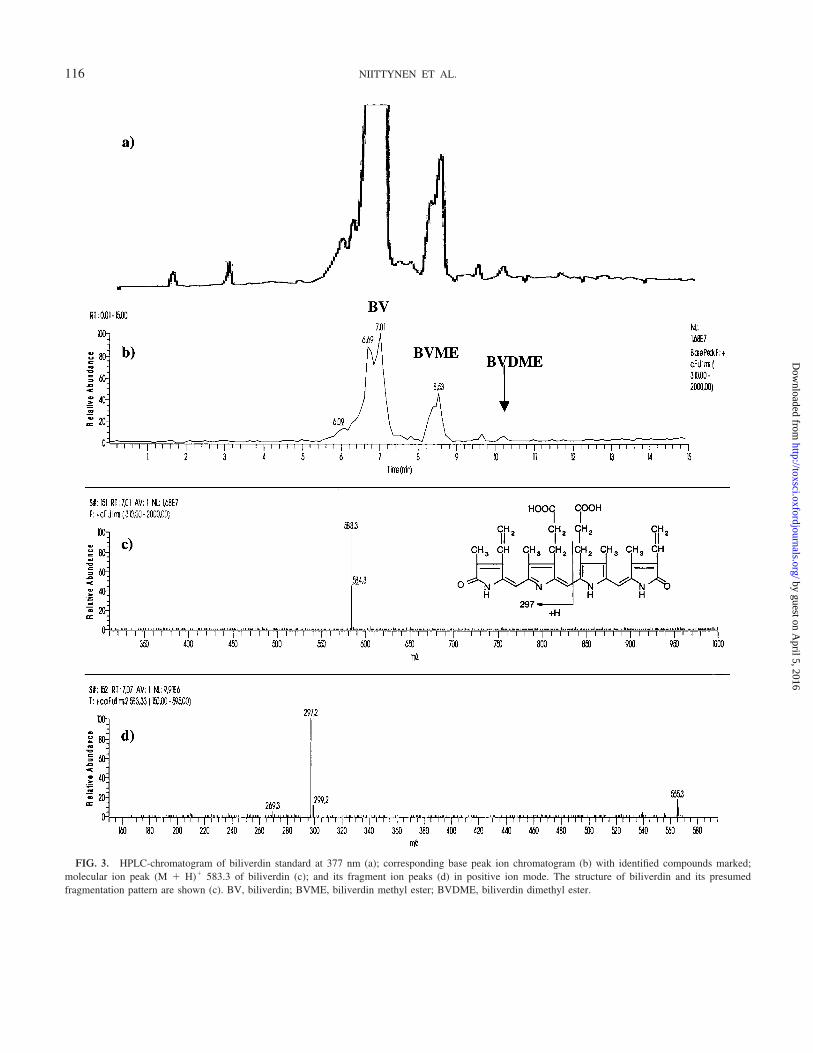
FIG. 3. HPLC-chromatogram of biliverdin standard at 377 nm (a); corresponding base peak ion chromatogram (b) with identified compounds marked;molecular ion peak (M � H)� 583.3 of biliverdin (c); and its fragment ion peaks (d) in positive ion mode. The structure of biliverdin and its presumedfragmentation pattern are shown (c). BV, biliverdin; BVME, biliverdin methyl ester; BVDME, biliverdin dimethyl ester.
116 NIITTYNEN ET AL.
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

sample from a TCDD-treated rat, but the color of this band wasreddish-brown without any green pigment.
Identification of Compounds
Parallel to MS detection, UV detection was also carried outat 377 nm, the absorption maximum of biliverdin hydrochlo-
ride in methanol (Lemberg and Legge, 1949). UV detectionwas used to confirm that the compounds detected by MS alsoabsorbed light at the same wavelength as biliverdin. This couldbe used as supporting information in the identification proce-dure. Only compounds absorbing light at 377 nm are consid-ered below.
FIG. 4. The analysis of the TLC-band 3. HPLC-chromatogram at 377 nm (a); corresponding base peak ion chromatogram (b) with identified compoundsmarked; molecular ion peak (M � H)� 583.3 of expected biliverdin (c); and its fragment ion peaks (d) in positive ion mode. BV, biliverdin; H, heme; BVME,biliverdin methyl ester; HME, heme methyl ester.
117TCDD AND BILIVERDIN ACCUMULATION IN RAT LIVER
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

The TLC band 3 was found to mainly consist of biliverdinand heme.2 The identification of biliverdin was based on asimilar retention time to an authentic standard and on theinformation from MS-MS tandem mass spectrometry. Bothparent and daughter ion spectra of the putative biliverdin insample were identical with the authentic standard (Figs. 3 and4). The positive ion mass spectra of biliverdin showed an (M �H)� ion at m/z 583.3 (calculated from atom weights: 583.7;Table 1). The other main component of TLC-band 3 wasexpected to be heme, which showed an [M(protoporphyrin IX)– 2H � Fe3�]� ion at m/z 616.3 (calculated 616.5).
The TLC band 1 was found to mainly consist of a compoundshowing an (M � H)� ion at m/z 759.3 (Fig. 5). This isproposed to be biliverdin monoglucuronide on the basis of thefollowing evidence. First, the calculated molecular mass ofbiliverdin monoglucuronide is 758.7, so it should show an(M � H)� ion at m/z 759.7. Second, there was a daughter ionwith the same m/z 583.2 as for biliverdin (Table 1). There wasalso a daughter ion at the m/z 297.2, which is also a daughterion of biliverdin. Third, on the basis of Rf-value and retentiontime the compound with m/z 759.3 (M � H)� is more polarthan biliverdin. Glucuronidation is a typical conjugation reac-tion in biotransformation of toxic compounds in the liver inorder to make them more soluble in water. Biliverdin accumu-lation in liver is an abnormal situation that could lead to theconjugation of biliverdin with glucuronate. Alternatively,biliverdin monoglucuronide might result from oxidation ofbilirubin monoglucuronide, a normal intermediate in hemecatabolism.
The main component of the TLC band 2 was a compoundshowing an (M � H)� ion at m/z 741.4 (compound I in Table1, Fig. 6). This is equivalent in size to a biliverdin monoglu-curonide with a loss of a hydroxyl group. Again, the daughterions at the m/z 583.3 and m/z 297.2 were found. Loss ofhydroxyl group could also explain why this compound isslightly less polar than biliverdin monoglucuronide.
The green pigment was also found to contain compoundsthat showed (M�H)� ions at m/z 597.3 and 773.4 (Table 1).We propose that these compounds are biliverdin methyl ester(mw � 596.7) and biliverdin monoglucuronide methyl ester(mw � 772.7), respectively (Fig. 7). Daughter ion patterns(Table 1) support this hypothesis. Because biliverdin methylester (and dimethyl ester) were also found in the standardsolution, it seems possible that methyl esters are formed spon-taneously in methanol. Furthermore, the relative amount of theputative methyl esters compared with free biliverdin remark-ably increased during storage of the standard solution.
As shown in Table 1, the putative biliverdin monoglucu-ronide methyl ester eluted at retention times of both 6.6 minand 7.7 min. Also daughter ion profiles differ at differentretention times. Because biliverdin monoglucuronide has twocarboxylic groups there are also two possible forms of biliv-erdin monoglucuronide methyl ester. Different polarities ofthese compounds can result in different retention times.
The areas corresponding to the TLC bands 1, 2, and 3 werealso scraped off the control TLC separation (Fig. 2), extractedand analyzed with LC-MS. The control extract of bands 1 and2 did not show any absorption peaks at 377 nm. The maincompounds found in the band 3 control extract were identifiedas heme and heme methyl ester on the basis of their retentiontimes and the information from MS-MS tandem mass spec-trometry. The putative heme methyl ester showed an [M(pro-toporphyrin IX methyl ester) – 2H � Fe3�]� ion at m/z 630.3(calculated 630.5). No biliverdin or its derivatives were foundin control samples.
DISCUSSION
TCDD is known to affect both the synthesis and degradationof heme giving rise to porphyria and jaundice, respectively(Pohjanvirta and Tuomisto, 1994; see Fig. 8 for flowchart ofheme metabolism). The porphyrinogenic effect of TCDD re-sults in accumulation of uroporphyrin in the liver due toinhibition of uroporphyrinogen decarboxylase. CYP1A2 ap-pears to have a critical role in the development of this uropor-
2 This analysis is qualitative in nature. Therefore only rough estimates of therelative amounts of compounds are given assuming an equal response of thesecompounds in MS detection.
TABLE 1Compounds Found in Green Liver Pigment Extracted from the Livers of Line B Female Rats 35 Days after a Single TCDD Dose
Retention time inHPLC (min) Compound TLC band (M�H)� ion Daughter ions
6.4 Biliverdin monoglucuronide 1 759.3 583.2; 741.3; 723.3; 297.16.6 Biliverdin monoglucuronide methyl ester 2 773.4 583.3; 297.2; 723.2; 755.36.9 Compound I 2 741.4 455.3; 723.3; 297.2; 583.37.0 Biliverdin 3 583.3 297.2; 565.37.7 Biliverdin monoglucuronide methyl ester 2 773.4 597.3; 755.38.0 Heme 3 616.3 557.2; 543.28.5 Biliverdin methyl ester 3 597.3 311.2; 297.3; 579.2; 313.28.7 Heme methyl ester 3 630.3 571.2; 557.2
Note. The dose was 300 �g/kg, intragastrically. The most abundant compound in each TLC-band is in bold print.
118 NIITTYNEN ET AL.
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

FIG. 5. The analysis of the TLC-band 1. HPLC-chromatogram at 377 nm (a); corresponding base peak ion chromatogram (b), identified compound marked;molecular ion peak (M � H)� 759.3 of putative biliverdin monoglucuronide (c); and its fragment ion peaks (d) in positive ion mode. The structure of biliverdinmonoglucuronide and presumed fragmentation pattern are shown (c). BVMG, biliverdin monoglucuronide.
119TCDD AND BILIVERDIN ACCUMULATION IN RAT LIVER
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

phyria (Smith et al., 2001). The mechanism of jaundice is moreobscure at the moment. TCDD elevates both total and conju-gated bilirubin levels in serum rapidly and progressively (Un-kila et al., 1994). The increase in serum bilirubin could be dueto diminished clearance (Choe and Yang, 1983; Yang et al.,1983) or augmented formation (Mitchell et al., 1990) of bili-rubin in the liver by TCDD. On the other hand, in congenitallyjaundiced Gunn rats a single dose of TCDD reduced serumbilirubin levels by inducing hepatic bilirubin catabolism (Co-hen et al., 1986).
The present study reveals a novel heme-related syndromecaused by TCDD. It is characterized by large black livers,
which accumulate a green pigment. The syndrome mainlyaffected the intermediately TCDD-resistant rat lines such asline B. Both sexes were affected with about equal frequency.An interesting observation was that the syndrome was neverseen—not even after massive doses of TCDD (up to 30,000�g/kg given to a single animal)—in line A or in other ratsexpressing only the H/W-type AhR. This suggests a pivotalinvolvement of AhR transactivation domain in the molecularmechanism of the syndrome. It also implies that the syndromebelongs to type II dioxin effects, where the magnitude of effectvaries broadly among different genotypes (Simanainen et al.,2002; Tuomisto et al., 1999).
FIG. 6. The analysis of the TLC-band 2. HPLC-chromatogram at 377 nm (a); corresponding base peak ion chromatogram (b), identified compounds marked;molecular ion peak (M � H)� 741.4 of compound I (c); and its fragment ion peaks (d) in positive ion mode. Comp. I, compound I; BVMG, biliverdinmonoglucuronide; BVMGME, biliverdin monoglucuronide methyl ester.
120 NIITTYNEN ET AL.
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

Based on our current findings, the green pigment accumu-lating in the livers of TCDD-treated rats is likely to consist ofbiliverdin and its derivatives (biliverdin monoglucuronide,compound I, and methyl esters of biliverdin and biliverdinmonoglucuronide). Of these, only biliverdin was identifiedhere with the aid of a standard compound. Further analysisusing standards (some of which are not readily available) forthe other compounds also would verify their nature. We as-sume that biliverdin monoglucuronide is formed in the livereither from biliverdin or from bilirubin monoglucuronide. It issomewhat surprising that no biliverdin diglucuronide wasfound in the pigment. The reason for this is not known.However, it has been shown that when bilirubin conjugatesexist abnormally in human serum, they are predominantlymonoglucuronides (Murray, 1988). The putative methyl estersof biliverdin and biliverdin monoglucuronide are probablyformed during sample preparation. The exact nature and for-mation of compound I are not known.
Biliverdin is an intermediate compound formed in hemebreakdown and oxidation in the reticuloendothelial system. Itis rapidly reduced to bilirubin, which is transported to liver,glucuronized, and excreted to bile (Fig. 8). Upstream inductionof heme degradation is thus one possible explanation for in-creased biliverdin levels. This pathway has also other functionsin addition to heme metabolism. Heme oxygenase (HO) is theonly enzyme capable of producing carbon monoxide (CO), animportant stress mediator and neurotransmitter. Biliverdin andiron are the by-products of CO production, which is catalyzedby two isozymes of HO, the inducible HO-1 and the constitu-tively expressed HO-2. Hexachlorobenzene stimulates HO-1 inrat liver (Stonard et al., 1998). Hexachlorobenzene and otherpolyhalogenated compounds are also thought to cause oxida-tive stress, a strong inducer of HO-1, via cellular hepatic ironand CYP1A (Maines, 1999; Stonard et al., 1998). On the other
hand, biliverdin reductase displays extensive microheterogene-ity in rat organs, and bromobenzene has been shown to selec-tively suppress the main variant in liver but not in spleen(Huang et al., 1989). Phosphorylation state critically regulatesbiliverdin reductase activity (Salim et al., 2001), and TCDDmodifies protein kinase activities (Enan and Matsumura, 1995).Thus, TCDD treatment could conceivably lead to an imbalancebetween the hepatic activities of HO and biliverdin reductase.The present findings warrant studying whether TCDD modu-lates the function of these enzymes.
Another possible source of biliverdin is oxidation frombilirubin (Fig. 8). A number of studies have suggested duringthe last few years that bilirubin is an important antioxidantacting as radical scavenger in various forms of oxidative stress(Elbirt and Bonkovsky, 1999; Galbraith, 1999; Ryter and Tyr-rell, 2000; Stocker et al., 1987). Oxidative stress might lead toincreased oxidation of bilirubin to biliverdin, instead of glucu-ronidation and excretion, although experimental data are con-flicting in this respect (Dudnik and Khrapova, 1998; Minetti etal., 1998).
To the best of our knowledge, the peliosis-like sinusoidaldistension found here has not been reported before in the liversof TCDD-treated rats. However, it has been recorded in ratstreated daily for 91 days with 3 or 10 �g/kg 2,3,7,8-TBDD,i.e., the corresponding brominated dioxin congener (Ivens etal., 1993). Furthermore, repeated exposure to hexachloroben-zene is capable of eliciting the lesion (Arnold et al., 1985;Smith et al., 1993), and iron overload potentiates the effect(Smith et al., 1993). Hepatic peliosis is frequently seen in agedLong-Evans Cinnamon rats (an animal model for copper ac-cumulation in the liver [Wilson’s disease]; Onaya et al., 2000).These findings suggest involvement of oxygen radicals in thepathogenesis of the condition. Another contributing factormight be the TCDD-induced wasting, since hepatic peliosis has
FIG. 7. Structures and presumedfragmentation patterns of biliverdinmethyl esters (a and b) and biliverdinmonoglucuronide methyl esters (c andd).
121TCDD AND BILIVERDIN ACCUMULATION IN RAT LIVER
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

been observed in patients with chronic wasting diseases such astuberculosis or cancer (Burger and Marcuse, 1952; Hamiltonand Lubitz, 1952). Thus, biliverdin accumulation and sinusoi-dal distension may share common pathogenetic factors. Theyfurther displayed similar temporal patterns. However, sinceonly one of the four peliotic rats at 35 days exhibited the blackliver syndrome, these two phenomena may be coinciding butcausally unrelated.
In the present study the incidence of the black liver syn-drome was very similar in both sexes, but the histopathologicalexamination was limited only to male rats. However, in aprevious study with hexachlorobenzene female rats were re-ported to be even more sensitive than males to hepatic peliosis(Arnold et al., 1985), the principal morphological finding ofthe present study. Therefore, it seems likely that females alsowould exhibit hepatic peliosis. Indeed, the macroscopic find-ings were the same in both sexes.
In conclusion, we have demonstrated that TCDD may, incertain conditions, cause a dramatic accumulation of bile pig-ments, notably biliverdin and its conjugates, and bring aboutpeliosis-like histopathological changes in rat liver.
ACKNOWLEDGMENTS
We thank Ms. Arja Tamminen and Ms. Minna Voutilainen for excellenttechnical assistance and Dr. Veli-Matti Kosma for work leading to the firsthistological observations of this syndrome. This study was supported by theAcademy of Finland, the Finnish Research Program on Environmental Health(Project 42551), and the European Commission (Contracts ENV4-CT96-0336and QLK4-1999-01446).
REFERENCES
Arnold, D. L., Moodie, C. A., Charbonneau, S. M., Grice, H. C., McGuire,P. F., Bryce, F. R., Collins, B. T., Zawidzka, Z. Z., Krewski, D. R., Nera,E. A., and Munro, I. C. (1985). Long-term toxicity of hexachlorobenzene inthe rat and the effect of dietary vitamin A. Food Chem. Toxicol. 23,779–793.
Bonkovsky, H. L., Wood, S. G., Howell, S. K., Sinclair, P. R., Lincoln, B.,Healey, J. F., and Sinclair, J. F. (1986). High-performance liquid chromato-graphic separation and quantitation of tetrapyrroles from biological materi-als. Anal. Biochem. 155, 56–64.
Burger, R. A., and Marcuse, P. M. (1952). Peliosis hepatis. Report of a case.Am. J. Clin. Pathol. 22, 569–573.
Cardenas-Vazquez, R., Yokosuka, O., and Billing, B. H. (1986). Enzymicoxidation of unconjugated bilirubin by rat liver. Biochem. J. 236, 625–633.
FIG. 8. Heme biosynthetic and degradation pathways. The known effect of TCDD in heme biosynthesis is marked. ALAD, �-aminolevulinate dehydratase;ALAS, �-aminolevulinate synthase; BRO, bilirubin oxidase; BVR, biliverdin reductase; CPO, coproporphyrinogen oxidase; FEC, ferrochelatase; HCB,hexachlorobenzene; HO, heme oxygenase; PBGD, porphobilinogen deaminase; PCB, polychlorinated biphenyl; PPO, protoporphyrinogen oxidase; UCoS,uroporphyrinogen III cosynthase; UDP-GT, UDP-glucuronyltransferase; UROD, uroporphyrinogen decarboxylase. The flowchart (including the effect of TCDD)is based on Maines (1999), Murray (1988), and Sassa and Maines (2001). aThe presence of bilirubin oxidase in rat liver has been demonstrated (Cardenas-Vazquez et al., 1986; Yokosuka and Billing, 1987) but the role of this enzyme in vivo is obscure.
122 NIITTYNEN ET AL.
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from

Choe, S. Y., and Yang, K. H. (1983). Effect of butylated hydroxytoluene(BHT) and butylated hydroxyanisole (BHA) on carbon tetrachloride andTCDD hepatotoxicity. Korean Biochem. J. 16, 315–321.
Cohen, A. N., Kapitulnik, J., Ostrow, J. D., and Webster, C. C. (1986). Effectof combined treatment with 2,3,7,8-tetrachlorodibenzo-p-dioxin and photo-therapy on bilirubin metabolism in the jaundiced Gunn rat. Hepatology 6,490–494.
Dudnik, L. B., and Khrapova, N. G. (1998). Characterization of bilirubininhibitory properties in free radical oxidation reactions. Membr. Cell Biol.12, 233–240.
Elbirt, K. K., and Bonkovsky, H. L. (1999). Heme oxygenase: Recent advancesin understanding its regulation and role. Proc. Assoc. Am. Physicians 111,438–447.
Enan, E., and Matsumura, F. (1995). Evidence for a second pathway in theaction mechanism of 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD). Signifi-cance of Ah-receptor mediated activation of protein kinase under cell-freeconditions. Biochem. Pharmacol. 49, 249–261.
Galbraith, R. (1999). Heme oxygenase: Who needs it? Proc. Soc. Exp. Biol.Med. 222, 299–305.
Hamilton, F. T., and Lubitz, J. M. (1952). Peliosis hepatis: A report of 3 caseswith discussion of pathogenesis. Arch. Pathol. 54, 564–572.
Huang, T. J., Trakshel, G. M., and Maines, M. D. (1989). Microheterogeneityof biliverdin reductase in rat liver and spleen: Selective suppression ofenzyme variants in liver by bromobenzene. Arch. Biochem. Biophys. 274,617–625.
Ivens, I. A., Loser, E., Rinke, M., Schmidt, U., and Mohr, U. (1993). Sub-chronic toxicity of 2,3,7,8-tetrabromodibenzo-p-dioxin in rats. Toxicology83, 181–201.
Lemberg, R., and Legge, J. W. (1949). Hematin Compounds and Bile Pig-ments. Interscience Publishers, New York.
Maines, M. D. (1999). Overview of heme degradation pathway. In CurrentProtocols in Toxicology (M. D. Maines, L. G. Costa, D. J. Reed, S. Sassa,and I. G. Sipes, Eds.), pp. 9.1.1–9.1.5. John Wiley & Sons, Inc., New York.
Minetti, M., Mallozzi, C., Di Stasi, A. M., and Pietraforte, D. (1998). Bilirubinis an effective antioxidant of peroxynitrite-mediated protein oxidation inhuman blood plasma. Arch. Biochem. Biophys. 352, 165–174.
Mitchell, D. Y., Madhu, C., and Klaassen, C. D. (1990). Hepatotoxicantselevate serum bilirubin by increasing formation not decreasing eliminationof bilirubin. Tox. Sci. 10(Suppl.), 61 (Abstract).
Murray, R. K. (1988). Porphyrins and bile pigments. In Harper’s Biochemis-try, 21st ed. (R. K. Murray, D. K. Granner, P. A. Mayes, and V. W. Rodwell,Eds.), pp. 319–334. Appleton & Lange, Norwalk, CT.
Onaya, H., Itai, Y., Yoshioka, H., Doy, M., and Mitsumori, F. (2000). Peliosishepatis and neoplastic/dysplastic lesions in aged male Long-Evans Cinna-mon rats: MR imaging with pathologic correlation. Magn. Reson. Imaging18, 143–150.
Pohjanvirta, R. (1990). TCDD resistance is inherited as an autosomal dominanttrait in the rat. Toxicol. Lett. 50, 49–56.
Pohjanvirta, R., Kulju, T., Morselt, A. F., Tuominen, R., Juvonen, R., Rozman,K., Mannisto, P., Collan, Y., Sainio, E.-L., and Tuomisto, J. (1989). Target
tissue morphology and serum biochemistry following 2,3,7,8-tetrachloro-dibenzo-p-dioxin (TCDD) exposure in a TCDD-susceptible and a TCDD-resistant rat strain. Fundam. Appl. Toxicol. 12, 698–712.
Pohjanvirta, R., and Tuomisto, J. (1994). Short-term toxicity of 2,3,7,8-tetrachlorodibenzo-p-dioxin in laboratory animals: Effects, mechanisms andanimal models. Pharmacol. Rev. 46, 483–549.
Pohjanvirta, R., Wong, J. M., Li, W., Harper, P. A., Tuomisto, J., and Okey,A. B. (1998). Point mutation in intron sequence causes altered carboxyl-terminal structure in the aryl hydrocarbon receptor of the most 2,3,7,8-tetrachlorodibenzo-p-dioxin-resistant rat strain. Mol. Pharmacol. 54, 86–93.
Ryter, S. W., and Tyrrell, R. M. (2000). The heme synthesis and degradationpathways: Role in oxidant sensitivity. Heme oxygenase has both pro- andantioxidant properties. Free Radical Biol. Med. 28, 289–309.
Salim, M., Brown-Kipphut, B. A., and Maines, M. D. (2001). Human biliv-erdin reductase is autophosphorylated, and phosphorylation is required forbilirubin formation. J. Biol. Chem. 276, 10929–10934.
Sassa, S., and Maines, M. D. (2001). Heme synthesis pathway. In CurrentProtocols in Toxicology (M. D. Maines, L. G. Costa, D. J. Reed, S. Sassa,and I. G. Sipes, Eds.), pp. 8.0.1–8.0.3. John Wiley & Sons, Inc., New York.
Simanainen, U., Tuomisto, J. T., Tuomisto, J., and Viluksela, M. (2002).Structure-activity relationships and dose responses of polychlorinateddibenzo-p-dioxins for short-term effects in 2,3,7,8-tetrachlorodibenzo-p-dioxin-resistant and -sensitive rat strains. Toxicol. Appl. Pharmacol. 181,38–47.
Smith, A. G., Carthew, P., Francis, J. E., Cabral, J. R., and Manson, M. M.(1993). Enhancement by iron of hepatic neoplasia in rats caused by hexa-chlorobenzene. Carcinogenesis 14, 1381–1387.
Smith, A. G., Clothier, B., Carthew, P., Childs, N. L., Sinclair, P. R., Nebert,D. W., and Dalton, T. P. (2001). Protection of the Cyp1a2(-/-) null mouseagainst uroporphyria and hepatic injury following exposure to 2,3,7,8-tetrachlorodibenzo-p-dioxin. Toxicol. Appl. Pharmacol. 173, 89–98.
Stocker, R., Yamamoto, Y., McDonagh, A. F., Glazer, A. N., and Ames, B. N.(1987). Bilirubin is an antioxidant of possible physiological importance.Science 235, 1043–1046.
Stonard, M. D., Poli, G., and De Matteis, F. (1998). Stimulation of liver hemeoxygenase in hexachlorobenzene-induced hepatic porphyria. Arch. Toxicol.72, 355–361.
Tuomisto, J. T., Viluksela, M., Pohjanvirta, R., and Tuomisto, J. (1999). TheAh receptor and a novel gene determine acute toxic responses to TCDD:Segregation of the resistant alleles to different rat lines. Toxicol. Appl.Pharmacol. 155, 71–81.
Unkila, M., Pohjanvirta, R., MacDonald, E., Tuomisto, J. T., and Tuomisto, J.(1994). Dose response and time course of alterations in tryptophan metab-olism by 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in the most TCDD-susceptible and the most TCDD-resistant rat strain: Relationship withTCDD lethality. Toxicol. Appl. Pharmacol. 128, 280–292.
Yang, K. H., Yoo, B. S., and Choe, S. Y. (1983). Effects of halogenateddibenzo-p-dioxins on plasma disappearance and biliary excretion of ouabainin rats. Toxicol. Lett. 15, 259–264.
Yokosuka, O., and Billing, B. (1987). Enzymatic oxidation of bilirubin byintestinal mucosa. Biochim. Biophys. Acta 923, 268–274
123TCDD AND BILIVERDIN ACCUMULATION IN RAT LIVER
by guest on April 5, 2016
http://toxsci.oxfordjournals.org/D
ownloaded from





























