Embed Size (px)
Citation preview

of July 16, 2022.This information is current as
Responses in Inflamed SkinBasophils Promote Innate Lymphoid Cell
and David ArtisNoti, Elia D. Tait Wojno, Thomas C. Fung, Masato KuboSaenz, Jonathan R. Brestoff, Laurel A. Monticelli, Mario Brian S. Kim, Kelvin Wang, Mark C. Siracusa, Steven A.
http://www.jimmunol.org/content/193/7/3717doi: 10.4049/jimmunol.1401307August 2014;
2014; 193:3717-3725; Prepublished online 25J Immunol
MaterialSupplementary
7.DCSupplementalhttp://www.jimmunol.org/content/suppl/2014/08/23/jimmunol.140130
Referenceshttp://www.jimmunol.org/content/193/7/3717.full#ref-list-1
, 17 of which you can access for free at: cites 50 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2014 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 16, 2022
http://ww
w.jim
munol.org/
Dow
nloaded from

The Journal of Immunology
Basophils Promote Innate Lymphoid Cell Responses inInflamed Skin
Brian S. Kim,*,†,‡,1 Kelvin Wang,*,†,‡ Mark C. Siracusa,*,†,2 Steven A. Saenz,*,†,3
Jonathan R. Brestoff,*,† Laurel A. Monticelli,*,† Mario Noti,*,†,4 Elia D. Tait Wojno,*,†
Thomas C. Fung,*,† Masato Kubo,x,{ and David Artis*,†,‖
Type 2 inflammation underlies allergic diseases such as atopic dermatitis, which is characterized by the accumulation of basophils
and group 2 innate lymphoid cells (ILC2s) in inflamed skin lesions. Although murine studies have demonstrated that cutaneous
basophil and ILC2 responses are dependent on thymic stromal lymphopoietin, whether these cell populations interact to regulate the
development of cutaneous type 2 inflammation is poorly defined. In this study, we identify that basophils and ILC2s significantly
accumulate in inflamed human and murine skin and form clusters not observed in control skin. We demonstrate that murine ba-
sophil responses precede ILC2 responses and that basophils are the dominant IL-4–enhanced GFP-expressing cell type in inflamed
skin. Furthermore, basophils and IL-4 were necessary for the optimal accumulation of ILC2s and induction of atopic dermatitis–
like disease. We show that ILC2s express IL-4Ra and proliferate in an IL-4–dependent manner. Additionally, basophil-derived IL-
4 was required for cutaneous ILC2 responses in vivo and directly regulated ILC2 proliferation ex vivo. Collectively, these data
reveal a previously unrecognized role for basophil-derived IL-4 in promoting ILC2 responses during cutaneous inflammation.
The Journal of Immunology, 2014, 193: 3717–3725.
Type 2 cytokine responses promote inflammation associ-ated with allergic rhinitis, food allergy, asthma, and atopicdermatitis (AD) that is characterized by IL-4, IL-5, and
IL-13 production, induction of IgE, and recruitment of gran-ulocytes (1–4). The epithelial cell–derived cytokine thymic stromallymphopoietin (TSLP) inititates type 2 immune responses viaeffects on multiple cellular targets, including CD4+ T cells, den-dritic cells (DCs), basophils, and group 2 innate lymphoid cells(ILC2s) (5–12). Polymorphisms in TSLP are associated with AD inhumans (13, 14), and TSLP expression is elevated in lesional skinand sera of AD patients (15, 16). In mice, TSLP/TSLP receptorinteractions promote the development of AD-like disease (17–20),supporting a role for TSLP in the pathogenesis of human andmurine skin inflammation.Recently, we demonstrated that murine basophils and ILC2s
accumulate in inflamed AD-like skin lesions in a TSLP-dependentmanner and contribute to type 2 cytokine-associated inflamma-
tion (9, 20). Basophils lack expression of cell lineage markersassociated with T and B cells, DCs, macrophages, and othergranulocytes, but they express FcεRIa and CD49b (21). Func-tionally, basophils express high levels of IL-4 in vivo and promotethe accumulation of other innate cells such as eosinophils in thecontext of chronic allergic dermatitis (9, 21, 22). ILC2s also lackexpression of lineage markers but can be identified by the ex-pression of CD25 and IL-33R (3). In contrast to basophils, whichpredominantly express IL-4, ILC2s express IL-5 and IL-13 (23–26). The differential effector cytokine expression profiles ofbasophils and ILC2s define their specialized functions in vivo (25),but whether functional interactions or cross-regulation occurs be-tween basophils and ILC2s remains unknown.In this study, we demonstrate that basophils and ILC2s accu-
mulate in close proximity to each other in the dermis of inflamedskin lesions isolated from AD patients and in AD-like murinelesions. Quantification of basophil–ILC2 clusters demonstrated
*Department of Microbiology, Perelman School of Medicine, University of Pennsyl-vania, Philadelphia, PA 19104; †Institute for Immunology, Perelman School of Medi-cine, University of Pennsylvania, Philadelphia, PA 19104; ‡Department of Dermatology,Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA 19104;xLaboratory for Cytokine Regulation, Research Center for Integrative Medical Science,RIKEN Yokohama Institute, Kanagawa 230-0045, Japan; {Division of Molecular Pa-thology, Research Institute for Biomedical Science, Tokyo University of Science, Chiba278-0022, Japan; and ‖Department of Pathobiology, School of Veterinary Medicine,University of Pennsylvania, Philadelphia, PA 19104
1Current address: Division of Dermatology, Department of Medicine, WashingtonUniversity School of Medicine, St. Louis, MO.
2Current address: Department of Medicine, Center for Immunity and Inflammation,Rutgers New Jersey Medical School, Newark, NJ.
3Current address: Immunology Research, Biogen Idec, Inc., Cambridge, MA.
4Current address: Department of Experimental Pathology, Institute of Pathology,University of Bern, Bern, Switzerland.
Received for publication May 21, 2014. Accepted for publication July 29, 2014.
This work was supported by National Institutes of Health (NIH) Grants AI061570,AI087990, AI074878, AI106697, AI095466, AI095608, AI102942, and AI097333 (toD.A.); NIH Grants T32-AR007465-28, 5-P30-AR-057217, KL2-RR024132, and K08-AR065577-01 (to B.S.K.); NIH Grants AI085828 and K22-AI110573 (to M.C.S.); NIH
Grants T-32A106516 and F30-AI112023 (to J.R.B.); NIH Grant T32-AI007532 (toL.A.M.); and NIH Grant F32-AI098365 (to E.D.T.W.). B.S.K. and K.W. were supportedby Mary Connolly Braun, the Arguild Foundation, and the Laffey-McHugh Foundation.K.W. was supported by a Benjamin Franklin Society research grant. T.C.F. was sup-ported by a Cancer Research Institute Student Training and Research in Tumor Immu-nology grant. M.N. was supported by a Swiss National Science Foundation prospectiveresearch fellowship (PA00P3-136468). This work was also supported by the BurroughsWellcome Fund, the National Center for Research Resources, the National Center forAdvancing Translational Sciences, NIH Grant KL2TR000139, and the Skin DiseaseResearch Center through NIH Grant SDRC 5-P30-AR-057217.
Address correspondence and reprint requests to Dr. David Artis, Perelman School ofMedicine, Department of Microbiology, Institute for Immunology, University ofPennsylvania, 421 Curie Boulevard, BRB II/III, Room 356, Philadelphia PA19104-6160. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this article: 7-AAD, 7-aminoactinomycin D; AD, atopic der-matitis; BM, bone marrow; DC, dendritic cell; eGFP, enhanced GFP; HPF, high-power field; i.d., intradermal(ly); IF, immunofluorescence; ILC2, group 2 innatelymphoid cell; KLRG1, killer-cell lectin like receptor G1; Lin, lineage; rm, recombi-nant murine; TSLP, thymic stromal lymphopoietin; WT, wild-type.
Copyright� 2014 by TheAmerican Association of Immunologists, Inc. 0022-1767/14/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1401307
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

a significant accumulation of these clusters in AD-associated skinin comparison with healthy control skin. Temporal analysesrevealed that the accumulation of basophils in murine skin pre-cedes that of ILC2s in the context of AD-like inflammation.Furthermore, loss- and gain-of-function studies demonstrated thatbasophils are required to promote cutaneous ILC2 responsesin vivo. Strikingly, employing IL-4–enhanced GFP (eGFP) (4get)reporter mice, we found that basophils were the dominant sourceof IL-4–eGFP expression in AD-like skin. Both basophils and IL-4 were necessary for the optimal accumulation of ILC2s andinduction of AD-like disease. Further examination of ILC2srevealed cell surface expression of IL-4Ra and their dependenceon IL-4 for proliferation in AD-like inflammation. Basophil-derived IL-4 was required for cutaneous ILC2 responses in vivoand directly regulated ILC2 proliferation ex vivo. Collectively,these studies identify that basophils are early regulators of ILC2responses in the context of cutaneous type 2 inflammation andAD-like disease.
Materials and MethodsPatients and samples
Human skin samples were assessed as previously described for flowcytometry, and basophils were defined as CD123+FcεRIa+ cells negativefor expression of CD3, CD11c, CD19, CD56, and c-Kit, whereas ILC2swere defined as CD25+IL-33R+ cells negative for expression of lineage(CD3, TCRab, CD5, CD11b, CD11c, CD19, CD56, and FcεRIa)markers (10, 20). Experiments using human skin samples were approvedby the University of Pennsylvania Institutional Review Board (protocol814945).
Mice and samples
C57BL/6 wild-type (WT), BALB/c WT, 4get, Rag12/2, and Il42/2 micewere purchased from The Jackson Laboratory. Il4ra2/2 mice werepurchased from Taconic. BaS-TRECK (BaS) mice were provided byDr. M. Kubo (Tokyo). All mice were treated with MC903 as previouslydescribed (9, 20, 27). Murine skin samples were assessed as previouslydescribed for flow cytometry, and basophils were defined as CD49b+
FcεRIa+ cells negative for expression of CD3, CD5, CD11c, CD19,NK1.1, and c-Kit, whereas ILC2s were defined as CD25+IL-33R+ cellsnegative for expression of lineage (CD3, CD5, CD11b, CD11c, B220,NK1.1, and FcεRIa) markers (10, 20). Splenic basophils were sort-purifiedfrom TSLP cDNA plasmid-treated WT or Il42/2 mice using a BD FAC-SAria cell sorter 3 wk after TSLP cDNA plasmid injection, as previouslydescribed (9). TSLP cDNA plasmid was provided by M.R. Comeau(Amgen). WT and BaS mice were treated with diphtheria toxin as previ-ously described (9). Basophils (10,000 cells) were suspended in 50 ml PBSand injected intradermally (i.d.) into naive WT mice. WT mice weretreated with 300 ng recombinant murine (rm)IL-33 (R&D Systems) dailyin 200 ml PBS i.p. for 7 d prior to sort-purification of ILC2s on a BDFACSAria cell sorter. ILC2s were sort-purified from pooled skin-draininglymph nodes, mesenteric lymph nodes, peritoneal cavity, and adiposetissue as previously described (20, 28). Annexin V, 7-aminoactinomycin D(7-AAD), killer-cell lectin-like receptor G1 (KLRG1), CD45, and Ki67staining of ILC2s was performed as previously described (29–31).Experiments were performed according to the guidelines of the Universityof Pennsylvania Institutional Animal Care and Use Committee.
Histology
For all human and murine immunofluorescence (IF) microscopy, paraffin-embedded 5-mm skin sections were rehydrated and incubated with pri-mary Ab at 4˚C overnight, followed by incubation with secondary Abs at37˚C for 30 min. For human samples, primary Abs against 2D7 (1:250,BioLegend, Ab mouse IgG1), IL-33R (1:250, MD Bioproducts, biotin-conjugated mouse IgG1), or CD3 (1:50, Dako, rabbit IgG) and second-ary Abs to mouse IgG conjugated with PE-Cy5 (1:300), streptavidin-Cy3(1:300), or rabbit IgG conjugated with Cy2 (1:300), respectively, wereused. Murine samples were stained with H&E or primary Abs againstMcpt8 (1:500, TUG8, BioLegend, rat IgG2) or IL-33R (1:300, DJ8, MDBioproducts, biotin-conjugated rat IgG1) and secondary Abs to rat IgGconjugated with PE-Cy5 (1:300) and streptavidin-Cy3 (1:300), respec-tively. Sections were placed in mounting medium with DAPI (Vectashield).
Histology score and ear thickness measurements
Histology score was determined by the following formula: (total numberof lymphocytes per high-power field [HPF] + thickness of the epidermismeasured in micrometers from the basement membrane to the top of thestratum corneum)/100. The ear thickness was measured on day 7 followingtreatment by measuring the entire width of the ear at the widest point inmicrometers from the top of one stratum corneum to the top of the stratumcorneum on the opposite side of the ear. All measurements were performedusing the National Institutes of Health ImageJ program.
In vitro/ex vivo cultures
Sort-purified basophils from the spleens of TSLP cDNA plasmid-treatedWT and Il42/2 mice were cultured for 24 h at 37˚C in supplementedDMEM with rmIL-3 (10 ng/ml, R&D Systems) as previously described(9). CFSE-labeled ILC2s (20,000 cells) from rmIL-33–treated mice werethen cultured in 200 ml of WT and Il42/2 basophil-conditioned media for4 d at 37˚C with rmIL-7 (R&D Systems) as previously described (20), andcell division was assessed by FACS analysis of CFSE dye dilution aspreviously described (32).
Basophil–ILC2, basophil–T cell, and ILC2–T cell clusteranalysis
Basophil–ILC2 (yellow circle), basophil–T cell (green circles), and ILC2–T cell (red circle) clusters were analyzed by drawing 25-mm colored cir-cles around the cell/cell clusters per HPF.
Statistical analysis
Data are means 6 SEM unless indicated otherwise. Statistical significancewas determined by an unpaired Student t test or one-way ANOVA formultiple comparisons using GraphPad Prism 6 software. The p values aredenoted for each analysis.
ResultsBasophils and ILC2s are enriched and found in close proximityto each other in the dermis of human AD lesions
We recently reported that murine basophils and ILC2s are elicitedin a TSLP-dependent manner during AD-like inflammation (9, 20).Consistent with this, basophils have been identified in skin lesionsfrom AD patients, and we recently showed that ILC2s are enrichedin the inflamed skin of patients (20, 33). However, whether baso-phils and ILC2s accumulate in the same lesions and whether thesecell types colocalize in inflamed human skin have not been exam-ined. To address these issues, 4-mm punch biopsies of normal anddiseased human skin were obtained from healthy control subjectsand AD patients. Strikingly, frequencies of CD123+FcεRIa+ baso-phils (Fig. 1A) and lineage (Lin)2CD25+IL-33R+ ILC2s (Fig. 1B)were markedly elevated in lesional human AD skin, indicating thatbasophil and ILC2 responses occur simultaneously in the sameinflamed lesion.To visualize the distribution of cutaneous basophils and ILC2s,
IF microscopy was employed. Basophils were identified by stainingwith an Ab against the basophil-specific granule marker 2D7 (34).Consistent with the flow cytometric analyses (Fig. 1A), 2D7+
basophils were undetectable in healthy control skin (SupplementalFig. 1A) but were readily identified in lesional AD skin (Fig. 1C,Supplemental Fig. 1B). To visualize ILC2s and identify non–T cellpopulations that express the IL-33R, sections were costained withanti-CD3 Ab and anti–IL-33R Ab. Although IL-33R+ cells andCD3+ T cells were rarely identified in healthy control skin(Supplemental Fig. 1C), both were enriched in lesional AD skinand accumulated in a perivascular distribution in the dermis (Fig.1D, Supplemental Fig. 1D). Most IL-33R+ cells were negative forCD3 (Fig. 1D, white arrows), suggesting that these IL-33R+ cellsare ILC2s. Finally, 2D7+ basophils and IL-33R+ ILC2s accumu-lated in close proximity to each other in the papillary dermis inlesional human AD skin compared with healthy control skin (Fig.1E–G, Supplemental Fig. 1E, 1F). Although our analyses demon-
3718 INNATE IMMUNE REGULATION OF SKIN INFLAMMATION
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
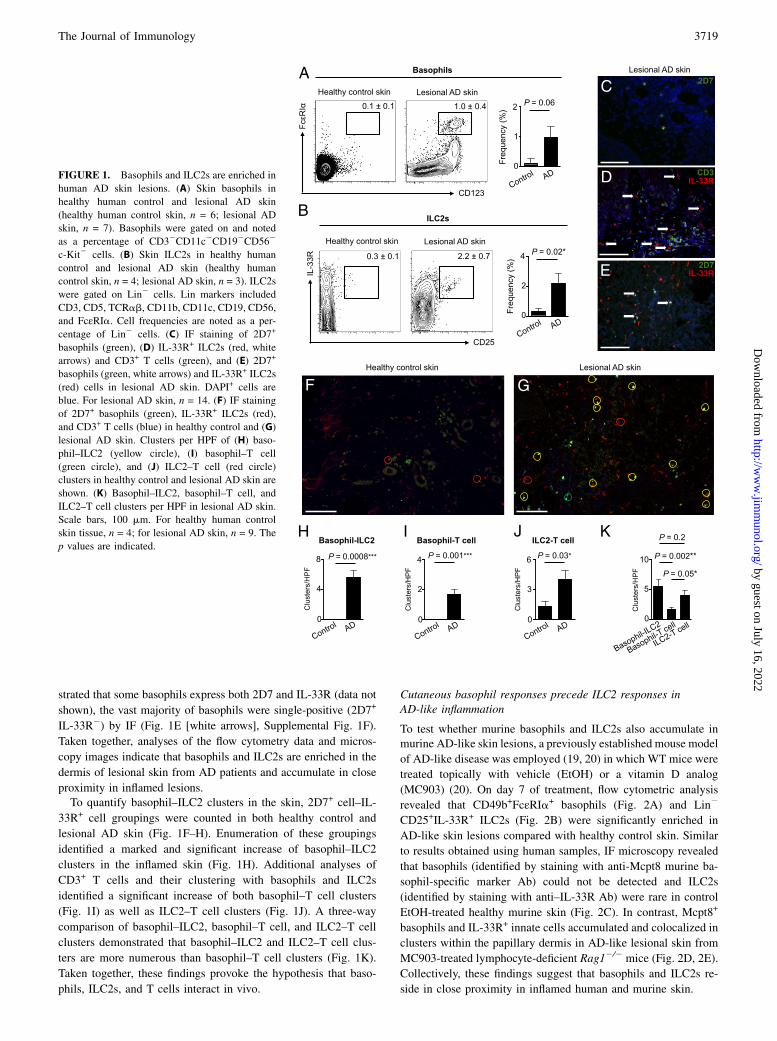
strated that some basophils express both 2D7 and IL-33R (data not
shown), the vast majority of basophils were single-positive (2D7+
IL-33R2) by IF (Fig. 1E [white arrows], Supplemental Fig. 1F).
Taken together, analyses of the flow cytometry data and micros-
copy images indicate that basophils and ILC2s are enriched in the
dermis of lesional skin from AD patients and accumulate in close
proximity in inflamed lesions.To quantify basophil–ILC2 clusters in the skin, 2D7+ cell–IL-
33R+ cell groupings were counted in both healthy control and
lesional AD skin (Fig. 1F–H). Enumeration of these groupings
identified a marked and significant increase of basophil–ILC2
clusters in the inflamed skin (Fig. 1H). Additional analyses of
CD3+ T cells and their clustering with basophils and ILC2s
identified a significant increase of both basophil–T cell clusters
(Fig. 1I) as well as ILC2–T cell clusters (Fig. 1J). A three-way
comparison of basophil–ILC2, basophil–T cell, and ILC2–T cell
clusters demonstrated that basophil–ILC2 and ILC2–T cell clus-
ters are more numerous than basophil–T cell clusters (Fig. 1K).
Taken together, these findings provoke the hypothesis that baso-
phils, ILC2s, and T cells interact in vivo.
Cutaneous basophil responses precede ILC2 responses inAD-like inflammation
To test whether murine basophils and ILC2s also accumulate inmurine AD-like skin lesions, a previously established mouse model
of AD-like disease was employed (19, 20) in which WT mice were
treated topically with vehicle (EtOH) or a vitamin D analog
(MC903) (20). On day 7 of treatment, flow cytometric analysis
revealed that CD49b+FcεRIa+ basophils (Fig. 2A) and Lin2
CD25+IL-33R+ ILC2s (Fig. 2B) were significantly enriched in
AD-like skin lesions compared with healthy control skin. Similar
to results obtained using human samples, IF microscopy revealed
that basophils (identified by staining with anti-Mcpt8 murine ba-
sophil-specific marker Ab) could not be detected and ILC2s
(identified by staining with anti–IL-33R Ab) were rare in control
EtOH-treated healthy murine skin (Fig. 2C). In contrast, Mcpt8+
basophils and IL-33R+ innate cells accumulated and colocalized in
clusters within the papillary dermis in AD-like lesional skin from
MC903-treated lymphocyte-deficient Rag12/2 mice (Fig. 2D, 2E).
Collectively, these findings suggest that basophils and ILC2s re-
side in close proximity in inflamed human and murine skin.
FIGURE 1. Basophils and ILC2s are enriched in
human AD skin lesions. (A) Skin basophils in
healthy human control and lesional AD skin
(healthy human control skin, n = 6; lesional AD
skin, n = 7). Basophils were gated on and noted
as a percentage of CD32CD11c2CD192CD562
c-Kit2 cells. (B) Skin ILC2s in healthy human
control and lesional AD skin (healthy human
control skin, n = 4; lesional AD skin, n = 3). ILC2s
were gated on Lin2 cells. Lin markers included
CD3, CD5, TCRab, CD11b, CD11c, CD19, CD56,
and FcεRIa. Cell frequencies are noted as a per-
centage of Lin2 cells. (C) IF staining of 2D7+
basophils (green), (D) IL-33R+ ILC2s (red, white
arrows) and CD3+ T cells (green), and (E) 2D7+
basophils (green, white arrows) and IL-33R+ ILC2s
(red) cells in lesional AD skin. DAPI+ cells are
blue. For lesional AD skin, n = 14. (F) IF staining
of 2D7+ basophils (green), IL-33R+ ILC2s (red),
and CD3+ T cells (blue) in healthy control and (G)
lesional AD skin. Clusters per HPF of (H) baso-
phil–ILC2 (yellow circle), (I) basophil–T cell
(green circle), and (J) ILC2–T cell (red circle)
clusters in healthy control and lesional AD skin are
shown. (K) Basophil–ILC2, basophil–T cell, and
ILC2–T cell clusters per HPF in lesional AD skin.
Scale bars, 100 mm. For healthy human control
skin tissue, n = 4; for lesional AD skin, n = 9. The
p values are indicated.
The Journal of Immunology 3719
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
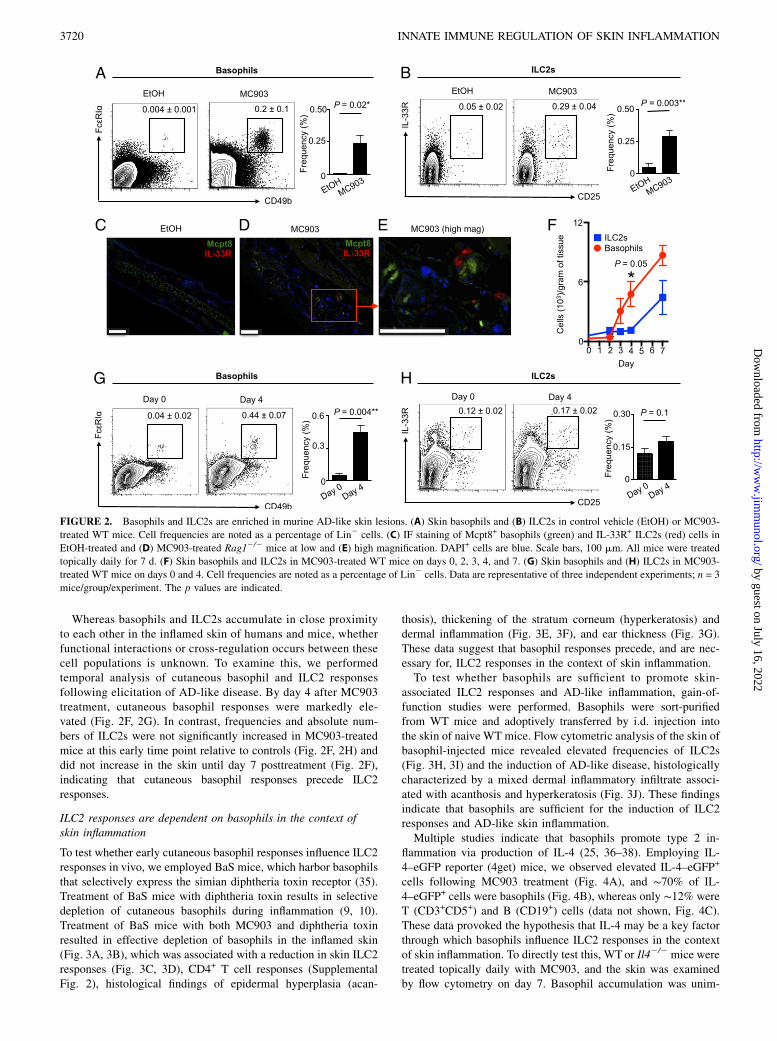
Whereas basophils and ILC2s accumulate in close proximityto each other in the inflamed skin of humans and mice, whetherfunctional interactions or cross-regulation occurs between thesecell populations is unknown. To examine this, we performedtemporal analysis of cutaneous basophil and ILC2 responsesfollowing elicitation of AD-like disease. By day 4 after MC903treatment, cutaneous basophil responses were markedly ele-vated (Fig. 2F, 2G). In contrast, frequencies and absolute num-bers of ILC2s were not significantly increased in MC903-treatedmice at this early time point relative to controls (Fig. 2F, 2H) anddid not increase in the skin until day 7 posttreatment (Fig. 2F),indicating that cutaneous basophil responses precede ILC2responses.
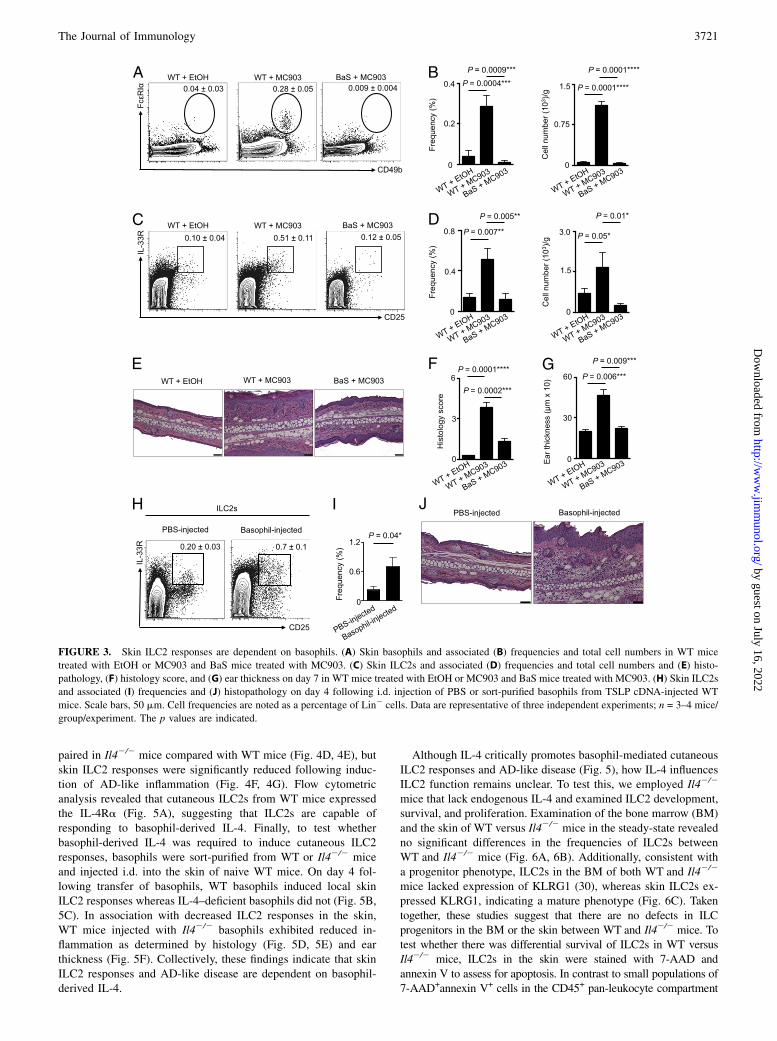
ILC2 responses are dependent on basophils in the context ofskin inflammation
To test whether early cutaneous basophil responses influence ILC2responses in vivo, we employed BaS mice, which harbor basophilsthat selectively express the simian diphtheria toxin receptor (35).Treatment of BaS mice with diphtheria toxin results in selectivedepletion of cutaneous basophils during inflammation (9, 10).Treatment of BaS mice with both MC903 and diphtheria toxinresulted in effective depletion of basophils in the inflamed skin(Fig. 3A, 3B), which was associated with a reduction in skin ILC2responses (Fig. 3C, 3D), CD4+ T cell responses (SupplementalFig. 2), histological findings of epidermal hyperplasia (acan-
thosis), thickening of the stratum corneum (hyperkeratosis) anddermal inflammation (Fig. 3E, 3F), and ear thickness (Fig. 3G).These data suggest that basophil responses precede, and are nec-essary for, ILC2 responses in the context of skin inflammation.To test whether basophils are sufficient to promote skin-
associated ILC2 responses and AD-like inflammation, gain-of-function studies were performed. Basophils were sort-purifiedfrom WT mice and adoptively transferred by i.d. injection intothe skin of naive WT mice. Flow cytometric analysis of the skin ofbasophil-injected mice revealed elevated frequencies of ILC2s(Fig. 3H, 3I) and the induction of AD-like disease, histologicallycharacterized by a mixed dermal inflammatory infiltrate associ-ated with acanthosis and hyperkeratosis (Fig. 3J). These findingsindicate that basophils are sufficient for the induction of ILC2responses and AD-like skin inflammation.Multiple studies indicate that basophils promote type 2 in-
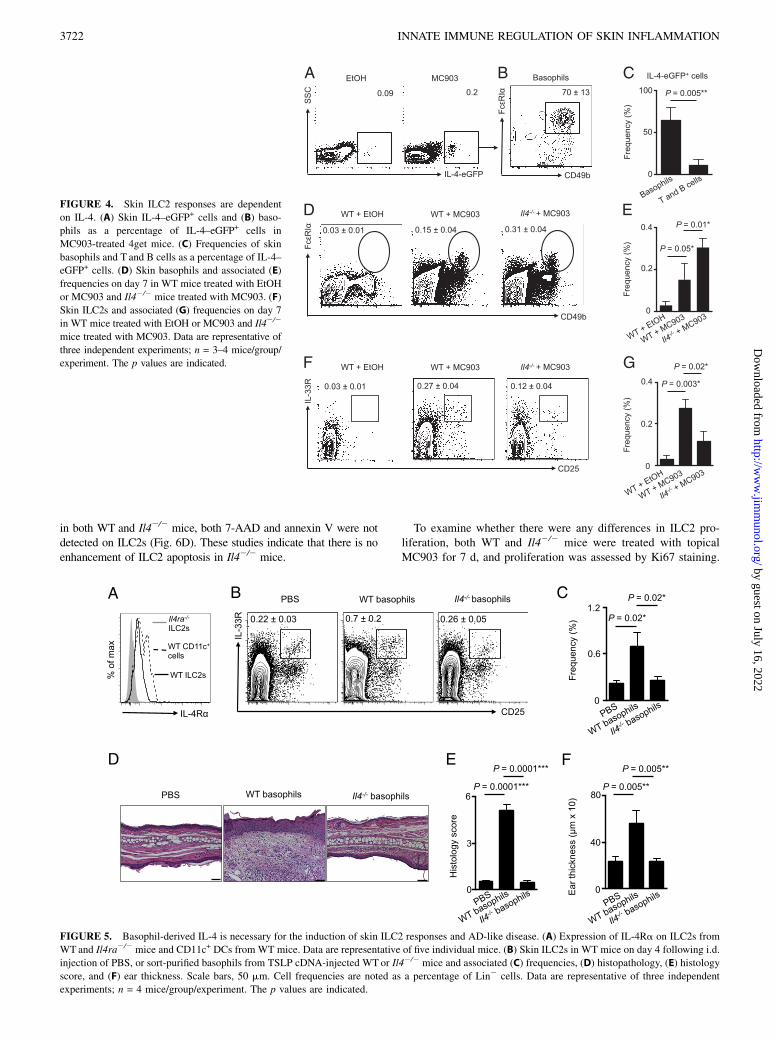
flammation via production of IL-4 (25, 36–38). Employing IL-4–eGFP reporter (4get) mice, we observed elevated IL-4–eGFP+
cells following MC903 treatment (Fig. 4A), and ∼70% of IL-4–eGFP+ cells were basophils (Fig. 4B), whereas only ∼12% wereT (CD3+CD5+) and B (CD19+) cells (data not shown, Fig. 4C).These data provoked the hypothesis that IL-4 may be a key factorthrough which basophils influence ILC2 responses in the contextof skin inflammation. To directly test this, WTor Il42/2 mice weretreated topically daily with MC903, and the skin was examinedby flow cytometry on day 7. Basophil accumulation was unim-
FIGURE 2. Basophils and ILC2s are enriched in murine AD-like skin lesions. (A) Skin basophils and (B) ILC2s in control vehicle (EtOH) or MC903-
treated WT mice. Cell frequencies are noted as a percentage of Lin2 cells. (C) IF staining of Mcpt8+ basophils (green) and IL-33R+ ILC2s (red) cells in
EtOH-treated and (D) MC903-treated Rag12/2 mice at low and (E) high magnification. DAPI+ cells are blue. Scale bars, 100 mm. All mice were treated
topically daily for 7 d. (F) Skin basophils and ILC2s in MC903-treated WT mice on days 0, 2, 3, 4, and 7. (G) Skin basophils and (H) ILC2s in MC903-
treated WT mice on days 0 and 4. Cell frequencies are noted as a percentage of Lin2 cells. Data are representative of three independent experiments; n = 3
mice/group/experiment. The p values are indicated.
3720 INNATE IMMUNE REGULATION OF SKIN INFLAMMATION
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
paired in Il42/2 mice compared with WT mice (Fig. 4D, 4E), butskin ILC2 responses were significantly reduced following induc-tion of AD-like inflammation (Fig. 4F, 4G). Flow cytometricanalysis revealed that cutaneous ILC2s from WT mice expressedthe IL-4Ra (Fig. 5A), suggesting that ILC2s are capable ofresponding to basophil-derived IL-4. Finally, to test whetherbasophil-derived IL-4 was required to induce cutaneous ILC2responses, basophils were sort-purified from WT or Il42/2 miceand injected i.d. into the skin of naive WT mice. On day 4 fol-lowing transfer of basophils, WT basophils induced local skinILC2 responses whereas IL-4–deficient basophils did not (Fig. 5B,5C). In association with decreased ILC2 responses in the skin,WT mice injected with Il42/2 basophils exhibited reduced in-flammation as determined by histology (Fig. 5D, 5E) and earthickness (Fig. 5F). Collectively, these findings indicate that skinILC2 responses and AD-like disease are dependent on basophil-derived IL-4.
Although IL-4 critically promotes basophil-mediated cutaneousILC2 responses and AD-like disease (Fig. 5), how IL-4 influencesILC2 function remains unclear. To test this, we employed Il42/2
mice that lack endogenous IL-4 and examined ILC2 development,survival, and proliferation. Examination of the bone marrow (BM)and the skin of WT versus Il42/2 mice in the steady-state revealedno significant differences in the frequencies of ILC2s betweenWT and Il42/2 mice (Fig. 6A, 6B). Additionally, consistent witha progenitor phenotype, ILC2s in the BM of both WT and Il42/2
mice lacked expression of KLRG1 (30), whereas skin ILC2s ex-pressed KLRG1, indicating a mature phenotype (Fig. 6C). Takentogether, these studies suggest that there are no defects in ILCprogenitors in the BM or the skin between WT and Il42/2 mice. Totest whether there was differential survival of ILC2s in WT versusIl42/2 mice, ILC2s in the skin were stained with 7-AAD andannexin V to assess for apoptosis. In contrast to small populations of7-AAD+annexin V+ cells in the CD45+ pan-leukocyte compartment
FIGURE 3. Skin ILC2 responses are dependent on basophils. (A) Skin basophils and associated (B) frequencies and total cell numbers in WT mice
treated with EtOH or MC903 and BaS mice treated with MC903. (C) Skin ILC2s and associated (D) frequencies and total cell numbers and (E) histo-
pathology, (F) histology score, and (G) ear thickness on day 7 in WT mice treated with EtOH or MC903 and BaS mice treated with MC903. (H) Skin ILC2s
and associated (I) frequencies and (J) histopathology on day 4 following i.d. injection of PBS or sort-purified basophils from TSLP cDNA-injected WT
mice. Scale bars, 50 mm. Cell frequencies are noted as a percentage of Lin2 cells. Data are representative of three independent experiments; n = 3–4 mice/
group/experiment. The p values are indicated.
The Journal of Immunology 3721
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
in both WT and Il42/2 mice, both 7-AAD and annexin V were notdetected on ILC2s (Fig. 6D). These studies indicate that there is noenhancement of ILC2 apoptosis in Il42/2 mice.
To examine whether there were any differences in ILC2 pro-liferation, both WT and Il42/2 mice were treated with topicalMC903 for 7 d, and proliferation was assessed by Ki67 staining.
FIGURE 4. Skin ILC2 responses are dependent
on IL-4. (A) Skin IL-4–eGFP+ cells and (B) baso-
phils as a percentage of IL-4–eGFP+ cells in
MC903-treated 4get mice. (C) Frequencies of skin
basophils and T and B cells as a percentage of IL-4–
eGFP+ cells. (D) Skin basophils and associated (E)
frequencies on day 7 in WT mice treated with EtOH
or MC903 and Il42/2 mice treated with MC903. (F)
Skin ILC2s and associated (G) frequencies on day 7
in WT mice treated with EtOH or MC903 and Il42/2
mice treated with MC903. Data are representative of
three independent experiments; n = 3–4 mice/group/
experiment. The p values are indicated.
FIGURE 5. Basophil-derived IL-4 is necessary for the induction of skin ILC2 responses and AD-like disease. (A) Expression of IL-4Ra on ILC2s from
WTand Il4ra2/2 mice and CD11c+ DCs from WT mice. Data are representative of five individual mice. (B) Skin ILC2s in WT mice on day 4 following i.d.
injection of PBS, or sort-purified basophils from TSLP cDNA-injected WT or Il42/2 mice and associated (C) frequencies, (D) histopathology, (E) histology
score, and (F) ear thickness. Scale bars, 50 mm. Cell frequencies are noted as a percentage of Lin2 cells. Data are representative of three independent
experiments; n = 4 mice/group/experiment. The p values are indicated.
3722 INNATE IMMUNE REGULATION OF SKIN INFLAMMATION
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
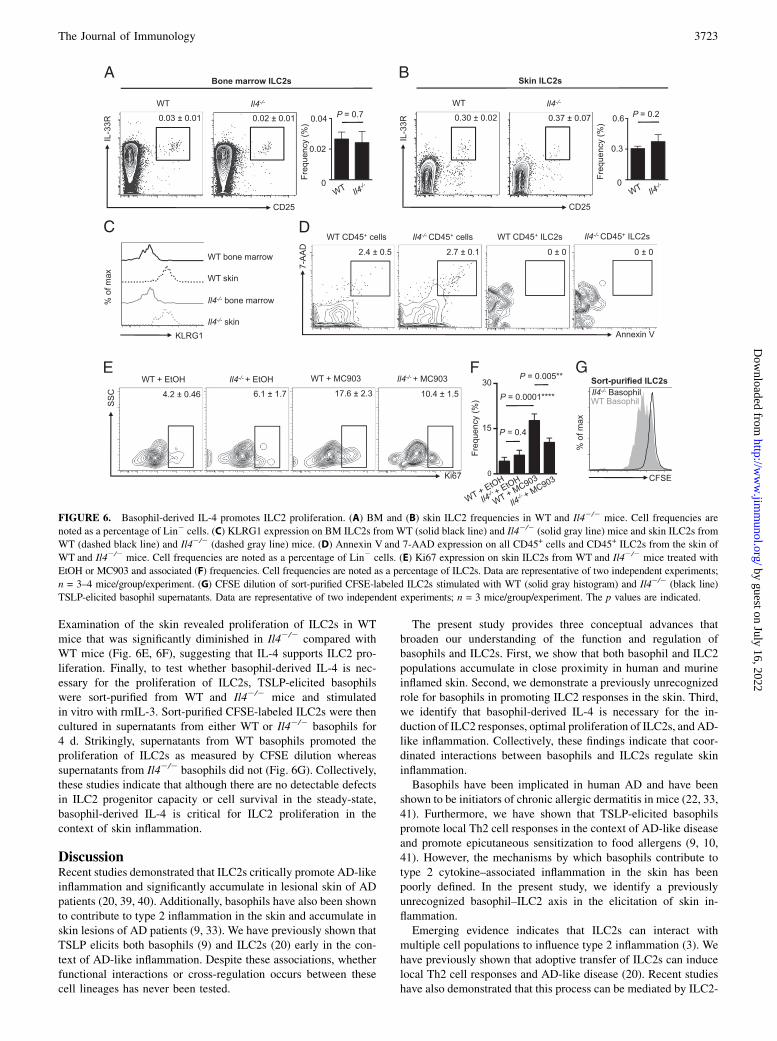
Examination of the skin revealed proliferation of ILC2s in WTmice that was significantly diminished in Il42/2 compared withWT mice (Fig. 6E, 6F), suggesting that IL-4 supports ILC2 pro-liferation. Finally, to test whether basophil-derived IL-4 is nec-essary for the proliferation of ILC2s, TSLP-elicited basophilswere sort-purified from WT and Il42/2 mice and stimulatedin vitro with rmIL-3. Sort-purified CFSE-labeled ILC2s were thencultured in supernatants from either WT or Il42/2 basophils for4 d. Strikingly, supernatants from WT basophils promoted theproliferation of ILC2s as measured by CFSE dilution whereassupernatants from Il42/2 basophils did not (Fig. 6G). Collectively,these studies indicate that although there are no detectable defectsin ILC2 progenitor capacity or cell survival in the steady-state,basophil-derived IL-4 is critical for ILC2 proliferation in thecontext of skin inflammation.
DiscussionRecent studies demonstrated that ILC2s critically promote AD-likeinflammation and significantly accumulate in lesional skin of ADpatients (20, 39, 40). Additionally, basophils have also been shownto contribute to type 2 inflammation in the skin and accumulate inskin lesions of AD patients (9, 33). We have previously shown thatTSLP elicits both basophils (9) and ILC2s (20) early in the con-text of AD-like inflammation. Despite these associations, whetherfunctional interactions or cross-regulation occurs between thesecell lineages has never been tested.
The present study provides three conceptual advances thatbroaden our understanding of the function and regulation ofbasophils and ILC2s. First, we show that both basophil and ILC2populations accumulate in close proximity in human and murineinflamed skin. Second, we demonstrate a previously unrecognizedrole for basophils in promoting ILC2 responses in the skin. Third,we identify that basophil-derived IL-4 is necessary for the in-duction of ILC2 responses, optimal proliferation of ILC2s, and AD-like inflammation. Collectively, these findings indicate that coor-dinated interactions between basophils and ILC2s regulate skininflammation.Basophils have been implicated in human AD and have been
shown to be initiators of chronic allergic dermatitis in mice (22, 33,41). Furthermore, we have shown that TSLP-elicited basophilspromote local Th2 cell responses in the context of AD-like diseaseand promote epicutaneous sensitization to food allergens (9, 10,41). However, the mechanisms by which basophils contribute totype 2 cytokine–associated inflammation in the skin has beenpoorly defined. In the present study, we identify a previouslyunrecognized basophil–ILC2 axis in the elicitation of skin in-flammation.Emerging evidence indicates that ILC2s can interact with
multiple cell populations to influence type 2 inflammation (3). Wehave previously shown that adoptive transfer of ILC2s can inducelocal Th2 cell responses and AD-like disease (20). Recent studieshave also demonstrated that this process can be mediated by ILC2-
FIGURE 6. Basophil-derived IL-4 promotes ILC2 proliferation. (A) BM and (B) skin ILC2 frequencies in WT and Il42/2 mice. Cell frequencies are
noted as a percentage of Lin2 cells. (C) KLRG1 expression on BM ILC2s from WT (solid black line) and Il42/2 (solid gray line) mice and skin ILC2s from
WT (dashed black line) and Il42/2 (dashed gray line) mice. (D) Annexin V and 7-AAD expression on all CD45+ cells and CD45+ ILC2s from the skin of
WT and Il42/2 mice. Cell frequencies are noted as a percentage of Lin2 cells. (E) Ki67 expression on skin ILC2s from WT and Il42/2 mice treated with
EtOH or MC903 and associated (F) frequencies. Cell frequencies are noted as a percentage of ILC2s. Data are representative of two independent experiments;
n = 3–4 mice/group/experiment. (G) CFSE dilution of sort-purified CFSE-labeled ILC2s stimulated with WT (solid gray histogram) and Il42/2 (black line)
TSLP-elicited basophil supernatants. Data are representative of two independent experiments; n = 3 mice/group/experiment. The p values are indicated.
The Journal of Immunology 3723
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

derived IL-13 influencing DC migration and/or contact-dependentstimulation of T cells by MHC class II or other factors (32, 42).Taken together, these studies highlight a role for ILC2s in pro-moting innate and adaptive immune responses. Furthermore,Roediger et al. (43) employed two-photon intravital microscopy inthe skin to show that ILC2s suppress the function of other gran-ulocytes such as mast cells in vivo. However, whether other up-stream innate cellular mechanisms influence ILC2 function in thecontext of AD was not known. We now show that basophils candirectly influence ILC2 function through the production of IL-4.Therefore, although ILC2s can be activated directly by epithelialcell–derived cytokines (3), it appears that hematopoietic cell–de-rived cytokines can also influence ILC2 function in the context ofinflammation.It has been appreciated that IL-4 is highly expressed in lesional
AD skin and contributes to the pathogenesis of AD (13, 44, 45).Furthermore, other common g-chain cytokines, such as IL-2 andIL-7, have shown to be critical for the development and survivalof ILCs (43, 46). However, we found that IL-4 was dispensablefor these processes but essential to promote ILC2 proliferationboth in vitro and in vivo. This is consistent with the role of IL-4 insupporting the growth and proliferation of multiple other celltypes such as T cells and B cells (47). However, our studies donot preclude other mechanisms by which basophils and IL-4 candirectly promote ILC2 function. For example, basophil-derivedIL-4 may influence the responsiveness of ILC2s to epithelial cell–derived cytokines such as IL-25, IL-33, and TSLP (3), or influ-ence the activation and/or recruitment of ILC2s into inflamedskin. Motomura et al. (48) recently demonstrated a role forbasophils in regulating ILC2 activation in the context of protease-induced airway inflammation. These findings, coupled with thepresent study, suggest that the capacity of basophils to promoteILC2 responses may be a conserved pathway at multiple barriersurfaces.In addition to IL-4, basophils produce a variety of factors such
as proinflammatory eicosanoids that can influence the functionof other immune cells that promote type 2 inflammation (21).Therefore, other endogenous factors derived from basophils mayact in concert with IL-4 to modulate ILC2 function. In support ofthis, recent studies have shown that both murine and human ILC2sdirectly respond to leukotrienes (49, 50). Moreover, both IL-33and TSLP are known to differentially regulate the production ofcytokines and chemokines from basophils (9). However, how thedifferential regulation of basophils influences ILC2 function andtype 2 inflammation remains to be fully defined. Notwithstandingthis, the present study demonstrates that TSLP-elicited basophilsand IL-4 production promote cutaneous ILC2 responses andproliferation, suggesting that selective targeting of TSLP andTSLP-dependent basophils could prove to be a beneficial strategyin the treatment of AD and related allergic diseases.
AcknowledgmentsWe thank members of the Artis laboratory for discussions and reading of the
manuscript, M.R. Comeau (Amgen) for providing TSLP cDNA plasmid,
C.J. Miller, J. Sobanko, X. Zhu, T. Nunnciato, N. Manogue, and A.S. Payne
for providing healthy human control samples through the Skin Disease Re-
search Center Core B, A.S. Van Voorhees and W.D. James for clinical sup-
port, and J. Molleston and S. Cherry for microscopy resources. We also
thank Mary Connolly Braun, the Arguild Foundation, and the Laffey-
McHugh Foundation for support of these studies.
DisclosuresThe authors have no financial conflicts of interest.
References1. Paul, W. E., and J. Zhu. 2010. How are TH2-type immune responses initiated and
amplified? Nat. Rev. Immunol. 10: 225–235.2. Kaplan, M. H. 2013. Th9 cells: differentiation and disease. Immunol. Rev. 252:
104–115.3. Kim, B. S., E. D. Wojno, and D. Artis. 2013. Innate lymphoid cells and allergic
inflammation. Curr. Opin. Immunol. 25: 738–744.4. Licona-Limon, P., L. K. Kim, N. W. Palm, and R. A. Flavell. 2013. TH2, allergy
and group 2 innate lymphoid cells. Nat. Immunol. 14: 536–542.5. Watanabe, N., S. Hanabuchi, V. Soumelis, W. Yuan, S. Ho, R. de Waal Malefyt,
and Y. J. Liu. 2004. Human thymic stromal lymphopoietin promotes dendriticcell-mediated CD4+ T cell homeostatic expansion. Nat. Immunol. 5: 426–434.
6. Ito, T., Y. H. Wang, O. Duramad, T. Hori, G. J. Delespesse, N. Watanabe,F. X. Qin, Z. Yao, W. Cao, and Y. J. Liu. 2005. TSLP-activated dendritic cellsinduce an inflammatory T helper type 2 cell response through OX40 ligand. J.Exp. Med. 202: 1213–1223.
7. Wang, Y. H., T. Ito, Y. H. Wang, B. Homey, N. Watanabe, R. Martin,C. J. Barnes, B. W. McIntyre, M. Gilliet, R. Kumar, et al. 2006. Maintenance andpolarization of human TH2 central memory T cells by thymic stromallymphopoietin-activated dendritic cells. Immunity 24: 827–838.
8. Omori, M., and S. Ziegler. 2007. Induction of IL-4 expression in CD4+ T cells bythymic stromal lymphopoietin. J. Immunol. 178: 1396–1404.
9. Siracusa, M. C., S. A. Saenz, D. A. Hill, B. S. Kim, M. B. Headley,T. A. Doering, E. J. Wherry, H. K. Jessup, L. A. Siegel, T. Kambayashi, et al.2011. TSLP promotes interleukin-3-independent basophil haematopoiesis andtype 2 inflammation. Nature 477: 229–233.
10. Noti, M., E. D. Wojno, B. S. Kim, M. C. Siracusa, P. R. Giacomin, M. G. Nair,A. J. Benitez, K. R. Ruymann, A. B. Muir, D. A. Hill, et al. 2013. Thymicstromal lymphopoietin-elicited basophil responses promote eosinophilic esoph-agitis. Nat. Med. 19: 1005–1013.
11. Halim, T. Y., R. H. Krauss, A. C. Sun, and F. Takei. 2012. Lung natural helpercells are a critical source of Th2 cell-type cytokines in protease allergen-inducedairway inflammation. Immunity 36: 451–463.
12. Mjosberg, J., J. Bernink, K. Golebski, J. J. Karrich, C. P. Peters, B. Blom,A. A. te Velde, W. J. Fokkens, C. M. van Drunen, and H. Spits. 2012. Thetranscription factor GATA3 is essential for the function of human type 2 innatelymphoid cells. Immunity 37: 649–659.
13. Gao, P. S., N. M. Rafaels, D. Mu, T. Hand, T. Murray, M. Boguniewicz, T. Hata,L. Schneider, J. M. Hanifin, R. L. Gallo, et al. 2010. Genetic variants in thymicstromal lymphopoietin are associated with atopic dermatitis and eczema her-peticum. J. Allergy Clin. Immunol. 125: 1403–1407.e4.
14. Margolis, D. J., B. Kim, A. J. Apter, J. Gupta, O. Hoffstad, M. Papadopoulos,and N. Mitra. 2014. Thymic stromal lymphopoietin variation, filaggrin loss offunction, and the persistence of atopic dermatitis. JAMA Dermatol. 150: 254–259.
15. Soumelis, V., P. A. Reche, H. Kanzler, W. Yuan, G. Edward, B. Homey,M. Gilliet, S. Ho, S. Antonenko, A. Lauerma, et al. 2002. Human epithelial cellstrigger dendritic cell mediated allergic inflammation by producing TSLP. Nat.Immunol. 3: 673–680.
16. Lee, E. B., K. W. Kim, J. Y. Hong, H. M. Jee, M. H. Sohn, and K. E. Kim. 2010.Increased serum thymic stromal lymphopoietin in children with atopic derma-titis. Pediatr. Allergy Immunol. 21: e457–e460.
17. Yoo, J., M. Omori, D. Gyarmati, B. Zhou, T. Aye, A. Brewer, M. R. Comeau,D. J. Campbell, and S. F. Ziegler. 2005. Spontaneous atopic dermatitis in miceexpressing an inducible thymic stromal lymphopoietin transgene specifically inthe skin. J. Exp. Med. 202: 541–549.
18. He, R., M. K. Oyoshi, L. Garibyan, L. Kumar, S. F. Ziegler, and R. S. Geha.2008. TSLP acts on infiltrating effector T cells to drive allergic skin inflam-mation. Proc. Natl. Acad. Sci. USA 105: 11875–11880.
19. Li, M., P. Hener, Z. Zhang, K. P. Ganti, D. Metzger, and P. Chambon. 2009.Induction of thymic stromal lymphopoietin expression in keratinocytes is nec-essary for generating an atopic dermatitis upon application of the active vitaminD3 analogue MC903 on mouse skin. J. Invest. Dermatol. 129: 498–502.
20. Kim, B. S., M. C. Siracusa, S. A. Saenz, M. Noti, L. A. Monticelli,G. F. Sonnenberg, M. R. Hepworth, A. S. Van Voorhees, M. R. Comeau, andD. Artis. 2013. TSLP Elicits IL-33-independent innate lymphoid cell responsesto promote skin inflammation. Sci. Transl. Med. 5: 170ra116.
21. Siracusa, M. C., B. S. Kim, J. M. Spergel, and D. Artis. 2013. Basophils andallergic inflammation. J. Allergy Clin. Immunol. 132: 789–801.
22. Obata, K., K. Mukai, Y. Tsujimura, K. Ishiwata, Y. Kawano, Y. Minegishi,N. Watanabe, and H. Karasuyama. 2007. Basophils are essential initiators ofa novel type of chronic allergic inflammation. Blood 110: 913–920.
23. Neill, D. R., S. H. Wong, A. Bellosi, R. J. Flynn, M. Daly, T. K. Langford,C. Bucks, C. M. Kane, P. G. Fallon, R. Pannell, et al. 2010. Nuocytes representa new innate effector leukocyte that mediates type-2 immunity. Nature 464:1367–1370.
24. Moro, K., T. Yamada, M. Tanabe, T. Takeuchi, T. Ikawa, H. Kawamoto,J. Furusawa, M. Ohtani, H. Fujii, and S. Koyasu. 2010. Innate production of TH2cytokines by adipose tissue-associated c-Kit(+)Sca-1(+) lymphoid cells. Nature463: 540–544.
25. Liang, H. E., R. L. Reinhardt, J. K. Bando, B. M. Sullivan, I. C. Ho, andR. M. Locksley. 2012. Divergent expression patterns of IL-4 and IL-13 defineunique functions in allergic immunity. Nat. Immunol. 13: 58–66.
26. Price, A. E., H. E. Liang, B. M. Sullivan, R. L. Reinhardt, C. J. Eisley, D. J. Erle,and R. M. Locksley. 2010. Systemically dispersed innate IL-13-expressing cellsin type 2 immunity. Proc. Natl. Acad. Sci. USA 107: 11489–11494.
3724 INNATE IMMUNE REGULATION OF SKIN INFLAMMATION
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from

27. Siracusa, M. C., S. A. Saenz, E. D. Wojno, B. S. Kim, L. C. Osborne,C. G. Ziegler, A. J. Benitez, K. R. Ruymann, D. L. Farber, P. M. Sleiman, et al.2013. Thymic stromal lymphopoietin-mediated extramedullary hematopoiesispromotes allergic inflammation. Immunity 39: 1158–1170.
28. Saenz, S. A., M. C. Siracusa, L. A. Monticelli, C. G. Ziegler, B. S. Kim,J. R. Brestoff, L. W. Peterson, E. J. Wherry, A. W. Goldrath, A. Bhandoola, andD. Artis. 2013. IL-25 simultaneously elicits distinct populations of innate lym-phoid cells and multipotent progenitor type 2 (MPPtype2) cells. J. Exp. Med. 210:1823–1837.
29. Turner, J. E., P. J. Morrison, C. Wilhelm, M. Wilson, H. Ahlfors, J. C. Renauld,U. Panzer, H. Helmby, and B. Stockinger. 2013. IL-9-mediated survival of type 2innate lymphoid cells promotes damage control in helminth-induced lung in-flammation. J. Exp. Med. 210: 2951–2965.
30. Hoyler, T., C. S. Klose, A. Souabni, A. Turqueti-Neves, D. Pfeifer,E. L. Rawlins, D. Voehringer, M. Busslinger, and A. Diefenbach. 2012. Thetranscription factor GATA-3 controls cell fate and maintenance of type 2 innatelymphoid cells. Immunity 37: 634–648.
31. Spencer, S. P., C. Wilhelm, Q. Yang, J. A. Hall, N. Bouladoux, A. Boyd,T. B. Nutman, J. F. Urban, Jr., J. Wang, T. R. Ramalingam, et al. 2014. Adap-tation of innate lymphoid cells to a micronutrient deficiency promotes type 2barrier immunity. Science 343: 432–437.
32. Mirchandani, A. S., A. G. Besnard, E. Yip, C. Scott, C. C. Bain, V. Cerovic,R. J. Salmond, and F. Y. Liew. 2014. Type 2 innate lymphoid cells drive CD4+
Th2 cell responses. J. Immunol. 192: 2442–2448.33. Ito, Y., T. Satoh, K. Takayama, C. Miyagishi, A. F. Walls, and H. Yokozeki.
2011. Basophil recruitment and activation in inflammatory skin diseases. Allergy66: 1107–1113.
34. Kepley, C. L., S. S. Craig, and L. B. Schwartz. 1995. Identification and partialcharacterization of a unique marker for human basophils. J. Immunol. 154:6548–6555.
35. Sawaguchi, M., S. Tanaka, Y. Nakatani, Y. Harada, K. Mukai, Y. Matsunaga,K. Ishiwata, K. Oboki, T. Kambayashi, N. Watanabe, et al. 2012. Role of mastcells and basophils in IgE responses and in allergic airway hyperresponsiveness.J. Immunol. 188: 1809–1818.
36. van Panhuys, N., M. Prout, E. Forbes, B. Min, W. E. Paul, and G. Le Gros. 2011.Basophils are the major producers of IL-4 during primary helminth infection. J.Immunol. 186: 2719–2728.
37. Ohnmacht, C., C. Schwartz, M. Panzer, I. Schiedewitz, R. Naumann, andD. Voehringer. 2010. Basophils orchestrate chronic allergic dermatitis and pro-tective immunity against helminths. Immunity 33: 364–374.
38. Ohnmacht, C., and D. Voehringer. 2010. Basophils protect against reinfectionwith hookworms independently of mast cells and memory Th2 cells. J. Immunol.184: 344–350.
39. Salimi, M., J. L. Barlow, S. P. Saunders, L. Xue, D. Gutowska-Owsiak, X. Wang,L. C. Huang, D. Johnson, S. T. Scanlon, A. N. McKenzie, et al. 2013. A role for
IL-25 and IL-33-driven type-2 innate lymphoid cells in atopic dermatitis. J. Exp.Med. 210: 2939–2950.
40. Imai, Y., K. Yasuda, Y. Sakaguchi, T. Haneda, H. Mizutani, T. Yoshimoto,K. Nakanishi, and K. Yamanishi. 2013. Skin-specific expression of IL-33 acti-vates group 2 innate lymphoid cells and elicits atopic dermatitis-like inflam-mation in mice. Proc. Natl. Acad. Sci. USA 110: 13921–13926.
41. Noti, M., B. S. Kim, M. C. Siracusa, G. D. Rak, M. Kubo, A. E. Moghaddam,Q. A. Sattentau, M. R. Comeau, J. M. Spergel, and D. Artis. 2014. Exposure tofood allergens through inflamed skin promotes intestinal food allergy through thethymic stromal lymphopoietin-basophil axis. J. Allergy Clin. Immunol. 133:1390–1399.e6.
42. Halim, T. Y., C. A. Steer, L. Matha, M. J. Gold, I. Martinez-Gonzalez,K. M. McNagny, A. N. McKenzie, and F. Takei. 2014. Group 2 innate lymphoidcells are critical for the initiation of adaptive T helper 2 cell-mediated allergiclung inflammation. Immunity 40: 425–435.
43. Roediger, B., R. Kyle, K. H. Yip, N. Sumaria, T. V. Guy, B. S. Kim,A. J. Mitchell, S. S. Tay, R. Jain, E. Forbes-Blom, et al. 2013. Cutaneousimmunosurveillance and regulation of inflammation by group 2 innate lymphoidcells. Nat. Immunol. 14: 564–573.
44. Renz, H., K. Jujo, K. L. Bradley, J. Domenico, E. W. Gelfand, and D. Y. Leung.1992. Enhanced IL-4 production and IL-4 receptor expression in atopic der-matitis and their modulation by interferon-gamma. J. Invest. Dermatol. 99: 403–408.
45. Kayserova, J., K. Sismova, I. Zentsova-Jaresova, S. Katina, E. Vernerova,A. Polouckova, S. Capkova, V. Malinova, I. Striz, and A. Sediva. 2012. A pro-spective study in children with a severe form of atopic dermatitis: clinical out-come in relation to cytokine gene polymorphisms. J. Investig. Allergol. Clin.Immunol. 22: 92–101.
46. Schmutz, S., N. Bosco, S. Chappaz, O. Boyman, H. Acha-Orbea, R. Ceredig,A. G. Rolink, and D. Finke. 2009. Cutting edge: IL-7 regulates the peripheralpool of adult RORg+ lymphoid tissue inducer cells. J. Immunol. 183: 2217–2221.
47. Li-Weber, M., and P. H. Krammer. 2003. Regulation of IL4 gene expression byT cells and therapeutic perspectives. Nat. Rev. Immunol. 3: 534–543.
48. Motomura, Y., H. Morita, K. Moro, S. Nakae, D. Artis, T. A. Endo, Y. Kuroki,O. Ohara, S. Koyasu, and M. Kubo. 2014. Basophil-derived interleukin-4 con-trols the function of natural helper cells, a member of ILC2s, in lung inflam-mation. Immunity 40: 758–771.
49. Doherty, T. A., N. Khorram, S. Lund, A. K. Mehta, M. Croft, and D. H. Broide.2013. Lung type 2 innate lymphoid cells express cysteinyl leukotriene receptor1, which regulates TH2 cytokine production. J. Allergy Clin. Immunol. 132: 205–213.
50. Barnig, C., M. Cernadas, S. Dutile, X. Liu, M. A. Perrella, S. Kazani,M. E. Wechsler, E. Israel, and B. D. Levy. 2013. Lipoxin A4 regulates naturalkiller cell and type 2 innate lymphoid cell activation in asthma. Sci Transl Med 5:174ra126.
The Journal of Immunology 3725
by guest on July 16, 2022http://w
ww
.jimm
unol.org/D
ownloaded from
















![[Monoclonal antibodies in clinical immunology]](https://img.pdfslide.net/doc/110x75/63375eff20d9c9602f0b4441/monoclonal-antibodies-in-clinical-immunology.jpg)












