Embed Size (px)
Citation preview

THROMBOSIS RESEARCH 34; 241-254, 1984 0049-3848/84 $3.00 + .OO Printed in the USA. Copyright (c) 1984 Pergamon Press Ltd. All rights reserved.
A DIRECT, PLASMIN-INDEPENDENT ASSAY FOR PLASMINOGEN ACTIVATOR
Luciana Mussoni', Dan Lawrence and David J. Loskutoff' Department of Immunology
Scripps Clinic and Research Foundation 10666 North Torrey Pines Road La Jolla, California 92037
USA
(Received 2.12.1983; Accepted in revised form 17.2.1984 by Editor T.S. Edgington)
ABSTRACT A direct assay for plasminogen activator (PA) was developed. It employed polyacrylamide gel electrophoresis in the oresence of sodium dodecyl sulfate and fi-mercaptoethanol to monitor PA-mediated conversion of single chain, lz51-plasminogen to two chain plasmin. By incorpo- rating Triton X-100, albumin and trasylol in the reaction buffer, we were able to minimize the adsorptive and autolytic loss of reactants frequently associated with similar approaches. Under these conditions, plasmin formation was linear for at least 24 hours, dose-dependent over a 20-fold range of urokinase concentrations, and at least lOO-fold more sensitive (0.05 units/ml) than previously reported direct assays for PA. The versatility of the assay was demonstrated by its ability to distinguish between urokinase-like and tissue-type PA, and to quan- titate the effects of agents like fibrin and c-amino caproic acid on their respective activities. The assay was readily adapted to detect inhibitors of PA in various samples, and was employed to demonstrate the presence of such inhibitors in both rabbit and bovine endothelial cells. Interestingly, the rabbit inhibitor was found to block the activity of urokinase but not that of tissue-type PA, while the bovine inhibitor neutralized the activities of both molecules. These results demonstratethatcleavage of 1251-plasminogen can be employed as a direct, sensitive and quantitative assay for various PAS, and thus offers a new approach for studying plasminogen activation and agents that stimulate or inhibit it.
Key words: Plasminogen activators, antiactivators, direct assay
-Current address: Milano, Italy.
Instituto Richerche Farmacologiche "Mario Negri",
‘To whom correspondence should be addressed.
241

242 ASSAY FOR PLASMINOGEN ACTIVATOR Vol.34, No.3
INTRODUCTION
The release of plasminogen activators(PAs) by cells is essential to the progression of a diverse number of biologically important processes (l-16), and may represent a general mechanism by which cells initiate localized pro- teolytic activity (17). Unfortunately, our understanding of the various mole- cules and events that influence the production and activity of cellular PAS is minimal, in large part because of the absence of direct assays for these extremely specific molecules. PAS frequently exist as trace cellular proteins, necessitating the development of two-stage, plasmin-dependent assays to amplify their activity (18,19). An alternative approach was suggested which appeared to avoid the ambiguities of interpretation inherent in such two-stage assays (20). This assay employed 1251-plasminogen as substrate, and polyacrylamide gel electrophoresis (PAGE) in the presence of sodium dodecyl sulfate (SDS), to monitor its conversion from an inactive, single chain polypeptide into an active, two chain molecule. This approach was successfully employed in a few instances (20,21). However, cleavage was not linear, thus severely reducing the sensitivity of the assay, making quantitation difficult, and in general restricting its use.
In the present investigation, we re-examined the use of plasminogen cleavage as a direct assay for PA. We found that both the linearity and sensitivity of the assay increased dramatically when Triton X-100 (TX-loo) and albumin were included in the reaction mixture. This modified assay can be used conveniently to study the effects of fibrin, E-amino caproic acid and molecules from endothelial cells on plasminogen activation.
MATERIALS AND METHODS
Cells. Endothelial cells were isolated from intact rabbit vena cava (REVC) or bovine aorta (BAE), and grown in falcon disposable plasticware as described (22). All cells used in these studies had been sub-cultured lo-20 times. Conditioned medium (CM) was prepared by incubating confluent BAEs for 16 hrs in serum-free medium (23), while cell cytosols were prepared by differential centrifugation of homogenates from confluent REVCs (23).
Fibrinolytic activity. Fibrinolytic activity was measured in 12'1-fibrin coated tissue culture wells (16 mm; Costar, Cambridge, MA) as described (23). The assay contained in 1 ml, 4 pg human plasminogen, 0.1% gelatin, and 0.1 M Tris HCl, pH 3.1 supplemented with 25 ~1 CM as a source of PA. Fibrinolysis was not observed when plasminogen or PA was omitted from the reaction mixture. The results are the average of duplicate samples and represent less than 15% variation.
Polyacrylamide gel electrophoresis and autoradiography. Sodium dodecyl sul- fate polyacrylamide gel electrophoresis (SDS-PAGE) was performed in slab gels according to Laemmli (24). The upper stacking gel consisted of 4% acrylamide while the lower separating gel contained 9% acrylamide. Samples were boiled for 3 min. in sample buffer (24) containing SDS (2%) and 6-mercaptoethanol (5%) and subjected to electrophoresis for 18 hours at 40V at room temperature. After electrophoresis, the gels were fixed and stained in 50% trichloroacetic acid, 0.1% Coomassie brilliant blue R-250 (Bio-Rad Laboratories, Richmond, CA) for 30 min., destained in 10% glacial acetic acid, and dried with a slab gel drier (Hoeffer Scientific Instruments, San Francisco, CA). The dried gels were placed on Kodak X-omat film at -70°C to localize the radiolabelled pro- tein. Molecular weight standards included human plasminogen (89,000), human serum albumin (65,000), ovalbumin (43,000), chymotrypsinogen (25,700).

Vo1.34, No.3 ASSAY FOR PLASMINOGEN ACTIVATOR 243
Ai51-plasminogen cleavage assay. The modified 1Z51-plasminogen cleavage assay system contained 0.1% Triton X-100 in O.lM phosphate buffer pH 7.2, supple- mented with 100 ug/ml of human albumin (5x crystallized, Sigma, St. Louis, MO), 1,000 units/ml of trasylol (Calbiochem-Behring, La Jolla, CA), 2 x lo6 cpm/ml of 1251-plasminogen, and a source of PA. Reactants were combined in a total volume of 100 ~1 in polypropylene tubes and then incubated for various times at 37°C. The reaction was stopped by boiling the samples in the presence of an equal volume of 2x concentrated sample buffer. A portion of this mixture was then analyzed by SDS-PAGE. After electrophoresis, the gels were stained, destained, dried and autoradiograms prepared. Quantitative data on lz51- plasminogen cleavage was achieved by removing the portions of the gel contain- ing radiolabelled plasminogen and plasmin (the position of the radiolabelled peptides was determined by placing the X-ray film on the dried gel), and counting them in a gamma counter. Inhibitor activity in this assay was indicated by the ability of samples to block PA-mediated cleavage of single chain plasminogen into the characteristic plasmin "heavy" and "light" chains (20,25) as revealed by SDS-PAGE. The effect of fibrin on plasminogen cleavage was determined as follows: Bovine fibrinogen was added to the complete standard assay containing PA, and the mixture incubated for 5 min at 37°C. Purified human ,-thrombin (a generous gift of Dr. J. Fenton, Albany, NY) was added to a final concentration of 1 unit/ml and the incubation continued as indicated. The mixtures were heated at 100°C for 3 min in the presence of sample buffer to dissolve the fibrin clots and then analyzed for plasminogen cleavage as described above.
Miscellaneous. Human plasminogen was prepared from outdated plasma by affinity chromatography on lysine Sepharose (26), and iodinated by the insolubilized lactoperoxidase method (27) to a specific activity of l-5 x lo6 cpm/pg protein. -amino caproic acid was obtained from Sigma. Radioactivity measurements were
performed in a Packard Gamma-Spectrophotometer. Bovine fibrinogen (fraction I, Calbiochem) was purified free of plasminogen by ethanol precipitation in the presence of lysine (28). Purified human tissue-type PA (t-PA) was isolated from the medium of a human melanoma cell line as described (29), while urokinase was a kind gift from Abbott Laboratories, Chicago, Illinois, and consisted of a mixture of both high and low M forms. All PAS were stored at -20°C in the presence of 0.5% Triton X-100 orr0.014 Tween 80. The BAE inhibitor was purified from CM as described (30) and stored at -20°C in PBS containing 0.01% Tween 80. Protein was determined by the method of Bradford (31) using bovine serum albumin as standard.
RESULTS
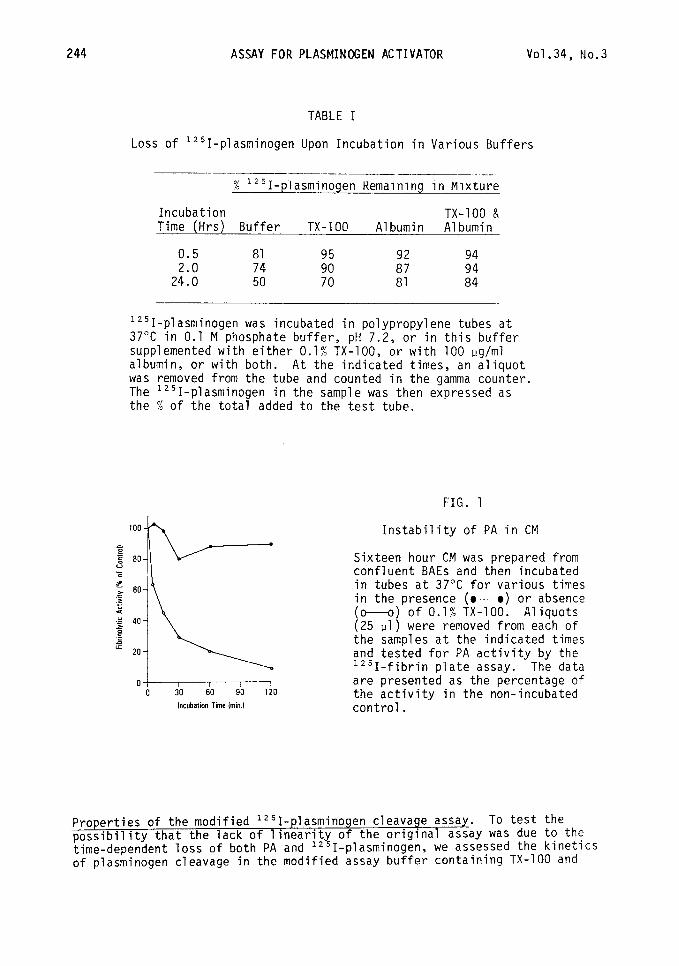
To develop plasminogen cleavage as a direct, sensitive, and reproducible assay for PA, we sought to identify the specific conditions of the original assay (20) that contributed to its lack of linearity. The results presented in Figure 1 and Table I indicate that upon incubation, there is a rapid loss of both PA activity and 1251-plasminogen itself. For example, when serum-free CM was incubated in tubes at 37"C, the PA activity decreased with a half-life of less than 15 min (Fig. 1). Similar results were obtained when purified urokinase and t-PA were tested (not shown), in agreement with the recent obser- vations of others (7,29,32). In addition, the amount of 1251-plasminogen recovered from the reaction mixture also decreased upon incubation, with only 50% of the input radioactivity remaining after 24 hrs (Table I). These losses were reduced dramatically when TX-100 and/or albumin were added to the reaction buffer. On the basis of these observations, the original assay system (20) was modified to contain 0.1% TX-100 and 100 ug/ml of albumin.
244 ASSAY FOR PLASMINOGEN ACTIVATOR vo1.34, No.3
TABLE I
Loss of 1251-plasminogen Upon Incubation in Various Buffers
% i'~I-plasminogen Kemaining in Mixture
Incubation TX-100 & Time (Hrs) Buffer TX-100 Albumin Albumin
0.5 81 95 2.0 74 8': ;: 24.0 50 81 84
1251-plasminogen was incubated in polypropylene tubes at 37°C in 0.1 M phosphate buffer, pH 7.2, or in this buffer supplemented with either 0.1% TX-100, or with 100 pg/ml albumin, or with both. At the indicated times, an aliquot was removed from the tube and counted in the gamma counter. The 1251-plasminogen in the sample was then expressed as the % of the total added to the test tube.
FIG. 1
Ob40 120 Incubation Time Imin.l
Instability of PA in CM
Sixteen hour CM was prepared from confluent BAEs and then incubated in tubes at 37°C for various times in the presence (M) or absence (o---o) of 0.1% TX-100. Aliquots (25 ~1) were removed from each of the samples at the indicated times and tested for PA activity by the lz51-fibrin plate assay. The data are presented as the percentage of the activity in the non-incubated control.
Properties of the modified 1251-plasminogen cleavage assay. To test the possibility that the lack of linearity of the original assay was due to the time-dependent loss of both PA and lz51-plasminogen, we assessed the kinetics of plasminogen cleavage in the modified assay buffer containing TX-100 and

Vo1.34, No.3 ASSAY FOR PLASMINOGEN ACTIVATOR
12 3
245
FIG. 2
89,000- 65,000- -z-:
25,000- *. ti
100
90
80
70
60
s 50 0 E
k
- 40
30
20
10
OETTT-F?O Minutes
Cleavage of 1251-plasminogen by Urokinase.
1*51-plasminogen (Lane 1) was mixed with 100 units/ml of uro- kinase and incubated at 37°C for TLi;$r3i5 min. (Lane 2) or 30 min
. The samples were boiled in the presence of SDS and -met-- captoethanol (sample buffer B and analyzed by SDS-PAGE. The figure shows an autoradiogram of the dried gel.
FIG. 3
Kinetics of Plasminogen Cleavage.
1251-plasminogen was incubated with urokinase (100 units/ml) at 37°C. Aliquots were removed at the indicated times, boiled in sample buffer, and then analyzed by SDS-PAGE and autoradiography. Regions of the gel containing native plasminogen (H), and the plasmin heavy (c---o) and light (M) chains were excised from the dried gel, and counted in a gamma counter. The radio- activity recovered from each of these portions of the gel was then expressed as a percentage of the total cpm recovered from the 3 regions. The total cpm recovered from the 3 regions (m) remained constant over the incuba- tion interval and represented approximately 70% of the total counts recovered from the gel.
246 ASSAY FOR PLASMINOGEN ACTIVATOR Vol.34, No.3
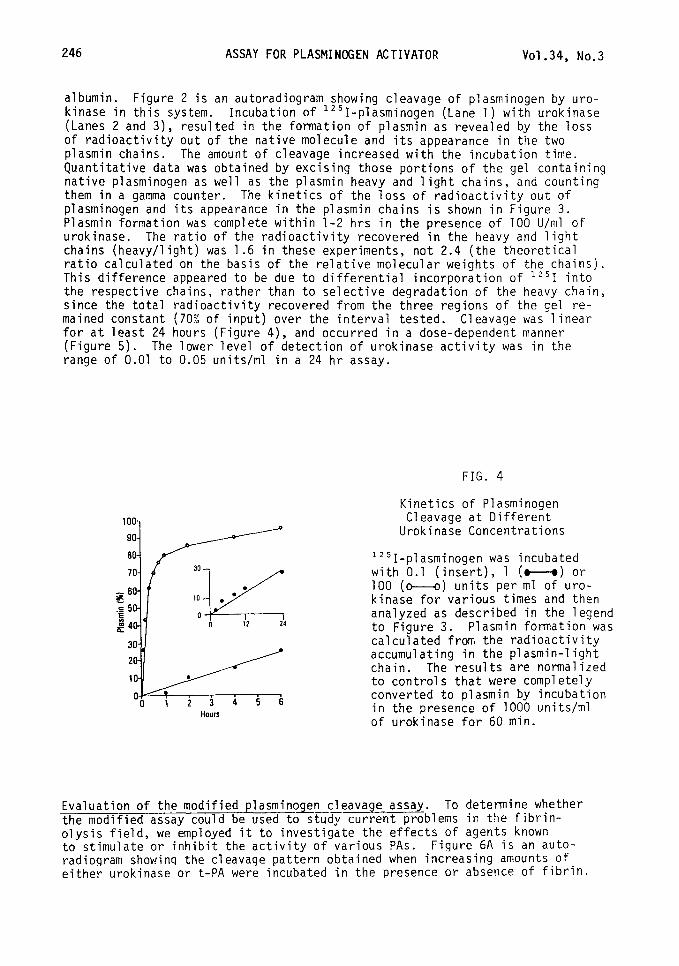
albumin. Figure 2 is an autoradiogram showing cleavage of plasminogen by uro- kinase in this system. Incubation of 1251-plasminogen (Lane 1) with urokinase (Lanes 2 and 3), resulted in the formation of plasmin as revealed by the loss of radioactivity out of the native molecule and its appearance in the two plasmin chains. The amount of cleavage increased with the incubation time. Quantitative data was obtained by excising those portions of the gel containing native plasminogen as well as the plasmin heavy and light chains, and counting them in a gamma counter. The kinetics of the loss of radioactivity out of plasminogen and its appearance in the plasmin chains is shown in Figure 3. Plasmin formation was complete within l-2 hrs in the presence of 100 U/ml of urokinase. The ratio of the radioactivity recovered in the heavy and light chains (heavy/light) was 1.6 in these experiments, not 2.4 (the theoretical ratio calculated on the basis of the relative molecular weights of the chains). This difference appeared to be due to differential incorporation of "'1 into the respective chains, rather than to selective degradation of the heavy chain, since the total radioactivity recovered from the three regions of the gel re- mained constant (70% of input) over the interval tested. Cleavage was linear for at least 24 hours (Figure 4), and occurred in a dose-dependent manner (Figure 5). The lower level of detection of urokinase activity was in the range of 0.01 to 0.05 units/ml in a 24 hr assay.
FIG. 4
Hours
Kinetics of Plasminogen Cleavage at Different
Urokinase Concentrations
1251-plasminogen was incubated with 0.1 (insert), 1 (M) or 100 (M) units per ml of uro- kinase for various times and then analyzed as described in the legend to Figure 3. Plasmin formation was calculated from the radioactivity accumulating in the plasmin-light chain. The results are normalized to controls that were completely converted to plasmin by incubation in the presence of 1000 units/ml of urokinase for 60 min.
Evaluation of the modified plasminogen cleavage assay. To determine whether --- the modified assay could be used to study current problems in the fibrin- olysis field, we employed it to investigate the effects of agents known to stimulate or inhibit the activity of various PAS. Fiqure 6A is an auto- radiogram showinq the cleavage pattern obtained when increasing amounts of either urokinase or t-PA were incubated in the presence or absence of fibrin.
vo1.34, No.3 ASSAY FOR PLASMINOGEN ACTIVATOR 247
0 7
0.01 0.1 1.0 Llnitslml
A. -Fibrin
I 234 567
6. +Fibrin
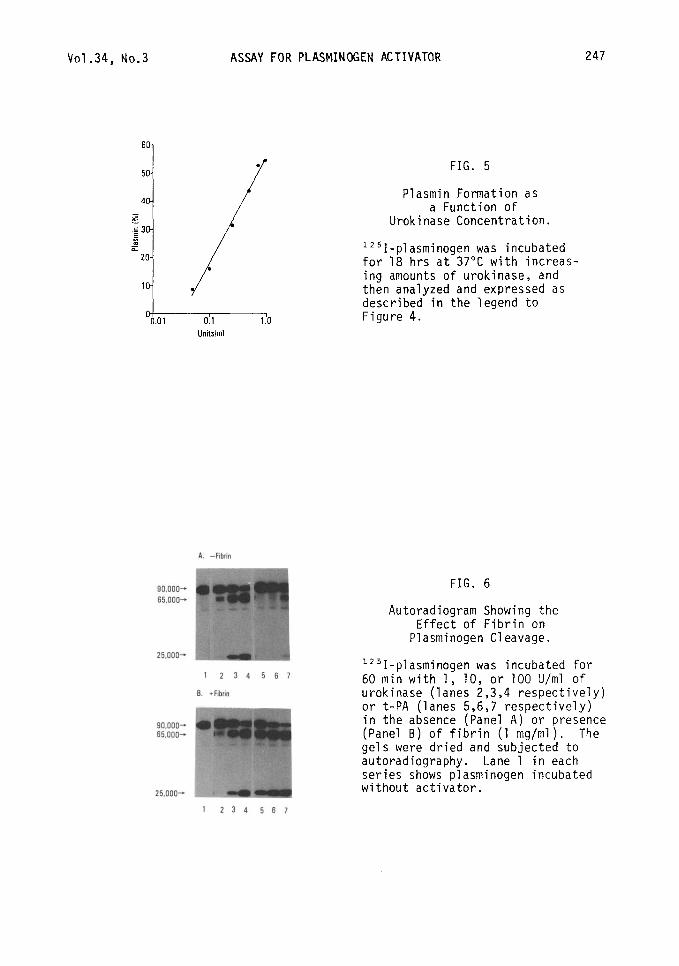
FIG. 5
Plasmin Formation as a Function of
Urokinase Concentration.
lz51-plasminogen was incubated for 18 hrs at 37°C with increas- ing amounts of urokinase, and then analyzed and expressed as described in the legend to Figure 4.
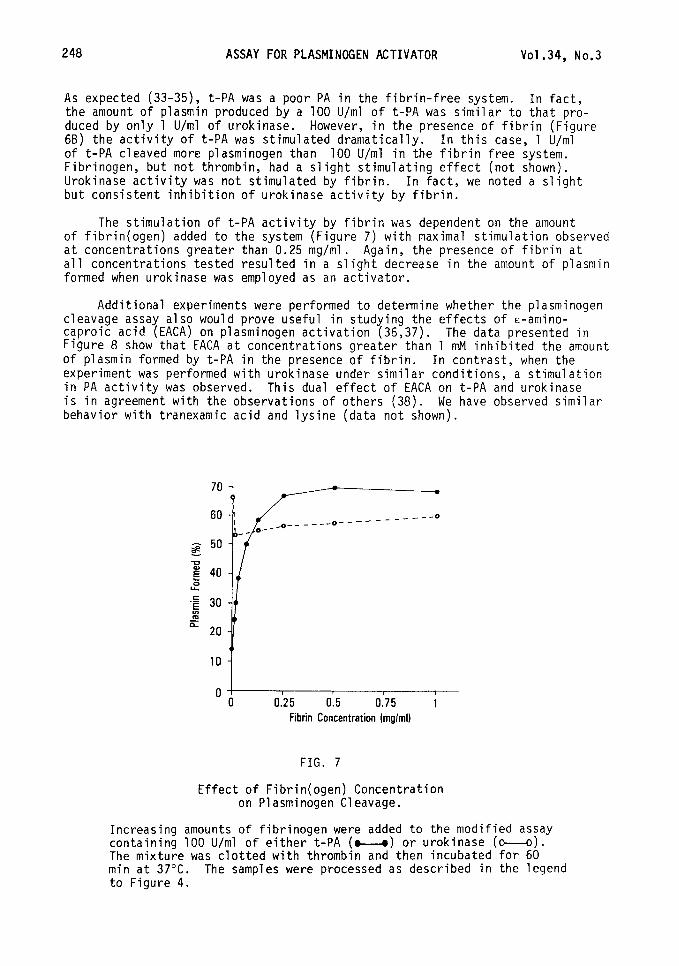
FIG. 6
Autoradiogram Showing the Effect of Fibrin on
Plasminogen Cleavage.
1Z51-plasminogen was incubated for 60 min with 1, 10, or 100 U/ml of urokinase (lanes 2,3,4 respectively) or t-PA (lanes 5,6,7 respectively) in the absence (Panel A) or presence (Panel B) of fibrin (1 mg/ml). The gels were dried and subjected to autoradiography. Lane 1 in each series shows plasminogen incubated without activator.
248 ASSAY FOR PLASMINOGEN ACTIVATOR Vo1.34, No.3
As expected (33-35), t-PA was a poor PA in the fibrin-free system. In fact, the amount of plasmin produced by a 100 U/ml of t-PA was similar to that pro- duced by only 1 U/ml of urokinase. However, in the presence of fibrin (Figure 68) the activity of t-PA was stimulated dramatically. In this case, 1 U/ml of t-PA cleaved more plasminogen than 100 U/ml in the fibrin free system. Fibrinogen, but not thrombin, had a slight stimulating effect (not shown). Urokinase activity was not stimulated by fibrin. In fact, we noted a slight but consistent inhibition of urokinase activity by fibrin.
The stimulation of t-PA activity by fibrin was dependent on the amount of fibrin(ogen) added to the system (Figure 7) with maximal stimulation observed at concentrations greater than 0.25 mg/ml. Again, the presence of fibrin at all concentrations tested resulted in a slight decrease in the amount of plasmin formed when urokinase was employed as an activator.
Additional experiments were performed to determine whether the plasminogen cleavage assay also would prove useful in studying the effects of E-amino- caproic acid (EACA) on plasminogen activation (36,37). The data presented in Figure 8 show that EACA at concentrations greater than 1 mM inhibited the amount of plasmin formed by t-PA in the presence of fibrin. In contrast, when the experiment was performed with urokinase under similar conditions, a stimulation in PA activity was observed. This dual effect of EACA on t-PA and urokinase is in agreement with the observations of others (38). We have observed similar behavior with tranexamic acid and lysine (data not shown).
70
60
Fibrin Concentration Imglml)
FIG. 7
Effect of Fibrin(ogen) Concentration on Plasminogen Cleavage.
Increasing amounts of fibrinogen were added to the modified assay containing 100 U/ml of either t-PA (u) or urokinase (o--o). The mixture was clotted with thrombin and then incubated for 60 min at 37°C. The samples were processed as described in the legend to Figure 4.
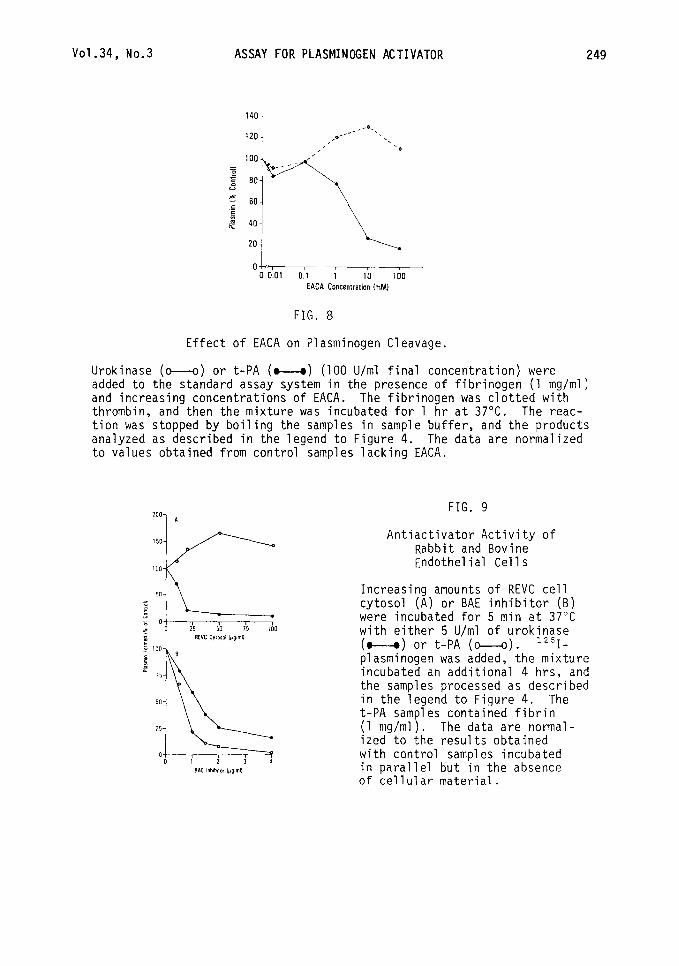
Vo1.34, No.3 ASSAY FOR PLASMINOGEN ACTIVATOR 249
OL 0 0.01 0.1 1 IO 100
EACA Concentration (mMI
FIG. 8
Effect of EACA on Plasminogen Cleavage.
Urokinase (o--o) or t-PA (w) (100 U/ml final concentration) were added to the standard assay system in the presence of fibrinogen (1 mg/ml) and increasing concentrations of EACA. The fibrinogen was clotted with thrombin, and then the mixture was incubated for 1 hr at 37°C. The reac- tion was stopped by boiling the samples in sample buffer, and the products analyzed as described in the legend to Figure 4. The data are normalized to values obtained from control samples lacking EACA.
200 1 FIG. 9
100 Endothelial Cells
Antiactivator Activity of Rabbit and Bovine
Increasing amounts of REVC cell cytosol (A) or BAE inhibitor (B) were incubated for 5 min at 37°C with either 5 U/ml of urokinase (o--o) or t-PA (o--o). 1251- plasminogen was added, the mixture incubated an additional 4 hrs, and the samples processed as described in the legend to Figure 4. The t-PA samples contained fibrin (1 mg/ml). The data are normal- ized to the results obtained with control samples incubated in parallel but in the absence of cellular material.

250 ASSAY FOR PLASMINOGEN ACTIVATOR Vo1.34, No.3
Use of the plasminogen cleavage assay to detect antiactivators. In previous experiments, we were able to employ this direct assay for PA to demonstrate unequivocally that the inhibitor activity associated with the cytosol of rabbit endothelial cells in culture (23) was due to the presence of an antiactivator, not an antiplasmin (39). Figure 9A demonstrates that although this inhibitor blocks the ability of urokinase to cleave plasminogen, it does not inhibit t-PA. In contrast, the inhibitor produced by cultured BAEs (30,40) neutralized the activity of both urokinase and t-PA (Fig. 9B).
DISCUSSION
The present investigation was undertaken with three specific goals in mind. First, we sought to identify the assay conditions responsible for the lack of kinetic linearity frequently encountered in assays that employ 1251- plasminogen as a direct substrate for PA (20). Once defined, we wished to modify the assay to improve its linearity and thus increase its sensitivity. Finally, we wished to establish whether the modified assay had both the sensi- tivity and specificity required to detect and quantitiate cellular PAS and their inhibitors. The results summarized here suggest that the limited linear- ity associated with this approach was due to the time-dependent loss of both PA activity (Fig. 1) and 1251-plasminogen itself (Table I) upon incubation at 37°C. These losses may result from adsorption of these trace proteins to the surface of the incubation tubes, or from their natural instability. In any case, it is clear that the losses were largely overcome by including TX-100 and albumin in the reaction mixture (Fig. 1; Table I), consistent with the observation of others (7,9,32). For example, in the absence of TX-100, the half-life of PA was less than 15 min. However, in its presence, essentially all the PA activity could be recovered, even after 120 min. incubation.
The modified assay had many of the properties required to study PAS present at low concentration in complex biological samples. Thus, cleavage of '*'I- plasminogen was linear for at least 24 hr (Fig. 4) as compared to 20 min in the original assay (20). As a direct consequence of this improved linearity, the sensitivity of the assay increased approximately 20-fold (from 1 U/ml to less than 0.05 U/ml), and the dose response was linear over a 20-fold range of enzyme concentrations (Fig. 5).
There are a number of additional features of the modified assay which suggest that it may provide a useful, general approach for studying PAS and factors that influence their activity. For example, in model studies to more completely evaluate this assay system, we demonstrated that it had the specificity to distinguish between urokinase-like and tissue-type PAS (Fig. 6), and, in addi- tion, could be used conveniently to study the effects of agents like fibrin (Fig. 6,7) and E-amino caproic acid (Fig. 8) on their respective activities. These agents also influence plasmin activity, making the 2-stage, plasmin-depen- dent assays difficult to interpret. Perhaps the greatest potential use of this approach, however, lies in its ability to detect cellular inhibitors of PA. It is in this area that the ambiguity of plasmin-dependent assays is greatest. Thus, although fibrinolytic inhibitors have been detected in various cells and tissues (8,22,23,41-44), it is still unclear whether these are antiactivators or antiplasmins. By employing the direct 1251-plasminogen cleavage assay we were able to demonstrate unequivocally that the inhibitor activity associated with the cytosol of rabbit endothelial cells in culture was due to the presence of an antiactivator and not an antiplasmin (39). Further, the cytosol preparations inhibited plasminogen cleavage initiated by urokinase, but not by t-PA (Fig. 9). The stimulation of t-PA activity may

Vo1.34, No.3 ASSAY FOR PLASMINOGEN ACTIVATOR 251
be due to the presence of denatured proteins (45) in the cytosol. The fibrin- olytic activity of plasmin was unaffected by any of the concentrations of cytosol tested (data not shown). BAEs also produce a fibrinolytic inhibitor (40) that we have purified (30) and begun to characterize. However, unlike the cytosol inhibitor of rabbit ECs, the BAE inhibitor was found to neutralize both urokinase and t-PA (Fig. 9B). Experiments are in progress to further compare and character ize these inhibitors. The results suggest that we should be able to employ the modified assay to establish accurate rate and inactivation constants, determine stoichiometry, and study specificity.
In summary, PAS have been shown to be synthesized and secreted by many different cell types, and may be involved in processes as diverse as tissue reorganization, neovascularization, embryo implantation, metastasis, and vas- cular haemostasis (l-4,17). Elevated levels have been found in transformed cells (1,10,16,18), activated macrophages (1,5), granulosa cells during ovula- tion (4), embryonic cells during differentiation (4,12), hormone-treated uteri (8), and confluent endothelial cells (11,12,46,47). Many of these cells may also produce inhibitors of PA (8,23,30,39,40,48-50). The modified 1251-plasmin- ogen cleavage assay should prove useful both for identifying and characterizing these molecules, and for investigating their interaction.
ACKNOWLEDGMENT
The authors wish to thank Karen Roegner for excellent technical assistance, and Gerry Josephs for typing the manuscript. This work was supported by NIH grant HL22289 to D.J.L. This is publication number 3261-IMM from the Scripps Clinic and Research Foundation.
1.
2.
3.
4.
5.
6.
7.
REFERENCES
REICH, E. Plasminogen activator: secretion by neoplastic cells and macro- phages. In: Proteases and Biological Control. E. Reich, D.B. Rifkin, and E. Shaw (Eds.) New York, Cold Spring Harbor Laboratory, 1975, pp. 333-341.
ASTRUP, T. Fibrinolysis. An overview. In: Progress in Chemical Fibrin- olysis and Thrombolysis, vol. 3. J.F. Davidson, R.M. Rowan, M.M. Samama,- and P.C. Desnoyers (Eds.) New York, Raven Press, 1978, pp. l-57.
COLLEN, D. On the regulation and control of fibrinolysis. Thrombos. Haemostas. 43, 77-89, 1980.
STRICKLAND, S. Plasminogen activator in early development. ment in Mammals.
In: Develop- M.H. Johnson (Ed.) Amsterdam, Elsevier/North Holland,
1980, pp. 81-100.
WERB, Z., MAINARDI, C., VATER, C. and HARRIS, E. Endogenous activation of latent collagenase by rheumatoid synovial cells. Evidence for a role of plasminogen activator. N. Engl. J. Med. 296, 1017-1023, 1977.
OSSOWSKI, L., BIEGEL, D. and REICH, E. Mammary plasminogen activator: correlation with involution, hormonal modulation and comparison between normal and neoplastic tissue. Cell 16, 929-940, 1979.
RIJKEN, D.C., WIJNGAARDS, G., ZAAL-DE JONG, M. and WELBERGEN, J. Purification and partial characterization of plasminogen activator from human uterine tissue. Biochim. Biophys. Acta 580, 140-153, 1979.

252 ASSAY FOR PLASMINOGEN ACTIVATOR Vo1.34, No.3
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
MULLINS, D., BAZER, F. and ROBERTS, R. Secretion of a progesterone- induced inhibitor of plasminogen activator by the porcine uterus. Cell 20, 865-872, 1980.
VETTERLEIN, D., BELL, T.E., YOUNG, P.L. and ROBLIN, R. Immunological quantitation and immunoadsorption of urokinase-like plasminogen activa- tors secreted by human cells. J. Biol. Chem. 225, 3665-3672, 1980.
WILSON, E.L., BECKER, M.L.B., HOAL, E.G. and DOWDLE, E.G. Molecular species of plasminogen activators secreted by normal and neoplastic human cells. Cancer Res. 40, 933-938, 1980.
LEVIN, E.G. and LOSKUTOFF, D.J. Cultured bovine endothelial cells produce both urokinase and tissue-type plasminogen activators. Cell 94, 631-636, 1982.
MAROTTI, K.R., BELIN, D. and STRICKLAND, S. The production of distinct forms of plasminogen activator by mouse embryonic cells. Develop. Biol. 90, 154-159, 1982.
SHARONI, Y., TOPAL, M.C., TUTTLE, P.R. and BERGER, H. A comparison of plasminogen activators derived from rat plasma, primary rat hepatocytes and isolated perfused rat liver. Thrombos. Haemostas. 47, 166-172, 1982.
GROSS, J.L., MOSCATELLI, D. and RIFKIN, D.B. Increased capillary endo- thelial cell protease activity in response to angiogenic stimuli in vitro. Proc. Natl.Acad. Sci. USA 80, 2623-2627, 1983.
GRANELLI-PIPERNO, A. and REICH, E. Plasminogen activators of the pitu- itary gland: enzyme characterization and hormonal regulation. J. Cell. Biol. 9J, 1029-1037, 1983.
WILSON, L., JACOBS, P. and DOWDLE, E.B. The secretion of plasminogen activators by human myeloid leukemic cells in vitro. Blood 6l_, 568-574, 1983.
REICH, E. Activation of plasminogen: A general mechanism for producing localized extracellular proteolysis. In: Molecular Basis of Biological Reproductive Processes. R.D. Berlin, M. Herrman, I.H. Lepow, and J.M. Tanzer (Eds.) New York, Academic Press, 1978, pp. 155-169.
UNKELESS, J.C., TOBIA, A., OSSOWSKI, L., QUIGLEY, J.P., RIFKIN, D.B. and REICH, E. An enzymatic function associated with transformation of fibro- blasts by oncogenic viruses. J. Exp. Med. 137, 85-111, 1973.
DRAPIER, J.C., TENU, J.P., LEMAIRE, G. and PETIT, J.F. Regulation of plasminogen activator secretion in mouse peritoneal macrophages. I. Role of serum studied by a new spectrophotometric assay for plasminogen activators Biochimie 6l_, 463-471, 1979.
DAN@, K. and REICH, E. Plasminogen activator from cells transformed by an oncogenic virus. Inhibitors of the activation reaction. Biochim. Biophys. Acta 566, 138-151, 1979. --
RIJKEN, D.C., HOYLAERTS, M. and COLLEN, 0. Fibrinolytic properties of one-chain and two-chain human extrinsic (tissue-type) plasminogen activator. J. Biol. Chem. 257, 2920-2925, 1982.

Vo1.34, No.3 ASSAY FOR PLASMINOGEN ACTIVATOR 253
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
LEVIN, E.G. and LOSKUTOFF, D.J. Comparative studies of the fibrinolytic activity of cultured vascular cells. Thromb. Res. fi, 869-878, 1979.
LOSKUTOFF, D.J. and EDGINGTON, T.S. Synthesis of a fibrinolytic activator and inhibitor by endothelial cells. Proc. Natl. Acad. Sci. USA 74, 3903-3907, 1977.
LAEMMLI, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685, 1970. ___-
SUMMARIA, L., HSIEH, B. and ROBBINS, K.C. The specific mechanism of activation of human plasminogen to plasmin. 4283, 1967.
J. Biol. Chem. 242, 4279-
DEUTSCH, D.G. and MERTZ, E.T. Plasminogen: purification from human plasma by affinity chromatography. Science 170, 1095-1096, 1970.
DAVID, G.S. and REISFELD, R.A. Protein iodination with solid state lactoperoxidase. Biochemistry 13, 1014-1021, 1976.
MOSESSON, M.W. The preparation of human fibrinogen free of plasminogen. Biochim. Biophys. Acta 57, 204-213, 1962.
RIJKEN, D.C. and COLLEN, D. Purification and characterization of the plasminogen activator secreted by human melanoma cells in culture. J. Biol. Chem. 256, 7035-7041, 1981.
VAN MOURIK, J.A., LAWRENCE, D.J. and LOSKUTOFF, D.J. Characterization of a novel fibrinolytic inhibitor synthesized by bovine aortic endothelial cells in culture. Thrombos. Haemostas. 3, 281, 1983.
BRADFORD, M.M. A rapid and sensitive method for the quantitation of microgram quantitaties of protein utilizing the principle of protein- dye binding. Anal. Biochem. 12, 248-254, 1976.
RADCLIFFE, R. and HEINZE, T. Isolation of plasminogen activator from human plasmaby chromatography on lysine-sepharose. Arch. Biochem. Biophys. l&, 185-194, 1978.
CAMIOLO, S., THORSEN, S. and ASTRUP, T. Fibrinogenolysis and fibrinolysis with tissue plasminogen activator, urokinase, streptokinase-activated human globulin, and plasmin. Proc. Sot. Exp. Biol. Med. 138, 277-280, 1971.
WALLEN, P. Activation of plasminogen with urokinase and tissue activator. In: Thrombosis and Urokinase. Paloetti, R., and S. Sherry (Eds.) New York, Academic Press, 1977, pp. 91-102.
HOYLAERTS, M., RIJKEN, D.C., LIJNEN, H.R. and COLLEN, D. Kinetics of the activation of plasminogen by human tissue plasminogen activator. Role of fibrin. J. Biol. Chem. 257, 2912-2919, 1982.
THORSEN, S. Influence of fibrin on the effect of 6-aminohexanoic acid on fibrinolysis caused by tissue plasminogen activator or urokinase. In: Progress in Chemical Fibrinolysis and Thrombolysis, vol. 3. Davidson, J.F., Rowan, R.M., Samama, M.M., and P.C. Desnoyers (Eds.) New York, Raven Press, 1978, pp. 269-283.

254 ASSAY FOR PLASMINOGEN ACTIVATOR Vol.34, No.3
37. VIOLAND, B.N., BYRNE, R. and CASTELLINO, F.J. The effect of a-, W- amino acids on human plasminogen structure and activation. J. Biol. Chem. 253, 5395-5401, 1978.
38. KOK, P. and ASTRUP, T. Differentiation by means of epsilon-aminocaproic acid. 77-87, 1972.
between plasminogen activators Thromb. Diath. Haemorrh. 27,
39. LOSKUTOFF, D.J. and EDGINGTON, T.S. An activator in rabbit endothelial cells. 1981.
inhibitor of plasminogen J. Biol. Chem. 2_5&, 4142-4145,
40.
41.
LOSKUTOFF, D.J., VAN MOURIK, J.A., ERICKSON, L.A. and LAWRENCE, D. Detection of an unusually stable fibrinolytic inhibitor produced by bovine endothelial cells. Proc. Natl. Acad. Sci. USA 80, 2956-2960, 1983.
MC;LLERTZ, S. Natural inhibitors of fibrinolysis. In: Progress in Chemical Fibrinolysis and Thrombolysis, vol. 3. Davidson, J.F., Rowan, R.M., Samama, M.M., and P.C. Desnoyers (Eds.)mrk, Raven Press, 1978, pp. 213-237.
42. NOORDHOEK HEGT, V. Localization and distribution of fibrinolysis inhibition in the walls of human arteries and veins. Thromb. Res. lo, 121-133, 1977.
43.
44.
45.
BERNIK, M.B. and KWAAN, H.C. Inhibitors of fibrinolysis in human tissues in culture. Am. J. Physiol. 221, 916-921, 1971.
CRUTCHLEY, D.J., CONANAN, L.B. and MAYNARD, J.R. Human fibroblasts produce inhibitor directed against plasminogen activator when treated with glucocorticoids. Ann. N.Y. Acad. Sci. 370, 604-616, 1981.
RADCLIFFE, R. and HEINZE, T. Stimulation of tissue plasminogen activator by denatured proteins and fibrin clots. A possible additional role for plasminogen activator? Arch. Biochem. Biophys. 211, 750-761, 1981.
46.
47.
48.
LAUG, W.E. Secretion of plasminogen activators by cultured bovine endothelial cells: partial purification, characterization and evidence for multiple forms. Thrombos. Haemostas. 3, 219-224, 1981.
BOOYSE, F.M., SCHEINBUKS, J., RADEK, J., OSIKOWICZ, G., FEDER, S. and QUARFOOT, A. Immunological identification and comparison of plasminogen activator forms in cultured normal human endothelial cells and smooth muscle cells. Thromb. Res. 4, 495-504, 1981.
EMEIS, J.J., VAN HINSBERGH, V.W.M., VERHEIJEN, J.H. and WIJNGAARDS, G. Inhibition of tissue-type plasminogen activator by conditioned medium from cultured human and porcine vascular endothelial cells. Biochem. Biophys. Res Comm. 110, 392-398, 1983.
49.
50.
CHAPMAN, H.A., VAVRIN, Z. and HIBBS, J.B. Macrophage fibrinolytic activity: Identification of two pathways of plasmin formation by intact cells and of a plasminogen activator inhibitor. Cell 28, 653-662, 1982.
LEVIN, E.G. Latent tissue plasminogen activator produced by human endo- thelial cells in culture: evidence for an enzyme-inhibitor complex. Proc. Natl. Acad. Sci. USA 80, 6804-6806, 1983.





























