Embed Size (px)
Citation preview

A-Kinase Anchor Protein 84/121 are Targeted toMitochondria and Mitotic Spindles by OverlappingAmino-terminal Motifs
Luca Cardone1, Tiziana de Cristofaro1, Adelina Affaitati1
Corrado Garbi1, Michael D. Ginsberg2, Michele Saviano3
Stelio Varrone1, Charles S. Rubin4, Max E. Gottesman2
Enrico V. Avvedimento1 and Antonio Feliciello1*
1Dipartimento di Biologia ePatologia Molecolare eCellulare, BioGem ConsortiumInstituto di Endocrinologia edOncologia Sperimentale CNRUniversita “Federico II”, via S.Pansini 5, 80131 Naples, Italy
2Institute of Cancer ResearchColumbia UniversityNew York, NY 10032, USA
3Centro Studi BiocristallografiaCNR, via Mezzocannone 6Naples, Italy
4Department of MolecularPharmacologyAtran LaboratoriesAlbert Einstein Collegeof Medicine, BronxNY 10461, USA
A-kinase anchor proteins (AKAPs) assemble multi-enzyme signaling com-plexes in proximity to substrate/effector proteins, thus directing andamplifying membrane-generated signals. S-AKAP84 and AKAP121 arealternative splicing products with identical NH2 termini. These AKAPsbind and target protein kinase A (PKA) to the outer mitochondrial mem-brane. Tubulin was identified as a binding partner of S-AKAP84 in ayeast two-hybrid screen. Immunoprecipitation and co-sedimentationexperiments in rat testis extracts confirmed the interaction between micro-tubules and S-AKAP84. In situ immunostaining of testicular germ cells(GC2) shows that AKAP121 concentrates on mitochondria in interphaseand on mitotic spindles during M phase. Purified tubulin binds directlyto S-AKAP84 but not to a deletion mutant lacking the mitochondrialtargeting domain (MT) at residues 1–30. The MT is predicted to form ahighly hydrophobic a-helical wheel that might also mediate interactionwith tubulin. Disruption of the wheel by site-directed mutagenesisabolished tubulin binding and reduced mitochondrial attachment of anMT-GFP fusion protein. Some MT mutants retain tubulin binding butdo not localize to mitochondria. Thus, the tubulin-binding motif lieswithin the mitochondrial attachment motif. Our findings indicate thatS-AKAP84/AKAP121 use overlapping targeting motifs to localizesignaling enzymes to mitochondrial and cytoskeletal compartments.
q 2002 Elsevier Science Ltd. All rights reserved
Keywords: A-kinase anchor proteins; mitochondria; S-AKAP84/AKAP121;mitotic spindles*Corresponding author
Introduction
Agonist-induced elevations of intracellularcAMP induce a rapid, sustained and reversibleactivation of protein kinase A (PKA). Phosphoryl-ation of PKA substrates regulates critical cellularfunctions, such as development, differentiation,
cell survival, and cycle progression.1,2 PKA isoften concentrated in particulate membranes andcellular organelles through interaction withA-kinase anchor proteins (AKAPs).3 – 6 Althoughthe members of the AKAP family share little over-all primary sequence similarity, their functionaldomains are highly conserved. Thus, AKAPspossess a targeting domain, which serves as ascaffold and membrane anchor, and an amphi-pathic helical tethering domain that binds regu-latory subunits of PKA. Although the preferredligand is RII/PKAII, several AKAPs also bind RI/PKAI.7,8 AKAPs assemble in discrete membranecompartments as a multisubunit complex contain-ing a variety of effectors. In addition to bindingPKAII, some AKAPs associate with protein kinase
0022-2836/02/$ - see front matter q 2002 Elsevier Science Ltd. All rights reserved
E-mail address of the corresponding author:[email protected]
Abbreviations used: AKAP, A-kinase anchor protein;C, catalytic subunit of PKA; ER, endoplasmic reticulum;GSH, reduced glutathione; GST, glutathione; MAP,microtubule-associated protein; MT, mitochondrialtargeting domain; PKA, cAMP-dependent proteinkinase; R, regulatory subunit of PKA; RBD, R-bindingdomain; SOD, superoxide dismutase.
doi:10.1016/S0022-2836(02)00479-5 available online at http://www.idealibrary.com onBw
J. Mol. Biol. (2002) 320, 663–675
C and Ser/Thr phosphatase.9 – 13 AKAPs function atkey nodal points for several signal transductionpathways.14 – 23 We believe that AKAP forms a“transduceosome”, an autonomous multivalentscaffold that assembles and integrates signalsderived from multiple pathways.6
S-AKAP84, AKAP121, D-AKAP1, and AKAP149derive from the same gene by alternative pre-mRNA splicing. They are expressed in the malegerm-cell lineage and in several other tissues,including thyroid and heart. All splice variantsshare the same 525-amino acid residue NH2-termi-nal core, which includes the anchoring domainand the R-binding domain, but diverge signifi-cantly at the C terminus.24 – 29 The first 30NH2-terminal residues (MT) mediate targeting ofS-AKAP84/AKAP121 to the outer membrane ofmitochondria, both in male germ cells and in trans-fected heterologous cells. D-AKAP 1 (dual speci-ficity A-kinase anchor protein) isoforms (N0, N1,C0, and C1) are generated by alternative splicing
occurring at the extreme 50 or 30-ends of the 3.2 kbmRNA transcript. Residues 317–338 of D-AKAP1mediate binding with both type I and type II regu-latory subunits. The targeting motif of N0 (1–30)is identical with S-AKAP84/AKAP121. N1 carries35 additional NH2-terminal residues that anchorthe variant to the endoplasmic reticulum. Thesefindings suggested that residues 1–35 of N1 sup-press mitochondrial targeting and expose an endo-plasmic reticulum (ER)-targeting motif of N0.29
This illustrates how AKAPs of the same familycan be targeted differentially to discrete subcellularcompartments by using overlapping targetingsignals.
S-AKAP84 accumulates in the outer membraneof mitochondria in spermatids at a late phase ofdevelopment.24 The transcription and accumu-lation of AKAP121 in thyroid and in testiculargerm cells are regulated positively by hor-mones that activate the cAMP/PKA pathway,implying a positive feed-back loop between
Figure 1. The NH2 terminus of S-AKAP84/AKAP121 interacts with b tubulin in a yeast two-hybrid sytem. (a). Arepresentation of rat AKAP121. The mitochondrial targeting motif (MT), RII/PKA tethering domain (RII) and putativeRNA-binding motif (KH) are boxed. The recombinant fusion (pGBD-A121N) constituted by a fusion between theDNA-binding domain of Gal4 (GAL4 BD) and the NH2 terminus of AKAP121 is indicated. (b) Yeast strain YRG-2was co-transformed with the indicated yeast expression vectors and grown for two days on selective plates. The histi-dine analog 3-aminotriazole (3-AT) was added to His2 plates. (c) Gal4-dependent transcription of lacZ was evaluatedin yeast strains co-transformed with vectors indicated above. The b-galactosidase activity is expressed as mean ^ S.E.of four independent experiments. (d) Protein extract (100 mg) from control or YRG-2 cells co-transformed with theindicated vectors was immunoblotted with anti-b tubulin and anti-DNA binding domain of GAL4 antibodies.
664 A-Kinase Anchor Protein 84/121
membrane-generated signals and downstreameffector molecules of cAMP/PKA signals.28 PKAanchored to mitochondria is known to enhancethe transmission of survival signals from the cellsurface to these organelles. Phosphorylation andinactivation of proapoptotic BAD depends on themitochondrial anchoring of PKA. A synthetic pep-tide spanning the a-helical wheel of thyroidAKAP (Ht-31) that delocalizes PKAII reduces thephosphorylation of BAD at Ser112 and stimulatescell death.30 Although PKA targeting controlscritical biological functions, the mechanism thatanchors S-AKAP84/PKA complexes to distalorganelles is unknown.
We report here the association betweenS-AKAP84/AKAP121 and cytoskeletal elements.We show that the amino terminus of S-AKAP84/AKAP121 interacts with mitochondrial membranesand with tubulin. Endogenous AKAP121 orexogenous S-AKAP84 fused to green fluorescentprotein (GFP) concentrate on mitochondria ininterphase and on mitotic spindles during meta-phase transition. These findings suggest a novelmechanism to convey cAMP signals to mito-chondrial and cytoskeletal effector molecules.
Results
In vivo interaction between S-AKAP84, tubulinand protein kinase A
To understand the molecular mechanism thatregulates sub-cellular targeting of the S-AKAP84/AKAP121 protein family, we identified proteinsthat mediate its interaction with target organelles.The 50 end of rat S-AKAP84 cDNA (nucleotides
þ1 to þ375) was cloned in the yeast bait plasmidpBD-Gal4 (Stratagene). The recombinant plasmidexpresses a protein with the DNA-binding domainof Gal4 fused to the NH2 terminus (residues1–125) of S-AKAP84, which includes the mito-chondrial targeting motif (Figure 1(a)). A cDNAlibrary derived from rat thyroid mRNA wasinserted into the yeast vector pGAD10 downstreamfrom DNA encoding the activation domain of theGal4 transcription factor (pAD-Gal4).31 Approxi-mately 106 independent clones were screened.Interaction between target fusion proteins andthe NH2-terminal region of S-AKAP84 reconsti-tutes Gal4 transcriptional activity and inducesexpression of his and lacZ reporter genes. cDNAsencoding AKAP-binding proteins were selectedfrom Trpþ Hisþ Leuþ Lacþ colonies. The screenyielded multiple isolates of two cDNAs, includingone clone that corresponded to full-length b tubu-lin cDNA. Coexpression of pGAD-b tubulin withpGBD-AKAP84 strongly activated transcription ofhis and lacZ reporter genes (Figure 1(b) and (c)).The two proteins (GAD-b tubulin and GBD-AKAP84) were expressed in our yeast strain atcomparable levels (Figure 1(d)). Co-immunopre-cipitation assays of rat testis extracts confirmedthat S-AKAP84 and tubulin interact physically invivo. Membrane proteins were solubilized withTriton X-100 and incubated with specific antibodiesto b tubulin, S-AKAP84, or RII. Antibodiesdirected against RII, b tubulin and S-AKAP84
Figure 2. S-AKAP84 interacts with cellular micro-tubules. Membrane proteins (2 mg) extracted from rattestis were immunoprecipitated with anti-RIIa, anti-S-AKAP84, or anti-b tubulin. Pre-immune serum wasused as control. Immunoprecipitates were size-fractionated in an SDS/8% polyacrylamide gel and sub-jected to an RII-overlay assay (left panel) or immunoblotanalysis with anti-S-AKAP84 antibodies (right panel).
Figure 3. Binding of S-AKAP84 to polymerized tubu-lin. Total proteins from rat testis were subjected to cyclesof tubulin polymerization and depolymerization asdescribed in Materials and Methods. Proteins fromsoluble (warm supernatant, WS) and polymerized tubu-lin fractions (warm pellet, WP) were size-fractionatedby SDS-PAGE (10% polyacrylamide gel) and immuno-blotted with antibodies directed against S-AKAP84, btubulin and Harvey-Ras (H-Ras). The numbers refer tothe fractions (supernatant, S, or pellet, P) obtained afterrounds of tubulin polymerization.
A-Kinase Anchor Protein 84/121 665
Figure 4. In vitro binding between S-AKAP84 and b tubulin A. His6-tagged S-AKAP84 was incubated with purifiedGST-b tubulin or with GST polypeptide. An S-AKAP84 mutant deleted for residues 1–30 (S-AKAP84 D1 – 30) was usedalso. The precipitates (bound, B) and supernatant (unbound, U) fractions were size-fractionated by SDS-PAGE (8%polyacrylamide gel) and analyzed by Western blot using anti-S-AKAP84 (upper panel) or anti-GST antibodies (lowerpanel). Representative sets of radiograms are presented. The right panel represents a densitometric analysis (MolecularDynamics) of the experiments indicated in (a). CNT indicates the level of binding of S-AKAP84 to GST beads. Thevalues are expressed as mean ^S.E. of three independent experiments that gave similar results. (b) In vitro bindingbetween His6-tagged S-AKAP84 and GST-b tubulin in the absence or presence of synthetic peptides spanning distinctsub-domains of the mitochondrial targeting motif of S-AKAP84 (residues 1–30). AKAP1 (residues 1–15), AKAP2(residues 10–25) AKAP3 (residues 20–35) peptides were used at concentrations ranging from 1 to 500 nM. The experi-ment shown was carried out at 500 nM peptide. Peptide solubility was monitored by measuring absorbance at 230–280 nm. A representative set of autoradiograms is presented. The right panel represents a densitometric analysis ofthe experiments indicated in (b). Values relative to the control (100%) are presented as mean ^S.E. of four independentexperiments that gave similar results. (c) Recombinant AKAP121 fused to maltose-binding protein (MBP) was purifiedextensively by maltose affinity chromatography and in vitro 32P-labeled using the PKA catalytic subunit. Maximalbinding to immobilized GST-b tubulin was determined by incubating different concentrations of AKAP121 to excessof GST-tubulin. Binding of [32P]AKAP121 to GST-tubulin was competed with increasing concentrations of unlabeledAKAP121. The binding over the range of AKAP concentrations was measured by densitometry of the autoradiograms.The values of the binding curve are expressed as mean ^ S.E. of four independent experiments.
666 A-Kinase Anchor Protein 84/121

efficiently co-immunoprecipitated an 84 kDaRII-binding protein (Figure 2, overlay, left panel).Western blot and immunodepletion analyses (datanot shown) of these immunoprecipitates identifiedthe 84 kDa RII-binding protein as S-AKAP84(Figure 2, right panel). Thus, S-AKAP84, tubulin,and RII form a stable complex in vivo. We nextasked whether S-AKAP84 binds to soluble and/orpolymerized tubulin. Cycles of tubulin polymeri-zation/depolymerization were performed on totalrat testis proteins. Figure 3 shows that S-AKAP84efficiently binds both polymerized microtubules(WP) and soluble tubulin (WS), after three cyclesof tubulin polymerization (see the legend to Figure3). A control protein, H-Ras, binds to neither formof tubulin.
In vitro binding between S-AKAP84 and tubulin
To study in detail the interaction between tubu-lin and S-AKAP84, we performed in vitro bindingexperiments using purified recombinant proteins.As shown in Figure 4(a), recombinant His6-taggedS-AKAP84 directly bound recombinant GST-btubulin immobilized on GSH beads. We nextasked if the tubulin and mitochondrial bindingmotifs of S-AKAP84 overlapped. The deletion ofthe first 30 NH2-terminal amino acid residues,which contains the mitochondrial targeting (MT)motif, abolished b tubulin binding (Figure 4(a)).To define further these motifs, b tubulin bindingwas measured in the presence of competitorsynthetic peptides spanning distinct sub-segmentsof the MT motif. As shown in Figure 4(b), pre-incu-bation of GST-b tubulin with AKAP peptide 2(residues 10–25) significantly inhibited S-AKAP84binding, whereas peptides 1 and 3 (residues 1–15and 20–35, respectively) did not (see the legend toFigure 4). These data suggest that S-AKAP84 resi-dues 10–25 are necessary for b tubulin binding.The binding affinity between b tubulin andAKAP121 was determined as described inMaterials and Methods. Recombinant 32P-labeledAKAP121 was bound to immobilized GST-tubulinin the presence of increasing amounts of unlabeledAKAP121. Binding was detected by auto-radiography and measured by densitometry. Ourdata indicate that b tubulin and AKAP121 interactwith half-maximal binding values of approxi-mately 50–70 nM (Figure 4(c)). Given the high con-centrations of cellular tubulin, the interaction withAKAP121 is likely to occur in vivo.
A secondary structure prediction of the MTmotif of S-AKAP84/AKAP121 was generated bythe 3D-PSSM web server Biomolecular ModelingLaboratory.32 The MT domain is predicted to forma highly hydrophobic a-helical wheel (Figure 5(a)and (b)). This structure is similar to that of the tar-geting domain of NADH-cytochrome b reductase,which also localizes to the outer mitochondrialmembrane.24,33 The helical wheel conformationmight be important for both tubulin binding andmitocondrial targeting. To test this idea, the inter-
action between S-AKAP84 and b tubulin wasstudied using fusions between S-AKAP84 or itsMT (residues 1–30) and green fluorescent protein(S-AKAP84-GFP and S-AKAP84(1 – 30)-GFP). Fusionbetween distinct sub-segments of the MT motif andGFP (S-AKAP84(1 –15)-GFP, S-AKAP84(10 – 25)-GFP,S-AKAP84(1 – 25)-GFP and S-AKAP84(16 – 30)-GFP)were also constructed. cDNAs encoding the GFPfusion proteins were subcloned in a eucaryoticexpression vector and transcribed and translatedin reticulocyte lysates in the presence of [35S]methi-onine (Figure 5(c)). The 35S-labeled GFP fusionswere tested in vitro for binding to recombinantGST-b tubulin immobilized on GSH beads. Figure5(d)–(f) shows that S-AKAP84-GFP, S-AKAP84(1–30)-GFP, S-AKAP84(10 – 25)-GFP, S-AKAP84(16 – 30)-GFPand S-AKAP84(1 – 25)-GFP were retained on thecolumn, suggesting that residues 16–25 are neces-sary and sufficient for association with b tubulin.We also generated a mutant (S-AKAP84(1 – 30)mut-GFP) in which hydrophobic residues within thetubulin-binding motif (Met16, Ala18, Leu20,Phe25) were substituted with aspartate. Thesemutations, which are predicted by structuralanalysis to disrupt the helical conformation of theprotein, abolished binding of the chimera to btubulin. These observations indicate that S-AKAP84,in contrast to other tubulin-binding proteins,binds to b tubulin via non-polar interactions.
S-AKAP84/AKAP121 associates with mitoticspindle microtubules in GC2 cells
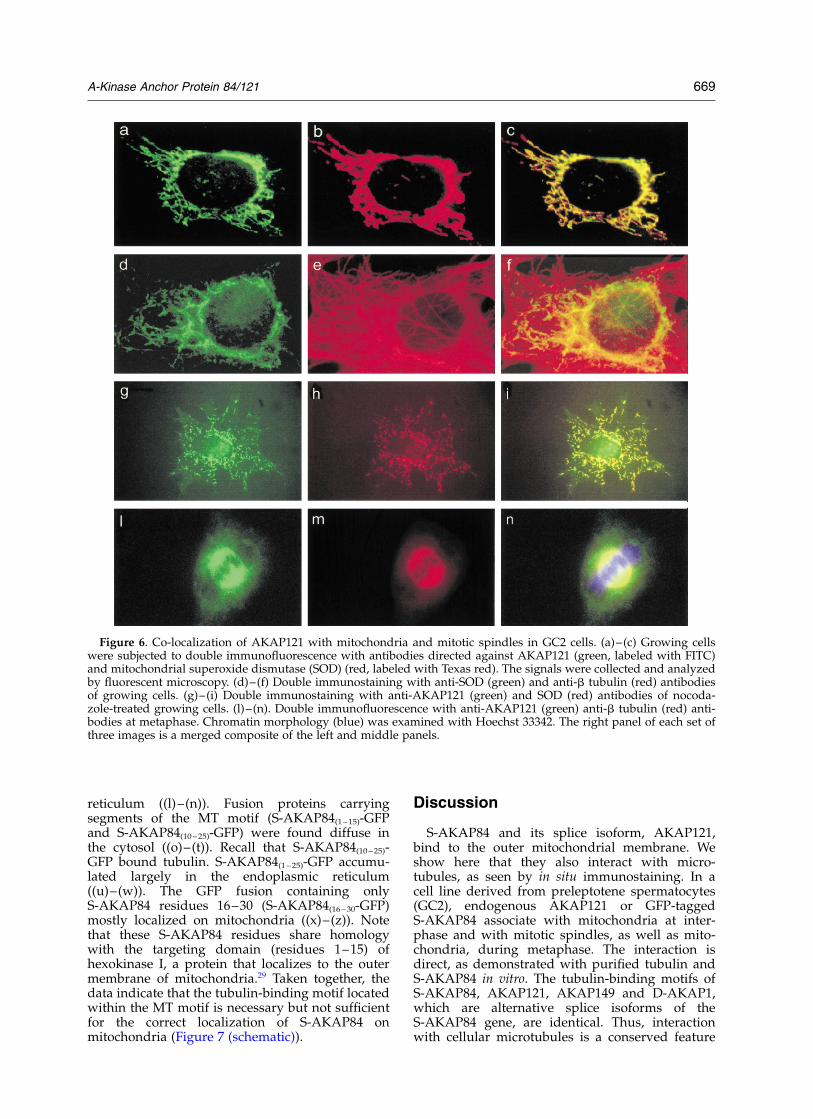
The subcellular distribution of S-AKAP84/AKAP121 and its association with microtubuleswas investigated by immunocytochemistry inGC2 spermatocytes, which express high levels ofAKAP121.28 As shown in Figure 6 (a)–(c)) and, asdemonstrated previously,25 AKAP121 colocalizeswith superoxide dismutase (Mn-SOD), a proteinthat accumulates selectively in mitochondria.34
Some codistribution of AKAP121 with micro-tubules could be detected using antibodies againstb tubulin (data not shown). Additionally, theimmunostains for mitochondria and for micro-tubules partly overlap (Figure 6 (d)–(f)). We thenasked if mitochondrial targeting was mediated bytubulin polymers. Cells were treated with nocoda-zole to depolymerize microtubules. Althoughnocodazole induced mitochondria to redistributethroughout the cytoplasm, it did not affect mito-chondrial localization of AKAP121 (Figure 6(g)–(i)).35 This experiment is consistent with the possi-bility that AKAP121 attaches to mitochondria viatubulin monomers.
We were particularly interested in knowingif AKAP121 localization was influenced byrearrangement of microtubules during meta-phase.36,37 This rearrangement plays a key role inthe formation of bipolar mitotic spindles that dic-tate the position of the cleavage plane (midzone)and allow the ingression of the cleavage furrowand completion of cytokinesis. These events
A-Kinase Anchor Protein 84/121 667
support the physical process of cell division andproper chromosome segregation. In metaphase,AKAP121 staining could be seen on mitotic spin-dles, with a distribution similar to that of tubulin(Figure 6(l)–(n)). AKAP121 association with mito-chondria, however, persists in metaphase, asshown by double labeling with AKAP121 andsuperoxide dismutase (SOD) antibodies (data notshown). Thus, S-AKAP84 appears to associatewith tubulin independently of mitochondria.
Mutations in the tubulin-binding motif ofS-AKAP84 impair mitochondrial targeting
We next studied the subcellular distribution ofthe S-AKAP84-GFP fusion proteins in cycling
cells. Expression vectors were transiently trans-fected into GC2 cells and the subcellular locationof the chimeric proteins was determined by fluor-escent microscopy. As shown in Figure 7, S-AKAP84-GFP, like endogenous AKAP121, accu-mulated selectively on mitochondria in interphase(Figure 7(a)–(c)), and with both mitotic spindlesand mitochondria during metaphase transition(Figure 7(d)–(f)).
To explore the relationship between tubulinbinding and mitochondrial anchoring, cDNAsencoding GFP fusions with all or part of the MTmotif were microinjected into GC2 cells (Figure7(g)–(z)). S-AKAP84(1 – 30)-GFP concentrated selec-tively on mitochondria ((g)–(i)). S-AKAP84(1 – 30)mut-GFP accumulated largely in the endoplasmic
Figure 5. Interaction of NH2 terminus (residues 1–30) of S-AKAP84 with tubulin. (a) A representation of S-AKAP84protein. MT (mitochondrial targeting domain), RBD (R binding domain). (b) Secondary structure prediction and thea-helical wheel analysis of the 14–26 segment of the MT. Hydrophobic residues of the putative tubulin-bindingdomain that have been mutagenized are indicated in bold. (c)–(f) In vitro translated and 35S-labeled S-AKAP84(1 – 30)-GFP, and other chimera (S-AKAP84(1 – 25)-GFP, S-AKAP84(1 –15)-GFP S-AKAP84(16 – 30)-GFP, S-AKAP84(10 – 25)-GFP,S-AKAP84(1 –30)mut-GFP) were subjected to pull-down experiments with recombinant GST-b tubulin immobilized onGSH beads, as indicated in Materials and Methods. GST polypeptide was used as control. The amounts (input) of35S-labeled chimera used for pull-down experiments were analyzed by SDS-PAGE (12% polyacrylamide gel).(c) The gel was fixed with methanol/acetic acid, enhanced with sodium salicylate, dried and exposed to X-ray film.(e) Chimera bound to the GST-tubulin beads were size-fractionated by SDS-PAGE (12% polyacryalmide gel) andvisualized by autoradiography. (d) The upper part of the same gel was immunoblotted with anti-GST antibody.(f) Densitometric analysis of the experiments indicated in (c)–(e). Values are expressed as mean ^ S.E. of threeindependent experiments that gave similar results.
668 A-Kinase Anchor Protein 84/121
reticulum ((l)–(n)). Fusion proteins carryingsegments of the MT motif (S-AKAP84(1 – 15)-GFPand S-AKAP84(10 – 25)-GFP) were found diffuse inthe cytosol ((o)–(t)). Recall that S-AKAP84(10 – 25)-GFP bound tubulin. S-AKAP84(1 – 25)-GFP accumu-lated largely in the endoplasmic reticulum((u)–(w)). The GFP fusion containing onlyS-AKAP84 residues 16–30 (S-AKAP84(16 –30-GFP)mostly localized on mitochondria ((x)–(z)). Notethat these S-AKAP84 residues share homologywith the targeting domain (residues 1–15) ofhexokinase I, a protein that localizes to the outermembrane of mitochondria.29 Taken together, thedata indicate that the tubulin-binding motif locatedwithin the MT motif is necessary but not sufficientfor the correct localization of S-AKAP84 onmitochondria (Figure 7 (schematic)).
Discussion
S-AKAP84 and its splice isoform, AKAP121,bind to the outer mitochondrial membrane. Weshow here that they also interact with micro-tubules, as seen by in situ immunostaining. In acell line derived from preleptotene spermatocytes(GC2), endogenous AKAP121 or GFP-taggedS-AKAP84 associate with mitochondria at inter-phase and with mitotic spindles, as well as mito-chondria, during metaphase. The interaction isdirect, as demonstrated with purified tubulin andS-AKAP84 in vitro. The tubulin-binding motifs ofS-AKAP84, AKAP121, AKAP149 and D-AKAP1,which are alternative splice isoforms of theS-AKAP84 gene, are identical. Thus, interactionwith cellular microtubules is a conserved feature
Figure 6. Co-localization of AKAP121 with mitochondria and mitotic spindles in GC2 cells. (a)–(c) Growing cellswere subjected to double immunofluorescence with antibodies directed against AKAP121 (green, labeled with FITC)and mitochondrial superoxide dismutase (SOD) (red, labeled with Texas red). The signals were collected and analyzedby fluorescent microscopy. (d)–(f) Double immunostaining with anti-SOD (green) and anti-b tubulin (red) antibodiesof growing cells. (g)–(i) Double immunostaining with anti-AKAP121 (green) and SOD (red) antibodies of nocoda-zole-treated growing cells. (l)–(n). Double immunofluorescence with anti-AKAP121 (green) anti-b tubulin (red) anti-bodies at metaphase. Chromatin morphology (blue) was examined with Hoechst 33342. The right panel of each set ofthree images is a merged composite of the left and middle panels.
A-Kinase Anchor Protein 84/121 669
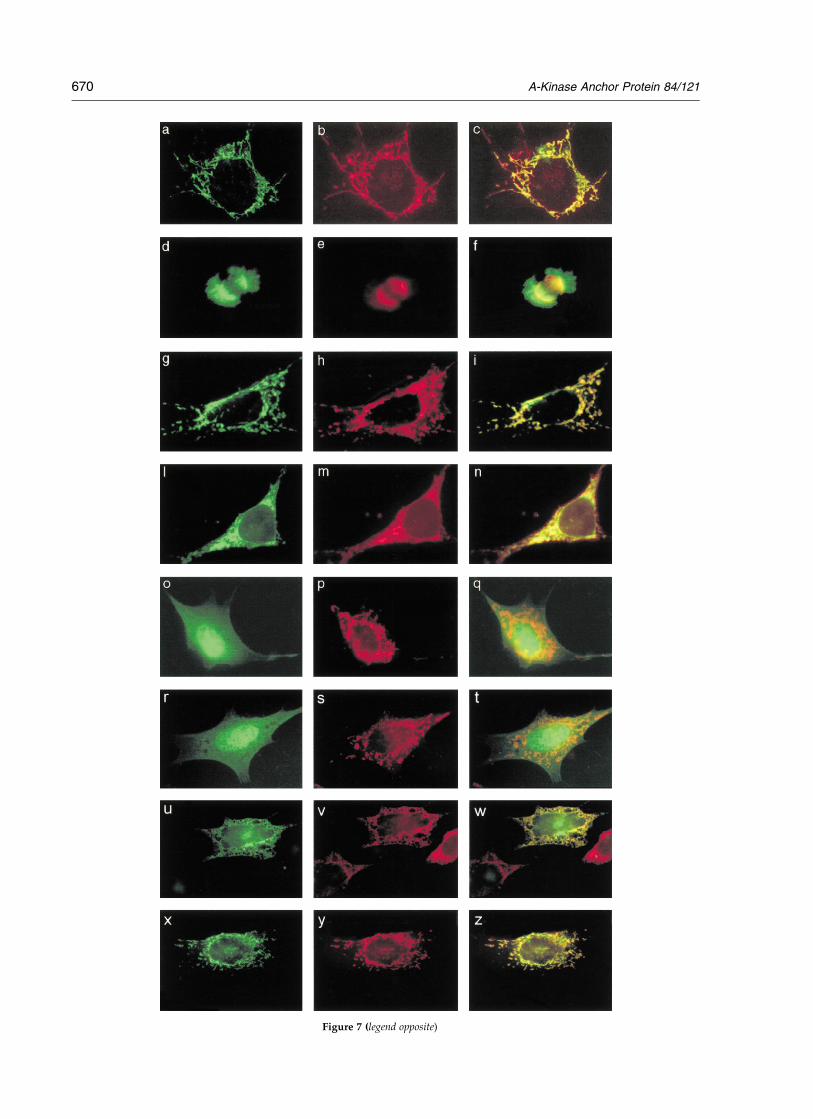
Figure 7 (legend opposite)
670 A-Kinase Anchor Protein 84/121

of these anchor proteins. Mutation of hydrophobicresidues located within this motif ablates binding.The tubulin-binding motif (residues 10–25) lieswithin the MT motif (residues 1–30). AlthoughS-AKAP84 binds tubulin polymers, we do notbelieve that they participate in mitochondrialtargeting. Thus, nocodazole, which depolymerizesmicrotubules, did not eliminate mitochondrialassociation. However, S-AKAP84 also bindssoluble tubulin, and it is possible that tubulinmonomers promote attachment of the anchorprotein to the mitochondria.
Tubulin, a heterodimer composed of two similaracidic isoforms (a and b), participates in theorganization of eukaryotic microtubule networks.Maintainance of microtubular architecture isassured by the dynamic assembly/disassembly oftubulin heterodimers. This mechanism, which con-trols cellular shape, organelle transport, motilityand cell division, requires the interaction of micro-tubules with several microtubule-associatedproteins.38,39 Microtubule-associated proteins(MAPs) such as tau, MAP2 and MAP4, share a
basic binding domain that interacts with the acidicC-terminal region of tubulin. The C-terminalregion lies on the outer face of the microtubuleprotofilament. It is highly variable among tubulinisoforms and plays an important role in the self-assembly of microtubule filaments.38,39 The findingthat hydrophobic residues in S-AKAP84 mediatethe interaction with tubulin was, therefore, quiteunexpected. It is possible that other, as yet unde-fined, microtubule-binding proteins might alsoutilize hydrophobic interactions. Prediction analy-sis of secondary structure suggests the formationof a-helical wheel located at the extreme NH2
terminus of S-AKAP84/121. The structure mightconstitute a novel binding module that mediatesthe interaction with tubulin. At present, we do notknow if S-AKAP84/AKAP121, like MAPs, bindsthe tubulin C terminus.
Several AKAP chimeras fail to associate withmitochondria or cytoskeleton and accumulatein the endoplasmic reticulum. This might be aconsequence of a misfolding of the fusion proteinor of the unmasking of a cryptic ER targeting
Figure 7. Tubulin binding and mitochondrial targeting motifs overlap. Opposite: (a) GC2 cells were transfectedtransiently with vectors carrying GFP fusion proteins (green) and subjected to immunofluorescence with different anti-bodies (red). (a)–(c) Immunostaining with anti-MnSOD antibodies of interphase cells transiently expressingS-AKAP84-GFP. (d)–(f) Cells as in a-c at metaphase transition. g-i Immunostaining with anti-MnSOD antibody ofgrowing cells transiently expressing S-AKAP84(1 – 30)-GFP chimera. (l)–(n) Immunostaining with anti-calreticulin anti-body of growing cells transiently expressing S-AKAP84(1 – 30)mut-GFP chimera. (o)–(q) Immunostaining with anti-MnSOD antibodies of growing cells transiently expressing S-AKAP84(10 –25)-GFP chimera. (r)–(t) Immunostainingwith anti-MnSOD antibody of growing cells transiently expressing S-AKAP84(1 – 15)-GFP chimera. (u)–(w) Immuno-staining with anti-calreticulin antibody of growing cells transiently expressing S-AKAP84(1 – 25)-GFP chimera. (x)–(z)Immunostaining with anti-MnSOD antibody of growing cells transiently expressing S-AKAP84(16– 30)-GFP chimera.The right panel of each set of three images is a merged composite of the left and middle panels. Above: Subcellularlocalization and tubulin-binding properties of distinct S-AKAP84-GFP chimera. Numbers indicate amino acid residuesof the mitochondrial targeting domain (MT) of S-AKAP84. Mit, mitochondrion; cyt, cytoplasm; ER, endoplasmicreticulum.
A-Kinase Anchor Protein 84/121 671

motif. However, a natural splice variant ofS-AKAP84, D-AKAP1, bearing 35 additional NH2-terminal amino acid residues accumulates in theER. It has been suggested that these residues maskthe MT motif and activate a cryptic ER localizationmotif.29 By analogy, mutations of the tubulin-binding motif might disrupt the mitochondrial tar-geting of S-AKAP84 and unmask the ER-targetingmotif.
Rearrangement of the microtubule cytoskeletonduring mitosis promotes the formation of bipolarmitotic spindles that dictate the position of thecleavage plane (midzone). Subsequently, the micro-tubules of the spindle midzone allow ingression ofthe cleavage furrow and completion of cytokinesis.These events support the physical process of celldivision and proper chromosome segregation.36,37
What might be the physiological role(s) of theinteraction between microtubules and S-AKAP84?AKAP/PKA complexes anchored to mitoticspindles might regulate the dynamic assembly/disassembly of microtubules during cell division.In this context, S-AKAP84/PKA might cooperatewith other microtubule-associated RII-binding pro-teins to create target sites of cAMP action.40 – 42 Forexample, PKA targeted to microtubules mightmodulate phosphorylation and activity of micro-tubular substrates in a cell-cycle-specific manner.Phosphorylation of certain MAPs by PKA blockstheir association with microtubules and, therefore,plays a pivotal role in the dynamic and functionalorganization of the microtubule network.
The interaction between S-AKAP84 and tubulinmight also provide a structural bridge for themovement of mitochondria across the micro-tubular network. In spermatogenic cells, theaccumulation of S-AKAP84 on mitochondria iscoincident with developmentally timed re-distri-bution of mitochondria along the microtubuleaxoneme.24 Tubulin heptamers are associated withthe outer mitochondrial membrane, implying aface-to-face contact between these organelles andthe microtubular tracks (J. Wolff, personalcommunication).43 – 45 It is possible that S-AKAP84might influence mitochondrial association withmicrotubules by enhancing the phosphorylation ofMAP2 and TAU by PKA. MAP2 and TAU areknown to affect the binding of purified micro-tubules to isolated mitochondria.46
The NH2 terminus of AKAP121 is unusual, sinceit contains overlapping motifs that bind mito-chondria and a cytoskeletal component, tubulin.We did not isolate mutants that attached to mito-chondria but failed to bind tubulin, suggestingthat tubulin interaction is important for mito-chondrial targeting. Further studies will show ifinteraction with tubulin is a prerequisite for otherproteins that bind to the mitochondrial outer mem-brane. We also note that we screened only onecDNA library for S-AKAP84 interacting proteins.It remains entirely possible that a screen of anotherlibrary, e.g. testis, will reveal additional proteinsthat bind to the MT domain.
Materials and Methods
Cell lines
GC2 cells were generously provided by J. L. Millanand M. C. Hofmann, La Jolla Cancer Research Foun-dation, CA. GC2 cells were derived from primarymouse preleptotene spermatocytes by stable co-transfec-tion with transgenes encoding SV40 large T-antigen anda temperature-sensitive variant of the p53 transcriptionalregulator protein.47 GC2 cells were grown at 37 8C inDulbecco’s modified Eagle’s medium (DMEM) supple-mented with 10% (v/v) fetal calf serum.
Plasmid and transfection
A fusion between full-length rat S-AKAP84 and greenfluorescent protein (GFP) was generated by subcloningHindIII-digested cDNA of S-AKAP84 in-frame withGFP in the pEGFP-N1 vector (Clontech) pre-digestedwith HindIII.28 Distinct sub-segments of the MT ofS-AKAP84 were appended to the NH2 terminus of GFPby sequential amplification of GFP cDNA template,using oligonucleotide primers spanning the extreme50-end of the S-AKAP84 gene. The primers contain anEco RI restriction site. The reverse oligonucleotide primerwas the same for all amplifications (GCTTTACTTGTAC-AGCTCGTCCATGCCGAGAGTGATC), while forwardprimers are the following: S-AKAP841 – 15-GFP, ATGGCA-ATCCAGTTCCGCTCACTCTTCCCCTTGGCCTTGCCT-GGAATGGTGAGCAAGGGCGAGGAG; S-AKAP8416 – 30-GFP, ATGCTAGCACTCCTTGGCTGGTGGTGGTTTTTC-TCTCGTAAAAAAGATATGGTGAGCAAGGGCGA-GGAG CTG; S-AKAP8410 – 25-GFP, ATGCCCTTGGCCTT-GCCTGGAATGCTAGCACTCCTTGGCTGGTGGTGGT-TTATGGTGAGCAAGGGCGAGGAG; S-AKAP841 – 30-GFP, ATGGCAATCCAGTTCCGCTCACTCTTCCCCTT-GGCCTTGCCTGGAATGCTAGCACTCCTTGGCTG-GTGG; S-AKAP8416 – 30-GFP cDNA was used as template;S-AKAP841 – 30mut-GFP, ATGGCAATCCAGTTCCGCTCA-CTCTTCCCCTTGGCCTTGCCTGGAGACCTAGACCT-CGACGGCTGGTGG; S-AKAP8416 – 30-GFP cDNA wasused as template; S-AKAP841 – 25-GFP, ATGGCAATCCA-GTTCCGCTCACTCTTCCCCTTGGCCTTGCCTGGAAT-GCTAGCACTC; S-AKAP8410 – 25-GFP cDNA was used astemplate.
PCR cDNA products were subcloned in a CMVexpression vector (pcDNA3, Invitrogen) and sequenced.DNA transfections were carried out by the calcium phos-phate procedure, as described.48 Microinjection of GFPexpression vectors was carried out as described.18 Briefly,the microinjection apparatus consisted of a phase con-trast microscope connected to a computer-aided imageanalyzer (AIS automatic injection system, Zeiss,Germany) and a computer-operated microinjector(Eppendorf, Germany). In each experiment 150 cells/dish and two dishes/DNA were injected. At 18 hoursafter the injection, the cells were subjected to immuno-fluorescence analysis.
Expression and purification of recombinantfusion proteins
Full-length or amino-terminal truncations ofS-AKAP84 and GST-b tubulin recombinant proteinswere prepared as follows. Full-length cDNA and50-deletion mutants for S-AKAP84 were generated byPCR using specific oligonucleotide primers. The primerswere designed to create HindIII restriction sites at the 50
672 A-Kinase Anchor Protein 84/121

and 30-ends of the product. PCR products were digestedwith HindIII restriction enzyme and cloned in thepRSET vector pre-digested with HindIII. Retention ofthe correct S-AKAP84-coding region was confirmed byDNA sequencing. Full-length b tubulin cDNA wasexcised from pAD-GAL4 positive clone of the libraryand subcloned in-frame with GST polypeptide usingpGEX-4 vector. Hexahistidine (His)6-tagged or GST-recombinant fusion proteins were expressed inEscherichia coli BL-21 (DE3). His6 or GST-tagged fusionproteins were purified following the manufacturer’sinstructions.
GST pull-down experiments
GST-b tubulin beads (20 ml) were incubated with 1 mgof recombinant His6-S-AKAP84 polypeptides in 200 mlof PBS1X containing 0.5% (v/v) Triton X-100 in rotationat 4 8C for three hours. The pellets were washed fourtimes in the binding buffer and eluted in Laemmli buffer.The eluted samples were resolved by PAGE (8% poly-acrylamide gel), transferred to polyvinylidene difluoridemembranes and immunoblotted with anti-b tubulin,anti-GST or anti-S-AKAP84 antibodies. 35S-labeledS-AKAP84-GFP fusions were in vitro synthesized byusing TnT quick-coupled transcription/translationsystem (Promega) in presence of 45 mCi of [35S]methio-nine. Where indicated, the filter was probed with an RIIoverlay before Western analysis.
Binding affinity
Full-length AKAP121 cDNA was subcloned in pMALvector (New England Biolabs). Recombinant AKAP121fused to maltose-binding domain was purifiedextensively by maltose affinity chromatography and32P-labeled in vitro with PKA catalytic subunit. Optimalbinding to GST-b tubulin was determined by incubatingdifferent concentrations of AKAP121 with an excess ofGST-tubulin. Binding of [32P]AKAP121 to GST-tubulinwas competed with increasing concentrations ofunlabeled AKAP121. Unbound [32P]AKAP121 waswashed off with binding buffer (see above). Bindingwas detected by autoradiography and measured bydensitometry.
Assay for RII-binding activity
Protein samples were size-fractionated by SDS-PAGE(8% polyacrylamide gel) as described above. Resolvedpolypeptides were transferred to polyvinylidene difluor-ide membranes (Immobilon P, Millipore). RII probeswere labeled with [g-32P]ATP and PKA catalytic subunit(one unit) (Sigma), as described.24
Immunoprecipitation and immunoblot analysis
Cells or rat tissues were homogenized in lysis buffer(20 mM Tris–HCl (pH 7.4), 0.15 M NaCl, 10 mM EDTA,0.25% Triton X-100, 0.05% (v/v) Tween-20, 0.02% (w/v)sodium azide) containing aprotinin (5 mg/ml), leupeptin(10 mg/ml), pepstatin (2 mg/ml) and 0.5 mM phenyl-methylsulfonyl fluoride (PMSF). The lysates werecleared by centrifugation at 15,000g for 15 minutes. Totallysate (1 mg) was subjected to immunoprecipitationwith the indicated antibodies. Total proteins (100 mg) orthe immunoprecipitates were resolved by SDS-PAGEand transferred to Immobilon P membrane. The filters
were rinsed in T-PBS (phosphate-buffered saline (PBS);10 mM Na2HPO4, 1.8 mM KH2PO4 (pH 7.4), 137 mMNaCl, 2.6 mM KCl), containing 0.1% Tween-20 andblocked for one hour at room temperature in 5% (w/v)non-fat dry milk. Blots were washed with T-PBS andimmunoblotted in the same buffer using specific anti-bodies. Where indicated, anti-Ha-Ras antibodies wereused at 1/1000 dilution (Upstate BiotechnologyIncorporation). The filters were washed three times inT-PBS for 30 minutes and incubated with peroxidase-conjugated anti-rabbit or anti-mouse IgG in T-PBS forone hour. The filters were washed and developed withECL reagents (Amersham). The films were analyzed byscanning densitometry (Molecular Dynamics). Mono-clonal antibodies directed versus b tubulin (Sigma) werediluted at 1/200 for immunoprecipitation and 1/5000for immunoblot analysis. Anti-RIIa and anti-S-AKAP84polyclonal antibodies were produced by immunizinganimals with the NH2 terminus of the protein. Since thisregion is conserved in AKAP121, the antibodies recog-nize both S-AKAP84 and AKAP121.25 Anti-RII and anti-S-AKAP84 polyclonal antibodies were used for immuno-precipitation (1/200) and immunoblot (1/1000) analyses.Total proteins from yeast strain (YRG2) were extracted asdescribed.31 Polyclonal antibodies directed against thebinding domain of GAL4 were purchased from SantaCruz Biotechonolgy.
In vitro S-AKAP84-tubulin co-polymerization assay
Cycles of cellular tubulin polymerization/depolymeri-zation were carried out as described.49 Briefly, rat testismaterial was homogenized in lysis buffer (100 mMPipes–NaOH (pH 6.9), 1 mM EGTA, 1 mM MgCl2, 20%(v/v) glycerol, 1 mM DTT). The homogenates werecentrifuged at 6500g for 15 minutes at 4 8C. The super-natant was decanted and centrifuged for 75 minutes at96,000g. The supernatant was decanted and incubatedfor 30 minutes at 37 8C in the presence of 1 mM GTP(polymerization cycle). Polymerized microtubules (WP,warm pellet) were sedimented by centrifugation at96,000g for 60 minutes at 37 8C and the supernatantdecanted (WS, warm supernatant). The WP fraction wasresuspended in lysis buffer and incubated on ice for 30minutes (depolymerization cycle). Polymerization/depolymerization cycles were repeated several times.Fractions of WP and WS (50 mg) were resolved by SDS-PAGE (10% polyacrylamide gel), transferred topolyvinylidene difluoride membranes and immuno-blotted with anti-tubulin, anti-S-AKAP84 or anti H-Rasantibodies.
Immunofluorescence analysis
Transiently transfected or microinjected GC2 cellswere rinsed in PBS and fixed in 3.7% (v/v) formalde-hyde for 20 minutes. After permeabilization with 0.5%Triton X-100 in PBS for five minutes, the cells were incu-bated with PBS, 0.1 mg/ml of bovine serum albumin forone hour at room temperature. Double immunofluores-cence was carried out with the following antibodies:anti-SOD monoclonal antibody (Walter Occhiena,1/200), anti-a/b tubulin monoclonal antibody (Sigma,1/200); anti-calreticulin polyclonal antibody (Stressgene,1/200) anti-S-AKAP84 (1/200) and RIIa (1/200) poly-clonal antibodies. Fluorescein or rhodamine-tagged anti-rabbit and anti-mouse IgG (Technogenetics) secondaryantibodies were used. Coverslips were analyzed by
A-Kinase Anchor Protein 84/121 673

Zeiss Axiomat microscope, as described.21 Where indi-cated, nocodazole was added (5 mM for 90 minutes) tothe medium of growing cells. Cells were synchronizedat mitosis by first arresting them in S phase by twosequential thymidine blocks (2 mM for 16 hours). Thecells were then released from S phase by removing thedrug and harvested at different times (four to six hours)after release. Cell-cycle progression was monitored byfluorescence-activated cell sorter analysis (FACS), asdescribed.21 Chromatin morphology was analyzed byincubating the fixed cells with Hoechst 33342 for fiveminutes at room temperature followed by three washeswith PBS.
Acknowledgments
The work was supported by grants from “AssociazioneItaliana per la Ricerca sul Cancro” (A.I.R.C.); TargetedProject Biotechnology CNR; and MURST (Italian Depart-ment of University and Research); National Institute ofHealth (grant #DK07328). Thanks to Dr C. Laezza forhelp in tubulin polymerization experiments. We grate-fully acknowledge Professor C. Pietropaolo, andDr G. Russo for helpful suggestions concerning themicroinjection experiments.
References
1. Edelman, A. M., Blumenthal, D. K. & Krebs, E. G.(1987). Protein serine/threonine kinases. Annu. Rev.Biochem. 56, 567–613.
2. Taylor, S., Buechler, J. A. & Yonemoto, Y. (1990).Structural framework for the protein kinase family.Annu. Rev. Biochem. 59, 971–1005.
3. Rubin, C. S. (1994). A kinase anchor proteins and theintracellular targeting of signals carried by cyclicAMP. Biochim. Biophys. Acta, 1224, 467–479.
4. Smith, F. D. & Scott, J. D. (2002). Signaling com-plexes: junctions on the intracellular informationsuper highway. Curr. Biol. 12, R32–R40.
5. Diviani, D. & Scott, J. D. (2001). AKAP signalingcomplexes at the cytoskeleton. J. Cell Sci. 114,1431–1437.
6. Feliciello, A., Gottesman, M. E. & Avvedimento, V. E.(2001). The biological function of A-kinase anchorproteins. J. Mol. Biol. 308, 99–114.
7. Miki, K. & Eddy, E. M. (1999). Single amino acidsdetermine specificity of binding of protein kinase Aregulatory subunits by protein kinase A anchoringproteins. J. Biol. Chem. 274, 29057–29062.
8. Angelo, R. G. & Rubin, C. S. (2000). Characterizationof structural features that mediate the tethering ofCaenorhabditis elegans protein kinase A to a novelA kinase anchor protein. Insights into the anchoringof PKAI isoforms. J. Biol. Chem. 275, 4351–4362.
9. Coghlan, V. M., Perrino, B. A., Howard, M., Lange-berg, L. K., Hicks, J. B., Gallatin, W. M. & Scott, J. D.(1995). Association of protein kinase A and proteinphosphatase 2B with a common anchoring protein.Science, 267, 108–111.
10. Klauck, T. M., Faux, M. C., Labudda, K., Langeberg,L. K., Jaken, S. & Scott, J. D. (1996). Coordination ofthree signaling enzymes by AKAP79, a mammalianscaffold protein. Science, 271, 1589–1592.
11. Schillace, R. V. & Scott, J. D. (1999). Association of thetype 1 protein phosphatase PP1 with the A-kinaseanchoring protein AKAP220. Curr. Biol. 9, 321–324.
12. Westphal, R. S., Tavalin, S. J., Lin, J. W., Alto, N. M.,Fraser, I. D., Langeberg, L. K. et al. (1999). Regulationof NMDA receptors by an associated phosphatase-kinase signaling complex. Science, 285, 93–96.
13. Steen, R. L., Martins, S. B., Tasken, K. & Collas, P.(2000). Recruitment of protein phosphatase 1 to thenuclear envelope by A-kinase anchoring proteinAKAP149 is a prerequisite for nuclear laminaassembly. J. Cell Biol. 150, 1251–1262.
14. Feliciello, A., Giuliano, P., Porcellini, A., Garbi, C.,Obici, S., Mele, E. et al. (1996). The v-Ki-Ras oncogenealters cAMP nuclear signaling by regulating thelocation and the expression of cAMP-dependentprotein kinase IIbeta. J. Biol. Chem. 271, 25350–25359.
15. Feliciello, A., Avvedimento, V. E., Li, Y., Gottesman,M., E, . & Rubin, C. S. (1997). A-kinase anchorprotein 75 increases the rate and magnitude ofcAMP signaling to the nucleus. Curr. Biol. 7,1011–1014.
16. Lester, L. B., Langeberg, L. K. & Scott, J. D. (1997).Anchoring of protein kinase A facilitates hormone-mediated insulin secretion. Proc. Natl Acad. Sci.USA, 94, 14942–14947.
17. Gray, P. C., Scott, J. D. & Catterall, W. A. (1998).Regulation of ion channels by cAMP-dependentprotein kinase and A-kinase anchoring proteins.Curr. Opin. Neurobiol. 8, 330–334.
18. Paolillo, M., Feliciello, A., Porcellini, A., Garbi, C.,Bifulco, M., Schinelli, S. et al. (1999). The type andthe localization of cAMP-dependent protein kinaseregulate transmission of cAMP signals to the nucleusin cortical and cerebellar granule cells. J. Biol. Chem.274, 6546–6552.
19. Westphal, R. S., Soderling, S. H., Alto, N. M.,Langeberg, L. K. & Scott, J. D. (2000). Scar/WAVE-1,a wiskott-aldrich syndrome protein, assembles anactin-associated multi-kinase scaffold. EMBO J. 19,4589–4600.
20. Colledge, M., Dean, R. A., Scott, G. K., Langeberg,L. K., Huganir, R. L. & Scott, J. D. (2000). Targetingof PKA to glutamate receptors through a MAGUK-AKAP complex. Neuron, 27, 107–119.
21. Feliciello, A., Gallo, A., Mele, E., Porcellini, A.,Troncone, G., Garbi, C. et al. (2000). The localizationand activity of cAMP-dependent protein kinaseaffect cell cycle progression in thyroid cells. J. Biol.Chem. 275, 303–311.
22. Lester, L. B., Faux, M. C., Nauert, J. B. & Scott, J. D.(2001). Targeted protein kinase A and PP-2B regulateinsulin secretion through reversible phosphorylation.Endocrinology, 142, 1218–1227.
23. Fink, M. A., Zakhary, D. R., Mackey, J. A., Desnoyer,R. W., Apperson-Hansen, C., Damron, D. S. & Bond,M. (2001). AKAP-mediated targeting of proteinkinase a regulates contractility in cardiac myocytes.Circ. Res. 88, 291–297.
24. Lin, R., Moss, S. B. & Rubin, C. S. (1995). Characteriz-ation of S-AKAP84, a novel developmentally regu-lated A kinase anchor protein of male germ cells.J. Biol. Chem. 270, 27804–27811.
25. Chen, Q., Lin, R. & Rubin, C. S. (1997). Organelle-specific targeting of protein kinase AII (PKAII).Molecular and in situ characterization of murine Akinase anchor proteins that recruit regulatorysubunits of PKAII to the cytoplasmic surface ofmitochondria. J. Biol. Chem. 272, 15247–15257.
674 A-Kinase Anchor Protein 84/121

26. Trendelenburg, G., Hummel, M., Riecken, E. O. &Hanski, C. (1996). Molecular characterization ofAKAP149, a novel A kinase anchor protein with aKH domain. Biochem. Biophys. Res. Commun. 225,313–319.
27. Huang, L. J., Durick, K., Weiner, J. A., Chun, J. &Taylor, S. S. (1997). Identification of a novel proteinkinase A anchoring protein that binds both type Iand type II regulatory subunits. J. Biol. Chem. 272,8057–8064.
28. Feliciello, A., Rubin, C. S., Avvedimento, E. V. &Gottesman, M. E. (1998). Expression of A kinaseanchor protein121 is regulated by hormones inthyroid and testicular germ cells. J. Biol. Chem. 273,23361–23366.
29. Huang, L. J., Wang, L., Ma, Y., Durick, K., Perkins, G.,Deerinck, T. J. et al. (1999). NH2-Terminal targetingmotifs direct dual specificity A-kinase-anchoringprotein 1 (D-AKAP1) to either mitochondria orendoplasmic reticulum. J. Cell Biol. 145, 951–959.
30. Harada, H., Becknell, B., Wilm, M., Mann, M.,Huang, L. J., Taylor, S. S. et al. (1999). Phosphoryl-ation and inactivation of BAD by mitochondria-anchored protein kinase A. Mol. Cell, 4, 413–422.
31. Feliciello, A., Cardone, L., Garbi, C., Ginsberg, M. D.,Varrone, S., Rubin, C. S. et al. (1999). Yotiao protein, aligand for the NMDA receptor, binds and targetscAMP-dependent protein kinase II. FEBS Letters,464, 174–178.
32. Kelley, L. A., MacCallum, R. M. & Sternberg, M. J. E.(2000). Enhanced genome annotation using struc-tural profiles in the program 3D-PSSM. J. Mol. Biol.299, 499–520.
33. Hahne, K., Haucke, V., Ramage, L. & Schatz, G.(1994). Incomplete arrest in the outer membranesorts NADH-cytochrome b5 reductase to two dif-ferent submitochondrial compartments. Cell, 79,829–839.
34. Weisiger, R. A. & Fridovich, I. (1973). Mitochondrialsuperoxide dismutase. Site of synthesis and intra-mitochondrial localization. J. Biol. Chem. 24,4793–4796.
35. Yaffe, M. (1999). The machinery of mitochondrialinheritance and behavior. Science, 283, 1493–1497.
36. Straight, A. F. & Field, C. M. (2000). Microtubules,membranes and cytokinesis. Curr. Biol. 10,R760–R770.
37. Piel, M., Nordberg, J., Euteneuer, U. & Bornens, M.(2001). Centrosome-dependent exit of cytokinesis inanimal cells. Science, 291, 1550–1553.
38. Nogales, E. (2000). Structural insights into micro-tubule function. Annu. Rev. Biochem. 69, 277–302.
39. Downing, K. H. & Nogales, E. (1998). New insightsinto microtubule structure and function from theatomic model of tubulin. Eur. Biophys. J. 27, 431–436.
40. Rubino, H. M., Dammerman, M., Shafit-Zagardo, B.& Erlichman, J. (1989). Localization and characteri-zation of the binding site for the regulatory subunitof type II cAMP-dependent protein kinase onMAP2. Neuron, 3, 631–638.
41. Vallee, R. B., DiBartolomeis, M. J. & Theurkauf, W. E.(1981). A protein kinase bound to the projectionportion of MAP 2 (microtubule-associated protein2). J. Cell Biol. 90, 568–576.
42. Diviani, D., Langeberg, L. K., Doxsey, S. J. & Scott,J. D. (2000). Pericentrin anchors protein kinase A atthe centrosome through a newly identified RII-bind-ing domain. Curr. Biol. 10, 417–420.
43. Bernier-Valentin, F. & Rousset, B. (1982). Interactionof tubulin with rat liver mitochondria. J. Biol. Chem.257, 7092–7099.
44. Jung, D., Filliol, D., Miehe, M. & Rendon, A. (1993).Interaction of brain mitochondria with microtubulesreconstituted from brain tubulin and MAP2 or TAU.Cell Motil. Cytoskel. 24, 245–255.
45. Langford, G. (1995). Actin- and microtubule-depen-dent organelle motors: interrelationships betweenthe two motility systems. Curr. Opin. Cell. Biol. 7,82–88.
46. Bernier-Valentin, F., Aunis, D. & Rousset, B. (1983).Evidence for tubulin-binding sites on cellularmembranes: plasma membranes, mitochondrialmembranes, and secretory granule membranes..J. Cell Biol. 97, 209–216.
47. Wolkolwicz, M. J., Coonrod, S. M., Reddi, P. P.,Millan, J. C., Hofmann, M. C. & Herr, J. R. (1996).Refinement of the differentiated phenotype of thespermatogenic cell line GC-2spd(ts). Biol. Reprod. 55,923–932.
48. Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989).Molecular Cloning: A Laboratory Manual, 2nd edit.,Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
49. Williams, R. C. & Lee, J. C. (1982). Preparation oftubulin from brain. Methods Enzymol. 85, 376–408.
Edited by M. Yaniv
(Received 6 February 2002; received in revised form 24 April 2002; accepted 13 May 2002)
A-Kinase Anchor Protein 84/121 675





























