Embed Size (px)
Citation preview

A Mollusk Retinoic Acid Receptor (RAR) OrthologSheds Light on the Evolution of Ligand Binding
Juliana Gutierrez-Mazariegos, Eswar Kumar Nadendla, Daniela Lima,Keely Pierzchalski, Jace W. Jones, Maureen Kane, Jun-Ichi Nishikawa,Youhei Hiromori, Tsuyoshi Nakanishi, Miguel M. Santos, L. Filipe C. Castro,William Bourguet, Michael Schubert,* and Vincent Laudet*
Molecular Zoology Team (J.G.-M., V.L.), Institut de Génomique Fonctionnelle de Lyon, Unité Mixte deRecherche 5242, Université Lyon 1, Centre National de la Recherche Scientifique, Ecole NormaleSupérieure de Lyon, 69364 Lyon Cedex 07, France; Institut National de la Santé et de la RechercheMédicale Unité 1054 (E.K.N., W.B.), Centre de Biochimie Structurale, Centre National de la RechercheScientifique Unité Mixte de Recherche 5048, Universités Montpellier 1 and 2, 34967 Montpellier, France;CAS in Crystallography and Biophysics (E.K.N.), University of Madras, 600-005 Chennai, India; Centre ofMarine and Environmental Research/Interdisciplinary Centre of Marine and Environmental Research (D.L.,M.M.S., L.F.C.C.), FCUP–Department of Biology, Faculty of Sciences, University of Porto, 4050-123 Porto,Portugal; Department of Pharmaceutical Sciences (K.P., J.W.J., M.K.), School of Pharmacy, University ofMaryland, Baltimore, Maryland 21201; Laboratory of Health Sciences (J.-I.N.), School of Pharmacy andPharmaceutical Sciences, Mukogawa Women’s University, Koshien, Nishinomiya, Hyogo 663-8179, Japan;Laboratory of Hygienic Chemistry and Molecular Toxicology (Y.H., T.N.), Gifu Pharmaceutical University, Gifu501-1196, Japan; and Laboratoire de Biologie du Développement de Villefranche-sur-Mer, Unité Mixte deRecherche 7009, Sorbonne Universités, Université Pierre et Marie Curie Paris 06, Centre National de laRecherche Scientifique, Observatoire Océanologique de Villefranche-sur-Mer, 06230 Villefranche-sur-Mer,France
Nuclear receptors are transcription factors that regulate networks of target genes in response to smallmolecules. There is a strong bias in our knowledge of these receptors because they were mainlycharacterized in classical model organisms, mostly vertebrates. Therefore, the evolutionary origins ofspecific ligand-receptor couples still remain elusive. Here we present the identification and character-ization of a retinoic acid receptor (RAR) from the mollusk Nucella lapillus (NlRAR). We show that thisreceptor specifically binds to DNA response elements organized in direct repeats as a heterodimer withretinoid X receptor. Surprisingly, we also find that NlRAR does not bind all-trans retinoic acid or anyother retinoid we tested. Furthermore, NlRAR is unable to activate the transcription of reporter genesin response to stimulation by retinoids and to recruit coactivators in the presence of these compounds.Three-dimensional modeling of the ligand-binding domain of NlRAR reveals an overall structure thatis similar to vertebrate RARs. However, in the ligand-binding pocket (LBP) of the mollusk receptor, thealterationofseveral residues interactingwiththeligandhasapparently ledtoanoveralldecreaseinthestrength of the interaction with the ligand. Accordingly, mutations of NlRAR at key positions withinthe LBP generate receptors that are responsive to retinoids. Altogether our data suggest that, inmollusks, RAR has lost its affinity for all-trans retinoic acid, highlighting the evolutionary plasticity ofits LBP. When put in an evolutionary context, our results reveal new structural and functional featuresof nuclear receptors validated by millions of years of evolution that were impossible to reveal in modelorganisms. (Endocrinology 155: 4275–4286, 2014)
ISSN Print 0013-7227 ISSN Online 1945-7170Printed in U.S.A.Copyright © 2014 by the Endocrine SocietyReceived March 3, 2014. Accepted August 6, 2014.First Published Online August 13, 2014
* M.S. and V.L. contributed equally to this work.Abbreviations: ATRA, all-trans RA; 9cRA, 9-cis RA; 13cRA, 13-cis RA; 3D, three-dimen-sional; DBD, DNA-binding domain; DR, direct repeat; ER, estrogen receptor; HEK, humanembryonic kidney; LBD, ligand-binding domain; LBP, ligand-binding pocket; LPA, limitedproteolysis assay; NlRAR, RAR from the mollusk Nucella lapillus; NlRXR, RXR from themollusk Nucella lapillus; NR, nuclear receptor; RA, retinoic acid; RAR, retinoic acid receptor;RARE, retinoic acid response element; RXR, retinoid X receptor; T-box, terminal box; TR,thyroid hormone receptor.
G E N E R A L E N D O C R I N O L O G Y
doi: 10.1210/en.2014-1181 Endocrinology, November 2014, 155(11):4275–4286 endo.endojournals.org 4275
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

Among all transcription factors, nuclear receptors(NRs) are unusual in that their ability to modulate
gene transcription is regulated by the binding of a ligand(1). We have a good understanding of how these receptorsare able to regulate transcription after ligand binding.However, our view of NR action is strongly biased to-wards vertebrate models and towards mammals in par-ticular. Therefore, key questions on the evolutionary or-igin of NRs and of their ligands still remain elusive. Inparticular, the evolutionary elaboration of specific ligand-receptor couples is still unclear (2, 3). Several observationshave suggested that, in contrast to what was initially ex-pected, a given ligand-receptor couple may not be stable inevolutionary time and that changes in ligand binding spec-ificity was quite frequent during animal diversification (4).Estrogen receptor (ER) orthologs from several protostomespecies, for example, have been shown to be insensitive toestradiol, as is the case for mollusk ERs (5, 6). In addition,the presence of the bona fide vertebrate ER ligand, 17�-estradiol, in nonchordate species has been questioned (7–9). Similarly, thyroid hormone receptors (TRs), present inbasal chordates, have been shown to recognize a differentligand than vertebrate TRs (5, 10). These examples clearlysuggest that the presence of the ortholog of a given NRgene does not necessarily imply similar molecular and bio-chemical functions.
Retinoic acid (RA) is a morphogen derived from vita-min A that controls key processes during vertebrate de-velopment. In particular, it controls the patterning of theanteroposterior axis and the differentiation of various celltypes, such as neurons or hematopoietic cells. RA is syn-thesized from retinol and can be found in the form of threeprincipal isomers, all-trans RA (ATRA), which is the ma-jor biologically active compound in vertebrates, 13-cis RA(13cRA), and 9-cis RA (9cRA), which has been proposedas a ligand for retinoid X receptor (RXR), but whose invivo presence is still debated (11). Like other NRs, retinoicacid receptor (RAR) and RXR proteins consist of twomain domains, the DNA-binding domain (DBD) and theligand-binding domain (LBD). RAR and RXR regulate thetranscription of their target genes by forming het-erodimers that bind specific response elements called reti-noic acid response element (RAREs) in the regulatory re-gions of target genes (12, 13). These elements consists oftwo copies of the consensus sequence (A/G)G(G/T)(G/T)(G/C)A organized as direct repeats (DRs) and separatedby a variable number of nucleotides (12, 13). In verte-brates, RAR/RXR heterodimers recognize DR1 (one nu-cleotide spacer), DR2, and DR5 elements (14–17),whereas the RXR homodimer recognizes almost exclu-sively DR1 (16). Recent genome-wide analyses of RAREsusing chromatin immunoprecipitation followed by high-
throughout sequencing have revealed that other elements,such as DR8 sequences, are also recognized by RAR/RXR(15).
Little is known about RA signaling outside of chor-dates, and evidence for biological functions of RA in non-chordate invertebrates is scarce (18). However, some stud-ies have shown that RA might have a physiological role inmollusks. In Lymnaea stagnalis, both ATRA and 9cRAare involved in neuronal differentiation (19, 20). In Thaisclavigera and Nucella lapillus, the injection of 9cRAcauses abnormalities in sex organ development (21, 22).Moreover, in several species, treatment of embryos withATRA affects eye formation (23). However, these resultsare based on treatments with exogenous RA and do notprove that endogenous retinoids are indeed present andactive in these animals. There is some evidence of the pres-ence of endogenous retinoids, such as ATRA and 9cRA, inmollusks (20, 24, 25). In addition, it was demonstratedthat an RXR from the mollusk Biomphalaria glabrata canbind 9cRA and activate transcription in response to thiscompound (26). Therefore, these data suggests that RAmight have a biological function in mollusks.
Using an in silico approach, key elements of the RAmachinery, such as RAR and RXR, have been identified inthe genomes of nonchordate species, including lophotro-chozoans, indicating that the RA signaling pathway maybe present and functional in these species (18). For exam-ple, rar genes have been identified in ambulacrarians, he-michordates, annelids, and mollusks (27–31). Further-more, a rar from the mollusk T. clavigera was recentlycloned and characterized. However, the protein encodedby this gene was unable to activate transcription in thepresence of RA (32). The available data therefore suggest,but do not unambiguously prove, that RA signaling maybe active in nonchordates, such as mollusks. Therefore,detailed characterizations of lophotrochozoan RARs areneeded to gain insights into the evolutionary origin of ret-inoid signaling in metazoans.
Here we describe a novel RAR from the gastropodmollusk N. lapillus. We show that this N. lapillus RAR(NlRAR) is able to heterodimerize with N. lapillus RXR(NlRXR) and bind specific DNA sequences based on theclassical direct repeat RARE consensus. Ligand-bindingand transactivation assays reveal that NlRAR does notbind to and is not activated by ATRA or other retinoids,despite the presence of these compounds in N. lapillustissues. Analysis of the ligand-binding pocket of NlRARrevealed a series of amino acid changes that may explaina reduction of its affinity for the bona fide vertebrate li-gands. Indeed, the mutation of these positions in theNlRAR LBD retrieves ATRA and 9cRA binding and ac-tivation of the mutated receptor. Taken together, our re-
4276 Gutierrez-Mazariegos et al Evolution of Retinoid Receptors Endocrinology, November 2014, 155(11):4275–4286
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

sults show that, even though N. lapillus possesses the ma-chinery necessary for RA signaling, this signaling pathwaymight be functional in a different manner in mollusks,when compared with vertebrates or even other lophotro-chozoans. This finding thus highlights the plasticity of theLBD in terms of ligand binding and reveals an unexpectedflexibility of RARs to respond to novel physiologicaland/or biochemical contexts.
Materials and Methods
Cloning of N. lapillus rar and plasmid constructsTotal RNA was extracted from N. lapillus prehatched juve-
niles and adult tissues and reverse transcribed. A N. lapillus rarcDNA, 1101 bp long encoding a protein of 367 amino acids, wascloned using a PCR strategy (Ref. 22 and Supplemental Mate-rials). Genes encoding the full-length receptors (RAR and RXR)were cloned into the pSG5 vector and a flag tag was added at theN terminus. The LBDs of the receptors were cloned intopG4MpolyII (33) to produce GAL4-LBD fusions. Full-lengthRAR and RXR protein sequences from different species (see Sup-plemental Materials) were aligned with Seaview (34), and thetree was calculated using PhyML with the JTT model and opti-mized invariable sites and 1000 bootstrap replicates (34). Gapswere treated as unknown characters.
Electrophoretic mobility shift assays (EMSAs)In vitro translation of tagged NlRAR and NlRXR labeled
with 35S-methionine was performed using the TNT wheat germextract system (Promega). The EMSAs were performed as pre-viously described (26) and details are listed in the SupplementalMaterials.
Ligands and peptidesATRA, 9cRA, 13cRA, and TTNPB were purchased from Sig-
ma-Aldrich. Stock solutions were prepared in ethanol at 10�2 M.The fluorescent peptides NCOR ID1, NCOR ID2, and SRC-1NR2 were purchased from EZBiolab. The fluorescent SMRTID1 peptide was obtained from Neosystem, and the fluorescentSMRT ID2 peptide was synthesized by Dr. J. F. Guichou (CentreNational de la Recherche Scientifique, Unité Mixte de Recherche5048, Montpellier, France). Their sequences are listed in theSupplemental Materials.
Transactivation assays in human embryonic kidney(HEK) 293T cells
HEK 293T cells were maintained in DMEM supplementedwith 10% fetal calf serum (Invitrogen). Transfection and treat-ments were performed as previously described (35) with 60 ng oftotal DNA, and the final concentration for the ligands were be-tween 10�7 and 10�9 M.
Limited proteolysis assays (LPAs)LPAs were performed as previously described (35) using the
receptors translated in vitro using the TNT-coupled reticulocytelysate system (Promega) and labeled with 35S-methionine. The
final concentrations tested for each ligand were between 10�5
and 10�7 M.
Ligand-binding assaysThe LBD of the human HsRAR� and NlRAR were cloned
into pGEM-4T (Amersham Biosciences), containing a gluta-thione-S-transferase tag. The fusion proteins were purified, andeach protein was then incubated with 10 nM all-trans-[11,12-3H] RA (PerkinElmer) with increasing concentrations of non-labeled ATRA (from 1 to 1000 nM). After an incubation at 4°Cfor 1 hour, hydroxyapatite was added to each tube. Hydroxy-apatite-bound radioactivity was determined by liquid scintilla-tion counting.
Fluorescence anisotropy measurementsThe NlRAR LBD was cloned into the pET-32a vector. Ex-
pression and purification of the LBDs of NlRAR and of the hu-man HsRAR� were performed as previously described (36). Flo-rescence anisotropy assays were performed using a Safire2microplate reader (TECAN) as described in (36).
Three-dimensional (3D) modeling of RAR LBDs andmutagenesis
The NlRAR model was constructed using the modeling me-taserver @TOME 2 (37) and the crystal structure of the LBD ofthe human RAR� (3KMR) as a template. Mutants were con-structed by PCR-assisted, site-directed mutagenesis using the PfuDNA polymerase (Promega). The DpnI enzyme (New EnglandBiolabs) was used to remove the parental DNA template.
Tissue retinoid determinationTissues were stored at �80°C until assayed. They were ho-
mogenized and extracted as previously described (38). RA iso-mers were quantified using liquid chromatography-tandem massspectrometry on an AB Sciex 5500 QTRAP in MRM mode usingAPCI in positive ion mode as previously described (38, 39). Ret-inol and retinyl esters (REs) were quantified by high-perfor-mance liquid chromatography with ultraviolet detection on aWaters Acquity as previously described (38, 40).
Results
Identification of a RAR ortholog in the molluskN. lapillus
We cloned a short fragment of a gene encoding RARfrom N. lapillus using a RT-PCR strategy (GenBank ac-cession number KJ410131). This rar clone contains anopen reading frame of 1101 bp that encodes a 367-aminoacid-long protein, called NlRAR. The predicted NlRARprotein is characterized by the overall domain structure ofNRs: a DBD with conserved zinc finger motifs importantfor DNA binding and a LBD that contains the main fea-tures present in other RARs implicated in ligand binding,dimerization with RXR, and recognition of transcrip-tional coactivators and corepressors (Supplemental Figure1). Amino acid alignments indicate that the NlRAR DBD
doi: 10.1210/en.2014-1181 endo.endojournals.org 4277
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

shares approximately 81%–83% sequence similaritywith chordate RARs and 93% with the DBD of the re-cently identified RAR from the mollusk T. clavigera. Con-cerning the LBD, the identity levels are 51%–54% withthe chordate RARs and 68% with T. clavigera RAR. In aphylogenetic tree, the NlRAR sequence is placed close tothe T. clavigera RAR and is associated with another pro-tostome RAR from the annelid Capitella teleta (Figure 1).As expected, these protostome RARs are an early offshootof the RAR branch, clearly suggesting that a unique RARortholog was present in the last common ancestor of allbilaterian animals.
It is interesting to compare the conservation of specificsequence features between NlRAR and other RARs (Sup-plemental Figure 1). Within the DBD, the proximal box,which is involved in determining the DNA binding spec-ificity (41), and the terminal box (T-box), which is re-quired for the heterodimerization with RXR on DR2 el-ements (42, 43), are 100% conserved, suggesting that theNlRAR recognizes the classical RARE core sequences as aheterodimeric binding partner with RXR. In contrast, thedistal box, which plays a role in the heterodimerization ofRAR and RXR on DR5 elements (44), is more divergent(Supplemental Figure 1).
The NlRAR also shares the general NR organizationwith 12 helices within the LBD (45). However, the NlRAR
LBD is characterized by a significant amount of aminoacid differences compared with the chordate RARs. Forexample, of the 24 amino acids that have been shown tointeract with ATRA in human RAR� (Supplemental Fig-ure 1) (46), seven are different in NlRAR. Among thesedivergent residues, four are conserved between the RARsfrom T. clavigera and N. lapillus, suggesting that this se-quence signature may be a conserved feature of mollusks.Nonetheless, this low degree of sequence conservation ofkey residues in the ligand-binding pocket (LBP) betweenmollusk and chordate RARs suggests that the ligand bind-ing properties of mollusk RARs, and thus of NlRAR,might be different from those of chordates RARs.
NlRAR and NlRXR are able to recognize specificDNA sequences
To characterize the DNA binding properties of NlRAR,we performed EMSA experiments using as a partner thepreviously described N. lapillus RXR (22) (Figure 2). ThisRXR gene encodes two isoforms, NlRXRa and NlRXRb,which differ by a small insertion in the T-box of NlRXRb.Because NlRXRa is the major isoform, we used this iso-form for our experiments.
Binding was assessed on consensus DR elements as de-fined in vertebrates (Figure 2 and Supplemental Figure 2).Using a radiolabeled DR2 probe, no binding of NlRAR
Figure 1. Phylogenetic position of NlRAR. Phylogenetic tree of the RAR cloned from N. lapillus. Numbers on each branch are bootstrap supportvalues in percentage of 1000 replicates. The tree was rooted with the RXR sequences. The species listed are the following: Branchiostoma floridae,Biomphalaria glabrata, Capitella teleta, Ciona intestinalis, Danio rerio, Homo sapiens, Nucella lapillus, Strongylocentrotus purpuratus, Thaisclavigera, and Xenopus tropicalis.
4278 Gutierrez-Mazariegos et al Evolution of Retinoid Receptors Endocrinology, November 2014, 155(11):4275–4286
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

alone could be detected (Figure 2). In contrast, whenNlRXRa was added, the NlRAR/NlRXRa heterodimerstrongly binds the DR2 element (Figure 2). This binding tothe labeled DR2 probe is lost in presence of a large excess(10- or 100-fold) of unlabeled DR1, DR2, and DR5 (Fig-ure 2). In contrast, the use of unlabeled DR3 or DR4 aswell as a nonspecific unlabeled probe added in excess wasnot able to compete for binding (Figure 2). Similar resultswere obtained, with a radiolabeled DR5 probe (Supple-mental Figure 2A). Therefore, we conclude that theNlRAR/NlRXRa heterodimer is able to bind specific se-quences of DNA, in particular DR1, DR2, or DR5 ele-ments, but not DR3 or DR4 sequences. As shown in Sup-plemental Figure 2B, NlRXRa is able to bind as ahomodimer to DR1 elements. In contrast, the NlRXRbisoform shows impaired DNA binding both as a ho-modimer and as a heterodimer with NlRAR. Interestingly,in T. clavigera, similar RXR isoforms have been observed,and it has been shown that the T-box insertion reduces thetranscriptional activity of the receptor (47).
Taken together, these results show that N. lapillus pos-sesses a RAR/RXR heterodimer that recognizes a set ofRAREs very similar to the ones recognized by chordateRAR/RXR heterodimers.
NlRAR does not bind ATRA and other retinoids,but NlRXR binds 9cRA
To obtain insights into the ligand binding properties ofthe RAR and RXR from N. lapillus, we tested the ability
of both NlRAR and NlRXR to bind to different retinoidsand to activate transcription in a ligand-dependent man-ner (Figure 3 and Supplemental Figures 3–5). Initially, theability of NlRAR to activate transcription in mammaliancells was assessed using transient transfection with GAL4-LBD-RAR fusion constructs. We observed that NlRARwas not able to activate the transcription of the luciferasereporter with any of the ligands tested (ATRA, 9cRA,13cRA, retinol, and retinal) at doses varying from 10�7 to10�5 M (Figure 3A and Supplemental Figure 3A, data notshown). In contrast, all of these compounds stimulated thetranscriptional activity of human RAR� (Figure 3A andSupplemental Figure 3A).
This inability of NlRAR to activate transcription, in thepresence of a ligand, may be due to the fact that we testeda mollusk protein in mammalian cells, even if the regionsof the LBD implicated in coactivator binding are relativelywell-conserved in NlRAR. We nonetheless tested the di-rect binding of different ligands to NlRAR using LPAs.This method allows the in vitro assessment of the confor-mational change after the ligand binding to the receptor(35, 48). Using this technique, we observed that the LBDof NlRAR was not protected from proteolysis by ATRA(at final concentrations ranging from 10�7 to 10�5 M) orby any other retinoid tested (9cRA, 13cRA, retinol, andretinal) (Figure 3B and Supplemental Figure 3B).
To further confirm these results, we performed a com-petitive binding assay using 3H-labeled ATRA and in-creasing concentrations of unlabeled ligand. We observedthat the addition of increasing concentrations of unlabeledATRA decreased the human RAR�-bound radioactivity(Figure 3C). The unlabeled competitor displaced 50% ofthe binding at a concentration of approximately 10 nM,which is in general agreement with the known affinity ofRAR� for ATRA. In contrast, NlRAR yielded only resid-ual radioactivity levels, and the addition of increasing con-centrations of unlabeled ATRA produced no reduction inradioactivity, suggesting that the weak binding observedin this assay is nonspecific (Figure 3C).
In addition, we investigated the ability of NlRAR torecruit coactivators by analyzing the titration of a fluo-rescein-labeled SRC-1 NR2 peptide by NlRAR in the pres-ence of different retinoids (ATRA, 9cRA, and 13cRA)(Figure 3, D and E). As expected, we observed that theaffinity of the human RAR� for the SRC-1 NR2 peptideincreases in the presence ATRA and 9cRA. When humanRAR� is analyzed in the presence of 13cRA, the receptordoes not recruit the SRC-1 NR2 peptide. For the molluskreceptor, we observed no binding of the SRC-1 NR2 pep-tide in the presence of any of the compounds tested (Figure3E). In contrast, we observed that the NlRAR LBD inter-acts efficiently with corepressor-derived peptides (Supple-
Figure 2. DNA recognition by NlRAR and NlRXRa. Binding of in vitrosynthesized NlRAR/NlRXRa heterodimers to a radioactive DR2 probe.Control lanes corresponding to unprogrammed reticulocytes (lane 1),NlRAR alone (lane 2), and NlRXR alone (lane 3). The NlRAR/NlRXRaheterodimers very strongly bind the radioactive DR2 probe (lane 4).Cold competitors correspond to 10-fold (�10) or 100-fold (�100)excess of unlabeled oligonucleotides (DR0 to DR5) (lanes 5–16). Anonspecific element (NS) was used as a negative control (lanes 17–18).
doi: 10.1210/en.2014-1181 endo.endojournals.org 4279
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

mental Figure 4A) and that none of the ligands tested isable to dissociate this interaction, suggesting that NlRARcould act as a constitutive repressor (Supplemental Figure4C). As expected, the dissociation of the corepressor pep-tides is observed with human RAR�, irrespective of whichligand is used (Supplemental Figure 4B). Because the N.lapillus RAR seems to behave like a transcriptional repres-sor, we tested whether this receptor has a competitive effecton transactivation mediated by a human RAR. As shown inSupplemental Figure 5, this is effectively the case: the intro-duction of increasing amounts of NlRAR shuts down theRA-dependent transactivation, mediated by human RAR, ofa RARE-containing reporter gene. Altogether, these resultsshow that NlRAR is not able to bind ATRA, as well as otherclassical retinoids, and that in the presence of these retinoids,NlRAR does not activate target gene transcription.
Contrasting the results we obtained for NlRAR, weobserved that, as expected, NlRXR is able to activate thetranscription of the luciferase reporter gene in a dose-de-pendent manner in the presence of 9cRA and other rexi-noids (Supplemental Figure 6A) (49, 50).
Specific amino acid differences are responsible forthe loss of NlRAR ligand binding
As shown above, the novel RAR identified in N. lapillusis unable to bind the different retinoids tested and to ac-tivate transcription in response to these compounds.Given that the sequence of the NlRAR LBD is globallyrelatively well conserved, with 52%–54% amino acididentity when compared with vertebrate RARs, which is inthe range of the conservation between vertebrate ER� andER�, both of which bind to and are activated by estrogens
Figure 3. NlRAR does not bind or activate transcription in the presence retinoids. A, The ability of NlRAR and human RAR� (HsRAR�) to activatetranscription of a luciferase reporter gene construct was tested in transfected HEK 293T cells in the presence of 10�5 M of ATRA, 9cRA, and13cRA. The GAL4 DNA binding domain alone was used as a negative control (Ctrl). B, The binding affinity of both receptors for ATRA, 9cRA, and13cRA was tested by limited proteolysis assays. Ligands were used at 10�5 M, and digested protein in the presence of ethanol was used as anegative control (lane �). Protected bands are indicated by arrows. C, Competitive binding assay with 10 nM 3H-labeled ATRA and increasingconcentrations of unlabeled ligand. The human RAR� (HsRAR�) was used as a positive control. D and E, Titration of fluorescein-labeled SRC-1 NR2peptide by the ligand-binding domains of human RAR� (HsRAR�) (D) and NlRAR (E) in the absence (apo) or presence (9cRA, ATRA, and 13cRA) ofdifferent ligands.
4280 Gutierrez-Mazariegos et al Evolution of Retinoid Receptors Endocrinology, November 2014, 155(11):4275–4286
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

(51), the incapacity of RA binding by NlRAR is very likelynot due to a general divergence of the LBD. This findingindicates that alterations of specific amino acids inside theNlRAR LBP strongly reduce the affinity for ATRA. In-deed, in human receptors, it was shown that the mutationofa small numberof residues involved in thebinding toRAcan abolish ligand binding and transactivation (52, 53).Sequence alignments and protein modeling thus revealedthat, of the 24 amino acids known to be in contact with theligand in vertebrate RARs, seven are different in NlRAR(T154 in helix 1, V183 and A190 in helix 3, M224 in helix5, A242 in the �-turn, L361 in the loop linking helix 11and helix 12, and V365 in helix 12). These seven aminoacid differences might thus be responsible for the reduc-tion of the affinity for retinoids that we identified inNlRAR (Figure 4A and Supplemental Figure 1).
The impact on ligand binding of three amino acid dif-ferences (V183F, A242S, and V365I) was evaluated byreplacing these residues in NlRAR by the correspondingamino acids present in human RAR� (Supplemental Fig-ure 1). In addition, we mutated V239 in the �-strand S1into the corresponding human RAR� methionine residue(Supplemental Figure 1). Although this residue does notestablish a direct contact with the ligand in human RARs,the presence in NlRAR of a valine residue at this position(a significantly smaller amino acid) might destabilize thehydrophobic network located in close proximity of theligand (Figure 4B). We first assessed the ability of the dif-ferent GAL4-LBD-mutant-RAR constructs to activatetranscription in the presence of different retinoids. Of thefour single mutants tested, two, V239M and A242S, ac-tivated transcription in the presence of high doses ofATRA and 9cRA (Figure 5A), but not in the presence of thesynthetic ligand TTNPB (Supplemental Table 1). This dif-ference is very likely due to the higher adaptability and
flexibility of natural retinoids when compared with theconstrained and rigid structures of synthetic ones. Alongthese lines, 3D modeling demonstrated that the A242Smutation restores an important hydrogen bond betweenthe receptor and the carboxylate moiety of the retinoids(Figure 4A). The transactivation assays further revealedthat the other two single mutants (V183F and V365I)could not activate transcription (Supplemental Table 1).
To confirm these results and extend the analysis of theLBP, we also constructed five double mutants (V183F-A242S, V183F-V365I, V239M-A242S, V239M-V365I,and A242S-V365I) (Supplemental Figure 1). Interestingly,all the double mutants can be activated by 9cRA and twoof them (V239M-A242S and A242S-V365I) can addition-ally be activated by ATRA (Figure 5B). Again, all the syn-thetic compounds tested were inactive (Supplemental Ta-ble 1). We further observed that the level of receptoractivation by ATRA and 9cRA greatly varies according tothe mutations introduced in NlRAR. The double mutantcombining the two single mutants that restored activity(V239M plus A242S) strongly activates transcription inthe presence of 9cRA and only weakly in the presence ofATRA. The double mutant A242S plus V365I is activatedmore efficiently by both ATRA and 9cRA than the cor-responding single mutants. We also observed that thecombination of the two single mutants that did not yielda transcriptional activation in the presence of retinoids(V183F plus V365I) gave rise to a mutant receptor that isactivated by 9cRA. These data suggest that a single mu-tation might be sufficient to restore at least a weak retinoidbinding capacity in NlRAR and indicate that a combina-tion of mutations within the LBP might potentiate eachother to further increase the affinity of NlRAR for retinoidbinding. The LBP of NlRAR is thus unable to bind ATRAand other retinoids due to a series of specific mutations
Figure 4. Model of the RAR ligand-binding pocket. A and B, Superimposition of the model of the NlRAR LBP (magenta) on the crystal structure ofthe human RAR� (HsRAR�) LBP (light blue) bound to the RAR�-selective agonist AM580 (green). The NlRAR model was constructed using themodeling meta-server @TOME 2 (37) and the crystal structure of the HsRAR� LBD (PDB code 3KMR) as a template. Divergent residues as well asNlRAR amino acids corresponding to HsRAR�-specific residues are indicated.
doi: 10.1210/en.2014-1181 endo.endojournals.org 4281
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

with additive effects that incrementally altered the affinityfor retinoids. Importantly, this also indicates that muta-tions of amino acids that are not in direct contact with theligand may also affect ligand binding, as observed withNlRAR V239M.
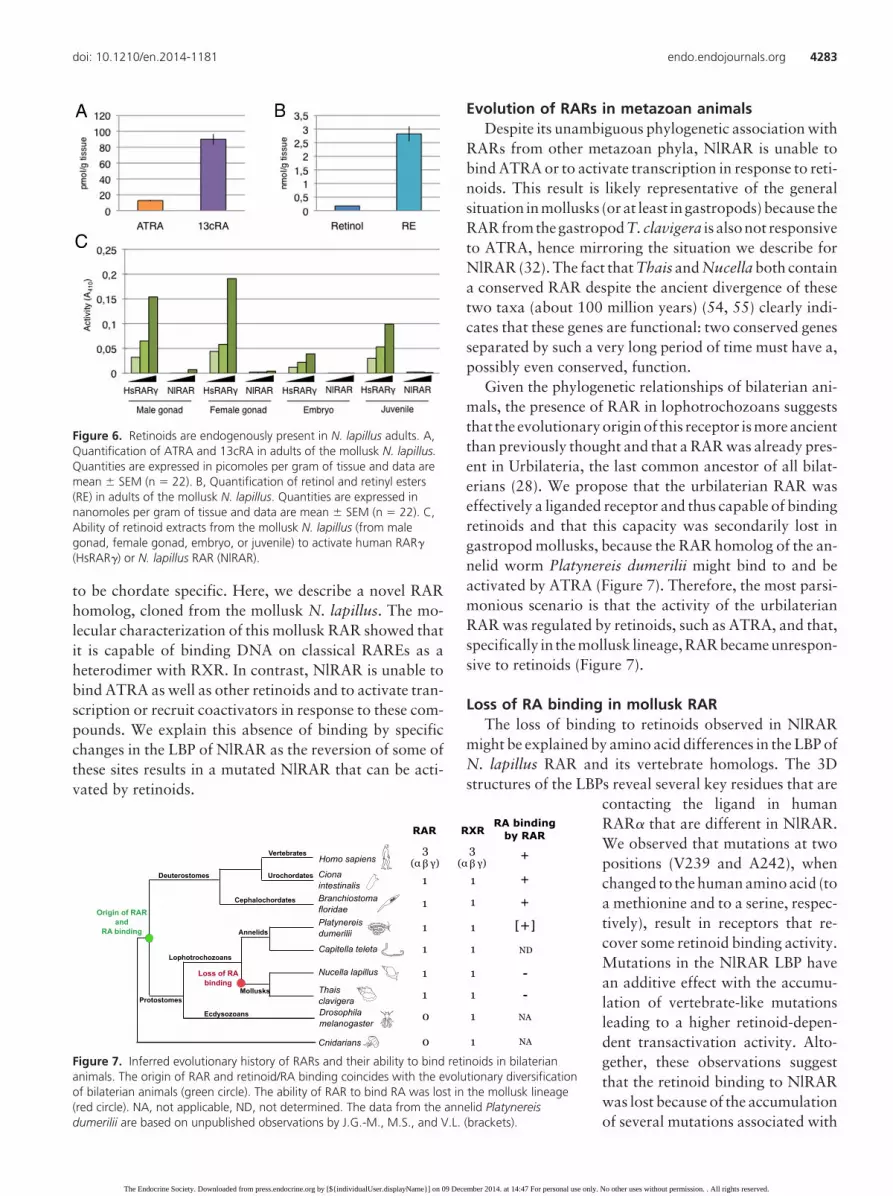
Endogenous retinoids in adult tissuesAnalysis of mollusk genomes has revealed the presence
of homologs of genes encoding enzymes responsible forRA synthesis, such as aldh1 and aldh8 (28). We thereforestudied the retinoids endogenously present in N. lapillusadults by tandem mass spectrometry (39). We were able todetect ATRA, 13cRA, retinol, and retinyl esters, but not9cRA. Concentrations of 13cRA were much higher thanthose of ATRA: 90.0 � 6.7 pmol/g tissue compared with12.8 � 0.9 pmol/g tissue, respectively (Figure 6A). In ad-dition, retinyl ester concentrations were at 2.8 � 0.3nmol/g tissue and those of retinol at 0.18 � 0.01 nmol/gtissue (Figure 6B). This shows that ATRA is present inadult tissues, but at levels much lower than 13cRA, whichseems to be the most abundant RA isomer. Interestingly,
as in most human and mouse tissues, 9cRA seems to beabsent from the tissues we tested.
Considering that NlRAR is unable to bind the differentretinoids tested and to activate transcription, we assessedwhether extracts from N. lapillus can activate NlRAR orone of its vertebrate homologs. We observed that, al-though N. lapillus extracts of male and female gonads, em-bryos, and juveniles can effectively activate the humanRAR�, which confirms the presence of active retinoids inthese extracts, NlRAR is not activated by any of these ex-tracts.Thisstronglysuggests thatNlRARisnonresponsivetothe retinoids contained in the N. lapillus tissue extracts (Fig-ure 6C).
Finally, we assessed the expression of the N. lapillus rargene in different adult tissues and observed that it is ex-pressed in a variety of tissues and organs with the highestlevels found in the kidney (Supplemental Figure 7).
Discussion
Very little is known about the evolutionary origin of theRA signaling pathway, which was believed until recently
Figure 5. Specific mutations recover retinoid binding of NlRAR. The ability of single (A) and double (B) amino acid mutants of NlRAR to activatetranscription of a luciferase reporter gene construct was tested in transfected HEK 293T cells in the presence of increasing concentrations (10�7,10�6, and 10�5 M) of ATRA and 9cRA. The GAL4 DNA binding domain alone was used as a negative control (Ctrl). The nature of the amino acidmutation is indicated for each mutant.
4282 Gutierrez-Mazariegos et al Evolution of Retinoid Receptors Endocrinology, November 2014, 155(11):4275–4286
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.
to be chordate specific. Here, we describe a novel RARhomolog, cloned from the mollusk N. lapillus. The mo-lecular characterization of this mollusk RAR showed thatit is capable of binding DNA on classical RAREs as aheterodimer with RXR. In contrast, NlRAR is unable tobind ATRA as well as other retinoids and to activate tran-scription or recruit coactivators in response to these com-pounds. We explain this absence of binding by specificchanges in the LBP of NlRAR as the reversion of some ofthese sites results in a mutated NlRAR that can be acti-vated by retinoids.
Evolution of RARs in metazoan animalsDespite its unambiguous phylogenetic association with
RARs from other metazoan phyla, NlRAR is unable tobind ATRA or to activate transcription in response to reti-noids. This result is likely representative of the generalsituation inmollusks (orat least ingastropods)because theRARfromthegastropod T. clavigera is alsonot responsiveto ATRA, hence mirroring the situation we describe forNlRAR (32). The fact that Thais and Nucella both containa conserved RAR despite the ancient divergence of thesetwo taxa (about 100 million years) (54, 55) clearly indi-cates that these genes are functional: two conserved genesseparated by such a very long period of time must have a,possibly even conserved, function.
Given the phylogenetic relationships of bilaterian ani-mals, the presence of RAR in lophotrochozoans suggeststhat the evolutionaryoriginof this receptor ismoreancientthan previously thought and that a RAR was already pres-ent in Urbilateria, the last common ancestor of all bilat-erians (28). We propose that the urbilaterian RAR waseffectively a liganded receptor and thus capable of bindingretinoids and that this capacity was secondarily lost ingastropod mollusks, because the RAR homolog of the an-nelid worm Platynereis dumerilii might bind to and beactivated by ATRA (Figure 7). Therefore, the most parsi-monious scenario is that the activity of the urbilaterianRAR was regulated by retinoids, such as ATRA, and that,specifically in the mollusk lineage, RAR became unrespon-sive to retinoids (Figure 7).
Loss of RA binding in mollusk RARThe loss of binding to retinoids observed in NlRAR
might be explained by amino acid differences in the LBP ofN. lapillus RAR and its vertebrate homologs. The 3Dstructures of the LBPs reveal several key residues that are
contacting the ligand in humanRAR� that are different in NlRAR.We observed that mutations at twopositions (V239 and A242), whenchanged to the human amino acid (toa methionine and to a serine, respec-tively), result in receptors that re-cover some retinoid binding activity.Mutations in the NlRAR LBP havean additive effect with the accumu-lation of vertebrate-like mutationsleading to a higher retinoid-depen-dent transactivation activity. Alto-gether, these observations suggestthat the retinoid binding to NlRARwas lost because of the accumulationof several mutations associated with
Figure 6. Retinoids are endogenously present in N. lapillus adults. A,Quantification of ATRA and 13cRA in adults of the mollusk N. lapillus.Quantities are expressed in picomoles per gram of tissue and data aremean � SEM (n � 22). B, Quantification of retinol and retinyl esters(RE) in adults of the mollusk N. lapillus. Quantities are expressed innanomoles per gram of tissue and data are mean � SEM (n � 22). C,Ability of retinoid extracts from the mollusk N. lapillus (from malegonad, female gonad, embryo, or juvenile) to activate human RAR�(HsRAR�) or N. lapillus RAR (NlRAR).
Figure 7. Inferred evolutionary history of RARs and their ability to bind retinoids in bilateriananimals. The origin of RAR and retinoid/RA binding coincides with the evolutionary diversificationof bilaterian animals (green circle). The ability of RAR to bind RA was lost in the mollusk lineage(red circle). NA, not applicable, ND, not determined. The data from the annelid Platynereisdumerilii are based on unpublished observations by J.G.-M., M.S., and V.L. (brackets).
doi: 10.1210/en.2014-1181 endo.endojournals.org 4283
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

the LBP, with each individual mutation probably havingonly minor effects on the ligand binding capacity of thereceptor. Therefore, the loss of retinoid binding by N.lapillus RAR was likely not sudden, but incremental,resulting from the accumulation of individual muta-tions over time.
It is interesting to note that, albeit likely not functioningas a ligand binding site (at least for retinoids), the LBDs ofthe N. lapillus and T. clavigera RAR are reasonably wellconserved, with 68% of sequence identity after approxi-mately 100 million years of divergence (54, 55). Further-more, 3D modeling of the T. clavigera RAR LBD revealedthat the loss of ligand-binding activity in this receptor isprobably not due to changes of the same sites as in the N.lapillus RAR. For example, C321 in the T. clavigera RARreplaces an arginine residue present in the C-terminal partof helix 5 in vertebrate RARs. This arginine is crucial forforming a stabilizing salt bridge with the acidic function ofretinoids. Another important difference between verte-brate RARs and the T. clavigera RAR is the replacementof a cysteine residue contacting the ligand by a smallerglycine residue (G280 in the T. clavigera RAR). Both ofthese amino acids are not conserved between the RARs ofT. clavigera and N. lapillus, and the C321 of T. clavigeraRAR even corresponds to an arginine in N. lapillus, as isthe case in vertebrates. These findings reinforce the notionthat the loss of retinoid binding of mollusk RARs occurredincrementally by the accumulation of several mutations,which might have taken place independently in differentmollusk lineages.
To better understand the loss of retinoid binding of theN. lapillus and T. clavigera RAR, we assessed which al-terations to the RAR sequence are specific to the mollusklineage; for this, we reconstructed the ancestral sequencesof the mollusk and protostome lineages (nodes 29 and 31,respectively, Supplemental Figure 8). We hence identified46 substitutions that are specific to the two mollusk RARs,of which 43 are in the LBD. In vertebrates, only two ofthese are in direct contact with the ligand: the threonine/serine 148 in helix 1 and the valine 365 in helix 12 (Sup-plemental Figure 8). These sites represent fundamentaldifferences that distinguish the mollusk RARs from allother known RAR sequences and are thus likely to haveoccurred early in the divergence of the mollusk lineage,at the onset of the reduction of the affinity for retinoidsof the mollusk RARs. This scenario for the stepwise lossof retinoid binding of the mollusk RARs is very similarto the mechanisms proposed for the acquisition of sub-strate specificity of the duplicated vertebrate steroid re-ceptors (2).
Retinoids in mollusksOur analysis of the retinoid content of N. lapillus re-
vealed the presence, at relatively high levels, of ATRA and13cRA and the apparent absence of 9cRA. In addition, wealso detected retinol and retinyl esters in N. lapillus. Thesefindings are somewhat different from previous retinoidmeasurements in N. lapillus, given that these previous re-ports were unable to detect retinol and retinyl esters in N.lapillus tissues (25). Also, these authors did not observeretinyl ester synthesis, when retinol was injected into theadult, suggesting that N. lapillus does not have the abilityto store retinoids in the form of retinyl esters. In addition,in this previous study, 9cRA was detected in N. lapillustissues, albeit at very low levels (25). These apparent dis-crepancies may be linked to the fact that the authors usedisolated tissues of N. lapillus, whereas we used extractsprepared from complete animals.
Importantly, though, there is one common feature be-tween these results: the detection of high levels of ATRAand 13cRA (25). These data thus suggest a paradoxicalreality: elevated concentrations of active retinoids arepresent in N. lapillus, but the putative cognate receptor,NlRAR, is unable to bind these compounds. It is possiblethat the biologically active ligand recognized by NlRAR isa different molecule, maybe even a different retinoid. Thefact that the N. lapillus and T. clavigera RAR LBD exhibit68% sequence identity and that at least some positions ofthe LBP are conserved between the two species (V228,M269, and V410) may suggest that a common ligand forthese receptors remains to be identified (54, 55), even ifextracts from N. lapillus were unable to activate NlRAR,in contrast to human RAR�. The fact that we detectedATRA and 13cRA in N. lapillus tissues and that molluskgenomes contain orthologs of the enzymes implicated inRA metabolism (28, 27, 28, 31) favors the notion thatretinoids have a specific function in members of this taxon.Studies of the biological functions of NlRAR are needed toobtain additional insights into the mechanisms of its ac-tivity and possible activation by a ligand.
ConclusionOur findings illustrate the plasticity of the LBD of nu-
clear receptors by illustrating the lineage-specific loss ofretinoid binding capacity by RARs. The characterizationof NR orthologs from various different animals has re-vealed that classical liganded vertebrate NRs, such asRARs, TRs, or ERs, are evolutionarily much more ancientthan expected, but also that the presence of these genes ina given genome does not automatically translate into anevolutionarily conserved function. Our work also illus-trates that the characterization of NRs from alternativemodel systems can shed new light on our understanding of
4284 Gutierrez-Mazariegos et al Evolution of Retinoid Receptors Endocrinology, November 2014, 155(11):4275–4286
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

the evolution of endocrine systems that might have neverbeen obtained from work on model organisms.
Acknowledgments
We thank Marie Sémon for help with sequence analyses, LaurentGuéguen for help with the reconstruction of ancestral sequences,andGuillaumeHolzer forperformingthecompetitionexperiments.
Address all correspondence and requests for reprints to: Vin-cent Laudet, Institut de Génomique Fonctionnelle de Lyon, EcoleNormale Supérieure de Lyon, 46 Allée d’Italie, 69364 Lyon Ce-dex 07, France. E-mail: [email protected]; or MichaelSchubert, Laboratoire de Biologie du Développement de Ville-franche-sur-Mer, Observatoire Océanologique de Villefranche-sur-Mer, 181 Chemin du Lazaret, 06230 Villefranche-sur-Mer,France. E-mail: [email protected].
V.L. is supported by the Centre National de la RechercheScientifique, the ENS de Lyon, and the MENRT. M.S. is sup-ported by the Centre National de la Recherche Scientifique andby Grant ANR-11-JSV2-002-01 awarded by the Agence Nation-ale de la Recherche. L.F.C.C. and M.M.S. are supported by theFCT (Grants PTDC/MAR/105199/2008 and PTDC/MAR/115199/2009). J.-I.N. is supported by the KAKENHI (Grant21590143). M.K. received support from the National Institute ofAllergy and Infectious Diseases Contract HHSN272202000046Cand from the University of Maryland, School of Pharmacy, MassSpectrometry Center (Grant SOP1841-IQB2014).
Disclosure Summary: The authors have nothing to disclose.
References
1. Laudet V, Gronemeyer H. The nuclear receptor facts book. SanDiego: Academic Press; 2002.
2. Harms MJ, Eick GN, Goswami D, et al. Biophysical mechanisms forlarge-effect mutations in the evolution of steroid hormone receptors.Proc Natl Acad Sci USA. 2013;110:11475–11480.
3. Bridgham JT, Keay J, Ortlund EA, Thornton JW. Vestigialization ofan allosteric switch: genetic and structural mechanisms for the evo-lution of constitutive activity in a steroid hormone receptor. PLoSGenet. 2014;10:e1004058.
4. Lecroisey C, Laudet V, Schubert M. The cephalochordate amphiox-us: a key to reveal the secrets of nuclear receptor evolution. BriefFunct Genomics. 2012;11:156–166.
5. Paris M, Pettersson K, Schubert M, et al. An amphioxus orthologueof the estrogen receptor that does not bind estradiol: insights intoestrogen receptor evolution. BMC Evol Biol. 2008;8:219.
6. Thornton JW, Need E, Crews D. Resurrecting the ancestral steroidreceptor: ancient origin of estrogen signaling. Science. 2003;301:1714–1717.
7. Markov GV, Tavares R, Dauphin-Villemant C, Demeneix BA,Baker ME, Laudet V. Independent elaboration of steroid hormonesignaling pathways in metazoans. Proc Nat Acad Sci USA. 2009;106:11913–11918.
8. Scott AP. Do mollusks use vertebrate sex steroids as reproductivehormones? Part I: critical appraisal of the evidence for the presence,biosynthesis and uptake of steroids. Steroids. 2012;77:1450–1468.
9. Scott AP. Do mollusks use vertebrate sex steroids as reproductive
hormones? II. Critical review of the evidence that steroids have bi-ological effects. Steroids. 2013;78:268–281.
10. Paris M, Escriva H, Schubert M, et al. Amphioxus postembryonicdevelopment reveals the homology of chordate metamorphosis.Curr Biol. 2008b;18:825–830.
11. Kane MA. Analysis, occurrence, and function of 9-cis-retinoic acid.Biochim Biophys Acta. 2012;1821:10–20.
12. Balmer JE, Blomhoff R. A robust characterization of retinoic acidresponse elements based on a comparison of sites in three species.J Steroid Biochem Mol Biol. 2005;96:347–354.
13. Chambon P. A decade of molecular biology of retinoic acid recep-tors. FASEB J. 1996;10:940–954.
14. Durand B, Saunders M, Leroy P, Leid M, Chambon P. All-trans and9-cis retinoic acid induction of CRABPII transcription is mediatedby RAR-RXR heterodimers bound to DR1 and DR2 repeated mo-tifs. Cell. 1992;71:73–85.
15. Moutier E, Ye T, Choukrallah M-A, et al. Retinoic acid receptorsrecognize the mouse genome through binding elements with diversespacing and topology. J Biol Chem. 2012;287:26328–26341.
16. Ross SA, McCaffery PJ, Drager UC, De Luca LM. Retinoids inembryonal development. Physiol Rev. 2000;80:1021–1054.
17. Smith WC, Nakshatri H, Leroy P, Rees J, Chambon, P. A retinoicacid response element is present in the mouse cellular retinol bindingprotein I (mCRBPI) promoter. EMBO J. 1991;10:2223–2230.
18. Gutierrez-Mazariegos J, Schubert M, Laudet V. Evolution of reti-noic acid receptors and retinoic acid signaling. Subcell Biochem.2014;70:55–73.
19. Dmetrichuk JM, Carlone RL, Spencer GE. Retinoic acid inducesneurite outgrowth and growth cone turning in invertebrate neurons.Dev Biol. 2006;294:39–49.
20. Dmetrichuk JM, Carlone RL, Jones TRB, Vesprini ND, Spencer GE.Detection of endogenous retinoids in the molluskan CNS and char-acterization of the trophic and tropic actions of 9-cis retinoic acid onisolated neurons. J Neurosci. 2008;28:13014–13024.
21. Nishikawa J, Mamiya S, Kanayama T, Nishikawa T, Shiraishi F,Horiguchi T. Involvement of the retinoid X receptor in the devel-opment of imposex caused by organotins in gastropods. Environ SciTechnol. 2004;38:6271–6276.
22. Castro FC, Lima D, Machado A, et al. Imposex induction is medi-ated through the retinoid X receptor signalling pathway in the neo-gastropod Nucella lapillus. Aquat Toxicol. 2007;85:57–66.
23. Creton R, Zwaan G, Dohmen R. Specific developmental defects inmollusks after treatment with retinoic acid during gastrulation. DevGrowth Differ. 1993;35:357–364.
24. Gesto M, Castro LF, Reis-Henriques MA, Santos MM. Retinol me-tabolism in the mollusk Osilinus lineatus indicates an ancient originfor retinyl ester storage capacity. PLoS One. 2012;7:e35138.
25. Gesto M, Castro LF, Santos M. Differences in retinoid levels andmetabolism among gastropod lineages: imposex-susceptible gastro-pods lack the ability to store retinoids in the form of retinyl esters.Aquat Toxicol. 2013;142–143:96–103.
26. Bouton D, Escriva H, de Mendonça RL, et al. A conserved retinoidX receptor (RXR) from the mollusk Biomphalaria glabrata trans-activates transcription in the presence of retinoids. J Mol Endocri-nol. 2005;34:567–582.
27. Albalat R, Cañestro C. Identification of Aldh1a, Cyp26 and RARorthologs in protostomes pushes back the retinoic acid genetic ma-chinery in evolutionary time to the bilaterian ancestor. Chem BiolInteract. 2009;178:188–196.
28. Campo-Paysaa F, Marlétaz F, Laudet V, Schubert M. Retinoic acidsignaling in development: tissue-specific functions and evolutionaryorigins. Genesis. 2008;46:640–656.
29. Cañestro C, Postlethwait JH, Gonzàlez-Duarte R, Albalat R. Is reti-noic acid genetic machinery a chordate innovation? Evol Dev. 2006;8:394–406.
30. Marlétaz F, Holland L, Laudet V, Schubert M. Retinoic acid sig-naling and the evolution of chordates. Int J. 2006;2:38–47.
doi: 10.1210/en.2014-1181 endo.endojournals.org 4285
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.

31. Simões-Costa MS, Azambuja AP, Xavier-Neto J. The search fornon-chordate retinoic acid signaling: lessons from chordates. J ExpZool B Mol Dev Evol. 2008;310:54–72.
32. Urushitani H, Katsu Y, Ohta Y, Shiraishi H, Iguchi T, Horiguchi T.Cloning and characterization of the retinoic acid receptor-like pro-tein in the rock shell, Thais clavigera. Aquat Toxicol. 2013;142–143:403–413.
33. Allenby G, Bocquel MT, Saunders M, et al. Retinoic acid receptorsand retinoid X receptors: interactions with endogenous retinoic ac-ids. Proc Natl Acad Sci USA. 1993;90:30–34.
34. Gouy M, Guindon S, Gascuel O. SeaView version 4: a multiplatformgraphical user interface for sequence alignment and phylogenetictree building. Mol Biol Evol. 2010;27:221–224.
35. Escriva H, Bertrand S, Germain P, et al. Neofunctionalization invertebrates: the example of retinoic acid receptors. PLoS Genet.2006;2:e102.
36. le Maire A, Teyssier C, Erb C, et al. A unique secondary-structureswitch controls constitutive gene repression by retinoic acid recep-tor. Nat Struct Mol Biol. 2010;17:801–807.
37. Pons JL, Labesse G. @TOME-2: a new pipeline for comparativemodeling of protein-ligand complexes. Nucleic Acids Res. 2009;37:W485–W491.
38. Kane MA, Napoli JL. Quantification of endogenous retinoids(Chapter 1). Retinoids: Methods and Protocols. In: Sun H, TravisGH, eds. Methods in Molecular Biology. Totowa: Humana Press;2010;652:1–54.
39. Kane MA, Folias AE, Wang C, Napoli JL. Quantitative profiling ofendogenous retinoic acid in vivo and in vitro by tandem mass spec-trometry. Anal Chem. 2008;80:1702–1708.
40. Kane MA, Folias AE, Napoli JL. HPLC/UV quantitation of retinal,retinol, and retinyl esters in serum and tissues. Anal Biochem. 2008;378:71–79.
41. Perlmann T, Rangarajan PN, Umesono K, Evans RM. Determinantsfor selective RAR and TR recognition of direct repeat HREs. GenesDev. 1993;7:1411–1422.
42. Wilson TE, Paulsen RE, Padgett KA, Milbrandt J. Participation ofnon-zinc finger residues in DNA binding by two nuclear orphanreceptors. Science. 1992;256:107–110.
43. Zechel C, Shen XQ, Chen JY, Chen ZP, Chambon P, GronemeyerH. The dimerization interfaces formed between the DNA binding
domains of RXR, RAR and TR determine the binding specificity andpolarity of the full-length receptors to direct repeats. EMBO J. 1994;13:1425–1433.
44. Zechel C, Shen XQ, Chambon P, Gronemeyer H. Dimerization in-terfaces formed between the DNA binding domains determine thecooperative binding of RXR/RAR and RXR/TR heterodimers toDR5 and DR4 elements. EMBO J. 1994;13:1414–1424.
45. Gronemeyer H, Gustafsson JA, Laudet V. Principles for modulationof the nuclear receptor superfamily. Nat Rev Drug Discov. 2004;3:950–964.
46. Renaud J, Rochel N, Ruff M, Vivat V. Crystal structure of theRAR-� ligand-binding domain bound to all-trans retinoic acid.Nature. 1995;378:681–689.
47. Urushitani H, Katsu Y, Ohta Y, Shiraishi H, Iguchi T, Horiguchi T.Cloning and characterization of retinoid X receptor (RXR) isoformsin the rock shell, Thais clavigera. Aquat Toxicol. 2011;103:101–111.
48. Leid M. Ligand-induced alteration of the protease sensitivity of ret-inoid X receptor �. J Biol Chem. 1994;269:14175–14181.
49. Egea PF, Mitschler A, Moras D. Molecular recognition of agonistligands by RXRs. Mol Endocrinol. 2002;16:987–997.
50. Nahoum V, Pérez E, Germain P, et al. Modulators of the structuraldynamics of the retinoid X receptor to reveal receptor function. ProcNat Acad Sci USA. 2007;104:17323–17328.
51. Kuiper GG, Enmark E, Pelto-Huikko M, Nilsson S, Gustafsson JA.Cloning of a novel receptor expressed in rat prostate and ovary. ProcNatl Acad Sci USA. 1996;93:5925–5930.
52. Ostrowski J, Hammer L, Roalsvig T, Pokornowski K, Reczek PR.The N-terminal portion of domain E of retinoic acid receptors � and� is essential for the recognition of retinoic acid and various analogs.Proc Natl Acad Sci USA. 1995;92:1812–1816.
53. Tate BF, Grippo JF. Mutagenesis of the ligand binding domain of thehuman retinoic acid receptor � identifies critical residues for 9-cis-retinoic acid binding. J Biol Chem. 1995;270:20258–20263.
54. Barco A, Claremont M, Reid DG, et al. A molecular phylogeneticframework for the Muricidae, a diverse family of carnivorous gas-tropods. Mol Phylogenet Evol. 2010;56:1025–1039.
55. Claremont M, Vermeij GJ, Williams ST, Reid DG. Global phylog-eny and new classification of the Rapaninae (Gastropoda: Muri-cidae), dominant molluscan predators on tropical rocky seashores.Mol Phylogenet Evol. 2013;66:91–102.
4286 Gutierrez-Mazariegos et al Evolution of Retinoid Receptors Endocrinology, November 2014, 155(11):4275–4286
The Endocrine Society. Downloaded from press.endocrine.org by [${individualUser.displayName}] on 09 December 2014. at 14:47 For personal use only. No other uses without permission. . All rights reserved.





























