Embed Size (px)
Citation preview

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
A new varanoid squamate from the Early Cretaceous (BarremianeAptian)of Burgos, Spain
Alexandra Houssaye a,b,*, Jean-Claude Rage a, Fidel Torcida Fernández-Baldor c,d,Pedro Huerta c,d, Nathalie Bardet a, Xabier Pereda Suberbiola e
aUMR 7207 du CNRS, Département Histoire de la Terre, Muséum National d’Histoire Naturelle, 57 rue Cuvier CP 38, 75005 Paris, Franceb Steinmann Institut für Geologie, Paläontologie und Mineralogie, Universität Bonn, Nussallee 8, 53115 Bonn, GermanycMuseo de Dinosaurios de Salas de los Infantes, 09600 Salas de los Infantes, Burgos, SpaindColectivo ArqueológicoePaleontológico Salense (CAS), Plaza Jesús Aparicio 9, 09600 Salas de los Infantes, Burgos, SpaineUniversidad del País Vasco/EHU, Facultad de Ciencia y Tecnología, Estratigrafía y Paleontología, Apartado 644, 48080 Bilbao, Spain
a r t i c l e i n f o
Article history:Received 7 May 2012Accepted in revised form 23 November 2012Available online 20 December 2012
Keywords:SquamataVaranoideaVertebraePaleoecologyVaranoid evolutionary history
a b s t r a c t
A new anguimorph lizard from the late BarremianeAptian of the Salas de los Infantes area (CamerosBasin, Iberian Peninsula) is described here as Arcanosaurus ibericus gen. et sp. nov. It corresponds to thefirst squamate remains from the Burgos Province. The unique combination of morphological andmicroanatomical characters enables us to erect a new genus and species that we tentatively assign toVaranoidea despite the strange seemingly absence on the axis of a posterior hypapophysis. Thecombination of both microanatomical and geological data suggests an essentially terrestrial mode of life.This discovery reveals itself particularly interesting concerning the evolutionary history of varanoids asthis new Spanish taxon might correspond to the oldest terrestrial varanoid known to date.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
The Salas de los Infantes area (Burgos Province, northern Spain)has yielded numerous fossil remains of dinosaurs, turtles andcrocodiles (Torcida Fernández-Baldor, 2006; Torcida Fernández-Baldor et al., 2011; Pérez-García et al., 2012; Pereda Suberbiolaet al., 2012). This study presents the first squamate remains fromthis region, which have been discovered in the Castrillo de la ReinaFormation (late BarremianeAptian) of Viajete.
Bolet and Evans (2010) stated that, as compared to other parts ofEurope, the Iberian Peninsula has a relatively good Mesozoicsquamate record. However, although the squamate fauna from theMesozoic of Iberia is one of the richest known, it is rather limited.One major difficulty is the relatively frequent impossibility toclassify at a precise level the material that is often too fragmentary.Another difficulty involves the field sampling because the materialis generally of small size, and collecting often requires screening-
washing techniques that are generally not used when fieldworkfocalizes on larger vertebrates.
Squamates have been recovered from various sites in EarlyCretaceous aged deposits of Spain (in the Iberian Range and Pyr-enean domain). In Las Hoyas (Cuenca Province, late Barremian) areknown the lacertoid scincomorph Meyasaurus diazromerali Evansand Barbadillo, 1997, the stem squamates Scandensia ciervensisEvans and Barbadillo, 1998 and Hoyalacerta sanzi Evans andBarbadillo, 1999; in Uña (Cuenca Province, late Barremian) Meya-saurus, two Paramacellodidae indet. and Cuencasaurus (Incertaesedis); in Buenache de la Sierra (Cuenca Province, late Barremian) atleast one yet undescribed squamate (Evans and Barbadillo, 1998;Bolet and Evans, 2010; Canudo et al., 2010); in Galve (Teruel Prov-ince, late Hauterivianeearly Barremian), Meyasaurus and para-macellodids (Evans and Barbadillo, 1997); in El Montsec (LleidaProvince, late Berriasianeearly Valanginian) Meyasaurus, the scin-cogekkonomorph Pedrerasaurus latifrontalis Bolet and Evans, 2010,and the stem squamate or stem scleroglossan EichstaettisaurusKuhn, 1958. The Spanish Early Cretaceous squamate assemblage isthus dominated by the scincomorph lizard Meyasaurus. These taxaare generally considered as terrestrial and as having lived close tothe edges of still waters (Evans and Barbadillo, 1998; Canudo et al.,2010).
* Corresponding author. Steinmann Institut für Geologie, Paläontologie undMineralogie, Universität Bonn, Nussallee 8, 53115 Bonn, Germany. Fax: +49 228 733509.
E-mail addresses: [email protected], [email protected] (A. Houssaye).
Contents lists available at SciVerse ScienceDirect
Cretaceous Research
journal homepage: www.elsevier .com/locate/CretRes
0195-6671/$ e see front matter � 2012 Elsevier Ltd. All rights reserved.http://dx.doi.org/10.1016/j.cretres.2012.11.005
Cretaceous Research 41 (2013) 127e135

Author's personal copy
This study reports the first discovery of squamate material fromthe Burgos Province of Spain, describes and diagnoses this newmaterial, and also provides important new data about the evolu-tionary history of squamates in Spain.
2. Geological setting
The Viajete site is located close to Villanueva de Carazo village,4 km apart from Salas de los Infantes (Burgos, Spain) (Fig. 1). Thesite occurs in the Castrillo de la Reina Formation, which belongs tothe fifth depositional sequence of the Cameros Basin (Martín-Closas and Alonso Millán, 1998). The Castrillo de la Reina Forma-tion is equivalent in the Salas de los Infantes area to part of theUrbión Group (Beuther, 1966). This basin was extensional duringthe Late JurassiceEarly Cretaceous and accumulated more than9000 m of non-marine deposits (Mas et al., 1993). The CamerosBasin was the more internal of the basins in the Iberian rift systemthat connected the Tethys with the North-Atlantic ocean (Salasand Casas, 1993; Salas et al., 2001), and although some foramini-fers and algal remains e interpreted as marine incursions e havebeen reported in the eastern part (Mas et al., 2002), the westernpart of the basin was non-marine until the Cenomanian. Thispaper follows the stratigraphic framework of Beuther (1966) andMartín-Closas and Alonso Millán (1998). In the western part of thebasin, the age of the fifth depositional sequence is late BarremianeAptian based on charophyte and ostracod biostratigraphy (Martín-Closas and Alonso Millán, 1998; Schudack and Schudack, 2009).The lack of pollen studies in the rocks of the fifth depositionalsequence, the absence of geochemical dating and the impossibilityof correlation with marine beds, do not allow a more precisedating of the Viajete site. During the last years, the stratigraphicframework of the basin has been reinterpreted (Mas et al., 2002;Arribas et al., 2003; Clemente, 2010). The main problem of thenew stratigraphic proposals is the lack of a geological map thatreflects the extension and distribution and allows a clear
distinction between the different lithostratigraphic units (groupsand formations).
The Castrillo de la Reina Formation is mainly composed of redmudstones and white to yellow sandstone beds with a channel orlenticular morphology. The sandstone beds occur as ribbon-shapedchannel bodies, 3e10 m thick, scattered in red mudstones. TheCastrillo de la Reina Formation in the Viajete area is dominated bythe red mudstones and, on the contrary to the stratotype, thelenticular sandstone bodies are very scarce. The sandstone bedshave tabular geometries, their thickness is less than 1.5 m and,instead of being white and yellow (like in the stratotype), they arered colored and have a high content in clay matrix.
The Castrillo de la Reina Formation is interpreted as a fluvialsystem that drained the Cameros Basin toward the NE. Althoughlarge sandstone ribbon-shaped bodies, interpreted as channel fills,occur relatively close in the Hacinas area, in Viajete the dominanceof redmudstones suggest that this areawas part of the floodplain ofthe fluvial system, lateral to the main fluvial channels. The tabularsandstones interbedded within the red mudstone beds are inter-preted as crevasse splay deposits. The high content in red claymatrix in theses sandstone beds indicates that the flow was notstrong enough to separate the bedload from the suspended load.
The paleoenvironment where the new varanoid remains werefound in the Viajete site is interpreted as a floodplain, which wasinundated during the floods of the fluvial system that drained thispart of the Cameros Basin. In the Salas de los Infantes area, differentskeletal remains of sauropod, ornithopod, and theropod dinosaurs,turtles, and crocodiles, as well as ichnites have been found in theCastrillo de la Reina fluvial system (see Torcida Fernández-Baldor,2006; Pereda Suberbiola et al., 2003a, b; Torcida Fernández-Baldor et al., 2011; Huerta et al., 2012; Pérez-García et al., 2012;Pereda Suberbiola et al., 2012).
Institutional abbreviations: MDS:Museo de Dinosaurios de Salas delos Infantes; VJ: Viajete site, Salas de los Infantes, Burgos, Spain.
Fig. 1. Location map, geological sketch of the Salas de los Infantes area (Burgos, Spain), and stratigraphic position of the Viajete fossiliferous level.
A. Houssaye et al. / Cretaceous Research 41 (2013) 127e135128

Author's personal copy
3. Material and methods
The material consists of 29 vertebrae discovered associated butdisarticulated in the late BarremianeAptian site of Viajete. Thesevertebrae do not display any notable taphonomic variation (e.g. ofcolor). Moreover, there is no ontogenetic variation. It can thus besuggested that they all belonged to the same individual. Thishypothesis is supported by the fact that there appears to bea coherent size variation along the vertebral column. However, aswe can not ascertain that there is indeed only one individual, wewill define a holotype and the rest of the material will consist ofparatypes. The vertebrae are housed in the collections of the Salasde los Infantes Museum under the references MDS-VJ 1-29.
In order to obtain microanatomical data and, especially, to testthe possible occurrence of osteosclerosis (increase in inner bonecompactness; not morphologically observable), the microanatomyof one of these vertebrae was investigated. The dorsal vertebraMDS-VJ 6 was mCT scanned. Third generation synchrotron micro-tomography was used at the European Synchrotron RadiationFacility (ESRF, Grenoble, France) on beamline ID19 during anexperiment realized in collaboration with P. Tafforeau (ESRF,France). Image visualization and mid-sagittal and neutral trans-verse thin sections (sensu Buffrénil et al., 2008) were made usingthe software VG StudioMax version 2.2. (Volume Graphics, Hei-delberg, Germany). The relative area of primary periosteal (¼cortical) bone in longitudinal section (PBCL) could be estimatedfollowing Houssaye et al. (2010) as the area occupied by primaryperiosteal bone multiplied by 100 and divided by the total area ofthe centrum, and the relative area of primary periosteal bone intransverse section (PBCT) as the area occupied by primary perios-teal bone multiplied by 100 and divided by the total vertebral areaminus the area occupied by the neural canal.
4. Systematic Paleontology
Reptilia Linnaeus, 1758Squamata Oppel, 1811Anguimorpha Fürbringer, 1900Varanoidea Gray, 1827Genus Arcanosaurus gen. nov.Derivation of name. From the Spanish arcano (Latin, arcanus):mysterious.Type and only species. Arcanosaurus ibericus sp. nov.
Diagnosis. As for species.
Arcanosaurus ibericus sp. nov.Derivation of name. From the Latin ibericus, referring to the IberianPeninsula.Holotype: Mid-dorsal vertebra MDS-VJ 5 (Fig. 3FeJ).Type Locality and Horizon: Viajete, near Salas de los Infantes (Bur-gos, Spain); Castrillo de la Reina Formation; Late BarremianeAptian(Martín-Closas and Alonso Millán, 1998; Schudack and Schudack,2009).Paratypes: Axis MDS-VJ 1 (Fig. 2AeF); cervical vertebrae MDS-VJ2,8,9,16,17 (Fig. 2GeJ; Fig. 3AeE); dorsal vertebrae MDS-VJ3,4,6,7,10-15 and caudal vertebrae MDS-VJ 18-29 (Fig. 3KeO)Differential diagnosis: Anguimorpha differing from all otherAnguimorpha, except Parviraptor, in the absence of secondaryhypapophysis on the axis; from the non-varanoidean anguimor-phans in the absence of autotomous septa, the presence of hypa-pophyses of central origin (i.e., originating from the centrum) withindividualized intercentral elements in cervical vertebrae, theobliquity of the axis of the cotyleecondyle system, the precondylar
constriction and the chevrons not in contact with the condyle.Presumed Varanoidea differing from Pythonomorpha in theabsence of a zygospheneezygantrum system; from Pachyvaranus inthe absence of a dorsally notched and subrectangularly shapedneural spine, the occurrence of aweak precondylar constriction andthe absence of pachyostosis; from Varanus in the smaller andsubrounded neural canal, the much less elongated axis centrum,the absence of anterior depression on each side of the neural spine,the less inclined cotyleecondyle system and the less well devel-oped precondylar constriction; from Gobiderma in the absence ofzygosphene and in the occurrence of a weak precondylarconstriction; from Necrosauridae (restricted to Necrosaurus, asoriginally defined) in the small size of the neural canal and themore inclined and markedly lower neural spine on dorsal verte-brae; from both Varanus and Necrosaurus in the absence of clearpars tectiformis; from Helodermatidae and Saniwa in lackinga subrectangular shape of the neural spine; from Gobiderma, Hel-odermatidae and Lanthanotidae in the triangular shape of thecentrum; from Saniwa in that cervical vertebrae are not longer thananterior dorsals, in the relatively shorter axis, in the absence ofa rudimentary pseudozygosphene and in the presence of trans-verse processes in the axis and third cervical; from Primaderma byits centrum clearly narrower anteriorly and by its more posteriorlylocated paradiapophyses; from Parasaniwa, Palaeosaniwa and Tel-masaurus in the absence of a rudimentary pseudozygosphene; fromParasaniwa and Palaeosaniwa in the absence of depressions on eachside of the neural arch, from Parasaniwa in the precondylarconstriction, and from Palaeosaniwa in its smaller zygapophysesand condyle, and the relatively narrower anterior part of thecentrum resulting in less depressed vertebrae.
5. Description
5.1. Introduction
Comparisons with extant varanoid lizards were used to assessthe original positions of the vertebrae along the vertebral columnand to analyze the intracolumnar variations. Despite this varia-bility, various characters are common to almost all vertebrae.
All centra (except of course the axis) are procoelous. In dorsalview, vertebrae are slightly wider anteriorly than posteriorly,resulting from the more important lateral projection of the pre-zygapophyses as compared to the postzygapophyses. The narrow-est part of the interzygapophyseal constriction is positioned atabout the mid-length of the vertebra (Fig. 3H). The pre-zygapophyseal articular surfaces are particularly large. They are notelongated but rather subrounded (Figs. 2H, 3I). There is no pre-zygapophyseal process and no zygospheneezygantrum system.Paradiapophyses do not extend laterally beyond the pre-zygapophyses (except in anterior cervicals; or maybe only in C3;Fig. 2H versus Fig. 3DeF). The neural spine rises progressivelyabove the neural arch (Fig. 3C, J, N, O) and posteriorly extendsmuchbeyond the level of the condyle (Fig. 3C, J). Rather thin anteriorly,the neural spine becomes thicker from about its mid-length to itsposterior end (Fig. 3H). Because of the antero-lateral projection ofthe prezygapophyses and absence of zygosphene, a “U” shapedembayment is observable above the anterior border of the neuralcanal in dorsal view (Fig. 3H). In anterior view, vertebrae aredepressed with a low neural arch (Fig. 3D,F). The neural canal issmall (much smaller in both height and width than the cotyle) andsubcircular to ovoid (dorso-ventrally elongated) in shape (Fig. 3D,F). Zygapophyses are markedly inclined, at about 45 degrees to thehorizontal (Fig. 3D, F). The cotyle and condyle are depressed(Fig. 3D, F). In lateral view, paradiapophyses seem to form a singlearticular facet. The cotyleecondyle system is inclined on the
A. Houssaye et al. / Cretaceous Research 41 (2013) 127e135 129
Author's personal copy
horizontal, with the cotyle facing anteroventrally and the condylebeing oriented posterodorsally (Fig. 3I, J). In ventral view, thecentrum is subtriangular in shape (Fig. 3A, I). There seems to bea slight precondylar constriction (Fig. 3A, M). In posterior view, theneural spine is thick (Fig. 3E, G). Only subcentral and lateralforamina are observable but their occurrence is variable, withoutany correlation with the position within the vertebral column.
Some vertebrae display a lateral thickening in ventral view. Thisfeature is regularly observed in pachyostotic squamates (Houssayeet al., 2008; A.H., pers. obs). However, it is unclear if this slightlateral thickening corresponds to a real increase in cortical bonedeposits (pachyostosis) or if it is related to the vertebralmorphology without peculiar osseous specialization. This questionis discussed below (Section 5.5) in the light of the microanatomicaldata.
As compared to adult varanoid lizards, these vertebrae displaya well-ossified condyle, the lateral borders of the neural canal arewell developed in their anterior part, the cotyle and condyle are notexcessively depressed and the neural canal is not excessively wide.These characters being those used to determine a juvenile stage
based on vertebrae in squamates (Scanlon et al., 2003; Rage andNéraudeau, 2004), this material is considered to belong to anadult individual.
5.2. Cervical vertebrae
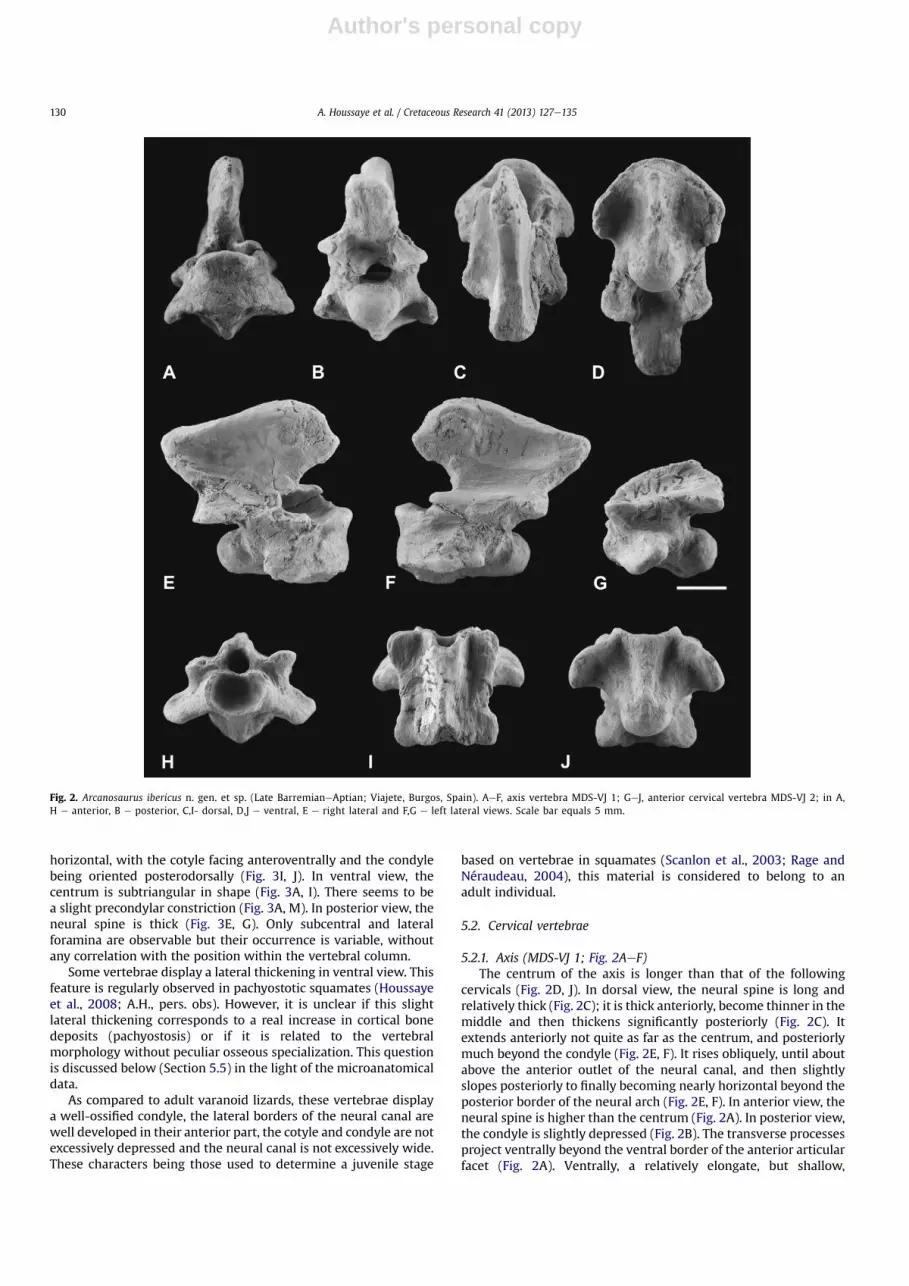
5.2.1. Axis (MDS-VJ 1; Fig. 2AeF)The centrum of the axis is longer than that of the following
cervicals (Fig. 2D, J). In dorsal view, the neural spine is long andrelatively thick (Fig. 2C); it is thick anteriorly, become thinner in themiddle and then thickens significantly posteriorly (Fig. 2C). Itextends anteriorly not quite as far as the centrum, and posteriorlymuch beyond the condyle (Fig. 2E, F). It rises obliquely, until aboutabove the anterior outlet of the neural canal, and then slightlyslopes posteriorly to finally becoming nearly horizontal beyond theposterior border of the neural arch (Fig. 2E, F). In anterior view, theneural spine is higher than the centrum (Fig. 2A). In posterior view,the condyle is slightly depressed (Fig. 2B). The transverse processesproject ventrally beyond the ventral border of the anterior articularfacet (Fig. 2A). Ventrally, a relatively elongate, but shallow,
Fig. 2. Arcanosaurus ibericus n. gen. et sp. (Late BarremianeAptian; Viajete, Burgos, Spain). AeF, axis vertebra MDS-VJ 1; GeJ, anterior cervical vertebra MDS-VJ 2; in A,H e anterior, B e posterior, C,I- dorsal, D,J e ventral, E e right lateral and F,G e left lateral views. Scale bar equals 5 mm.
A. Houssaye et al. / Cretaceous Research 41 (2013) 127e135130

Author's personal copy
Fig. 3. Arcanosaurus ibericus n. gen. et sp. (Late BarremianeAptian; Viajete, Burgos, Spain). AeB, posterior cervical vertebra MDS-VJ 17; C, posterior cervical vertebra MDS-VJ 9; DeE,posterior cervical vertebra MDS-VJ 8; FeJ, dorsal vertebra (holotype) MDS-VJ 5; KeL, caudal vertebra MDS-VJ 19; M, caudal vertebra MDS-VJ 20; N, caudal vertebra MDS-VJ 23 and O,caudal vertebra MDS-VJ 28; in A, I, K, M e ventral, B, C, J, N e left lateral, D, F, L e anterior, E, G e posterior, H e dorsal, and O e right lateral views. Scale bar equals 5 mm.
A. Houssaye et al. / Cretaceous Research 41 (2013) 127e135 131

Author's personal copy
hypapophyseal peduncle fused to the centrum is observable in theanterior portion of the centrum (Fig. 2D, F). Its articular facet for thedistal element of the hypapophysis, which is not preserved, isrelatively flat and narrow, though broader anteriorly than posteri-orly (length: 4.1 mm). There does not seem to be any articularsurface for an eventual hypapophyseal peduncle located moreposteriorly, just anteriorly to the condyle (Fig. 2D). Pre-zygapophyses are much reduced but postzygapophyses are welldeveloped; the articular facets are subhorizontal (Fig. 2B, C, E).
5.2.2. Anterior cervical vertebra (MDS-VJ 2; Fig. 2GeJ)The vertebra MDS-VJ 2 corresponds to a much anterior cervical
vertebra (probably C3, as suggested by its morphology, morespecifically its paradiapophyses). A small knob, corresponding toa small hypapophysis directed ventro-posteriorly is locatedventrally just anterior to the condyle (Fig. 2G, J). Unfortunately, thepresence or absence of an articulating surface for individualizedintercentral elements on this vertebra (as well as on the othercervical vertebrae) cannot be determined (as the distal surface isslightly eroded). It is prolonged anteriorly by a slight keel until theanterior border of the centrum (Fig. 2G, J). In anterior view, thecotyle is slightly depressed (Fig. 2H). The paradiapophyses projectlaterally and posteriorly and slightly ventrally (Fig. 2G, H, J) but notbeyond the ventral border of the centrum (Fig. 2G, H); they arerelatively short dorso-ventrally and the dorsal part is notablyinclined posteriorly (Fig. 2G). They do not seem to bear any articularsurface (Fig. 2G). The neural spine rises much obliquely above theneural arch and, though broken, appears relatively short antero-posteriorly (Fig. 2G, I). The cotyleecondyle system is slightly obli-que (Fig. 2G). Prezygapophyses are strongly oblique (at about 45�;Fig. 2H). Their articular facets are large and antero-laterally directed(Fig. 2I).
5.2.3. Posterior cervical vertebrae (MDS-VJ 8,9,16,17; Fig. 3AeE)In these vertebrae, the paradiapophyses, which still project, but
to a much lesser extent, postero-laterally, extend low on thecentrum but do not project beyond its ventral border (Fig. 3D). Theyare located anteriorly, in the first half of the centrum, at the level ofthe posterior limit of the prezygapophyses and their dorsal part isstill inclined posteriorly (Fig. 3B, C). The paradiapophyses are muchelongated dorso-ventrally. The distance between the top of theparadiapophyses and the postzygapophyses is particularly short(Fig. 3B, C). When preserved, the neural spine rises much obliquelyfrom the level of the posterior limit of the prezygapophyses; itbecomes subhorizontal and thickens above the postzygapophysesand extends much more posteriorly than the posterior border ofthe neural arch and the tip of condyle (Fig. 3C); it is higher than thecentrum (about same height as between the ventral border of thecentrum and the top of the neural canal; Fig. 3C, E). It appears thatthe cotyle becomes less ovoid and more rectangular in the moreposterior cervicals, its dorsal and ventral borders becoming almoststraight and parallel (Fig. 3D). In vertebrae MDS-VJ 9 and 17, there isstill the slight keel observable in ventral view (Fig. 3A) and thereseems to have been a small knob anterior to the condyle in ventralview (Fig. 3A, C).
5.3. Dorsal vertebrae (MDS-VJ 3-7,10-15; Fig. 3FeJ)
Dorsal vertebrae are larger and more elongate than cervicals.The ventral face of the centrum is flat and it does not stronglywiden anteriorly, as a result of the relatively weak lateral projectionof the paradiapophyses (Fig. 3I). The latter are located as anteriorlybut more dorsally than in cervicals so that they are clearly wellabove the ventral border of the centrum (Fig. 3F, J). As a conse-quence, their dorso-ventral extension is much reduced (Fig. 3F, J).
This feature, in addition to the lengthening of the centrum,lengthen the distance between the dorsal border of the para-diapophyses and the postzygapophyses so that the inter-zygapophyseal ridge is more distinct (Fig. 3J). The neural spine isslightly smaller but the obliquity of the cotyleecondyle system ismuch more important than in cervicals (Fig. 3J). There is noimportant intracolumnar variability along the dorsal series. Thecentrum becomes more elongated in the more posterior dorsals,paradiapophyses become dorso-ventrally shorter and their ventralborder becomes close to the level of the dorsal limit of the cotyle.The posteriorly directed inclination of the dorsal border of theparadiapophyses increases in the more posterior dorsal vertebrae.
5.4. Caudal vertebrae (MDS-VJ 18-29; Fig. 3KeO)
There is no pygal vertebra, i.e., an anterior caudal with trans-verse processes but without haemal peduncles.
All caudal vertebrae available display both transverse processesand haemal peduncles, even the posteriormost ones. Transverseprocesses are antero-posteriorly elongated at their base (ratherthan dorso-ventrally like for paradiapophyses) and originate clearlyabove the dorsal border of the cotyle (Fig. 3K, L, N). They areespecially laterally but also slightly posteriorly directed (Fig. 3K).Along the tail, transverse processes migrate posteriorly along thecentrum and their antero-posterior extension decreases so thattheir lateral end is a thin point in more posterior caudals (Fig. 3M,N). Caudal vertebrae do not display autotomous septa. In ventralview, two ventral peduncles (Fig. 3K), onwhich articulated chevronbones, are located posteriorly, against the condyle. They are pro-longed anteriorly by keels that define a large sagittal groove(Fig. 3K). Prezygapophyses are more vertically inclined on thehorizontal than in dorsals (Fig. 3L). MDS-VJ 18 and 19, which arerelatively anterior caudal vertebrae, are much shorter and muchmore robust than posterior dorsals (Fig. 3K). Caudal vertebraebecome more and more elongated posteriorly (the width of thecentrum becoming much reduced as compared to its length;Fig. 3M). The neural spine, which is well observable in MDS-VJ 28(Fig. 3O), is relatively high (higher than the centrum) but verynarrow antero-posteriorly.
5.5. Microanatomical features
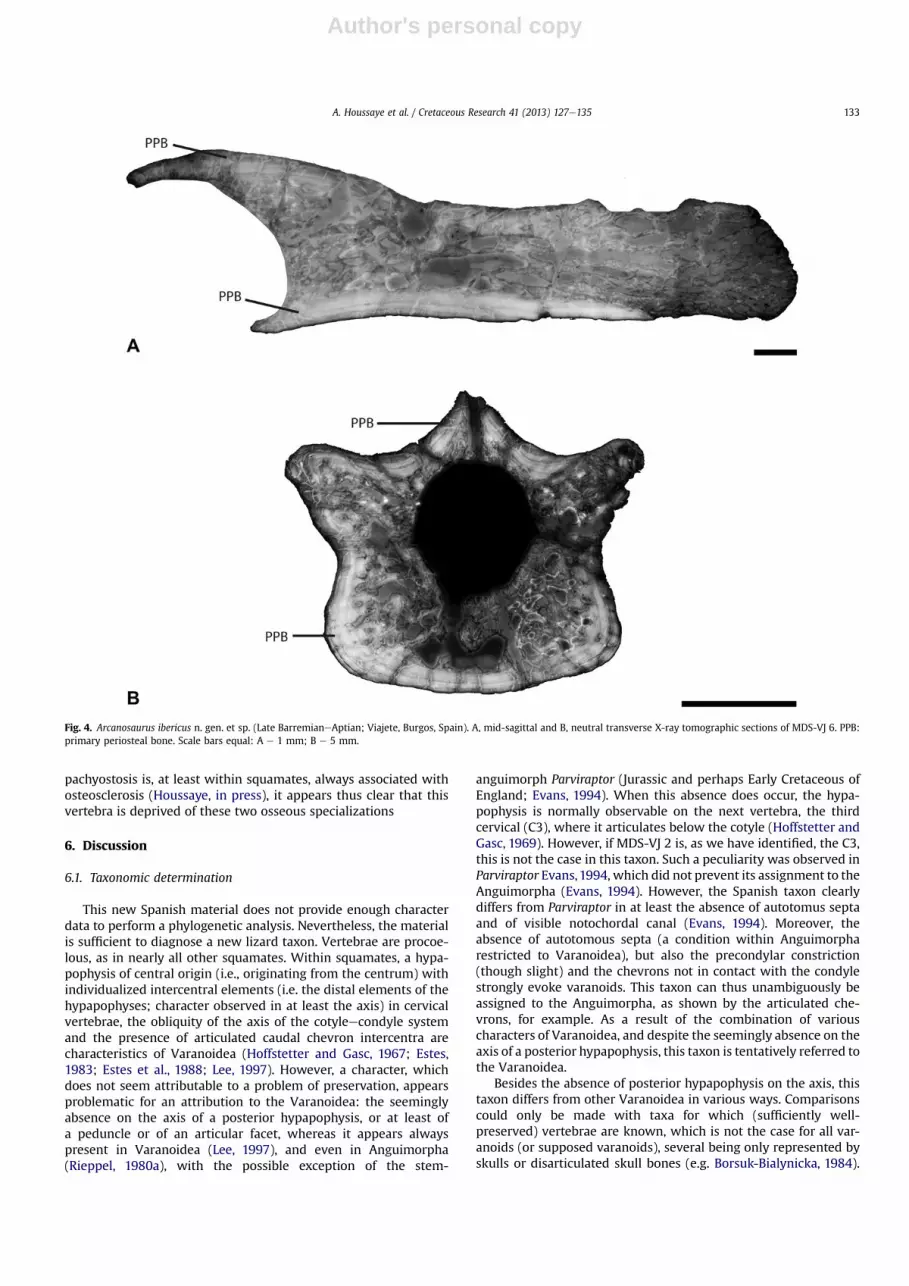
In longitudinal section, primary periosteal bone (PPB) is locatedat the ventral border of the centrum and above the cotyle. The restof the section consists of a spongiosa (Fig. 4A). Because of diagen-esis (and especially because remodeled bone and cavities filledwith sediment can not precisely be distinguished), compactnessindices could not be calculated but compactness appears similar tothat observed in extant squamates (cf. Houssaye et al., 2010). Intransverse section, a thick layer of primary periosteal bone isobserved along the bone periphery (Fig. 4B). Although the micro-anatomy is badly preserved above the centrum and around theneural canal, the vertebral organization seems similar to thepseudo-tubular structure in double rings e one peripheral and onesurrounding the neural canal, separated by trabecule, observed inextant squamates (cf. Houssaye et al., 2010; Fig. 4B).
The estimated amount of primary bone (PBCL about 23%; PBCTabout 50%) is consistent with what is observed in extant squamates(PBCL about 26%; PBCT about 51%; cf. Houssaye et al., 2010). In bothlongitudinal and transverse sections, the microanatomical organi-zation appears similar to that observed in extant squamates, withno particular thickening of the cortex, which suggests that theslight lateral thickening in ventral view observed in some vertebraedoes not correspond to pachyostosis, but is rather linked to thevertebral morphology. In addition, there is no osteosclerosis. As
A. Houssaye et al. / Cretaceous Research 41 (2013) 127e135132
Author's personal copy
pachyostosis is, at least within squamates, always associated withosteosclerosis (Houssaye, in press), it appears thus clear that thisvertebra is deprived of these two osseous specializations
6. Discussion
6.1. Taxonomic determination
This new Spanish material does not provide enough characterdata to perform a phylogenetic analysis. Nevertheless, the materialis sufficient to diagnose a new lizard taxon. Vertebrae are procoe-lous, as in nearly all other squamates. Within squamates, a hypa-pophysis of central origin (i.e., originating from the centrum) withindividualized intercentral elements (i.e. the distal elements of thehypapophyses; character observed in at least the axis) in cervicalvertebrae, the obliquity of the axis of the cotyleecondyle systemand the presence of articulated caudal chevron intercentra arecharacteristics of Varanoidea (Hoffstetter and Gasc, 1967; Estes,1983; Estes et al., 1988; Lee, 1997). However, a character, whichdoes not seem attributable to a problem of preservation, appearsproblematic for an attribution to the Varanoidea: the seeminglyabsence on the axis of a posterior hypapophysis, or at least ofa peduncle or of an articular facet, whereas it appears alwayspresent in Varanoidea (Lee, 1997), and even in Anguimorpha(Rieppel, 1980a), with the possible exception of the stem-
anguimorph Parviraptor (Jurassic and perhaps Early Cretaceous ofEngland; Evans, 1994). When this absence does occur, the hypa-pophysis is normally observable on the next vertebra, the thirdcervical (C3), where it articulates below the cotyle (Hoffstetter andGasc, 1969). However, if MDS-VJ 2 is, as we have identified, the C3,this is not the case in this taxon. Such a peculiarity was observed inParviraptor Evans,1994, which did not prevent its assignment to theAnguimorpha (Evans, 1994). However, the Spanish taxon clearlydiffers from Parviraptor in at least the absence of autotomus septaand of visible notochordal canal (Evans, 1994). Moreover, theabsence of autotomous septa (a condition within Anguimorpharestricted to Varanoidea), but also the precondylar constriction(though slight) and the chevrons not in contact with the condylestrongly evoke varanoids. This taxon can thus unambiguously beassigned to the Anguimorpha, as shown by the articulated che-vrons, for example. As a result of the combination of variouscharacters of Varanoidea, and despite the seemingly absence on theaxis of a posterior hypapophysis, this taxon is tentatively referred tothe Varanoidea.
Besides the absence of posterior hypapophysis on the axis, thistaxon differs from other Varanoidea in various ways. Comparisonscould only be made with taxa for which (sufficiently well-preserved) vertebrae are known, which is not the case for all var-anoids (or supposed varanoids), several being only represented byskulls or disarticulated skull bones (e.g. Borsuk-Bialynicka, 1984).
Fig. 4. Arcanosaurus ibericus n. gen. et sp. (Late BarremianeAptian; Viajete, Burgos, Spain). A, mid-sagittal and B, neutral transverse X-ray tomographic sections of MDS-VJ 6. PPB:primary periosteal bone. Scale bars equal: A e 1 mm; B e 5 mm.
A. Houssaye et al. / Cretaceous Research 41 (2013) 127e135 133

Author's personal copy
The absence of a zygospheneezygantrum system excludes thistaxon from Pythonomorpha (Lee, 1997).Within non pythonomorphVaranoidea, this specimen differs from Pachyvaranus (Pachyvar-anidae; Maastrichtian of Morocco) in: the absence of a dorsallynotched and subrectangularly shaped neural spine, the occurrenceof a weak precondylar constriction and the absence of pachyostosis(Houssaye et al., 2011). It differs from Varanus (Varanidae) in thesize and shape of the neural canal (bigger and rather triangular inthe former), the fact that the axis centrum is not elongated, theneural arch with no anterior depression on each side of the neuralspine, the less inclined cotyleecondyle system and the less welldeveloped precondylar constriction. It differs from Gobiderma(Monstersauria; Late Cretaceous of Mongolia and China) in theabsence of zygosphene and in the occurrence of aweak precondylarconstriction (Conrad et al., 2011), and from Necrosauridae(restricted to Necrosaurus, as originally defined) in the small size ofthe neural canal, in its more inclined and markedly lower neuralspine on dorsal vertebrae, in the ventral projection of the trans-verse processes of the axis, and in its much less elongated cervicalvertebrae (Augé, 2005). It differs from both Varanus andNecrosaurus (PaleoceneeOligocene of Europe) in the absence ofclear pars tectiformis (J.-C.R, pers. obs.). It differs from Varanus,Gobiderma, Helodermatidae and from Saniwa (Varanidae; Eoceneof North America, Europe and Asia, and perhaps late Cretaceous ofAsia and Oligocene of North America; Alifanov, 1993; Estes,1983) inthe non-subrectangular shape of the neural spine and from Helo-dermatidae and Lanthanotidae in the triangular shape of thecentrum (Rieppel, 1980b; J.-C.R., pers. obs.). It also differs fromSaniwa in that cervical vertebrae are not longer than anteriordorsals, in the length of the axis (relatively shorter in Saniwa) and inthe absence of a rudimentary pseudozygosphene. The latter wasreported in Saniwa by Hoffstetter (1968) and Rieppel and Grande(2007). Moreover, in Saniwa the axis and the third cervicalvertebra lack transverse processes (Rieppel and Grande, 2007),which is not the case here. It also differs from Telmasaurus (stem-varanid; Late Cretaceous of Mongolia), Parasaniwa (Monstersauria;CretaceouseEocene of North America) and Palaeosaniwa (Mons-tersauria; Late Cretaceous of North America) in the absence ofa rudimentary pseudozygosphene; it further differs from Para-saniwa and Palaeosaniwa in lacking depressions on each side of theneural arch, from Parasaniwa in having a precondylar constriction(Gao and Fox, 1996), and from Palaeosaniwa in its smallerzygapophyses and condyle, and the relatively thinner anterior partof the centrum resulting in less depressed vertebrae (Gilmore,1928; Estes, 1964). The poorly preserved vertebrae of the stem-helodermatid Primaderma (AlbianeCenomanian of North Amer-ica) have a centrum that markedly more widens anteriorly andparadiapophyses located more anteriorly (Nydam, 2000).
A vertebra from the late Albianeearly Cenomanian of CentralAsia appears relatively similar to that of the Burgos cervical verte-brae. It was assigned by Nessov (1988) to a Simoliophiidae, but themorphology is totally distinct from that of Simoliophis (Pachyo-phiidae; Cenomanian of Europe and North Africa; A.H. and J.C.R.;pers. obs.). However, as only the ventral view is available, thiscomparison is only speculative.
6.2. Paleoecology
The geological setting, interpreted as a floodplain, might beambiguous concerning the life environment of Arcanosaurus iber-icus, which could be either terrestrial, as for the various dinosaursfound at the same locality, or aquatic. However, no anatomicalcharacter suggests an adaptation to an aquatic life. Moreover, themicroanatomical data reveal that this taxon does not display any ofthe two adaptations to an aquatic life observed in the
microanatomy of exclusively aquatic subsurface swimmers: bonemass increase or a spongious microorganization (Ricqlès andBuffrénil, 2001; Houssaye, 2009). As a consequence, though it cannot be rejected that this taxon might have been occasionallyaquatic, its mode of life is considered essentially terrestrial.
7. Conclusion
The unique combination of characters observed in Arcanosaurusibericus supports a tentative attribution to the Varanoidea. Theanalysis of the microanatomical and geological data suggests anessentially terrestrial mode of life for this Spanish new taxon. Theoldest Varanoidea is the pythonomorph Kaganaias from theValanginianeHauterivian (Early Cretaceous) of Japan, thought tohave lived in freshwater swampy environment far from the sea(Evans et al., 2006). The other oldest varanoids are known from theLate Cretaceous, inmarine (pythonomorphs; review in Houssaye, inpress) or terrestrial (e.g. Norell et al., 1992; Gao and Fox, 1996;Nydam, 2000; Laurent et al., 2002; Pereda Suberbiola, 2009)deposits. If Arcanosaurus ibericus, from the late BarremianeAptianof the Burgos Province, indeed belongs to the Varanoidea, itwould thus be the oldest terrestrial form known to date. Thediscovery of this new taxon in Spain is thus of particular interestregarding the evolutionary history of varanoids.
Acknowledgments
We would like to thank J. Izquierdo Molinero, who discoveredthe site, the Museo de Dinosaurios de Salas de los Infantes, whichfacilitated access to the fossils as well as their transfer for thesession of photographs and the analyses carried out outside itslaboratories and Z. Herrera (Universidad de Zaragoza, Spain) for thephotographs. We are particularly grateful to P. Tafforeau (ESRF,Grenoble, France) and the ESRF for providing beamtime andsupport for the resort to synchrotron-radiation microtomography.We are thankful to M. Augé (Muséum National d’Histoire naturelle,Paris, France) and R.L. Nydam (Midwestern University, Arizona,U.S.A.) for constructive comments that improved the quality of themanuscript and to E.A.M. Koutsoukos for editorial work. A.H.acknowledges financial support from the Muséum National d’His-toire Naturelle (Special financial support for the organization of theThird Mosasaur Meeting from C. de Muizon to N. Bardet) and theA. v. Humboldt Foundation. Research of X.P.S. is supported bythe projects CGL2010e18851/BTE (Ministerio de Economía yCompetitividad of Spain) and ITe320e10 (Gobierno Vasco/EuskoJaurlaritza). The work of N.B., J.-C.R. and X.P.S. resulted froma collaboration agreement between the UPV/EHU (Bilbao), theCNRS (Paris), and the MNHN (Paris).
References
Alifanov, V.A., 1993. Some peculiarities of the Cretaceous and Paleogene lizardfaunas of the Mongolian People’s Republic. Kaupia 3, 9e13.
Arribas, J., Alonso, A., Mas, R., Tortosa, A., Rodas, M., Barrenechea, J.F., Alonso-Azcárate, J., Artigas, R., 2003. Sandstone petrography of continental depositionalsequences of an intraplate rift basin: western Cameros basin (north Spain).Journal of Sedimentary Research 73, 309e327.
Augé, M.L., 2005. Evolution des lézards du Paléogène en Europe. Mémoires duMuséum National d’Histoire Naturelle, Paris 192, 1e369.
Beuther, A., 1966. Geologische Untersuchungen in Wealden und Utrillas-Schichtenin Westteil der Sierra de los Cameros (Nordwestliche Iberische Ketten). Bei-hefte zum Geologischen Jahrburch 44, 103e121.
Bolet, A., Evans, S.E., 2010. A new lizard from the early Cretaceous of Catalonia(Spain), and the Mesozoic lizards of the Iberian Peninsula. Cretaceous Research31, 447e457.
Borsuk-Bialynicka, M., 1984. Anguimorphans and related lizards from the LateCretaceous of the Gobi Desert, Mongolia. Palaeontologica Polonica 46, 5e105.
A. Houssaye et al. / Cretaceous Research 41 (2013) 127e135134

Author's personal copy
Buffrénil, V. de, Bardet, N., PeredaSuberbiola, X., Bouya, B., 2008. Specializationof bonestructure in Pachyvaranus crassispondylus Arambourg, 1952, an aquatic squamatefrom the Late Cretaceous of the southern Tethyan margin. Lethaia 41, 59e69.
Canudo, J.I., Gasca, J.M., Aurell, M., Badiola, A., Blain, H.-A., Cruzado-Caballero, P.,Gómez-Fernández, D., Moreno-Azanza, M., Parrilla, J., Rabal-Garcés, R., Ruiz-Omeñaca, J.I., 2010. La Cantalera: an exceptional window onto the vertebratebiodiversity of the Hauterivian-Barremian transition in the Iberian Peninsula.Journal of Iberian Geology 36, 205e224.
Clemente, P., 2010. Review of the Upper Jurassic-Lower Cretaceous stratigraphy inWestern Cameros basin, Northern Spain. Revista de la Sociedad Geológica deEspaña 23, 101e143.
Conrad, J.C., Rieppel, O., Gauthier, J.A., Norell, M.A., 2011. Osteology of Gobidermapulchrum (Monstersauria, Lepidosauria, Reptilia). Bulletin of the AmericanMuseum of Natural History 362, 1e88.
Estes, R., 1964. Fossil vertebrates from the Late Cretaceous Lance formation, easternWyoming. University of California Publications in Geological Sciences 49, 1e180.
Estes, R., 1983. The fossil record and early distribution of lizards. In: Rhodin, A.,Miyata, K. (Eds.), Advances in Herpetology and Evolutionary Biology. HarvardUniversity, Cambridge, pp. 365e398.
Estes, R., De Queiroz, K., Gauthier, J., 1988. Phylogenetic relationships withinSquamata. In: Estes, R., Pregill, G. (Eds.), Phylogenetic Relationships of the LizardFamilies: Essays Commemorating C.L. Camp. Stanford University Press, Stan-ford, pp. 119e281.
Evans, S.E., 1994. A new anguimorph lizard from the Jurassic and Lower Cretaceousof England. Palaeontology 37, 33e49.
Evans, S.E., Barbadillo, L.J., 1997. Early Cretaceous lizards from Las Hoyas, Spain.Zoological Journal of the Linnean Society 119, 1e27.
Evans, S.E., Barbadillo, L.J., 1998. An unusual lizard (Reptilia: Squamata) from theEarly Cretaceous of Las Hoyas, Spain. Zoological Journal of the Linnean Society124, 235e266.
Evans, S.E., Barbadillo, L.J., 1999. A short-limbed lizard from the Lower Cretaceous ofSpain. Special Papers in Palaeontology 60, 73e85.
Evans, S.E., Manabe, M., Noro, M., Isaji, S., Yamaguchi, M., 2006. A long-bodied lizardfrom the Lower Cretaceous of Japan. Palaeontology 49, 1143e1165.
Fürbringer, M., 1900. Zur vergleichenden Anatomie des Brustschulterapparates undder Schultermuskeln. Jenaische Zeitschrift 34, 215e718.
Gao, K.-Q., Fox, R.C., 1996. Taxonomy and evolution of Late Cretaceous lizards(Reptilia: Squamata) from western Canada. Bulletin of the Carnegie Museum ofnatural History 33, 1e107.
Gilmore, C.W., 1928. Fossil lizards of North America. Memoirs of the NationalAcademy of Sciences 22, 1e201.
Gray, J.E., 1827. Reptilia. In: King, P.P. (Ed.), Narrative of a survey of the intertropicaland western coasts of Australia. Performed between the years 1818 and 1822,with an appendix containing various subjects. J. Murray, London, pp. 424e434.
Hoffstetter, R., 1968. Présence de Varanidae (Reptilia, Sauria) dans le Miocène deCatalogne. Considérations sur l’histoire de la famille. Bulletin du MuséumNational d’Histoire Naturelle 40, 1051e1064.
Hoffstetter, R., Gasc, J.P., 1967. Observations sur le squelette cervical et spécialementsur les hypapophyses des sauriens varanoides actuels et fossiles. Bulletin duMuséum National d’Histoire Naturelle 39, 1028e1043.
Hoffstetter, R., Gasc, J.-P., 1969. Vertebrae and ribs of modern Reptiles. In:Gans, C. (Ed.), Biology of the Reptilia. Academic Press, London and New York,pp. 201e310.
Houssaye, A., 2009. “Pachyostosis” in aquatic amniotes: a review. IntegrativeZoology 4, 325e340.
Houssaye, A. Palaeoecological and morphofunctional interpretation of bone massincrease: an example in Late Cretaceous shallow marine squamates. BiologicalReviews, in press.
Houssaye, A., Buffrénil, V. de, Rage, J.-C., Bardet, N., 2008. An analysis of vertebral‘pachyostosis’ in Carentonosaurus mineaui (Mosasauroidea, Squamata) fromthe Cenomanian (early Late Cretaceous) of France, with comments on itsphylogenetic and functional significance. Journal of Vertebrate Paleontology28, 685e691.
Houssaye, A., Mazurier, A., Herrel, A., Volpato, V., Tafforeau, P., Boistel, R.,Buffrénil, V. de, 2010. Vertebral microanatomy in squamates: structure, growthand ecological correlates. Journal of Anatomy 217, 715e727.
Houssaye, A., Bardet, N., Rage, J.-C., Pereda Suberbiola, X., Amaghzaz, M., Amalik, M.,2011. A review of Pachyvaranus crassispondylus Arambourg, 1952, a pachyostoticmarine squamate from the latest Cretaceous Phosphates of the margin of theSouthern Tethys. Geological Magazine 148, 237e249.
Huerta, P., Fernández-Baldor, F.T., Farlow, J.O., Montero, D., 2012. Exceptionalpreservation processes of 3D dinosaur footprint casts in Costalomo (LowerCretaceous, Cameros Basin, Spain). Terra Nova 24, 136e141.
Kuhn, O., 1958. Ein neuer lacertilier aus dem fränkischen Lithographieschiefer.Neues Jahrbuch für Geologie und Paläontologie, Monatshefte 1958, 437e440.
Laurent, Y., Bilotte, M., Le Loeuff, J., 2002. Late Maastrichtian continental vertebratesfrom southwestern France: correlation with marine fauna. Palaeogeography,Palaeoclimatology, Palaeoecology 187, 121e135.
Lee, M.S.Y., 1997. The phylogeny of varanoid lizards and the affinities of snakes.Philosophical Transactions of the Royal Society 352, 53e91.
Linnaeus, C., 1758. Systema naturae per regna tria naturae, secundum classes,ordines, genera, species, cum characteribus, differentiis, synonymis, locis.Tomus I. Edito decima, reformata. Holmiae, 823 pp.
Martín-Closas, C., Alonso Millán, A., 1998. Estratigrafía y bioestratigrafía(Charophyta) del Cretácico inferior en el sector occidental de la Cuenca deCameros (Cordillera Ibérica). Revista de la Sociedad Geológica de España 11,253e270.
Mas, R., Alonso, A., Guimerá, J., 1993. Evolución tectonosedimentaria de una cuencaextensional intraplaca: la cuenca finijurásica-eocretácica de Los Cameros (LaRioja e Soria). Revista de la Sociedad Geológica de España 6, 129e144.
Mas, J.R., Moreno, B., Isabel, M., Arribas Mocoroa, J., Serrano, A., Guimera Rosso, J.,Alonso Millán, A., Alonso Azcárate, J., 2002. La Cuenca de Cameros: desde laextensión Finijurásica-Eocretácica a la inversión terciaria-implicaciones en laexploración de hidrocarburos. Zubia Monográfico 14, 9e64.
Nessov, L.A., 1988. Late Mesozoic amphibians and lizards of Soviet Middle Asia. ActaZoologica Cracoviensia 31, 475e486.
Norell, M.A., McKenna, M.C., Novacek, M.J., 1992. Estesia mongoliensis, a new fossilvaranoid from the Late Cretaceous Barun Goyot Formation of Mongolia.American Museum Novitates 3045, 24.
Nydam, R.L., 2000. A new taxon of helodermatid-like lizard from the Albian-Cenomanian of Utah. Journal of Vertebrate Paleontology 20, 285e294.
Oppel, M., 1811. Die Ordnungen, Familien und Gattungen der Reptilien als Prodromeiner Naturgeschichte derselben. Lindauer, München, 87 pp.
Pereda Suberbiola, X., 2009. Biogeographical affinities of Late Cretaceous conti-nental tetrapods of Europe: a review. Bulletin de la Société Géologique deFrance 180, 57e71.
Pereda Suberbiola, X., Torcida Fernández-Baldor, F., Izquierdo, L.A., Huerta, P.,Montero, D., Pérez, G., 2003a. First rebbachisaurid dinosaur (Sauropoda, Dip-lodocoidea) from the Early Cretaceous of Spain: palaeobiogeographical impli-cations. Bulletin de la Société Géologique de France 174, 471e479.
Pereda Suberbiola, X., Galton, P.M., Torcida Fernández-Baldor, F., Huerta, P.,Izquierdo, L.A., Montero, D., Pérez, G., Urién, V., 2003b. First StegosaurianDinosaur remains from the Early Cretaceous of Burgos (Spain), with a review ofCretaceous stegosaurs. Estudios Geológicos 55, 267e272.
Pereda Suberbiola, X., Ruiz-Omeñaca, J.I., Canudo, J.I., Torcida, F., Sanz, J.L., 2012.Dinosaur faunas from the Early Cretaceous (Valanginian-Albian) of Spain. In:Godefroit, P. (Ed.), Bernissart Dinosaurs and Early Cretaceous Ecosystems.Indiana University Press, Bloomington & Indianapolis, pp. 378e407.
Pérez-García, A., Murelaga, X., Huerta, P., Torcida Fernández-Baldor, F., 2012. Turtlesfrom the Lower Cretaceous of the Cameros Basin (Iberian Range, Spain).Cretaceous Research 33, 146e158.
Rage, J.-C., Néraudeau, D., 2004. A new pachyostotic squamate reptile from theCenomanian of France. Palaeontology 47, 1195e1210.
Ricqlès, A. de, Buffrénil, V. de, 2001. Bone histology, heterochronies and thereturn of tetrapods to life in water: where are we? In: Mazin, J.M.,Buffrénil, V. de (Eds.), Secondary Adaptation of Tetrapods to life in Water.München, pp. 289e310.
Rieppel, O., 1980a. The phylogeny of anguinomorph lizards. Denkschriften derSchweizerischen Naturforschenden Gesellschaft 94, 1e86.
Rieppel, O., 1980b. The postcranial skeleton of Lanthanotus borneensis (Reptilia,Lacertilia). Amphibia-Reptilia 1, 95e112.
Rieppel, O., Grande, L., 2007. The anatomy of the fossil varanid lizard Saniwa ensi-dens Leidy, 1870, based on a newly discovered complete skeleton. Journal ofPaleontology 81, 643e665.
Salas, R., Casas, A., 1993. Mesozoic extensional tectonics, stratigraphy and crustalevolution during the Alpine cycle of the eastern Iberian basin. Tectonophysics228, 33e35.
Salas, R., Guimerá, J., Mas, R., Martín-Closas, C., Meléndez, A., Alonso, A., 2001.Evolution of the Mesozoic Central Iberian Rift System and its Cainozoic inver-sion (Iberian chain). In: Ziegler, P.A., Cavazza, W., Robertson, A.H.F., Crasquin-Soleau, S. (Eds.), Peri-Tethys Memoir: 6, Peri-Thetyan Rift/Wrench Basins andPassive Margins. Muséum national d’histoire naturelle, Paris, pp. 145e185.
Scanlon, J.D., Lee, M.S.Y., Archer, M., 2003. Mid-Tertiary elapid snakes (Squamata,Colubroidea) from Riversleigh, northern Australia: early steps in a continent-wide adaptive radiation. Geobios 36, 573e601.
Schudack, U., Schudack, M., 2009. Ostracod biostratigraphy in the Lower Cretaceousof the Iberian chain (eastern Spain). Journal of Iberian Geology 35, 141e168.
Torcida Fernández-Baldor, F., 2006. Restos directos de dinosaurios en Burgos(Sistema Ibérico): un balance provisional. In: Colectivo Arqueológico-Pale-ontológico de Salas (Ed.), III Jornadas Internacionales sobre Paleontología delos Dinosaurios y su Entorno. Actas, Salas de los Infantes, Burgos, Spain,pp. 105e128.
Torcida Fernández-Baldor, F., Canudo, J.I., Huerta, P., Montero, D., PeredaSuberbiola, X., Salgado, L., 2011. Demandasaurus darwini, a new rebbachisauridsauropod from the early Cretaceous of the Iberian Peninsula. Acta Palae-ontologica Polonica 56, 535e552.
A. Houssaye et al. / Cretaceous Research 41 (2013) 127e135 135












![Dientes de dinosaurios en el Barremiense de Allepuz, Teruel [Barremian dinosaur teeth from Allepuz, Teruel, Spain]](https://img.pdfslide.net/doc/110x75/635ccac1a3fa66b45c0e162b/dientes-de-dinosaurios-en-el-barremiense-de-allepuz-teruel-barremian-dinosaur.jpg)
















