Embed Size (px)
Citation preview

CURRENT THERAPEUTIC RESEARCH@ VOL. 56, NO. 2, FEBRUARY 1995
A PLASMID MEDIATING PRODUCTION OF A BETA-LACTAMASE BY STENOTROPHOMONAS MALTOPHIL.lA*
MICHAEL D. KELLY,l JOEL E. MORTENSEN,2t3 BARBARA A. KONKLE,l AND TERRENCE L. STULL*
‘Cardeza Foundation for Hematological Research, Thomas Jefferson University, 2Section of Infectious Diseases, Department of Pediatrics, Temple University School of Medicine,
3Department of Laboratories, St. Christopher~s Hospital for Children, and 4Department of Pediatrics and Mi~robio~ogyfImmunology, Medical College of Pennsylvania,
Philadelphia, Pennsylvania
ABSTRACT
To delineate the mechanisms that contribute to the high level of an- timicrobial resistance often demonstrated by Ste~ot~o~~orno~~ mal- to~~~Z~~, DNA was purified from five clinical isolates and analyzed for the presence of plasmid DNA. Purified plasmid DNA from a single isolate contained a 6.5-kb plasmid, designated pXM222, and a 5.6-kb plasmid, designated pTHB. Transformation of pTHB into ~~c~er~c~~~ coli K-12 HB101 resulted in the expression of resistance to the peni- cillins and to cefazolin.
INTRODUCTION
Xunthomonas maltophilia, recently reclassified as Stenotrophomonas mal- tophiZia,l has been isolated from river and well water, raw milk, rabbit feces, and frozen fish? This species is a common commensal, an occasional contaminant of clinical specimens, and part of the transient flora of hos- pitalized patients3 Although it has been documented as a cause of bacter- emia, urinary tract infections, respiratory tract infections, meningitis, serious wound infections, mastoiditis, epididymitis, conjunctivitis, and en- docarditis, S maltophilia is isolated predominantly from respiratory tract secretions.4-7
The reservoir of these organisms and their method of transmission to patients remain unknown3 S m~ZtophiZi~ is generally acquired in the hospital and is most commonly isolated from patients who have serious underlying diseases or have had tracheostomy, endotracheal intubation, or genitourinary instrumentation. Klinger and Thomassen5 showed that the
robial Resistance- A Crisis in Health
of Laboratories 19134-1095.
, St. Christopher’s Hos-
152

M. D. KELLY Err AL.
frequency of isolation of S maltophilia from respiratory tract cultures from patients with cystic fibrosis is increasing. In addition, the use of empiric, broad-spectrum antimicrobiai therapy for febrile neutropenic patients has emerged as a significant predisposing factor to infections due to S maltophilia, s
Resistance of S maltophilia to beta-lactam drugs had been attributed to low outer-membrane permeability and production of two chromosome- encoded beta-lactamases, s-12 The outer membrane of S maltophilia is rel- atively impermeable to numerous agents, a characteristic that prevents antimicrobial drugs from reaching their target site. Saino et al z1'12 de- scribed two separate beta-lactamases, LI and L2, produced by S malto- philia. L1 is a.zinc-containing penicillinase that hydrolyzes imipenem and is not inhibited by clavulanic acid. L2 is a cephalosporinase that is active against ceftizoxime and aztreonam, both known to have considerable sta- bility in the presence of other beta-lactamases. Iz'Is L2 is not active against imipenem and is inhibited by clavulanic acid and sulbactam. Production of L1 and L2 beta-lactamases is rapidly derepressed when S maltophilia is grown in the presence of beta-lactams. Although movement of resistance markers using conjugation experiments on filter paper has been re- ported, z3 plasmids have not been documented in S maltophilia.
In this study we analyzed clinical and environmental isolates of S maltophilia for the presence of plasmid DNA. Recovery of plasmids sug- gested a mechanism for the coding and transfer of antimicrobial resistance determinants. We report here the isolation ofa 6.5-kb plasmid and a 5.6-kb plasmid from S maltophilia. Characterization of one of these plasmids was performed by transformation, restriction endonuclease, and antimicrobial susceptibility studies.
MATERIALS AND METHODS
Random stock isolates of S maltophilia were obtained from a collection at St. Christopher's Hospital for Children (isolates #40, 42, 176, 222), and the American Type Culture Collection (ATCC 13637) (isolate #1). All strains were identified using the Uni-N/F-Tek System (Remel Inc., Lenexa, Kan- sas). Competent strains of Escherichia coli K-12 HB101 were used for transformation experiments (Promega Corp., Madison, Wisconsin). All strains were subcultured and maintained on Luria-Bertani (LB) agar (Difco Laboratories, ' Inc., Detroit, Michigan) at 37 °C. Transformants were selected on LB agar containing 50 ttg/mL of ampicillin (Sigma Diagnostics, St. Louis, Missouri).
Plasmid DNA was isolated by using the rapid alkaline extraction method as described by Birnboim, z4 with some modifications. 15 Cultures from single colonies were grown for 18 to 24 hours to an optical density of
153

BETA-LACTAMASE PRODUCTION BY STENOTROPHOMONAS MALTOPHILIA
0.3 to 0.7 at 540 nm. One milliliter of the culture was centrifuged, and the pelleted cells were resuspended in 200 ~L of 50 mM glucose, 10 mM ethylene diamine-tetraacetic acid (EDTA) (pH 8.0), 25 mM TRIS (pH 8.0), and 1 mg/mL lysozyme (Fisher Scientific Co., Pittsburgh, Pennsylvania) on ice for 5 minutes. A total of 400 ~L of sodium dodecyl sulfate (SDS) was added to the sample, which was then mixed by inversion and placed on ice for 5 minutes; 300 ~L of a high-salt solution (3 M potassium acetate, 1 M formic acid) was added. After placing the sample on ice for 15 minutes, it was centrifuged for 5 minutes, and 600 ~LL of the supernatant was trans- ferred to a fresh tube. The supernatant was treated with 2.5 ~L of RNAse A (10 mg/mL) for 15 minutes at 37 °C. The sample was extracted with an equal volume of phenol/chloroform/isoamyl alcohol (25:24:1), and the DNA was recovered via ethanol precipitation. After centrifugation, the pellet was resuspended in 100 ~L of acetate-3-[N-morpholino] propanesulfonic acid (MOPS) (0.1 M sodium acetate, 0.05 M MOPS, adjusting the pH to 8.0 with sodium hydroxide) and reprecipitated with ethanol. This washing was repeated, and the final pellet was resuspended in 40 ~L of TRIS EDTA buffer (pH 7.5).
To provide highly purified plasmid DNA for transformation experi- ments, large-scale plasmid preparations were performed via the "lysis by alkali" method 16 with the following modifications: S rnaltophilia cultures were grown in 1 L of LB broth with 5 ~g/mL ampicillin at 30 °C for 48 hours, shaking at 100 rpm. The DNA was purified by equilibrium centrif- ugation in continuous cesium chloride/ethidium bromide gradients. The ethidium bromide was removed from the purified DNA by extraction with an equal volume of 1-butanol.
Restriction endonuclease digestion of the resulting plasmid DNA was performed according to the manufacturer's instructions. The DNA was subjected to electrophoresis through 1% agarose (Gibco BRL Life Technol- ogies, Gaithersburg, Maryland) at 100 V, 40 mA for 1 to 3 hours at room temperature in TBE buffer (89 mM TRIS base, 89 mM boric acid, 2.5 mM disodium EDTA; pH 8.0). The molecular size of the plasmid DNA frag- ments was calculated by comparison of endonuclease digested preparations with HindIII fragments of lambda DNA and HaeIII fragments of phiX174 RF DNA (New England Biolabs, Inc., Beverly, Massachusetts). Double enzyme digests were used for mapping experiments. 16
For bacterial transformation experiments, competent E coli K-12 HB101 cells were incubated with plasmid DNA on ice for 15 minutes, then placed at 37 °C for 5 minutes. One milliliter of LB medium was added, and the sample was returned to 37 °C, shaking at 225 rpm for 1 hour to allow for resistance expression. The volume was increased to 5 mL with LB medium, and the sample was filtered through a nitrocellulose membrane (0.45 ~m, 82 mm diameter) (Schleicher & Schuell, Inc., Keene, New Hamp- shire). The filters were placed on LB agar plates containing 50 ~g/mL
1 5 4

M, D. KELLY ET AL.
ampicillin and incubated 18 to 24 hours at 37 °C. pGEM plasmid 7zf(- ) was used as a control to test bacterial cell competency, and untransformed E coli K-12 HB101 served as a negative control.
For Southern blotting experiments, DNA was transferred to a nylon membrane (Hybond-N; Amersham Corp., Arlington Heights, Illinois). The DNA was fixed to the membrane with ultraviolet irradiation. Blots were prehybridized in 1 M sodium chloride, 0.1% SDS, 1.5 mg/mL herring sperm DNA, and 10% dextran for 3 hours at 65 °C. The blots were then hybridized with the appropriate radiolabeled probe at 65 °C for 18 to 24 hours in the prehybridization solution with the addition of 1.5 mg/mL herring sperm DNA. The blots were washed in high stringency conditions--0.1X SSC (2.25 M sodium chloride, 0.225 M sodium citrate), 0.1% SDS, I mM EDTA (pH 8.0), 10 mM sodium phosphate (pH 6.8) at 65 °C--and analyzed by using autoradiography. The probes were labeled with 32p deoxycytidine triphosphate (Dupont New England Nuclear Research Products, Boston, Massachusetts) by using random hexamer priming. 17
For susceptibility testing, cultures were grown on Mueller-Hinten agar at 37 °C. The minimum inhibitory concentrations (MICs) were deter- mined by using a broth microdilution method with the UniSept MIC an- timicrobial susceptibility test system (Analytab Products, Plainview, New York) following the information on the package insert. Pseudomonas aeruginosa ATCC 27853 and Staphylococcus aureus ATCC 29213 were used as controls during each testing procedure. The cefinase test (Becton Dickinson Microbiology Systems, Cockeysville, Maryland) was used to de- tect beta-lactamase production employing standard methods.
R E S U L T S
Primary plasmid mini-preparations were performed on five laboratory stock strains of S maltophilia. Sample #222 contained a unique band with a mobility consistent with approximately 6.5 kb plasmid DNA (Figure 1). Analysis after restriction endonuclease digestion with Bgl I, Hanf I, Rsa I, and Sty I revealed a plasmid, pXM222, of 6.5 kb.
Competent E coli K-12 HB101 cells were transformed with purified plasmid DNA from strain #222, and ampicillin-resistant colonies were used for plasmid mini-preparations. In contrast to the 6.5-kb plasmid in the S maltophilia host, agarose gel electrophoresis of plasmid preparations from resistant colonies revealed a plasmid, designated pTHB, of approxi- mately 5.6 kb (Figure 2). Endonuclease digestion analysis of the two plas- mids revealed different banding patterns (Figure 3). To determine if the plasmids shared homology, the DNA was subjected to Southern blotting and probed with pTHB. The autoradiograph demonstrated that none of the pXM222 bands shared homology with pTHB (Figure 4). However, the lane containing pXM222 also contained hybridization bands with a pattern
155

BETA-LACTAMASE PRODUCTION BY STENOTROPHOMONAS MALTOPHILIA
23.0 - 9 . 1 - 6 . 5 -
4 . 3 -
1 40 42 176 222 " 0 J I
¢ " i
STUDY STOCK #
Figure 1. Agarose gel (1%) electrophoresis of DNA preparations from isolates of Stenotroph- omonas maltophilia. A HindIII digest of lambda phage DNA is shown as a molec- ular weight standard.
identical to pTHB, suggesting that the plasmid preparations from S mal- tophilia contained at least two plasmids, To investigate this hypothesis, the plasmid preparation from S maltophilia was subjected to agarose gel electrophoresis. Although there was no detectable band comigrating with pXM222, the putative location of pTHB was adjacent to pXM222. This adjacent area was cut out and subjected to electroelution. E coli K-12 HB101 was transformed with the resultant solution. Ampicillin-resistant transformants were grown for large-scale plasmid preparations. The re- sulting plasmid DNA had the same banding pattern as pTHB, and the restriction map confirms that the size of pTHB is approximately 5.6 kb (Figure 5).
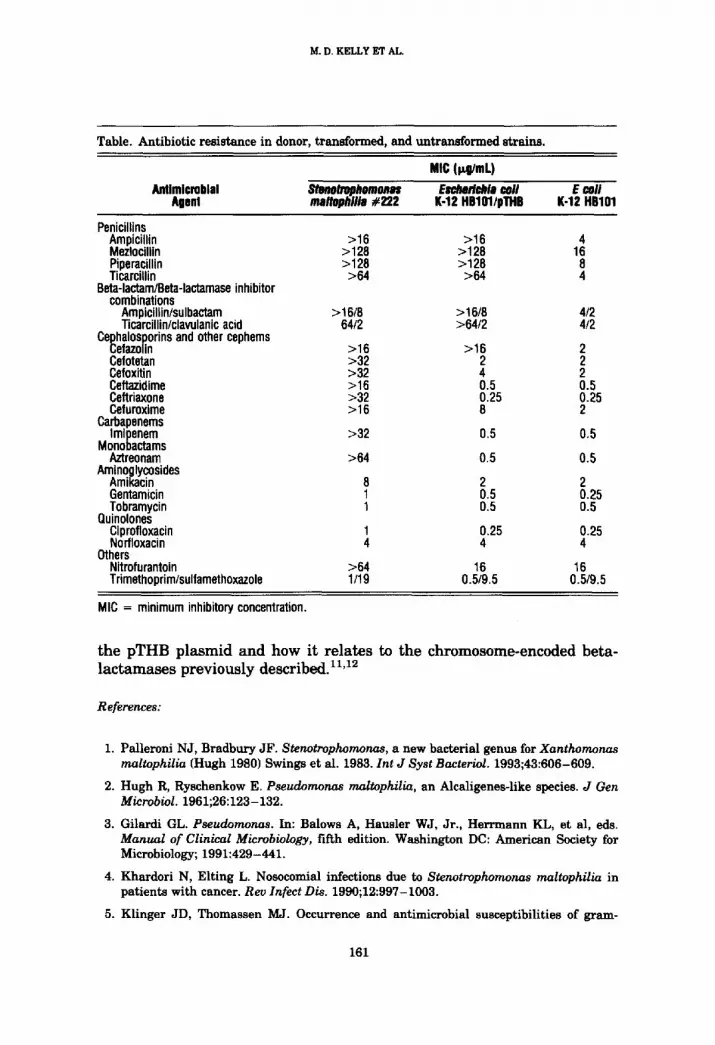
Beta-lactamase activity was detected in the transformed E coli using the cefinase test. The MIC values of various antimicrobial agents for the three experimental strains are listed in the table. The MICs for HB101/
156

M. D. KELLY ET AL.
23 .0 - 9 . 1 - 6 . 5 -
4 . 3 -
04 CD 04 "1"
"O 04 b- r- ~ O. " r X
Figure 2. Comparison of plasmids pXM222 and pTHB in 1% agarose electrophoresis gel. Lane 1 = lambda phage DNA digested with HindIII; lane 2 = pXM222, 6.5 kb; lane 3 = pTHB, 5.6 kb.
pTHB were higher for the penicillins and cefazolin compared with the untransformed HB101. The susceptibility patterns of these two strains were otherwise similar. The penicillin and cefazolin resistance values ac- quired by the transformed E coli K-12 HB101 were comparable to those of S maltophilia #222.
DISCUSSION
The resistance of S maltophilia to beta-lactam antibiotics has been previ- ously a t t r ibuted to two chromosome-encoded beta-lactamases. Mecha- nisms of S maltophilia resistance to aminoglycosides and other classes of antibiotics are less well delineated. Plasmid-mediated resistance to ami- noglycosides has been described; however, the isolation and characteriza- tion of an element responsible for the resistance were not addressed. 12
157

BETA-LACTAMASE PRODUCTION BY STENOTROPHOMONAS MALTOPHILIA
I | I I I i
X C I .
Figure 3. Endonuclease digestion ofpXM222 and pTHB in 1% agarose electrophoresis gel a t 100 V, 40 mA for 2 hours.
We isolated plasmid DNA from S maltophilia to begin characterizing the mechanisms that contribute to the widespread antimicrobial resis- tance observed in this species. A 6.5-kb plasmid was recovered from an environmental isolate of S maltophilia. Selection of competent E coli K-12 HB101 cells transformed with this 6.5-kb plasmid yielded a 5.6-kb plasmid conferring ampicillin resistance. Comparison of the two plasmids by re- striction endonuclease digestion and Southern blot analysis indicated that they are distinct. The initial plasmid preparation contained two plasmids, the 6.5-kb and 5.6-kb species, and only the 5.6-kb species was expressed in the transformed E coli K-12 HB101 cells.
Plasmid DNA of a comparable size has been consistently isolated in a study of Pseudomonas cepacia strains. 15 Transformed E coli K-12 HB101
158

M. D. KELLY ET AL.
cs~ ~ m~ l l l,,, I
I t
cq O4
X
Figure 4. Southern blotting analysis of plasmid preparation from SCermtrophorrmno~ roxdto- phili~ and pTttB (Figure 3) probed with 32p deoxyeytidine triphosphat~-labeled pTI-IB isolated from transformed Escheriehia eoli K-12 HB101. pTHB eomigrates with the hybridization band of S maltophilia plasmid preparation in Figure 3.
that demonstrated ampicillin and tetracycline resistance, however, yielded a larger plasmid DNA. Is
In the current study, antimicrobial susceptibilities of S maltophilia, E coli HB101/pTHB, and E coli K-12 HB101 demonstrated that pTHB en- codes for a beta-lactamase with primary activity against penicillins and cefazolin. The pattern of susceptibilities suggests that this enzyme is re- lated to the L1 penicillinase previously reported by Saino et al. 11 The pTHB beta-lactamase activity varies from that reported by Saino et a111 however, in that it has no activity against imipenem, suggesting that it is a new type ofS maltophilia beta-lactamase. Saino et a111 reported a lack of
159

BETA-LACTAMASE PRODUCTION BY STENOTROPHOMONAS MA~TOFH~~A
Figure 5. Restriction endonuclease map of plasmid pTHB.
PLASMID
a close relationship between the kinetic constants of Ll beta-lactamase for various beta-la&am antibiotics, and differences in the drug resistance lev- els of the Ll-producing strains. A study by Akova et al9 showed that either inducible or high-level constitutive expression of Ll and L2 beta- lactamase contributed to the insensitivity of S ~~Z~o~~~~~. The authors also showed that these enzymes are not the only factors involved in the resistance of the organism, since the basal mutant strains 7-def and 9-def retained substantial resistance to many penicillins and cephalosporins in spite of their loss of these two beta-lactamases. An explanation for this level of resistance to beta-la&am antibiotics may reside in the presence of an element such as the pTHB plasmid. The beta-lactamase product(s) en- coded by the pTHB plasmid may extend the resistance spectrum beyond the chromosome-encoded beta-lactamases in S m~~tophi~ia.
Future studies of the resistance mechanisms of S maltophilia will include further characterization of the beta-lactamase enzyme encoded by
160
M. D. KELLY ET AL.
Table. Antibiotic resistance in donor, transformed, and untransformed strains.
MIC (;zg/mL)
Angmicroblal $tenotrophornons Esclmrlcbla wli E coil Agent malWhilla #222 K-12 HBIO1/pTHB K-12 HBI01
Penicillins Ampicillin >16 >16 4 Mezlocillin >128 >128 16 Piperacillin >128 >128 8 Ticarcillin >64 >64 4
Beta-lactam/Beta-lactamase inhibitor combinations
Ampicillin/sulbactam >16/8 >16/8 4/2 Ticarcillin/clavulanic acid 64/2 >64/2 4/2
Cephalosporins and other cephems Cefazolin >16 >16 2 Cefotetan >32 2 2 Cefoxitin >32 4 2 Ceftazidime > 16 0.5 0.5 Ceftriaxone >32 0.25 0.25 Cefuroxime >16 8 2
Carbapenems Imipenem >32 0.5 0.5
Monobactams Aztreonam >64 0.5 0.5
Aminoglycosides Amikacin 8 2 2 Gentamicin 1 0.5 0.25 Tobramycin 1 0.5 0.5
Quinolones Ciprofloxacin 1 0.25 0.25 Norfloxacin 4 4 4
Others Nitrofurantoin >64 16 16 Trimethoprim/sulfamethoxazole 1/19 0.5/9.5 0.5/9.5
MIC = minimum inhibitory concentration.
the pTHB plasmid and how it relates to the chromosome-encoded beta- lactarnases previously described, ii'i2
References:
1. Palleroni NJ, Bradbury JF. Stenotrophomonas, a new bacterial genus for Xanthomonas maltophilia (Hugh 1980) Swings et al. 1983. Int J Syst Bacteriol. 1993;43:606-609.
2. Hugh R, Ryschenkow E. Pseudomonas maltophilia, an Alcaligenes-like species. J Gen Microbiol. 1961;26:123-132.
3. Gilardi GL. Pseudomonas. In: Balows A, Hausler WJ, Jr., Herrmann KL, et al, eds. Manual of Clinical Microbiology, fifth edition. Washington DC: American Society for Microbiology; 1991:429-441.
4. Khardori N, Elting L. Nosocomial infectious due to Stenotrophomonas maltophilia in patients with cancer. Rev Infect Dis. 1990;12:997-1003.
5. Klinger JD, Thomassen MJ. Occurrence and antimicrobial susceptibilities of gram-
161

BETA-LACTAMASE PRODUCTION BY STENOTROPHOMONAS MALTOPHILIA
negative nonfermentative bacilli in cystic fibrosis patients. Diagn Microbiol Infect Dis. 1985;3:149-158.
6. Yu VL, Rumans LW, Wing WJ, et al. Pseudomonas maltophilia causing heroin- associated infective endocarditis. Arch Intern Med. 1989;138:1667-1671.
7. Zuravleff J, Yu Y. Infections caused by Pseudomonas maltophilia with emphasis on bacteremia: Case reports and a review of the literature. Rev Infect Dis. 1982;4:1236- 1246.
8. Garcia-Rodriguez JA, Garcia-Sanchez JE. Antibiotic susceptibility profile of Stenotroph- omonas maltophilia. Diagn Microbiol Infect Dis. 1991;14:239-243.
9. Akova M, Bonfiglio G, Livermore DM. Susceptibility to ~-lactam antibiotics of mutant strains of Stenotrophomonas maltophilia with high and low level constitutive expression of L1 and L2 ~-lactamases. J Med Microbiol. 1991;35:208-213.
10. Mett H, Rosta S, Schacher B, Frei R. Outer membrane permeability and ~-lactamase content in Pseudomonas maltophilia clinical isolates and laboratory mutants. Rev Infect Dis. 1988;10:765-769.
11. Saino Y, Inoue M, Mitsuhashi S. Purification and properties of an inducible cephalospo- rinase from P seudomonas maltophilia GN12873. A ntirnicrob Agents C hemother. 1984;25: 362-365.
12. Saino Y, Kobayashi F, Inoue M, Mitsuhashi S. Purification and properties of inducible penicillin fl-lactamases isolated from Pseudomonas maltophilia. Antirnicrob Agents Chemother. 1982;22:564-570.
13. Krcmery V, Antal M, Langsadl L, Knothe H. Transferable amikacin resistance in Pseudomonas maltophilia and Acinetobacter calcoaceticus. Infection. 1985;13:89-90. Letter.
14. Birnboim HC. A rapid alkaline extraction method for the isolation of plasmid DNA. Methods Enzymol. 1983;100:243-255.
15. Lennon E, DeCicco BJ. Plasmids ofPseudomonas cepacia strains of diverse origins. Appl Environ Microbiol. 1991;57:2345-2350.
16. Sambrook J, Fritsch EF, Maniatis T, eds. Molecular Cloning, A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory; 1989.
17. Feinberg AP, Vogelstein B. A technique for radiolabeling DNA restriction endonuclear fragments to high specific activity. Anal Biochem. 1983;132:6-13.
162





























