Embed Size (px)
Citation preview

Neurobiology of Disease 45 (2012) 692–700
Contents lists available at SciVerse ScienceDirect
Neurobiology of Disease
j ourna l homepage: www.e lsev ie r .com/ locate /ynbd i
A prolonged experimental febrile seizure results in motor map reorganizationin adulthood
Aylin Y. Reid a,b,⁎, Quentin J. Pittman a,c, G. Campbell Teskey a,c,d,e
a Hotchkiss Brain Institute, University of Calgary, Calgary, Alberta, Canada T2N 4N1b Department of Clinical Neurosciences, University of Calgary, Calgary, Alberta, Canada T2N 4N1c Department of Physiology and Pharmacology, University of Calgary, Calgary, Alberta, Canada T2N 4N1d Department of Cell Biology and Anatomy, University of Calgary, Calgary, Alberta, Canada T2N 4N1e Department of Psychology, University of Calgary, Calgary, Alberta, Canada T2N 4N1
⁎ Corresponding author at: Health Sciences Center, FaCalgary, 3330Hospital Drive NW, Calgary, Alberta, Canada T
E-mail address: [email protected] (A.Y. Reid).Available online on ScienceDirect (www.scienced
0969-9961/$ – see front matter © 2011 Elsevier Inc. Alldoi:10.1016/j.nbd.2011.10.013
a b s t r a c t
a r t i c l e i n f oArticle history:
Received 16 August 2011Revised 22 September 2011Accepted 16 October 2011Available online 25 October 2011Keywords:Intracortical microstimulationFAST ratGABAGlutamateNKCC1KCC2Cation-chloride co-transporterLipopolysaccharideKainic acid
Introduction: Clinical studies have suggested that children experiencing a febrile seizure (FS) before the age of1 year have persistent deficits, but it is unknown whether these seizures lead to permanent cortical reorga-nization and alterations in function. A FS on the background of increased genetic seizure susceptibility mayalso lead to negative long-term consequences. Alterations in neocortical motor map expression provide ameasure of neocortical reorganization and have been reported in both adults with frontal lobe epilepsyand following seizure induction in experimental models. The objectives of the present study were to deter-mine whether 1) an infantile FS leads to changes to motor map expression in adulthood; 2) long-term corti-cal reorganization is a function of the age at FS or genetic seizure susceptibility; and 3) different levels ofGABAA or glutamate receptor subunits or cation-chloride-co-transporters (CCCs) at the time of FS correlatewith alterations to motor map expression.Materials and methods: FSs were induced in postnatal day 10 (P10) or P14 Long–Evans (LE) rats or in P14seizure-prone FAST rats by the administration of the bacterial endotoxin lipopolysaccharide (LPS) and a sub-convulsant dose of kainic acid. Ten weeks later intracortical microstimulation was performed to generatemotor maps of forelimb movement representations. Sensorimotor neocortex samples were also dissected
from naïve P10 FAST and P10 and P14 LE pups for western blotting with antibodies against various GABAA,NMDA, and AMPA receptor subunits and for CCCs.Results: Adult FAST rats had larger motor maps with lower stimulation thresholds after a FS at P14, whileadult LE rats had significantly lower map stimulation thresholds but similar sized maps after a FS at P10 com-pared to controls. There were no differences in neocortical motor map size or stimulation thresholds in adultLE rats after a FS at P14. Both P10 LE and P14 FAST rats had significantly lower levels of the GABAA receptorα1 subunit, higher levels of the α2 subunit, and a higher NKCC1/KCC2 ratio in the sensorimotor cortex com-pared with the P14 LE rat. In addition, the P14 FAST rats had lower levels of the GluR2 and NR2A receptorsubunits in the sensorimotor cortex compared with the P14 LE rats.Conclusions: A single infantile FS can have long-term effects on neocortical reorganization in younger individ-uals and those with underlying seizure susceptibility. These changes may be related to an increased level ofexcitability in the neocortex of younger or genetically seizure-prone rats, as suggested by immaturity of theirGABAergic and CCC systems. Given the high incidence of FSs in children, it will be important to gain a betterunderstanding of how age and genetic seizure predisposition may contribute to the long-term sequelae ofthese events.© 2011 Elsevier Inc. All rights reserved.
Introduction
Febrile seizures (FSs) are the most common type of childhood sei-zures, affecting 2–5% of children between the ages of 6 months and
culty of Medicine, University of2N 4N1. Fax:+1 403 283 2700.
irect.com).
rights reserved.
5 years in the United States and Western Europe (Berg and Shinnar,1996). Nonetheless, they are still poorly understood and many longterm sequelae remain unknown.While alterations in hippocampal func-tion (Werboff and Havlena, 1963; Nealis et al., 1978; Kornelsen et al.,1996; Chang et al., 2003; Mesquita et al., 2006; Lemmens et al., 2009)and the subsequent development of spontaneous temporal lobe seizures(Dubé et al., 2006; Scantlebury et al., 2005) after experimental FSs havebeen reported, a compelling question is whether or not seizure inducedchanges may occur in other areas of the juvenile brain and lead to

693A.Y. Reid et al. / Neurobiology of Disease 45 (2012) 692–700
permanent alterations in neuronal function. There has been recent inter-est in evaluating the effect of repetitive neonatal seizures on neocorticalfunction (Isaeva et al., 2009, 2010), but studies investigating the effect ofprolonged FSs on the neocortex are lacking. While most cases of febrilestatus epilepticus involve seizures of focal origin, one third of cases aregeneralized seizures involving the neocortex (Shinnar et al., 2008).Given that even minor interventions in infancy can have profoundchanges on neuronal function that persist into adulthood (Bilbo et al.,2005; Boissé et al., 2005; Galic et al., 2008; Harré et al., 2008; Spenceret al., 2006) and that multiple neonatal seizures alter neocortical excit-ability (Isaeva et al., 2009, 2010) we hypothesized that a single early-life FS would induce neocortical changes in adult animals.
One such change is seizure-induced alterations in neocortical motormap expression, previously demonstrated in both humanswith epilepsy(Branco et al., 2003; Chlebus et al., 2004; Hamer et al., 2005; Labyt et al.,2007; Lado et al., 2002; Stoeckel et al., 2002; Uematsu et al., 1992;Urasaki et al., 1994), and with electrical and chemical seizure models inmature rats (Henderson et al., 2011; Teskey et al., 2002; van Rooyenet al., 2006; Younget al., 2009). Alterations inmovement representationsreflect reorganization of the neocortex that results from a change in thebalance between excitation and inhibition (see Teskey et al., 2008).
In evaluating the effect of FSs on neocortical function it is also impor-tant to determinewhether age at FS or an underlying genetic increase inseizure susceptibility may play a role. In humans there is evidence thatcomplex FSs at an earlier age, namely within the first year of life, havemore profound consequences than FSs occurring after the age of oneyear (Chang et al., 2001). Experimental FSs have been induced at differ-ent developmental stages, but there have been no direct comparisons ofthe effects of FSs at different ages. Evidence suggests experimental FSsmay have different long-term consequences depending on the geneticbackground (Zhao et al., 1985a, 1985b), which may also hold true inFS-induced changes in neocortical function. It is possible that thelong-termeffects found in younger or genetically seizure-prone animalsare related to maturity of various neurotransmitter and ion transportersystems at the time of the FS.
The objectives of the present study were 1) to determine whetheran infantile FS leads to changes in adult motor map expression; 2) todetermine whether the age when FS occurs or genetic seizure suscep-tibility relate to motor map expression in adulthood; and 3) to deter-mine whether different levels of expression of GABAA or glutamatereceptor subunits or cation-chloride co-transporters at the time ofFS correlate with plasticity of neocortical motor maps.
Materials and methods
To assess whether age at FS plays a role in the long-term conse-quences, we induced FSs at either P10 or P14 in Long–Evans rats. P10was chosen as this age is often used in other models of FSs that resultin significant alterations of neuronal function in adulthood and eventhe generation of spontaneous seizures (Bender et al., 2003; Brewsteret al., 2002, 2005; Chen et al., 1999; Dubé et al., 2006; Richichi et al.,2008). P14 was chosen as an age representative of a human infant at amore mature stage (Romijn et al., 1991; Talos et al., 2006a, 2006b)and when we have previously found that FSs lead to altered hippocam-pal excitability in adulthood (Heida et al., 2005). In addition, mindful ofthe fact that genetic alterations are thought to underlie at least sometypes of seizures, we also induced FSs at P14 in the seizure-proneFAST rat strain. FAST rats have been selectively bred to kindle morequickly (Racine et al., 1999) and demonstrate increased sensitivity tochemical convulsants (Racine et al., 2003), although there have notbeen published reports of spontaneous seizures.
Animals
Pregnant female LE rats (control strain) were obtained fromCharles River (Montreal, Que., Canada) and pregnant female FAST
rats were obtained from the breeding facility at the University of Cal-gary (FAST rat colony courtesy of Dr. Dan McIntyre, Carleton Univer-sity). Animals were maintained in standard, specified pathogen freeenvironmental conditions on a 12 h light/dark cycle at 22 °C withfood and water available ad libitum. Animals were monitored forthe day of parturition, which was taken as postnatal day (P)0. Onlymale pups were used for experimental procedures, and a minimumof four litters were used at a time in all experiments. All procedureswere approved by the local animal care committee and met theguidelines of the Canadian Council of Animal Care and EC Directive86/609/EEC for animal experiments.
Induction of febrile seizures
FSs were induced in P10 (LE) or P14 (LE and FAST) male pups aspreviously described (Heida et al., 2004). Briefly, pups were separatedfrom their dams and placed in groups of five to six in a temperature-controlled room maintained at 30 °C. After an hour in this environ-ment rats received either an i.p. injection of lipopolysaccharide(LPS) (Escherichia coli serotype 026:B6; 200 μg/kg) or pyrogen-freesaline. Two and a half hours later, during the increase in body temper-ature, rats that had previously received an injection of LPS received ani.p. injection of what is normally a subconvulsant dose of KA(1.75 mg/kg) when given alone; all other rats received another injec-tion of physiological saline. This combination of KA and LPS has previ-ously been found to elicit FS in approximately fifty percent of P14 rats(Heida et al., 2004). The rats were then videotaped for 3 h. The video-tapes were later analyzed by a trained, blind observer, for signs of be-havioral convulsions consisting of “wet-dog shakes”, loss of posturalcontrol, clonic movement of the extremities, and tonus, using a mod-ified Racine seizure scale. Based on the presence of those behaviorsrats were assigned to an experimental group: 1) saline controls(SC); 2) LPS/KA no seizure; or 3) LPS/KA+seizure. After rats returnedto normal behavior they were returned to their dams. At P21 ratswere weaned from their dam and maintained two or three animalsper cage until intracortical microstimulation (ICMS) was performed.
Intracortical microstimulation
Standard ICMS techniques (Kleim et al., 1998; Nudo et al., 1990)were used to generate detailed maps of the sensorimotor neocorticalforelimb regions ten to twelve weeks after induction of FS, when therats were 12 to 14 weeks old. The forelimb representation area waschosen as it is easily quantified because it is bordered by non-forelimb points, it is well-studied, and shows plasticity to seizures(see Teskey et al., 2008). LE and FAST rats were food deprived 12 hbefore ICMS surgery with free access to water. On the day of surgeryrats received an initial i.p. injection of ketamine (100 mg/kg) andxylazine (5 mg/kg). Supplemental injections of ketamine alone(25 mg/kg) or a cocktail of ketamine (17 mg/kg) and xylazine(2 mg/kg) were delivered i.p. as required throughout the surgicalprocedure to maintain a constant level of anesthesia as indicated bybreathing rate, vibrissae whisking, and withdrawal reflex in responseto a gentle pinch of the hind foot. Amounts of anesthetic wererecorded to allow later verification that experimental groups did notrequired different amounts.
A craniotomy approximately 7×5 mm in size was performed overthe left sensorimotor neocortex. The window extended approximate-ly 4 mm anterior to and 3 mm posterior to bregma, and from nearmidline to 5 mm lateral of midline. The cisterna magnumwas openedto reduce intracranial pressure. The dura over the exposed sensori-motor neocortex was carefully removed and body-temperature sili-cone fluid (Factor II Inc, Lakeside, AZ) was applied to cover thecortical surface. A 32× digital image of the exposed portion of theneocortex was acquired with a Stemi 2000-C stereomicroscope (CarlZeiss, Thornwood, NY) and digital camera (Cohu, San Diego, CA)

Table 1Antibodies. Primary and secondary antibodies used for western blotting.
Primary antibodies
Protein Antibody Dilution
Β-actin Mouse monoclonalMillipore, Temecula, CA
1:30,000
Rabbit polyclonalSigma-Aldrich, St Louis, MO
1:30,000
GluR2 Mouse monoclonalUC Davis/NINDS/NIMH NeuroMab Facility
1:500
NR2A Rabbit polyclonalAbcam, Cambridge, MA
1:300
GABAA α1 Mouse monoclonalUC Davis/NINDS/NIMH NeuroMab Facility
1:200
GABAA α2 Rabbit polyclonalAbcam, Cambridge, MA
1:200
KCC2 Mouse monoclonalUC Davis/NINDS/NIMH NeuroMab Facility
1:250
Secondary antibodies
Channel Antibody Dilution
680 nm Goat anti-mouseThermoscientific, Waltham, MA
1:10,000
Goat anti-rabbitThermoscientific, Waltham, MA
1:10,000
800 nm Goat anti-mouseThermoscientific, Waltham, MA
1:10,000
Goat anti-rabbitThermoscientific, Waltham, MA
1:10,000
694 A.Y. Reid et al. / Neurobiology of Disease 45 (2012) 692–700
and displayed on a computer monitor using the imaging softwareCanvas (version 9.0.1, ACD Systems Inc, Miami, FL). A grid composedof 500 μm squares was overlaid on the digital image. Penetrationpoints were chosen at the intersections of the grid lines and at a cen-tral point in the middle of each square, except when located over ablood vessel.
Microelectrodes were made from borosilicate glass capillaries(World Precision Instruments, Sarasota, FL) on a micropipette puller(Kopf, Tujunga, CA) and were filled with 3.5 M NaCl and beveled ata 30° angle to yield a 3 μm tip diameter with impedance values be-tween 1.0 and 1.5 MΩ. Neocortical penetrations by the microelec-trodes were guided by a microdrive (Narashige, Tokyo, Japan) to adepth of 1550 (+/−25)μm from the neocortical surface, correspond-ing to the cell-body region of neocortical layer V. Electrical stimula-tion was delivered with an isolated stimulator (A–M Systems,Carlsborg, WA) and consisted of 13 monophasic cathodal pulses,each 200 μs in duration, delivered at a frequency of 333 Hz, repeatedevery second. Rats were maintained in a prone position throughoutsurgery with the limb contralateral to the side of stimulation beingsupported below the elbow joint, with the forelimb elevated, to aidvisual detection of forelimb movements. The minimum threshold re-quired to elicit a movement was recorded for each penetration site,and a color-coded dot was placed on the digital image correspondingto the type of movement seen. The minimum threshold was deter-mined by starting with a current intensity of 0 μA, rapidly increasingthe intensity until a movement was elicited, then decreasing the in-tensity again until no movement was seen. No more than 10 trainsof pulses were delivered to a single site to determine movements.Penetration sites that did not elicit any movement up to the maxi-mum stimulation intensity of 60 μA were defined as nonresponsive.The border of the forelimb motor map was determined first, charac-terized by non-forelimbmovements or non-responsive sites. The cen-tral forelimb motor map points were then determined in order toreduce the likelihood of ICMS affecting the border points of the map(Nudo et al., 1990). The level of anesthesia was assessed by revisitingpositive-response sites later in the mapping procedure to check forchanges in movement thresholds. After the mapping session wascomplete the rats were euthanized with an intracardiac injection of0.4 ml of 240 mg/ml sodium pentobarbital.
Canvas imaging software (ACD Systems) was used to calculate thesurface area of both caudal and rostral forelimb representation areas.As some animals did not display a clear border between these twoareas both were included in all analyses for all animals.
Western blotting
Experimentally naïve P10 and P14 LE rats and P14 FAST rats wereanesthetised with pentobarbital (60 mg/kg) and perfused transcar-dially with cold phosphate-buffered saline. In order to look for differ-ences present at the time of FS induction which may account forchanges in motor map expression in adulthood, the sensorimotor cor-tex were quickly dissected and frozen at −80 °C for further proces-sing. Proteins were chosen for analysis based on previous reports oftheir roles in epileptogenesis and plasticity: glutamatergic receptorsubunits GluR2 (Richichi et al., 2008) and NR2B (Tang et al., 1999),GABAA receptor subunits α1 (Ni et al., 2004) and α5 (Houser andEsclapez, 2003), and the cation-chloride co-transporters NKCC1(Kang et al., 2002; Okabe et al., 2002) and KCC2 (Jin et al., 2005;Pathak et al., 2007). Tissue samples from each rat were homogenizedand total cellular protein was extracted using RIPA lysis buffer withprotease inhibitor cocktail (Roche, Mississauga, Ontario). After dis-ruption with a mechanical tissue homogenizer the samples were cen-trifuged at 10,000×g for 10 min at 4 °C and the supernatant wascollected. Protein levels were quantified by the Bradford ProteinAssay (Bio-Rad, Mississauga, Ontario), and concentrations were ad-justed with RIPA buffer. Samples were reduced with 6X Laemlli
buffer, denatured at 60 °C for 5 min, and used immediately. Proteinswere separated on either 4–12% or 4–20% Tris-glycine buffered poly-acrylamide gels (PAGEr Gold Precast Gels, Lonza, Rockland, ME),depending on the size of the protein of interest, and then transferredonto polyvinylidene fluoride membranes using a semi-dry transfermethod. Membranes were blocked by incubation in Seablock reagent(Calbiochem, EMD Biosciences Inc., La Jolla, CA) for 2 h, and then in-cubated overnight at 4 °C with one primary antibody against the pro-tein of interest and a second primary antibody against β-actin as aloading control (Table 1). Actin, GluR2, and GABAA α1 Abs wereshown to be specific by the manufacturer as demonstrated by the ab-sence of staining in knock-out animals; this information was not pro-vided by the manufacturers for the other Abs, so specificity of stainingwas evaluated by presence of a band at the expected molecularweight of the antigen and by reference to previous publicationsreporting use and specificity of the Ab. The next day membraneswere washed and incubated for 2 h in infrared fluorescent dye-conjugated secondary Abs against the hosts of the two primary Abs(Table 1). Protein bands were visualized using a Li-Cor Odyssey dualwavelength far infrared scanner (Li-Cor Biosciences, Lincoln, NE)with imaging in the 800 nm channel for the protein of interest andin the 700 nm channel for β-actin. The area under the intensity profilecurve of a given band was quantified (Quantity One, Bio-Rad) and theratio of optical density values for the protein of interest/β-actin wascalculated. The mean optical density ratios for any given protein inthe P14 LE pups were normalized to a value of 1.0, and the ratiosfor the P10 LE and P14 FAST pups were then normalized to the P14LE group.
Real-time RT-PCR
Testing of three commercially available Abs for NKCC1 all showednon-selective staining, therefore real-time RT-PCR was performed onsensorimotor cortex samples to determine mRNA levels for NKCC1and KCC2. Tissue was prepared for mRNA extraction using the RNeasyPlus kit (Qiagen, Mississauga, ON) according to the manufacturer's in-structions. Extracted RNA was used to synthesize cDNA according tomanufacturer's instructions for the Quantitect Reverse Transcription

695A.Y. Reid et al. / Neurobiology of Disease 45 (2012) 692–700
kit (Qiagen, Mississauga, ON). Gene-specific probes designed forNKCC1 and KCC2 were synthesized at the University of Calgary(Table 2), and have been previously used in our laboratory (Reidet al., 2009). RT-PCR was performed on a Mastercycler Realplex 4(Eppendorf, USA). Amplification of the genes of interest was detectedusing the fluorescent dye SYBR-green (SYBR-green PCR kit: Qiagen,Mississauga, ON). To perform semi-quantitative analysis, a thresholdcycle (CT) value was calculated for each gene of interest by determin-ing the point at which fluorescence exceeded a threshold limit. Datawas normalized against mRNA levels of an endogenous reference gene,GAPDH. mRNA levels in the P14 FAST pups were expressed relative toP14 LE pups using the “delta–delta CT” method (Livak and Schmittgen,2001). As mRNA levels of GAPDH differed significantly in the P10 LEpups, the mRNA levels of the genes of interest in the P10 pups couldnot be normalized against the P14 animals.
Statistical analysis
The Kruskal–Wallis test was conducted to assess differences inICMS anesthetic levels, ICMS stimulation thresholds, map areas, andwestern blot protein levels within each strain or age. Dunn's MultipleComparison post-hoc test was conducted to assess differences be-tween the saline, LPS/KA no seizure, and LPS/KA+seizure groupsfor ICMS parameters, and between the P10 LE, P14 LE, and P14 FASTgroups for western blotting. The Mann–Whitney U test was con-ducted to assess differences in levels of mRNA expression for NKCC1and KCC2 between the P14 LE and FAST pups. Statistical significancewas set at pb0.05.
Results
Induction of febrile seizures
FSs were induced in LE pups at P10 or P14, and at P14 in FASTpups. The distribution of pups per litter to each experimental groupis shown in Table 3. There were no significant differences in the pro-portion of animals receiving LPS/KA that had a behavioral seizure orin the duration of FSs between groups (p>0.05). All behavioral sei-zures lasted at least 30 min, meeting the criteria for complex FS.
P10 LE pupsSix LE pups received saline injections, none of which demonstrat-
ed behavioral FS. Fourteen animals received injections of LPS and KA,of which eight developed behavioral FS. The seizures started withwet-dog shakes, and then some animals developed periods of immo-bility with rats laying on their backs or sides. All animals in the FSgroup progressed to fully generalized seizures. Seizures began5–15 min after KA injection, lasting a range of 35 to 65 min.
P14 LE pupsSix LE pups received saline injections, none of which demonstrated
behavioral FS. Thirteen animals received injections of LPS and KA, ofwhich six developed behavioral FS. The seizures were similar to thoseseen in the P10 LE pups in both character and duration (35 to50 min). All animals in the FS group progressed to at least forelimb
Table 2RT-PCR probes. Probes and their forward (3′ to 5′) and reverse sequences (5′ to 3′)used in reverse-transcription polymerase chain reaction.
Primer Sequence
GAPDH-for GCA TGG CCT TCC GTG TTC CTA CCCGAPDH-rev GGC CGC CTG CTT CAC CAC CTT CTNKCC1-for TGG TGT CAG GAT TTG CAC CCNKCC1-rev GCC CAC CAG TTA ATG ACA AAC CKCC2-for CGG TGA TGG CAA CCC CAA AGKCC2-rev GGC TCC TCG AAT GGG CAC CTT
tonus/clonus, with twenty-five percent of animals progressing to fullygeneralized seizures.
P14 FAST pupsSeven FAST pups received saline injections, none of which demon-
strated behavioral FS. Fourteen animals received injections of LPS andKA, of which eight developed behavioral FS. The seizures were similarto those seen in LE pups, although periods of tonus were generallycontinuous once they began, and the rats occasionally regainedsome mobility, had further wet-dog shakes, and then returned to aperiod of clonus. All animals in the FS group progressed to fully gen-eralized seizures. Seizures began 5–15 min after injection of KA, andlasted a range of 40 to 90 min.
Intra-cortical microstimulation and movement representation analysis
P10 LE ratsThere were no significant differences in the mean amounts of keta-
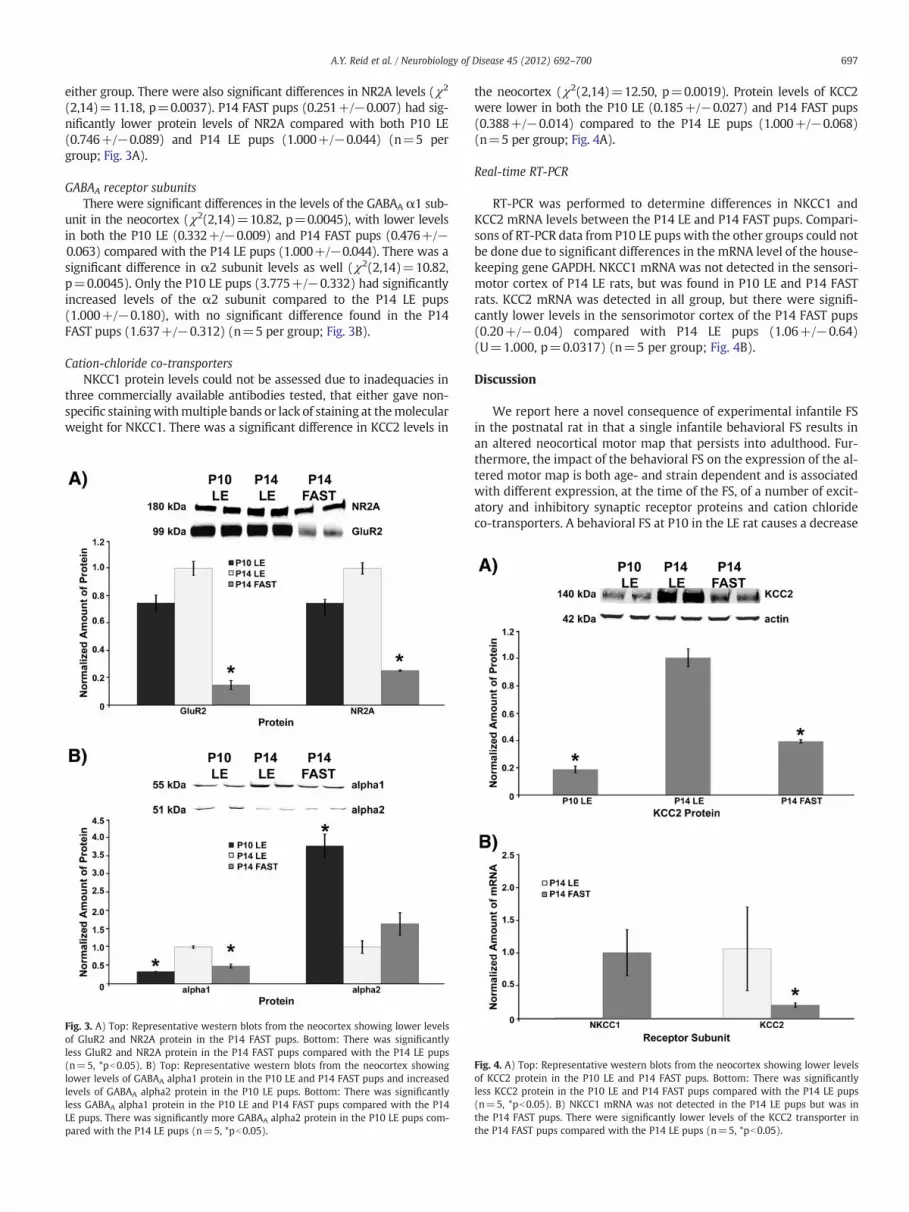
mine or xylazine used as a function of bodyweight and duration of sur-gery between the saline (n=6), LPS/KA no seizure (n=6) or LPS/KA+seizure groups (n=8). The mean amounts of ketamine (+/−SEM)used during ICMS in mg/kg/min were: 0.916+/−0.078 (saline),1.050+/−0.093 (LPS/KA no seizure), and 0.991+/−0.025 (LPS/KA+seizure). The mean amounts of xylazine (+/−SEM) in mg/kg/minwere: 0.0410+/−0.0033 (saline), 0.0472+/−0.0031 (LPS/KA no sei-zure), and 0.0451+/−0.0008 (LPS/KA+seizure). There was a signifi-cant difference in the minimum thresholds required to elicit forelimbresponses between groups (χ2(2,20)=8.982, p=0.0112). The LPS/KA+seizure group had a significantly lower meanminimum threshold(22.5+/−1.0) than either the saline (27.5+/−0.9) and LPS/KA no sei-zure groups (27.3+/−1.2) (Fig. 1A). There were no significant differ-ences in the mean size of neocortical forelimb representation areasbetween any of the P10 LE groups (saline=5.19+/−0.59; LPS/KA noseizure=5.12+/−0.64; LPS/KA+seizure=5.41+/−0.36) (Fig. 1B).A representative sample of a neocortical forelimb motor map from aP10 LE LPS/KA+seizure animal is shown in Fig. 2A.
P14 LE ratsThere were no significant differences in the mean amount of anes-
thetics used during the ICMS procedure between the saline (n=6), LPS/KA no seizure (n=7) or LPS/KA+seizure (n=6) groups. The meanamounts of ketamine (+/−SEM) in mg/kg/min were: 0.898+/−0.061(saline), 0.934+/−0.086 (LPS/KA no seizure), and 1.004+/−0.066(LPS/KA+seizure). The mean amounts of xylazine used were:0.0417+/−0.0016 (saline), 0.0460+/−0.0017 (LPS/KA no sei-zure), and 0.0445+/−0.0015 (LPS/KA+seizure). There were no signif-icant differences in the minimum thresholds (saline=27.0+/−1.1;LPS/KA no seizure=27.0+/−0.9; LPS/KA+seizure=28.4+/−1.0)(Fig. 1A) or the size of the neocortical forelimb area (saline=4.78+/−0.98; LPS/KA no seizure=5.73+/−0.89; LPS/KA+seizure=3.90+/−0.66) (Fig. 1B) between any of the P14 LE groups. A representative sam-ple of a neocortical forelimb motor map from a P14 LE LPS/KA+seizureanimal is shown in Fig. 2B.
P14 FAST ratsThere were no significant differences in the mean amount of keta-
mine or xylazine administered during ICMS as a function of bodyweightand duration of surgery between the saline (n=7), LPS/KA no seizure(n=6) or LPS/KA+seizure (n=8) groups. The mean amountsof ketamine (+/−SEM) in mg/kg/min used were: 0.983+/−0.052 (sa-line), 1.015+/−0.077 (LPS/KA no seizure), and 1.073+/−0.071 (LPS/KA+seizure). The mean amounts of xylazine (+/−SEM) used were:0.04050+/−0.0019 (saline), 0.0429+/−0.0029 (LPS/KA no seizure),and 0.0455+/−0.0026 (LPS/KA+seizure). There was a significant dif-ference in the minimum threshold required to elicit forelimb move-ments (χ2(2,20)=11.94, p=0.0026). The LPS/KA+seizure group

Table 3Antibodies. Distribution of litters to experimental groups.
P10 Long Evans pups P14 Long Evans pups P14 Fast pups
Group Litters Number of pups LITTERS Number of pups Litters Number of pups
Saline A 2 E 1 I 1B 1 F 2 J 2C 1 G 2 K 2D 2 H 1 L 1
M 1Total=4 litters Total=6 pups Total=4 litters Total=6 pups Total=5 litters Total=7 pups
LPS/KA no seizure A 1 E 2 I 1B 2 F 1 J 1C 1 G 2 K 1D 2 H 2 L 1
M 2Total=4 litters Total=6 pups Total=4 litters Total=7 pups Total=5 litters Total=6 pups
LPS/KA+seizure A 2 E 2 I 2B 2 F 2 J 2C 3 G 1 K 1D 1 H 1 L 2
M 1Total=4 litters Total=8 pups Total=4 litters Total=6 pups Total=5 litters Total=8 pups
696 A.Y. Reid et al. / Neurobiology of Disease 45 (2012) 692–700
(21.2+/−0.9) had a significantly lowermeanminimum threshold thaneither the saline (27.1+/−1.1) or LPS/KA no seizure groups (26.9+/−1.0) (Fig. 1A). Mean forelimb representation areas also differedsignificantly between groups (χ2(2,20)=12.18, p=0.0023). Themean neocortical forelimb motor area in the LPS/KA+seizure group(8.84+/−1.01) was significantly larger than the mean area for the sa-line (4.79+/−0.64) or LPS/KA no seizure groups (4.91+/−0.58)
Fig. 1. A) P10 LE rats in the LPS/KA+seizure group had significantly lower thresholdsthan the P10 LE saline and LPS/KA no seizure rats. P14 FAST rats in the LPS/KA+seizuregroup had significantly lower thresholds than the P14 FAST saline and LPS/KA no seizurerats. B) Only P14 FAST rats in the LPS/KA+seizure group had significantly larger mapsthan the saline and LPS/KA no seizure groups (n=7, *pb0.05).
(Fig. 1B). The total neocortical forelimb area in FS rats was 185% ofthat of the saline group and 180% of that of the LPS/KA no seizuregroup. Representative samples of neocortical forelimb motor mapsfrom a P14 FAST saline and a P14 FAST LPS/KA+seizure animal areshown in Fig. 2C and D, respectively.
Western blotting
Glutamate receptor subunitsWestern blots of sensorimotor neocortical samples showed signifi-
cant differences in GluR2 levels (χ2(2,14)=12.02, p=0.0025), withlower protein levels of GluR2 in P14 FAST pups (0.146+/−0.036) com-pared to P14 LE pups (1.000+/−0.053). The protein level of GluR2 inP10 LE pups (0.748+/−0.059) was not significantly different than
Fig. 2. Representation motor maps from a: A) P10 LE LPS/KA+seizure; B) P14 LE LPS/KA+seizure; C) P14 FAST saline control; and D) P14 FAST LPS/KA+seizure animal.
697A.Y. Reid et al. / Neurobiology of Disease 45 (2012) 692–700
either group. There were also significant differences in NR2A levels (χ2
(2,14)=11.18, p=0.0037). P14 FAST pups (0.251+/−0.007) had sig-nificantly lower protein levels of NR2A compared with both P10 LE(0.746+/−0.089) and P14 LE pups (1.000+/−0.044) (n=5 pergroup; Fig. 3A).
GABAA receptor subunitsThere were significant differences in the levels of the GABAA α1 sub-
unit in the neocortex (χ2(2,14)=10.82, p=0.0045), with lower levelsin both the P10 LE (0.332+/−0.009) and P14 FAST pups (0.476+/−0.063) compared with the P14 LE pups (1.000+/−0.044). There was asignificant difference in α2 subunit levels as well (χ2(2,14)=10.82,p=0.0045). Only the P10 LE pups (3.775+/−0.332) had significantlyincreased levels of the α2 subunit compared to the P14 LE pups(1.000+/−0.180), with no significant difference found in the P14FAST pups (1.637+/−0.312) (n=5 per group; Fig. 3B).
Cation-chloride co-transportersNKCC1 protein levels could not be assessed due to inadequacies in
three commercially available antibodies tested, that either gave non-specific stainingwithmultiple bands or lack of staining at themolecularweight for NKCC1. There was a significant difference in KCC2 levels in
Fig. 3. A) Top: Representative western blots from the neocortex showing lower levelsof GluR2 and NR2A protein in the P14 FAST pups. Bottom: There was significantlyless GluR2 and NR2A protein in the P14 FAST pups compared with the P14 LE pups(n=5, *pb0.05). B) Top: Representative western blots from the neocortex showinglower levels of GABAA alpha1 protein in the P10 LE and P14 FAST pups and increasedlevels of GABAA alpha2 protein in the P10 LE pups. Bottom: There was significantlyless GABAA alpha1 protein in the P10 LE and P14 FAST pups compared with the P14LE pups. There was significantly more GABAA alpha2 protein in the P10 LE pups com-pared with the P14 LE pups (n=5, *pb0.05).
the neocortex (χ2(2,14)=12.50, p=0.0019). Protein levels of KCC2were lower in both the P10 LE (0.185+/−0.027) and P14 FAST pups(0.388+/−0.014) compared to the P14 LE pups (1.000+/−0.068)(n=5 per group; Fig. 4A).
Real-time RT-PCR
RT-PCR was performed to determine differences in NKCC1 andKCC2 mRNA levels between the P14 LE and P14 FAST pups. Compari-sons of RT-PCR data from P10 LE pups with the other groups could notbe done due to significant differences in the mRNA level of the house-keeping gene GAPDH. NKCC1 mRNA was not detected in the sensori-motor cortex of P14 LE rats, but was found in P10 LE and P14 FASTrats. KCC2 mRNA was detected in all group, but there were signifi-cantly lower levels in the sensorimotor cortex of the P14 FAST pups(0.20+/−0.04) compared with P14 LE pups (1.06+/−0.64)(U=1.000, p=0.0317) (n=5 per group; Fig. 4B).
Discussion
We report here a novel consequence of experimental infantile FSin the postnatal rat in that a single infantile behavioral FS results inan altered neocortical motor map that persists into adulthood. Fur-thermore, the impact of the behavioral FS on the expression of the al-tered motor map is both age- and strain dependent and is associatedwith different expression, at the time of the FS, of a number of excit-atory and inhibitory synaptic receptor proteins and cation chlorideco-transporters. A behavioral FS at P10 in the LE rat causes a decrease
Fig. 4. A) Top: Representative western blots from the neocortex showing lower levelsof KCC2 protein in the P10 LE and P14 FAST pups. Bottom: There was significantlyless KCC2 protein in the P10 LE and P14 FAST pups compared with the P14 LE pups(n=5, *pb0.05). B) NKCC1 mRNA was not detected in the P14 LE pups but was inthe P14 FAST pups. There were significantly lower levels of the KCC2 transporter inthe P14 FAST pups compared with the P14 LE pups (n=5, *pb0.05).

698 A.Y. Reid et al. / Neurobiology of Disease 45 (2012) 692–700
in the minimum stimulation intensity in the neocortex required toelicit forelimb movements in adulthood, while a single behavioralFS at P14 in the genetically seizure-prone FAST rat causes both a de-crease in minimum stimulation threshold as well as an increase inthe size of the neocortical forelimb representation area. However, asingle behavioral FS at P14 in the LE rat did not lead to changes in ei-ther minimum thresholds or map size in adulthood. We propose thatdifferences in abundance of various neurotransmitter subreceptorsand in CCC balance in P10 LE and P14 FAST rats compared with P14LE rats, reflect a brain that responds to an early-life behavioral FS inan altered manner, leading to more persistent alterations in neocorti-cal neuronal function in adulthood.
The larger motor maps observed in adulthood after a behavioral FSin the P14 FAST pups indicate an overall increase in excitability inneocortical layer V. Synapse number and dendritic morphology inlayer V correlate with synaptic efficacy in that larger cortical motormaps (Kleim et al., 2002; Teskey et al., 2002) have increased numbersof excitatory synapses (Henry et al., 2008). In contrast, smaller motormaps are associated with fewer excitatory perforated synapses andmore inhibitory synapses (Teskey et al., 2007). Dendritic hypertrophyin layer V has also been found in association with larger motor maps(Monfils et al., 2004), while smaller maps are associated with a de-crease in dendritic length in layer V (Monfils and Teskey, 2004).Seizure-induced enlargement of the forelimb motor map correlateswith forelimb behavioral deficits (Flynn et al., 2010; Henry et al.,2008).
The lower ICMS stimulation thresholds seen in adulthood after abehavioral FS in P10 LE and P14 FAST pups are also likely dependanton overall increased excitability in neocortical layer V. The normal in-crease in ICMS-generated motor map size that occurs during post-natal development is inversely related to minimum stimulationthresholds (Young et al., 2011). There is evidence that the GABAergicsystem is involved in determining threshold levels, as enhancedGABAergic inhibition induced by diazepam during the ICMS proce-dure leads to smaller motor maps and higher minimum thresholds(Young et al., 2011), while injection of the GABA antagonist bicucul-line directly into layer V leads to responsive map points at an agewhen neocortical motor maps are not yet revealed with standardICMS procedures (Young et al., 2011). Evidence also exists for a roleof the glutamatergic system in determining threshold levels, as ratsdeeply anesthetized with the NMDA antagonist ketamine have in-creased ICMS stimulation thresholds (Tandon et al., 2008; Younget al., 2011). It is likely that the lowered thresholds related to an over-all increase in excitability, but whether this balance was altered aftera FS in the P10 LE and P14 FAST secondary to changes in inhibition,excitation, or both is unknown.
Given the role of excitation and inhibition in sculpting adult motormap size and sensitivity, we postulated that differences in maturity ofthe neocortex may be responsible for the differences we observed inthe age and strain dependent effects of behavioral FS on adult motormaps. Our findings of differences in the expression of various GABAA
and glutamate receptor subunits and CCCs in the P10 LE and P14 FASTneocortex compared to the P14 LE neocortex are consistent with thispostulate. In particular, we found lower levels of GABAA receptor α1subunits, higher levels of GABAA receptor α2 subunits, higher levels ofNKCC1, and lower levels of KCC2 in both the P10 LE and P14 FASTpups compared with the P14 LE pups. Lower ICMS stimulation thresh-olds were found in both of these groups after a behavioral FS. It is diffi-cult to interpret the impact of different GABAA receptor subunit levels,which in themselves are associated with different gating and currentcarrying properties (Lüddens and Wisden, 1991; Vicini, 1991; Wisdenand Seeburg, 1992). However, the elevated expression of NKCC1 rela-tive to that of KCC2 would favor GABA having a more excitatory actionduring the FS in the P14 FAST rat. The difference we see with respect tothe P10 and P14 LE rats would have a similar impact and is also consis-tent with the previously reported switch-over of these transporters
between P10 and P14 (Miles, 1999; Rivera et al., 1999) It is also likelythat a FS elicited at these times when NKCC1 activity is elevatedwould also favor NMDA receptor activation by excitatory GABAergic ac-tion (Wang and Kriegstein, 2008). Seizure activity during this time pe-riod may have set up for altered neocortical GABAergic transmissionin adulthood, similar to the long-term suppression of neocorticalGABAergic activity after recurrent flurothyl-induced neonatal seizures(Isaeva et al., 2009, 2010), and in chronically injured, epileptogenic sen-sorimotor neocortex (Li and Prince, 2002).
In addition to this likely scenario of increased excitatory tone inthe more immature (either genetically or age-wise) neocortex as apredisposition to long term effects of a behavioral FS, P14 FAST pupsalso had lower levels of the AMPA receptor GluR2 subunit and theNMDA receptor NR2A subunit in the neocortex, and were the only an-imals to show larger motor maps in adulthood after a FS. Neuronalcalcium entry occurs after activation of GluR2-lacking AMPA recep-tors, which can lead to endocytosis of GABAA receptors (Khalilov etal., 2003), internalization of potassium channels (Bernard et al.,2004), and AMPA receptor mediated potentiation (Rakhade et al.,2008). Increased neuronal calcium entry after activation of GluR2-lacking AMPA receptors during a FS in the P14 FAST pup may haveled to some of these previously documented changes, causing in-creased excitability and leading to larger ICMS-generated motormaps. In addition, while NR2B levels were comparable in all of the ex-perimentally naïve pups, the lower expression of NR2A would lead toa lower NR2A/NR2B ratio in the P14 FAST neocortex. The NR2B subu-nit allows for enhanced activation of the NMDA channel, which in-creases its ability to strengthen synapses (Tang et al., 1999).Therefore, having a lower NR2A/NR2B ratio of subunits at the timeof behavioral FS may have also allowed for increased neuronal calci-um entry in the P14 FAST pups, with similar consequences as activa-tion of GluR2-lacking AMPA receptors.
We have found that a behavioral FS at a younger age (P10) leads topersistent changes in neocortical excitability in adulthood, namelylower ICMS stimulation thresholds that are not seen with a FS at aslightly older age (P14). While it is difficult to correlate the age of arat with the same developmental period in humans, a FS at P10 likelycorresponds to the first year of human life, while a FS at P14 mightcorrelate with the beginning of the toddler years (Romijn et al.,1991; Talos et al., 2006a, 2006b). In regard to the cerebral cortexstudies we have made comparisons of a full-term human baby to aPND 15 rat in terms of synapse formation, a PND 7–9 rat for the devel-opment of glutamic acid decarboxylase activity, a PND 20 rat for thedevelopment of choline acetyltransferase activity, and a PND 12–13rat for the developmental pattern of electrocortical activity (seeRomijn et al., 1991). There is evidence for a role of age in the conse-quences of FSs in humans as well. While many studies in school-agechildren with a previous FS have demonstrated no deficits (Changet al., 2000; Ellenberg and Nelson, 1978; Verity et al., 1998), or evenimprovements (Chang et al., 2001) in memory and scholastic perfor-mance, age does appear to be a factor. If the FS occurred in the firstyear of life then the children had deficits in learning, consolidation,and delayed recognition (Chang et al., 2001) and required specialschooling more often than those with later FSs (Verity et al., 1998).Also, children with a FS before the age of one are more likely tohave recurrent FSs (van Stuijvenberg et al., 1998), with a 2.5-fold in-crease in the risk of recurrence versus children with a FS after the ageof three years (Verity et al., 1985). The children having a FS at thisyounger age may be more susceptible to them in the first place, orperhaps a FS within that time period has different consequencesand leads to a level of hyperexcitability that is not seen with a FS oc-curring after the age of one year. The ongoing prospective FEBSTATstudy has found that children presenting in febrile SE are more likelyto be of a younger age (Shinnar et al., 2008). As further results fromthis study become available it will be important to see what roleage may play in the long-term consequences of these seizures.

699A.Y. Reid et al. / Neurobiology of Disease 45 (2012) 692–700
We have also found that a behavioral FS that occurs on the back-ground of increased seizure susceptibility (FAST pup) leads to moreprofound consequences in neocortical function, with both loweredICMS stimulation thresholds and enlarged motor maps. Previousstudies have also shown that when rats genetically prone to audio-genic seizures are exposed to FSs they demonstrate altered seizuresusceptibility in adulthood (Zhao et al., 1985a) and will kindle withrepetitive FSs while rats that are not genetically seizure-prone donot (Zhao et al., 1985b). As clinical studies favor complex heredityof FSs (Rich et al., 1987; Tsuboi, 1976), it is difficult to study specificgenetic factors. However, a positive family history of FSs confers agreater risk of focal or recurrent FSs (Bethune et al., 1993; van Eschet al., 1994; Pavlidou et al., 2008) and is a risk factor for developinggeneralized epilepsy after a FS regardless of the clinical presentationof the FS (Birca et al., 2005).
This study relied on behavioral outcomes rather than electroen-cephalogram recordings to determine whether rats had a seizure inresponse to LPS/KA. It is possible that animals in the LPS/KA no sei-zure group had subclinical seizures that were not detected. However,we have previously recorded electroencephalograms from the hippo-campus of a subset of pups receiving LPS/KA and did not find anyelectrical seizures in the pups that did not demonstrate behavioralseizures (Reid et al., 2009). As well, previous experiments in our lab-oratory using the LPS/KA model have demonstrated significant differ-ences between LPS/KA no seizure and LPS/KA+seizure groups whenlooking at cytokine levels (Heida and Pittman, 2005) or electrical kin-dling parameters (Heida et al., 2005), suggesting it is the behavioralseizure which is causing the effect. However it is possible that less se-vere, subclinical seizure activity thatwe did not detect causes long-termalterations in the brain as well. Likewise, since we were observing be-havioral seizureswe cannot state definitively that the severity of the be-havioral FSs, where they existed, was identical among animals in termsof electroencephalogramactivity, so there is the possibility that changesseen among groups in adulthood reflect difference in FS severity aspups.
The demonstration of sensorimotor neocortical changes in anadult after a single infantile behavioral FS is a novel finding. Recentstudies have demonstrated suppression of neocortical GABAergic ac-tivity (Isaeva et al., 2009) and long-term increases in neocortical ex-citability (Isaeva et al., 2010) after flurothyl-induced neonatalseizures, but these were recurrent seizures. One study has shown al-terations in the development of spatio-temporal receptive field prop-erties of neurons in the primary visual cortex after a single FS, but thiswas in animals that also had a focal cortical dysplasia (Prevost et al.,2010). We have demonstrated altered neocortical neuronal functionafter a single early-life seizure, which appears to depend on a youngage of seizure onset or an underlying seizure predisposition, corre-sponding to an increased baseline level of excitation in the brain atthe time of the seizure. Given previous findings of altered cognitionand global brain development in children with a FS within the firstyear of life, and recently found changes in the visual cortex, it is likelythese changes are not restricted to the motor cortex but representmore widespread modifications. With the high incidence of FSs inchildren, it will be important to gain a better understanding of howage and genetic seizure predisposition may contribute to the long-term sequelae of these events.
Acknowledgments
This work was supported by Canadian Institutes of Health Re-search and Natural Sciences and Engineering Research Council grantsto G.C.T. and Q.J.P., and by personnel awards from the Hotchkiss BrainInstitute and Killam Trust (A.Y.R.), the Savoy Foundation (K.R.) andthe Alberta Heritage Foundation for Medical Research (A.Y.R., K.R.,and Q.J.P.). Thanks to Dr. Mio Tsutsui, Lorenzo Bauce, Bob Winkfein,
and Bonita Ma for technical support and to Dr. Wee Yong for theuse of the Li-Cor Odyssey scanner.
References
Bender, R.A., Soleymani, S.V., Brewster, A.L., Nguyen, S.T., Beck, H., Mathern, G.W.,Baram, T.Z., 2003. Enhanced expression of a specific hyperpolarization-activatedcyclic nucleotide-gated cation channel (HCN) in surviving dentate gyrus granulecells of human and experimental epileptic hippocampus. J. Neurosci. 23,6826–6836.
Berg, A.T., Shinnar, S., 1996. Unprovoked seizures in children with febrile seizures:short-term outcome. Neurology 47, 562–568.
Bernard, C., Anderson, A., Becker, A., Poolos, N.P., Beck, H., Johnston, D., 2004. Acquireddendritic channelopathy in temporal lobe epilepsy. Science 305, 532–535.
Bethune, P., Gordon, K., Dooley, J., Camfield, C., Camfield, P., 1993. Which child willhave a febrile seizure? Am. J. Dis. Child 147, 35–39.
Bilbo, S.D., Levkoff, L.H., Mahoney, J.H.,Watkins, L.R., Rudy, J.W., Maier, S.F., 2005. Neonatalinfection inducesmemory impairments following an immune challenge in adulthood.Behav. Neurosci. 119, 293–301.
Birca, A., Guy, N., Fortier, I., Cossette, P., Lortie, A., Carmant, L., 2005. Genetic influenceon the clinical characteristics and outcome of febrile seizures — a retrospectivestudy. Eur. J. Paediatr. Neurol. 9, 339–345.
Boissé, L., Spencer, S.J., Mouihate, A., Vergnolle, N., Pittman, Q.J., 2005. Neonatal immunechallenge alters nociception in the adult rat. Pain 119, 133–141.
Branco, D.M., Coelho, T.M., Branco, B.M., Schmidt, L., Calcagnotto,M.E., Portuguez,M., Neto,E.P., et al., 2003. Functional variability of the human cortical motor map: electricalstimulation findings in perirolandic epilepsy surgery. J. Clin. Neurophysiol. 20, 17–25.
Brewster, A., Bender, R.A., Chen, Y., Dube, C., Eghbal-Ahmadi,M., Baram, T.Z., 2002. Develop-mental febrile seizures modulate hippocampal gene expression of hyperpolarization-activated channels in an isoform- and cell-specific manner. J. Neurosci. 22, 4591–4599.
Brewster, A.L., Bernard, J.A., Gall, C.M., Baram, T.Z., 2005. Formation of heteromerichyperpolarization-activated cyclic nucleotide-gated (HCN) channels in thehippocampusis regulated by developmental seizures. Neurobiol. Dis. 19, 200–207.
Chang, Y.C., Guo, N.W., Huang, C.C., Wang, S.T., Tsai, J.J., 2000. Neurocognitive attentionand behavior outcome of school-age children with a history of febrile convulsions:a population study. Epilepsia 41, 412–420.
Chang, Y.C., Guo, N.W., Wang, S.T., Huang, C.C., Tsai, J.J., 2001. Working memory ofschool-aged children with a history of febrile convulsions: a population study.Neurology 57, 37–42.
Chang, Y.C., Huang, A.M., Kuo, Y.M., Wang, S.T., Chang, Y.Y., Huang, C.C., 2003. Febrileseizures impair memory and cAMP response-element binding protein. Ann. Neu-rol. 54, 706–718.
Chen, K., Baram, T.Z., Soltesz, I., 1999. Febrile seizures in the developing brain result inpersistent modification of neuronal excitability in limbic circuits. Nat. Med. 5,888–894.
Chlebus, P., Brázdil, M., Hlustík, P., Mikl, M., Pazourková, M., Krupa, P., 2004. Handednessshift as a consequence ofmotor cortex reorganization after early functional impairmentin left temporal lobe epilepsy— an fMRI case report. Neurocase 10, 326–329.
Dubé, C., Richichi, C., Bender, R.A., Chung, G., Litt, B., Baram, T.Z., 2006. Temporal lobeepilepsy after experimental prolonged febrile seizures: prospective analysis.Brain 129, 911–922.
Ellenberg, J.H., Nelson, K.B., 1978. Febrile seizures and later intellectual performance.Arch. Neurol. 35, 17–21.
Flynn, C., Young, N.A., Teskey, G.C., 2010. Seizures, but not lowered seizure thresholds,results in larger neocortical motor maps and concomitant disruptions in skilledmotor behaviour. Behav. Brain Res. 214, 60–65.
Galic, M.A., Riazi, K., Heida, J.G., Mouihate, A., Fournier, N.M., Spencer, S.J., Kalynchuk, L.E.,et al., 2008. Postnatal inflammation increases seizure susceptibility in adult rats.J. Neurosci. 28, 6904–6913.
Hamer, H.M., Reis, J., Mueller, H.H., Knake, S., Overhof, M., Oertel, W.H., Rosenow, F.,2005. Motor cortex excitability in focal epilepsies not including the primarymotor area — a TMS study. Brain 128, 811–818.
Harré, E.M., Galic, M.A., Mouihate, A., Noorbakhsh, F., Pittman, Q.J., 2008. Neonatal inflamma-tion produces selective behavioural deficits and alters N-methyl-D-aspartate receptorsubunit mRNA in the adult rat brain. Eur. J. Neurosci. 27, 644–653.
Heida, J.G., Pittman, Q.J., 2005. Causal links between brain cytokines and experimentalfebrile convulsions in the rat. Epilepsia 46, 1906–1913.
Heida, J.G., Boissé, L., Pittman, Q.J., 2004. Lipopolysaccharide-induced febrile convul-sions in the rat: short-term sequelae. Epilepsia 45, 1317–1329.
Heida, J.G., Teskey, G.C., Pittman,Q.J., 2005. Febrile convulsions inducedby the combination oflipopolysaccharide and low-dose kainic acid enhance seizure susceptibility, not epilepto-genesis, in rats. Epilepsia 46, 1898–1905.
Henderson, A.K., Galic, M.A., Fouad, K., Dyck, R.H., Pittman, Q.J., Teskey, G.C., 2011. Largercortical motor maps after seizures. Eur. J. Neurosci. 34, 615–621.
Henry, L.C., Goertzen, C.D., Lee, A., Teskey, G.C., 2008. Repeated seizures lead to alteredskilled behaviour and are associatedwithmorehighly efficacious excitatory synapses.Eur. J. Neurosci. 27, 2165–2176.
Houser, C.R., Esclapez, M., 2003. Downregulation of the alpha5 subunit of the GABA(A) re-ceptor in the pilocarpinemodel of temporal lobe epilepsy. Hippocampus 13, 633–645.
Isaeva, E., Isaev, D., Khazipov, R., Holmes, G.L., 2009. Long-term suppression of GABAergicactivity by neonatal seizures in rat somatosensory cortex. Epilepsy Res. 87, 286–289.
Isaeva, E., Isaev, D., Savrasova, A., Khazipov, R., Holmes, G.L., 2010. Recurrent neonatalseizures result in long-term increases in neuronal network excitability in the ratneocortex. Eur. J. Neurosci. 31, 1446–1455.

700 A.Y. Reid et al. / Neurobiology of Disease 45 (2012) 692–700
Jin, X., Huguenard, J.R., Prince, D.A., 2005. Impaired Cl- extrusion in layer V pyramidalneurons of chronically injured epileptogenic neocortex. J. Neurophysiol. 93,2117–2126.
Kang, T.C., An, S.J., Park, S.K., Hwang, I.K., Bae, J.C., Suh, J.G., Oh, Y.S., et al., 2002. Changesin Na(+)-K(+)-Cl(−) cotransporter immunoreactivity in the gerbil hippocampusfollowing spontaneous seizure. Neurosci. Res. 44, 285–295.
Khalilov, I., Holmes, G.L., Ben-Ari, Y., 2003. In vitro formation of a secondary epileptogenicmirror focus by interhippocampal propagation of seizures. Nat. Neurosci. 6, 1079–1085.
Kleim, J.A., Barbay, S., Nudo, R., 1998. J. Functional reorganization of the rat motor cortexfollowing motor skill learning. J. Neurophysiol. 80, 3321–3325.
Kleim, J.A., Barbay, S., Cooper, N.R., Hogg, T.M., Reidel, C.N., Remple, M.S., Nudo, R.J.,2002. Motor learning-dependent synaptogenesis is localized to functionally reor-ganized motor cortex. Neurobiol. Learn. Mem. 77, 63–77.
Kornelsen, R.A., Boon, F., Leung, L.S., Cain, D.P., 1996. The effects of a single neonatallyinduced convulsion on spatial navigation, locomotor activity and convulsion sus-ceptibility in the adult rat. Brain Res. 706, 155–159.
Labyt, E., Houdayer, E., Cassim, F., Bourriez, J.L., Derambure, P., Devanne, H., 2007.Motor representation areas in epileptic patients with focal motor seizures: a TMSstudy. Epilepsy Res. 75, 197–205.
Lado, F.A., Legatt, A.D., LaSala, P.A., Shinnar, S., 2002. Alteration of the corticalmotormap ina patient with intractable focal seizures. J. Neurol. Neurosurg. Psychiatry 72, 812–815.
Lemmens, E.M., Aendekerk, B., Schijins, O.E., Blokland, A., Beuls, E.A., Hoogland, G.,2009. Long-term behavioral outcome after early-life hyperthermia-induced sei-zures. Epilepsy Behav. 14, 309–315.
Li, H., Prince, D.A., 2002. Synaptic activity in chronically injured, epileptogenic sensory-motor neocortex. J. Neurophysiol. 88, 2–12.
Livak, K.J., Schmittgen, T.D., 2001. Analysis of relative gene expression data using real-timequantitative PCR and the 2(-Delta Delta C(T)) method. Methods 25, 402–408.
Lüddens, H., Wisden, W., 1991. Function and pharmacology of multiple GABAA recep-tor subunits. Trends Pharmacol. Sci. 12, 49–51.
Mesquita, A.R., Tavares, H.B., Silva, R., Sousa, N., 2006b. Febrile convulsions in develop-ing rats induce a hyperanxious phenotype later in life. Epilepsy Behav. 9, 401–406.
Miles, R., 1999. Neurobiology. A homeostatic switch. Nature 397, 215–216.Monfils, M.H., Teskey, G.C., 2004. Induction of long-term depression is associated with
decreased dendritic length and spine density in layers III and V of sensorimotorneocortex. Synapse 53, 114–121.
Monfils, M.H., VandenBerg, P.M., Kleim, J.A., Teskey, G.C., 2004. Long-term potentiationinduces expanded movement representations and dendritic hypertrophy in layerV of rat sensorimotor neocortex. Cereb. Cortex 14, 586–593.
Nealis, J.G., Rosman, N.P., De Piero, T.J., Ouellette, E.M., 1978. Neurologic sequelae of ex-perimental febrile convulsions. Neurology 28, 246–250.
Ni, H., Jiang, Y.W., Bo, T., Wang, J.M., Pan, H., Wu, X.R., 2004. Long-term effects of neonatalseizures on subsequent N-methyl-D-aspartate receptor-1 and gamma-aminobutyricacid receptor A-alpha 1 receptor expression in hippocampusof theWistar rat. Neurosci.Lett. 368, 254–257.
Nudo, R.J., Jenkins, W.M., Merzenich, M.M., 1990. Repetitive microstimulation alters thecortical representation ofmovements in adult rats. Somatosens. Mot. Res. 7, 463–483.
Okabe, A., Ohno, K., Toyoda, H., Yokokura,M., Sato, K., Fukuda, A., 2002. Amygdala kindlinginduces upregulation of mRNA for NKCC1, a Na(+), K(+)-2Cl(−) cotransporter, inthe rat piriform cortex. Neurosci. Res. 44, 225–229.
Pathak, H.R., Weissinger, F., Terunuma, M., Carlson, G.C., Hsu, F.C., Moss, S.J., Coulter, D.A.,2007. Disrupted dentate granule cell chloride regulation enhances synaptic excitabilityduring development of temporal lobe epilepsy. J. Neurosci. 27, 14012–14022.
Pavlidou, E., Tzitiridou, M., Kontopoulos, E., Panteliadis, C.P., 2008. Which factors deter-mine febrile seizure recurrence? A prospective study. Brain Dev. 30, 7–13.
Prevost, F., Costa, M., Carmant, L., Lepore, F., Guillemot, J.P., 2010. Effects of hyperthermic sei-zures on the developing primary visual cortex of the rat. Neuroscience 171, 1120–1130.
Racine, R.J., Steingart, M., McIntyre, D.C., 1999. Development of kindling-prone andkindling-resistant rats: selective breeding and electrophysiological studies. EpilepsyRes. 35, 183–195.
Racine, R.J., Steingart,M., Bureau, Y.,McIntyre, D.C., 2003. Differential sensitivity of geneticallyfast vs. slow kindling rats strains to GABAergic convulsive agents. Neuropharmacology45, 918–924.
Rakhade, S.N., Zhou, C., Aujla, P.K., Fishman, R., Sucher, N.J., Jensen, F.E., 2008. Early alterationsof AMPA receptors mediate synaptic potentiation induced by neonatal seizures. J. Neu-rosci. 28, 7979–7990.
Reid, A.Y., Teskey, G.C., Pittman,Q.J., 2009. AlteredmRNAexpression of GABA andglutamatereceptor subunits and cation chloride co-transporters is associated with increased sei-zure susceptibility after experimental febrile seizure. Epilepsia 50, 323.
Rich, S.S., Annegers, J.F., Hauser, W.A., Anderson, V.E., 1987. Complex segregation analysisof febrile convulsions. Am. J. Hum. Genet. 41, 249–257.
Richichi, C., Brewster, A.L., Bender, R.A., Simeone, T.A., Zha, Q., Yin, H.Z.,Weiss, J.H., et al., 2008.Mechanisms of seizure-induced “transcriptional channelopathy” of hyperpolarization-activated cyclic nucleotide gated (HCN) channels. Neurobiol. Dis. 29, 297–305.
Rivera, C., Voipio, J., Payne, J.A., Ruusuvuori, E., Lahtinen, H., Lamsa, K., Pirvola, U.,Saarma, M., Kaila, K., 1999. The K+/Cl- co-transporter KCC2 renders GABA hyper-polarizing during neuronal maturation. Nature 397, 251–255.
Romijn, H.J., Hofman, M.A., Gramsbergen, A., 1991. At what age is the developing cerebralcortex of the rat comparable to that of the full-term newborn human baby? EarlyHum. Dev. 26, 61–67.
Scantlebury, M.H., Gibbs, S.A., Foadjo, B., Lema, P., Psarropoulou, C., Carmant, L., 2005.Febrile seizures in the predisposed brain: a new model of temporal lobe epilepsy.Ann. Neurol. 58, 41–49.
Shinnar, S., Hesdorffer, D.C., Nordli, D.R., Pellock, J.M., O'Dell, C., Lewis, D.V., Frank, L.M.,Moshé, S.L., Epstein, L.G., Marmarou, A., Bagiella., E., FEBSTAT Study Team, 2008.Phenomenology of prolonged febrile seizures: results of the FEBSTAT study. Neu-rology 71, 170–176.
Spencer, S.J., Auer, R.N., Pittman, Q.J., 2006. Rat neonatal immune challenge alters adultresponses to cerebral ischaemia. J. Cereb. Blood Flow Metab. 26, 456–467.
Stoeckel, M.C., Kleinschmidt, A., Ebner, A., Witte, O.W., Seitz, R.J., 2002. Reorganization ofmotor representation in a patient with epilepsia partialis continua as shown by[O15]-labeled butanol positron emission tomography and functional magnetic reso-nance imaging. J. Neuroimaging 12, 276–281.
Talos, D.M., Fishman, R.E., Park, H., Folkerth, R.D., Follett, P.L., Volpe, J.J., Jensen, F.E., 2006a.Developmental regulation of alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propionicacid receptor subunit expression in forebrain and relationship to regional susceptibilityto hypoxic/ischemic injury. I. Rodent cerebral whitematter and cortex. J. Comp. Neurol.497, 42–60.
Talos, D.M., Follett, P.L., Folkerth, R.D., Fishman, R.E., Trachtenberg, F.L., Volpe, J.J., Jensen, F.E.,2006b. Developmental regulation of alpha-amino-3-hydroxy-5-methyl-4-isoxazole-propionic acid receptor subunit expression in forebrain and relationship to regional sus-ceptibility to hypoxic/ischemic injury. II. Human cerebral white matter and cortex.J. Comp. Neurol. 497, 61–77.
Tandon, S., Kambi, N., Jain, N., 2008. Overlapping representations of the neck and whis-kers in the rat motor cortex revealed by mapping at different anaesthetic depths.Eur. J. Neurosci. 27, 228–237.
Tang, Y.P., Shimizu, E., Dube, G.R., Rampon, C., Kerchner, G.A., Zhuo, M., Liu, G., et al.,1999. Genetic enhancement of learning and memory in mice. Nature 401, 63–69.
Teskey, G.C., Monfils, M.H., VandenBerg, P.M., Kleim, J.A., 2002. Motor map expansionfollowing repeated cortical and limbic seizures is related to synaptic potentiation.Cereb. Cortex 12, 98–105.
Teskey, G.C., Young, N.A., van Rooyen, F., Larson, S.E.M., Flynn, C., Monfils, M.H., Kleim, J.A.,et al., 2007. Induction of neocortical long-termdepression results in smallermovementrepresentations, fewer excitatory perforated synapses, and more inhibitory synapses.Cereb. Cortex 17, 434–442.
Teskey, G.C., Monfils, M.H., Flynn, C., Young, N.A., van Rooyen, F., Henry, L.C., Ozen,L.J., et al., 2008. Motor maps, seizures, and behaviour. Can. J. Exp. Psychol. 62,132–139.
Tsuboi, T., 1976. Polygenic inheritance of epilepsy and febrile convulsions: analysisbased on a computational model. Br. J. Psychiatry 129, 239–242.
Uematsu, S., Lesser, R., Fisher, R.S., Gordon, B., Hara, K., Krauss, G.L., Vining, E.P., et al., 1992.Motor and sensory cortex in humans: topography studied with chronic subduralstimulation. Neurosurgery 31, 59–71.
Urasaki, E., Uematsu, S., Gordon, B., Lesser, R.P., 1994. Cortical tongue area studied bychronically implanted subdural electrodes — with special reference to parietalmotor and frontal sensory responses. Brain 117, 117–132.
van Esch, A., Steyerberg, E.W., Berger, M.Y., Offringa, M., Derksen-Lubsen, G., Habbema,J.D., 1994. Family history and recurrence of febrile seizures. Arch. Dis. Child 70,395–399.
van Rooyen, F., Young, N.A., Larson, S.E.M., Teskey, G.C., 2006. Hippocampal kindlingleads to motor map expansion. Epilepsia 47, 1383–1391.
van Stuijvenberg, M., Steyerberg, E.W., Derksen-Lubsen, G., Moll, H.A., 1998. Tempera-ture, age, and recurrence of febrile seizure. Arch. Pediatr. Adolesc. Med. 152,1170–1175.
Verity, C.M., Butler, N.R., Golding, J., 1985. Febrile convulsions in a national cohort fol-lowed up from birth. I–Prevalence and recurrence in the first five years of life.Br. Med. J. (Clin Res Ed) 290, 1307–1310.
Verity, C.M., Greenwood, R., Golding, J., 1998. Long-term intellectual and behavioraloutcomes of children with febrile convulsions. N. Engl. J. Med. 338, 1723–1728.
Vicini, S., 1991. Pharmacologic significance of the structural heterogeneity of theGABAA receptor-chloride ion channel complex. Neuropsychopharmacology 4,9–15.
Wang, D.D., Kriegstein, A.R., 2008. GABA regulates excitatory synapse formation in theneocortex via NMDA receptor activation. J. Neurosci. 28, 5547–5558.
Werboff, J., Havlena, J., 1963. Febrile convulsions in infant rats, and later behavior. Science142, 684–685.
Wisden, W., Seeburg, P.H., 1992. GABAA receptor channels: from subunits to functionalentities. Curr. Opin. Neurobiol. 2, 263–269.
Young, N.A., Vuong, J., Ozen, L.J., Flynn, C., Teskey, G.C., 2009. Motor map expansion inthe pilocarpine model of temporal lobe epilepsy is dependent on seizure severityand rat strain. Exp. Neurol. 217, 421–428.
Young, N.A., Vuong, J., Flynn, C., Teskey,G.C., 2011. Optimal parameters formicrostimulationderived forelimb movement thresholds and motor maps in rats and mice. J. Neurosci.Methods 196, 60–69.
Zhao, D.Y., Wu, X.R., Pei, Y.Q., Zuo, Q.H., 1985a. Long-term effects of febrile convulsionon seizure susceptibility in P77PMC rat-resistant to acoustic stimuli but susceptibleto kainate-induced seizures. Exp. Neurol. 88, 688–695.
Zhao, D.Y., Wu, X.R., Pei, Y.Q., Zuo, Q.H., 1985b. Kindling phenomenon of hyperthermicseizures in the epilepsy-prone versus the epilepsy-resistant rat. Brain Res. 358,390–393.





























