Embed Size (px)
Citation preview

Metab Brain Dis (2006) 21:189–199DOI 10.1007/s11011-006-9012-7
ORIGINAL PAPER
Acetylsalicylic acid and acetaminophen protect againstoxidative neurotoxicity
H. Maharaj · D. S. Maharaj · S. Daya
Received: 9 May 2005 / Accepted: 12 August 2005 / Published online: 20 July 2006C© Springer Science+Business Media, Inc. 2006
Abstract Due to the implication of oxidative stress in neurodegenerative disorders wedecided to investigate the antioxidant properties of acetylsalicylic acid and acetaminopheneither alone or in combination. The thiobarbituric acid assay (TBA) and the nitroblue tetra-zolium (NBT) assay were used to investigate quinolinic acid (QA)-induced: lipid peroxida-tion and superoxide anion generation in the rat hippocampus, in vivo. The study also shows,using cresyl violet staining, the preservation of structural integrity of neuronal cells follow-ing treatment with acetylsalicylic acid and acetaminophen in QA-lesioned rat hippocampus.Furthermore the study sought to determine whether these agents have any effect on endoge-nous (QA) formation. This study shows that acetylsalicylic acid and acetaminophen inhibitQA-induced superoxide anion generation, lipid peroxidation and cell damage, in vivo, inthe rat hippocampus. In addition these agents inhibit the enzyme, 3-hydroxyanthranilic acidoxygenase (3-HAO), responsible for the synthesis of endogenous QA.
Keywords Quinolinic acid . Oxidative stress . Free radicals . Necrosis . And lipidperoxidation
Introduction
Quinolinic acid (2,3-pyridine dicarboxylic acid) (QA) is a neuroactive metabolite of thetryptophan-kynurenine pathway (Stone, 1993). It is present in both the human and ratbrain and has been implicated in the pathogenesis of a broad spectrum of degenerative,infectious, inflammatory and non-inflammatory human neurological diseases (Heyes, 1996).The neurotoxic effects of QA are well known with administration of quinolinate, in vivo,inducing seizures in various species of mammals (Lapin et al., 1998).
H. Maharaj · D. S. Maharaj · S. Daya (�)Division of Pharmacology, Faculty of Pharmacy, Rhodes University,Grahamstown, 6139 South Africae-mail: [email protected]
Springer

190 Metab Brain Dis (2006) 21:189–199
Free radicals have been postulated to be important mediators of tissue injury in severalneurodegenerative models (Bautista and Spitzer, 1990). The brain is particularly susceptibledue to its high utilization of oxygen and its relatively low concentration of antioxidantenzymes and free radical scavengers (Reiter, 1995). One of the most vulnerable regions ofthe brain to quinolate-induced neurotoxicity is the hippocampus (Schwarcz and Koler, 1983).Rios and Santamaria (1991) have described the lipoperoxidant effect of QA in brain tissue.Lipid peroxidation is believed to be related to free radical formation which results in cellulardamage therefore evidence suggests that QA-induced toxicity could be partially attenuatedby free radical scavengers due to the pro-oxidant properties of QA (Stone, 1993).
The role of endogenous QA in the brain remains obscure and controversial, but thereis increasing evidence that glia can synthesize and release relatively high concentrationsof QA, via the kynurenine pathway (Heyes, 1996). The increase of QA could produce orenhance the amount of neuronal damage produced by primary brain insult. Quinolinic acidinduced neurotoxicity results from the activation of ionic channels through which sodium,potassium and calcium flood into the cell (Stone, 1993). The increased intracellular calciumsets off a cascade of events that culminate in the generation of free radicals. The reductionof endogenous QA synthesis could, therefore, limit the amount of brain damage. A novelapproach to preventing the synthesis of QA is to inhibit the enzyme, 3-hydroxyanthranillicacid oxygenase (3-HAO) (Schwarcz, 2004).
There have been conflicting reports as to whether COX inhibitors such as acetylsali-cylic acid are effective in protecting neurons against neurotoxicity (Aubin et al., 1998).Acetaminophen has been shown to rescue neuronal cells from mitochondrial redox impair-ment, apoptotic DNA fragmentation, lipoperoxidative products and the generation of mal-ondialdehyde (MDA) (Bisaglia et al., 2002). Furthermore acetaminophen has been shown toalso reduce the cytoplasmic accumulation of peroxides and inhibition of NF-κB activationwhich is a multisubunit transcription factor involved in cellular responses to viral infectionand inflammation (Bisaglia et al., 2002). The use of COX-inhibitors has been reported toreduce the incidence of neurodegenerative disorders such as Alzheimer’s disease and Parkin-son’s disease. The mode of action by which acetylsalicylic acid and acetaminophen elicitneuroprotection is however unclear as many mechanisms of action have been inconclusivelypostulated.
Thus the aim of this study was to elucidate whether acetylsalicylic acid and acetaminophenare able to reduce QA-induced oxidative neurotoxicity. This study investigated the effectsof these agents alone and in combination as many pharmaceutical analgesic preparationscontain both drugs. In addition, this study investigates whether acetylsalicylic acid andacetaminophen are capable of reducing endogenous QA synthesis in the brain by inhibiting3-HAO synthesis.
Materials and methods
Chemicals and reagents
Acetylsalicylic acid, acetaminophen, quinolinic acid, 3-hydroxyanthranilic acid (3-HA),nitro-blue tetrazolium (NBT), nitro-blue diformazan (NBD), 2-thiobarbituric acid (98%)(TBA), 1, 1, 3, 3-tetramethoxypropane (98%), and butylated hydroxytoluene (BHT) werepurchased from Sigma Chemical Corporation, St. Louis, MO, U.S.A. Glacial acetic acidand trichloroacetic acid (TCA) were purchased from Saarchem (PTY) Ltd, Krugersdorp(South Africa). Methanol (HPLC grade) and cresyl violet stain were purchased from BDH
Springer

Metab Brain Dis (2006) 21:189–199 191
Laboratory Supplies, Poole, England and distrene plasticizer in xylene (DPX) was purchasedfrom Philip Harris.
Animals
Adult male rats of the Wistar strain, weighing between 200–250 g were used in this exper-iment. The rats were randomly assembled into groups of five and housed in separate cagesin a controlled environment. The Rhodes University Animal Ethics Committee approved allprotocols for the experiments.
Drug treatment
For the superoxide anion, lipid peroxidation and histology assays animals were divided intofive groups (I–V), each group containing five animals. Each animal was anaesthetized withchloral hydrate (400 mg/kg/day, i.p.). QA, dissolved in phosphate buffered saline (PBS),pH = 7.4, (120 nmol in 2 µl) was infused bilaterally into the hippocampi employing a ratbrain stereotaxic apparatus (Stoelting, IL, USA). The stereotaxic coordinates derived fromthe bregma and according to the Konig and Klippel (1963) Atlas were 4.0 mm caudal to thebregma, 2.5 mm lateral to the saggital suture, and 3.2 mm ventral of the dura.
Two µl of PBS was infused bilaterally into the hippocampi, in sham control animals(group I). Group II was infused with QA while Group III-V were infused with QA andreceived intraperitoneal injections of acetylsalicylic acid (100 mg/kg/day), acetaminophen(100 mg/kg/day) or a combination of acetylsalicylic acid and acetaminophen, respectively,20 min prior to receiving the QA and thereafter, each day for 7 days. The animals in group Iand II received the vehicle for the drugs, viz. 40% ethanol and saline solution.
In the 3-HAO study the animals were divided into four groups (n = 5) and were injectedintraperitoneally with acetylsalicylic acid and/or acetaminophen (a total of 100 mg/kg), everyhour for three hours and killed on the fourth hour by decapitation and their livers rapidlyexcised. The liver was used as the lipid source for this assay as it contains a high concentrationof the enzyme, 3-HAO (Mehler, 1956; Heyes, 1996). The dosages of the drugs were chosenfrom previous literature sources where these agents were shown to have neuroprotectiveeffects (Daya et al., 2000; Maharaj et al., 2004).
Dissection of rat hippocampii and homogenate preparation
On the seventh day the rats treated for superoxide anion, lipid peroxidation and histology as-says were killed by decapitation, the brains were quickly removed and the hippocampii weredissected on an ice-cooled dish according to Glowinski and Iversen (1966). Hippocampiiwere homogenized in a glass Teflon homogenizer with 0.1 M phosphate-buffered salinebuffer (PBS), pH 7.4 so as to give a final concentration of 10% w/v. This method of homoge-nizing was necessary to prevent lysosomal damage of the tissue. (Palkovits and Brownstein,1983).
Nitroblue tetrazolium (NBT) assay
A modified method of Sagar et al. (1992) and Das et al. (1990) was used for this assay. Briefly,the rat hippocampal homogenate (1 mL) was incubated with 0.4 ml of a 0.1% NBT solutionin an oscillating water bath for 60 min at 37◦C. Termination of the assay and extraction ofreduced NBT was carried out by centrifugation of the suspensions at 2000 × g for 10 min.
Springer

192 Metab Brain Dis (2006) 21:189–199
The supernatant was decanted and the pellet was resuspended with 2 ml glacial acetic acid.The relative absorbance of the glacial acetic acid fraction was measured at 560 nm andconverted to µmoles Diformazan using a standard curve generated from NBD. Final resultsare expressed as µmoles Diformazan/mg protein.
The thiobarbituric acid (TBA) test
Malondialdehyde (MDA) is a degraded oxidative lipid product from cell membranes, and istaken as a reliable indicator of oxidative stress (Reiter et al., 1995). The foundation of theTBA test is based on the reaction of one molecule of MDA with two molecules of TBA toyield a pink coloured chromagen. A modified method of Sagar et al. (1992) and Das et al.(1990) was used in this assay. An aliquot of 1ml hippocampal homogenate was incubatedin a shaking water bath for 1 h at 37◦C. At the end of the incubation 0.5 ml butylatedhydroxytoluene (BHT) (0.5 g/L in absolute methanol) and 1 ml 25% trichloroacetic acid(TCA) were added to the mixture. The samples were centrifuged at 2000 × g for 20 min at4◦C to remove insoluble proteins. Following centrifugation, 2 ml of protein free supernatantwas removed from each tube and a 0.5 ml aliquot of 0.33% TBA was added to this fraction.All tubes were heated for 1 h at 95◦C in a water bath. After cooling, the TBA-MDA complexeswere extracted with 2 ml of butanol. The absorbance was read at 532 nm and MDA levelswere determined from a standard curve generated from 1, 1, 3, 3-tetramethoxypropane. Thefinal results were expressed as nmol MDA/mg tissue.
Histology
The brains used for histology studies were fixed in Davidson’s fixative prior to embeddingin paraffin wax. The tissue was then processed and sectioned 10 µm thick using a rotarymicrotome. The sections were Nissl stained using cresyl violet. The slides were then mountedwith DPX and viewed under the light microscope and photographed using a digital camera.
3-Hydroxyanthranillic acid oxygenase assay
The activity of the enzyme was assayed spectrophotometrically in liver homogenate asdescribed by Mehler (1956). The homogenate was centrifuged at 12 000 × g for 10 min at 4◦C.An aliquot of 1.5 ml of the cell homogenate supernatant was added to the reaction mixture.The reaction mixture consisted of 1.5 ml of phosphate buffer 0.1M (pH 7.5) containing 10 µMascorbate, 6 mM FeSO4, and 10 µM 3-hydroxyanthranillic acid which was the substrate. Thebuffer extracts were then incubated for 30 min at 37◦C. After the incubation the reaction wasimmediately terminated by the addition of 2 ml, 3M HCl. The mixture was then centrifugedat 12000 × g for 10 min at 4◦C. An aliquot of 3ml of supernatant was removed and used tomeasure enzymatic activity. Enzymatic activity was calculated from the difference betweenproduct content before (blanks) and after incubation. The absorbance was read at 360 nm witha Shimadzu UV 160A UV-visible recording spectrophotometer and an extinction coefficientof 47,500 M−1 cm−1 for the reaction product (α-amino-β-carboxymuconic ε-semialdehyde)was used to calculate the concentration of α-amino-β-carboxymuconic ε-semialdehyde.
Springer
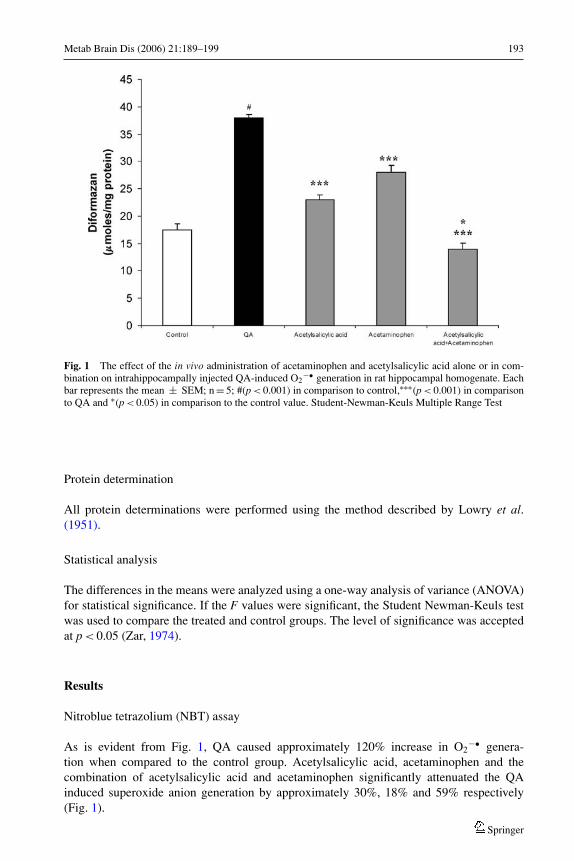
Metab Brain Dis (2006) 21:189–199 193
Fig. 1 The effect of the in vivo administration of acetaminophen and acetylsalicylic acid alone or in com-bination on intrahippocampally injected QA-induced O2
−• generation in rat hippocampal homogenate. Eachbar represents the mean ± SEM; n = 5; #(p < 0.001) in comparison to control,∗∗∗(p < 0.001) in comparisonto QA and ∗(p < 0.05) in comparison to the control value. Student-Newman-Keuls Multiple Range Test
Protein determination
All protein determinations were performed using the method described by Lowry et al.(1951).
Statistical analysis
The differences in the means were analyzed using a one-way analysis of variance (ANOVA)for statistical significance. If the F values were significant, the Student Newman-Keuls testwas used to compare the treated and control groups. The level of significance was acceptedat p < 0.05 (Zar, 1974).
Results
Nitroblue tetrazolium (NBT) assay
As is evident from Fig. 1, QA caused approximately 120% increase in O2−• genera-
tion when compared to the control group. Acetylsalicylic acid, acetaminophen and thecombination of acetylsalicylic acid and acetaminophen significantly attenuated the QAinduced superoxide anion generation by approximately 30%, 18% and 59% respectively(Fig. 1).
Springer
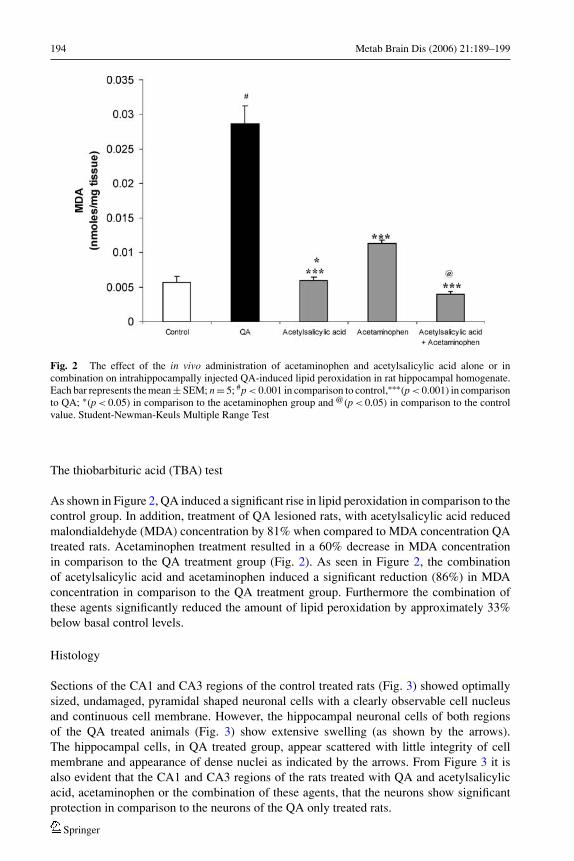
194 Metab Brain Dis (2006) 21:189–199
Fig. 2 The effect of the in vivo administration of acetaminophen and acetylsalicylic acid alone or incombination on intrahippocampally injected QA-induced lipid peroxidation in rat hippocampal homogenate.Each bar represents the mean ± SEM; n = 5; #p < 0.001 in comparison to control,∗∗∗(p < 0.001) in comparisonto QA; ∗(p < 0.05) in comparison to the acetaminophen group and @(p < 0.05) in comparison to the controlvalue. Student-Newman-Keuls Multiple Range Test
The thiobarbituric acid (TBA) test
As shown in Figure 2, QA induced a significant rise in lipid peroxidation in comparison to thecontrol group. In addition, treatment of QA lesioned rats, with acetylsalicylic acid reducedmalondialdehyde (MDA) concentration by 81% when compared to MDA concentration QAtreated rats. Acetaminophen treatment resulted in a 60% decrease in MDA concentrationin comparison to the QA treatment group (Fig. 2). As seen in Figure 2, the combinationof acetylsalicylic acid and acetaminophen induced a significant reduction (86%) in MDAconcentration in comparison to the QA treatment group. Furthermore the combination ofthese agents significantly reduced the amount of lipid peroxidation by approximately 33%below basal control levels.
Histology
Sections of the CA1 and CA3 regions of the control treated rats (Fig. 3) showed optimallysized, undamaged, pyramidal shaped neuronal cells with a clearly observable cell nucleusand continuous cell membrane. However, the hippocampal neuronal cells of both regionsof the QA treated animals (Fig. 3) show extensive swelling (as shown by the arrows).The hippocampal cells, in QA treated group, appear scattered with little integrity of cellmembrane and appearance of dense nuclei as indicated by the arrows. From Figure 3 it isalso evident that the CA1 and CA3 regions of the rats treated with QA and acetylsalicylicacid, acetaminophen or the combination of these agents, that the neurons show significantprotection in comparison to the neurons of the QA only treated rats.
Springer
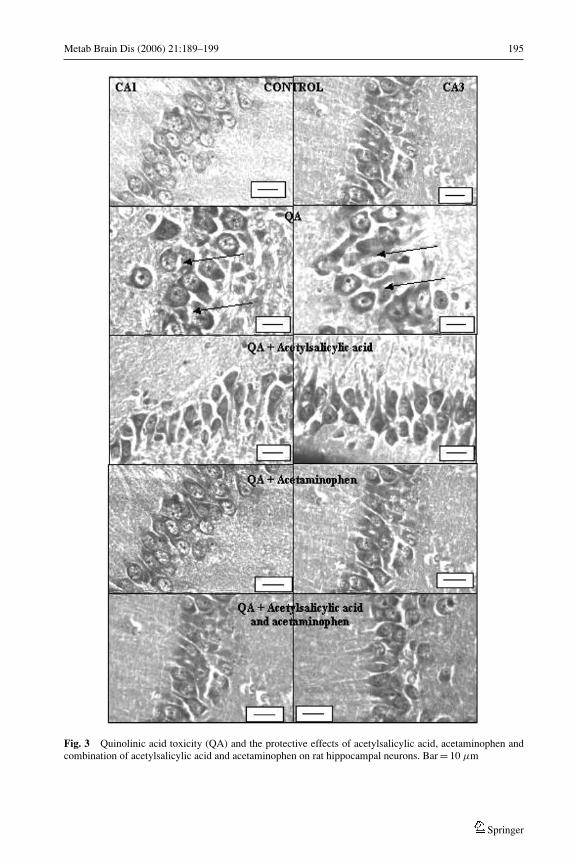
Metab Brain Dis (2006) 21:189–199 195
Fig. 3 Quinolinic acid toxicity (QA) and the protective effects of acetylsalicylic acid, acetaminophen andcombination of acetylsalicylic acid and acetaminophen on rat hippocampal neurons. Bar = 10 µm
Springer
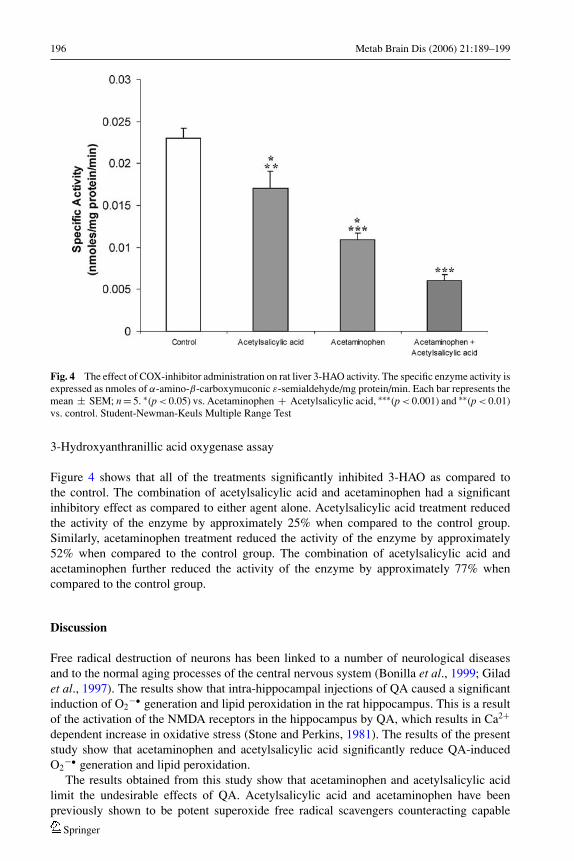
196 Metab Brain Dis (2006) 21:189–199
Fig. 4 The effect of COX-inhibitor administration on rat liver 3-HAO activity. The specific enzyme activity isexpressed as nmoles of α-amino-β-carboxymuconic ε-semialdehyde/mg protein/min. Each bar represents themean ± SEM; n = 5. ∗(p < 0.05) vs. Acetaminophen + Acetylsalicylic acid, ∗∗∗(p < 0.001) and ∗∗(p < 0.01)vs. control. Student-Newman-Keuls Multiple Range Test
3-Hydroxyanthranillic acid oxygenase assay
Figure 4 shows that all of the treatments significantly inhibited 3-HAO as compared tothe control. The combination of acetylsalicylic acid and acetaminophen had a significantinhibitory effect as compared to either agent alone. Acetylsalicylic acid treatment reducedthe activity of the enzyme by approximately 25% when compared to the control group.Similarly, acetaminophen treatment reduced the activity of the enzyme by approximately52% when compared to the control group. The combination of acetylsalicylic acid andacetaminophen further reduced the activity of the enzyme by approximately 77% whencompared to the control group.
Discussion
Free radical destruction of neurons has been linked to a number of neurological diseasesand to the normal aging processes of the central nervous system (Bonilla et al., 1999; Giladet al., 1997). The results show that intra-hippocampal injections of QA caused a significantinduction of O2
−• generation and lipid peroxidation in the rat hippocampus. This is a resultof the activation of the NMDA receptors in the hippocampus by QA, which results in Ca2+
dependent increase in oxidative stress (Stone and Perkins, 1981). The results of the presentstudy show that acetaminophen and acetylsalicylic acid significantly reduce QA-inducedO2
−• generation and lipid peroxidation.The results obtained from this study show that acetaminophen and acetylsalicylic acid
limit the undesirable effects of QA. Acetylsalicylic acid and acetaminophen have beenpreviously shown to be potent superoxide free radical scavengers counteracting capable
Springer

Metab Brain Dis (2006) 21:189–199 197
inducers such as KCN (Maharaj et al., 2004). In addition, the histological analysis showsthat these COX-inhibitors are able to prevent the QA-induced alteration to cellular integrityin the CA1 and CA3 regions of the rat hippocampus. This effect has been described in asimilar study were the authors stated that acute toxicity of QA occurred because of the rapidinflux of Na+ ions into the neurons, which causes passive water and Cl− entry via osmoticpressure. This toxic process may be associated with abnormalities in membrane permeabilityand may be lethal, via osmotic lysis (Southgate, 1999).
The results obtained from this study, using QA as the neurotoxin, provided confirmation,of the antioxidant properties demonstrated by acetylsalicylic acid and acetaminophen, in thein vitro study performed by our laboratory (Maharaj et al., 2004). Acetylsalicylic acid andacetaminophen either alone or in combination have demonstrated their ability to preventO2
−• generation in vivo.Acetylsalicylic acid and acetaminophen are drugs with wide spectrums of pharmacologi-
cal activities and multiple sites of action. Possible mechanisms by which acetylsalicylic acidexhibits neuroprotective properties include the reduction of oxidative stress (Maharaj et al.,2004) and the inhibition of the activation transcription factor, NF–κB (Grilli et al., 1996).Furthermore it has been shown that acetylsalicylic acid rectifies calcium (Ca2+) homeostasisand therefore decreases ROS in human endothelial cells, in vitro (Dragomir et al., 2004). Thealteration in Ca2+ levels by QA, which results in the generation of ROS, could therefore beattenuated by acetylsalicylic acid, which would also explain the potent action exhibited bythis agent in Figures 1–3. The relative extent of each drug preventing oxidative stress doescorrelate with the degree of prevention of structural changes as evident by the highly struc-tured, neuronal cells in acetylsalicylic acid and acetaminophen treated rats when comparedto the QA treated rats. Furthermore the additive effects of the combination of acetylsalicylicacid and acetaminophen as seen in Figures 1 and 2 is due to both these agents acting as freeradical scavengers.
The enzyme cyclooxygenase II (COX-II) as well as inflammatory mediators such as nitricoxide have been reported to be increased in neurodegenerative diseases such as Parkinson’sdisease (Iravani et al., 2002). The non-selective COX-inhibitor, aspirin and the COX-2 pref-erential inhibitor meloxicam have been reported to confer neuroprotection in MPTP-induceddopamine depletion in mice (Teismann and Ferger, 2001). However, other COX activityinhibitors like paracetamol, indomethacin, diclofenac or COX expression inhibitor dexam-ethasone were found to be ineffective in protecting neurons against MPTP neurotoxicity(Aubin et al., 1998). These findings explain the results presented in this study which showsacetylsalicylic acid to be more effective in preventing QA-induced free radical generation. Inaddition, Stipek et al (1997) demonstrated that quinolinate does not have a direct peroxidativeeffect, but that it modulates lipid peroxidation via its interaction with Fe2+. Acetylsalicylicacid has been shown to complex with Fe2+ and Fe3+ (Kotrly and Sucha, 1985). Thus,the chelation of ASA with iron would serve to further explain the greater protective ef-fects of this agent than when compared to acetaminophen against QA-induced free radicalgeneration.
In an attempt to develop therapies for the treatment of neurodegeneration, the kynure-nine pathway has been manipulated in several ways and the most recent approach, is theinhibition of the activity of the enzyme 3-HAO which is responsible for synthesizingthe NMDA receptor agonist, QA (Stone, 2000). The inhibition of this enzyme was aptlyachieved with the use of acetylsalicylic acid and acetaminophen either individually or incombination and thus promotes further research in the possible therapeutic use of theseagents in diseases in which QA is implicated such as Alzheimer’s disease and Huntington’sdisease.
Springer

198 Metab Brain Dis (2006) 21:189–199
Although the results of the present study are encouraging, with respect to these agentspossessing antioxidant properties, the study was limited to just a single dose response andfurther investigations, using lower dosages, are currently being undertaken in our laboratory.Furthermore, the non-quantitative histological data described in this study only provides anindication of QA-induced neuronal change and further studies are required to confirm theneuroprotective role of these drugs.
Conclusion
Acetylsalicylic acid and acetaminophen have been shown, in the present study, to possesspotent antioxidant activity at inhibiting QA-induced superoxide anion generation, lipid per-oxidation and maintained cell integrity in rat hippocampus, in vivo. Furthermore the noveluse of these agents extends to their ability to inhibit the enzyme responsible for the endoge-nous synthesis of QA. This study suggests that the antioxidant properties of these agentscould be exploited as a possible therapeutic approach against neurodegenerative disorders.
Acknowledgements This study was made possible by a grant from the South African National ResearchFoundation to Prof. S. Daya. HM and DSM thanks the Medical Research Council (South Africa) fortheir Post Doctoral scholarships. The authors would like to thank Dave and Sally Morley for technicalassistance.
References
Aubin N, Curet O, Deffois A, Carter C (1998) Aspirin and salicylate protect against MPTP-induced dopaminedepletion in mice. J Neurochem 71:1635–1642
Bautista AP, Spitzer JJ (1990) Superoxide anion generation by in situ perfused rat liver: effect of invivoendotoxin. Am J Physiol 259:G907–G912
Bisaglia M, Venezia V, Piccioli P, Stanzione S, Porcile C, Russo C, Mancini F, Milanese C, Schettini G (2002)Acetaminophen protects hippocampal neurons and PC12 cultures from amyloid beta-peptides inducedoxidative stress and reduces NF-kappaB activation. Neurochem Int 41:43–54
Bonilla E, Tanji K, Hirano M, Vu TH, DiiMauro S, Schon EA (1999) Mitochondrial involvement inAlzheimer’s disease. Biochim Biophys Acta 1410:171–182
Das UN, Padma M, Sagar PS, Ramesh G, Koratkar R (1990) Stimulation of free radical generation in humanleukocytes by various agents including tumor necrosis factor is a calmodulin-dependent process. BiochemBiophys Res Commun 167:1030–1036
Daya S, Walker RB, Anoopkumar-Dukie S (2000) Cyanide-induced free radical production and lipid peroxi-dation in rat brain homogenate is reduced by aspirin. Met Brain Dis 15:203–209
Dragomir E, Manduteanu I, Voinea M, Costache G, Manea A, Simionescu M (2004) Aspirin rectifies calciumhomeostasis, decreases reactive oxygen species, and increases NO production in high glucose-exposedhuman endothelial cells. J Diabetes Complications 18(5):289–299
Gilad E, Cuzzocrea S, Zingarelli B, Salzman AL, Szabo C (1997) Melatonin is a scavenger of peroxynitrite.Life Sci 60:169–174
Glowinski J, Iversen LL (1966) Regional studies of catecholamines in the rat brain. I. The dispositionof [3H]norepinephrine, [3H]dopamine and [3H]dopa in various regions of the brain. J Neurochem13:655–669
Grilli M, Pizzi M, Menio M, Spano F (1996) Neuroprotection by aspirin and sodium salicylate throughblockade of NF-kappaB activation. Science 274:1383–1385
Heyes MP (1996) The kynurenine pathway and neurological disease. Adv Exp Med Biol 398:125–129Iravani MM, Kashefi K, Mander P, Rose S, Jenner P (2002) Involvement of inducible nitric oxide synthase in
inflammation-induced dopaminergic neurodegeneration. Neuroscience 110:49–58Konig JFR, Klippel RA (1963) The Rat Brain. Williams and Wilkins, Baltimore MD, 108–114Kotrly S, Sucha L (1985) Handbook of Chemical Equilibria in Analytical Chemistry, Ellis Horwood Limited,
Chichester, UK, p 163
Springer

Metab Brain Dis (2006) 21:189–199 199
Lapin IP, Mirzaev SM, Rysov IV, Oxenkrug GF (1998) Anticonvulsant activity of melatonin against seizuresinduced by quinolate, kainite, glutamate, NMDA, and pentylenetetrazole in mice. J Pineal Res 24:215–218
Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the folin phenol reagent.J Biol Chem 193:265–267
Maharaj DS, Saravanan KS, Maharaj H, Mohanakumar KP, Daya S (2004) Acetaminophen and aspirin inhibitsuperoxide anion generation and lipid peroxidation, and protect against 1-methyl-4-phenyl pyridinium-induced dopaminergic neurotoxicity in rats. Neurochem Int 44:355–360
Mehler AH (1956) Formation of picolinic and quinolinic acids following enzymatic oxidation of3-hydroxyanthranilic acid. J Biol Chem 218:241–54
Palkovits M, Brownstein MJ (1983) Microdissection of brain areas by punch technique. In: Cuello AC (Ed.),Brain Microdissection Techniques. Wiley, New York, pp 1–36
Paxinos G, and Watson C (1998) The Rat Brain in Sterotaxic Coordinates. Academic Press, New York, pp41–41
Reiter RJ (1995) Oxidative processes and antioxidant defense mechanisms in the ageing brain. J FASEB9:526–533
Reiter RJ, Melchiori D, Poeggeler B, Barlow Walden L, Chuang J, Oritz GG, Acuna-Castrviejo D (1995) Areview of the evidence supporting melatonin’s role as an antioxidant. J Pineal Res 18:1–11
Rios C, Santamaria A (1991) Quinolinic acid is a potent lipid peroxidant in rat brain homogenates. NeurochemRes 16:1139–1143
Sagar PS, Das UN, Koratkar R, Ramesh G, Padma M, Kumar GS (1992) Cytotoxic action of cis-unsaturatedfatty acids on human cervical carcinoma (HeLa) cells: relationship to free radicals and lipid peroxidationand its modulation by calmodulin antagonists. Cancer Lett 63:189–198
Schwarcz R (2004) The kynurenine pathway of tryptophan degradation as a drug target. Curr Opin Pharmacol4:12–7
Schwarcz R, Koler C (1983) Differential vulnerability of central neurons of the rat to quinolinic acid. NeurosciLett 38:85–90
Southgate G, Daya S (1999) Melatonin reduces quinolinic acid-induced lipid peroxidation in rat brain ho-mogenate. Metab Brain Dis 14:165–71
Stipek S, Stastny F, Platenik J, Crkovska J, Zima T (1997) The effect of quinolinate on rat brain lipidperoxidation is dependent on iron. Neurochem Int 30:233–237
Stone TW (1993) Neuropharmacology of quinolinic and kynurenic acids. Pharmacol Rev 45:309Stone TW (2000) Development and therapeutic potential of kynurenic acid and kynurenine derivatives for
neuroprotection. TIPS 21:149–54Stone TW, Perkins MN (1981) Quinolinic acid: A potent endogenous excitant at amino acid receptors in CNS.
European J Pharmacol 72:411–412Teismann P, Ferger B (2001) Inhibition of the cyclooxygenase isoenzymes COX-1 and COX-2 provide
neuroprotection in the MPTP-mouse model of Parkinson’s disease. Synapse 39:167–174Zar JH (1974) Biostatistical Analysis, 1st edn. Prentice Hall, Englewood Cliffs, NJ, pp 151–466
Springer





























