Embed Size (px)
Citation preview

0306-4522(94)00339-4
Neurosrience Vol. 62, No. 4, pp. I t 03- 1120, 1994 Elsevier Science Ltd
Copyright Q 1994 IBRO Printed in Great Britain. All rights reserved
0306-4522/94 $7.00 + 0.00
ACTIVATION OF DOPAMINERGIC NEURONS MODULATES VENTRAL PALLIDAL RESPONSES EVOKED
BY AMYGDALA STIMULATION
R. J. MASLOWSKI-COBUZZI and T. C. NAPIER*
Neurosience Program, and Department of Pharmacology and Experimental Therapeutics, Loyola University Chicago, Stritch School of Medicine, Maywood, IL 60153, U.S.A.
Abstract-The ventral pallidurn is a basal forebrain region that is thought to integrate cognitive processes with motoric behaviors. These functions are influenced by ventral pallidal inputs, which include projections from the amygdala and the ventral tegmental a~a/substantia nigra zona compacta. By examining the consequences of this convergence at the neuronal level, the present study indicates that electrical activation of ventral tegmental regions releases dopamine in the ventral pallidum which subsequently modulates pallidal electrophysiological responses evoked by stimulating the amygdala.
Stimulation-evoked responses were characterized for extracellular single unit recordings of spon- taneously active ventral pallidal neurons from chloral hydrate anesthetized rats. Stimulation of the amygdala evoked short latency (G 12 ms; possibly monosynaptic) and/or long latency (> 12 ms; polysy- naptic) responses in all ventral pallidal neurons tested. Fifty-nine per cent of the tested neurons responded to ventral tegmental stimulation with short latency inhibition, and these neurons were often sensitive to microiontophoreti~lly applied dopamine. Iontophoresis of dopamine antagonists SCH23390 (a Dl antagonist) or sulpiride (a D2 antagonist) attenuated the ventral tegmental-indu~d inhibitions. These observations indicate that the evoked responding was the result of endogenously released dopamine, and that Dl and D2 receptors were involved in this effect.
Ninety-two per cent of the ventral pallidal neurons that demonstrated short latency responses to amygdala stimulation also exhibited short latency responses to activation of the ventral tegmentum. This suggests that these inputs often converge onto the same pallidal neurons. Amygdala-evoked responses were consistently attenuated by prior stimulation of the ventral tegmentum. Similarly, microiontophoretic ejection of dopamine attenuated amygdala-evoked effects. These results indicate that dopamine modulates amygdala-evoked pallidal responses, Such modulation may contribute to the integrative functions of the ventral pallidurn,
The ventral pallidum (VP) largely comprises the sub- commissural extension of the globus pallidus and the rostra1 aspects of the sublenticular substantia innomi- nata.29,30 The VP receives inputs from several limbic brain regions, inctuding the amygdala, the nucleus accumbens (NA), and the limbic cortex (for review see Ref. 88). The VP also is innervated by the ventral tegmental area (VTA) and substantia nigra pars com- pacta (SNc) (for reviews see Ref. 53, 57, 88). These midbrain regions are associated with mesolimbic and extrapyramidal motoric functions, respectively. VP efferent projections include medial dorsal thalamuss6 and brainstem regions74’76 that govern motoric behav- iors4,” and limbic cortical areas5.7’ that are criticaf for mnemonic processes (for review see Ref. 60). This circuitry has led to the proposal that the VP is critical to the ‘“limbic-motor integration” necessary for com- plex behavior performance (for review see 46).
*To whom correspondence should be addressed. Abbreoiufions: AMN. amygdaloid nuclei; DA, dopamine;
NA, nucleus accumbens; SNc, substantia nigra zona compacta; VP, ventral pallidum: VTA, ventral tegmental area.
Dopamine (DA) is a major transmitter within the ascending projection from the VTA/SNc, and bio- chemical studies demonstrated the presence of DA and its metabolites within the VP.2a.55 A role for DA in VP function is likely, for DA administered directly within the VP alters neuronal firing52.ss.57 and loco- motion34,51 in rats. Electrical stimulation of the SNc evokes short latency inhibitory VP neuronal re- sponses53 that are suggestive of monosynaptic effects, but it is unknown whether VTA stimulation similarly affects VP activity, nor has it been verified that DA is the transmitter responsible for this inhibition.
Recent studies using molecular biological tech- niques have described at least six distinct genes coding for DA receptor subtypes within the brain (for reviews see Ref. 9, IO). The protein products of these genes fall into two pharmacologically identifiable “families”. The D,-like family can be labeled by the agonist SKF38393 and the antagonist SCH23390. The D,-like family can be identified by the agonist quinpirole and the antagonist sulpiride. Since the degree of discrimination by present pharmacologic tools is at the level of these receptor families and not at specific subtypes, Dl will be used hereafter to
1103

1104 R. J. Maslowski-Cobuzzi and T. C. Napier
denote the Dl-like family and D2 will denote the D2-like family. Radiohgands with high affinity to D, or D, receptors have been used to confirm the presence of both groups within the VP.‘.“.57 Adminis- tration of SKF38393 or quinpirole alters firing of VP neurons,43.52 however, the involvement of the DA receptor subtypes in the responses of VP neurons to endogenously released DA is unknown.
Clues to the synaptic physiology of VP DA can be obtained from previous studies of other do- paminoceptive brain regions. For example, the NA receives glutamatergic inputs from amygdaloid nuclei (~~~)18,36.64.68 and electrical stimulation of AMN often evokes short latency excitation in the NA that may be mediated monosynaptically. Iontophoretic applications of DA within the NA, or VTA activation (which releases endogenous DA) prior to stimulating AMN, attenuates AMN-evoked excitatory responses in the NA.84 These findings agree with biochemical studies demonstrating the ability of DA to inhibit presynaptic release of glutamate in striatal re- gions. ‘5.44.45~58.66~80 The VP projection from AMN also is [email protected] and contains glutamate (and/or aspartate) as one of its transmitters.‘.r8 These anatom- ical observations are supported by electrophysiologi- cal studies showing that iontophoretic applications of glutamate increases VP firing8,j7 and AMN stimu- lation can evoke short latency excitatory effects on VP neuronal activity. 85 Since midbrain dopaminergic and AMN efferents converge within the VP, then like the NA, DA modulation of AMN-evoked signals may occur within the VP. The present study was designed to test this hypothesis and to address current paucities in the literature regarding DA inputs to the VP. These objectives were accomplished by answering the following questions: (1) does stimu- lation of the VTA evoke similar responses in VP neurons as SNc stimulation? (2) Are the VP-evoked responses due to the endogenous release of DA? (3) Can convergence of AMN and VTA/SNc inputs onto single VP neurons be demonstrated electro- physiologically? (4) Does VTA/SNc stimulation alter AMN-evoked activity of VP neurons? (5) Does ex- ogenous DA applied within the VP mimic the effects of VTA/SNc stimulation on VP neuronal activity? (6) Can DA act as a neuromodulator of AMN-evoked VP activity?
EXPERIMENTAL PROCEDURES
Surgical preparation of animals
Male Sprague-Dawley rats (Harlan Inc., Indianapolis, IN) weighing 250-35Og were anesthetized with chloral hydrate (400 mg/kg, i.p.; Sigma Chem. Co., St Louis, MO). A lateral tail vein was cannulated for intravenous injection of anesthetic supplements to maintain surgical levels of anesthesia. The animals then were placed into a stereotaxic apparatus (David Kopf Instruments, Tujunga, CA) with the nose piece set at - 3.3 mm. The skull was exposed and holes were drilled through the skull overlying the VP, AMN, VTA and SNc. Stereotaxic coordinates used for recording from the VP were 0.5 mm posterior to Bregma (P), 2.5 mm
lateral to the midline (L) and 7.5-8.5 mm below dura (V). Stimulating electrode coordinates were as follows: AMN, 2.8 P, 4.8 L and 7.2-7.5 V; VTA, 5.7 P, 1.0 L and 7.8-8.0 V; and SNc, 5.7 P, 1.6-2.0 L and 7.6-7.8 V. Rectal temperature of the animals was monitored throughout the experiments, and maintained at 35537°C with a thermo- statically controlled heating pad (Fintronics Inc., Orange, CT)
Exrracellular single-neuron recordings
Extracellular recording of individual VP neurons was accomplished with single barrel glass micropipettes (A-M Systems, Inc., Everett, WA) that were heat-pulled (Narishige PE-2 vertical electrode puller, Tokyo, Japan), the tips broken back to approximately 2pm, and then were heat-bent to allow attachment to the microiontophoretic pipettes (see below). The microelectrodes were filled with 2 M NaCl saturated with fast green dye (Fisher Scientific Co., St Louis, MO), which produced an in uirro impedance of 36 MR, measured at 165 Hz with a Winston Electronics microelectrode tester (San Francisco, CA). A hydraulic microdrive (Trent Wells, South Gate, CA) was used to advance the electrode assembly, and spontaneously firing VP neurons were sampled. Action potentials were filtered from background (2OOHz and 2 kHz) with a high- impedance amplifier (Fintronics, Inc., Orange, CT), and were monitored on a storage oscilloscope (Tektronix Inc., Beaverton, OR) and an audiomonitor (Grass Instru- ments Co., Quincy, MA). To quantify the neuronal activity, the digital output from the window discriminator (Fintron- its, Inc.) was transmitted to an IBM AT compatible com- puter, and Brainstorm Systems Spikes to Stats software was used to display, store and analyse data.
Microionlophoresis and marerials
To allow for microiontophoretic drug application, pre-as- sembled five-barrel glass pipettes (A-M Systems, Inc.) were heat pulled, and the tips broken back to 8--12pm. The pipettes were glued to the recording microelectrodes so that the pulled portions were in parallel, and the microelectrode tip extended 8-15 pm below the multibarrel tip. The center barrel of the pipette was filled with a 2 M NaCl solution and was used for automatic current balancing of the other barrels. The remaining barrels each contained one of the following drugs: dopamine hydrochloride (DA; 0.2 M, pH 4; Sigma, St Louis, MO), SCH23390 ((R)-( + )-8-chloro- i,3,4,5-tetrahydro-3-methyi-5-phenyl-lH:j-benzazepin-7-o1 hemimaleate: 0.01 M. oH 4: Scherina-Plouah. Bloomfield. NJ), or sulpiride (5(aminosulfony&N-[(I-ethyl-2-pyrro- lidinyl)methyl]-2-methoxybenzamide; 0.02 M, pH 4; Sigma). The concentration of DA and its antagonists was selected to allow the passage of ionized drug through high resistance (20-60 MR) barrels of the microiontophoretic pipette. ‘).‘O In some experiments, the D,/D, dopamine receptor antagonist, flupentixol (4-[3-[2-(trifluoromethyl)- 9H-thioxanthen-9-ylidenelpropyll-1-piperazine ethanol di- hydrochloride; Research Biochemicals International, Natick, MA) was injected through a lateral tail vein cannula (0.54 mg/kg). All drugs were dissolved in sterile deionized water, except SCH23390 and sulpiride. These were dissolved with HCI, then diluted with deionized water, and droplet addition of ION NaOH was used to establish a pH of 4.
At the start of each penetration into the brain, a six-chan- nel current generator and programmer (Fintronics, Inc.) was used to apply maximal ejection currents to each barrel for at least 30min to concentrate drugs at the pipette tips. Drugs were ejected with cationic currents, and anionic currents of 10 nA were used to retain the drugs within the pipette. Neuronal waveforms were monitored continu- ously for the duration of each experiment, and data were excluded if, on the rare occasion, iontophoretic drug appli- cation reduced the amplitude or widened duration of the

Dopamine attenuates amygdala-evoked ventral pallidal activity 1105
action potential (indicative of non-specific membrane effects).
Microiontophoretic application of dopaminergic antag- onists (SCH23390 and sulpiride) was used to determine if VP activity evoked by stimulation of VTA/SNc was a consequence of DA being released into the VP. Since antagonists have no efficacy, changes in VTA/SNc-evoked responses during VP iontophoresis of these antagonists would be the result of competitive antagonism with the endogenous neurotransmitter. The ejection current selected for antagonist application was that which attenuated VP responses to iontophoresed DA or the homotypic DA agonist.”
Electrical stimulation
For stimulation of the AMN or VTA/SNc, stainless steel concentric bipolar electrodes (NEX-100; 0.5 mm outer lead diameter, 0.2 mm inner lead diameter, with 0.5 mm exposed on the tips that are separated by 0.5 mm; David Kopf Instr.) were used to deliver current generated by Grass Instr. (Quincy, MA) S88 stimulators, each coupled to a Grass stimulation isolation unit (SW 5) and a Grass constant current unit (CCU I). For single pulse stimulation, 0.1 ms monophasic pulses were applied at 1 Hz with a current range from 0.05-1.5 mA. The VTA/SNc were also stimulated with a train consisting of 10,O. 1 ms pulses. The pulses within the train occurred at IO Hz, and the train was delivered at 0.5 Hz.
A peristimulus rate histogram was generated for each sample of 128 stimulation epochs, using a bin width of 2 ms. The mean number of action potentials (or counts) per bin occurring 80-100 ms before the stimulation was considered as the control interstimulus baseline. The onset of an evoked response was delineated by the first of three consecutive bins with counts that were greater than 1 S.D. from the control mean; offset was determined by the same criterion. Counts occurring within this evoked response period were com- pared to the number of counts occurring within the same time period during “treatment”. Treatments were (I) DA antagonist administration for VTA/SNc-induced evoked responses; and (2) DA application and VTA/SNc stimu- lation for AMN-evoked responses. Such treatments altered the interstimulus baseline in some VP neurons. To compen- sate for any influence this baseline change might have on action potential occurrence during the evoked response period (which would bias interpretation of treatment effects on evoked responding), an “expected value” was deter- rnined.s,% The expected value was calculated by multiplying the number of counts within an evoked component of the control sample by the ratio of the treatment prestimulus mean to the control prestimulus mean. The expected value (expressed as mean counts/bin) was compared to the mean counts/bin actually observed during the evoked period of the treatment sample. If the observed mean differed from the expected value by at least 25%, the treatment was con- sidered effective.
To evaluate responses of VP neurons to electrical acti- vation of the AMN and/or the VTA/SNc, the following protocol was used: (1) a spontaneously active VP neuron was isolated and a stable baseline rate obtained. (2) AMN or VTA/SNc stimulation-evoked effects on VP activity were characterized using a current that was shown in preliminary stimulation experiments to be effective for these regions (i.e., 0.5 and 0.3 mA for AMN and VTA/SNc, respectively). (3) If an evoked response occurred in the VP neuron, the procedure was repeated at lower stimulation currents to determine the response threshold. In contrast, if the VP neuron was insensitive, currents up to 1.5 mA were used, after which an unresponsive neuron was con- sidered insensitive, and was not studied further. (4) The VP neuron was assessed for sensitivity to stimulation of the other afferent brain region to determine if the neuron
exhibited convergence. (5) DA was microiontophoretically- applied as described above. If VP neuronal activity was altered by DA, SCH23390 or sulpiride was co-applied with DA, using an antagonist ejection current level which is known to attenuate the homotypic agonist, but not the heterotypic agonist, in the VP.52 The ability of the antagonist to attenuate VTA/SNc evoked VP response also was tested. If the antagonist(s) did not attenuate the DA agonist-induced effect, the maximal ejection current that did not produce nonspecific membrane effects was used for antagonist ejection during the period of VTA/SNc stimulation. (6) During antagonist application, VTAjSNc stimulation was repeated using a stimulation current that evoked a response of approximately 50% of the maximum observed with the range of stimulation currents tested, or ECuSO.
For those VP neurons that responded to both AMN and VTA/SN stimulation, the physiology of this convergence was evaluated as follows: (1) DA was microiontophoreti- tally-applied during AMN stimulation. (2) The VTA/SNc was stimulated with a train consisting of 10 pulses. (Even though midbrain dopaminergic neurons fire with either single spikes or in bursts of up to 10 pulse~,*~~*~ Yim and MogensonW observed that while both single or train stimu- lation of the VTA is sufficient to evoke NA responses, only train stimulation is capable of attenuating the NA excitatory responses to AMN stimulation. This may be due to the fact that the burst firing pattern elicits higher concentrations of extracellular DA.22*23,4’) (3) Following a delay of 100 ms, a single pulse was delivered to the AMN using the Ecu50 level, and VP neuronal activity was recorded for 900ms. (4) This entire VTA/SNc train-AMN single pulse sequence was repeated at a rate of once every 2 s for 128 stimulation epochs.
Histology
At the end of each experiment, the rat received an overdose of chloral hydrate i.v. The location of the last recording site was marked with fast green dye by passing anionic current through the recording barrel of the electrode for It%20min. The brain was removed, mounted, frozen and then cut in 50 pm sections with a microtome. The brain sections were stained with cresyl violet and the location of each recording site was determined using the fast green spot and electrode tracks as references.
Several comparisons were made between placement of the midbrain and AMN stimulating electrodes, and the anatom- ical location and profile of the electrophysiological response of stimulation-evoked VP neurons. To describe the topogra- phy of sensitive VP neurons, the VP was divided into subregions according to previously described differences in histochemical markers and circuitry.W”’ The infracommis- sural VP was divided into ventromedial and dorsolateral regions, and the caudal areas into the sublenticular VP and substantia innominata.
Statistics
Parametric data are presented as mean f S.E.M. Chi- square analysis was used to detect changes in (1) the number of VTA/SNc-evoked VP responses that were sensitive or insensitive to the different antagonist treatments, and (2) the number of AMN-evoked VP responses that were attenuated or insensitive to DA application versus VTA/SNc stimu- lation. Criterion of significance for all statistical tests was P < 0.05.
RESULTS
Characteristics of spontaneously active ventral paNida
neurons
The data for this study were assessed from record-
ings of 146 neurons that were histologically verified
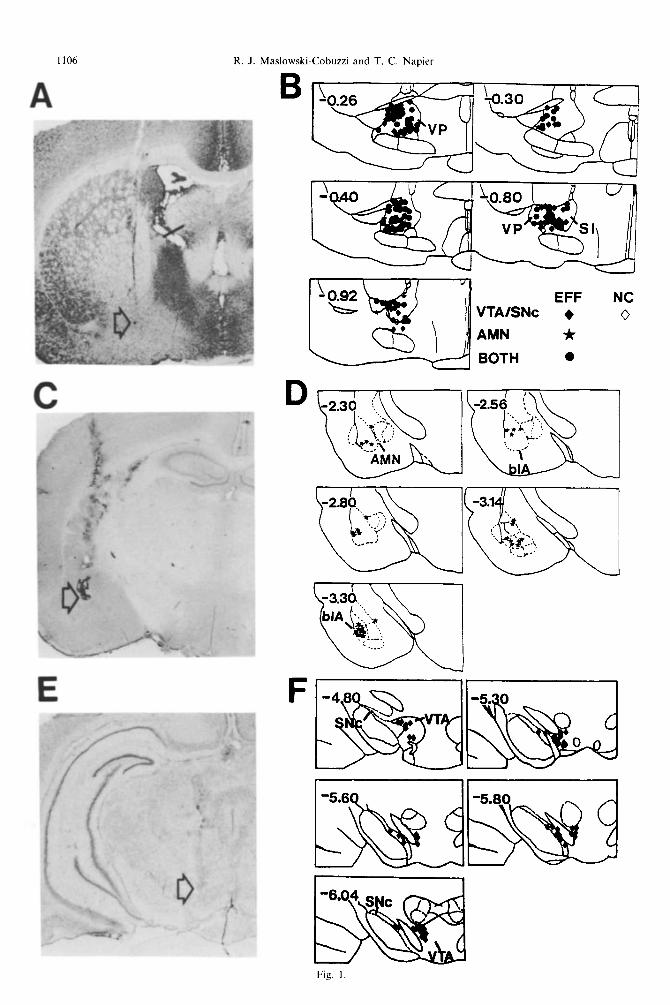
1106 R. J. P vlaslowski-Cobuzzi and T. C. Napier
B
EFF NC
o
Fig. 1

Dopamine attenuates amygdala-evoked ventral pallidal activity 1107
Table I. Summary of VP evoked responses to stimulation of the amygdaloid nuclei and ventral tegmental area substantia nigra zona compacta
Response category*
Region stimulated
No. neurons sensitive to
stimulation vs no. tested
Short latency Long latency (<12ms) (>12ms)
Inhibition Excitation Inhibition Excitation
AMN 86/86 39186 42186 67186 52186 (45%) (49%) (78%) (60%)
VTA/SNc 135/138 80/135 41/135 701135 70/135 (59%) (30%) (52%) (52%)
*Response category was defined by the ms latency to the onset of a particular evoked response, as described in Experimental Procedures. Data are presented as the occurrence of each response in the indicated category as the numerator and the number of neurons evoked by stimulating the indicated region as the denominator. Eighty per cent of VP neurons responding to AMN stimulation and 60% to VTA/SNc activation exhibited a complex response sequence consisting of more than one evoked response category (e.g., inhibition followed by excitation; consequently, this one neuron would be counted in two response categories). Thus, the ratio in each response category indicates the proportion of the cells that exhibited that particular response, making the sum of the percents across all four categories greater than 100
to be within the VP region (Fig. lA, B). All neurons
were spontaneously active with a mean discharge rate of 13.1 + 0.83 spikes per second. The most frequently encountered recording demonstrated a firing pattern with a broad interspike interval histogram denoting an irregular distribution of action potential occur- rence intervals. Of the recorded neurons, 132 (90%) displayed biphasic and 14 (10%) had triphasic action potentials. One hundred and twenty six recordings (86%) exhibited initially negative action potential waveforms, while the remaining 20 (14%) were in-
itially positive. The duration of the action potentials
was 1.1 f 0.02 ms, with a peak to peak amplitude of
331 f 12 pV. The recordings obtained in this study
had similar characteristics to those from previous studies of the ~~~8~32~42~43~48.50.52.57.81.82
Responses of ventral pallidal neurons to stimulation of amygdaloid nuclei
Figure 1B illustrates the location of VP neurons affected by stimulating the AMN. Most of the AMN stimulation sites were within the basolateral nucleus (Fig. lC, D), and it was from this nucleus that VP responses were evoked with the lowest stimulation
intensity. No trends in the electrophysiological profile of VP responses could be attributed to the anterior/posterior placement of the stimulating electrode within the basolateral nucleus.
The distribution of latencies of evoked VP excit- atory responses was bimodal, peaking at 4-12 ms, and from 16-18 ms. Latency distribution for the evoked inhibitions also was bimodal, with one peak between 2-6 ms, and another from 1618 ms; a distri- bution which agrees with that reported by Yim and Mogenson.85 Thus, the VP responses (both excitatory and inhibitory) were categorized as short (< 12 ms) or long (> 12 ms) latency (Table 1). The most frequently observed AMN-evoked VP response category was a long latency inhibition (Table 1) but since AMN stimulation often induced a complex sequence of evoked responses, this long latency inhibition often occurred in conjunction with evoked excitations. For many VP neurons tested, raising the stimulation intensity increased the complexity of the response pattern (Fig. 2A, B). The AMN-evoked responses did not appear to be mediated antidromically since the criteria of constant onset latency and the ability to follow pulse-pair stimulation at high frequency4” were not fulfilled.
Fig. 1. Location of recording sites in the VP or sublenticular substantia innominata (SI), and stimulation sites within the AMN, in particular within the basolateral nucleus (blA), and the VTA or SNc. (A, C, E) are cresyl violet-stained coronal sections that are representative photomicrographs of the recording and stimulation sites. The arrow in (A) indicates the fast green dye deposited at the recording microelectrode tip; the arrows in (B) and (E) indicate the position of the stimulating electrode tips. The right column illustrates the location of individual recording and stimulation sites on maps redrawn from Paxinos and Watson6* and many recording sites overlapped. Number in the upper left corner indicates the distance (mm) from Bregma. The sections in (A, C, E) are illustrated in the -0.80, -2.56 and -5.30 stereotaxic maps in (B, D, F), respectively. In (B), the filled symbols indicate effective (EFF) and the open symbols indicate ineffective (NC for no change) stimulation of AMN (stars), VTA/SNc (diamonds) or convergence of AMN and VTA/SNc (circles) on the activity of VP neurons. All VP neurons evaluated with AMN
stimulation responded to at least one of the tested stimulation currents.
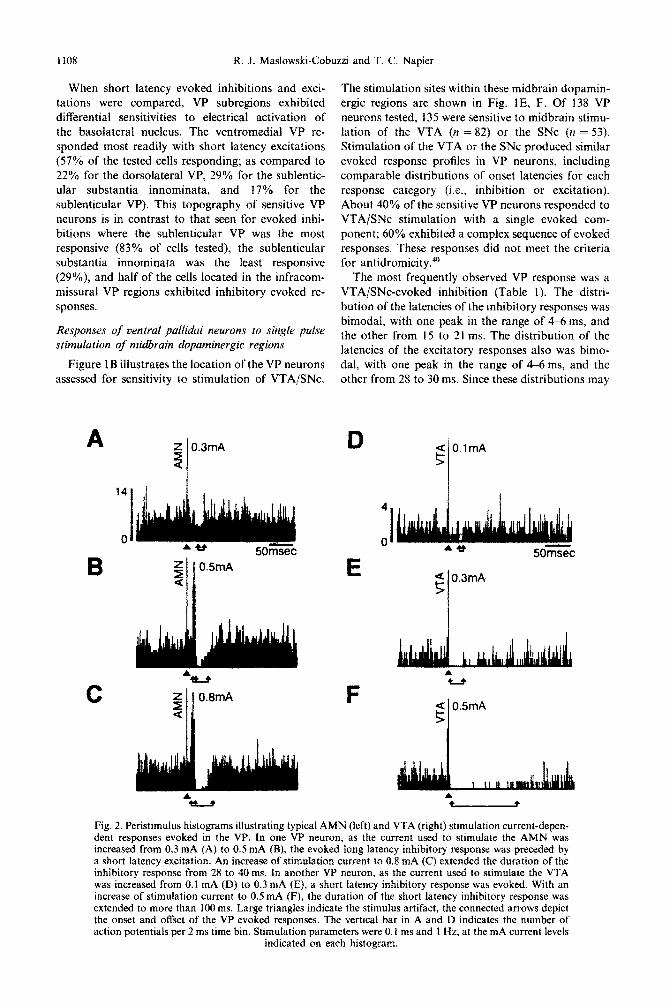
I108 R. J. Maslowski-Cobuzzi and T. C. Napier
When short latency evoked inhibitions and exci- tations were compared, VP subregions exhibited differential sensitivities to electrical activation of the basoiateral nucleus. The ventromedial VP re- sponded most readily with short latency excitations (57% of the tested cells responding; as compared to 22% for the dorsolateral VP, 29% for the sublentic- ular substantia innominata, and 17% for the sublenticular VP). This topography of sensitive VP neurons is in contrast to that seen for evoked inhi- bitions where the sublenti~ular VP was the most responsive (83% of cells tested), the sublenticular substantia innominata was the least responsive (29%), and half of the cells located in the infracom- missural VP regions exhibited inhibitory evoked re- sponses.
Responses of ventral pall~dai neurons to single pulse stimulation of midbrain dopaminergic regions
Figure 1 B illustrates the location of the VP neurons assessed for sensitivity to stimulation of VTA/SNc.
C
03mA
50msec 3 0.5mA a
I
The stimulation sites within these midbrain dopamin- ergic regions are shown in Fig. 1 E, F. Of 138 VP neurons tested, 135 were sensitive to midbrain stimu- lation of the VTA (n = 82) or the SNc (n = 53). Stimulation of the VTA or the SNc produced similar evoked response profiles in VP neurons, including comparable distributions of onset latencies for each response category (i.e., inhibition or excitation). About 40% of the sensitive VP neurons responded to VTA/SNc stimulation with a single evoked com- ponent; 60% exhibited a complex sequence of evoked responses. These responses did not meet the criteria for antidromicity.40
The most frequently observed VP response was a VTA/SNc-evoked inhibition (Table 1). The distri- bution of the latencies of the inhibitory responses was bimodal, with one peak in the range of 4-6 ms, and the other from 15 to 21 ms. The distribution of the latencies of the excitatory responses also was bimo- dal, with one peak in the range of 4-6ms, and the other from 28 to 30 ms. Since these distributions may
D
E
F
O.lmA
0.3mA
Y + 9
Fig. 2. Peristimulus histograms illustrating typical AMN (left) and VTA (right) stimulation current-depen- dent responses evoked in the VP. In one VP neuron, as the current used to stimulate the AMN was increased from 0.3 mA (A) to 0.5 mA (B), the evoked long latency inhibitory response was preceded by a short latency excitation. An increase of stimulation current to 0.8 mA (C) extended the duration of the inhibitory response from 28 to 40 ms. In another VP neuron, as the current used to stimulate the VTA was increased from 0. I mA (D) to 0.3 mA (E), a short latency inhibitory response was evoked. With an increase of stimulation current to 0.5 mA (F), the duration of the short latency inhibitory response was extended to more than 100 ms. Large triangles indicate the stimulus artifact, the connected arrows depict the onset and offset of the VP evoked responses. The vertical bar in A and D indicates the number of action potentials per 2 ms time bin. Stimulation parameters were 0. I ms and I Hz, at the mA current levels
indicated on each histogram.
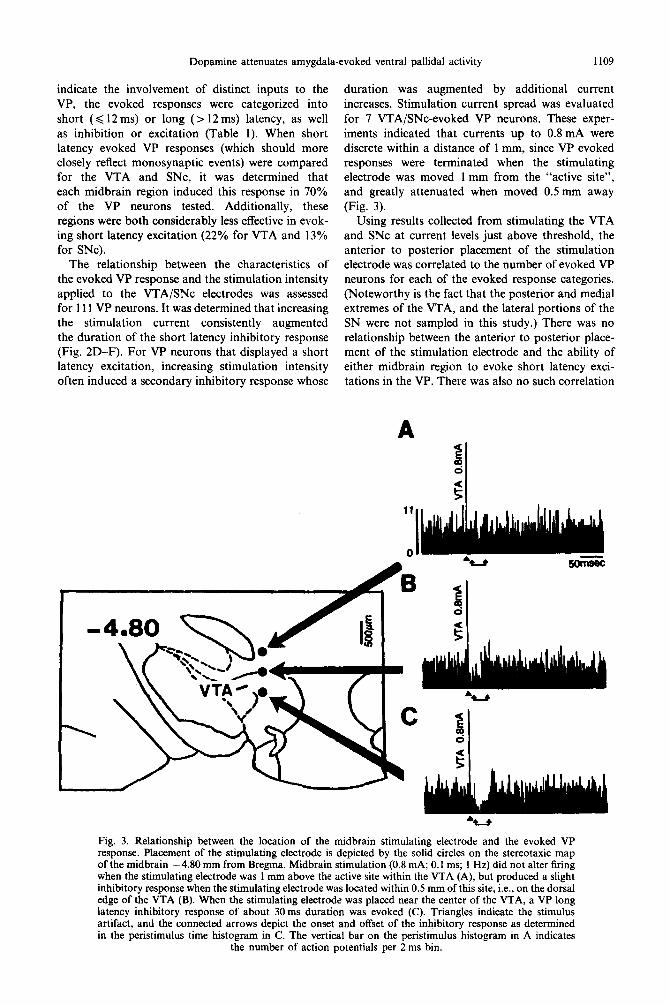
Dopamine attenuates amygdala-evoked ventral pallidal activity 1109
indicate the involvement of distinct inputs to the VP, the evoked responses were categorized into short (< 12 ms) or long (> 12ms) latency, as well as inhibition or excitation (Table 1). When short latency evoked VP responses (which should more closely reflect monosynaptic events) were compared for the VTA and SNc, it was determined that each midbrain region induced this response in 70% of the VP neurons tested. Additionally, these regions were both considerably less effective in evok- ing short latency excitation (22% for VTA and 13% for SNc).
The relationship between the characteristics of the evoked VP response and the stimulation intensity applied to the VTA/SNc electrodes was assessed for 111 VP neurons. It was determined that increasing the stimulation current consistently augmented the duration of the short latency inhibitory response (Fig. 2D-F). For VP neurons that displayed a short latency excitation, increasing stimulation intensity often induced a secondary inhibitory response whose
duration was augmented by additional current increases. Stimulation current spread was evaluated for 7 VTA/SNc-evoked VP neurons. These exper- iments indicated that currents up to 0.8 mA were discrete within a distance of 1 mm, since VP evoked responses were terminated when the stimulating electrode was moved 1 mm from the “active site”, and greatly attenuated when moved 0.5 mm away (Fig. 3).
Using results collected from stimulating the VTA and SNc at current levels just above threshold, the anterior to posterior placement of the stimulation electrode was correlated to the number of evoked VP neurons for each of the evoked response categories. (Noteworthy is the fact that the posterior and medial extremes of the VTA, and the lateral portions of the SN were not sampled in this study.) There was no relationship between the anterior to posterior place- ment of the stimulation electrode and the ability of either midbrain region to evoke short latency exci- tations in the VP. There was also no such correlation
f d
s
Fig. 3. Relationship between the location of the midbrain stimulating electrode and the evoked VP response. Placement of the stimulating electrode is depicted by the solid circles on the stereotaxic map of the midbrain -4.80 mm from Bregma. Midbrain stimulation (0.8 mA; 0.1 ms; 1 Hz) did not alter firing when the stimulating electrode was 1 mm above the active site within the VTA (A), but produced a slight inhibitory response when the stimulating electrode was located within 0.5 mm of this site, i.e., on the dorsal edge of the VTA (B). When the stimulating electrode was placed near the center of the VTA, a VP long latency inhibitory response of about 30ms duration was evoked (C). Triangles indicate the stimulus artifact, and the connected arrows depict the onset and offset of the inhibitory response as determined in the peristimulus time histogram in C. The vertical bar on the peristimulus histogram in A indicates
the number of action potentials per 2 ms bin.
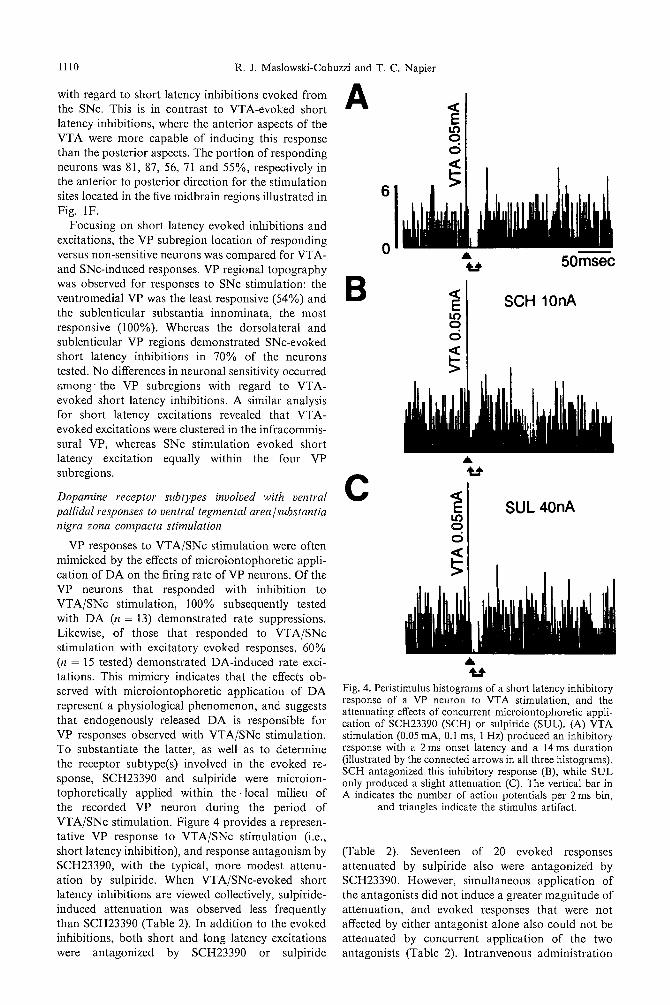
1110 R. J. Maslowski-Cobuzzi and T. C. Napier
with regard to short latency inhibitions evoked from the SNc. This is in contrast to VTA-evoked short latency inhibitions, where the anterior aspects of the VTA were more capable of inducing this response than the posterior aspects. The portion of responding neurons was 81, 87, 56, 71 and 55%, respectively in the anterior to posterior direction for the stimulation sites located in the five midbrain regions illustrated in Fig. 1F.
Focusing on short latency evoked inhibitions and excitations, the VP subregion location of responding versus non-sensitive neurons was compared for VTA- and SNc-induced responses. VP regional topography was observed for responses to SNc stimulation: the ventromedial VP was the least responsive (54%) and the sublenticular substantia innominata, the most responsive (100%). Whereas the dorsolateral and sublenticular VP regions demonstrated SNc-evoked short latency inhibitions in 70% of the neurons tested. No differences in neuronal sensitivity occurred among the VP subregions with regard to VTA- evoked short latency inhibitions. A similar analysis for short latency excitations revealed that VTA- evoked excitations were clustered in the infracommis- sural VP, whereas SNc stimulation evoked short latency excitation equally within the four VP subregions.
Dopamine receptor subtypes involved with ventral pallidal responses to ventral tegmental area /substantia nigra zona compacta stimulation
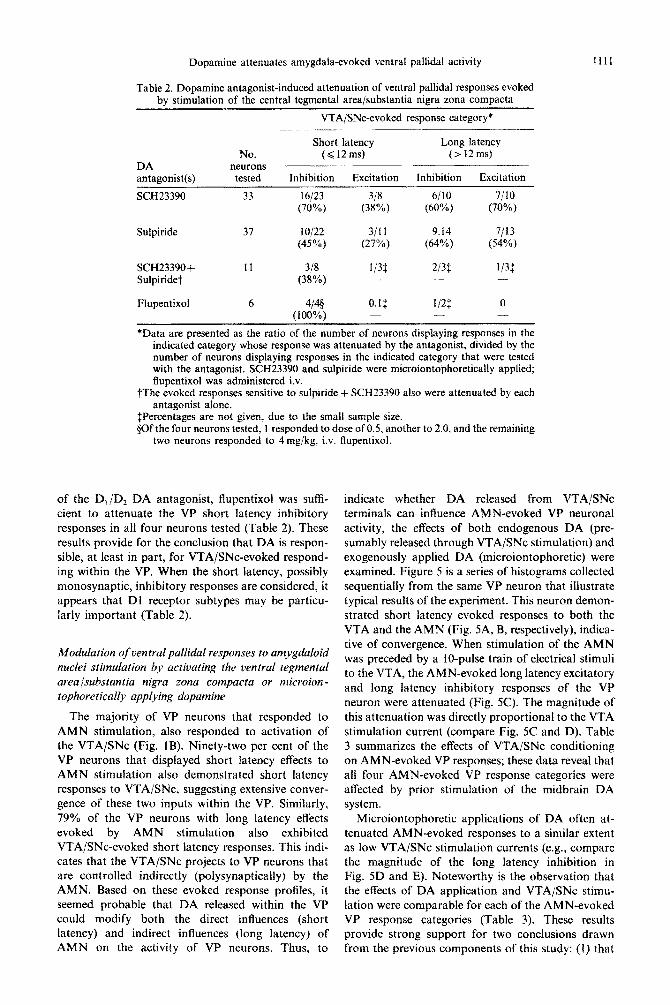
VP responses to VTA/SNc stimulation were often mimicked by the effects of microiontophoretic appli- cation of DA on the firing rate of VP neurons. Of the VP neurons that responded with inhibition to VTA/SNc stimulation, 100% subsequently tested with DA (n = 13) demonstrated rate suppressions. Likewise, of those that responded to VTA/SNc stimulation with excitatory evoked responses, 60% (n = 15 tested) demonstrated DA-induced rate exci- tations. This mimicry indicates that the effects ob- served with microiontophoretic application of DA represent a physiological phenomenon, and suggests that endogenously released DA is responsible for VP responses observed with VTA/SNc stimulation. To substantiate the latter, as well as to determine the receptor subtype(s) involved in the evoked re- sponse, SCH23390 and sulpiride were microion- tophoretically applied within t h e local milieu of the recorded VP neuron during the period of VTA/SNc stimulation. Figure 4 provides a represen- tative VP response to VTA/SNc stimulation (i.e., short latency inhibition), and response antagonism by SCH23390, with the typical, more modest attenu- ation by sulpiride. When VTA/SNc-evoked short latency inhibitions are viewed collectively, sulpiride- induced attenuation was observed less frequently than SCH23390 (Table 2). In addition to the evoked inhibitions, both short and long latency excitations were antagonized by SCH23390 or sulpiride
A
0 = 5 0 m s e c
B o.,on O d
A
Fig. 4. Peristimulus histograms of a short latency inhibitory response of a VP neuron to VTA stimulation, and the attenuating effects of concurrent microiontophoretic appli- cation of SCH23390 (SCH) or sulpiride (SUL). (A) VTA stimulation (0.05 mA, 0.1 ms, 1 Hz) produced an inhibitory response with a 2 ms onset latency and a 14 ms duration (illustrated by the connected arrows in all three histograms). SCH antagonized this inhibitory response (B), while SUL only produced a slight attenuation (C). The vertical bar in A indicates the number of action potentials per 2 ms bin,
and triangles indicate the stimulus artifact.
(Table 2). Seventeen of 20 evoked responses attenuated by sulpiride also were antagonized by SCH23390. However, simultaneous application of the antagonists did not induce a greater magnitude of attenuation, and evoked responses that were not affected by either antagonist alone also could not be attenuated by concurrent application of the two antagonists (Table 2). Intranvenous administration
Dopamine attenuates amygdaia-evoked ventral pallidal activity
Table 2. Dopamine antagonist-induced attenuation of ventral pallidal responses evoked by stimulation of the central tegmental area/substantia nigra zona compacta
VTA/SNc-evoked response category*
Short latency Long latency No. (< 12ms) (> 12ms)
DA neurons -
antagonist(s) tested Inhibition Excitation Inhibition Excitation
SCH23390 33 16123 318 6/10 7/10 (70%) (38%) (60%) (70%)
Sulpiride 37 lOj22 3111 9.14 7/13 (45%) (27%) (64%) (54%)
SCH23390+ 11 3/8 l/3$ 213i l/3$ Sulpiridet (38%) - - -
Flupentixol 6 4/4# O.l$ I/2$ 0 (100%) -
*Data are presented as the ratio of the number of neurons displaying responses in the indicated category whose response was attenuated by the antagonist, divided by the number of neurons displaying responses in the indicated category that were tested with the antagonist. SCH23390 and sulpiride were microiontophoretically applied; flupentixol was administered iv.
TThe evoked responses sensitive to sulpiride + SCH23390 also were attenuated by each antagonist alone.
ipercentages are not given, due to the small sample size. EjOf the four neurons tested, 1 responded to dose of 0.5, another to 2.0, and the remaining
two neurons responded to 4 mg/kg, iv. flupentixol.
1111
of the D,/D, DA antagonist, flu~ntixol was sufh- cient to attenuate the VP short latency inhibitory responses in all four neurons tested (Table 2). These results provide for the conclusion that DA is respon- sible, at least in part, for VTA/SNc-evoked respond- ing within the VP. When the short latency, possibly monosynaptic, inhibitory responses are considered, it appears that Dl receptor subtypes may be particu- larly important (Table 2).
Modulation of ven tral pallidal responses to amygdaloid nuclei stimulation by activating the ventral tegmental arealsubstantia nigra zona compacta or microion- tophoret~cal~y appiying dopam~~e
The majority of VP neurons that responded to AMN stimulation, also responded to activation of the VTA/SNc (Fig. IB). Ninety-two per cent of the VP neurons that displayed short latency effects to AMN stimulation also demonstrated short latency responses to VTA/SNc, suggesting extensive conver- gence of these two inputs within the VP. Similarly, 79% of the VP neurons with long latency effects evoked by AMN stimulation also exhibited VTA/SNc-evoked short latency responses. This indi- cates that the VTA/SNc projects to VP neurons that are controlled indirectly (polysynapticaily) by the AMN. Based on these evoked response profiles, it seemed probable that DA released within the VP could modify both the direct influences (short latency) and indirect influences (long latency) of AMN on the activity of VP neurons. Thus, to
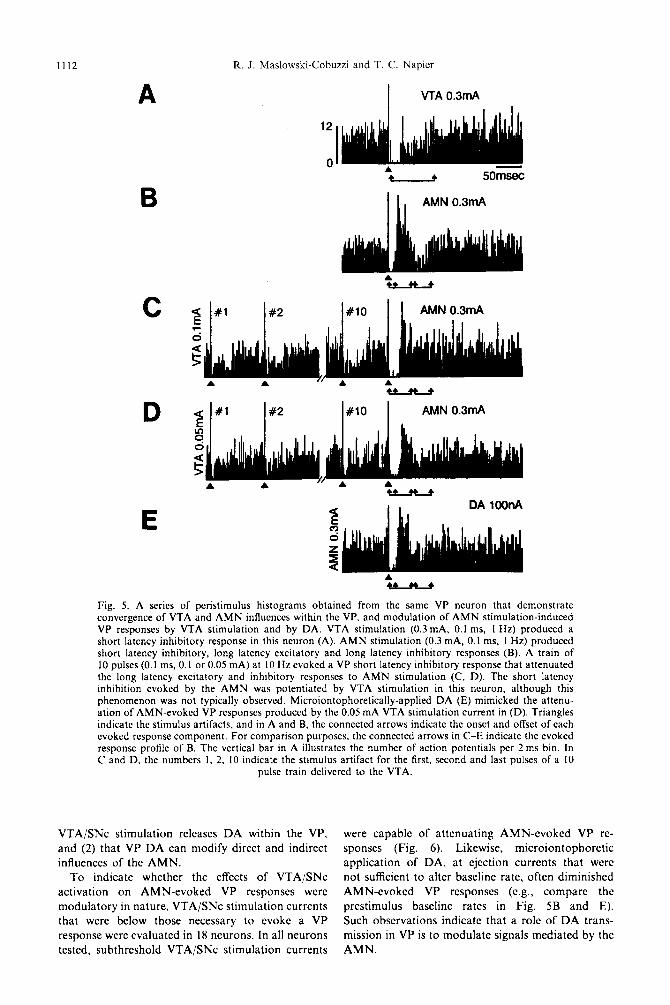
indicate whether DA released from VTA/SNc terminals can influence AMN-evoked VP neuronat activity, the effects of both endogenous DA (pre- sumably released through VTA/SNc stimulation) and exogenously applied DA (microiontophoretic) were examined. Figure 5 is a series of histograms collected sequentially from the same VP neuron that illustrate typical results of the experiment. This neuron demon- strated short latency evoked responses to both the VTA and the AMN (Fig. 5A, B, respectively), indica- tive of convergence. When stimulation of the AMN was preceded by a lo-pulse train of electrical stimuli to the VTA, the AMN-evoked long latency excitatory and long latency inhibitory responses of the VP neuron were attenuated (Fig. 5C). The magnitude of this attenuation was directly proportional to the VTA stimulation current (compare Fig. SC and D). Table 3 summarizes the effects of VTA/SNc conditioning on AMN-evoked VP responses; these data reveal that all four AMN-evoked VP response categories were affected by prior stimulation of the midbrain DA system.
Microiontophoretic applications of DA often at- tenuated AMN-evoked responses to a similar extent as low VTA/SNc stimulation currents (e.g., compare the magnitude of the long latency inhibition in Fig. 5D and E). Noteworthy is the observation that the effects of DA application and VTA/SNc stimu- lation were comparable for each of the AMN-evoked VP response categories (Table 3). These results provide strong support for two conclusions drawn from the previous components of this study: (1) that
l112 R.J . Maslowski-Cobuzzi and T. C. Napier
A VTA 0.3mA
'il ,~ ,p 50msec
B
C
A A & A
D I I I ,MNo.3 d
E Z
Fig. 5. A series of peristimulus histograms obtained from the same VP neuron that demonstrate convergence of VTA and AMN influences within the VP, and modulation of AMN stimulation-induced VP responses by VTA stimulation and by DA. VTA stimulation (0.3 mA, 0.1 ms, 1 Hz) produced a short latency inhibitory response in this neuron (A). AMN stimulation (0.3 mA, 0.1 ms, 1 Hz) produced short latency inhibitory, long latency excitatory and long latency inhibitory responses (B). A train of 10 pulses (0.1 ms, 0.1 or 0.05 mA) at 10 Hz evoked a VP short latency inhibitory response that attenuated the long latency excitatory and inhibitory responses to AMN stimulation (C, D). The short latency inhibition evoked by the AMN was potentiated by VTA stimulation in this neuron, although this phenomenon was not typically observed. Microiontophoretically-applied DA (E) mimicked the attenu- ation of AMN-evoked VP responses produced by the 0.05 mA VTA stimulation current in (D). Triangles indicate the stimulus artifacts, and in A and B, the connected arrows indicate the onset and offset of each evoked response component. For comparison purposes, the connected arrows in C-E indicate the evoked response profile of B. The vertical bar in A illustrates the number of action potentials per 2 ms bin. In C and D, the numbers 1, 2, 10 indicate the stimulus artifact for the first, second and last pulses of a 10
pulse train delivered to the VTA.
VTA/SNc s t imulat ion releases D A within the VP, and (2) tha t VP D A can modify direct and indirect influences of the A M N .
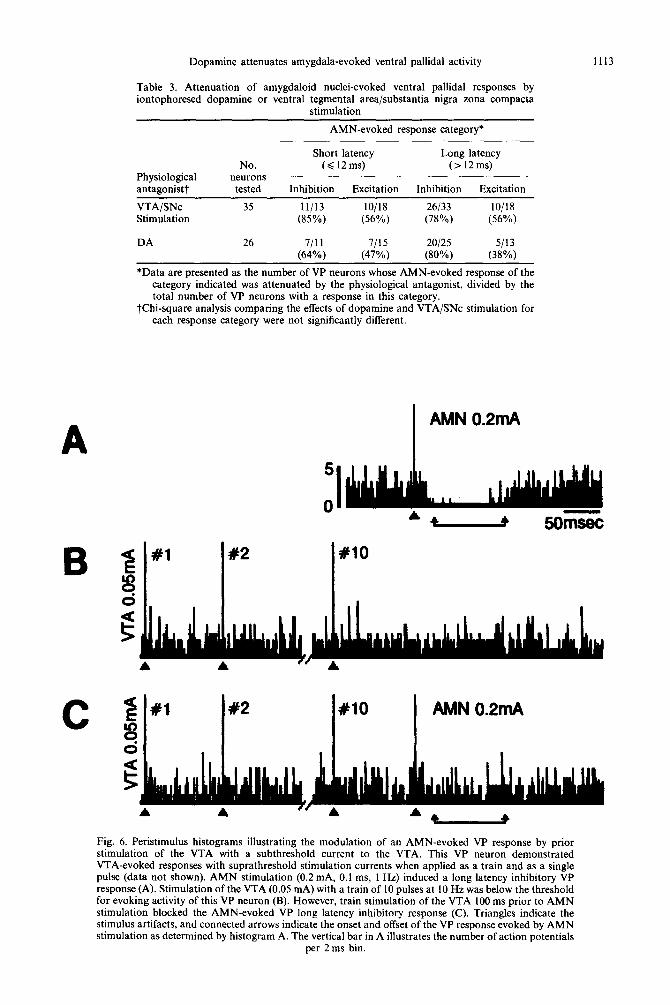
To indicate whether the effects of VTA/SNc act ivat ion on A M N - e v o k e d VP responses were modula to ry in nature, VTA/SNc s t imulat ion currents tha t were below those necessary to evoke a VP response were evaluated in 18 neurons. In all neurons tested, subthreshold VTA/SNc s t imulat ion currents
were capable of a t tenua t ing A M N - e v o k e d VP re- sponses (Fig. 6). Likewise, micro ion tophore t ic appl icat ion of DA, at ejection currents tha t w e r e not sufficient to alter baseline rate, often diminished A M N - e v o k e d VP responses (e.g., compare the prest imulus baseline rates in Fig. 5B and E). Such observat ions indicate tha t a role of D A trans- mission in VP is to modula te signals media ted by the A M N .
Dopamine attenuates amygdala-evoked ventral pallidal activity 1113
Table 3. Attenuation of amygdaloid nuclei-evoked ventral pallidal responses by iontophoresed dopamine or ventral tegmental area/substantia nigra zona compacta
stimulation
AMN-evoked response category*
A
Physiological antagonistt
No. neurons
tested
Short latency Long latency (<12ms) (>12ms)
Inhibition Excitation Inhibition Excitation
VTA/SNc Stimulation
DA
35 II/13 lO/lS 26133 lo/18 (85%) (56%) (78%) (56%)
26 7/11 7115 20125 5113 (64%) (47%) (80%) (38%)
*Data are presented as the number of VP neurons whose AMN-evoked response of the category indicated was attenuated by the physiological antagonist, divided by the total number of VP neurons with a response in this category.
tChi-square analysis comparing the effects of dopamine and VTA/SNc stimulation for each response category were not significantly different.
C #2 x10 AMN 0.2mA
Fig. 6. Peristimulus histograms illustrating the modulation of an AMN-evoked VP response by prior stimulation of the VTA with a subthreshold current to the VTA. This VP neuron demonstrated VTA-evoked responses with suprathreshold stimulation currents when applied as a train and as a single pulse (data not shown). AMN stimulation (0.2 mA, 0. I ms, 1 Hz) induced a long latency inhibitory VP response (A). Stimulation of the VTA (0.05 mA) with a train of 10 pulses at 10 Hz was below the threshold for evoking activity of this VP neuron (B). However, train stimulation of the VTA 100 ms prior to AMN stimulation blocked the AMN-evoked VP long latency inhibitory response (C). Triangles indicate the stimulus artifacts, and connected arrows indicate the onset and offset of the VP response evoked by AMN stimulation as determined by histogram A. The vertical bar in A illustrates the number of action potentials
per 2 ms bin.

1114 K. J. Maslowski-Cobu~i and T. C. Napier
DISCUSSION
The present study demonstrated that VTA/SNc activation evokes neuronal responses within the VP that are temporally and pharmacologically con- sistent with a monosynaptic dopaminergic input. This work also revealed that VP DA can modulate AMN influences on VP neuronal activity. The func- tional consequences of these findings are discussed below.
~i~~i~ur~ties between the qfjkcts of’ ~nd~g~n~us and
exogenous dopatnine on activity of ventral pallidul neurons
With the demonstration that VTA/SNc stjmulation alters the activity of VP neurons, the present results provide functional relevance for anatomical reports of dopaminergic innervation of the VP from the VTA/SNC,2X.31.h7.7?.87 and concurs with previous find- ings that VP neurons often are inhibited by SNc stimulation.s’
Many of the VTASNc-evoked VP responses ex- hibited latencies that were less than I2 ms. The distance from the VTA/SNc to the VP (as calculated from stereotaxic coordinates) is 5-6 mm. With a conduction velocity of dopaminergic fibers of ap- proximately 0.5 m/s”.” and a synaptic delay account- ing for another 0.5 ms,” the onset of monosynaptic DA-mediated effects can occur within 12ms. In VP neurons with slower firing rates, the onset of evoked responses appeared to occur immediately after the stimulus artifact because the interspike interval of the spontaneous activity was longer than the delay for orthodromic conduction, whereas neur- ons with faster firing rates displayed action potentials prior to the evoked response that were not driven by the stimulus. Short latency VTA/SNc-evoked re- sponses were attenuated by intra-VP application of DA antagonists, providing strong evidence that at least some of the VTA/SNc inputs to the VP are dopaminergic.
The topographical relationship between the mid- brain stimulation site and the anatomical location of VP neurons responding with a short latency inhi- bition corresponds to the topography of the ascend- ing dopdminergic projection. Electrical activation of either region altered VP neuronal firing, and anatom- ical reports demonstrate that both the VTA and the SNc provide afferents to the VP (for reviews see Refs 53, 57, 8X). No differences in neuronal sensitivity occurred among the VP subregions with regard to VTA-evoked short latency inhibitions. Since the medial most aspect of the VTA was not sampled in the present study, this observation functionally supports anatomical reports that the lateral aspects of the VTA project to both the ventro- medial and dorsolateral VP.14 In contrast to the VTA, SNc-evoked short latency inhibitions were more readily observed in the dorsolateral and sublenticular regions than in the ventromedial VP. This agrees
the observation by Klitenick et al.“’ that the SNc projection to the VP avoids the ventromedial region. The topography of VTA/SNc-evoked short latency excitations did not follow the aforementioned trend, The reason for this disparity is unknown, but may relate either to an involvement of a non-dopamin- ergic projection with a different trajectory, or simply to the difficulty of this type of analysis with the relatively small sample size obtained in the present study for the short latency excitation category. Nevertheless, given that VP neuronal activity was altered by both VTA and SNc stimulation, it is likely that limbic (VTA) and extrapyramidal motor (SNc) dopaminergic projections are able to direct VP signal- ing.
According to criteria established by Werman7’ for potential neurotransmitters, exogenously applied DA should mimic the response to evoking the endogen- ous system. The present results fulfill this criterion, since all of the inhibitory, and 60% of the excitatory VP responses produced by V’I’A/SNc stimulation were mimicked by microiontophoretic applications of DA within the VP. In addition, SCH23390 or sulpiride applied within the VP antagonized the evoked inhibitions (present study), as well as DA-in- duced VP rate suppressions.i’ Thus. it appears that DA is released within the VP during electrical stimu- lation of the VTASNc and where it alters VP firing through the activation of D, and DZ receptor sub- types. These observations agree with reports of VP sensitivity to locally administered Dl and D2 agonists.“.ch
VTA/SNc stimulation also evoked VP responses with long onset latencies characteristic of polysynap- tic transmission. These polysynaptic events may be mediated through interneurons within the VP or through afferents to the VP. Regions that input the VP, including the NA and AMN, receive midbrain dopaminergic projections.‘“.” Thus, VTA/SNc- evoked responses of the VP may be mediated in-
directly through alterations in neuronal activity of the NA and/or AMN. The effects of DA transmission within the AMN on VP neuronal activity have not yet been evaluated, but Mogenson and co11eaguesR’~X5 have studied the potential involvement of the NA. NA neurons display a variety of responses to VTA stimulation, and 50% of the responses are attenuated by systemic administration of the DA antagonist,
haloperidokn4 suggesting that DA is involved in the VTA-evoked NA suppression. Microinjections of DA into the NA alters VP neuronal firing.** These reports indicate that DA released into the NA following VTA activation can indirectly regulate VP neuronal ac- tivity. However, the present study revealed that long latency responses of VP neurons to VTA/SNc stimu- lation were readily attenuated by intra-VP appli- cation of DA antagonists. These results suggest that polysynaptically mediated VTA/SNc-evoked VP re- sponses can also be controlled by DA at the level of the VP.

Dopamine attenuates amygdala-evoked ventral pallidal activity Ill5
Effects of endogenous and exogenous dopamine on amygdaloid nuclei-evoked responses of ventral pallidal neurons
Both VTA/SNc and AMN projections terminate within the VP. The present study indicates that this convergence is extensive and occurs at the level of individual VP neurons since 92% of the VP neurons tested displayed short latency (monosynaptic) evoked responses to VTA/SNc and AMN stimulation. Eighty five per cent of the short latency inhibitory, and 56% of the short latency excitatory AMN- evoked VP responses were attenuated by prior stimu- lation of the VTA/SNc suggesting that VTA/SNc activation can function as a physiological antagonist of the monosynaptic AMN input to the VP. Intra-VP DA application attenuated 64% of the inhibitory, and 47% of the excitatory short latency VP responses to AMN stimulation, suggesting that DA may be the physiologic antagonist.
Since actual VP synaptic concentrations of the putative endogenously released DA or exogenously administered DA cannot be determined with the approaches employed in this study (i.e., electrical stimulation of VTA/SNc and DA microiontophore- sis, respectively), the possibility exists that concen- trations were excessive and not physiologically relevant. To address this concern, the prospect was tested that AMN-evoked VP responses could be attenuated even at VTA/SNc stimulation current and DA iontophoretic ejection current levels that were lower than those needed to directly alter VP activity. It was demonstrated that the subthreshold current levels for both VTA/SNc stimulation and DA iotophoresis were sufficient to profoundly antagonize AMN-evoked VP responding. These results provide strong evidence that the protocols employed in this study likely reflect normal physiological processing in the VP. More central to the hypothesis of this study, these results also indicate that DA is a modulator of AMN influences on the VP.
AMN-evoked VP responses occurred with the low- est stimulation current intensity from electrodes placed within the basolateral nucleus. This agrees with anatomical reports that the basolateral nucleus sends the most substantial amygdaloid projection to the VP.36 While the topography of VP afferents from the basolateral nucleus has not been described anatomically, it was observed in the present study that stimulating the basolateral nucleus induced short latency excitations most readily from neurons located in the ventromedial VP subregion, whereas the sublenticular VP responded more often with short latency inhibitions.
Anatomical studies suggest that glutamate may mediate the AMN-evoked short latency excitatory responses in the VP.‘.‘* Microiontophoresis of gluta- mate increases VP neuronal firing and this excitation is readily attenuated by co-application of DA.57 It is therefore likely that VTA/SNc activation releases DA
in the VP which attenuates AMN-induced excitations mediated by glutamate. The mechanism underlying this effect may include presynaptic inhibition, for DA is known to attenuate glutamate release in the striatum, 15A4.45.58.66kKl
The transmitter responsible for the short latency AMN-evoked inhibitions is not known, but indirect evidence suggests that GABA is a reasonable candi- date. For example, though not evaluated for the basolateral nucleus, GABAergic projections have been demonstrated to originate from the intercalated cell mass of the amygdala and project to the sublenticular substantia innominata.6’ Like the intercalated nuclei, the basolateral nucleus contains a large number of perikarya that stain for GABA markers59 and mi- croiontophoresis of GABA induces a rapid and robust inhibition VP neuronal activity,32,38.57 both of which provide for the possibility that an amygdaloid GABA- ergic projection inhibits VP neuronal activity. The observation that DA can modulate AMN-induced VP rate suppressions agrees with biochemical reports on striatal regions where DA suppresses GABA re- lease.3,‘2.77 This also is in accord with the electrophysio- logical demonstration that iontophoretically applied DA attenuates GABA-induced decreases in globus pallidus cell firing.2
In addition to short latency effects, AMN stimu- lation readily evoked long latency excitatory and inhibitory VP responses, with the later being the most frequently observed of all AMN-evoked effects. These responses may involve the NA, for the NA is directly innervated by the AMN,‘8.33~3h,64,6s and the
NA provides both inhibitory and excitatory inputs to the VP (for review see Ref. 8). Yim and Mogensonx5 observed that procaine-induced inactivation of the NA attenuates 54% of the AMN-evoked long latency suppression of VP activity. AMN-evoked long latency inhibitory responses of VP neurons also are attenuated by the VTA stimulation,*5 (and present study) and by D-amphetamine injected within the NA.85 These observations suggest that DA released in the NA can mediate some of the VTA stimulation- induced attenuation of AMN-evoked long latency VP responses. The present study extended these findings to reveal that (1) 79% of the VP neurons exhibiting AMN-evoked long latency responses were modulated monosynaptically by VTA/SNc stimulation, and (2) intra-VP DA application attenuated 80% of the long latency inhibitory, and 38% of the long latency excitatory VP responses evoked by AMN stimu- lation. Thus, the ascending dopaminergic system can govern AMN influences over the VP both at the level of the NABS and within the VP itself (present study).
Functional signljicance of the convergence of ventral tegmental arealsubstantia nigra zona compacta and amygdaloid nuclei influences within the ventral pallidurn
AMN regulate a number of limbic-related func- tions included reward- and emotion-associated be-

1116 R. J. Maslowski-Cobuzzi and T. C. Napier
haviors.6.2’,4y VP responding to AMN stimulation of AMN signals provides a cellular basis for a
conveys limbic influences not only to the VP, but to modulatory role for DA in the brain, and indicates
VP target sites as well. For example, more than that DA may take on the attributes of VP function
two-thirds of VP neurons that are antidromically by altering VP inputs. This modulation, along activated by stimulation of the pedunculopontine with that occurring at the level of the NA,47 provides
nucleus of the mesencephalic locomotor region, DA with a powerful influence over limbic-directed
exhibit AMN-evoked responses.76 Thus, AMN pro- behavior. iections to the VP may provide a limbic bias on VP efferents to brainstem regions associated with
locomotor activity. The behavioral repertoire influenced by forebrain
DA systems is particularly broad in scope (for review see Ref. 14). Neuronal mechanisms responsible for this diversity may involve the ability of the mono- amine to regulate transmitter signals from several brain regions. Le Moal and Simon” argue that the behavioral manifestation of DA transmission reflects the function encompassed by the brain region which DA innervates. This argument is consistent with the idea that DA biases, or determines the gain of,
inputs.26,73 Results from the present work support
these proposals. It is thought that the VP participates in what is
termed “reward-mediated behaviors”; for example, the execution of appropriate motor responses to stimuli associated with a positive reinforcer.35~46~63~“5~79
Many of the components of these complex behaviors also are attributed to DA. The demonstration that DA within the VP can modify the effectiveness
CONCLUSIONS
Analysis of evoked-responding experiments of the present study indicate that VP DA can modify (1) the polysynaptic influences recruited by an activated midbrain dopaminergic system, (2) the polysynaptic influences called into action by stimulating of AMN, and (3) the direct monosynaptically-mediated effects of AMN stimulation. These effects are apparent even at synaptic concentrations of DA that are below those necessary to induce direct effects on neuronal activity. It therefore appears that DA within the VP acts as a significant neuromodulator of limbic signals.
Acknowledgements-The authors thank Drs Sheryl Beck and James Chrobak for their advice and assistance, Drs Robert Cobuzzi and Byron Heidenreich for their helpful comments on the manuscript, and Mr Ronald Price, Jr for his computer expertise. Gratitude also is extended to Scher- ing-Plough, Corp. for the generous gift of SCH23390. This research was supported by USPHS grants MH45180 and DA05255 to TCN.
I.
2.
3.
4.
5.
6. 7.
8.
9.
10.
Il.
12.
13. 14.
15.
16.
17.
REFERENCES
Beckstead R. M., Wooten G. F. and Trugman J. M. (1988) Distribution of D, and D, dopamine receptors in the basal ganglia of the cat determined by quantitative autoradiography. J. camp. Neurol. 268, 131-145. Bergstrom D. A. and Walters J. R. (1984) Dopamine attenuates the effects of GABA on single unit activity in the globus pallidus. Brain Res. 310, 23-33. Brase D. A. (1980) Pre- and postsynaptic striatal dopamine receptors: differential sensitivity to apomorphine inhibition of [‘Hldopamine and [“‘CIGABA release in vitro. J. Pharm. Pharmac. 32, 432433. Brudzynski S. M. and Mogenson G. J. (1985) Association of the mesencephalic locomotor region with locomotor activity induced by injections of amphetamine into the nucleus accumbens. Brain Res. 334, 77-84. Butcher L. L. and Woolf N. J. (1986) Central cholinergic systems: synopsis of anatomy and overview of physiology and pathology. In The Biological Substrates of Alzheimer’s Disease (eds Scheibel A. B., and Wechsler A. F.), pp. 73-86, Academic Press, New York. Carlsen J. (1989) Organization of the basolateral amygdala, Acta Neurol. Sand. 122, Suppl: l-27. Carnes K. M., Fuller T. A. and Price J. L. (1990) Sources of presumptive glutamatergic/aspartatergic afferents to the magnocellular basal forebrain in the rat. J. camp. Neural. 302, 8244852. Chrobak J. J. and Napier T. C. (1993) Opioid and GABA modulation of accumbens-evoked ventral pallidal activity. J. Neural Transm. 93, 123-143. Civelli O., Bunzow J. R. and Grandy D. K. (1993) Molecular diversity of the dopamine receptors. A. Rev. Pharmac. Toxicol. 32, 281-307. Civelli O., Bunzow J. R., Grandy D. K., Zhou Q. Y. and Van To1 H. H. M. (1991) Molecular biology of the dopamine receptors. Eur. J. Pharmac. 207, 277-286. Contreras P. C., Quirion R., Gehlert D. R., Contreras M. L. and O’Donohue T. L. (1987) Autoradiographic distribution of non-dopaminergic binding sites labeled by [‘HI haloperidol in rat brain. Neurosci. Lett. 75, 133-140. de Belleroche J. and Gardiner L. M. (1983) Action of apomorphine, bromocriptine and lergotrile on y-aminobutyric acid and acetylcholine release in nucleus accumbens and corpus striatum. J. Neurol Transm. 58, 1533168. Duggan A. W. (1983) Microiontophoresis and the central nervous system. J. Electrophysiol. Tech. 9, loll1 12. Dunnett S. B. and Robbins T. W. (1992) The functional role of mesotelencephalic dopamine systems. Eiol. Rev. 67, 491-518. Earle M. L. and Davies J. A. (1991) The effect of methamphetamine on the release of glutamate from striatal slices. J. Neural Transm. 86, 217-222. Fallon J. H., Koziell D. A. and Moore R. Y. (1978) Catecholamine innervation of the basal forebrain. II. Amygdala, suprarhinal cortex and entorhinal cortex. J. camp. Neurol. 180, 509-532. Fallon J. H. and Moore R. Y. (1978) Catecholamine innervation of the forebrain. IV. Topography of the dopamine projection to the basal forebrain and neostriatum. J. camp. Neurol. 180, 5455580.

Dopamine attenuates amygdala-evoked ventral pallidal activity 1117
18. Fuller T. A., Russchen F. T. and Price J. L. (1987) Sources of presumptive glutamergic/aspartergic afferents to the rat ventral striatopallidal region. J. camp. Neural. 258, 317-338.
20. Geula C. and Slevin J. T. (1989) Substantia nigra 6-hydroxydopamine lesions alter dopaminergic synaptic markers in the nucleus basalis magnocellularis and striatum of rats. Synapse 4, 248-253.
21. Gloor P. (1978) Inputs and outputs of the amygdala: what the amygdala is trying to tell the rest of the brain. In Limbic Mechanisms: The Continuing Evolution of the Limbic System Concept (eds Livingston K. and Hornykiewicz 0.) pp. 189-209. Plenum Press, New York.
22. Gonon F. G. (1988) Nonlinear relationship between impulse flow and dopamine released by rat midbrain dopaminergic neurons as studied by in vivo electrochemistry. Neuroscience 24, 19-28.
23. Gonon F. G. and Buda M. (1985) Regulation of dopamine release by impulse flow and by autoreceptors as studied by in vivo voltammetry in the rat striatum. Neuroscience 14, 7655774.
24. Grace A. A. and Bunney B. S. (1984) The control of firing pattern in nigral dopamine neurons: single spike firing. J. Neurosci. 4, 2866-2876.
25. Grace A. A. and Bunney B. S. (1984) The control of firing pattern in nigral dopamine neurons: burst firing. J. Neurosci. 4, 2877-2890.
26. Graybiel A. M. (1990) Neurotransmitters and neuromodulators in the basal ganglia. Trends Neurosci. 13, 244-254. 27. Guyenet P. G. and Aghajanian G. K. (1978) Antidromic identification of dopaminergic and other output neurons of
the rat substantia nigra. Brain Res. 150, 69-84. 28. Haring J. H. and Wang R. Y. (1986) The identification of some sources of afferent input to the rat nucleus basalis
magnocellularis by retrograde transport of horseradish peroxidase. Bruin Res. 266, 152-158. 29. Heimer L. and Alheid G. F. (1991) Piecing together the puzzle of basal forebrain anatomy. In The Basal Forebrain:
Anaromy to Function (eds Napier T. C., Kalivas P. W. and Hanin I.), Vol. 295, pp. l-42. Plenum Press, New York. 30. Heimer L. and Wilson R. D. (1975) The subcortical projections of allocortex: similarities in the neural associations
of the hippocampus, the piriform cortex and the neocortex. In Golgi Centennial Symposium Proceedings (ed. Santini M.), pp. 177-193. Raven Press, New York.
31. Jones B. E. and Cue110 A. C. (1989) Afferents to the basal forebrain cholinergic cell area from pontomesencephalic, catecholamine, serotonin, and acetylcholine neurons. Neuroscience 31, 3761.
32. Jones D. L. and Mogenson G. J. (1980) Nucleus accumbens to globus pallidus GABA projection: electrophysiological and iontophoretic investigations. Brain Res. 188, 93-105.
33. Kelley A. E., Domesick V. B. and Nauta W. J. H. (1982) The amygdalostriatal projection in the rat-an anatomical study by a terograde and retrograde tracing methods. Neuroscience 7, 615630.
34. Klitenick A., Deutch A. Y., Churchill L. and Kalivas P. W. (1992) Topography and functional role of dopaminergic 4 projections from the ventral mesencephalic tegmentum to the ventral pallidum. Neuroscience 50, 371-386.
35. Koob G. F., Swerlow N. R., Vaccarino F., Hubner C., Pulvirenti L. and Weiss F. (1991) Functional output of the basal forebrain. In The Basul Forebrain: Anaromy to Function (eds Napier T. C., Kalivas P. W. and Hanin I.), Vol. 295, pp. 291-305. Plenum Press, New York.
36. Krettek J. E. and Price J. L. (1978) Amygdaloid projections to subcortical structures within the basal forebrain and brainstem in the rat and cat. J. camp. Neural. 178, 225-254.
37. Kuffler S. W., Nicolls J. G. and Martin A. R. (1984) Release of chemical transmitters. In From Neuron to Bruin: A Cellular Approach to the Function of the Nervous System (eds Kuffler S. W., Nicolls J. G. and Martin A. R.), pp. 241-261. Sinauer Assoc. Inc., Sunderland, U.K.
38. Lamour Y., Dutar P., Rascal 0. and Jobert A. (1986) Basal forebrain neurons projecting to the rat frontoparietal cortex: electrophysiological and pharmacological properties. Brain Res. 362, 1222131.
39. Le Moal M. and Simon H. (1991) Mesocorticolimbic dopaminergic network: functional and regulatory roles. Physiol. Rev. 71, 155-234.
40. Lipiski J. (1991) Antidromic activation of neurons as an analytic tool in the study of the central nervous system. J. Neurosci. Meth. 4, l-32.
41. Manley L. D., Kuczenski R., Segal D. S., Young S. J. and Groves P. M. (1992) Effects of frequency and pattern of medial forebrain bundle stimulation on caudate dialysate dopamine and serotonin. J. Neurochem. 58, 1491-1498.
42. Maslowski R. J. and Napier T. C. (1991) Dopamine D, and D, receptor agonists induce opposite changes in the firing rate of ventral pallidal neurons. Eur. J. Pharmac. 200, 103-l 12.
43. Maslowski R. J. and Napier T. C. (1991) Effects of D, and D, antagonists on apomorphine-induced responses of ventral pallidal neurons. Neuroreport 2, 451454.
44. Maura G., Giardi A. and Raiteri M. (1988) Release-regulating D-2 dopamine receptors are located on striatal glutamatergic nerve terminals. J. Pharmac. exp. Ther. 247, 680-684.
45. Mitchell P. R. and Doggett N. S. (1980) Modulation of striatal [rH]glutamic acid release by dopaminergic drugs. Life Sci. 26, 2073-2081.
46. Mogenson G. J. and Yang C. R. (1991) The contribution of basal forebrain to limbic-motor integration and the mediation of motivation to action. In The Basal Forebrain: Anatomy CO Function (eds Napier T. C., Kalivas P. W. and Hanin I.), pp. 267-290. Plenum Press, New York.
47. Mogenson G. J., Yang C. R. and Yim C. Y. (1988) Influence of dopamine on limbic inputs to the nucleus accumbens. Ann. N. Y. Acad. Sci. 537, 86100.
48. Mogenson G. J. M., Swanson L. W. and Wu M. (1983) Neural projections from nucleus accumbens to globus pallidus, substantia innominata, and lateral preoptic-lateral hypothalamic area: an anatomical and electrophysiological investigation in the rat. J. Neurosci. 3, 189-202.
49. Nakano Y., Lenard L., Oomura Y., Nishino H., Aou S. and Yamamoto T. (1987) Functional involvement of catecholamines in reward-related neuronal activity of the monkey amygdala. J. Neurophysiol. 57, 72-91.
50. Napier T. C. (1992) Contribution of the amygdala and nucleus accumbens to ventral pallidal responses to dopamine agonists. Synapse 10, 110-l 19.
51. Napier T. C. and Chrobak J. J. (1992) Evaluations of ventral pallidal dopamine receptor activation in behaving rats. Neuroreport 3, 609-611.
52. Napier T. C. and Maslowski-Cobuzzi (1994) Electrophysiological verification for the presence of Dl and D2 dopamine receptors within the ventral pallidurn. Synapse 17, 160-166.

1118 R. J. Maslowski-Cobuzzi and T. C. Napier
53.
54.
55.
56.
51.
58.
59.
60.
61.
62. 63.
64. 65.
66.
61.
68.
70.
71.
12.
73.
74.
75.
76.
77.
78.
79.
80.
81.
82.
83.
84.
85.
86.
87.
Napier T. C., Muench M. B., Maslowski R. J. and Battaglia G. (1991) Is dopamine a neurotransmitter within the ventral pallidum/substantia innominata? In The Basal Forebrain: Anatomy to Funclion (eds Napier T. C., Kalivas P. W. and Hanin I.), Vol. 295, pp. 1833195. Plenum Press, New York. Napier T. C., Pirch J. H. and Strahlendorf H. K. (1983) Naloxone antagonizes striatally-induced suppression of globus pallidus unit activity. Neuroscience 9, 53-59. Napier T. C. and Potter P. E. (1989) Dopamine in the rat ventral pallidum/substantia innominata: biochemical and electrophysiological studies. Neuropharmacology 28, 757-760. Napier T. C. and Rehman F. (1992) Motoric analysis of dopamine receptor subtype activation within the ventral pallidum and dorsal globus pallidus. Sot. Neurosci. Absfr. 18, 994. Napier T. C., Simson P. E. and Givens B. S. (1991) Dopamine electrophysiology of ventral pallidal/substantia innominata neurons: comparison with the dorsal globus pallidus. J. Pharmac. exp. Ther. 258, 249-262. Nieoullon A., Kerkerian L. and Dusticier N. (1983) Presynaptic dopaminergic control of high affinity glutamate uptake in the striatum. Neurosci. LRII. 43, 19lll96. Nitecka L. and Frotscher M. (1989) Organization and synaptic interconnections of GABAergic and cholinergic elements in the rat amygdaloid nuclei: single- and double-immunolabeling studies. J. camp. Neurol. 279, 470-488. Olton D., Markowska A., Voytko M. L., Givens B., Gorman L. and Wenk G. (1991) Basal forebrain cholinergic system: a functional analysis. In The Basal Forebrain: Anatomy IO Funcrion (eds Napier T. C., Kalivas P. W. and Hanin I.), Vol. 295, pp. 3533372. Plenum Press, New York. Pare D., Smith Y. and Pare J.-F. (1993) GABAergic projections from the intercalated cell masses (ICMs) to the centromedial (CM) amygdaloid complex and basal forebrain (BF). Sot. Neurosci. Abstr. 19, 1440. Paxinos G. and Watson C. (1986) The Rat Brain in Stereotaxic Coordinu/es. Academic Press, Orlando, FL, U.S.A. Pirch J.. Rigdon G., Rucker H. and Turco K. (1991) Basal forebrain modulation of cortical cell activitv during conditioning: In The Basal Forebruin: Anatomy to Fur&on (eds Napier T. C.. Kalivas P. W. and Hanin I.), Vol. 295: pp. 219-232. Plenum Press, New York. Price J. L. (1986) Subcortical projections from the amygdaloid complex. Adv. Exp. Med. Biol. 203, 19-~33. Richardson R. T. and DeLong M. R. (1991) Electrophysiological studies of the functions of the nucleus basalis in primates. In The Busul Forebrain: Anafomy /o Funclion (eds Napier T. C., Kalivas P. W. and Hanin I.), Vol. 295. pp. 2333252. Plenum Press, New York. Rowlands G. J. and Roberts P. J. (1980) Activation of dopamine receptors inhibits calcium-dependent glutamate release from cortico-striatal terminals in oitro. Eur. J. Pharmac. 62, 241-242. Russchen F. T., Amaral D. G. and Price J. L. (1985) The afferent connections of the substantia innominata in the monkey, Macaca fascicularis. J. camp. Neural. 242, l--21. Russchen F. T. and Price J. L. (1984) Amygdalostriatal projections in the rat. Topographical organization and fiber morphology shown using the lectin PHA-L as an anterogrdde tracer. Neurosci. Let/. 47, 15-22. Salmoiraghi G. C. and Weight F. (1967) Micromethods in neuropharmacology: an approach to the study of anesthetics. Anesthesiology 28, 5444. Saper C. B. (1984) Organization of cerebral cortical afferent systems in the rat. I. Magnocellular basal nucleus. J. romp. Neural. 222, 3 13-342. Semba K., Reiner P. B., McGeer E. G. and Fibiger H. C. (1988) Brainstem afferents to the magnocellular basal forebrain studied by axonal transport, immunohistochemistry, and electrophysiology in the rat. J. camp. Neural. 267, 433453. Servan-Schreiber D., Printz H. and Cohen J. D. (1990) A network model of catecholamine effects: gain, signal-to-noise ratio, and behavior. Science 249, 892.-895. Swanson L. W., Mogenson G. J., Gerfen C. R. and Robinson P. (1984) Evidence for a projection from the lateral preoptic area and substantia innominata to the “mesencephalic locomotor region” in the rat. Brain Res. 295, 161 178. Swerdlow N. R. and Koob G. F. (1987) Lesions of the dorsomedial nucleus of the thalamus, medial prefrontal cortex and pedunculopontine nucleus: effects on locomotor activity mediated by nucleus accumbens-ventral pallidal circuitry. Brain Res. 412, 233.-243. Tsai C. T., Mogenson G. J., Wu M. and Yang C. R. (1989) A comparison of the effects of electrical stimulation of the amygdala and hippocampus on subpallidal output neurons to the pedunculopontine nucleus. Brain Res. 494,22-29. van der Heyden J. A. M.. Venema K. and Korf J. (1980) In rioo release of endogenous GABA from rat striatum: inhibition by dopamine. J. Neurochem. 34, 1338-1341. Werman R. (1966) A review--criteria for identification of a central nervous system transmitter. Comp. Biochem. Phpsio/. 18, 7455766. Wilson F. A. W. (1991) The relationship between learning, memory and neuronal responses in the primate basal forebrain. In The Basal Forebrain: Anatomy fo Function (eds Napier T. C., Kalivas P. W. and Hanin I.), Vol. 295, pp. 253-266. Plenum Press, New York. Yamamoto B. K. and Davy S. (1992) Dopdminergic modulation of glutamate release in striatum as measured by microdialysis. J. Neurochem. 58, 17361742. Yang C. R. and Mogenson G. J. (1985) An electrophysiological study of the neural projections from the hippocampus to the ventral pallidum and the subpallidal areas by way of the nucleus accumbens. Neuroscience 15, 1015~1024. Yang C. R. and Mogenson G. J. (1989) Ventral pallidal responses to dopamine receptor stimulation in the nucleus accumbens. Bruin Res. 489, 237-246. Yim C. Y. and Mogenson G. J. (1980) Electrophysiological studies of neurons in the ventral tegmental area of Tsai. Brain Res. 181, 301.-313. Yim C. Y. and Mogenson G. J. (1982) Response of nucleus accumbens neurons to amygdala stimulation and its modification by dopamine. Bruin Re.r. 239, 401-415. Yim C. Y. and Mogenson G. J. (1983) Response of ventral pallidal neurons to amygdala stimulation and its modulation by dopamine projections to nucleus accumbens. J. Neurophysiol. 50, 148-161. Young W. S., Alheid G. F. and Heimer L. (1984) The ventral pallidal projection to the mediodorsal thalamus; a study with fluorescent retrograde tracers and immunohistofluorescence. J. Neurosci. 4, 16261638. Zaborszky L. (1989) kfferent connections of the forebrain cholinergic projection neurons. with special reference to monoaminergic and peptidergic fibers. In Central Cholinergic Synaptic Transmission (eds Frotscher M. and Misgeld U.). pp. 12-32. Birkhauser, Base].

Dopamine attenuates amygdala-evoked ventral pallidal activity 1119
88. Zaborszky L., Cullinan W. E. and Braun A. (1991) Afferents to basal forebrain choIinergic projection neurons: an update. In The Basal Forebrain: Anaromy IO Function (eds Napier T. C., Kalivas P. W. and Hanin I.), Vol. 295, pp. 43-100. Plenum Press, New York.
89. Zaborszky L., Leranth C. S. and Heimer L. (1984) Ultrastructural evidence of amygdalofugal axons terminating on cholinergic cells of the rostra1 forebrain. Neurosci. Lett. 52, 219-225.
90. Zahm D. S. (1989) The ventral striatopallidal parts of the basal ganglia in the rat-II. Compartmentation of ventral pallidal efferents. Neuroscience 30, 33-50.
91. Zahm D. S. and Heimer L. (1988) Ventral striatopallidal parts of the basal ganglia in the rat: I. Neurochemical compartmentation as reflected by the distributions of neurotensin and substance P immunoreactivity. J. camp. Neural. 272. 516-535.
(Accepted 22 February 1994)





























