Embed Size (px)
Citation preview

Adenosine-induced neuroprotection: involvement of glia cells and cytokines

Financiële ondersteuning:
School of Behavioral and Cognitive Neurosciences
Rijksuniversiteit Groningen
Lay-out: Dick Visser (Rijksuniversiteit Groningen)Ontwerp omslag: Saskia van der Schaaf
Printed by: Van Denderen b.v., Groningen
ISBN: 90-367-2132-6

RIJKSUNIVERSITEIT GRONINGEN
Adenosine-induced neuroprotection: involvement of glia cells and cytokines
PROEFSCHRIFT
ter verkrijging van het doctoraat in de Medische Wetenschappen
aan de Rijksuniversiteit Groningenop gezag van de
Rector Magnificus, Dr. F. Zwarts,in het openbaar te verdedigen op
woensdag 29 september 2004om 16:15 uur
door
Maria Catharina Wittendorp
geboren op 20 april 1977te Emmen

Promotor: prof. dr. H.W.G.M. Boddeke
Copromotor: dr. K. Biber
Beoordelingscommissie: prof. dr. D. van Calkerprof. dr. A.P. IJzermanprof. dr. J. Zaagsma

Contents
List of Abbreviations 6
Chapter 1 Introduction 7
Chapter 2 The role of glia cells in purine receptor-induced neuroprotection 39
Chapter 3 Adenosine A3 receptor-induced CCL2 synthesis in cultured 57mouse astrocytes
Chapter 4 The mouse brain adenosine A1 receptor: functional expression 75and pharmacology
Chapter 5 Interleukin-6 is involved in upregulation of adenosine A1 89receptor expression after seizures
Chapter 6 Summary and discussion 107
Nederlandse samenvatting 115
Dankwoord 119

6
List of Abbreviations
ATP adenosine triphosphateADP adenosine diphosphateAMP adenosine monophosphatebFGF basic fibroblast growth factorBDNF brain-derived neurotrophic factor cAMP cyclic adenosine monophosphateCCL2 CC chemokine ligand 2CHA cyclohexyladenosineCHO Chinese Hamster Ovary (cells)CNS central nervous systemCNTF ciliary neurotrophic factorCT-1 cardiotrophin-1IL interleukinEDTA ethylene diaminotetra acetic acidGDNF glial-derived neurotrophic factorGFAP glial fibrillary acidic proteinG-CSF granulocyte colony-stimulating factorGTP guanine triphosphateJAK Janus kinaseLIF leukemia inhibiting factorMAPK mitogen activated protein kinaseMCP monocyte chemoattractant proteinmRNA messenger ribonucleic acidNGF nerve growth factorNMDA N-methyl-D-aspartateNFκB nuclear factor kappa BNT neurotrophinOSM oncostatin MPTX pertussis toxinPTX3 pentraxin 3PTZ pentylenetetrazoleRANTES regulated on activation, normal T-cell expressed and secreted TGF-β transforming growth factor betaTM7 7 trans-membrane spanning receptors TNF-α tumor necrosis factor alphaTrk tyrosine kinaseUTP uridine triphosphateUDP uridine diphosphateVEGF vascular endothelial growth factor

7
Introduction
Chapter 1

The adenosinergic system
General aspects
Adenosine and ATP belong to a group of molecules, called purines, which havesimilar molecular structure. Purines are important constituents of living cells, forexample adenine and guanine are basic components of nucleic acids. ATP is the universal “currency” of free energy in the cell, acting as a energy donorin most cellular activities. Besides their role in energy transfer, purines alsofunction as important intercellular signaling molecules [65]. When cells useenergy, ATP is hydrolyzed into ADP, AMP and finally into adenosine. Underphysiological conditions the production and consumption of energy are balancedand the amount of intracellular adenosine is tightly regulated. Since underphysiological conditions ATP concentrations in the cell are high (about 3 mM),adenosine concentrations rise sharply if a small amount of ATP is metabolized.Thus, in situations where cells are impaired in their ability to synthesize ATP, thelevels of intracellular adenosine will therefore increase rapidly. Adenosine is transported passively across the cell membrane by facilitateddiffusion transporters, which equilibrate the concentration of extra- andintracellular adenosine. Rising intracellular levels of adenosine will thus lead tothe release of adenosine, Under basal conditions the concentration of extra-cellular adenosine in all biological fluids is estimated at 30-300 nM, but as aresult of decreased energy supply or increased metabolic activity the concen-tration can rise to 10 µM or higher [86, 178, 189]. Another source of adenosine isthe extracellular breakdown of ATP by ecto-nucleotidases, but this has beensuggested to be a minor contributor to the amount of extracellular adenosine[65].The two main mechanisms responsible for the clearing of adenosine from theextracellular space are transformation into inosine by adenosine deaminase or byreuptake into cells, which occurs by facilitated diffusion or by active transport[43, 65]. The breakdown rate of adenosine in the extracellular space is very high due to thehigh expression of the appropriate enzymes. Thus the extracellular concen-trations of adenosine can rise and decrease rapidly, which makes adenosine anideal signaling molecule. Figure 1.1 describes pathways of adenosine release,production and degradation and the enzymes involved.
Purinoreceptors
Purinoreceptors have been subdivided into P1 receptors, which bind adenosine asnatural ligand and P2 receptors which can bind ATP, ADP, adenine dinucleotidesbut also pyrimidines like UTP and UDP [32]. P1 or adenosine receptors have
Chapter 1
8

initially been divided in two subtypes. This classification between A1 and A2 wasbased on their effect on cAMP [219]. Currently, the adenosine receptor familycontains the four subtypes, A1, A2A, A2B and A3, which all couple to G-proteinsand have the typical seven-transmembrane structure as shown in figure 1.2 [85].Originally it was reported that adenosine A1 and A3 interact primarily with Gi-proteins and induce inhibition of adenylyl cyclase whereas adenosine A2A and A2B
receptors couple preferentially to Gs-proteins and thus stimulate adenylyl cyclaseand increase cAMP levels [86, 152, 218]. Adenosine receptors, however, have alsobeen reported to interact with other G-proteins and signal through various otherpathways, independent of adenylyl cyclase, as reviewed recently [189]. In table1.1 an overview of the second messenger pathways induced by adenosinereceptors has been provided. Several reviews addressing the structure,
Introduction
9
adenylylcyclase
ecto-5'-nucleotidase
cAMP
creatinekinase
adenylatekinase
ATP
ADP
5'-AMP
Adenosine
nucleotidase
cytosolic5'-nucleotidase
phospodiesterase
ATPase
adenosinekinase
nucleotidase
ecto-ATPase
ATP
ADP
5'-AMP
Adenosine
release
fac. transport
ado. reuptake
adenosinedeaminase
Inosineextracellularintracellular
Figure 1.1. Main pathways for the formation of adenosine. ATP is being released from damagedcells, where it can be metabolized into adenosine by ectonucleotidases. Intracellularly, ATP canalso be metabolized into adenosine, which is then being released by facilitated transport.Adenosine can be cleared from the extracellular space by reuptake into the cell or by degradationinto inosine. (Derived from refs 65, 237).

classification and pharmacology of adenosine receptors have been published [2,86, 152, 153, 157, 201].P2 receptors are divided in a family of ligand gated ion channels, P2X receptorsand G-protein coupled receptors termed P2Y receptors. Several subtypes of bothfamilies have been cloned and characterized [170].
Physiological functions of adenosine in the periphery
In situations of increased energy use or a decreased energy supply the consumptionof ATP overrides its’generation. As a result the balance is shifted towards higherlevels of adenosine, which is rapidly released from the cell. The resultingincreased extracellular levels of adenosine and the subsequent stimulation of cell-surface adenosine receptors will generally result in an inhibition of the cellmetabolism (Figure 1.3). Thus adenosine-based phosphate metabolism provides avery basic feedback system linking energy demand to energy supply, whichcontrols the metabolic rate in order to prevent energy depletion and subsequentcellular damage [58, 65, 98, 157]. Already since 1929, when adenosine was described to be involved in cardiovascu-lar regulation [61], extensive research on the many physiological functions ofadenosine has been performed [22, 63, 111, 202, 230].
Chapter 1
10
Agonist & antagonistrecognition site
extracellular
intracellular
76
4
5
3 21
COOH
NH2
Figure 1.2. Structure of the adenosine A1 receptor. As other G-protein coupled receptors, theadenosine A1 receptor has 7 transmembrane domains (1-7) which have an α-helix structure. Themost important region for agonist and antagonist binding is indicated. (Adapted from ref 170).

In keeping with the already mentioned role of adenosine, coupling energyconsumption to energy demand, adenosine receptors are distributed in almost allbiological tissues and in many different species [147, 172, 173]. Althoughadenosine may have different actions depending on cell type, the ultimate resultof the action is the control of metabolic rate. Thus, adenosine has several actions,which directly modulate energy supply. Adenosine induces relaxation of vascularsmooth muscle cells causing vasodilatation, thereby increasing blood flow. In thekidney, adenosine causes vasoconstriction thus reducing renal blood flow andindirectly regulating blood pressure [100, 231]. Adenosine plays a role in themodulation of cardiac and respiratory function as well [144, 198, 208]. Forexample, adenosine is involved in hypoxia-induced angiogenesis, therebycounteracting the effects of a reduced energy supply [135]. In addition, adenosine
Introduction
11
Table 1.1. Overview of G-protein coupling and second messenger signaling of adenosinereceptors. (Adapted from refs 86, 189).
adenosine G-protein effects of MAPK signaling pathway receptor G-protein coupling subtypesubtype
A1 Gi1/2/3 cAMP ERK1/2 Gi/0 > βγ > Tyr kinasea >IP3/DAG (PLC) P13K> MEK1Arachidonate (PLA2)choline, DAG (PLD)K+ channelsQ,P, N type Ca2+ channels
G0
A2A Gs cAMP ERK1/2 Gs > cAMP > PKA > Rap1b
Golf cAMP > B-Raf > MEK1G15/16 IP3 ERK1/2 Gαs > cAMP > PKA > Src >
Ras
A2B Gs cAMP ERK1/2 Gs > cAMP > P13K> MEK1Gq/11 IP3/DAG (PLC) p38 Gs > cAMP > PKA
A3 Gi2/3 cAMP ERK1/2 Gi/0 > βγ > P13K> Ras >IP3/DAG (PLC) MEK1choline, DAG (PLD)K+ -ATP channelsCl- channels
Gq/11 IP3/DAG (PLC)
←←
←
←←←
←←
←←←←
←←
←←
←

Chapter 1
12
ATP
adenosine(extracellular)
adenosine
adenosine receptorstimulation
use ofenergy
AR
Figure 1.3. The adenosinergic system forms a negative feedback loop to regulate cell metabolism.Adenosine levels rise in cases of excessive energy use. Adenosine is transported out of the cell,binding to adenosine receptors on the surface. Activation of adenosine receptors slows downenergy usage, which will eventually result in reduced formation of adenosine.
Table 1.2. Overview of physiological systems/pathological conditions in which adenosine plays arole.
physiological system/pathology references
central nervous system [65, 175, 202]sleep [17, 168, 169]circadian rhythm [69]anxiety [82, 110]drugs of abuse induced actions [64, 203]pain modulation [110, 166, 184, 185]cardiac system [14, 144, 154, 208] blood flow [58, 164, 208]angiogenesis [62, 94, 135, 136]platelet aggregation [126, 183]respiratory system [95, 198]mast cell degranulation, asthma [83, 125]immune system [47, 126]kidney [42, 100]gastrointestinal tract [177]lipolysis [103, 186, 212]cell growth, proliferation [33]apoptosis [4, 151]embryogenesis [119]

is involved in several other physiological functions, which are not directly linkedto energy control. Adenosine is known to mediate anti-inflammatory effects,which could protect tissues from damage [77, 126, 204]. Furthermore, adenosineis involved in platelet aggregation, gastrointestinal mobility, mast celldegranulation, pain modulation, induction of sleep, cell growth, proliferation andapoptosis. In table 1.2 an overview of all the different functions in whichadenosine plays a role has been provided.
Physiological functions of adenosine in the nervous system
Generally the brain consumes approximately 20% of our total energy.Consequently, the brain is very vulnerable to fluctuations in energy supply. Inthis respect adenosine plays an important role by coupling energy use to energydemand. Large amounts of adenosine are produced and released duringconditions of increased energy use such as high neuronal activity during seizures,or under conditions of reduced energy supply like in ischemia or hypoglycemia.[180]. Stimulation of cell surface adenosine receptors in the brain, that are mainlyof the A1 subtype, protects neurons by retaining neuronal firing and inhibitingthe release of excitatory neurotransmitters, including glutamate [202]. Theseactions are primarily described for neurons in the brain but adenosine seems toexert similar effects in the spinal cord [56].Whereas under pathological conditions adenosine is neuroprotective, underphysiological conditions adenosine acts as a neuromodulator by regulating ageneral inhibitory tone in the brain. This neuromodulatory role at the synapselevel is mediated by stimulation of inhibitory A1 receptors and facilitatory A2A
receptors [48, 231].Besides neuromodulatory and neuroprotective effects, adenosine also inducestrophic effects in neurons and glia cells. Adenosine stimulates neurite outgrowth[36], increases glia cell proliferation and promotes myelination [200]. Theseactions are more extensively reviewed in chapter 2.
Adenosine is also involved in other functions in the central nervous system. Forinstance adenosine is known to induce sleep [168], which explains the activatingproperties of the unspecific adenosine antagonist caffeine [87]. There is someevidence that adenosine may play a role in the effects of drug abuse, since opiates,benzodiazepines as well as ethanol inhibit adenosine reuptake [65]. Sinceadenosine is known to have analgesic effects [66, 110], it has been suggested thatanalgesic effects of for example morphine are actually caused by its effect onadenosine [63, 64, 203].
Introduction
13

Functions of different adenosine receptors in the nervous system
Although adenosine receptors are found throughout the brain, their expressionvaries in specific brain regions [55, 175] (Figure 1.4). The specific expressionpattern of the different receptor subtypes is related to their specific functions. Thefunctions of adenosine receptors have been analyzed by using selective adenosinereceptor antagonists and by the generation of mouse strains with targeteddeletions of adenosine A1, A2A and A3 receptor subtypes [110, 124, 149, 235].
Adenosine A1 receptorsAdenosine A1 receptors are clearly involved in neuroprotection. These receptorsare found throughout the brain, but show especially high expression in vulnerableareas like for example the hippocampus. Adenosine A1 receptors are found bothpre- and postsynaptically in neurons, where they play an important role ininhibiting the release of excitatory neurotransmitters and inducing hyper-polarization respectively. Thus presynaptically, adenosine inhibits the release ofthe excitotoxic neurotransmitters glutamate, probably through the inhibition ofCa2+-influx [63, 65, 70]. Postsynaptically, adenosine counteracts depolarizationby stabilization of the Mg2+ blockade of NMDA receptors [178, 181]. Adenosinealso actively reduces postsynaptic Ca2+-influx, probably by an inhibition of N-type voltage-dependent Ca2+-channels [60, 129, 148, 159, 195, 210]. Moreover,stimulation of adenosine A1 receptors causes hyperpolarization of the postsynapticresting membrane potential via G-protein-dependent activation of inwardly
Chapter 1
14
amygdala
A1olfactory
bulb
striatum
cortex
substantianigra
nucleustractus
solitarius
spinal cord(dorsal horn)
thalamushippocampus
cerebellum
A2AA1
A2A
A1A2A
A2AA1
A1A1A2A A1
A1
A2A
A1
A1
Figure 1.4. Distribution of adenosine A1 and A2A receptors in the brain. Bigger fonts indicatehigh levels of expression. (Adapted from ref 175).

rectifying K+-channels (GIRKs) [160, 214]. In addition, adenosine A1 receptorstimulation enhances a calcium-dependent potassium current, in much the sameway as GABAB receptor stimulation, although different G-proteins might beinvolved [90, 98].
Adenosine A2A receptorsIn general adenosine A2A receptors are involved in the facilitation of neuronalfiring. In close interaction with adenosine A1 receptors they modulate synapsefunction. Although A2A receptors have been found in all brain regions they areparticularly expressed in the nucleus accumbens, olfactory tubercle and striatum,where they are co-localized with dopamine D2 receptors [88, 122, 143, 155, 170].It has been shown that antagonistic interactions between adenosine A2A anddopamine D2 receptors and also between adenosine A1 and dopamine D1
receptors are partly responsible for the motor stimulant effects of adenosinereceptor antagonists like caffeine [88]. Furthermore, involvement of adenosineA2A receptors in locomotion, anxiety, aggression, motivation and reward in drugaddiction and psychotic-like behavior have been suggested [37, 39,124,143].Most of these functions have been revealed using adenosine A2A receptor knockout mice. [37, 38, 52, 143].Adenosine A2A receptors are also involved in the control of cerebral blood flow[58, 170].
Adenosine A2B receptorsAdenosine A2B receptors have been found in most tissues but are generallyexpressed at low levels. Low expression levels were also found throughout thebrain [59]. Since selective ligands for the adenosine A2B subtype are lacking, andno A2B knock mouse has been generated yet, less is known on its physiologicalrole. It has been suggested that A2B receptors play a role in vascularization andcontrol of cerebral blood flow [93, 170, 193]. Stimulation of A2B receptors hasbeen shown to induce release of vascular endothelial growth factor in bothperipheral and cerebral endothelial cells [73, 78, 93, 94]. There are indicationsthat A2B receptors are involved in neuroexcitatory actions and that stimulation ofthese receptors would aggravate tissue injury [72]. A2B receptors are alsoexpressed in glia cells where they have been shown to induce release ofinterleukin-6 [76, 191].
Adenosine A3 receptorsAdenosine A3 receptors are widely distributed in the brain, but its physiologicalrole is largely unknown [170]. Adenosine A3 receptors mediate inhibition ofsynaptic transmission in neurons in concert with A1 receptors [31] and they playa role in modulating synaptic plasticity [46]. More extensive research on the role
Introduction
15

of the A3 receptor in the brain has been done by the group of Von Lubitz [224,226-228]. Von Lubitz and colleagues showed that stimulation of A3 receptorsinduces apoptosis of brain tissue and they therefore suggested that the A3
receptor acts as a “death receptor”. Inducing apoptosis of badly damaged neuronsin stroke would be beneficial since it would limit neuroinflammation and infarctsize [228]. In contrast, stimulation of the A3 receptor with low concentrations of adenosineseems to induce neuroprotective effects [3, 4, 71, 106]. These seeminglyconflicting actions do have physiological significance. High levels of adenosine inthe core area of an ischemic insult would induce apoptosis through action of A3
receptors, while in the surrounding brain tissue lower adenosine levels exertneuroprotective effects mediated by the same receptors. Furthermore, severalreports indicate that adenosine A3 receptor stimulation in glia cells leads toneuroprotection by inducing cytoskeleton rearrangement [1, 5].
The four different adenosine receptor subtypes have a different affinity foradenosine. Whereas A1 and A2A receptors have relatively high (nanomolar range)affinities for adenosine, A2B and A3 receptors have a much lower affinity and areonly activated at micromolar concentrations [65]. These differences in affinitymay reflect functional significance. Thus different receptors with differentfunctional responses are activated by varying extracellular concentrations ofadenosine. Moreover, adenosine at varying concentrations not only activatesdifferent receptor subtypes, but also induces multiple, sometimes even oppositeeffects by activation of the same receptor subtype. These observations show thatthe adenosinergic system regulates a complex interplay of biological activities.
Adenosine in pathology and therapy
Adenosine-based treatment of disorders
Since adenosine is involved in many physiological functions, drugs that interactwith the adenosinergic system (so called “adenosine-based drugs”) could bedeveloped to treat a variety of pathological conditions. But at the same time theseadenosine-based drugs cause serious side effects because adenosine receptors areso widely distributed. This explains why presently only very few adenosine-baseddrugs are used in the clinic, even though extensive research on the physiologicalroles of adenosine has been done since 1929 [61].Currently, adenosine is only therapeutically used as intravenous application totreat patients suffering from supraventricular tachycardias (see [176] andwww.adenocard.com). Adenosine-based treatment of other disorders is still at anearly stage of investigation.
Chapter 1
16

High levels of extracellular adenosine have been associated with thepathophysiology of lung diseases [24]. Adenosine A2B and A3 receptors play arole in adenosine-induced mast cell degranulation and bronchoconstriction andhave therefore been associated with the pathophysiology of asthma [74, 171, 185,236]. In order to block adenosine-induced bronchoconstriction asthmatic patientsuse theophylline, a non-specific adenosine antagonist [72]. High doses oftheophylline, however, can lead to seizure activity, so more specific adenosinereceptor antagonists, which show fewer side effects are preferable. Since specificantagonists for A2B receptors are not available, currently only adenosine A3
antagonists are under investigation as possible anti-asthmatic drugs [75, 83,197].It has been reported that adenosine and adenosine analogues induce apoptosis invarious types of tumor cells [13, 35, 115, 139, 188]. Furthermore, it has beenreported that particularly adenosine A3 receptors are beneficial in the treatmentof cancer [79, 140, 151]. In addition to inducing apoptosis in tumor cells,stimulation of adenosine A3 receptors protects tissue from damage by chemo-therapy and induces the release of granulocyte colony-stimulating factor (G-CSF)which stimulates the proliferation of bone marrow cells [16, 80]. Currently, 2-chlorodeoxyadenosine is tested in clinical trials for the treatment of glioma (seewww.clinicaltrials.gov). Adenosine is also known to mediate anti-inflammatory effects, like suppressionof phagocytosis and reduction of free radical generation, which could protecttissues from damage [77, 101, 126, 204].
Adenosine and the treatment of neurological diseases
It is well established that adenosine induces neuroprotective activity in the brain[53, 111, 178, 202]. These effects of adenosine might have significant therapeuticpotential in acute brain injuries like brain trauma and stroke, but also in a widerange of chronic neurological diseases including seizures, Alzheimer’s disease,Parkinson’s disease, Huntington’s disease and multiple sclerosis [10, 27, 65, 175,179, 216, 230].Before considering possible pharmacological tools to manipulate the adeno-sinergic system it is important to realize that prolonged stimulation of adenosinereceptors leads to receptor desensitization. This process involves uncoupling ofthe activated receptor from its G-protein by receptor phosphorylation mediatedby G-protein kinases (GRK’s). Internalization of receptors into intracellularcompartments may also occur [28]. Ligand stimulation for hours to days causesreceptor down regulation. In this case degradation of receptors leads to a decreasein actual receptor number.Desensitization of adenosine A1 receptors in several tissues including brain,requires exposure to agonist for at least 15 minutes to hours or even days, while
Introduction
17

adenosine A3 receptors in astrocytes undergo significant desensitization alreadyafter several minutes after stimulation [44, 170, 213]. Long-term stimulation ofadenosine receptors with antagonists generally leads to an increase in receptornumber [170]. Stimulation of adenosine A1 receptors as well as inhibition of adenosine A2A
receptors reduces neuronal damage when administered acutely [51]. Accordingly,mice lacking adenosine A2A receptors show less neuronal damage in ischaemiamodels [30, 38]. Surprisingly, it has also been reported that chronic stimulationof adenosine A1 receptors or chronic inhibition of adenosine A2A receptors,aggravates neuronal damage [53, 107]. These contradictory results, a phenomenoncalled “effect inversion”, may be caused by desensitization and up-regulation ofadenosine receptors due to the chronic agonists and antagonists treatment,respectively [53, 107].
Possible pharmacological approaches to increase the neuroprotective effects ofadenosineIt is clear that the adenosinergic system is important to maintain a healthynervous system. It may thus be attractive to evaluate therapeutic approaches toseveral neurological diseases based on the adenosinergic system. Severalpharmacological approaches to manipulate the adenosinergic system are availableand can be divided in two categories: synthetic adenosine derivatives that directlystimulate adenosine receptors or factors that indirectly increase the effectivenessof endogenous adenosine. Table 1.3 shows a summary of neuroprotective effectsof different pharmacological approaches in various models of brain pathology.
STIMULATION OF ADENOSINE RECEPTORS
Stable synthetic adenosine derivatives that can cross the blood-brain barrier makemuch better candidates for clinical use than adenosine, which is instantly degraded.For clinical use it is also mandatory that these compounds are effective evenwhen administered hours after the pathological event, e.g. stroke. The adenosineA1 agonists CHA and R-PIA and the adenosine-amine congener ADAC showedneuroprotective effects 30 minutes to several hours after cerebral ischaemia [108,130, 225].However, due to the widespread distribution of adenosine receptors throughoutthe body, especially the A1 subtype, peripheral side effects often occur whenusing adenosine A1 receptor agonists [202]. This could be prevented by usingadenosine A2A receptor antagonists, which have less effect on heart rate andblood pressure than adenosine A1 receptor agonists [202].
Chapter 1
18
Table 1.3. Neuroprotective effects of adenosine by using different therapeutic approaches.

Introduction
19
Experimental model Drug Mechanism Effect refs
Kainic acid injection in hippocampus adenosine agonist protection [133]Kainic acid induced toxicity 2-CA agonist protection [11]rat hippocampal cell culture CPA A1 agonist protection [142]
cell injuryvessel occlusion in rats (forebrain R-PIA A1 agonist protection [26]
ischaemia)/ KA induced seizurescarotid artery occlusion in gerbils ADAC A1 agonist protection [223]
hypoxia, ischaemiacarotid artery ligation in newborn rats PD 81,273 allosteric enhancer protection [99]
hypoxia, ischaemia of A1 receptor binding hyperglycemic cerebral ischaemia PD 81,273 allosteric enhancer protection [138]
of A1 receptor bindingpreconditioning MCA occlusion followed DPCPX A1 antagonist reduction of [146]
by longer MCA occlusion protective effecthypoxia, ischaemia of preconditioning
quinolic acid injection# combined with SCH 58261 A2A antagonist protection [19]free radicals (xanthine) in hippocampus ZM 241358
β-amyloid toxicity in cultured rat neurons caffeine antagonist protection [51]ZM 241358 A2A antagonist
carotid artery occlusion in newborn rats theofylline antagonist protection [30]hypoxia, ischaemia SCH 58261 A2A antagonist
quinolic acid induced neurotoxicity# SCH 58261 A2A antagonist protection [167]MPTP induced neurotoxicity* SCH 58261 A2A antagonist protection [40]MPTP induced neurotoxicity* KW-6002 A2A antagonist protection [104]MCA occlusion in rats GP683 adenosine kinase protection [211]
hypoxia, ischaemia inhibitorMCA occlusion in rats 5’d-5IT adenosine kinase protection [109]
hypoxia, ischaemia inhibitorcell death by stimulation with propentofylline adenosine uptake protection [81]
macrophage/microglial products inhibitorcarotid artery occlusion propentofylline adenosine uptake increased cerebral [217]
hypoxia, ischaemia inhibitor blood flowsubclavian and brachiocephalic artery NBTI adenosine uptake protection against [91]
occlusion. ischaemia inhibitor reperfusion injurybilateral artery occlusion in gerbils deoxyco- adenosine deaminase protection [165]
ischaemia formycin inhibitor
# animal model for Huntington’s disease, * animal model for Parkinson’s disease. 2-CA = 2-chloroadenosine,CPA= N6- cyclopentyladenosine, R-PIA = R- N6-phenylisopropyladenosine, ADAC = adenosine aminecongener, PD 81,273 = a 2-amino-3-benzylthiophene(no details given in paper), DPCPX = 8-cyclopentyl-1,3-dipropylxanthine, SCH 58261 = 7-(2-phenylethyl)-5-amino-2-(2-furyl)pyrazolo-[4,3-e]-1,2,4-triazolo[1,5-c]pyrimidine, ZM 241358 = 4-(2-[7-amino-2-{2-furyl}{1,2,4}triazolo{2,3-a}{1,3,5}triazin-5-yl-amino]ethyl)phenol, KW-6002 = (E)-1,3-diethyl-8-(3,4-dimethoxystyryl)-7-methyl-3,7-dihydro-1H-purine-2,6-dione, GP 683 = 4-(N-phenylamino)-5-phenyl-7-(5’-deoxy β- D-ribofurasonyl)pyrrolo[2,3-d]pyrimidine,5’d-5IT = 5’-deoxy-5-iodotubercidin, NBTI = nitrobenzylthioinosine, KA = kainic acid.

INCREASING THE EFFECTIVENESS OF ENDOGENOUS ADENOSINE
Compounds that increase the effectiveness of endogenous adenosine will onlyenhance effects at sites of high extracellular adenosine levels. Presumably, thisapproach would induce tissue/region-specific effects without much peripheralside effects. Inhibition of enzymes that metabolize adenosine, like adenosinedeaminase or adenosine kinase have been shown to increase the neuroprotectiveeffects of adenosine [53, 109, 165]. The extracellular concentration of adenosinecan also be increased by inhibition of adenosine reuptake [81]. Yet anotherapproach to increase the effectiveness of adenosine is the use of factors, so-calledallosteric enhancers, that do not activate the adenosine receptor itself butenhance the binding of endogenous adenosine to the receptor [99].
Alzheimer’s diseaseIt has been suggested that chronic neurological diseases as well as acute braininjuries could be treated with trophic factors like nerve growth factor (NGF),brain-derived neurotrophic factor (BDNF) and glial-derived neurotrophic factor(GDNF). However, poor penetration of the blood-brain barrier and theoccurrence of side effects limits the use of exogenous application of these factors.Therefore stimulating the local production of trophic factors seems a moreattractive approach [43].Two synthetic purine derivatives, propentofylline and AIT-082, are currentlyunder evaluation in clinical trials for the treatment of Alzheimer’s disease. Bothcompounds have been shown in vivo to increase the mRNA expression for NGF,neurotrophin-3 and basic fibroblast growth factor (bFGF) in vivo. As NGF isconsidered to protect cholinergic neurons, which degenerate in Alzheimer’sdisease, it has been suggested that propentofylline and AIT-082 might have aneuroprotective effect in Alzheimer’s disease [43, 57, 96, 118].Propentofylline acts as an adenosine uptake inhibitor thereby maintaining highconcentrations of adenosine in the extracellular space. Furthermore, it has beendemonstrated that propentofylline stimulates the production of NGF in culturedmouse astrocytes [194]. In addition, the capacity of propentofylline to improvecerebral blood flow presumably also contributes to its’ neuroprotective effect [43,217]. It is not yet known whether propentofylline has been successful in clinicaltrials since the pharmaceutical company involved is reluctant to publish theresults. A meta-analysis of clinical trial results published so far, did not show anybeneficial effect of propentofylline in patients with Alzheimer’s disease [84].AIT-082 is under evaluation in clinical trials as a memory-enhancing agent since itincreases NGF release from glia cells, it enhances NGF induced neurite outgrowthin a neuron-like cell line and it protects neuronal tissue from damage in vivo [57,141, 174]. The mechanism of AIT-082-induced protection remains to be elu-cidated, but it has been suggested that AIT-082 increases the release of adenosine
Chapter 1
20

from astrocytes. First clinical trials have been performed with AIT-082, nowrenamed as NeotrofinTM, to investigate pharmacokinetics and tolerability [96].
Parkinson’s diseaseParkinson’s disease is caused by degeneration of dopaminergic neurons in thesubstantia nigra that innervate the striatum. The subsequent decreased levels ofdopamine in the striatum lead to a disturbed regulation of motor behavior causingthe symptoms typically observed in Parkinson’s disease. In the striatum dopamineD2 receptors are co-localized with adenosine A2A receptors whereas dopamine D1
receptors are in close proximity of adenosine A1 receptors. Through a system ofreceptor cross talk, adenosine counteracts the actions of the neuro-transmitterdopamine. Whereas stimulation with dopamine or other dopamine D2 receptoragonists enhances motor activity, stimulation of adenosine A2A receptors reducesthis effect [88]. Likewise, stimulation with adenosine A1 receptor agonistscounteracts the enhancing effect of dopamine D1 receptor agonists on motorbehavior. These interactions are probably responsible for the motor stimulanteffects of adenosine receptor antagonists like caffeine. Furthermore, adenosineA2A receptor antagonists have been reported to attenuate the neurotoxicityobserved in a mouse model of Parkinson’s disease [40, 104]. In addition,adenosine A2A receptor antagonists were found not only to diminish thesymptoms of Parkinson’s disease but also to potentiate the effect of L-DOPA[229]. L-DOPA, a dopamine precursor, which is currently used to treatParkinson’s disease, shows significant side effects like dyskinesia that are observedespecially in patients that receive high dosages of L-DOPA. If adenosine A2A
receptor antagonists indeed increase the efficacy of L-DOPA, lower doses of L-DOPA could be used and less side effects would occur [25]. All these findingssuggest that adenosine A2A receptor antagonists could be useful in the treatmentof Parkinson’s disease [192]. Recently, phase II clinical trials of the adenosine A2A
receptor antagonist, KW-6002 (IstradefyllineR) have been performed and showedrelief of Parkinson’s disease motor symptoms without side effects [102, 114, 120].Phase III clinical trials will start shortly (Schwarzschild, personal communication).
Adenosine A1 receptor expression in pathological events
During seizure activity, cerebral hypoxia and ischemia elevated extracellularconcentrations of adenosine have been found in brain tissue [65, 67]. It has beenassumed that these elevated levels of extracellular adenosine cause endogenousanticonvulsant activity as well as neuroprotection [66, 178]. Since the mainprotective actions of adenosine are mediated via the A1 receptor, it is likely thatthe expression level of this receptor has a significant influence on the efficiency ofneuroprotection by adenosine. The study of adenosine A1 receptor expression indisease is therefore of particular interest.
Introduction
21

Several conflicting reports addressing the level of adenosine A1 receptor expressionin epilepsy have been published. Some reports show that adenosine A1 receptors arechronically reduced in epilepsy [65, 92, 150]. Others have found an upregulationof these receptors [9, 221]. It is thus not clear whether changes in adenosine A1
receptor expression might be a causal factor in the pathophysiology of epilepsy. Several reports describe modification of adenosine A1 receptor activity that isrelated to the aging process [41, 128]. This modification is most likely due to adecrease in adenosine A1 receptor expression, which has been observed in agedrats and mice [49, 50, 68, 156, 196]. A reduction of adenosine A1 receptors has also been observed in autopsy andpost-mortem samples of patients with Alzheimer’s disease [112] as well asdementia with sclerosis type pathology [54].
Glia cells
Most of the research on adenosine receptors in the brain has been focussed onneurons. Mechanisms of neuroprotection induced through neuronal adenosinereceptors have been well described. In addition glia cells form a considerablecomponent of the nervous system and play an important role in for example braindevelopment, nerve tissue maintenance, modulation of synaptic transmission,formation of the blood-brain barrier and brain immune function [113]. This thesiswill partly focus on the role of glia cells in adenosine-induced neuroprotection. Itherefore briefly describe here the properties and functions of glia cells.
Microglia
Unlike neurons and astrocytes, microglia are derived from bone-marrow andmigrate to the brain during development. Microglia are the primaryimmunocompetent cells of the brain. Under physiological conditions microgliaare resting ramified cells, with many large processes to monitor theirsurroundings. If damage in the CNS occurs, microglia become activated, retracttheir processes and adapt a round macrophage-like morphology [199]. Microgliacan orchestrate neuroinflammation by producing cytokines and by presentingantigens. Furthermore microglia can phagocytose both cells and cell debris [7].Since microglia can produce neurotoxic substances, activation of microglia, so-called reactive microgliosis has been linked to the pathophysiology of many typesof brain pathology [12, 15, 205]. In addition to their detrimental activity,microglia can also assume a neurosupportive role by producing neurotrophins[145, 205, 206]. Whether microglia mediate detrimental or beneficial effectsprobably depends on a variety of factors. Indeed it has been shown that adenosinereceptors are involved in the regulation of microglia activation [105].
Chapter 1
22

Astrocytes
Astrocytes are the most abundant cells in the brain; by estimation the number ofneurons in the human brain is outnumbered ten times by astrocytes. Duringearly brain development astrocytes form growth tracts to guide the migration ofneurons. In this development phase astrocytes also produce trophic factors thussupporting neurons in their development. In the adult brain, astrocytes play anessential role in the maintenance of the nervous system. Astrocytes that surroundglutamatergic synapses regulate the extracellular glutamate concentration byactively taking up glutamate (Figure 1.5). This mechanism prevents the occurren-ce of glutamate excitotoxicity [8]. Since the active uptake of glutamate requires alarge amount of energy, it is clear that a balanced cell metabolism in astrocytessurrounding the synaptic cleft is of the utmost importance. It is likely that besidestheir involvement in inhibiting neurotransmitter release from the presynapticneuron, adenosine receptors also play an essential role in regulating cellmetabolism in astrocytes surrounding the synaptic cleft.
Introduction
23
GLUTAMATE
Na+
Glutamatergic synapse Astrocyte
Ca2+ Na+
ATP
ADP
Na+
ATPase/K+
K+
OH-/HCO3-
GLUTAMINEGlutamate
GlnNa+
G
Glutamate receptorsMetabotropic Ionotropic
K+
Figure 1.5. Regulation of glutamate concentration in the synaptic cleft by astrocytes. Glutamatereleased presynaptically stimulates glutamate receptors on the postsynaptic neuron leading todepolarization. Excessive stimulation of glutamate receptors can lead to an increase of theintracellular Ca2+ concentration, resulting in neuronal damage. Astrocytes control the action ofglutamate by actively taking up glutamate from the synaptic cleft. Glutamate is cotransported withNa+, leading to an increase of the Na+ concentration in astrocytes. Na+ concentrations inastrocytes are regulated by the energy consuming Na+/ K+ - ATPase (Adapted from ref 215).

During physiological conditions but also in brain pathology, astrocytes are knownto produce and release neuroprotective substances. Furthermore, astrocytesconstitute an important component of the blood-brain barrier, a structure thatprevents antigens from entering the brain.
Both astrocytes and microglia fulfill various important roles in regulation ofneuronal function. It is likely that adenosine is widely involved in the molecularmechanisms underlying the multiple functions of these glia cells [43]. Thereforepart of this thesis is focussed on glial adenosine receptors and their involvementin glial function is being reviewed in chapter 2.
Interleukin-6 in the brain
The cytokine interleukin-6 (IL-6), like adenosine is released duringneuropathological conditions and has been shown to mediate neuroprotectiveeffects. IL-6 belongs to the family of neuropoietic cytokines, which consists ofciliary neurotrophic factor (CNTF), leukemia inhibiting factor (LIF), oncostatin M(OSM), cardiotrophin-1 (CT-1), IL-6 and IL-11 [209].This family of cytokines is involved in several biological functions includingimmune responsivity and hematopoiesis [117]. IL-6 does not only elicit itsfunctions in the peripheral immune system. Since IL-6 is also produced byneurons, astrocytes and microglia, various additional functions of IL-6 in thebrain have been proposed [21, 97, 182, 187, 220, 233]. Under physiologicalconditions IL-6 levels in the brain are very low or undetectable, but duringpathological events like neuroinflammation, ischaemia and seizures, IL-6 levelsrise dramatically [97, 127, 134, 161, 222]. IL-6 mediates contrasting effects in thebrain [89]. IL-6 is involved in the neuroimmune response, causing neuronaldegeneration. Therefore, IL-6 has been associated with the pathophysiology ofneurodegenerative disorders like Alzheimer’s disease [18, 158, 234]. On theother hand, IL-6 plays a role in neuronal and glial differentiation and survival [89,97]. Numerous reports show neuroprotective effects of IL-6. In vitro, IL-6 protectsneurons during ischemia or induced excitotoxicity [123, 137, 232]. In vivo, IL-6shows neuroprotective effects in a number of different animal models [6, 20, 127,132]. IL-6 deficient mice showed increased neuronal death in animal models likeexperimentally induced brain-injury, the MPTP (Parkinson’s disease) model andafter kainic acid-induced seizures [29, 162, 163, 207].IL-6 elicits effects in CNS cells by binding to the IL-6 receptor, which is asso-ciated with the transmembrane transduction peptide gp130. Two forms of the IL-6 receptor have been identified; an extracellular soluble form and a membranebound form. Under both conditions the receptor can induce signal transduction
Chapter 1
24

after binding to gp130 [116]. The gp130-IL6 receptor complex can activatesecond messenger pathways involving JAK kinases and homodimerization ofSTAT3, which then act on the IL-6 response element to activate genetranscription leading to protein synthesis [190]. An alternative pathway involvesthe RAS/MAPK cascade and the activation of the nuclear factor NF-IL-6 [97]. Although the signaling pathway involved in IL-6 action has been described, thecomplete signaling cascade leading to the neuroprotective effects of IL-6 is stilllargely unknown. IL-6 might have neuroprotective properties due to induction ofother neuroprotective substances since IL-6 has been found to induce the expres-sion of vascular growth factor, a factor involved in angiogenesis [45]. Furthermore,IL-6 was found to induce the release of NGF from astrocytes [34, 121, 131].
Aim of the thesis
Adenosine is released during pathological conditions and has significantneuroprotective effects mainly by stimulating adenosine A1 receptors in neurons.These neuroprotective effects are increased following upregulation of adenosineA1 receptors. Much research has been performed to enhance the neuroprotectiveeffects of adenosine experimentally. Since direct interference with theadenosinergic system causes side effects, it would be preferable to find ways toincrease the neuroprotective effects of adenosine indirectly, for example byfinding factors that increase adenosine A1 receptor expression.
Like adenosine, the proinflammatory cytokine interleukin-6 (IL-6) is releasedduring pathological conditions and IL-6 is also known to reduce neuronal damageand mortality. In contrast to adenosine, however, little is known so far regardingthe mechanism of IL-6 mediated neuroprotection. Recent findings in vitro haveshown that IL-6 is being released by cultured astrocytes after adenosine receptorstimulation [76, 191]. Furthermore, it has been shown that stimulation with IL-6increases the expression of adenosine A1 receptors in nervous tissue, which impliesthat the neuroprotective effect of IL-6 might partially be due to upregulation ofadenosine A1 receptors [23]. From these findings we propose a model for inter-actions between the adenosinergic system and IL-6 (Figure 1.6).
The aim of the current thesis is to further investigate the interactions between theadenosinergic system and IL-6 and to check whether IL-6 has an effect on adenosine A1
receptor expression in pathological conditions.
While the mechanisms of direct adenosine-induced neuroprotection in neuronsare well understood, less is known on putative neuroprotective effects of
Introduction
25

adenosine that are mediated by glia cells. Therefore part of this thesis focuses onthe role of glia cells in adenosine-induced neuroprotection. In chapter 2 we have reviewed current knowledge on the neuroprotective sub-stances that are released by glia cells after adenosine receptor stimulation. Inaddition, in chapter 3 we show that the chemokine CCL2 is another factor with“presumed” neuroprotective effects that is released after stimulation of glialadenosine receptors.Most research on receptor pharmacology has been performed on rat and humanadenosine receptors while mouse adenosine receptors have not been fullycharacterized. In order to study the adenosinergic system in knock out models inmice, it was necessary to investigate pharmacological properties of mouseadenosine receptors. In chapter 4 we therefore present a pharmacologicalcharacterization of the mouse adenosine A1 receptor using functional studies andradioligand binding assays.In chapter 5 we have investigated the effect of IL-6 on the regulation ofadenosine A1 receptor expression during seizures. Finally, the results have beensummarized and discussed in chapter 6.
Chapter 1
26
upregulation?
Neuropathologicalevent
(4)
stimulation(3)
Adenosine
Adenosine A1receptors
stimulation (2)
Adenosine A2Breceptors
release? (3)
IL-6
Neuroprotection
variousmechanisms? (2)(5)
release (1)
Figure 1.6. Proposed model of interactions between adenosine and IL-6.Neuropathological events lead to an increase of extracellular adenosine concentration (1). Adeno-sine A1 receptors in neurons are stimulated, resulting in membrane hyperpolarization andinhibition of neurotransmitter release, which will protect neurons (2). Stimulation of adenosineA2B receptors on glial cells leads to a release of interleukin-6 (3), which will subsequently resultin adenosine A1 receptor upregulation (in neurons) (4) and thereby increasing adenosine’sneuroprotective effects. Question marks (3,4) indicate that it is not yet known if theseinteractions exist in vivo. IL-6 has neuroprotective effects but the mechanisms are unknown (5).

References
Introduction
27
1. Abbracchio, M. P., Camurri, A., Ceruti, S., Cattabeni, F., Falzano, L., Giammarioli, A. M., Jacobson, K. A., Trincavelli, L., Martini, C., Malorni, W., and Fiorentini, C. 2001. The A3 adenosine receptor induces cytoskeleton rearrangement in human astrocytoma cells via a specific action on Rho proteins.Ann.N.Y.Acad.Sci. 939: 63-73
2. Abbracchio, M. P., Cattabeni, F., Fredholm, B. B., and Williams, M. 1993. Purinoceptor Nomenclature: A status report. Drug development research 28: 207-213
3. Abbracchio, M. P., Ceruti, S., Brambilla, R., Barbieri, D., Camurri, A., Franceschi, C., Giammarioli, A. M., Jacobson, K. A., Cattabeni, F., and Malorni, W. 2003. Adenosine A3 receptors and viability of astrocytes. Drug development research 45: 379-386
4. Abbracchio, M. P., Ceruti, S., Brambilla, R., Franceschi, C., Malorni, W., Jacobson, K. A., von Lubitz, D. K., and Cattabeni, F. 1997. Modulation of apoptosis by adenosine in the central nervous system: a possible role for the A3 receptor. Pathophysiological significance and therapeutic implications for neurodegenerative disorders. Ann.N.Y.Acad.Sci. 825: 11-22
5. Abbracchio, M. P., Rainaldi, G., Giammarioli, A. M., Ceruti, S., Brambilla, R., Cattabeni, F., Barbieri, D., Franceschi, C., Jacobson, K. A., and Malorni, W. 1997. The A3 adenosine receptor mediates cell spreading, reorganization of actin cytoskeleton, and distribution of Bcl-XL: studies in human astroglioma cells. Biochem.Biophys.Res.Commun. 241(2): 297-304
6. Ali, C., Nicole, O., Docagne, F., Lesne, S., MacKenzie, E. T., Nouvelot, A., Buisson, A., and Vivien, D. 2000. Ischemia-induced interleukin-6 as a potential endogenous neuroprotective cytokine againstNMDA receptor-mediated excitotoxicity in the brain. J Cereb.Blood Flow Metab 20(6): 956-966
7. Aloisi, F. 2001. Immune function of microglia. Glia 36(2): 165-1798. Anderson, C. M. and Swanson, R. A. 2000. Astrocyte glutamate transport: review of properties,
regulation, and physiological functions. Glia 32(1): 1-149. Angelatou, F., Pagonopoulou, O., Maraziotis, T., Olivier, A., Villemeure, J. G., Avoli, M., and
Kostopoulos, G. 1993. Upregulation of A1 adenosine receptors in human temporal lobe epilepsy: a quantitative autoradiographic study. Neurosci Lett 163(1): 11-14
10. Angulo, E., Casado, V., Mallol, J., Canela, E. I., Vinals, F., Ferrer, I., Lluis, C., and Franco, R. 2003. A1 adenosine receptors accumulate in neurodegenerative structures in Alzheimer disease and mediate both amyloid precursor protein processing and tau phosphorylation and translocation. Brain Pathol. 13(4): 440-451
11. Arvin, B., Neville, L. F., Pan, J., and Roberts, P. J. 1989. 2-chloroadenosine attenuates kainic acid-induced toxicity within the rat straitum: relationship to release of glutamate and Ca2+ influx.Br J Pharmacol 98(1): 225-235
12. Aschner, M., Allen, J. W., Kimelberg, H. K., LoPachin, R. M., and Streit, W. J. 1999. Glial cells in neurotoxicity development. Annu.Rev.Pharmacol.Toxicol. 39: 151-173
13. Avery, T. L., Rehg, J. E., Lumm, W. C., Harwood, F. C., Santana, V. M., and Blakley, R. L. 1989. Biochemical pharmacology of 2-chlorodeoxyadenosine in malignant human hematopoietic cell lines and therapeutic effects of 2-bromodeoxyadenosine in drug combinations in mice. Cancer Res. 49(18):4972-4978
14. Baines, C. P., Cohen, M. V., and Downey, J. M. 1999. Signal transduction in ischemic preconditioning:the role of kinases and mitochondrial K(ATP) channels. J Cardiovasc.Electrophysiol. 10(5): 741-754
15. Banati, R. B., Gehrmann, J., Schubert, P., and Kreutzberg, G. W. 1993. Cytotoxicity of microglia. Glia 7(1): 111-118
16. Bar-Yehuda, S., Madi, L., Barak, D., Mittelman, M., Ardon, E., Ochaion, A., Cohn, S., and Fishman, P. 2002. Agonists to the A3 adenosine receptor induce G-CSF production via NF-kappaB activation: a new class of myeloprotective agents. Exp.Hematol. 30(12): 1390-1398
17. Basheer, R., Rainnie, D. G., Porkka-Heiskanen, T., Ramesh, V., and McCarley, R. W. 2001.Adenosine, prolonged wakefulness, and A1-activated NF-kappaB DNA binding in the basal forebrain of the rat. Neuroscience 104(3): 731-739

Chapter 1
28
18. Bauer, J., Strauss, S., Volk, B., and Berger, M. 1991. IL-6-mediated events in Alzheimer's disease pathology. Immunol.Today 12(11): 422
19. Behan, W. M. and Stone, T. W. 2002. Enhanced neuronal damage by co-administration of quinolinicacid and free radicals, and protection by adenosine A2A receptor antagonists. Br J Pharmacol 135(6): 1435-1442
20. Bensadoun, J. C., de Almeida, L. P., Dreano, M., Aebischer, P., and Deglon, N. 2001. Neuroprotectiveeffect of interleukin-6 and IL6/IL6R chimera in the quinolinic acid rat model of Huntington's syndrome. Eur J Neurosci. 14(11): 1753-1761
21. Benveniste, E. N., Sparacio, S. M., Norris, J. G., Grenett, H. E., and Fuller, G. M. 1990. Induction and regulation of interleukin-6 gene expression in rat astrocytes. J Neuroimmunol. 30(2-3): 201-212
22. Berne, R. M. 1963. Cardiac nucleotides in hypoxia: possible role in regulation of coronary blood flow. Am.J Physiol 204: 317-322
23. Biber, K., Lubrich, B., Fiebich, B. L., Boddeke, H. W., and van Calker, D. 2001. Interleukin-6 enhances expression of adenosine A(1) receptor mRNA and signaling in cultured rat cortical astrocytes and brain slices. Neuropsychopharmacology 24(1): 86-96
24. Blackburn, M. R. 2003. Too much of a good thing: adenosine overload in adenosine-deaminase-deficient mice. Trends Pharmacol.Sci. 24(2): 66-70
25. Blandini, F. 2003. Adenosine receptors and L-DOPA-induced dyskinesia in Parkinson's disease: potential targets for a new therapeutic approach. Exp.Neurol 184(2): 556-560
26. Blondeau, N., Plamondon, H., Richelme, C., Heurteaux, C., and Lazdunski, M. 2000. K(ATP) channel openers, adenosine agonists and epileptic preconditioning are stress signals inducing hippocampal neuroprotection. Neuroscience 100(3): 465-474
27. Blum, D., Hourez, R., Galas, M. C., Popoli, P., and Schiffmann, S. N. 2003. Adenosine receptors andHuntington's disease: implications for pathogenesis and therapeutics. Lancet Neurol 2(6): 366-374
28. Bohm, S. K., Grady, E. F., and Bunnett, N. W. 1997. Regulatory mechanisms that modulate signalling by G-protein-coupled receptors. Biochem.J 322 ( Pt 1): 1-18
29. Bolin, L. M., Strycharska-Orczyk, I., Murray, R., Langston, J. W., and Di Monte, D. 2002. Increased vulnerability of dopaminergic neurons in MPTP-lesioned interleukin-6 deficient mice. J Neurochem. 83(1): 167-175
30. Bona, E., Aden, U., Gilland, E., Fredholm, B. B., and Hagberg, H. 1997. Neonatal cerebral hypoxia-ischemia: the effect of adenosine receptor antagonists. Neuropharmacology 36(9): 1327-1338
31. Brand, A., Vissiennon, Z., Eschke, D., and Nieber, K. 2001. Adenosine A(1) and A(3) receptors mediate inhibition of synaptic transmission in rat cortical neurons. Neuropharmacology 40(1): 85-95
32. Burnstock, G. 1978. A basis for distinguishing two types of purinergic receptor. In Cell membrane receptors for drugs and hormones: A multidisciplinary approach, Bollis, L and Straub, R. W. 107-118.New York: Raven Press.
33. Burnstock, G. 2002. Purinergic signaling and vascular cell proliferation and death. Arterioscler. Thromb.Vasc.Biol. 22(3): 364-373
34. Carlson, N. G., Wieggel, W. A., Chen, J., Bacchi, A., Rogers, S. W., and Gahring, L. C. 1999. Inflammatory cytokines IL-1 alpha, IL-1 beta, IL-6, and TNF-alpha impart neuroprotection to an excitotoxin through distinct pathways. J Immunol 163(7): 3963-3968
35. Ceruti, S., Franceschi, C., Barbieri, D., Malorni, W., Camurri, A., Giammarioli, A. M., Ambrosini, A., Racagni, G., Cattabeni, F., and Abbracchio, M. P. 2000. Apoptosis induced by 2-chloro-adenosineand 2-chloro-2'-deoxy-adenosine in a human astrocytoma cell line: differential mechanisms and possible clinical relevance. J Neurosci Res 60(3): 388-400
36. Charles, M. P., Adamski, D., Kholler, B., Pelletier, L., Berger, F., and Wion, D. 2003. Induction of neurite outgrowth in PC12 cells by the bacterial nucleoside N6-methyldeoxyadenosine is mediated through adenosine A2a receptors and via cAMP and MAPK signaling pathways. Biochem.Biophys.Res.Commun. 304(4): 795-800
37. Chen, J. F., Beilstein, M., Xu, Y. H., Turner, T. J., Moratalla, R., Standaert, D. G., Aloyo, V. J., Fink, J. S., and Schwarzschild, M. A. 2000. Selective attenuation of psychostimulant-induced behavioral responses in mice lacking A(2A) adenosine receptors. Neuroscience 97(1): 195-204

Introduction
29
38. Chen, J. F., Huang, Z., Ma, J., Zhu, J., Moratalla, R., Standaert, D., Moskowitz, M. A., Fink, J. S., andSchwarzschild, M. A. 1999. A(2A) adenosine receptor deficiency attenuates brain injury induced by transient focal ischemia in mice. J Neurosci 19(21): 9192-9200
39. Chen, J. F., Moratalla, R., Impagnatiello, F., Grandy, D. K., Cuellar, B., Rubinstein, M., Beilstein, M. A., Hackett, E., Fink, J. S., Low, M. J., Ongini, E., and Schwarzschild, M. A. 2001. The role of the D(2) dopamine receptor (D(2)R) in A(2A) adenosine receptor (A(2A)R)-mediated behavioral and cellular responses as revealed by A(2A) and D(2) receptor knockout mice. Proc Natl Acad Sci U S A98(4): 1970-1975
40. Chen, J. F., Xu, K., Petzer, J. P., Staal, R., Xu, Y. H., Beilstein, M., Sonsalla, P. K., Castagnoli, K., Castagnoli, N., Jr., and Schwarzschild, M. A. 2001. Neuroprotection by caffeine and A(2A) adenosine receptor inactivation in a model of Parkinson's disease. J Neurosci 21(10): RC143.
41. Cheng, J. T., Liu, I. M., Juang, S. W., and Jou, S. B. 2000. Decrease of adenosine A-1 receptor gene expression in cerebral cortex of aged rats. Neurosci Lett 283(3): 227-229
42. Churchill, P. C. and Bidani, A. K. 1990. Adenosine and renal function. In Adenosine and adenosine receptors, Williams, M. 335-380. Clifton, NJ: The Humana Press.
43. Ciccarelli, R., Ballerini, P., Sabatino, G., Rathbone, M. P., D'Onofrio, M., Caciagli, F., and Di Iorio, P. 2001. Involvement of astrocytes in purine-mediated reparative processes in the brain.Int J Dev Neurosci 19(4): 395-414
44. Ciruela, F., Saura, C., Canela, E. I., Mallol, J., Lluis, C., and Franco, R. 1997. Ligand-induced phos-phorylation, clustering, and desensitization of A1 adenosine receptors. Mol.Pharmacol. 52(5): 788-797
45. Cohen, T., Nahari, D., Cerem, L. W., Neufeld, G., and Levi, B. Z. 1996. Interleukin 6 induces the expression of vascular endothelial growth factor. J Biol.Chem. 271(2): 736-741
46. Costenla, A. R., Lopes, L. V., de Mendonca, A., and Ribeiro, J. A. 2001. A functional role for adenosineA3 receptors: modulation of synaptic plasticity in the rat hippocampus. Neurosci Lett 302(1): 53-57
47. Cronstein, B. N. 1994. Adenosine, an endogenous anti-inflammatory agent. J Appl.Physiol 76(1): 5-1348. Cunha, R. A. 2001. Adenosine as a neuromodulator and as a homeostatic regulator in the nervous
system: different roles, different sources and different receptors. Neurochem Int 38(2): 107-12549. Cunha, R. A., Constantino, M. C., Sebastiao, A. M., and Ribeiro, J. A. 1995. Modification of A1 and
A2a adenosine receptor binding in aged striatum, hippocampus and cortex of the rat. Neuroreport6(11): 1583-1588
50. Cunha, R. A., Constantino, M. D., Fonseca, E., and Ribeiro, J. A. 2001. Age-dependent decrease in adenosine A1 receptor binding sites in the rat brain. Effect of cis unsaturated free fatty acids. Eur J Biochem 268(10): 2939-2947
51. Dall'lgna, O. P., Porciuncula, L. O., Souza, D. O., Cunha, R. A., and Lara, D. R. 2003. Neuroprotectionby caffeine and adenosine A2A receptor blockade of beta-amyloid neurotoxicity. Br.J Pharmacol. 138(7): 1207-1209
52. Dassesse, D., Massie, A., Ferrari, R., Ledent, C., Parmentier, M., Arckens, L., Zoli, M., and Schiffmann, S. N. 2001. Functional striatal hypodopaminergic activity in mice lacking adenosine A(2A) receptors. J Neurochem 78(1): 183-198
53. de Mendonca, A., Sebastiao, A. M., and Ribeiro, J. A. 2000. Adenosine: does it have a neuropro-tective role after all? Brain Res Brain Res Rev 33(2-3): 258-274
54. Deckert, J., Abel, F., Kunig, G., Hartmann, J., Senitz, D., Maier, H., Ransmayr, G., and Riederer, P. 1998. Loss of human hippocampal adenosine A1 receptors in dementia: evidence for lack of specificity. Neurosci.Lett. 244(1): 1-4
55. Deckert, J., Morgan, P. F., Bisserbe, J. C., Jacobson, K. A., Kirk, K. L., Daly, J. W., and Marangos, P. J. 1988. Autoradiographic localization of mouse brain adenosine receptors with an antagonist ([3H]xanthine amine congener) ligand probe. Neurosci Lett 86(2): 121-126
56. Deuchars, S. A., Brooke, R. E., and Deuchars, J. 2001. Adenosine A1 receptors reduce release from excitatory but not inhibitory synaptic inputs onto lateral horn neurons. J Neurosci. 21(16): 6308-6320
57. Di Iorio, P., Virgilio, A., Giuliani, P., Ballerini, P., Vianale, G., Middlemiss, P. J., Rathbone, M. P., and Ciccarelli, R. 2001. AIT-082 is neuroprotective against kainate-induced neuronal injury in rats. Exp.Neurol 169(2): 392-399

Chapter 1
30
58. Dirnagl, U., Niwa, K., Lindauer, U., and Villringer, A. 1994. Coupling of cerebral blood flow to neuronal activation: role of adenosine and nitric oxide. Am J Physiol 267(1 Pt 2): 296-301
59. Dixon, A. K., Gubitz, A. K., Sirinathsinghji, D. J., Richardson, P. J., and Freeman, T. C. 1996. Tissuedistribution of adenosine receptor mRNAs in the rat. Br.J Pharmacol. 118(6): 1461-1468
60. Dolphin, A. C., Forda, S. R., and Scott, R. H. 1986. Calcium-dependent currents in cultured rat dorsal root ganglion neurones are inhibited by an adenosine analogue. J Physiol 373: 47-61
61. Drury, A. N. and Szent-Gyorgyi, A. 1929. The physiological activity of adenine compounds with especial reference to their action upon the mammalian heart. J Physiol (Lond) 68: 213-237
62. Dubey, R. K., Gillespie, D. G., and Jackson, E. K. 2002. A(2B) adenosine receptors stimulate growthof porcine and rat arterial endothelial cells. Hypertension 39(2 Pt 2): 530-535
63. Dunwiddie, T. V. 1985. The physiological role of adenosine in the central nervous system.Int Rev Neurobiol 27: 63-139
64. Dunwiddie, T. V. 1999. Adenosine and ethanol: is there a caffeine connection in the actions of ethanol? In The "Drunken" Synapse: Studies of Alcohol-Related Disorders, Liu, Y. and Hunt, W. A.119-133. New York: Kluwer/Plenum.
65. Dunwiddie, T. V. and Masino, S. A. 2001. The role and regulation of adenosine in the central nervous system. Annu Rev Neurosci 24: 31-55
66. Dunwiddie, T. V. and Worth, T. 1982. Sedative and anticonvulsant effects of adenosine analogs in mouse and rat. J.Pharmacol.Exp.Ther. 220(1): 70-76
67. During, M. J. and Spencer, D. D. 1992. Adenosine: a potential mediator of seizure arrest and postictal refractoriness. Ann.Neurol 32(5): 618-624
68. Ekonomou, A., Pagonopoulou, O., and Angelatou, F. 2000. Age-dependent changes in adenosine A1
receptor and uptake site binding in the mouse brain: an autoradiographic study. J Neurosci Res 60(2):257-265
69. Elliott, K. J., Todd, Weber E., and Rea, M. A. 2001. Adenosine A1 receptors regulate the response ofthe hamster circadian clock to light. Eur J Pharmacol. 414(1): 45-53
70. Eschke, D., Brand, A., Scheibler, P., Hess, S., Eger, K., Allgaier, C., and Nieber, K. 2001. Effect of anadenosine A(1) receptor agonist and a novel pyrimidoindole on membrane properties and neurotransmitter release in rat cortical and hippocampal neurons. Neurochem Int 38(5): 391-398
71. Fedorova, I. M., Jacobson, M. A., Basile, A., and Jacobson, K. A. 2003. Behavioral characterization ofmice lacking the A3 adenosine receptor: sensitivity to hypoxic neurodegeneration. Cell Mol.Neurobiol.23(3): 431-447
72. Feoktistov, I. and Biaggioni, I. 1997. Adenosine A2B receptors. Pharmacol Rev 49(4): 381-40273. Feoktistov, I., Goldstein, A. E., Ryzhov, S., Zeng, D., Belardinelli, L., Voyno-Yasenetskaya, T., and
Biaggioni, I. 2002. Differential expression of adenosine receptors in human endothelial cells: role of A2B receptors in angiogenic factor regulation. Circ Res 90(5): 531-538
74. Feoktistov, I., Ryzhov, S., Goldstein, A. E., and Biaggioni, I. 2003. Mast cell-mediated stimulation ofangiogenesis: cooperative interaction between A2B and A3 adenosine receptors. Circ.Res. 92(5): 485-492
75. Feoktistov, I., Wells, J. N., and Biaggioni, I. 1998. Adenosine A2B receptors as therapeutic targets. Drug development research 45: 198-206
76. Fiebich, B. L., Biber, K., Gyufko, K., Berger, M., Bauer, J., and van Calker, D. 1996. Adenosine A2b
receptors mediate an increase in interleukin (IL)-6 mRNA and IL-6 protein synthesis in human astroglioma cells. J Neurochem 66(4): 1426-1431
77. Firestein, G. S. 1996. Anti-inflammatory effects of adenosine kinase inhibitors in acute and chronic inflammation. Drug development research 39: 371-376
78. Fischer, S., Sharma, H. S., Karliczek, G. F., and Schaper, W. 1995. Expression of vascular permeability factor/vascular endothelial growth factor in pig cerebral microvascular endothelial cellsand its upregulation by adenosine. Brain Res Mol Brain Res 28(1): 141-148
79. Fishman, P., Bar-Yehuda, S., Barer, F., Madi, L., Multani, A. S., and Pathak, S. 2001. The A3
adenosine receptor as a new target for cancer therapy and chemoprotection. Exp.Cell Res. 269(2): 230-236

Introduction
31
80. Fishman, P., Bar-Yehuda, S., Farbstein, T., Barer, F., and Ohana, G. 2000. Adenosine acts as a chemoprotective agent by stimulating G-CSF production: a role for A1 and A3 adenosine receptors.J Cell Physiol 183(3): 393-399
81. Flavin, M. P. and Ho, L. T. 1999. Propentofylline protects neurons in culture from death triggered by macrophage or microglial secretory products. J Neurosci Res 56(1): 54-59
82. Florio, C., Prezioso, A., Papaioannou, A., and Vertua, R. 1998. Adenosine A1 receptors modulate anxiety in CD1 mice. Psychopharmacology (Berl) 136(4): 311-319
83. Forsythe, P. and Ennis, M. 1999. Adenosine, mast cells and asthma. Inflamm.Res. 48(6): 301-30784. Frampton, M., Harvey, R. J., and Kirchner, V. 2003. Propentofylline for dementia.
Cochrane.Database.Syst.Rev.(2): CD00285385. Fredholm, B. B., Abbracchio, M. P., Burnstock, G., Daly, J. W., Harden, T. K., Jacobson, K. A., Leff,
P., and Williams, M. 1994. Nomenclature and classification of purinoceptors. Pharmacol.Rev. 46(2): 143-156
86. Fredholm, B. B., IJzerman, A.P., Jacobson, K. A., Klotz, K. N., and Linden, J. 2001. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol Rev 53(4): 527-552
87. Fredholm, B. B., Battig, K., Holmen, J., Nehlig, A., and Zvartau, E. E. 1999. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol Rev 51(1):83-133
88. Fuxe, K., Ferre, S., Zoli, M., and Agnati, L. F. 1998. Integrated events in central dopamine transmission as analyzed at multiple levels. Evidence for intramembrane adenosine A2A/dopamine D2 and adenosine A1/dopamine D1 receptor interactions in the basal ganglia. Brain Res.Brain Res.Rev.26(2-3): 258-273
89. Gadient, R. A. and Otten, U. H. 1997. Interleukin-6 (IL-6)--a molecule with both beneficial and destructive potentials. Prog Neurobiol 52(5): 379-390
90. Gerber, U. and Gahwiler, B. H. 1994. GABAB and adenosine receptors mediate enhancement of the K+ current, IAHP, by reducing adenylyl cyclase activity in rat CA3 hippocampal neurons. J Neurophysiol 72(5): 2360-2367
91. Gidday, J. M., Kim, Y. B., Shah, A. R., Gonzales, E. R., and Park, T. S. 1996. Adenosine transport inhibition ameliorates postischemic hypoperfusion in pigs. Brain Res. 734(1-2): 261-268
92. Glass, M., Faull, R. L., Bullock, J. Y., Jansen, K., Mee, E. W., Walker, E. B., Synek, B. J., and Dragunow, M. 1996. Loss of A1 adenosine receptors in human temporal lobe epilepsy. Brain Res. 710(1-2): 56-68
93. Grant, M. B., Davis, M. I., Caballero, S., Feoktistov, I., Biaggioni, I., and Belardinelli, L. 2001. Proliferation, migration, and ERK activation in human retinal endothelial cells through A(2B) adenosine receptor stimulation. Invest Ophthalmol Vis Sci 42(9): 2068-2073
94. Grant, M. B., Tarnuzzer, R. W., Caballero, S., Ozeck, M. J., Davis, M. I., Spoerri, P. E., Feoktistov, I.,Biaggioni, I., Shryock, J. C., and Belardinelli, L. 1999. Adenosine receptor activation induces vascular endothelial growth factor in human retinal endothelial cells. Circ Res 85(8): 699-706
95. Griffiths, T. L. and Holgate, S. T. 1990. The role of adenosine receptors in respiratory physiology. In Adenosine and adenosine receptors, Williams, M. 381-422. Clifton, NJ: The Humana Press.
96. Grundman, M., Capparelli, E., Kim, H. T., Morris, J. C., Farlow, M., Rubin, E. H., Heidebrink, J., Hake, A., Ho, G., Schultz, A. N., Schafer, K., Houston, W., Thomas, R., and Thal, L. J. 2003. A multicenter, randomized, placebo controlled, multiple-dose, safety and pharmacokinetic study of AIT-082 (Neotrofin) in mild Alzheimer's disease patients. Life Sci. 73(5): 539-553
97. Gruol, D. L. and Nelson, T. E. 1997. Physiological and pathological roles of interleukin-6 in the central nervous system. Mol Neurobiol 15(3): 307-339
98. Haas, H. L. and Selbach, O. 2000. Functions of neuronal adenosine receptors. Naunyn Schmiedebergs Arch Pharmacol 362(4-5): 375-381
99. Halle, J. N., Kasper, C. E., Gidday, J. M., and Koos, B. J. 1997. Enhancing adenosine A1 receptor binding reduces hypoxic-ischemic brain injury in newborn rats. Brain Res 759(2): 309-312
100. Hansen, P. B. and Schnermann, J. 2003. Vasoconstrictor and vasodilator effects of adenosine in the kidney. Am.J Physiol Renal Physiol 285(4): F590-F599

Chapter 1
32
101. Hasko, G., Deitch, E. A., Szabo, C., Nemeth, Z. H., and Vizi, E. S. 2002. Adenosine: a potential mediator of immunosuppression in multiple organ failure. Curr Opin Pharmacol 2(4): 440-444
102. Hauser, R. A., Hubble, J. P., and Truong, D. D. 2003. Randomized trial of the adenosine A(2A) receptor antagonist istradefylline in advanced PD. Neurology 61(3): 297-303
103. Hoffman, B. B., Prokocimer, P., Thomas, J. M., Vagelos, R., Chang, H., and Reaven, G. M. 1989. Cellular tolerance to adenosine receptor-mediated inhibition of lipolysis: altered adenosine 3',5'-monophosphate metabolism and protein kinase activation. Endocrinology 124(5): 2434-2442
104. Ikeda, K., Kurokawa, M., Aoyama, S., and Kuwana, Y. 2002. Neuroprotection by adenosine A2A
receptor blockade in experimental models of Parkinson's disease. J Neurochem 80(2): 262-270105. Inoue, K. 2002. Microglial activation by purines and pyrimidines. Glia 40(2): 156-163106. Jacobson, K. A. 1998. Adenosine A3 receptors: novel ligands and paradoxical effects.
Trends Pharmacol Sci 19(5): 184-191107. Jacobson, K. A., von Lubitz, D. K., Daly, J. W., and Fredholm, B. B. 1996. Adenosine receptor
ligands: differences with acute versus chronic treatment. Trends Pharmacol Sci 17(3): 108-113108. Januszewicz von Lubitz, D. K., Dambrosia, J. M., and Redmond, D. J. 1989. Protective effect of
cyclohexyl adenosine in treatment of cerebral ischemia in gerbils. Neuroscience 30(2): 451-462109. Jiang, N., Kowaluk, E. A., Lee, C. H., Mazdiyasni, H., and Chopp, M. 1997. Adenosine kinase
inhibition protects brain against transient focal ischemia in rats. Eur J Pharmacol 320(2-3): 131-7110. Johansson, B., Halldner, L., Dunwiddie, T. V., Masino, S. A., Poelchen, W., Gimenez-Llort, L.,
Escorihuela, R. M., Fernandez-Teruel, A., Wiesenfeld-Hallin, Z., Xu, X. J., Hardemark, A., Betsholtz,C., Herlenius, E., and Fredholm, B. B. 2001. Hyperalgesia, anxiety, and decreased hypoxic neuro-protection in mice lacking the adenosine A1 receptor. Proc Natl Acad Sci U S A 98(16): 9407-9412
111. Kaiser, S. M. and Quinn, R. J. 1999. Adenosine receptors as potential therapeutic targets. Drug Discov Today 4(12): 542-551
112. Kalaria, R. N., Sromek, S., Wilcox, B. J., and Unnerstall, J. R. 1990. Hippocampal adenosine A1
receptors are decreased in Alzheimer's disease. Neurosci.Lett. 118(2): 257-260113. Kandel ER, Schwartz JH and Jessell TH. Principles of neural science. London: McGraw-Hill, 2000.114. Kase, H., Aoyama, S., Ichimura, M., Ikeda, K., Ishii, A., Kanda, T., Koga, K., Koike, N., Kurokawa,
M., Kuwana, Y., Mori, A., Nakamura, J., Nonaka, H., Ochi, M., Saki, M., Shimada, J., Shindou, T., Shiozaki, S., Suzuki, F., Takeda, M., Yanagawa, K., Richardson, P. J., Jenner, P., Bedard, P., Borrelli, E., Hauser, R. A., and Chase, T. N. 2003. Progress in pursuit of therapeutic A2A antagonists: the adenosine A2A receptor selective antagonist KW6002: research and development toward a novel nondopaminergic therapy for Parkinson's disease. Neurology 61(11 Suppl 6): S97-S100
115. Kim, S. G., Ravi, G., Hoffmann, C., Jung, Y. J., Kim, M., Chen, A., and Jacobson, K. A. 2002. p53-Independent induction of Fas and apoptosis in leukemic cells by an adenosine derivative, Cl-IB-MECA. Biochem Pharmacol 63(5): 871-880
116. Kishimoto, T., Akira, S., and Taga, T. 1992. IL-6 receptor and mechanism of signal transduction. Int.J Immunopharmacol. 14(3): 431-438
117. Kishimoto, T., Akira, S., and Taga, T. 1992. Interleukin-6 and its receptor: a paradigm for cytokines.Science 258(5082): 593-597
118. Kittner, B., Rossner, M., and Rother, M. 1997. Clinical trials in dementia with propentofylline. Ann.N.Y.Acad.Sci. 826: 307-316
119. Knudsen, T. B. and Elmer, W. A. 1987. Evidence for negative control of growth by adenosine in the mammalian embryo: induction of Hmx/+ mutant limb outgrowth by adenosine deaminase. Differentiation 33(3): 270-279
120. Knutsen, L. J. and Weiss, S. M. 2001. KW-6002 (Kyowa Hakko Kogyo). Curr.Opin.Investig.Drugs2(5): 668-673
121. Kossmann, T., Hans, V., Imhof, H. G., Trentz, O., and Morganti-Kossmann, M. C. 1996. Interleukin-6 released in human cerebrospinal fluid following traumatic brain injury may trigger nerve growth factor production in astrocytes. Brain Res 713(1-2): 143-152
122. Kull, B., Ferre, S., Arslan, G., Svenningsson, P., Fuxe, K., Owman, C., and Fredholm, B. B. 1999. Reciprocal interactions between adenosine A2A and dopamine D2 receptors in Chinese hamster ovary cells co-transfected with the two receptors. Biochem Pharmacol 58(6): 1035-1045

Introduction
33
123. Kunioku, H., Inoue, K., and Tomida, M. 2001. Interleukin-6 protects rat PC12 cells from serum deprivation or chemotherapeutic agents through the phosphatidylinositol 3-kinase and STAT3 pathways. Neurosci.Lett. 309(1): 13-16
124. Ledent, C., Vaugeois, J. M., Schiffmann, S. N., Pedrazzini, T., El Yacoubi, M., Vanderhaeghen, J. J., Costentin, J., Heath, J. K., Vassart, G., and Parmentier, M. 1997. Aggressiveness, hypoalgesia and high blood pressure in mice lacking the adenosine A2a receptor. Nature 388(6643): 674-678
125. Linden, J. 1994. Cloned adenosine A3 receptors: pharmacological properties, species differences and receptor functions. Trends Pharmacol.Sci. 15(8): 298-306
126. Linden, J. 2001. Molecular approach to adenosine receptors: receptor-mediated mechanisms of tissue protection. Annu Rev Pharmacol Toxicol 41: 775-787
127. Loddick, S. A., Turnbull, A. V., and Rothwell, N. J. 1998. Cerebral interleukin-6 is neuroprotective during permanent focal cerebral ischemia in the rat. J Cereb Blood Flow Metab 18(2): 176-179
128. Lopes, L. V., Cunha, R. A., and Ribeiro, J. A. 1999. Cross talk between A(1) and A(2A) adenosine receptors in the hippocampus and cortex of young adult and old rats. J Neurophysiol 82(6): 3196-3203
129. MacDonald, R. L., Skerritt, J. H., and Werz, M. A. 1986. Adenosine agonists reduce voltage-dependent calcium conductance of mouse sensory neurones in cell culture. J Physiol 370: 75-90
130. MacGregor, D. G., Miller, W. J., and Stone, T. W. 1993. Mediation of the neuroprotective action of R-phenylisopropyl-adenosine through a centrally located adenosine A1 receptor. Br.J Pharmacol. 110(1): 470-476
131. Marz, P., Heese, K., Dimitriades-Schmutz, B., Rose-John, S., and Otten, U. 1999. Role of interleukin-6 and soluble IL-6 receptor in region-specific induction of astrocytic differentiation and neurotrophin expression. Glia 26(3): 191-200
132. Matsuda, S., Wen, T. C., Morita, F., Otsuka, H., Igase, K., Yoshimura, H., and Sakanaka, M. 1996. Interleukin-6 prevents ischemia-induced learning disability and neuronal and synaptic loss in gerbils. Neurosci Lett 204(1-2): 109-112
133. Matsuoka, Y., Okazaki, M., Takata, K., Kitamura, Y., Ohta, S., Sekino, Y., and Taniguchi, T. 1999. Endogenous adenosine protects CA1 neurons from kainic acid-induced neuronal cell loss in the rat hippocampus. Eur J Neurosci 11(10): 3617-3625
134. Matsuzono, Y., Narita, M., Akutsu, Y., and Togashi, T. 1995. Interleukin-6 in cerebrospinal fluid of patients with central nervous system infections. Acta Paediatr. 84(8): 879-883
135. Meininger, C. J. and Granger, H. J. 1990. Mechanisms leading to adenosine-stimulated proliferation of microvascular endothelial cells. Am.J Physiol 258(1 Pt 2): H198-H206
136. Meininger, C. J., Schelling, M. E., and Granger, H. J. 1988. Adenosine and hypoxia stimulate proliferation and migration of endothelial cells. Am.J Physiol 255(3 Pt 2): H554-H562
137. Mendonca Torres, P. M. and de Araujo, E. G. 2001. Interleukin-6 increases the survival of retinal ganglion cells in vitro. J Neuroimmunol 117(1-2): 43-50
138. Meno, J. R., Higashi, H., Cambray, A. J., Zhou, J., D'Ambrosio, R., and Winn, H. R. 2003. Hippocampal injury and neurobehavioral deficits are improved by PD 81,723 following hyperglycemic cerebral ischemia. Exp.Neurol 183(1): 188-196
139. Merighi, S., Mirandola, P., Milani, D., Varani, K., Gessi, S., Klotz, K. N., Leung, E., Baraldi, P. G., and Borea, P. A. 2002. Adenosine receptors as mediators of both cell proliferation and cell death of cultured human melanoma cells. J.Invest Dermatol. 119(4): 923-933
140. Merighi, S., Mirandola, P., Varani, K., Gessi, S., Leung, E., Baraldi, P. G., Tabrizi, M. A., and Borea, P. A. 2003. A glance at adenosine receptors: novel target for antitumor therapy. Pharmacol.Ther. 100(1): 31-48
141. Middlemiss, P. J., Glasky, A. J., Rathbone, M. P., Werstuik, E., Hindley, S., and Gysbers, J. 1995. AIT-082, a unique purine derivative, enhances nerve growth factor mediated neurite outgrowth from PC12 cells. Neurosci.Lett. 199(2): 131-134
142. Mitchell, H. L., Frisella, W. A., Brooker, R. W., and Yoon, K. W. 1995. Attenuation of traumatic celldeath by an adenosine A1 agonist in rat hippocampal cells. Neurosurgery 36(5): 1003-1007
143. Moreau, J. L. and Huber, G. 1999. Central adenosine A(2A) receptors: an overview. Brain Res Brain Res Rev 31(1): 65-82

Chapter 1
34
144. Mubagwa, K. and Flameng, W. 2001. Adenosine, adenosine receptors and myocardial protection: anupdated overview. Cardiovasc Res 52(1): 25-39
145. Nakajima, K., Honda, S., Tohyama, Y., Imai, Y., Kohsaka, S., and Kurihara, T. 2001. Neurotrophin secretion from cultured microglia. J Neurosci Res 65(4): 322-331
146. Nakamura, M., Nakakimura, K., Matsumoto, M., and Sakabe, T. 2002. Rapid tolerance to focal cerebral ischemia in rats is attenuated by adenosine A1 receptor antagonist. J Cereb Blood Flow Metab22(2): 161-170
147. Neary, J.T., Rathbone, M. P., Cattabeni, F., Abbracchio, M. P., and Burnstock, G. 1996.Trophic actionsof extracellular nucleotides and nucleosides on glial and neuronal cells. Trends Neurosci 19(1): 13-18
148. Noguchi, J. and Yamashita, H. 2000. Adenosine inhibits voltage-dependent Ca2+ currents in rat dissociated supraoptic neurones via A1 receptors. J Physiol 526 Pt 2: 313-326
149. Nyce, J. W. 1999. Insight into adenosine receptor function using antisense and gene-knockout approaches. Trends Pharmacol Sci 20(2): 79-83
150. Ochiishi, T., Takita, M., Ikemoto, M., Nakata, H., and Suzuki, S. S. 1999. Immunohistochemical analysis on the role of adenosine A1 receptors in epilepsy. Neuroreport 10(17): 3535-3541
151. Ohana, G., Bar-Yehuda, S., Barer, F., and Fishman, P. 2001. Differential effect of adenosine on tumor and normal cell growth: focus on the A3 adenosine receptor. J.Cell Physiol 186(1): 19-23
152. Olah, M. E. and Stiles, G. L. 1995. Adenosine receptor subtypes: characterization and therapeutic regulation. Annu Rev Pharmacol Toxicol 35: 581-606
153. Olah, M. E. and Stiles, G. L. 2000. The role of receptor structure in determining adenosine receptor activity. Pharmacol Ther 85(2): 55-75
154. Olsson, R. A. and Pearson, J. D. 1990. Cardiovascular purinoceptors. Physiol Rev. 70(3): 761-845155. Ongini, E. and Fredholm, B. B. 1996. Pharmacology of adenosine A2A receptors. Trends Pharmacol.Sci.
17(10): 364-372156. Pagonopoulou, O. and Angelatou, F. 1992. Reduction of A1 adenosine receptors in cortex,
hippocampus and cerebellum in ageing mouse brain. Neuroreport 3(9): 735-737157. Palmer, T. M. and Stiles, G. L. 1995. Adenosine receptors. Neuropharmacology 34(7): 683-694158. Papassotiropoulos, A., Hock, C., and Nitsch, R. M. 2001. Genetics of interleukin 6: implications for
Alzheimer's disease. Neurobiol.Aging 22(6): 863-871159. Park, K. S., Jeong, S. W., Cha, S. K., Lee, B. S., Kong, I. D., Ikeda, S. R., and Lee, J. W. 2001.
Modulation of N-type Ca2+ currents by A1-adenosine receptor activation in male rat pelvic ganglion neurons. J Pharmacol Exp Ther 299(2): 501-508
160. Pavenstadt, H., Ruh, J., Greger, R., and Schollmeyer, P. 1994. Adenosine-induced hyperpolarization of the membrane voltage in rat mesangial cells in primary culture. Br J Pharmacol 113(1): 7-12
161. Peltola, J., Hurme, M., Miettinen, A., and Keranen, T. 1998. Elevated levels of interleukin-6 may occur in cerebrospinal fluid from patients with recent epileptic seizures. Epilepsy Res. 31(2): 129-133
162. Penkowa, M., Giralt, M., Carrasco, J., Hadberg, H., and Hidalgo, J. 2000. Impaired inflammatory response and increased oxidative stress and neurodegeneration after brain injury in interleukin-6-deficient mice. Glia 32(3): 271-285
163. Penkowa, M., Molinero, A., Carrasco, J., and Hidalgo, J. 2001. Interleukin-6 deficiency reduces the brain inflammatory response and increases oxidative stress and neurodegeneration after kainic acid-induced seizures. Neuroscience 102(4): 805-18
164. Phillis, J. W. 1989. Adenosine in the control of the cerebral circulation. Cerebrovasc.Brain Metab Rev. 1(1): 26-54
165. Phillis, J. W. and O'Regan, M. H. 1989. Deoxycoformycin antagonizes ischemia-induced neuronal degeneration. Brain Res.Bull. 22(3): 537-540
166. Poon, A. and Sawynok, J. 1998. Antinociception by adenosine analogs and inhibitors of adenosine metabolism in an inflammatory thermal hyperalgesia model in the rat. Pain 74(2-3): 235-245
167. Popoli, P., Pintor, A., Domenici, M. R., Frank, C., Tebano, M. T., Pezzola, A., Scarchilli, L., Quarta, D., Reggio, R., Malchiodi-Albedi, F., Falchi, M., and Massotti, M. 2002. Blockade of striatal adenosine A2A receptor reduces, through a presynaptic mechanism, quinolinic acid-induced excitotoxicity: possible relevance to neuroprotective interventions in neurodegenerative diseases of the striatum. J Neurosci 22(5): 1967-1975

Introduction
35
168. Porkka-Heiskanen, T., Alanko, L., Kalinchuk, A., and Stenberg, D. 2002. Adenosine and sleep. Sleep Med.Rev. 6(4): 321-332
169. Porkka-Heiskanen, T., Strecker, R. E., Thakkar, M., Bjorkum, A. A., Greene, R. W., and McCarley, R. W. 1997. Adenosine: a mediator of the sleep-inducing effects of prolonged wakefulness. Science276(5316): 1265-1268
170. Ralevic, V. and Burnstock, G. 1998. Receptors for purines and pyrimidines. Pharmacol.Rev. 50(3): 413-492
171. Ramkumar, V., Stiles, G. L., Beaven, M. A., and Ali, H. 1993. The A3 adenosine receptor is the unique adenosine receptor which facilitates release of allergic mediators in mast cells. J Biol.Chem. 268(23): 16887-16890
172. Rathbone, M. P., Christjanson, L., Deforge, S., Deluca, B., Gysbers, J. W., Hindley, S., Jovetich, M., Middlemiss, P., and Takhal, S. 1992. Extracellular purine nucleosides stimulate cell division and morphogenesis: pathological and physiological implications. Med.Hypotheses 37(4): 232-240
173. Rathbone, M. P., Deforge, S., Deluca, B., Gabel, B., Laurenssen, C., Middlemiss, P., and Parkinson, S. 1992. Purinergic stimulation of cell division and differentiation: mechanisms and pharmacologicalimplications. Med.Hypotheses 37(4): 213-219
174. Rathbone, M. P., Middlemiss, P. J., Gysbers, J., Diamond, J., Holmes, M., Pertens, E., Juurlink, B. H., Glasky, A., Ritzman, R., Glasky, M., Crocker, C. E., Ramirez, J. J., Lorenzen, A., Fein, T., Schulze, E., Schwabe, U., Ciccarelli, R., Di Iorio, P., and Caciagli, F. 1998. Physiology and Pharmacology of Natural and Synthetic Nonadenine-Based Purines in the Nervous System. Drug development research 45: 356-372
175. Ribeiro, J. A., Sebastiao, A. M., and de Mendonca, A. 2002. Adenosine receptors in the nervous system: pathophysiological implications. Prog.Neurobiol. 68(6): 377-392
176. Roach, A. C. 1991. Adenosine-Adenocard: a new intravenous antiarrhythmic agent for supraventricular tachycardia. Crit Care Nurse 11(7): 78-79
177. Roman, R. M. and Fitz, J. G. 1999. Emerging roles of purinergic signaling in gastrointestinal epithelial secretion and hepatobiliary function. Gastroenterology 116(4): 964-979
178. Rudolphi, K. A. and Schubert, P. 1996. Purinergic interventions in traumatic and ischemic injury. InNovel Thearapies for CNS Injuries, Peterson, P. L. and Willis, P. W. 327-342. Boca Raton, New York, London, Tokyo: CRC Press Inc.
179. Rudolphi, K. A. and Schubert, P. 1997. Modulation of neuronal and glial cell function by adenosine and neuroprotection in vascular dementia. Behav Brain Res 83(1-2): 123-128
180. Rudolphi, K. A., Schubert, P., Parkinson, F. E., and Fredholm, B. B. 1992. Adenosine and brain ischemia. Cerebrovasc.Brain Metab Rev. 4(4): 346-369
181. Rudolphi, K. A., Schubert, P., Parkinson, F. E., and Fredholm, B. B. 1992. Neuroprotective role of adenosine in cerebral ischaemia. Trends Pharmacol Sci 13(12): 439-445
182. Sallmann, S., Juttler, E., Prinz, S., Petersen, N., Knopf, U., Weiser, T., and Schwaninger, M. 2000. Induction of interleukin-6 by depolarization of neurons. J Neurosci. 20(23): 8637-8642
183. Sandoli, D., Chiu, P. J., Chintala, M., Dionisotti, S., and Ongini, E. 1994. In vivo and ex vivo effects of adenosine A1 and A2 receptor agonists on platelet aggregation in the rabbit. Eur J Pharmacol. 259(1): 43-49
184. Sawynok, J. and Liu, X. J. 2003. Adenosine in the spinal cord and periphery: release and regulation of pain. Prog.Neurobiol. 69(5): 313-340
185. Sawynok, J., Zarrindast, M. R., Reid, A. R., and Doak, G. J. 1997. Adenosine A3 receptor activation produces nociceptive behaviour and edema by release of histamine and 5-hydroxytryptamine. Eur J Pharmacol. 333(1): 1-7
186. Schimmel, R. J. and McCarthy, L. 1984. Role of adenosine as an endogenous regulator of respirationin hamster brown adipocytes. Am.J Physiol 246(3 Pt 1): C301-C307
187. Schobitz, B., de Kloet, E. R., Sutanto, W., and Holsboer, F. 1993. Cellular localization of interleukin 6 mRNA and interleukin 6 receptor mRNA in rat brain. Eur J Neurosci. 5(11): 1426-1435
188. Schrier, S. M., van Tilburg, E. W., van der, Meulen H., IJzerman, A. P., Mulder, G. J., and Nagelkerke,J. F. 2001. Extracellular adenosine-induced apoptosis in mouse neuroblastoma cells: studies on involvement of adenosine receptors and adenosine uptake. Biochem.Pharmacol. 61(4): 417-425

Chapter 1
36
189. Schulte, G. and Fredholm, B. B. 2003. Signalling from adenosine receptors to mitogen-activated protein kinases. Cell Signal. 15(9): 813-827
190. Schumann, G., Huell, M., Machein, U., Hocke, G., and Fiebich, B. L. 1999. Interleukin-6 activates signal transducer and activator of transcription and mitogen-activated protein kinase signal transduction pathways and induces de novo protein synthesis in human neuronal cells. J Neurochem. 73(5): 2009-2017
191. Schwaninger, M., Neher, M., Viegas, E., Schneider, A., and Spranger, M. 1997. Stimulation of interleukin-6 secretion and gene transcription in primary astrocytes by adenosine. J Neurochem69(3): 1145-1150
192. Schwarzschild, M. A., Chen, J. F., and Ascherio, A. 2002. Caffeinated clues and the promise of adenosine A(2A) antagonists in PD. Neurology 58(8): 1154-1160
193. Shin, H. K., Shin, Y. W., and Hong, K. W. 2000. Role of adenosine A(2B) receptors in vasodilation of rat pial artery and cerebral blood flow autoregulation. Am J Physiol Heart Circ Physiol 278(2):339-344
194. Shinoda, I., Furukawa, Y., and Furukawa, S. 1990. Stimulation of nerve growth factor synthesis/secretion by propentofylline in cultured mouse astroglial cells. Biochem.Pharmacol. 39(11): 1813-1816
195. Song, W. J., Tkatch, T., and Surmeier, D. J. 2000. Adenosine receptor expression and modulation of Ca(2+) channels in rat striatal cholinergic interneurons. J Neurophysiol 83(1): 322-332
196. Sperlagh, B., Zsilla, G., Baranyi, M., Kekes-Szabo, A., and Vizi, E. S. 1997. Age-dependent changes of presynaptic neuromodulation via A1-adenosine receptors in rat hippocampal slices. Int.J Dev.Neurosci. 15(6): 739-747
197. Spicuzza, L., Bonfiglio, C., and Polosa, R. 2003. Research applications and implications of adenosinein diseased airways. Trends Pharmacol.Sci. 24(8): 409-413
198. Spyer, K. M. and Thomas, T. 2000. A role for adenosine in modulating cardio-respiratory responses: a mini-review. Brain Res.Bull. 53(1): 121-124
199. Stence, N., Waite, M., and Dailey, M. E. 2001. Dynamics of microglial activation: a confocal time-lapse analysis in hippocampal slices. Glia 33(3): 256-266
200. Stevens, B., Porta, S., Haak, L. L., Gallo, V., and Fields, R. D. 2002. Adenosine: a neuron-glial trans-mitter promoting myelination in the CNS in response to action potentials. Neuron 36(5): 855-868
201. Stiles, G. L. 1992. Adenosine receptors. J.Biol.Chem. 267(10): 6451-6454202. Stone, T. W. 2002. Purines and neuroprotection. Adv.Exp.Med.Biol. 513: 249-280203. Stone, T. W., Fredholm, B. B., and Phillis, J. W. 1989. Adenosine and morphine. Trends
Pharmacol.Sci. 10(8): 316204. Straub, R. H., Pongratz, G., Gunzler, C., Michna, A., Baier, S., Kees, F., Falk, W., and Scholmerich, J.
2002. Immunoregulation of IL-6 secretion by endogenous and exogenous adenosine and by exogenous purinergic agonists in splenic tissue slices. J Neuroimmunol 125(1-2): 73-81
205. Streit, W. J. 2002. Microglia as neuroprotective, immunocompetent cells of the CNS. Glia 40(2): 133-139
206. Streit, W. J., Walter, S. A., and Pennell, N. A. 1999. Reactive microgliosis. Prog.Neurobiol. 57(6): 563-581
207. Swartz, K. R., Liu, F., Sewell, D., Schochet, T., Campbell, I., Sandor, M., and Fabry, Z. 2001. Inter-leukin-6 promotes post-traumatic healing in the central nervous system. Brain Res 896(1-2): 86-95
208. Tabrizchi, R. and Bedi, S. 2001. Pharmacology of adenosine receptors in the vasculature. Pharmacol Ther 91(2): 133-147
209. Taga, T. and Kishimoto, T. 1997. Gp130 and the interleukin-6 family of cytokines. Annu.Rev.Immunol. 15: 797-819
210. Tanaka, E., Yasumoto, S., Hattori, G., Niiyama, S., Matsuyama, S., and Higashi, H. 2001. Mechanisms underlying the depression of evoked fast EPSCs following in vitro ischemia in rat hippocampal CA1 neurons. J Neurophysiol 86(3): 1095-1103
211. Tatlisumak, T., Takano, K., Carano, R. A., Miller, L. P., Foster, A. C., and Fisher, M. 1998. Delayed treatment with an adenosine kinase inhibitor, GP683, attenuates infarct size in rats with temporary middle cerebral artery occlusion. Stroke 29(9): 1952-1958

Introduction
37
212. Tatsis-Kotsidis, I. and Erlanger, B. F. 1999. A1 adenosine receptor of human and mouse adipose tissues: cloning, expression, and characterization. Biochem Pharmacol 58(8): 1269-1277
213. Trincavelli, M. L., Tuscano, D., Marroni, M., Falleni, A., Gremigni, V., Ceruti, S., Abbracchio, M. P., Jacobson, K. A., Cattabeni, F., and Martini, C. 2002. A3 adenosine receptors in human astrocytoma cells: agonist-mediated desensitization, internalization, and down-regulation. Mol.Pharmacol. 62(6): 1373-1384
214. Trussell, L. O. and Jackson, M. B. 1985. Adenosine-activated potassium conductance in cultured striatal neurons. Proc Natl Acad Sci U S A 82(14): 4857-4861
215. Tsacopoulos, M. and Magistretti, P. J. 1996. Metabolic coupling between glia and neurons.J Neurosci.16(3): 877-885
216. Tsutsui, S., Schnermann, J., Noorbakhsh, F., Henry, S., Yong, V. W., Winston, B. W., Warren, K., and Power, C. 2004. A1 adenosine receptor upregulation and activation attenuates neuroinflammation and demyelination in a model of multiple sclerosis. J Neurosci. 24(6): 1521-1529
217. Turcani, P. and Tureani, M. 2001. Effect of propentofylline on cerebral blood flow in a gerbil focal cerebral ischemia. J Neurol Sci 183(1): 57-60
218. van Calker, D., Muller, M., and Hamprecht, B. 1978. Adenosine inhibits the accumulation of cyclic AMP in cultured brain cells. Nature 276(5690): 839-841
219. van Calker, D., Muller, M., and Hamprecht, B. 1979. Adenosine regulates via two different types of receptors, the accumulation of cyclic AMP in cultured brain cells. J.Neurochem. 33(5): 999-1005
220. Van Wagoner, N. J. and Benveniste, E. N. 1999. Interleukin-6 expression and regulation in astrocytes. J Neuroimmunol 100(1-2): 124-139
221. Vanore, G., Giraldez, L., Rodriguez de Lores, Arnaiz G., and Girardi, E. 2001. Seizure activityproduces differential changes in adenosine A1 receptors within rat hippocampus. Neurochem.Res.26(3): 225-230
222. Vollenweider, F., Herrmann, M., Otten, U., and Nitsch, C. 2003. Interleukin-6 receptor expression and localization after transient global ischemia in gerbil hippocampus. Neurosci.Lett. 341(1): 49-52
223. von Lubitz, D. K., Lin, R. C., Bischofberger, N., Beenhakker, M., Boyd, M., Lipartowska, R., and Jacobson, K. A. 1999. Protection against ischemic damage by adenosine amine congener, a potent and selective adenosine A1 receptor agonist. Eur J Pharmacol 369(3): 313-317
224. von Lubitz, D. K., Lin, R. C., Boyd, M., Bischofberger, N., and Jacobson, K. A. 1999. Chronic administration of adenosine A3 receptor agonist and cerebral ischemia: neuronal and glial effects. Eur J Pharmacol 367(2-3): 157-163
225. von Lubitz, D. K., Lin, R. C., Paul, I. A., Beenhakker, M., Boyd, M., Bischofberger, N., and Jacobson, K. A. 1996. Postischemic administration of adenosine amine congener (ADAC): analysis of recoveryin gerbils. Eur J Pharmacol. 316(2-3): 171-179
226. von Lubitz, D. K., Lin, R. C., Popik, P., Carter, M. F., and Jacobson, K. A. 1994. Adenosine A3
receptor stimulation and cerebral ischemia. Eur.J Pharmacol. 263(1-2): 59-67227. von Lubitz, D. K., Simpson, K. L., and Lin, R. C. 2001. Right thing at a wrong time? Adenosine A3
receptors and cerebroprotection in stroke. Ann N Y Acad Sci 939: 85-96228. von Lubitz, D. K., Ye, W., McClellan, J., and Lin, R. C. 1999. Stimulation of adenosine A3 receptors
in cerebral ischemia. Neuronal death, recovery, or both? Ann N Y Acad Sci 890: 93-106229. Wardas, J., Konieczny, J., and Lorenc-Koci, E. 2001. SCH 58261, an A(2A) adenosine receptor
antagonist, counteracts parkinsonian-like muscle rigidity in rats. Synapse 41(2): 160-171230. Williams, M. and Jarvis, M. F. 2000. Purinergic and pyrimidinergic receptors as potential drug
targets. Biochem Pharmacol 59(10): 1173-1185231. Williams, Michael. 1989. Adenosine: the prototypic neuromodulator. Neurochemistry international
14(3): 249-264232. Yamada, M. and Hatanaka, H. 1994. Interleukin-6 protects cultured rat hippocampal neurons
against glutamate-induced cell death. Brain Res. 643(1-2): 173-180233. Ye, S. M. and Johnson, R. W. 1999. Increased interleukin-6 expression by microglia from brain of
aged mice. J Neuroimmunol 93(1-2): 139-148

Chapter 1
38
234. Zhang, Y., Hayes, A., Pritchard, A., Thaker, U., Haque, M. S., Lemmon, H., Harris, J., Cumming, A., Lambert, J. C., Chartier-Harlin, M. C., St Clair, D., Iwatsubo, T., Mann, D. M., and Lendon, C. L. 2004. Interleukin-6 promoter polymorphism: risk and pathology of Alzheimer's disease. Neurosci.Lett. 362(2): 99-102
235. Zhao, Z., Makaritsis, K., Francis, C. E., Gavras, H., and Ravid, K. 2000. A role for the A3 adenosine receptor in determining tissue levels of cAMP and blood pressure: studies in knock-out mice. Biochim Biophys Acta 1500(3): 280-290
236. Zhong, H., Shlykov, S. G., Molina, J. G., Sanborn, B. M., Jacobson, M. A., Tilley, S. L., and Blackburn, M. R. 2003. Activation of murine lung mast cells by the adenosine A3 receptor. J Immunol. 171(1): 338-345
237. Zimmermann, H. and Braun, N. 1996. Extracellular metabolism of nucleotides in the nervous system. J Auton.Pharmacol. 16(6): 397-400

39
Submitted
The role of glia cellsin purine receptor-induced neuroprotection
M.C. Wittendorp, H.W.G.M. Boddeke and K. Biber
Chapter 2

Summary
Introduction
Purines have been divided into two classes: guanine-based and adenine-basedpurines. Whereas there is functional evidence for the existence of receptors forthe guanine-based nucleotide GTP and its nucleoside guanosine; these receptorsstill await their discovery [21, 126]. In contrast, a large variety of receptors foradenine-based purines has been described and cloned. Thus P1 or adenosinereceptors are activated by adenosine and P2 receptors are activated by the adeninenucleotides ATP and ADP (for excellent review see: [18, 42, 81]). Adenosine binds and activates four subtypes of adenosine or P1 receptors that arenamed adenosine A1, A2A, A2B and A3 receptors. These receptors all belong to thefamily of 7 trans-membrane receptors (TM7) that predominantly signal throughactivation of G-proteins [42]. Although adenosine A1 and A3 receptors wereoriginally classified to couple to Gi/o proteins and the adenosine A2 receptors toGs proteins, it is today clear that this is not exclusively the case [42]. Adenosinereceptors have been cloned from various species and their pharmacological profileis based on a variety of specific agonists and antagonists [42, 63]. P2 receptorsexist as two families: the P2Y receptors that belong to the TM7 receptors and theP2X receptors that are ligand-gated ion-channels. Currently 11 members of theP2Y receptor family have been cloned of which six are molecular and functionallycharacterized (P2Y1, P2Y2, P2Y4, P2Y6, P2Y11 and P2Y12). From the eight clonedmembers of the P2X family are functional responses known from seven members(P2X1-7) (for review see: [18, 58]). P2 receptors are not only activated by the
Chapter 2
40
Adenosine and its nucleotides (purines) are structurally closely related, can be produced ondemand and released to the extracellular environment. Today purines are regarded as thephylogenetically most ancient epigenetic factors for cellular control. Purines act via P1(adenosine) and P2 (ATP) receptor subtypes and have many important functions in the centralnervous system. In neurons adenosine is known for its function as a neuromodulator, whereasATP acts as a transmitter with profound effects in fast neurotransmission. Since purines aremassively released under pathological conditions in the CNS, it is generally assumed that purine-signaling in brain is aimed to protect neurons. Purine receptors are not only found in neurons, they are also abundantly expressed in glia cells(astrocytes and microglia). Apart from numerous purinergic effects on glial physiology, there isincreasing evidence that purine receptor signaling significantly contributes to the control of glialactivity under pathological conditions in the CNS. Purines control the proliferation of glia cellsand stimulation of glial purine receptors leads to synthesis and release of various neuroprotectivefactors. These findings suggest that, in addition to their direct effects on neurons, purines mayalso have indirect neuroprotective functions by activating glia cells. We here thus review thefunction of purines in neuroprotection with a main emphasis in purine-signaling in glia cells.

purine nucleotides (ATP and ADP) but also by the pyrimidine nucleotides (UTPand UDP) and their pharmacological characterization is based on the rank-orderof potency of the various agonists. Selective and potent antagonists for P2receptors, however are still lacking [58, 81].Adenosine and its nucleotides are structurally closely related and are regarded asthe phylogenetically most ancient epigenetic factors for cellular control. Theyshare a metabolic route, are both produced on demand and can be released to theextracellular environment. Adenosine and its nucleotides are present in all celltypes, making the broad and abundant expression of P1 and P2 receptors in alltissues conceivable [18, 58]. Since these molecules and their receptors areinvolved in both numerous physiological processes and disease pathologies thereare various therapeutic concepts and approaches that are based on purinesignaling [18, 42, 58, 136]. Activation of purine receptors leads to a variety of effects in the central nervoussystem [18, 24, 41]. Adenosine is well known for its role as neuromodulatorunder physiological conditions and ATP plays a role in fast neurotransmission.Under pathological conditions (e.g. ischaemia, trauma, and excitotoxicity), highconcentrations of adenosine and/or ATP are released into the extracellular spaceas a result of an imbalance in energy metabolism. High levels of extracellularadenosine protect neurons from excitatory damage by hyperpolarization andinhibition of neuronal transmitter release (see below). Although mostextracellular ATP is rapidly metabolized into adenosine, there is recent evidencethat ATP itself may influence the survival of neurons [131]. Brain damage in general also activates glia cells, mostly microglia and astrocytes.The prominent presence of activated glia cells in the injured brain has ledapproximately 10 years ago to the assumption that glial activation might be a keyfeature in neurodegenerative processes [73, 80, 85]. This assumption has nowbeen refined. Clear evidence that activated glia cells support damaged neuronsand promote survival has been achieved in the last couple of years [15]. Thesedata fit more in the general and old concept of glia as neuron supporting cells[119]. Thus glia activation per se is not detrimental; this probably is only the casewhen glia activation is very strong and prolonged as observed in chronicneurodegeneration. The control of glia activity is therefore crucial for the finaloutcome. Like other cells, glia cells express a variety of purine receptors. Thesereceptors are involved in several aspects of glial physiology such as control of ion-channel expression, induction of intracellular calcium transients and regulation ofproliferation and apoptosis [39, 54, 57, 67, 89]. Here we will review the currentliterature on purinergic effects in the CNS with a main emphasis on glia cells andtheir contribution to neuroprotection.
Glia cells in purine receptor-induced neuroprotection
41

Effects of purines in neurons
Expression of all types of adenosine receptors has been found in neurons.Adenosine A1 receptors are highly expressed especially in cortex, thalamus, cere-bellum and hippocampus [88, 122]. Adenosine A2A receptors are predominantlyfound in the striatum, where they are co-localized with dopamine D2 receptors.Moreover, A2A receptors are abundantly expressed in the nucleus accumbens andin the olfactory tubercle [33, 78]. The expression of the other two adenosinereceptor subtypes, A2B and A3, is not restricted to specific brain areas [36, 132]. Under physiological conditions adenosine has several prominent effects in thebrain. First adenosine is a key modulator of energy homeostasis, a function whichis common to all tissues [23, 137]. Moreover, adenosine in the brain plays aneuromodulatory role at the synaptic level, since it modulates the release ofneurotransmitter by a balanced activation of presynaptic inhibitory A1 receptorsand facilitatory A2A receptors [23, 107]. In addition, it has been suggested thatadenosine mediates direct trophic functions since it stimulates neurite outgrowth,through activation of adenosine A2A receptors [20, 96].Like in other tissues, adenosine is also in the brain essentially involved in thecoupling of energy consumption and energy supply. In order to maintain a stablecellular energy content, adenosine can suppress neuronal firing (reduce energyconsumption) and increase the cerebral blood flow (support energy supply) [28,50]. These effects are induced by the large amounts of adenosine that are releasedunder pathological conditions like seizures and ischaemia. Under these conditionsthe extracellular concentration of adenosine rapidly rises from nanomolar tomicromolar levels [31, 99], which subsequently leads to a significant protectionof neurons [25, 41, 99, 101, 133, 138]. Since the neuroprotective mechanisms ofneuronal adenosine A1 receptors have been reviewed extensively in a number ofexcellent articles they will only be discussed briefly here [31, 32, 50, 99].Presynaptically, adenosine inhibits the release of the excitatory neurotransmittersglutamate [31, 34]. Postsynaptically, adenosine causes hyperpolarization of thecell membrane, mainly by increasing K+ conductance [46, 94, 128], bystabilization of the Mg2+ blockade of NMDA receptors [99, 101] and by areduction of Ca2+-influx, probably caused by an inhibition of N-type voltage-dependent Ca2+ channels [29, 71, 86, 92, 114, 125]. Thus the sum of theseeffects will finally lead to a decrease of neuronal excitability, thereby preventingneurons from death caused by overstimulation, called excitotoxicity. The neuro-protective effects of adenosine A1 receptors are increased following receptorupregulation, suggesting that an enhancement of neuronal adenosine A1 receptorexpression attenuates neuronal damage in pathological conditions [99]. Investigations that studied the expression of P2 receptor mRNAs found abundantand ubiquitous expression of P2X1, P2X2, P2X4 and P2X6 mRNA in the brain,
Chapter 2
42

whereas other P2X mRNA were more restricted to special brain regions [24].Furthermore, the expression of P2Y1, P2Y2, P2Y4 and P2Y6 mRNA has beenobserved in brain tissue without further characterization of its localization [24].Protein studies, however, did not always corroborate the findings on mRNAexpression, which makes the issue of P2 receptor expression in brain a field forfurther investigation. Based on the work of G. Burnstock (1970) who was the firstto postulate a signaling function for nucleotides, it is widely accepted today thatnucleotides such as ATP or UTP are extracellular signaling molecules [17]. Theyact in the brain as neurotransmitter or co-transmitter with various effects on fastneurotransmission, dependent on the nucleotide and receptor subtype involved[18, 24, 81, 136]. Despite the fact that much progress has been made, the effectsof P2 receptors in neurons are still less well described than those of adenosinereceptors [70, 116], which is mostly due to the lack of selective ligands for mostP2 receptor subtypes [81].
Effects of purines in glia cells
Purine receptor expression in glia cells
Adenosine receptors are also present in glia cells [21]. The expression of alladenosine receptor subtypes has been described in both rat and mouse astrocytes[14, 36, 78]. Whereas consistent results have been published on the expressionof adenosine A1 and A3 receptors in cultured microglia, conflicting data exist onthe expression of adenosine A2A and A2B receptors in these cells. This differencemay be due to species differences [38, 51, 84, 132, 140].
Both astrocytes and microglia express P2X and P2Y receptor subtypes [84]. Theexpression of P2X receptors in hippocampal astrocytes was recently described in adetailed study showing that P2X4 and P2X6 were abundantly expressed in thesecells. Moderate expression of P2X1, P2X2, P2X3 and P2X7 was found inastrocytes, whereas astrocytic P2X5 protein was not detected [65]. Similar resultswere observed in cultured cortical astrocytes with the exception that the authorsdid not find expression of P2X6 but detected P2X5 expression [43]. RT-PCRanalysis and functional studies have also provided evidence for the expression ofseveral P2Y receptors (P2Y1, P2Y2, P2Y4, P2Y6 and P2Y12) in cultured corticalastrocytes [8, 43]. The situation concerning expression of P2 receptors inmicroglia is less complex. A variety of functional studies as well asimmunohistochemical evidence indicates that microglia express P2Y7, P2Y2 andP2Y12 (see for review: [57]). The expression of P2Y12 seems to be unique formicroglia, as compared to other macrophage-like cell types and could thereforeaccount as a microglia specific marker in immunological studies [103].
Glia cells in purine receptor-induced neuroprotection
43

Purinergic control of glia cell proliferation
Microglia and astrocytes rapidly respond to any pathological event in the CNS.The activation of these cells (gliosis) which inevitably occurs whenever neuronsare endangered is most likely aimed to reduce the damage and to protectneuronal cells [119]. However, highly activated, overreacting glia cells mightinduce a so-called bystander reaction that can lead to the degradation of healthytissue, thereby increasing the damage. It is therefore of particular interest tounderstand how the “double-edged sword” of glial activation is balanced, how theactivity of glia cells is controlled. Since ATP and adenosine are rapidly releasedunder pathological conditions and glia cells functionally express purine receptors,various groups have investigated the possible involvement of purines in thecontrol of glial activity.In this respect, it has been reported that adenosine as well as ATP can activatemicroglia and increase microglia proliferation [38, 45, 57], while others havefound that adenosine and ATP inhibit microglial proliferation and activation oreven induce apoptosis in these cells [13, 89, 112, 141]. These, apparentlycontradictory data, might be related to the different activation stages of microglia.It is known that activated microglia can act as brain macrophages and releaseneurotoxic substances, thereby increasing neuronal damage [3, 4, 120]. On theother hand increasing evidence suggests that activated microglia provide favorableconditions for neuronal repair since it has been shown that activated microgliahave direct neuroprotective effects or that they can trigger T-cells to mount aneuroprotective response [108, 119].Similarly astrocytes undergo profound functional changes in response to CNSinjury. This reaction, called reactive astrogliosis is characterized by increasedastrocyte proliferation, astrocyte hypertrophy and an increased expression of glialfibrillary acidic protein [74, 87]. Since reactive astrogliosis has been observed forexample in the aging brain or Alzheimer’s disease [129] it has been linked to thepathogenesis of these conditions and it has been assumed that astrogliosis wouldimpair neuronal regeneration in the CNS. However, there are some indicationsthat reactive gliosis may also be beneficial for neuronal repair. In addition,reactive astrocytes could actually support neural outgrowth [1, 84, 97, 135].Similar to the onset of microglia activity, several reports have shown that purinescontribute to the onset of astrogliosis, whereas blockade of adenosine receptorswas found to prevent this reaction [2, 14, 54, 62]. Accordingly, it has beensuggested that adenosine, by stimulating the proliferation of astrocytes wouldenhances their protective effects, activities such as the clearance of neurotoxiclevels of glutamate from the extracellular space [100]. On the other handadenosine has recently been described to induce apoptosis of astrocytes viaadenosine A3 receptors, indicating again a complex function of purines in theregulation of astroglial activity [26].
Chapter 2
44

Thus whereas the molecular identity of the involved purine receptor subtypes isnot always known, there are contradictory purinergic effects on the proliferationof both glia cell types, microglia and astrocytes [21]. As mentioned aboveproliferation of microglia and astrocytes may have neuroprotective or “neuro-detrimental” effects. It is thus tempting to speculate that purines may stimulatethe protective side of glial proliferation whereas the destructive side of glialproliferation is inhibited by purine signaling. Further experiments are clearlyneeded to investigate whether this hypothesis is true.
Purinergic control of glial secretion of neuroprotective substances
The release of various neurotrophic factors from astrocytes during developmenthas long been known [82]. Various reports have recently provided additionalevidence for a neuron-supportive role of astrocytes and microglia during CNSpathology. These cells are now known to release various factors aimed to protectneurons from the threats of damage. Among these factors are neurotrophicfactors and cytokines [82]. Neurotrophic factors constitute a large family ofrelated proteins such as nerve growth factor (NGF), neurotrophins 3 and 4 (NT3,NT4), brain-derived-neurotrophic factor (BDNF), glial-derived neurotrophicfactor (GDNF) and vascular endothelial growth factor (VEGF) [60, 124, 134].Next to these factors a number of glial derived cytokines have also been describedto have neuroprotective properties; IL-1α, IL-1β, IL-6, IL-10, TNF-α, TGF-β andthe chemokine CCL2 [12, 19, 115, 118, 121]. How the synthesis and release ofthese factors from glia cells is controlled has not been completely understood yet,but experimental evidence strongly suggests that glial purine receptors areimportant regulators here.
Release of neurotrophic factors from glia cells
The activity of nerve growth factor was initially associated with the differentiationand survival of young neurons. NGF is, however, also involved in maintenance ofmature neurons and neuronal repair. NGF exerts its neurotrophic effects throughseveral second messenger pathways, via the activation of tyrokinase A (TrkA)receptors. The essential mechanism involved in NGF-induced neuroprotection isbased on intervention of the apoptotic pathways. For an excellent review on NGFfunction see [113].Cultured mouse astrocytes have been shown to release NGF after stimulation ofadenosine A1 receptors [22], treatment with propentofylline (adenosine reuptakeinhibitor) [111] or with guanosine and GTP [77]. In the same line of evidence itwas described that exposure to AIT-082, a hypoxanthine derivative, stimulatedthe production of mRNA for NGF, neurotrophin-3 and basic fibroblast growthfactor (bFGF) in cultured mouse astrocytes [47, 96] as well as in vivo [48]. Themechanism underlying the effects induced by AIT-082 is not yet fully explained,
Glia cells in purine receptor-induced neuroprotection
45

but it has been suggested that AIT-082 might enhance the release of adenosinefrom astrocytes [27]. An effect of adenosine receptor stimulation on the release ofNGF was also found in microglia, but in contrast to astrocytes this effect was dueto the stimulation of adenosine A2A receptors [52].
S100BS100 proteins are small calcium- binding proteins that play a role in manyintracellular functions like protein phosphorylation, enzyme activities, cellproliferation and differentiation and intracellular calcium homeostasis [30].Unlike most other S100 proteins, S100B is secreted by cells, mainly astrocytes,and promotes of neuronal survival and neurite outgrowth [5, 79, 104]. Extra-cellular S100B exerts these effects probably by binding to the receptor foradvanced glycation end products (RAGE), nuclear translocation of NF-κB andstimulation of Bcl-2 expression [30]. The release of S100B from astrocytes is,among other receptors, induced by stimulation with adenosine A1 receptors [21,22].
CytokinesAlthough cytokines were believed to be solely involved in regulating immunefunction in the periphery and inflammatory responses in the nervous system, it isnow obvious that these molecules also have potent neuroprotective effects [19,59, 117]. The pro-inflammatory cytokine IL-6 was initially believed to induceprimarily destructive effects in the brain. Since expression of IL-6 in brain isincreased during brain pathologies like seizures, Alzheimer’s disease, as well asnormal aging, it was believed that IL-6 would play a causal role in the patho-physiology of these conditions [7, 44, 49, 130]. However, increasing evidence hasshown that IL-6 is neuroprotective, plays a role in differentiation andregeneration of nervous tissue and is essential for maintenance of homeostasis inthe CNS. Therefore, the higher levels of IL-6 observed under pathologicalconditions may also indicate an intrinsic response to counteract brain damage[44, 59]. Several reports clearly show the beneficial effects of IL-6 in differentmodels for neurotoxicity [9, 66, 69]. Moreover, IL-6-deficient mice depict acompromised inflammatory response in the CNS, an increase of oxidative stressand a delayed post-traumatic healing after brain injury [95, 123]. Next to otherfactors that stimulate the release of IL-6 from glia cells, purine receptors areinvolved. Stimulation of adenosine A2B receptors led to an increase of IL-6 mRNAand IL-6 protein synthesis in human astroglioma cells and cultured mouseastrocytes [37, 106]. Similar effects were observed in the microglia cell line MG5.However, in contrast to astrocytes, stimulation of adenosine A2B receptors had noeffect in MG5 cells, the fast IL-6 release here was induced by stimulation with ATPvia pertussis-toxin insensitive P2Y receptors [90, 110]. The question, which P2Y-
Chapter 2
46

subtype in microglia mediates this effects, however, remains to be answered [57]. Although the neuroprotective actions of IL-6 are now well established, theunderlying mechanisms remain to be elucidated. The effects of IL-6 on NGFproduction and enhancement of NGF-mediated effects most likely contribute tothe neuroprotective effects of the cytokine [19, 64, 72]. Based on our data,however, we propose an additional, purine-receptor-based mechanism. In 1999Schumann and colleagues found a marked increase of de novo protein synthesisin human neuronal cells after stimulation with IL-6 [105]. We could recentlydemonstrate that adenosine A1 receptors are among the newly synthesizedproteins in nervous tissue in response to IL-6 stimulation [10]. Since it is welldescribed that upregulation of adenosine A1 receptors in brain enhances theneuroprotective effects of adenosine, it has been proposed that the indirect effectof IL-6 on adenosine A1 receptors might be essential for the neuroprotectivefunction of IL-6 [10]. A clear dual role has now been acknowledged for TNF-α in brain [98]. Itsexpression can induce further damage to brain tissue during ischemia [6, 75] orprotect neurons from various insults [61, 83, 109]. An explanation for theseconflicting results has been addressed in two recent papers using mouse modelsdeficient for the two different TNF-α receptors TNF-R1 and TNF-R2. Both groupsfound that the detrimental effects of TNF-α were due to the activity of TNF-R1,whereas activation of TNF-R2 leads to neuroprotection and trophic effects [40,142]. Microglia cells are a major source for TNF-α in brain. Among other factorsthat control the release of TNF-α from microglia increasing evidence suggeststhat also purine receptors are involved. The receptor responsible for this effect isP2X7 that needs relatively high concentrations of ATP indicating its activity underacute situations [53, 57, 121].Another important cytokine in brain is the pro-inflammatory chemokine CCL2.Expression of this chemokine in glia cells has been associated with a variety ofneurodegenerative diseases [11, 68]. The expression of CCL2 in brain most likelyorchestrates the infiltration of the CNS by blood macrophages [55, 56, 127],however, recent results, showing various effects of CCL2 in brain cells indicateadditional functions of CCL2 in brain [11]. Recently, two reports show markedprotection of neurons by CCL2 stimulation after the neurons were treated withNMDA or HIV-tat [16, 35]. Although the mechanism of CCL2-induced neuro-protection is unknown, it has been hypothesized that CCL2 might modulate theuptake, release and metabolism of neurotransmitters like glutamate throughpresynaptic chemokine receptors [35, 76]. Interestingly, purine receptor stimu-lation has also been shown to be involved in the release of CCL2 from astrocytes.Two different purine receptors are involved. Thus astrocytic release of CCL2 canbe induced by stimulation of P2X7 receptors [91] as well as by stimulation ofadenosine A3 receptors [139].
Glia cells in purine receptor-induced neuroprotection
47

The findings described above suggest a general role of glial purine receptors inregulating the synthesis and release of various neuroprotective factors (Figure2.1). Thus, purines not only protect neurons indirectly, they also could haveindirect effects by inducing the release of neuroprotective factors from glia cells.Interestingly, the synthesis and release of neurotrophins, cytokines andchemokines in glia cells is controlled by several different purine receptors, namelyall four adenosine receptor subtypes, A1, A2A, A2B and A3, and the ATP receptor
Chapter 2
48
ATP
IL-6
Neuronal injuryischaemia
(2)
(1)
ATP
AdoAdo
(1) (3)
Ca2+-influxNT release
K+ ←←←
A1AR
NGF
S100BCCL2
A2AAR
P2Y
(8)
(9) (7)
(5)(5)
(4)
(8)
A1AR
A2BAR
A3AR
P2X7
(6)
(6)
(5)
(4)astrocyte
microglia
neuron
Figure 2.1. Overview of neuroprotective substances released by glia cells and its effects onneurons. (1) As a result of energy shortage ATP is broken down to adenosine. Adenosine istransported out of the cell by facilitated diffusion transporters. (2) ATP can be transported out ofthe cell co-localized with certain neurotransmitters or by ATP-conducting ion-channels. ATP is alsoreleased when severely damaged cells release all of their contents. (3) Extracellular ATP is brokendown by nucleotidases into adenosine. (4) ATP stimulates P2X7 receptors in astrocytes and P2Yreceptors in microglia resulting in the increased production of CCL2 and IL-6 respectively. (5)Adenosine stimulates adenosine A1 receptors in neurons resulting in a reduction of Ca2+ influx, aninhibition of neurotransmitter release and an increase of K+ influx, causing hyperpolarization.Adenosine stimulates A1, A2B and A3 receptors in astrocytes resulting in an increased production ofNGF, S100B, IL-6 and CCL2. Adenosine stimulates A2A receptors in microglia increasing theproduction of NGF. (6) S100B, CCL2, IL-6 and NGF are released by astrocytes. (7) NGF and IL-6are released by microglia. (8) S100B, CCL2, NGF exert neuroprotective effects in neurons. (9) IL-6exerts neuroprotection possibly through the upregulation of adenosine A1 receptors.

subtypes P2X7 and P2Y. Since these receptors have different affinities for theirnatural ligands, it is tempting to speculate that different receptor subtypes will beactivated depending on the extracellular concentration of adenosine and/or ATP.Accordingly, the release of different neuroprotective substances from glia cellswould be precisely regulated by varying extracellular concentrations of purines.
Conclusions
Purines, ATP and adenosine are released in high concentrations in theendangered CNS. In this review we have discussed an indirect neuroprotectivefunction of purine signaling in the CNS, by activating astrocytes and microglia.Purine signaling in glia cells might influence the proliferation and apoptosis ofthese cells, which could be of importance to initiate or stop an inflammatoryreaction in the CNS. Moreover purine signaling activates glia cells to releasefactors that induce neuroprotective functions (see table 2.1 for an overview).Interestingly, different factors are released by stimulation of specific purinereceptor subtypes, indicating that the purine-induced release of neurotrophicfactors is carefully regulated. Since purine research in glia cells is a relativelynovel research area, future investigations are likely to find even more neuropro-tective substances to be released by glia cells and might create opportunities forthe development of new therapeutic approaches in neuroinflammation.
Glia cells in purine receptor-induced neuroprotection
49
Table 2.1. Overview of substances released by glia cells upon purine receptor stimulation.
ligand receptor substance released cell type reference
C-CPA adenosine A1 NGF, S100B mouse astrocytes [22]CGS 21680 adenosine A2A NGF mouse microglia [52]
NECA adenosine A2B IL-6 human astroglioma [38]2-chloroadenosine adenosine A2B IL-6 mouse astrocytes [106]
Cl-IB-MECA adenosine A3 CCL2 mouse astrocytes [139]propentofylline unknowna NGF mouse astrocytes [111]
AIT-082 not specificb NGF, NT-3, bFGF mouse astrocytes [47, 96]ATP P2Y IL-6 MG5 cells [110]
(microglia cell line)ATP P2X7 IL-1b mouse microglia [102]ATP P2X7 TNF-a rat microglia [53]ATP P2X7 CCL2 rat astrocytes [91]
guanosine/GTP unknown NGF mouse astrocytes [77]
a propentofylline acts as an adenosine transport inhibitor and a weak adenosine receptor antagonist [93] b AIT-082 might enhance the release of adenosine from astrocytes [27]

References
Chapter 2
50
1. Abbracchio, M. P., Ceruti, S., Brambilla, R., Franceschi, C., Malorni, W., Jacobson, K. A., von Lubitz,D. K., and Cattabeni, F. 1997. Modulation of apoptosis by adenosine in the central nervous system: a possible role for the A3 receptor. Pathophysiological significance and therapeutic implications for neurodegenerative disorders. Ann.N.Y.Acad.Sci. 825: 11-22
2. Abbracchio, M. P., Saffrey, M. J., Hopker, V., and Burnstock, G. 1994. Modulation of astroglial cell proliferation by analogues of adenosine and ATP in primary cultures of rat striatum. Neuroscience59(1): 67-76
3. Aschner, M., Allen, J. W., Kimelberg, H. K., LoPachin, R. M., and Streit, W. J. 1999. Glial cells in neurotoxicity development. Annu.Rev.Pharmacol.Toxicol. 39: 151-173
4. Banati, R. B., Gehrmann, J., Schubert, P., and Kreutzberg, G. W. 1993. Cytotoxicity of microglia. Glia 7(1): 111-118
5. Barger, S. W. and Van Eldik, L. J. 1992. S100 beta stimulates calcium fluxes in glial and neuronal cells. J Biol.Chem. 267(14): 9689-9694
6. Barone, F. C., Arvin, B., White, R. F., Miller, A., Webb, C. L., Willette, R. N., Lysko, P. G., and Feuerstein, G. Z. 1997. Tumor necrosis factor-alpha. A mediator of focal ischemic brain injury. Stroke 28(6): 1233-1244
7. Bauer, J., Strauss, S., Volk, B., and Berger, M. 1991. IL-6-mediated events in Alzheimer's disease pathology. Immunol.Today 12(11): 422
8. Bennett, G. C., Ford, A. P., Smith, J. A., Emmett, C. J., Webb, T. E., and Boarder, M. R. 2003. P2Y receptor regulation of cultured rat cerebral cortical cells: calcium responses and mRNA expression in neurons and glia. Br.J Pharmacol. 139(2): 279-288
9. Bensadoun, J. C., de Almeida, L. P., Dreano, M., Aebischer, P., and Deglon, N. 2001. Neuroprotective effect of interleukin-6 and IL6/IL6R chimera in the quinolinic acid rat model of Huntington's syndrome. Eur J Neurosci. 14(11): 1753-1761
10. Biber, K., Lubrich, B., Fiebich, B. L., Boddeke, H. W., and van Calker, D. 2001. Interleukin-6 enhances expression of adenosine A(1) receptor mRNA and signaling in cultured rat cortical astrocytes and brain slices. Neuropsychopharmacology 24(1): 86-96
11. Biber, K., Zuurman, M. W., Dijkstra, I. M., and Boddeke, H. W. 2002. Chemokines in the brain: neuroimmunology and beyond. Curr.Opin.Pharmacol. 2(1): 63-68
12. Boche, D., Cunningham, C., Gauldie, J., and Perry, V. H. 2003. Transforming growth factor-beta 1-mediated neuroprotection against excitotoxic injury in vivo. J Cereb.Blood Flow Metab 23(10):1174-1182
13. Boucsein, C., Zacharias, R., Farber, K., Pavlovic, S., Hanisch, U. K., and Kettenmann, H. 2003. Purinergic receptors on microglial cells: functional expression in acute brain slices and modulation of microglial activation in vitro. Eur J Neurosci. 17(11): 2267-2276
14. Brambilla, R., Cottini, L., Fumagalli, M., Ceruti, S., and Abbracchio, M. P. 2003. Blockade of A2A
adenosine receptors prevents basic fibroblast growth factor-induced reactive astrogliosis in rat striatal primary astrocytes. Glia 43(2): 190-194
15. Bruce-Keller, A. J. 1999. Microglial-neuronal interactions in synaptic damage and recovery.J Neurosci.Res. 58(1): 191-201
16. Bruno, V., Copani, A., Besong, G., Scoto, G., and Nicoletti, F. 2000. Neuroprotective activity of chemokines against N-methyl-D-aspartate or beta-amyloid-induced toxicity in culture. Eur J Pharmacol 399(2-3): 117-121
17. Burnstock, G., Campbell, G., Satchell, D., and Smythe, A. 1970. Evidence that adenosine triphosphate or a related nucleotide is the transmitter substance released by non-adrenergic inhibitory nerves in the gut. Br.J Pharmacol. 40(4): 668-688
18. Burnstock, G. and Williams, M. 2000. P2 purinergic receptors: modulation of cell function and therapeutic potential. J Pharmacol.Exp.Ther. 295(3): 862-869

Glia cells in purine receptor-induced neuroprotection
51
19. Carlson, N. G., Wieggel, W. A., Chen, J., Bacchi, A., Rogers, S. W., and Gahring, L. C. 1999. Inflammatory cytokines IL-1 alpha, IL-1 beta, IL-6, and TNF-alpha impart neuroprotection to an excitotoxin through distinct pathways. J Immunol 163(7): 3963-3968
20. Charles, M. P., Adamski, D., Kholler, B., Pelletier, L., Berger, F., and Wion, D. 2003. Induction of neurite outgrowth in PC12 cells by the bacterial nucleoside N6-methyldeoxyadenosine is mediated through adenosine A2a receptors and via cAMP and MAPK signaling pathways. Biochem.Biophys.Res.Commun. 304(4): 795-800
21. Ciccarelli, R., Ballerini, P., Sabatino, G., Rathbone, M. P., D'Onofrio, M., Caciagli, F., and Di Iorio, P. 2001. Involvement of astrocytes in purine-mediated reparative processes in the brain. Int J Dev Neurosci 19(4): 395-414
22. Ciccarelli, R., Di Iorio, P., Bruno, V., Battaglia, G., D'Alimonte, I., D'Onofrio, M., Nicoletti, F., and Caciagli, F. 1999. Activation of A(1) adenosine or mGlu3 metabotropic glutamate receptors enhances the release of nerve growth factor and S-100beta protein from cultured astrocytes. Glia27(3): 275-281
23. Cunha, R. A. 2001. Adenosine as a neuromodulator and as a homeostatic regulator in the nervous system: different roles, different sources and different receptors. Neurochem Int 38(2): 107-125
24. Cunha, R. A. and Ribeiro, J. A. 2000. ATP as a presynaptic modulator. Life Sci. 68(2): 119-13725. de Mendonca, A., Sebastiao, A. M., and Ribeiro, J. A. 2000. Adenosine: does it have a
neuroprotective role after all? Brain Res Brain Res Rev 33(2-3): 258-27426. Di Iorio, P., Kleywegt, S., Ciccarelli, R., Traversa, U., Andrew, C. M., Crocker, C. E., Werstiuk, E. S.,
and Rathbone, M. P. 2002. Mechanisms of apoptosis induced by purine nucleosides in astrocytes. Glia 38(3): 179-190
27. Di Iorio, P., Virgilio, A., Giuliani, P., Ballerini, P., Vianale, G., Middlemiss, P. J., Rathbone, M. P., and Ciccarelli, R. 2001. AIT-082 is neuroprotective against kainate-induced neuronal injury in rats. Exp.Neurol 169(2): 392-399
28. Dirnagl, U., Niwa, K., Lindauer, U., and Villringer, A. 1994. Coupling of cerebral blood flow to neuronal activation: role of adenosine and nitric oxide. Am J Physiol 267(1 Pt 2): 296-301
29. Dolphin, A. C., Forda, S. R., and Scott, R. H. 1986. Calcium-dependent currents in cultured rat dorsal root ganglion neurones are inhibited by an adenosine analogue. J Physiol 373: 47-61
30. Donato, R. 2003. Intracellular and extracellular roles of S100 proteins. Microsc.Res.Tech. 60(6):540-551
31. Dunwiddie, T. V. 1985. The physiological role of adenosine in the central nervous system. Int Rev Neurobiol 27: 63-139
32. Dunwiddie, T. V. and Masino, S. A. 2001. The role and regulation of adenosine in the central nervous system. Annu Rev Neurosci 24: 31-55
33. El Yacoubi, M., Ledent, C., Parmentier, M., Ongini, E., Costentin, J., and Vaugeois, J. M. 2001. In vivo labelling of the adenosine A2A receptor in mouse brain using the selective antagonist [3H]SCH 58261. Eur J Neurosci 14(9): 1567-1570
34. Eschke, D., Brand, A., Scheibler, P., Hess, S., Eger, K., Allgaier, C., and Nieber, K. 2001. Effect of anadenosine A(1) receptor agonist and a novel pyrimidoindole on membrane properties and neurotransmitter release in rat cortical and hippocampal neurons. Neurochem Int 38(5): 391-398
35. Eugenin, E. A., D'Aversa, T. G., Lopez, L., Calderon, T. M., and Berman, J. W. 2003. MCP-1 (CCL2) protects human neurons and astrocytes from NMDA or HIV-tat-induced apoptosis. J Neurochem.85(5): 1299-1311
36. Feoktistov, I. and Biaggioni, I. 1997. Adenosine A2B receptors. Pharmacol Rev 49(4): 381-40237. Fiebich, B. L., Biber, K., Gyufko, K., Berger, M., Bauer, J., and van Calker, D. 1996. Adenosine A2b
receptors mediate an increase in interleukin (IL)-6 mRNA and IL-6 protein synthesis in human astroglioma cells. J Neurochem 66(4): 1426-1431
38. Fiebich, B. L., Biber, K., Lieb, K., van Calker, D., Berger, M., Bauer, J., and Gebicke-Haerter, P. J. 1996. Cyclooxygenase-2 expression in rat microglia is induced by adenosine A2a-receptors. Glia18(2): 152-160
39. Fields, R. D. and Stevens, B. 2000. ATP: an extracellular signaling molecule between neurons and glia. Trends Neurosci. 23(12): 625-633

Chapter 2
52
40. Fontaine, V., Mohand-Said, S., Hanoteau, N., Fuchs, C., Pfizenmaier, K., and Eisel, U. 2002. Neurodegenerative and neuroprotective effects of tumor Necrosis factor (TNF) in retinal ischemia: opposite roles of TNF receptor 1 and TNF receptor 2. J Neurosci. 22(7): RC216
41. Fredholm, B. B., Abbracchio, M. P., Burnstock, G., Daly, J. W., Harden, T. K., Jacobson, K. A., Leff, P., and Williams, M. 1994. Nomenclature and classification of purinoceptors. Pharmacol.Rev. 46(2): 143-156
42. Fredholm, B. B., IJzerman, A.P., Jacobson, K. A., Klotz, K. N., and Linden, J. 2001. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol Rev 53(4): 527-552
43. Fumagalli, M., Brambilla, R., D'Ambrosi, N., Volonte, C., Matteoli, M., Verderio, C., and Abbracchio, M. P. 2003. Nucleotide-mediated calcium signaling in rat cortical astrocytes: Role of P2X and P2Y receptors. Glia 43(3): 218-230
44. Gadient, R. A. and Otten, U. H. 1997. Interleukin-6 (IL-6)--a molecule with both beneficial and destructive potentials. Prog Neurobiol 52(5): 379-390
45. Gebicke-Haerter, P. J., Christoffel, F., Timmer, J., Northoff, H., Berger, M., and van Calker, D. 1996.Both adenosine A1- and A2-receptors are required to stimulate microglial proliferation. Neurochem.Int. 29(1): 37-42
46. Gerber, U. and Gahwiler, B. H. 1994. GABAB and adenosine receptors mediate enhancement of the K+ current, IAHP, by reducing adenylyl cyclase activity in rat CA3 hippocampal neurons.J Neurophysiol 72(5): 2360-2367
47. Glasky, A. J., Kirat, S., Middlemiss, P. J., Gysbers, J. W., and Rathbone, M. P. 1995. A novel purine derivative AIT-082 increases synthesis of NGF, FGF-2, and NT-3 mRNA in astrocytes. Abstr.Soc.Neurosci. 21(295): (no. 1.25.10)
48. Glasky, A. J., Ritzman, R. F., Prisecaru, J., Santos, S., Kati, K., Rathbone, M. P., Middlemiss, P. J., and Crocker, C. E. 1996. Elevation of brain mRNA for neurotrophins by oral AIT-082 in mice. Abstr.Soc.Neurosci. 22(751): (no.301.2)
49. Godbout, J. P. and Johnson, R. W. 2004. Interleukin-6 in the aging brain. J Neuroimmunol. 147 (1-2): 141-144
50. Haas, H. L. and Selbach, O. 2000. Functions of neuronal adenosine receptors. Naunyn Schmiedebergs Arch Pharmacol 362(4-5): 375-381
51. Hammarberg, C., Schulte, G., and Fredholm, B. B. 2003. Evidence for functional adenosine A3
receptors in microglia cells. J Neurochem. 86(4): 1051-105452. Heese, K., Fiebich, B. L., Bauer, J., and Otten, U. 1997. Nerve growth factor (NGF) expression in rat
microglia is induced by adenosine A2a-receptors. Neurosci Lett 231(2): 83-8653. Hide, I., Tanaka, M., Inoue, A., Nakajima, K., Kohsaka, S., Inoue, K., and Nakata, Y. 2000.
Extracellular ATP triggers tumor necrosis factor-alpha release from rat microglia. J Neurochem. 75(3):965-972
54. Hindley, S., Herman, M. A., and Rathbone, M. P. 1994. Stimulation of reactive astrogliosis in vivoby extracellular adenosine diphosphate or an adenosine A2 receptor agonist. J Neurosci.Res. 38(4): 399-406
55. Huang, D., Han, Y., Rani, M. R., Glabinski, A., Trebst, C., Sorensen, T., Tani, M., Wang, J., Chien, P., O'Bryan, S., Bielecki, B., Zhou, Z. L., Majumder, S., and Ransohoff, R. M. 2000. Chemokines and chemokine receptors in inflammation of the nervous system: manifold roles and exquisite regulation. Immunol.Rev. 177: 52-67
56. Huang, D. R., Wang, J., Kivisakk, P., Rollins, B. J., and Ransohoff, R. M. 2001. Absence of monocytechemoattractant protein 1 in mice leads to decreased local macrophage recruitment and antigen-specific T helper cell type 1 immune response in experimental autoimmune encephalomyelitis.J Exp.Med. 193(6): 713-726
57. Inoue, K. 2002. Microglial activation by purines and pyrimidines. Glia 40(2): 156-16358. Jacobson, K. A., Jarvis, M. F., and Williams, M. 2002. Purine and pyrimidine (P2) receptors as drug
targets. J Med.Chem. 45(19): 4057-409359. Jankowsky, J. L. and Patterson, P. H. 2001. The role of cytokines and growth factors in seizures and
their sequelae. Prog.Neurobiol. 63(2): 125-149

Glia cells in purine receptor-induced neuroprotection
53
60. Jin, K. L., Mao, X. O., and Greenberg, D. A. 2000. Vascular endothelial growth factor: direct neuroprotective effect in in vitro ischemia. Proc Natl Acad Sci U S A 97(18): 10242-10247
61. Kim, G. M., Xu, J., Xu, J., Song, S. K., Yan, P., Ku, G., Xu, X. M., and Hsu, C. Y. 2001. Tumor necrosis factor receptor deletion reduces nuclear factor-kappaB activation, cellular inhibitor of apoptosis protein 2 expression, and functional recovery after traumatic spinal cord injury. J Neurosci.21(17): 6617-6625
62. Kim, J. K., Rathbone, M. P., Middlemiss, P. J., Hughes, D. W., and Smith, R. W. 1991. Purinergic stimulation of astroblast proliferation: guanosine and its nucleotides stimulate cell division in chick astroblasts. J Neurosci.Res. 28(3): 442-455
63. Klotz, K. N., Hessling, J., Hegler, J., Owman, C., Kull, B., Fredholm, B. B., and Lohse, M. J. 1998. Comparative pharmacology of human adenosine receptor subtypes - characterization of stably transfected receptors in CHO cells. Naunyn Schmiedebergs Arch Pharmacol 357(1): 1-9
64. Kossmann, T., Hans, V., Imhof, H. G., Trentz, O., and Morganti-Kossmann, M. C. 1996. Interleukin-6 released in human cerebrospinal fluid following traumatic brain injury may trigger nerve growth factor production in astrocytes. Brain Res 713(1-2): 143-152
65. Kukley, M., Barden, J. A., Steinhauser, C., and Jabs, R. 2001. Distribution of P2X receptors on astrocytes in juvenile rat hippocampus. Glia 36(1): 11-21
66. Kunioku, H., Inoue, K., and Tomida, M. 2001. Interleukin-6 protects rat PC12 cells from serum deprivation or chemotherapeutic agents through the phosphatidylinositol 3-kinase and STAT3 pathways. Neurosci.Lett. 309(1): 13-16
67. Kust, B. M., Biber, K., van Calker, D., and Gebicke-Haerter, P. J. 1999. Regulation of K+ channel mRNA expression by stimulation of adenosine A2a-receptors in cultured rat microglia. Glia 25(2): 120-130
68. Little, A. R., Benkovic, S. A., Miller, D. B., and O'Callaghan, J. P. 2002. Chemically induced neuronaldamage and gliosis: enhanced expression of the proinflammatory chemokine, monocyte chemoattractant protein (MCP)-1, without a corresponding increase in proinflammatory cytokines(1). Neuroscience 115(1): 307-320
69. Loddick, S. A., Turnbull, A. V., and Rothwell, N. J. 1998. Cerebral interleukin-6 is neuroprotective during permanent focal cerebral ischemia in the rat. J Cereb Blood Flow Metab 18(2): 176-179
70. Luthardt, J., Borvendeg, S. J., Sperlagh, B., Poelchen, W., Wirkner, K., and Illes, P. 2003. P2Y(1) receptor activation inhibits NMDA receptor-channels in layer V pyramidal neurons of the rat prefrontal and parietal cortex. Neurochem.Int. 42(2): 161-172
71. MacDonald, R. L., Skerritt, J. H., and Werz, M. A. 1986. Adenosine agonists reduce voltage-dependent calcium conductance of mouse sensory neurones in cell culture. J Physiol 370: 75-90
72. Marz, P., Heese, K., Dimitriades-Schmutz, B., Rose-John, S., and Otten, U. 1999. Role of interleukin-6 and soluble IL-6 receptor in region-specific induction of astrocytic differentiation and neurotrophin expression. Glia 26(3): 191-200
73. McGeer, P. L., Rogers, J., and McGeer, E. G. 1994. Neuroimmune mechanisms in Alzheimer diseasepathogenesis. Alzheimer Dis.Assoc.Disord. 8(3): 149-158
74. McGraw, J., Hiebert, G. W., and Steeves, J. D. 2001. Modulating astrogliosis after neurotrauma.J Neurosci.Res. 63(2): 109-115
75. Meistrell, M. E., III, Botchkina, G. I., Wang, H., Di Santo, E., Cockroft, K. M., Bloom, O., Vishnubhakat, J. M., Ghezzi, P., and Tracey, K. J. 1997. Tumor necrosis factor is a brain damaging cytokine in cerebral ischemia. Shock 8(5): 341-348
76. Meucci, O., Fatatis, A., Simen, A. A., Bushell, T. J., Gray, P. W., and Miller, R. J. 1998. Chemokines regulate hippocampal neuronal signaling and gp120 neurotoxicity. Proc Natl Acad Sci U S A 95(24): 14500-14505
77. Middlemiss, P. J., Gysbers, J. W., and Rathbone, M. P. 1995. Extracellular guanosine and guanosine-5'-triphosphate increase: NGF synthesis and release from cultured mouse neopallial astrocytes. BrainRes 677(1): 152-156
78. Moreau, J. L. and Huber, G. 1999. Central adenosine A(2A) receptors: an overview. Brain Res Brain Res Rev 31(1): 65-82

Chapter 2
54
79. Mrak, R. E. and Griffinbc, W. S. 2001. The role of activated astrocytes and of the neurotrophic cytokine S100B in the pathogenesis of Alzheimer's disease. Neurobiol.Aging 22(6): 915-922
80. Mrak, R. E., Sheng, J. G., and Griffin, W. S. 1995. Glial cytokines in Alzheimer's disease: review and pathogenic implications. Hum.Pathol. 26(8): 816-823
81. Muller, C. E. 2002. P2-pyrimidinergic receptors and their ligands. Curr.Pharm.Des 8(26): 2353-236982. Muller, H. W., Junghans, U., and Kappler, J. 1995. Astroglial neurotrophic and neurite-promoting
factors. Pharmacol.Ther. 65(1): 1-1883. Nawashiro, H., Tasaki, K., Ruetzler, C. A., and Hallenbeck, J. M. 1997. TNF-alpha pretreatment
induces protective effects against focal cerebral ischemia in mice. J Cereb.Blood Flow Metab 17(5): 483-490
84. Neary, J. T., Rathbone, M. P., Cattabeni, F., Abbracchio, M. P., and Burnstock, G. 1996. Trophic actions of extracellular nucleotides and nucleosides on glial and neuronal cells. Trends Neurosci19(1): 13-18
85. Nieto-Sampedro, M. and Mora, F. 1994. Active microglia, sick astroglia and Alzheimer type dementias. Neuroreport 5(4): 375-380
86. Noguchi, J. and Yamashita, H. 2000. Adenosine inhibits voltage-dependent Ca2+ currents in rat dissociated supraoptic neurones via A1 receptors. J Physiol 526 Pt 2: 313-326
87. Norton, W. T., Aquino, D. A., Hozumi, I., Chiu, F. C., and Brosnan, C. F. 1992. Quantitative aspectsof reactive gliosis: a review. Neurochem.Res. 17(9): 877-885
88. Ochiishi, T., Chen, L., Yukawa, A., Saitoh, Y., Sekino, Y., Arai, T., Nakata, H., and Miyamoto, H. 1999. Cellular localization of adenosine A1 receptors in rat forebrain: immunohistochemical analysis using adenosine A1 receptor-specific monoclonal antibody. J Comp Neurol 411(2): 301-316
89. Ogata, T. and Schubert, P. 1996. Programmed cell death in rat microglia is controlled by extracellular adenosine. Neurosci Lett 218(2): 91-94
90. Ohsawa, K., Imai, Y., Nakajima, K., and Kohsaka, S. 1997. Generation and characterization of a microglial cell line, MG5, derived from a p53-deficient mouse. Glia 21(3): 285-298
91. Panenka, W., Jijon, H., Herx, L. M., Armstrong, J. N., Feighan, D., Wei, T., Yong, V. W., Ransohoff, R. M., and MacVicar, B. A. 2001. P2X7-like receptor activation in astrocytes increases chemokine monocyte chemoattractant protein-1 expression via mitogen-activated protein kinase. J Neurosci21(18): 7135-7142
92. Park, K. S., Jeong, S. W., Cha, S. K., Lee, B. S., Kong, I. D., Ikeda, S. R., and Lee, J. W. 2001. Modulation of N-type Ca2+ currents by A1-adenosine receptor activation in male rat pelvic ganglion neurons. J Pharmacol Exp Ther 299(2): 501-508
93. Parkinson, F. E. and Fredholm, B. B. 1991. Effects of propentofylline on adenosine A1 and A2
receptors and nitrobenzylthioinosine-sensitive nucleoside transporters: quantitative autoradiographic analysis. Eur J Pharmacol. 202(3): 361-366
94. Pavenstadt, H., Ruh, J., Greger, R., and Schollmeyer, P. 1994. Adenosine-induced hyperpolarization of the membrane voltage in rat mesangial cells in primary culture. Br J Pharmacol 113(1): 7-12
95. Penkowa, M., Giralt, M., Carrasco, J., Hadberg, H., and Hidalgo, J. 2000. Impaired inflammatory response and increased oxidative stress and neurodegeneration after brain injury in interleukin-6-deficient mice. Glia 32(3): 271-285
96. Rathbone, M. P., Middlemiss, P. J., Gysbers, J. W., Andrew, C., Herman, M. A., Reed, J. K., Ciccarelli, R., Di Iorio, P., and Caciagli, F. 1999. Trophic effects of purines in neurons and glial cells.Prog Neurobiol 59(6): 663-690
97. Ridet, J. L., Malhotra, S. K., Privat, A., and Gage, F. H. 1997. Reactive astrocytes: cellular and molecular cues to biological function. Trends Neurosci. 20(12): 570-577
98. Rothwell, N. J. and Luheshi, G. N. 1996. Brain TNF: damage limitation or damaged reputation? Nat.Med. 2(7): 746-747
99. Rudolphi, K. A. and Schubert, P. 1996. Purinergic interventions in traumatic and ischemic injury. InNovel Thearapies for CNS Injuries, Peterson, P. L. and Willis, P. W. 327-342. Boca Raton, New York, London, Tokyo: CRC Press Inc.
100. Rudolphi, K. A. and Schubert, P. 1997. Modulation of neuronal and glial cell function by adenosine and neuroprotection in vascular dementia. Behav Brain Res 83(1-2): 123-128

Glia cells in purine receptor-induced neuroprotection
55
101. Rudolphi, K. A., Schubert, P., Parkinson, F. E., and Fredholm, B. B. 1992. Neuroprotective role of adenosine in cerebral ischaemia. Trends Pharmacol Sci 13(12): 439-445
102. Sanz, J. M. and Di Virgilio, F. 2000. Kinetics and mechanism of ATP-dependent IL-1 beta release from microglial cells. J Immunol. 164(9): 4893-4898
103. Sasaki, Y., Hoshi, M., Akazawa, C., Nakamura, Y., Tsuzuki, H., Inoue, K., and Kohsaka, S. 2003. Selective expression of Gi/o-coupled ATP receptor P2Y12 in microglia in rat brain. Glia 44(3):242-250
104. Schafer, B. W. and Heizmann, C. W. 1996. The S100 family of EF-hand calcium-binding proteins: functions and pathology. Trends Biochem.Sci. 21(4): 134-140
105. Schumann, G., Huell, M., Machein, U., Hocke, G., and Fiebich, B. L. 1999. Interleukin-6 activates signal transducer and activator of transcription and mitogen-activated protein kinase signal transduction pathways and induces de novo protein synthesis in human neuronal cells. J Neurochem.73(5): 2009-2017
106. Schwaninger, M., Neher, M., Viegas, E., Schneider, A., and Spranger, M. 1997. Stimulation of interleukin-6 secretion and gene transcription in primary astrocytes by adenosine. J Neurochem69(3): 1145-1150
107. Sebastiao, A. M. and Ribeiro, J. A. 2000. Fine-tuning neuromodulation by adenosine. Trends Pharmacol Sci 21(9): 341-346
108. Shaked, I., Porat, Z., Gersner, R., Kipnis, J., and Schwartz, M. 2004. Early activation of microglia as antigen-presenting cells correlates with T cell-mediated protection and repair of the injured central nervous system. J Neuroimmunol. 146(1-2): 84-93
109. Shen, Y., Li, R., and Shiosaki, K. 1997. Inhibition of p75 tumor necrosis factor receptor by antisenseoligonucleotides increases hypoxic injury and beta-amyloid toxicity in human neuronal cell line. J Biol.Chem. 272(6): 3550-3553
110. Shigemoto-Mogami, Y., Koizumi, S., Tsuda, M., Ohsawa, K., Kohsaka, S., and Inoue, K. 2001. Mechanisms underlying extracellular ATP-evoked interleukin-6 release in mouse microglial cell line,MG-5. J Neurochem 78(6): 1339-1349
111. Shinoda, I., Furukawa, Y., and Furukawa, S. 1990. Stimulation of nerve growth factor synthesis/secretion by propentofylline in cultured mouse astroglial cells. Biochem.Pharmacol. 39(11): 1813-1816
112. Si, Q. S., Nakamura, Y., Schubert, P., Rudolphi, K., and Kataoka, K. 1996. Adenosine and propentofylline inhibit the proliferation of cultured microglial cells. Exp.Neurol 137(2): 345-349
113. Sofroniew, M. V., Howe, C. L., and Mobley, W. C. 2001. Nerve growth factor signaling, neuroprotection, and neural repair. Annu.Rev.Neurosci. 24: 1217-1281
114. Song, W. J., Tkatch, T., and Surmeier, D. J. 2000. Adenosine receptor expression and modulation of Ca(2+) channels in rat striatal cholinergic interneurons. J Neurophysiol 83(1): 322-332
115. Spera, P. A., Ellison, J. A., Feuerstein, G. Z., and Barone, F. C. 1998. IL-10 reduces rat brain injury following focal stroke. Neurosci.Lett. 251(3): 189-192
116. Sperlagh, B., Kofalvi, A., Deuchars, J., Atkinson, L., Milligan, C. J., Buckley, N. J., and Vizi, E. S. 2002. Involvement of P2X7 receptors in the regulation of neurotransmitter release in the rat hippocampus. J Neurochem. 81(6): 1196-1211
117. Stoll, G., Jander, S., and Schroeter, M. 2000. Cytokines in CNS disorders: neurotoxicity versus neuroprotection. J Neural Transm.Suppl 59: 81-89
118. Stoll, G., Jander, S., and Schroeter, M. 2002. Detrimental and beneficial effects of injury-induced inflammation and cytokine expression in the nervous system. Adv.Exp.Med.Biol. 513: 87-113
119. Streit, W. J. 2002. Microglia as neuroprotective, immunocompetent cells of the CNS. Glia 40(2): 133-139
120. Streit, W. J., Walter, S. A., and Pennell, N. A. 1999. Reactive microgliosis. Prog.Neurobiol. 57(6): 563-581
121. Suzuki, T., Hide, I., Ido, K., Kohsaka, S., Inoue, K., and Nakata, Y. 2004. Production and release of neuroprotective tumor necrosis factor by P2X7 receptor-activated microglia. J Neurosci. 24(1): 1-7
122. Swanson, T. H., Drazba, J. A., and Rivkees, S. A. 1995. Adenosine A1 receptors are located predominantly on axons in the rat hippocampal formation. J Comp Neurol 363(4): 517-531

Chapter 2
56
123. Swartz, K. R., Liu, F., Sewell, D., Schochet, T., Campbell, I., Sandor, M., and Fabry, Z. 2001. Inter-leukin-6 promotes post-traumatic healing in the central nervous system. Brain Res 896(1-2): 86-95
124. Tabakman, R., Lecht, S., Sephanova, S., Arien-Zakay, H., and Lazarovici, P. 2004. Interactions between the cells of the immune and nervous system: neurotrophins as neuroprotection mediators in CNS injury. Prog.Brain Res. 146: 387-401
125. Tanaka, E., Yasumoto, S., Hattori, G., Niiyama, S., Matsuyama, S., and Higashi, H. 2001. Mechanisms underlying the depression of evoked fast EPSCs following in vitro ischemia in rat hippocampal CA1 neurons. J Neurophysiol 86(3): 1095-1103
126. Traversa, U., Bombi, G., Di Iorio, P., Ciccarelli, R., Werstiuk, E. S., and Rathbone, M. P. 2002. Specific [(3)H]-guanosine binding sites in rat brain membranes. Br.J Pharmacol. 135(4): 969-976
127. Trebst, C., Staugaitis, S. M., Tucky, B., Wei, T., Suzuki, K., Aldape, K. D., Pardo, C. A., Troncoso, J.,Lassmann, H., and Ransohoff, R. M. 2003. Chemokine receptors on infiltrating leucocytes in inflammatory pathologies of the central nervous system (CNS). Neuropathol.Appl.Neurobiol. 29(6): 584-595
128. Trussell, L. O. and Jackson, M. B. 1985. Adenosine-activated potassium conductance in cultured striatal neurons. Proc Natl Acad Sci U S A 82(14): 4857-4861
129. Unger, J. W. 1998. Glial reaction in aging and Alzheimer's disease. Microsc.Res.Tech. 43(1): 24-28130. Van Wagoner, N. J. and Benveniste, E. N. 1999. Interleukin-6 expression and regulation in
astrocytes. J Neuroimmunol 100(1-2): 124-139131. Volonte, C., Amadio, S., Cavaliere, F., D'Ambrosi, N., Vacca, F., and Bernardi, G. 2003. Extracellular
ATP and neurodegeneration. Curr.Drug Targets.CNS.Neurol Disord. 2(6): 403-412132. von Lubitz, D. K. 1999. Adenosine and cerebral ischemia: therapeutic future or death of a brave
concept? [corrected and republished article originally printed in Eur J Pharmacol 1999 Jan 15;365(1):9-25]. Eur J Pharmacol 371(1): 85-102
133. von Lubitz, D. K., Lin, R. C., Bischofberger, N., Beenhakker, M., Boyd, M., Lipartowska, R., and Jacobson, K. A. 1999. Protection against ischemic damage by adenosine amine congener, a potent and selective adenosine A1 receptor agonist. Eur J Pharmacol 369(3): 313-317
134. Wang, Y., Chang, C. F., Morales, M., Chiang, Y. H., and Hoffer, J. 2002. Protective effects of glial cell line-derived neurotrophic factor in ischemic brain injury. Ann.N.Y.Acad.Sci. 962: 423-437
135. Weinstein, D. E., Shelanski, M. L., and Liem, R. K. 1991. Suppression by antisense mRNA demonstrates a requirement for the glial fibrillary acidic protein in the formation of stable astrocyticprocesses in response to neurons. J Cell Biol. 112(6): 1205-1213
136. Williams, M. 2000. Purines: from premise to promise. J Auton.Nerv.Syst. 81(1-3): 285-288137. Williams, M. 1989. Adenosine: the prototypic neuromodulator. Neurochemistry international 14(3):
249-264138. Williams, M. 1995. Purinoceptors in Central Nervous System Function: targets for Therapeutic
intervention. In Psychopharmacology: The Fourth Generation of Progress, Bloom FE, Kupfer DJ 643-656. New York: Raven Press. Ltd.
139. Wittendorp, M. C., Boddeke, H. W., and Biber, K. 2004. Adenosine A3 receptor-induced CCL2 synthesis in cultured mouse astrocytes. Glia 46(4): 410-418
140. Wittendorp, M. C., Von Frijtag Drabbe Kunzel J., IJzerman, A. P., Boddeke, H. W. G. M., and Biber, K. 2004. The mouse brain adenosine A(1) receptor: functional expression and pharmacology. Eur J Pharmacol. 487(1-3): 73-79
141. Wollmer, M. A., Lucius, R., Wilms, H., Held-Feindt, J., Sievers, J., and Mentlein, R. 2001. ATP and adenosine induce ramification of microglia in vitro. J Neuroimmunol 115(1-2): 19-27
142. Yang, L., Lindholm, K., Konishi, Y., Li, R., and Shen, Y. 2002. Target depletion of distinct tumor necrosis factor receptor subtypes reveals hippocampal neuron death and survival through different signal transduction pathways. J Neurosci. 22(8): 3025-3032

57
Published in Glia. 2004. 46 (4): 410-418
Adenosine A3 receptor-inducedCCL2 synthesis in cultured mouse astrocytes
M.C. Wittendorp, H.W.G.M. Boddeke and K. Biber
Chapter 3

Summary
Introduction
Adenosine is essentially involved in the coupling of cellular metabolism to energysupply. In order to control metabolic rate, adenosine, for example, suppressesneuronal firing and increases blood flow [18, 31, 50]. The effects of adenosine aremediated by four types of G-protein coupled adenosine receptors; A1, A2A, A2B
and A3 [27]. In the brain adenosine acts as a physiological neuromodulator [16,66]. Under pathological conditions like seizures, ischaemia and hypoxia theextracellular adenosine concentration rapidly rises from nanomolar to micromolarlevels [20, 57]. This large amount of adenosine has a significant neuroprotectiveeffect [17, 57, 58, 64, 67] mainly by stimulating neuronal adenosine A1 receptors[27]. In neurons, adenosine inhibits the release of excitatory neurotransmittersand causes hyperpolarization of the membrane potential [20, 57]. Recentevidence however, suggests that stimulation of glial adenosine receptors leads tothe synthesis of various neuroprotective substances. It has for example beenshown that adenosine A1 receptor stimulation in astrocytes induces release ofnerve growth factor and S100B protein [13]. Moreover, it was shown thatstimulation of adenosine A2B receptors in astrocytes induces synthesis and releaseof interleukin-6 (IL-6) [23, 59]. The induction of IL-6 synthesis might alsocontribute to the neuroprotective actions of adenosine since there is accumulatingevidence that IL-6 is a neuroprotective cytokine [45-47, 53, 63]. Thus theneuroprotective properties of adenosine might be direct in neurons and indirectin glia cells by stimulating the synthesis of neuroprotective substances.Chemokines are small chemotactic cytokines of approximately 10kDa, that areessential elements in the peripheral immune system (for review see [5, 51, 56]).
Chapter 3
58
During neuropathological conditions high concentrations of adenosine are released stimulatingadenosine receptors in neurons and glia cells. It has recently been shown that stimulation ofadenosine receptors in glia cells induces the release of neuroprotective substances like NGF,S100B and interleukin-6 (IL-6). It has therefore been suggested that glial adenosine receptors areinvolved in neuroprotection. Since recently neuroprotective effects of the chemokine CCL2(formerly known as MCP-1) have been reported, we investigated the possible effect of adenosinereceptor stimulation on glial CCL2 synthesis. Here we show that stimulation of cultured murineastrocytes with the selective adenosine A3 receptor agonist 2-chloro-N6-(3-iodobenzyl)-N-methyl-5’-carbamoyladenosine (Cl-IB-MECA) induced the release of CCL2. Specific ligands for adenosineA1 or A2 receptors did not affect CCL2 release. Furthermore, Cl-IB-MECA-induced CCL2synthesis was inhibited by adenosine A3 receptor antagonists. These results show that stimulationof adenosine A3 receptors in astrocytes induced the release of CCL2, thus supporting theassumption that adenosine receptors in glia cells regulate the synthesis of neuroprotectivesubstances.

It is now clear that chemokines are also found in brain where they are pre-dominantly expressed in glia cells [3, 4, 6]. The expression of CCL2 in glia cells, which is evident in most neurodegenerativeconditions, controls the infiltration of the brain by blood leukocytes [25, 37, 38,60]. On the other hand the presence of functional chemokine receptors in alltypes of endogenous brain cells (neurons, astrocytes, oligodendrocytes andmicroglia) led to the assumption that chemokines may function as intercellularmessengers in the brain (for review see [7, 36]). In line with this assumption arethe effects of CCL2 on astroglial synthesis of matrix metalloproteinases andcytokine release [10, 15, 19]. Moreover, recent results show direct neuro-protective properties of CCL2 stimulation in NMDA or HIV-tat-treated neurons[11, 21]. Although it is clear that astrocytes express and release CCL2, little is yet knownon the regulation of glial CCL2 synthesis. The findings, that astroglial adenosinereceptors stimulate expression of various neuroprotective substances and thatCCL2 has direct neuroprotective properties, prompted us to investigate possibleeffects of glial adenosine receptor stimulation on CCL2 synthesis.
Materials and methods
All cell culture media and supplements were obtained from Gibco-BRL Lifetechnologies (Breda, the Netherlands). Reverse transcriptase enzyme and bufferwere purchased from Promega (Madison, WI, USA). Taq-polymerase and bufferfor PCR were obtained from Eppendorf (Boulder, CO, USA). N6-cyclopenty-ladenosine (CPA), N-ethylcarboxamidoadenosine (NECA), 1,3-dimethyl-8-phenylxanthine (8PT) and 3-propyl-6-ethyl-5-[(ethylthio)carbony]-2 phenyl-4-propyl-3-pyridine carboxylate (MRS 1523), Lipopolysaccharide (E.Coli, 0128:B12)and pertussis toxin were obtained from Sigma-Aldrich (St.Louis, MO, U.S.A.), 2-chloro-N6-(3-iodobenzyl)-N-methyl-5’-carbamoyladenosine (Cl-IB-MECA) and 2-[p-(2-carboxyethyl)phenyl-ethylamino]-5'-N-ethylcarboxamidoadenosine(CGS21680) were purchased from Tocris Cookson (Ellisville, MO, USA).
Glial cultures
Astrocyte cell cultures were established as described previously [8]. In brief,mouse cortex was dissected from new-born mouse pups (<1 day). Brain tissuewas gently dissociated by trituration in phosphate-buffered saline (PBS) andfiltered through a cell strainer (70 mm Ø, Falcon) into Dulbecco’s Modified Eaglemedium (DMEM). After two washing steps (200 x g for 10 min), cells wereseeded in culture flasks (75 cm2, Greiner; 10 x 106 cells/flask). Cultures weremaintained up to 4 weeks in DMEM containing 10% fetal calf serum with 0.01%
Adenosine A3 receptor-induced CCL2 release
59

penicillin and 0.01% streptomycin in a humidified atmosphere (5% CO2) at 37ºC.Culture medium was changed the second day after preparation and every 6 daysthereafter. For in situ hybridization mixed astrocyte cultures were incubated with0.25% w/v porcine trypsin at 37ºC for 10 min and seeded on poly-L-lysine-coatedglass coverslips in 6-well culture dishes. Floating microglia were harvested fromconfluent mixed astrocyte cultures and plated on new culture dishes. Microgliacultures were pure (> 95%) as tested by cell specific markers (F4/80 and Mac-1).For in situ hybridization microglia were seeded on poly-L-lysine-coated glass cover-slips in 6-well culture dishes.
CCL2 ELISA
For ELISA experiments glia cells were seeded directly after preparation in 24-welldishes at a density of 1.0~1.5 x 105 cells/well and experiments were performed12-14 days thereafter. Microglia were removed from these mixed astrocytescultures by tapping and rinsing the plates yielding cultures that contained > 90%astrocytes. Harvested microglia were plated in 24-well plates at a density of 1.0 x105 cells/well The medium was changed two hours before stimulation withDMEM containing 0.1% fetal calf serum supplemented with 0.01% penicillin and0.01% streptomycin. Cells were incubated 24 hours with adenosine receptoragonists, which were added to the culture medium. Antagonists were added 30minutes before agonist treatment and were left in the medium the following 24hours. Control cells (unstimulated) received solvent only. In some experiments,cells were pre-incubated 18 hours with 200 ng/ml pertussis toxin. The mediumwas then changed followed by incubation with the adenosine agonist.Supernatants were then removed, centrifuged at 10.000 g for 2 minutes andstored at –80ºC until analysis. A commercial ELISA kit (R&D systems, Minneapolis, MN, USA) was used todetermine CCL2 levels. For ELISA analysis supernatants from astrocytes werediluted 1:4 in stimulation medium (DMEM + 0.1% fetal calf serum, 0.01%penicillin and streptomycin); supernatants from microglia were not diluted.ELISA experiments were carried out according to the manufacturer’s protocolwith some modifications due to the low activity of the streptavidin horseradishperoxidase supplied in the ELISA kit. After incubation with the secondarybiotinylated antibody, plates were washed and incubated for 30 min withstreptavidin conjugated to poly-horseradish peroxidase (Strepta-E from CentraalLaboratorium voor Bloedtransfusie, Amsterdam, The Netherlands) at aconcentration of 125 ng/ml in incubation buffer. A final washing step wasfollowed by the chromogen reaction: 1 mg of 3’, 3’, 5’, 5’-tetramethylbenzidin(Roth, Germany) in 0.3 ml 96% EtOH was added to 11 ml 0.1 M Na-acetate, pH6, and 0.004% H2O2 and plates were incubated for ca. 5 min with 0.1 ml of thechromogen solution until the reaction was stopped by addition of 1 M H2SO4.
Chapter 3
60

The absorption at 450 nm minus 570 nm was measured in a microplate reader(Labsystems©, Breda, The Netherlands), and the concentrations were calculatedusing SOFTmax PRO software (Molecular Devices, Sunnyvale, CA).
Reverse transcription-polymerase chain reaction (RT-PCR)
Cultured glia cells were lysed in guanidinium isothiocyanate/mercaptoethanolbuffer and total RNA was extracted with one phenol/chloroform step,precipitated according to Chomczynski and Sacchi [12]. Subsequently, RNA wastreated with RNAse free DNAse (Sigma-Aldrich, Bornhem, Belgium) for 2 hoursand precipitated. Reverse transcription (RT): 1 µg of total RNA was transcribedinto cDNA as described [8]. The quality of the cDNA was controlled usingglyceraldehyde-3-phosphate dehydrogenase (GAPDH) primers [9] and potentialcontamination by genomic DNA was checked for by running the reactionswithout reverse transcriptase and using GAPDH primers in subsequent poly-merase chain reaction (PCR) amplifications. PCR: 2 µl of the RT reaction product was used in the subsequent PCR ampli-fication as described [8]. Primer sequences used are listed in table 3.1. Cyclenumbers were 35 and annealing temperature was 58ºC for adenosine receptorprimers and 28 cycles and 60ºC for GAPDH primers.
In situ hybridization
For in situ hybridization PCRII vectors containing mouse adenosine receptor PCRproducts were linearized with BamHI or XhoI. Adenosine A3 receptor sense andantisense probes were synthesized by run off transcription and the use ofdigoxigenin-conjugated UTP according to the manufacturer’s protocol (BoehringerMannheim, Mannheim, Germany).
Adenosine A3 receptor-induced CCL2 release
61
Table 3.1. Primer sequences for mouse adenosine receptors and GAPDH
Gene Primer sequences (5’-3’) PCR product (bp)
A1 ACTTCTTCGTCTGGGTGCTG 349AGAAAGGTGACCCGGAACTT
A2A TCATGTACCTGGCCATCATC 349TGCTCCTGGGTAAGAAGCTC
A2B ATGTGGTCCCCATGAGCTAC 351AGGCATAGACAATGGGGTTG
A3 ATGGAAGCCGACAACACC 309CAGCAAGGACATGATGGAAG
GAPDH CATCCTGCACCACCAACTGCTTAG 312GCCTGCTTCACCACCTTCTTGATG

Cultured glia cells were seeded on poly-L-lysine-coated glass cover-slides andfixed in 4% paraformaldehyde for 30 min. The in situ hybridization procedure wascarried out as described earlier [14]; alkaline phosphatase-conjugated sheep anti-digoxigenin was used for the immunological detection of the digoxigenin- labeledRNA-RNA complex. Control experiments included hybridization with digoxigenin-labeled sense probes and hybridization with β-actin probes.
Intracellular calcium measurements
For calcium measurements, astrocytes were plated on glass coverslips two daysbefore the experiment. Cells were loaded 1 hour at 37ºC with Fura-PE3 (TEFLabs, Austin, USA) in loading buffer containing 120 mM NaCl, 5 mM HEPES, 6mM KCl, 2 mM CaCl2, 1 mM MgCl2, 5 mM glucose, 22 mM NaHCO3, 0.005 mMFura; pH 7.4. Subsequently, the coverslips were fixed in a perfusion chamber andmounted on an inverted microscope. Fluorimetric measurements were done usinga sensicam CCD camera supported by AxolabR 2.1 imaging software. Digitalimages of the cells were obtained at an emission wavelength of 510 nm usingpaired exposures to 340 and 380 nm excitation wavelength sampled at afrequency of 1 Hz. Fluorescence values representing spatial averages from adefined pixel area were recorded on-line. Increases in intracellular calciumconcentrations were expressed as the 340/380 ratio of the emission wavelengths.Compounds were administered with a pipette directly into the medium coveringthe cells. In pertussis toxin experiments, astrocytes were pre-incubated with 200ng/ml pertussis toxin overnight before the calcium measurements.
Data analysis
The results of the ELISA experiments are presented as mean ± S.E.M. of 3-4experiments per ligand. Statistical comparisons were performed using ANOVAfollowed by Tukey post hoc test. Significance refers to results where p < 0.05 wasobtained.
Results
Adenosine A3 receptor stimulation leads to CCL2 release from astrocytes
Similar to previous reports [30, 34], constitutive synthesis of CCL2 was observedin unstimulated astrocyte cultures. The CCL2 levels varied from 900-1800 pg/mlbetween various preparations. The adenosine A1 and A2A receptor agonists CPAand CGS 21680 (10 nM – 1 µM) did not affect the constitutive CCL2 synthesis inthese cultures (Figure 3.1A). A significant increase in CCL2 protein release inastrocyte cultures was observed in response to stimulation with the selective A3
adenosine receptor agonist Cl-IB-MECA and the non-selective adenosine receptor
Chapter 3
62

Adenosine A3 receptor-induced CCL2 release
63
CI-IB-MECANECA
CGS 21680
0
200
300
400
500
600
700
100
CCL2
pro
tein
(% o
f con
trol
)
control -7 -6log [agonist] (M)
LPS
0
40
80
120
CCL2
pro
tein
(% o
f CL-
IB-M
ECA
(10-7
))
-8 -7 -6log [antagonist] (M)
140
60
180
220
100
-8 -7 -6log [agonist]
-5
*
***
-∞
CCL2
pro
tein
(% o
f con
trol
)
control
*
v
8PTMRS 1523
CPA
CPANECACGS 21680CI-IB-MECA
A
B
C
Figure 3.1. Effect of adenosine receptor agonists and antagonists on CCL2 release in culturedmurine astrocytes and microglia. A) Effect of various concentrations (10 nM – 10 µM) of adenosinereceptor agonists on CCL2 release in cultured mixed astrocytes. Data represented as percentage ofcontrol (unstimulated) value, which varied between 900 pg/ml and 1800 pg/ml in differentpreparations. *, p < 0.05 for Cl-IB-MECA induced CCL2 release versus control value. ^, p < 0.05for NECA induced CCL2 release versus control value. B) Effect of adenosine receptor antagonistson CCL2 release in cultured mixed astrocytes induced by 100 nM Cl-IB-MECA. Data representedas percentage of value stimulated with 100 nM Cl-IB-MECA. Data are given as mean ± S.E.M.(n=4). *, significantly different from control p < 0.05. C) Lack of effect of various adenosine recep-tor agonists on CCL2 release in pure microglial cultures. Stimulation cultured microglia for 24hwith 100 nM or 1 µM adenosine receptor ligands did not change basal CCL2 release (300 pg/ml),whereas stimulation with LPS (1 ng/ml for 24h) pronouncedly induced CCL2 in these cultures.Data are given as mean ± S.E.M. (n=4).

agonist NECA, respectively (Figure 3.1A). Significant induction of CCL2 releaseby Cl-IB-MECA was observed at a concentration of 100 nM (p < 0.05). At con-centrations 10 nM and 1 µM CCL2 induction just failed to reach significance. Thenon-selective adenosine receptor antagonist NECA stimulated CCL2 release at aconcentration of 1 µM. The specific induction of CCL2 release with Cl-IB-MECA suggested the involve-ment of adenosine A3 receptors. In subsequent experiments astrocyte cultureswere pre-incubated with different concentrations (10 nM, 100 nM and 1 µM) ofthe antagonists 8-PT and MRS 1523 and then stimulated with 100 nM Cl-IB-MECA (Figure 3.1B). The non-selective adenosine antagonist 8-PT, which has avery low affinity for the adenosine A3 receptor, did not influence Cl-IB-MECA-induced CCL2 release. In contrast, the selective A3 receptor antagonist MRS 1523significantly inhibited Cl-IB-MECA-induced CCL2 release (p < 0.05) (Figure 3.1B),whereas CCL2 levels in unstimulated cells were unaffected by treatment with 1µM MRS 1523 (116 ± 18% from unstimulated controls).
No involvement of adenosine A3 receptors in CCL2 release from microglia
Since astrocyte cultures used in our experiments still contained microglia cells,similar ELISA experiments were performed in pure microglia cultures in order toinvestigate whether adenosine receptor stimulation would induce CCL2 release inthese cells. CCL2 ELISA analysis revealed a basal release of CCL2 in culturedmicroglia that was strongly induced by stimulation with LPS (1 ng/ml) (Figure3.1C). In contrast to the situation in astrocyte cultures, CCL2 release from puremicroglia was not significantly changed by stimulation with CPA, NECA,CGS21680 or Cl-IB-MECA (Figure 3.1C).
Adenosine receptor mRNA expression in glia cells
In order to check whether adenosine A3 receptor mRNA is expressed both inastrocyte cultures and pure microglia cultures, RT-PCR experiments wereperformed. Results of the RT-PCR experiments (Figure 3.2A) showed mRNAexpression of the adenosine A1, A2B and A3 receptor subtypes in astrocytecultures. Expression of adenosine A2A receptor mRNA was not detectable (Figure3.2A). In pure microglial cultures mRNA expression for adenosine A1, A2B and A3
adenosine receptor mRNA were found. Adenosine A2A receptor mRNA express-ion in cultured microglia was only detected after 2 hour stimulation with bacteriallipopolysaccharide (LPS) (100 ng/ml). In contrast to microglia, LPS did notinduce the expression of A2A receptor mRNA in mixed astrocyte cultures (datanot shown). Since adenosine A3 receptor mRNA was detected in both mixedastrocyte- and pure microglial cultures in situ hybridization experiments using aprobe for adenosine A3 receptor mRNA were performed. In situ hybridizationanalysis, using the antisense probe for adenosine A3 receptor mRNA, showed
Chapter 3
64

positive staining in cultured astrocytes (Figure 3.2B) and microglia (Figure 3.2D).Approximately 50% of the microglia and 75% of the astrocytes expressedadenosine A3 receptor mRNA. No staining was found using the sense controlprobe in astrocytes (Figure 3.2C) or microglia (Figure 3.2E).
Adenosine A3 receptor-induced CCL2 release
65
Figure 3.2. Analysis of adenosine receptor mRNA expression in cultured mixed astrocytes andpure microglial cultures. A) RT-PCR analysis of adenosine receptor mRNA expression in culturedmixed astrocytes and microglia. MM, molecular marker; highlighted band represents 500 bp. Onthe right results of the GAPDH control experiment of astrocytes (1), unstimulated microglia (2)and microglia stimulated with LPS (3). Results are shown from a typical experiment, similarresults were obtained from 4 independent experiments.In situ hybridization analysis showing the expression of adenosine A3 receptor mRNA in culturedastrocytes and microglia. B) Specific in situ hybridization signal in cultures astrocytes using theantisense riboprobe. C) Control hybridization with the sense probe revealed no staining in culturedastrocytes. D) Specific in situ hybridization signal in cultured microglia using the antisenseriboprobe. E) Control hybridization with the sense probe revealed no staining in culturedmicroglia. In B and C the bar represents 50 µm, in D and E it represents 20 µm.
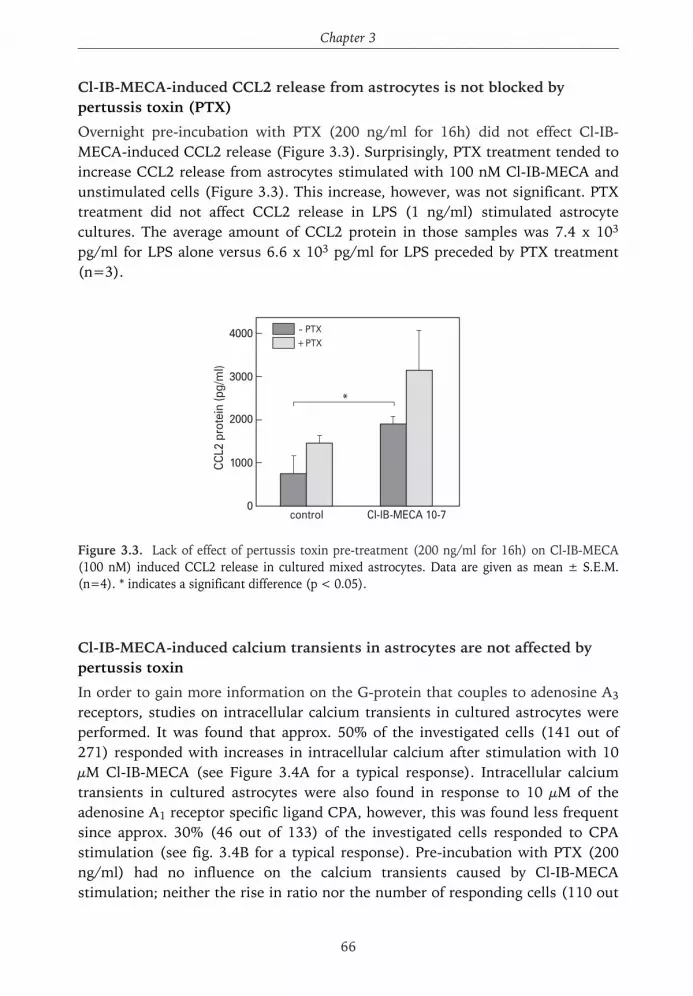
Cl-IB-MECA-induced CCL2 release from astrocytes is not blocked bypertussis toxin (PTX)
Overnight pre-incubation with PTX (200 ng/ml for 16h) did not effect Cl-IB-MECA-induced CCL2 release (Figure 3.3). Surprisingly, PTX treatment tended toincrease CCL2 release from astrocytes stimulated with 100 nM Cl-IB-MECA andunstimulated cells (Figure 3.3). This increase, however, was not significant. PTXtreatment did not affect CCL2 release in LPS (1 ng/ml) stimulated astrocytecultures. The average amount of CCL2 protein in those samples was 7.4 x 103
pg/ml for LPS alone versus 6.6 x 103 pg/ml for LPS preceded by PTX treatment(n=3).
Cl-IB-MECA-induced calcium transients in astrocytes are not affected bypertussis toxin
In order to gain more information on the G-protein that couples to adenosine A3
receptors, studies on intracellular calcium transients in cultured astrocytes wereperformed. It was found that approx. 50% of the investigated cells (141 out of271) responded with increases in intracellular calcium after stimulation with 10µM Cl-IB-MECA (see Figure 3.4A for a typical response). Intracellular calciumtransients in cultured astrocytes were also found in response to 10 µM of theadenosine A1 receptor specific ligand CPA, however, this was found less frequentsince approx. 30% (46 out of 133) of the investigated cells responded to CPAstimulation (see fig. 3.4B for a typical response). Pre-incubation with PTX (200ng/ml) had no influence on the calcium transients caused by Cl-IB-MECAstimulation; neither the rise in ratio nor the number of responding cells (110 out
Chapter 3
66
0
2000
3000
4000
1000CCL2
pro
tein
(pg/
ml)
control CI-IB-MECA 10-7
- PTX+ PTX
*
Figure 3.3. Lack of effect of pertussis toxin pre-treatment (200 ng/ml for 16h) on Cl-IB-MECA(100 nM) induced CCL2 release in cultured mixed astrocytes. Data are given as mean ± S.E.M.(n=4). * indicates a significant difference (p < 0.05).

of 177) was changed after PTX treatment (Figure 3.4C and 3.4D). In strikingcontrast pre-treatment with PTX (200 ng/ml) completely abolished the inductionof intracellular calcium transients in astrocytes in response to 10 µM CPA (Figure3.4D). Only 4 out of 151 investigated cells still responded and showed a smallerrise in ratio compared to untreated cells (Data not shown).
Adenosine A3 receptor-induced CCL2 release
67
A B
20 sec
0.02
CPA
0
40
60
80
20
resp
ondi
ng c
ells
(% o
f tot
al)
CPACI-IB-MECA
- PTX+ PTX
*20 sec
0.02
CI-IB-MECA+ PTX
C D
20 sec
0.02
CI-IB-MECA
Figure 3.4. Effect of adenosine receptor stimulation and PTX treatment on intracellular calciumtransients in cultured murine astrocytes. A) Typical intracellular calcium response induced by 10µM Cl-IB-MECA. B) Stimulation with 10 µM CPA also induced intracellular calcium transients inmurine astrocytes. C) Typical intracellular calcium response induced by 10 µM Cl-IB-MECA afterpre-treatment with PTX (200 ng/ml). D) Bar graph showing the percentage of cells responding toCl-IB-MECA and CPA in the presence or absence of PTX pre-treatment. Data in D are representedas mean ± S.E.M. from 6-12 experiments. *, significant effect of PTX treatment (p < 0.05). Thearrow in A-C indicates the moment of stimulation.

Discussion
The neuromodulator adenosine has prominent neuroprotective effects inneurodegenerative diseases by directly inhibiting excitatory neurotransmitterrelease and by inducing neuronal hyperpolarization [20, 57]. Recent evidencesuggests that neuroprotection by adenosine might also occur indirectly byinduction of the release of neuroprotective substances from glia cells [13, 23, 35,59]. In damaged neural tissue the chemokine CCL2 is predominately expressed inglia cells (see for example [29]) and recently direct neuroprotective effects of thischemokine have been published [11, 21]. However, little is yet known on theregulation of glial CCL2 expression. We therefore investigated the possible effectof adenosine receptor stimulation on glial CCL2 synthesis. We show thatstimulation of mixed mouse astrocyte cultures with the selective adenosine A3
receptor ligand Cl-IB-MECA induces a significant release of CCL2. Other selectiveadenosine receptor agonists (CPA; adenosine A1 receptors and CGS 21680;adenosine A2A receptors) did not have any effect on CCL2 release in mixedastrocytic cultures, indicating that CCL2 release is induced by stimulation ofadenosine A3 receptors. Most likely, the effect of the non-selective adenosineagonist NECA is also due to activation of adenosine A3 receptors, since NECAhas an intermediate affinity for this receptor subtype [27, 28, 68]. Antagoniststudies further corroborated the involvement of adenosine A3 receptors. The non-selective antagonist 8PT, which has high affinity for A1- and A2A-, intermediateaffinity for A2B- but very low affinity for A3 adenosine receptors [27], did noteffect Cl-IB-MECA-induced CCL2 release. In contrast, 10 nM of the selectiveadenosine A3 receptor antagonist, MRS 1523 [26, 49, 55], inhibited the effect ofCl-IB-MECA on CCL2 release. The affinity of MRS 1523 at mouse adenosine A3
receptors is not known yet. Ki -values for MRS 1523 at rat and human A3
receptors were 113 and 18.9 nM respectively [44], indicating species dependentdifferences of the affinity of MRS 1523. Similar to our findings a maximal effect ofMRS 1523 already with 10 nM was reported by Reshkin and colleagues [55] inA6 cells, a renal cell line derived from Xenopus Laevis. Thus, MRS 1523 can blockadenosine A3 receptor-mediated responses at low concentrations. Taken together the results obtained by RT-PCR analysis and in situ hybridizationand the pharmacological data confirm the expression of the adenosine A3 receptorin mouse astrocytes and its involvement in the release of CCL2. The effect ofadenosine A3 receptor ligands on the release of CCL2 was cell type specific.Microglia that are known to release CCL2 upon a variety of stimuli (for reviewsee [7]) did not release CCL2 in response to Cl-IB-MECA, although adenosine A3
receptors are expressed in these cells and functional as published recently (thiswork and [24, 32]).
Chapter 3
68

Cl-IB-MECA-induced CCL2 release in cultured astrocytes was not inhibited bypre-incubation with pertussis toxin, suggesting that Cl-IB-MECA-induced CCL2release through a Gi protein independent pathway. This is in agreement withprevious studies that describe various pertussis toxin insensitive adenosine A3
receptor mediated effects [43, 55]. Adenosine A3 receptors are reported to couplemainly to Gi or Gq proteins [52]. Since the involvement of Gi proteins in Cl-IB-MECA mediated CCL2 release in astrocytes seems to be unlikely, Gq proteinsmight be important here. The observed effects of Cl-IB-MECA on intracellularcalcium transients in astrocytes, which were completely insensitive to PTXtreatment, corroborate the assumption that adenosine A3 receptors in murineastrocytes are coupled to Gq proteins. On the other hand we also observedintracellular calcium transients in response to the adenosine A1 receptor agonistCPA. This effect, however, was most likely mediated by Gi proteins since it wascompletely blocked by PTX pre-treatment.Little is yet known about the physiological role of adenosine A3 receptors. Inperipheral tissues both beneficial as well as harmful effects of adenosine A3
receptor stimulation have been reported [22, 33]. This also is the case in CNStissues. In animal experiments it has been found that adenosine A3 receptorligands are neuroprotective in cerebral ischaemia models [65]. These findingshave recently been confirmed in adenosine A3 receptor deficient mice [22].Adenosine A3 receptor stimulation has been reported to regulate the survival ofglia cells. It was thus reported that administration of adenosine A3 receptorligands at nanomolar concentrations protected cultured astrocytes cells from celldeath, whereas stimulation with micromolar concentrations induced cell death[40, 48]. Also CCL2 induces both detrimental and beneficial effects in the brain. ThusCCL2 is clearly involved in neuroinflammation and several lines of evidencesuggest a function of CCL2 in the pathogenesis of neurodegenerative diseases[37, 41, 42, 54, 61, 62]. On the other hand there are also reports showing directneuroprotective effects of CCL2 in culture [11, 21]. To what extent the effects of adenosine A3 receptor stimulation in vivo are causedby increased release of CCL2 is still unclear. This should be further investigatedin adenosine A3 receptor deficient- or CCL2 deficient mice [1, 2, 39]. In conclusion, the data presented here show that stimulation of adenosine A3
receptors in cultured astrocytes induced the release of CCL2. This effect is mostlikely mediated via Gq proteins and suggests a new function of adenosine A3
receptors in astrocytes. The here presented results furthermore corroborate theassumption that adenosine receptors in glia cells regulate the release of variousneuroprotective substances as it was already shown for NGF and IL-6 [23, 35,59].
Adenosine A3 receptor-induced CCL2 release
69

References
Chapter 3
70
1. Abbracchio, M. P., Ceruti, S., Brambilla, R., Barbieri, D., Camurri, A., Franceschi, C., Giammarioli, A. M., Jacobson, K. A., Cattabeni, F., and Malorni, W. 2003. Adenosine A3 receptors and viability of astrocytes. Drug development research 45: 379-386
2. Abbracchio, M. P., Saffrey, M. J., Hopker, V., and Burnstock, G. 1994. Modulation of astroglial cell proliferation by analogues of adenosine and ATP in primary cultures of rat striatum. Neuroscience59(1): 67-76
3. Asensio, V. C. and Campbell, I. L. 1999. Chemokines in the CNS: plurifunctional mediators in diverse states. Trends Neurosci. 22(11): 504-512
4. Bacon, K. B. and Harrison, J. K. 2000. Chemokines and their receptors in neurobiology: perspectivesin physiology and homeostasis. J Neuroimmunol. 104(1): 92-97
5. Baggiolini, M. 1998. Chemokines and leukocyte traffic. Nature 392(6676): 565-5686. Bajetto, A., Bonavia, R., Barbero, S., and Schettini, G. 2002. Characterization of chemokines and
their receptors in the central nervous system: physiopathological implications. J Neurochem. 82(6): 1311-1329
7. Biber, K. 2002. Microglial chemokines and chemokine receptors. In Universes in delicate balance: chemokines and the central nervous system, Ransohoff, R., Suzuki, K., Proudfoot, A. E. I., Hickey, W. F., and Harrison, J. K. 289-299. Amsterdam: Elsevier Science.
8. Biber, K., Klotz, K. N., Berger, M., Gebicke-Harter, P. J., and van Calker, D. 1997. Adenosine A1
receptor-mediated activation of phospholipase C in cultured astrocytes depends on the level of receptor expression. J Neurosci 17(13): 4956-4964
9. Biber, K., Sauter, A., Brouwer, N., Copray, S. C., and Boddeke, H. W. 2001. Ischemia-induced neuronal expression of the microglia attracting chemokine Secondary Lymphoid-tissue Chemokine (SLC). Glia 34(2): 121-133
10. Boddeke, E. W., Meigel, I., Frentzel, S., Gourmala, N. G., Harrison, J. K., Buttini, M., Spleiss, O., and Gebicke-Harter, P. 1999. Cultured rat microglia express functional beta-chemokine receptors.J Neuroimmunol. 98(2): 176-184
11. Bruno, V., Copani, A., Besong, G., Scoto, G., and Nicoletti, F. 2000. Neuroprotective activity of chemokines against N-methyl-D-aspartate or beta-amyloid-induced toxicity in culture.Eur J Pharmacol 399(2-3): 117-121
12. Chomczynski, P. and Sacchi, N. 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal.Biochem. 162(1): 156-159
13. Ciccarelli, R., Di Iorio, P., Bruno, V., Battaglia, G., D'Alimonte, I., D'Onofrio, M., Nicoletti, F., and Caciagli, F. 1999. Activation of A(1) adenosine or mGlu3 metabotropic glutamate receptors enhancesthe release of nerve growth factor and S-100beta protein from cultured astrocytes. Glia 27(3): 275-281
14. Copray, J. C. and Brouwer, N. 1994. Selective expression of neurotrophin-3 messenger RNA in muscle spindles of the rat. Neuroscience 63(4): 1125-1135
15. Cross, A. K. and Woodroofe, M. N. 1999. Chemokine modulation of matrix metalloproteinase and TIMP production in adult rat brain microglia and a human microglial cell line in vitro. Glia 28(3): 183-189
16. Cunha, R. A. 2001. Adenosine as a neuromodulator and as a homeostatic regulator in the nervous system: different roles, different sources and different receptors. Neurochem Int 38(2): 107-125
17. de Mendonca, A., Sebastiao, A. M., and Ribeiro, J. A. 2000. Adenosine: does it have a neuroprotective role after all? Brain Res Brain Res Rev 33(2-3): 258-274
18. Dirnagl, U., Niwa, K., Lindauer, U., and Villringer, A. 1994. Coupling of cerebral blood flow to neuronal activation: role of adenosine and nitric oxide. Am J Physiol 267(1 Pt 2): 296-301
19. Dorf, M. E., Berman, M. A., Tanabe, S., Heesen, M., and Luo, Y. 2000. Astrocytes express functionalchemokine receptors. J Neuroimmunol. 111(1-2): 109-121
20. Dunwiddie, T. V. 1985. The physiological role of adenosine in the central nervous system. Int Rev Neurobiol 27: 63-139

Adenosine A3 receptor-induced CCL2 release
71
21. Eugenin, E. A., D'Aversa, T. G., Lopez, L., Calderon, T. M., and Berman, J. W. 2003. MCP-1 (CCL2) protects human neurons and astrocytes from NMDA or HIV-tat-induced apoptosis. J Neurochem. 85(5): 1299-1311
22. Fedorova, I. M., Jacobson, M. A., Basile, A., and Jacobson, K. A. 2003. Behavioral characterization ofmice lacking the A3 adenosine receptor: sensitivity to hypoxic neurodegeneration. Cell Mol.Neurobiol.23(3): 431-447
23. Fiebich, B. L., Biber, K., Gyufko, K., Berger, M., Bauer, J., and van Calker, D. 1996. Adenosine A2b
receptors mediate an increase in interleukin (IL)-6 mRNA and IL-6 protein synthesis in human astroglioma cells. J Neurochem 66(4): 1426-1431
24. Fiebich, B. L., Biber, K., Lieb, K., van Calker, D., Berger, M., Bauer, J., and Gebicke-Haerter, P. J. 1996. Cyclooxygenase-2 expression in rat microglia is induced by adenosine A2a-receptors.Glia 18(2): 152-160
25. Fife, B. T., Huffnagle, G. B., Kuziel, W. A., and Karpus, W. J. 2000. CC chemokine receptor 2 is critical for induction of experimental autoimmune encephalomyelitis. J Exp.Med. 192(6): 899-905
26. Fishman, P., Bar-Yehuda, S., Barer, F., Madi, L., Multani, A. S., and Pathak, S. 2001. The A3
adenosine receptor as a new target for cancer therapy and chemoprotection. Exp.Cell Res. 269(2): 230-236
27. Fredholm, B. B., Abbracchio, M. P., Burnstock, G., Daly, J. W., Harden, T. K., Jacobson, K. A., Leff, P., and Williams, M. 1994. Nomenclature and classification of purinoceptors. Pharmacol.Rev. 46(2): 143-156
28. Fredholm, B. B., IJzerman, A.P., Jacobson, K. A., Klotz, K. N., and Linden, J. 2001. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors.Pharmacol Rev 53(4): 527-552
29. Glabinski, A. R., Balasingam, V., Tani, M., Kunkel, S. L., Strieter, R. M., Yong, V. W., and Ransohoff, R. M. 1996. Chemokine monocyte chemoattractant protein-1 is expressed by astrocytes after mechanical injury to the brain. J.Immunol. 156(11): 4363-4368
30. Guillemin, G. J., Croitoru-Lamoury, J., Dormont, D., Armati, P. J., and Brew, B. J. 2003. Quinolinic acid upregulates chemokine production and chemokine receptor expression in astrocytes.Glia 41(4): 371-381
31. Haas, H. L. and Selbach, O. 2000. Functions of neuronal adenosine receptors. Naunyn Schmiedebergs Arch Pharmacol 362(4-5): 375-381
32. Hammarberg, C., Schulte, G., and Fredholm, B. B. 2003. Evidence for functional adenosine A3
receptors in microglia cells. J Neurochem. 86(4): 1051-105433. Harrison, G. J., Cerniway, R. J., Peart, J., Berr, S. S., Ashton, K., Regan, S., Paul Matherne, G., and
Headrick, J. P. 2002. Effects of A(3) adenosine receptor activation and gene knock-out in ischemic-reperfused mouse heart. Cardiovasc Res 53(1): 147-155
34. Hayashi, M., Luo, Y., Laning, J., Strieter, R. M., and Dorf, M. E. 1995. Production and function of monocyte chemoattractant protein-1 and other beta-chemokines in murine glial cells.J Neuroimmunol 60(1-2): 143-150
35. Heese, K., Fiebich, B. L., Bauer, J., and Otten, U. 1997. Nerve growth factor (NGF) expression in ratmicroglia is induced by adenosine A2a-receptors. Neurosci Lett 231(2): 83-86
36. Hesselgesser, J. and Horuk, R. 1999. Chemokine and chemokine receptor expression in the central nervous system. J Neurovirol. 5(1): 13-26
37. Huang, D. R., Wang, J., Kivisakk, P., Rollins, B. J., and Ransohoff, R. M. 2001. Absence of monocytechemoattractant protein 1 in mice leads to decreased local macrophage recruitment and antigen-specific T helper cell type 1 immune response in experimental autoimmune encephalomyelitis.J Exp.Med. 193(6): 713-726
38. Izikson, L., Klein, R. S., Charo, I. F., Weiner, H. L., and Luster, A. D. 2000. Resistance to experimental autoimmune encephalomyelitis in mice lacking the CC chemokine receptor (CCR)2J Exp.Med. 192(7): 1075-1080
39. Jacobson, K. A. 1998. Adenosine A3 receptors: novel ligands and paradoxical effects.Trends Pharmacol Sci 19(5): 184-191

Chapter 3
72
40. Jacobson, K. A., Moro, S., Kim, Y. C., and Li, A. H. 2003. A3 adenosine receptors: Protective vs. damaging effects identified using novel agonists and antagonists. Drug development research 45: 113-124
41. Karpus, W. J. and Ransohoff, R. M. 1998. Chemokine regulation of experimental autoimmune encephalomyelitis: temporal and spatial expression patterns govern disease pathogenesis.J Immunol. 161(6): 2667-2671
42. Kelder, W., McArthur, J. C., Nance-Sproson, T., McClernon, D., and Griffin, D. E. 1998. Beta-chemokines MCP-1 and RANTES are selectively increased in cerebrospinal fluid of patients with human immunodeficiency virus-associated dementia. Ann.Neurol. 44(5): 831-835
43. Lee, J. E., Bokoch, G., and Liang, B. T. 2001. A novel cardioprotective role of RhoA: new signaling mechanism for adenosine. FASEB J. 15(11): 1886-1894
44. Li, A. H., Moro, S., Melman, N., Ji, X. D., and Jacobson, K. A. 1998. Structure-activity relationships and molecular modeling of 3, 5-diacyl-2,4-dialkylpyridine derivatives as selective A3 adenosine receptor antagonists. J.Med.Chem. 41(17): 3186-3201
45. Loddick, S. A., Turnbull, A. V., and Rothwell, N. J. 1998. Cerebral interleukin-6 is neuroprotective during permanent focal cerebral ischemia in the rat. J Cereb Blood Flow Metab 18(2): 176-179
46. Matsuda, S., Wen, T. C., Morita, F., Otsuka, H., Igase, K., Yoshimura, H., and Sakanaka, M. 1996. Interleukin-6 prevents ischemia-induced learning disability and neuronal and synaptic loss in gerbils. Neurosci Lett 204(1-2): 109-112
47. Mendonca Torres, P. M. and de Araujo, E. G. 2001. Interleukin-6 increases the survival of retinal ganglion cells in vitro. J Neuroimmunol 117(1-2): 43-50
48. Merighi, S., Mirandola, P., Milani, D., Varani, K., Gessi, S., Klotz, K. N., Leung, E., Baraldi, P. G., and Borea, P. A. 2002. Adenosine receptors as mediators of both cell proliferation and cell death of cultured human melanoma cells. J.Invest Dermatol. 119(4): 923-933
49. Mitchell, C. H., Peterson-Yantorno, K., Carre, D. A., McGlinn, A. M., Coca-Prados, M., Stone, R. A., and Civan, M. M. 1999. A3 adenosine receptors regulate Cl- channels of nonpigmented ciliary epithelial cells. Am.J.Physiol 276(3 Pt 1): C659-C666
50. Mubagwa, K. and Flameng, W. 2001. Adenosine, adenosine receptors and myocardial protection: anupdated overview. Cardiovasc Res 52(1): 25-39
51. Murphy, P. M., Baggiolini, M., Charo, I. F., Hebert, C. A., Horuk, R., Matsushima, K., Miller, L. H., Oppenheim, J. J., and Power, C. A. 2000. International union of pharmacology. XXII. Nomenclature for chemokine receptors. Pharmacol.Rev. 52(1): 145-176
52. Palmer, T. M., Gettys, T. W., and Stiles, G. L. 1995. Differential interaction with and regulation of multiple G-proteins by the rat A3 adenosine receptor. J Biol.Chem. 270(28): 16895-16902
53. Penkowa, M., Molinero, A., Carrasco, J., and Hidalgo, J. 2001. Interleukin-6 deficiency reduces the brain inflammatory response and increases oxidative stress and neurodegeneration after kainic acid-induced seizures. Neuroscience 102(4): 805-18
54. Ransohoff, R. M., Hamilton, T. A., Tani, M., Stoler, M. H., Shick, H. E., Major, J. A., Estes, M. L., Thomas, D. M., and Tuohy, V. K. 1993. Astrocyte expression of mRNA encoding cytokines IP-10 and JE/MCP-1 in experimental autoimmune encephalomyelitis. Faseb J 7(6): 592-600
55. Reshkin, S. J., Guerra, L., Bagorda, A., Debellis, L., Cardone, R., Li, A. H., Jacobson, K. A., and Casavola, V. 2000. Activation of A(3) adenosine receptor induces calcium entry and chloride secretion in A(6) cells. J Membr Biol 178(2): 103-113
56. Rollins, B. J. 1997. Chemokines. Blood 90(3): 909-92857. Rudolphi, K. A. and Schubert, P. 1996. Purinergic interventions in traumatic and ischemic injury. In
Novel Thearapies for CNS Injuries, Peterson, P. L. and Willis, P. W. 327-342. Boca Raton, New York, London, Tokyo: CRC Press Inc.
58. Rudolphi, K. A., Schubert, P., Parkinson, F. E., and Fredholm, B. B. 1992. Neuroprotective role of adenosine in cerebral ischaemia. Trends Pharmacol Sci 13(12): 439-445
59. Schwaninger, M., Neher, M., Viegas, E., Schneider, A., and Spranger, M. 1997. Stimulation of interleukin-6 secretion and gene transcription in primary astrocytes by adenosine. J Neurochem69(3): 1145-1150

Adenosine A3 receptor-induced CCL2 release
73
60. Siebert, H., Sachse, A., Kuziel, W. A., Maeda, N., and Bruck, W. 2000. The chemokine receptor CCR2 is involved in macrophage recruitment to the injured peripheral nervous system.J Neuroimmunol. 110(1-2): 177-185
61. Sorensen, T. L., Sellebjerg, F., Jensen, C. V., Strieter, R. M., and Ransohoff, R. M. 2001. Chemokines CXCL10 and CCL2: differential involvement in intrathecal inflammation in multiple sclerosis. Eur J Neurol 8(6): 665-672
62. Sorensen, T. L., Tani, M., Jensen, J., Pierce, V., Lucchinetti, C., Folcik, V. A., Qin, S., Rottman, J., Sellebjerg, F., Strieter, R. M., Frederiksen, J. L., and Ransohoff, R. M. 1999. Expression of specific chemokines and chemokine receptors in the central nervous system of multiple sclerosis patients.J Clin.Invest 103(6): 807-815
63. Swartz, K. R., Liu, F., Sewell, D., Schochet, T., Campbell, I., Sandor, M., and Fabry, Z. 2001. Interleukin-6 promotes post-traumatic healing in the central nervous system. Brain Res 896(1-2): 86-95
64. von Lubitz, D. K., Lin, R. C., Bischofberger, N., Beenhakker, M., Boyd, M., Lipartowska, R., and Jacobson, K. A. 1999. Protection against ischemic damage by adenosine amine congener, a potent and selective adenosine A1 receptor agonist. Eur J Pharmacol 369(3): 313-317
65. von Lubitz, D. K., Lin, R. C., Popik, P., Carter, M. F., and Jacobson, K. A. 1994. Adenosine A3
receptor stimulation and cerebral ischemia. Eur.J Pharmacol. 263(1-2): 59-6766. Williams, Michael. 1989. Adenosine: the prototypic neuromodulator. Neurochemistry international
14(3): 249-26467. Williams, M. 1995. Purinoceptors in Central Nervous System Function: targets for Therapeutic
intervention. In Psychopharmacology: The Fourth Generation of Progress, Bloom FE, Kupfer DJ 643-656. New York: Raven Press. Ltd.
68. Zhou, Q. Y., Li, C., Olah, M. E., Johnson, R. A., Stiles, G. L., and Civelli, O. 1992. Molecular cloningand characterization of an adenosine receptor: the A3 adenosine receptor. Proc.Natl.Acad.Sci.U.S.A 89(16): 7432-7436

74

75
Published in European Journal of Pharmacology. 2004. 487: 73-79.
The mouse brain adenosine A1 receptor:functional expression and
pharmacology
M.C. Wittendorp, J. Von Frijtag Drabbe Künzel, A.P. IJzerman,H.W.G.M. Boddeke and K. Biber
Chapter 4

Summary
Introduction
The four distinct adenosine receptor subtypes, A1, A2A, A2B and A3, belong to thefamily of G-protein coupled receptors [11]. Originally adenosine A1 and A3
receptors were described to interact mainly with Gi-proteins and induceinhibition of adenylyl cyclase whereas adenosine A2A and A2B receptors arecoupled mainly to Gs-proteins, consequently stimulate adenylyl cyclase andincrease cAMP levels [12, 32]. However, since adenosine receptors have also beenreported to interact with different G-proteins and signal through various otherpathways, independent of adenylyl cyclase, as reviewed recently [37] the abovementioned classification should be regarded as historical and may not solelyreflect the situation in vivo.Since adenosine was first described to be involved cardiovascular regulation [9],extensive research has been performed and a large number of physiologicalfunctions of adenosine has been described (for reviews see: [10, 17, 22, 28, 35,38, 42]). After the cloning of the first adenosine receptor [20], differentadenosine receptor subtypes have been characterized. The adenosine A1 receptorhas been cloned from several species: [1, 21, 25, 27, 29, 31, 33, 43, 45]. Fewerreports described the cloning of adenosine A3 [29, 43, 45], adenosine A2A [26]and adenosine A2B [33] receptor subtypes.Mouse strains with targeted deletions of adenosine A1, A2A and A3 receptorsubtypes have been generated [16, 19, 44]. These transgenic mice prove to be auseful tool to unravel the function of these adenosine receptor subtypes [30].Despite the large interest in mouse adenosine A1 receptors, its pharmacologicalproperties are largely unknown [12]. This is in contrast to the human and ratadenosine receptors, which have been well characterized in pharmacologicalstudies. In order to elucidate the pharmacological properties of the mouse
Chapter 4
76
The adenosinergic system is involved in many important physiological functions. Adenosineexerts its extracellular effects through four types of G-protein coupled receptors: A1, A2A, A2B andA3. Adenosine acts as an important regulator of metabolic processes. In the brain adenosinemediates prominent neuroprotective functions via the adenosine A1 receptor. Whereas thepharmacological characteristics of the rat and human adenosine A1 receptor have been intensivelystudied, the mouse adenosine A1 receptor has not yet been characterized. Accordingly, we havecloned the mouse brain adenosine A1 receptor and present here a pharmacologicalcharacterization of the mouse adenosine A1 receptor using functional studies and radioligandbinding assays. The results show that the binding affinities of several ligands for the mouseadenosine A1 receptor are similar to the affinities for the rat and human adenosine A1 receptorwith some exceptions.

adenosine A1 receptor, we have cloned the adenosine A1 receptor from mousebrain and expressed it in Chinese Hamster Ovary (CHO) cells for pharma-cological characterization and functional studies. Additionally, further bindingstudies were performed on rat and human adenosine A1 receptor in order tocompare the binding affinities for all three species.
Materials and methods
Reagents
Phosphate-buffered saline, Dulbecco’s Modified Eagle medium (DMEM), DMEM-F12, fetal calf serum and penicillin/streptomycin were obtained from Gibco.Reverse transcriptase enzyme and buffer were purchased from Promega. Taq-polymerase and buffer for polymerase chain reaction (PCR) were obtained fromEppendorf (Boulder, CO, U.S.A). PCR-primers were ordered from Genset oligo’s.N6-cyclopentyladenosine (CPA), R-N6-phenylisopropyladenosine (R-PIA), cyclo-pentyltheophylline (CPT), N6-cyclopentyl-9-methyladenine (N-0840) and 8-cyclopentyl-1,3-dipropylxanthine (DPCPX) were obtained from RBI (Natick, MA,U.S.A.). N-ethylcarboxamidoadenosine (NECA), 2-chloro-N6-cyclopentylade-nosine (CCPA) and forskolin were purchased from Sigma (St.Louis, MO, U.S.A.),while adenosine deaminase was from Roche Biochemicals (Mannheim, Germany).2-[p-(2-carboxyethyl)phenyl-ethylamino]-5'-N-ethylcarboxamidoadenosine(CGS21680) was from Ciba Geigy. 4-(2-[7-amino-2-(2-furyl)[1,2,4]triazolo[2,3-a][1,3,5]triazin-5-ylamino]ethyl)phenol (ZM241358) was from Zeneca Pharma-ceuticals. [3H]DPCPX was purchased from Amersham, whereas [3H]cAMP wasfrom NEN (Du Pont Nemours, ‘s-Hertogenbosch, NL). GTP was obtained fromAldrich. 2-chloro- N6-(3-iodobenzyl)-N-methyl-5’-carbamoyladenosine (Cl-IB-MECA) was a kind gift from K.A. Jacobson, NIH, USA.
Glial cultures
Mixed astrocyte cell cultures were established as described previously [3]. Inbrief, mouse cortex was dissected from new-born mouse pups (<1 day). Braintissue was gently dissociated by trituration in phosphate-buffered saline andfiltered through a cell strainer (70 mm Ø, Falcon) into Dulbecco’s Modified Eaglemedium (DMEM). After two washing steps (200 x g for 10 min), cells wereseeded in culture flasks (75 cm2, Greiner; 10 x 106 cells/flask). Cultures weremaintained up to 4 weeks in DMEM containing 10% fetal calf serum with 0.01%penicillin and 0.01% streptomycin in a humidified atmosphere (5% CO2) at 37ºC.Culture medium was changed the second day after preparation and every 6 daysthereafter.
The mouse brain adenosine A1 receptor
77

Reverse transcription-polymerase chain reaction (RT-PCR)
Cells were lysed in guanidinium isothiocyanate/mercaptoethanol buffer and totalRNA was extracted with one phenol/chloroform step, precipitated according toChomczynski and Sacchi [5]. Subsequently RNA was treated with RNAse freeDNAse (Sigma-Aldrich, Bornhem, Belgium) for 2 hours and precipitated. Reversetranscription (RT): 1 µg of total RNA was transcribed into cDNA in a totalvolume of 25 µl containing 0.5 µl of M-MLV reverse transcriptase, 1 µl of RNaseinhibitor, 1 µl of random hexamers (0.2 mM), 5 µl of 5x buffer, 5 µl ofdeoxynucleosidetriphosphates (dNTPs) (2.5 mM) and H2O adjusted to 25 µl.After 60 min at 42°C, the reaction was stopped by heating at 95°C for 5 min. Thequality of the cDNA was controlled using glyceraldehyde-3-phosphatedehydrogenase (GAPDH) primers [4] and potential contamination by genomicDNA was checked for by running the reactions without reverse transcriptase andusing GAPDH primers in subsequent polymerase chain reaction (PCR)amplifications. PCR: 2 µl of the RT reaction was used in subsequent PCRamplification as described previously [3]. In brief, the following reagents wereadded: 5 µl 10x PCR-buffer, 2.5 µl MgCl2 (50 mM), 0.5 µl dNTPs (10 mM), 1 µlof each primer, 38 µl H2O and 0.1 µl Taq-polymerase. Primer sequences formouse full-length adenosine A1 receptor:forward primer: 5’-CATGCCGCCGTACATCTC-3’;backward primer: 5’-TCTAGTCCTCAGCTTTCTCCTC-3’.Cycle numbers were 35 and annealing temperature was 58ºC. The sequence ofthe PCR product was verified by TA cloning into pCRII (Invitrogen). In brief PCRproducts were stored on ice after the amplification. 20 µl of the resulting PCRproduct was checked by gel electrophoresis and 2 µl was used for ligation intolinearized pCRII (Invitrogen) according to the instructions of the manufacturer.The ligation product was used to transform competent bacteria (TOP-10F,Invitrogen) and resulting bacterial colonies were grown overnight in 5 ml LBmedium. Plasmid preparation was done by standard methods and positiveplasmids were sequenced by ALF (sequencing facility; University of Groningen).A XhoI /BamHI fragment, containing the full-length adenosine A1 receptorcoding region, was excised from the pCRII vector clone and ligated into a XhoI/BamHI digested pcDNA3.1(-) expression vector (Invitrogen), incorporating aneomycin resistance gene. Sequence analysis was performed to check theorientation of the adenosine A1 receptor insert.
Stable expression in Chinese hamster ovary (CHO) cells
CHO cells were cultured at 37ºC in cell culture flasks (25 cm2, Corning),containing 5 ml DMEM-F12 medium, supplemented with 10% fetal calf serumand 2 mM L-glutamine in a humidified atmosphere (5% CO2). Cells were seededinto a six-well dish and transfected with 1 µg of the pcDNA3.1(-)-mouse
Chapter 4
78

adenosine A1 receptor expression vector, using 3 µl of fuGENE 6 transfectionreagent (Roche). After transfection, cells were seeded in cell culture flasks. After2 weeks of selection with 50 µg/ml G 418, monoclonal CHO cell lines, stablyexpressing the mouse adenosine A1 receptor (CHO-mA1), were generated andadenosine A1 receptor mRNA expression levels of 16 clones were determined byRT-PCR. The CHO-mA1 cell line with highest expression levels was used forbinding studies and cAMP measurements.
Radioligand binding assay
For membrane preparation CHO-mA1 cells were cultured in cell culture flasks(75 cm2, Greiner), grown to confluency and harvested by trypsinization for 30 secwith 0.25% w/v porcine trypsin at 37°C. The cell suspension was spun for 10min at 500 g. After removing the supernatant, the pellet was resuspended in 50mM Tris/HCl pH 7.4 (at 25°C, approx. 10 x 106 cells/ml) and spun for 40 min(4°C) at 18,000 g.After discarding the supernatant, the pellet was resuspended in 50 mM Tris/HClpH 7.4 (at 4°C, approx. 20 x 106 cells/ml) and stored at –80°C until analysis.Rat cortical membranes were prepared according to the method of Lohse andcolleagues [23] and incubated with 2 IU/ml adenosine deaminase for 30 min at37°C before storage [34]. CHO-hA1 cell membranes were prepared as describedpreviously [7]. For displacement studies, membranes (75 µg) were incubated for1 hour at 25°C in 50 mM TrisHCl (pH 7.4) in the presence of 1.6 nM[3H]DPCPX and different concentrations of ligand to determine the Ki. Todetermine non-specific binding 100 µM CPA was used. Total volume duringincubation was 200 µl.Incubations were stopped by rapid dilution with 1 ml ice-cold buffer and boundradioligand was subsequently recovered by filtration through Whatman GF/Bfilters using either Millipore system or Brandel Harvester under reduced pressure.Filters were then washed three times with 2 ml buffer. The retained radioactivitywas measured by liquid scintillation counting (LKB Wallac, 1219 Rackbeta).Saturation experiments were carried out under the similar conditions. Increasingconcentrations of [3H]DPCPX (0 – 4 nM) were used. Filters were washed fivetimes with 2.5 ml buffer to remove excess of radioligand.
cAMP-accumulation assay
CHO-mA1 cells were seeded in 24 wells plate at a density of 2 x 105 cells/well.The next day, growth medium was aspirated and washed twice withDMEM/HEPES (pH 7.4). Cells were then incubated for 30 min at 37°C withDMEM/HEPES supplemented with adenosine deaminase (2 IU/ml), cilostamide(50 µM) and rolipram (50 µM). After incubation different concentrations of CPA,ranging from 1 nM till 100 µM, were added for another 10 min. Subsequently,
The mouse brain adenosine A1 receptor
79

forskolin (10 µM) was added. After another 15 min, the cAMP generation wasstopped by aspirating incubation medium and adding 200 µl ice-cold 0.1 N HCl tothe cells.The amount of cAMP was determined by competition with [3H]cAMP for proteinkinase A binding protein (PKA). Briefly, sample or cAMP standard (0 – 16 pmol),1.8 nM [3H]cAMP and PKA-solution were incubated on ice for at least 2.5 hours.The incubation was stopped by dilution with ice-cold TrisHCl buffer (pH 7.4).Bound radioactivity was recovered by filtration through Whatman GF/C filtersusing a Brandel harvester.
Data analysis
All radioligand binding data were analyzed using the Software program PRISM3.0 (GraphPad, San Diego, CA), based on non-linear curve fitting procedures,yielding KD and Bmax values in case of saturation binding experiments and Ki
values in case of displacement studies. Data derived from cAMP determinationswere also analyzed using PRISM (GraphPad, San Diego, CA), providing EC50
values.
Results
Sequence
The sequence that was obtained for the present adenosine A1 receptor genecloned from mouse brain (Figure 4.1, accession number (AC): AJ555877) showed99% similarity with the sequence suggested to be the mouse adenosine A1
receptor previously cloned by Marquardt and colleagues ([26], AC: U05671), 95%similarity with the rat adenosine A1 receptor (AC: M69045) and 89% similaritywith the human adenosine A1 receptor (AC: S56143). See Figure 4.1 for analignment of the protein sequences of the mouse, rat and human adenosine A1
receptor. The previously cloned mouse adenosine A1 receptor [26] differs fromthe present receptor sequence at position 10. We describe here an alanine, whichis also found in the rat and human sequence, whereas the previously describedsequence showed a glycine at this position.
Radioligand binding
Saturation binding experiments were performed at CHO-mA1 membranes, usingthe ligand [3H]DPCPX. A typical experiment is shown in Figure 4.2A. Analysis ofthree experiments resulted in a KD of 0.53 nM with a 95% confidence limit of0.44-0.63 nM and a Bmax of 97 ± 60 (standard deviation) fmol/mg. Competitionbinding experiments were performed with the following ligands: the selectiveadenosine A1 receptor agonists CCPA, CPA and R-PIA, the non-selective adenosine
Chapter 4
80

receptor agonist NECA and the selective adenosine A1 receptor antagonists CPT,DPCPX and N0840. Furthermore binding affinities towards the mouse adenosineA1 receptor were determined for ligands selective for other adenosine receptors:the adenosine A2A receptor agonist CGS21680, the adenosine A2A receptorantagonist ZM241385 and the selective adenosine A3 receptor agonist Cl-IB-MECA. Figure 4.2B shows a representative competition experiment of increasingconcentrations of selective agonists CPA and R-PIA with the radioligand[3H]DPCPX. Adding GTP (1 mM) shifted the displacement curve of CPA to theright, resulting in a Ki value of 4.2 ± 2.7 nM in the control situation in contrast toa Ki of 17 ± 8 nM in the presence of GTP. This finding suggested agonisticactivity of CPA at CHO-mA1 membranes, which was further corroborated byfunctional studies (see below). An overview of the affinity values of the bindingassays is provided in Table 4.1.
The mouse brain adenosine A1 receptor
81
mouse MPPYISAFQA AYIGIEVLIA LVSVPGNVLV IWAVKVNQAL RDATFCFIVS 50
mouse LAVADVAVGA LVIPLAILIN IGPQTYFHTC LMVACPVLIL TQSSILALLA 100
mouse IAVDRYLRVK IPLRYKTVVT QRRAAVAIAG CWILSLVVGL TPMFGWNNLS 150
mouse EVEQAWIANG SVGEPVIKCE FEKVISMEYM VYFNFFVWVL PPLLLMVLIY 200
mouse LEVFYLIRKQ LNKKVSASSG DPQKYYGKEL KIAKSLALIL FLFALSWPPL 250
mouse HILNCITLFC PTCQKPSILI YIAFLTHGN SAMNPIVYAF RIHKFRVTFL 300
mouse KIWNDHFRCQ PKPPIEEDIP EEKAED 326
rat MPPYISAFQA AYIGIEVLIA LVSVPGNVLV IWAVKVNQAL RDATFCFIVS
rat LAVADVAVGA LVIPLAILIN IGPQTYFHTC LMVACPVLIL TQSSILALLA
rat IAVDRYLRVK IPLRYKTVVT QRRAAVAIAG CWILSLVVGL TPMFGWNNLS
rat VVEQDWRANG SVGEPVIKCE FEKVISMEYM VYFNFFVWVL PPLLLMVLIY
rat LEVFYLIRKQ LNKKVSASSG DPQKYYGKEL KIAKSLALIL FLFALSWPPL
rat HILNCITLFC PTCQKPSILI YIAFLTHGN SAMNPIVYAF RIHKFRVTFL
rat KIWNDHFRCQ PKPPIDEDLP EEKAED
human MPPSISAFQA AYIGIEVLIA LVSVPGNVLV IWAVKVNQAL RDATFCFIVS
human LAVADVAVGA LVIPLAILIN IGPQTYFHTC LMVACPVLIL TQSSILALLA
human IAVDRYLRVK IPLRYKMVVT PRRAAVAIAG CWILSFVVGL TPMFGWNNLS
human AVERAWAANG SMGEPVIKCE FEKVISMEYM VYFNFFVWVL PPLLLMVLIY
human LEVFYLIRKQ LNKKVSASSG DPQKYYGKEL KIAKSLALIL FLFALSWPPL
human HILNCITLFC PSCHKPSILT YIAFLTHGN SAMNPIVYAF RIQKFRVTFL
human KIWNDHFRCQ PAPPIDEDLP EERPDD
Figure 4.1. Alignment of adenosine A1 receptor amino acid sequences for mouse (AC: AJ555877),rat (AC: M69045) and human (AC: S56143). Differences between the mouse and rat sequence areshown in black. Differences between the human sequence compared to rat and/or mouse areshaded. The mouse protein sequence presented here was identical to the previously describedmouse sequence (AC:U05671) except at position 10 (underlined) where a glycine was found at thisposition instead of the alanine in the rat, human and the mouse sequence presented here.

Chapter 4
82
non-specific binding
B
00
2500
5000
7500[3 H
]DPC
PX b
indi
ng (d
pm)
1 2 3 4concentration [3H]DPCPX (nM)
Atotal binding
0
25
50
75
% s
peci
fic b
indi
ng [3 H
]DPC
PX]
-12 -10 -8 -6log [ligand] (M)
CPACPA + GTPR-PIA
-4
100specific binding
Figure 4.2. A) Saturation analysis of [3H]DPCPX binding to membranes of CHO cells expressingthe mouse adenosine A1 receptor. A representative experiment is shown. B) Displacement studiesof specific [3H]DPCPX binding (1.6 nM) from membranes of CHO cells expressing the mouseadenosine A1 receptor by CPA, CPA + GTP and by R-PIA. A representative experiment is shown.
Table 4.1. Binding affinities (Ki values ± S.E.M. in nM or % displacement) of reference ligandsfor adenosine A1 receptors from mouse, rat and human.
mousea ratb humanc
CCPA 21 ± 2 8.1 ± 0.4 6.4 ± 1.8d
CPA 4.2 ± 2.7 6.1 ± 0.8 10 ± 1R-PIA 14 ± 3 11 ± 5e n.d.NECA 238 ± 55 98 ± 22 12 (9.6-15)d
DPCPX 1.4 ± 0.1 0.73 ± 0.08 1.6 ± 0.18-CPT 12 ± 2 11 ± 3 36 ± 2N0840 303 ± 70 182 ± 53 1081 ± 69
ZM 241385 116 ± 18 n.d. n.d.CGS 21680 3 - 5 % (10-6)f n.d. n.d.
18 – 20 % (10-5)f n.d. n.d.Cl-IB-MECA 10 - 12 % (10-6)f n.d. n.d.
45 - 49 % (10-5)f n.d. n.d.
a membranes of CHO cells expressing the mouse adenosine A1 receptorb rat brain cortical membranesc membranes of CHO cells expressing the human adenosine A1 receptord Ki value with 95% confidence interval; value taken from [2]e value taken from [41]f % displacement of specific binding at the designated molar concentration (n=2)n.d. = not determined

cAMP measurements
In recombinant CHO cells stably expressing the mouse adenosine A1 receptorbaseline cAMP levels were too low to allow analysis of A1 receptor-inducedinhibition of cAMP production. Therefore, cAMP levels were raised by addingforskolin at a concentration of 10 µM. Co-administration of 1 nM up to 100 µM ofthe selective adenosine A1 receptor agonist CPA concentration-dependentlyinhibited the forskolin-induced cAMP accumulation in CHO-mA1 cells (Figure4.3). An EC50 value of 16.7 ± 2.2 (S.E.M.) nM was calculated from 3 independentexperiments.
Discussion
The adenosinergic system has been studied intensively for several decades. Alarge number of physiological and metabolic functions of adenosine have beendescribed. Thus adenosine is essentially involved in the coupling of metabolicrequirement to energy supply. In order to control metabolic rate, adenosine forexample, retains neuronal firing, increases heart rate and increases blood flow byinducing vasodilatation of smooth muscle cells [6, 8, 15, 28, 39]. Furthermore,adenosine is also involved in other functions such as inducing sleep and causinganalgesic effects and anti-inflammatory effects [10, 13, 22]. Especially in the brain adenosine shows its importance as a metabolic controlfactor. Adenosine inhibits the release of excitatory neurotransmitters and causesa hyperpolarization of the membrane potential in neurons. Adenosine exertsthese neuroprotective effects mainly by stimulation of adenosine A1 receptors
The mouse brain adenosine A1 receptor
83
0
20
40
80
% o
f for
skol
in-in
duce
d cA
MP
prod
uctio
n
-10 -8 -6log [CPA] (M)
-4
100
60
Figure 4.3. Inhibition of forskolin-stimulated cAMP production by CPA in CHO cells expressingthe mouse adenosine A1 receptor. A representative experiment is shown.

present throughout the brain [10, 36]. Although adenosine is obviously involvedin essential physiological functions, studies show that adenosine A1 receptordeficient mice function relatively normal compared to wild type mice [14, 16].However, exposed to pathophysiological conditions like hypoxia, mice lackingadenosine A1 receptors show more neuronal damage and have a lower survivalrate. It is therefore concluded that adenosine A1 receptors are primarily importantin mediating effects of adenosine during pathophysiological conditions [14, 16].Despite the fact that human and rat adenosine receptors have been extensivelycharacterized, the pharmacological properties of the mouse adenosine receptorshave not yet been investigated in detail compared to other species [12]. The twoprevious studies that investigated binding affinities of adenosine A1 receptorligands in membrane fractions from fresh material, used only a limited set ofadenosine receptor ligands [24, 40]. Therefore, we have cloned the adenosine A1
receptor from mouse brain, expressed it in CHO cells, a cell line that has beensuccessfully used in a detailed study on rat and human adenosine receptors [18]and studied its characteristics in radioligand binding assays and secondmessenger studies.The sequence of the present adenosine A1 receptor gene cloned from mouse brainwas almost identical to the mouse adenosine A1 receptor gene that waspreviously described by Marquardt and colleagues [26] and the sequence wassimilar to the rat and human adenosine A1 receptor sequence. The radioligandbinding assays revealed that the binding characteristics of the mouse receptor inthis study are similar to the characteristics of the rat and human adenosine A1
receptor (Table 4.1), although the non-selective ligand NECA showed a slightlylower affinity for the mouse receptor than for the rat receptor. Moreover,inhibition of cAMP production was observed after receptor stimulation with CPAwith an EC50 value of 16.7 ± 2.2 nM. The results of the radioligand binding andcAMP assays show that ligands commonly used to study the rat and humanadenosine A1 receptor have similar binding affinities to the mouse adenosine A1
receptor. It is thus concluded that these ligands can be used to study pharma-cological and physiological effects of the mouse adenosine A1 receptor.In summary, we have cloned the mouse adenosine A1 receptor, which showed tobe functional after expression in CHO cells. Our results provide a specification ofmouse adenosine A1 receptor pharmacology. This information will be of use forthe characterization of physiological effects of the adenosine A1 receptor inmouse models.
Chapter 4
84

References
The mouse brain adenosine A1 receptor
85
1. Aguilar, J. S., Tan, F., Durand, I., and Green, R. D. 1995. Isolation and characterization of an avian A1 adenosine receptor gene and a related cDNA clone. Biochem.J. 307 ( Pt 3): 729-734
2. Beukers, M. W., Wanner, M. J., von Frijtag Drabbe Kunzel JK, Klaasse, E. C., IJzerman, A. P., and Koomen, G. J. 2003. N6-cyclopentyl-2-(3-phenylaminocarbonyltriazene-1-yl)adenosine (TCPA), a very selective agonist with high affinity for the human adenosine A1 receptor. J Med.Chem. 46(8): 1492-1503
3. Biber, K., Klotz, K. N., Berger, M., Gebicke-Harter, P. J., and van Calker, D. 1997. Adenosine A1
receptor-mediated activation of phospholipase C in cultured astrocytes depends on the level of receptor expression. J Neurosci 17(13): 4956-4964
4. Biber, K., Sauter, A., Brouwer, N., Copray, S. C., and Boddeke, H. W. 2001. Ischemia-induced neuronal expression of the microglia attracting chemokine Secondary Lymphoid-tissue Chemokine (SLC). Glia 34(2): 121-133
5. Chomczynski, P. and Sacchi, N. 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal.Biochem. 162(1): 156-159
6. Cunha, R. A. 2001. Adenosine as a neuromodulator and as a homeostatic regulator in the nervous system: different roles, different sources and different receptors. Neurochem Int 38(2): 107-125
7. Dalpiaz, A., Townsend-Nicholson, A., Beukers, M. W., Schofield, P. R., and IJzerman, A. P. 1998. Thermodynamics of full agonist, partial agonist, and antagonist binding to wild-type and mutant adenosine A1 receptors. Biochem.Pharmacol. 56(11): 1437-1445
8. Dirnagl, U., Niwa, K., Lindauer, U., and Villringer, A. 1994. Coupling of cerebral blood flow to neuronal activation: role of adenosine and nitric oxide. Am J Physiol 267(1 Pt 2): 296-301
9. Drury, A. N. and Szent-Gyorgyi, A. 1929. The physiological activity of adenine compounds with especial reference to their action upon the mammalian heart. J Physiol (Lond) 68: 213-237
10. Dunwiddie, T. V. 1985. The physiological role of adenosine in the central nervous system. Int Rev Neurobiol 27: 63-139
11. Fredholm, B. B., Abbracchio, M. P., Burnstock, G., Daly, J. W., Harden, T. K., Jacobson, K. A., Leff, P., and Williams, M. 1994. Nomenclature and classification of purinoceptors. Pharmacol.Rev. 46(2): 143-156
12. Fredholm, B. B., IJzerman, A.P., Jacobson, K. A., Klotz, K. N., and Linden, J. 2001. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors.Pharmacol Rev 53(4): 527-552
13. Fredholm, B. B., Battig, K., Holmen, J., Nehlig, A., and Zvartau, E. E. 1999. Actions of caffeine in the brain with special reference to factors that contribute to its widespread use. Pharmacol Rev 51(1):83-133
14. Gimenez-Llort, L., Fernandez-Teruel, A., Escorihuela, R. M., Fredholm, B. B., Tobena, A., Pekny, M., and Johansson, B. 2002. Mice lacking the adenosine A1 receptor are anxious and aggressive, but are normal learners with reduced muscle strength and survival rate. Eur J Neurosci. 16(3): 547-550
15. Haas, H. L. and Selbach, O. 2000. Functions of neuronal adenosine receptors. Naunyn Schmiedebergs Arch Pharmacol 362(4-5): 375-381
16. Johansson, B., Halldner, L., Dunwiddie, T. V., Masino, S. A., Poelchen, W., Gimenez-Llort, L., Escorihuela, R. M., Fernandez-Teruel, A., Wiesenfeld-Hallin, Z., Xu, X. J., Hardemark, A., Betsholtz,C., Herlenius, E., and Fredholm, B. B. 2001. Hyperalgesia, anxiety, and decreased hypoxic neuro-protection in mice lacking the adenosine A1 receptor. Proc Natl Acad Sci U S A 98(16): 9407-9412
17. Kaiser, S. M. and Quinn, R. J. 1999. Adenosine receptors as potential therapeutic targets. Drug Discov Today 4(12): 542-551
18. Klotz, K. N., Hessling, J., Hegler, J., Owman, C., Kull, B., Fredholm, B. B., and Lohse, M. J. 1998. Comparative pharmacology of human adenosine receptor subtypes - characterization of stably transfected receptors in CHO cells. Naunyn Schmiedebergs Arch Pharmacol 357(1): 1-9

Chapter 4
86
19. Ledent, C., Vaugeois, J. M., Schiffmann, S. N., Pedrazzini, T., El Yacoubi, M., Vanderhaeghen, J. J., Costentin, J., Heath, J. K., Vassart, G., and Parmentier, M. 1997. Aggressiveness, hypoalgesia and high blood pressure in mice lacking the adenosine A2a receptor. Nature 388(6643): 674-678
20. Libert, F., Parmentier, M., Lefort, A., Dinsart, C., Van Sande, J., Maenhaut, C., Simons, M. J., Dumont, J. E., and Vassart, G. 1989. Selective amplification and cloning of four new members of theG protein-coupled receptor family. Science 244(4904): 569-572
21. Libert, F., Van Sande, J., Lefort, A., Czernilofsky, A., Dumont, J. E., Vassart, G., Ensinger, H. A., andMendla, K. D. 1992. Cloning and functional characterization of a human A1 adenosine receptor. Biochem.Biophys.Res.Commun. 187(2): 919-926
22. Linden, J. 2001. Molecular approach to adenosine receptors: receptor-mediated mechanisms of tissue protection. Annu Rev Pharmacol Toxicol 41: 775-787
23. Lohse, M. J., Lenschow, V., and Schwabe, U. 1984. Interaction of barbiturates with adenosine receptors in rat brain. Naunyn Schmiedebergs Arch.Pharmacol. 326(1): 69-74
24. Maemoto, T., Finlayson, K., Olverman, H. J., Akahane, A., Horton, R. W., and Butcher, S. P. 1997. Species differences in brain adenosine A1 receptor pharmacology revealed by use of xanthine and pyrazolopyridine based antagonists. Br J Pharmacol 122(6): 1202-1208
25. Mahan, L. C., McVittie, L. D., Smyk-Randall, E. M., Nakata, H., Monsma, F. J., Jr., Gerfen, C. R., andSibley, D. R. 1991. Cloning and expression of an A1 adenosine receptor from rat brain. Mol.Pharmacol. 40(1): 1-7
26. Marquardt, D. L., Walker, L. L., and Heinemann, S. 1994. Cloning of two adenosine receptor subtypes from mouse bone marrow-derived mast cells. J Immunol 152(9): 4508-4515
27. Meng, F., Xie, G. X., Chalmers, D., Morgan, C., Watson, S. J., Jr., and Akil, H. 1994. Cloning and characterization of a pharmacologically distinct A1 adenosine receptor from guinea pig brain.Brain Res.Mol.Brain Res. 26(1-2): 143-155
28. Mubagwa, K. and Flameng, W. 2001. Adenosine, adenosine receptors and myocardial protection: anupdated overview. Cardiovasc Res 52(1): 25-39
29. Murrison, E. M., Goodson, S. J., Edbrooke, M. R., and Harris, C. A. 1996. Cloning and characterisation of the human adenosine A3 receptor gene. FEBS Lett. 384(3): 243-246
30. Nyce, J. W. 1999. Insight into adenosine receptor function using antisense and gene-knockout approaches. Trends Pharmacol Sci 20(2): 79-83
31. Olah, M. E., Ren, H., Ostrowski, J., Jacobson, K. A., and Stiles, G. L. 1992. Cloning, expression, and characterization of the unique bovine A1 adenosine receptor. Studies on the ligand binding site by site-directed mutagenesis. J.Biol.Chem. 267(15): 10764-10770
32. Olah, M. E. and Stiles, G. L. 1995. Adenosine receptor subtypes: characterization and therapeutic regulation. Annu Rev Pharmacol Toxicol 35: 581-606
33. Pierce, K. D., Furlong, T. J., Selbie, L. A., and Shine, J. 1992. Molecular cloning and expression of anadenosine A2b receptor from human brain. Biochem.Biophys.Res.Commun. 187(1): 86-93
34. Pirovano, I. M., IJzerman, A. P., Van Galen, P. J., and Soudijn, W. 1989. Influence of the molecular structure of N6-(omega-aminoalkyl)adenosines on adenosine receptor affinity and intrinsic activity. Eur.J Pharmacol. 172(2): 185-193
35. Ribeiro, J. A., Sebastiao, A. M., and de Mendonca, A. 2002. Adenosine receptors in the nervous system: pathophysiological implications. Prog.Neurobiol. 68(6): 377-392
36. Rudolphi, K. A. and Schubert, P. 1996. Purinergic interventions in traumatic and ischemic injury. InNovel Thearapies for CNS Injuries, Peterson, P. L. and Willis, P. W. 327-342. Boca Raton, New York, London, Tokyo: CRC Press Inc.
37. Schulte, G. and Fredholm, B. B. 2003. Signalling from adenosine receptors to mitogen-activated protein kinases. Cell Signal. 15(9): 813-827
38. Stone, T. W. 2002. Purines and neuroprotection. Adv.Exp.Med.Biol. 513: 249-28039. Tabrizchi, R. and Bedi, S. 2001. Pharmacology of adenosine receptors in the vasculature. Pharmacol
Ther 91(2): 133-14740. Tatsis-Kotsidis, I. and Erlanger, B. F. 1999. A1 adenosine receptor of human and mouse adipose
tissues: cloning, expression, and characterization. Biochem Pharmacol 58(8): 1269-1277

The mouse brain adenosine A1 receptor
87
41. Van der Wenden, E. M., Hartog-Witte, H. R., Roelen, H. C., von Frijtag Drabbe Kunzel JK, Pirovano, I. M., Mathot, R. A., Danhof, M., Van Aerschot, A., Lidaks, M. J., IJzerman, A. P., and . 1995. 8-substituted adenosine and theophylline-7-riboside analogues as potential partial agonists for the adenosine A1 receptor. Eur J Pharmacol. 290(3): 189-199
42. Williams, M. and Jarvis, M. F. 2000. Purinergic and pyrimidinergic receptors as potential drug targets. Biochem Pharmacol 59(10): 1173-1185
43. Zhao, Z., Francis, C., and Ravid, K. 1999. Characterization of the mouse A3 adenosine receptor gene: exon/intron organization and promoter activity. Genomics 57(1): 152-155
44. Zhao, Z., Makaritsis, K., Francis, C. E., Gavras, H., and Ravid, K. 2000. A role for the A3 adenosine receptor in determining tissue levels of cAMP and blood pressure: studies in knock-out mice. Biochim Biophys Acta 1500(3): 280-290
45. Zhou, Q. Y., Li, C., Olah, M. E., Johnson, R. A., Stiles, G. L., and Civelli, O. 1992. Molecular cloningand characterization of an adenosine receptor: the A3 adenosine receptor. Proc.Natl.Acad.Sci.U.S.A 89(16): 7432-7436

88

89
To be submitted
Interleukin-6 is involved in upregulationof adenosine A1 receptor expression
after seizures
M.C. Wittendorp, J. Von Frijtag Drabbe Künzel, J.N. Keijser,M.A. Hensens, N. Brouwer, A.P. IJzerman,
H.W.G.M. Boddeke and K. Biber
Chapter 5

Summary
Introduction
Adenosine plays an important role in coupling energy supply to energy demandand exerts its effects via four subtypes of adenosine receptors named adenosineA1, A2A, A2B and A3 receptors. These receptors all belong to the family of 7 trans-membrane receptors (TM7) that predominantly signal through activation of G-proteins [17]. Although adenosine A1 and A3 receptors were originally reportedto couple to Gi/o protein and the adenosine A2 receptors to Gs protein, it is nowclear that this is not exclusively the case [17]. Adenosine receptors have beencloned from various species and their pharmacological profile is based on avariety of specific agonists and antagonists [17, 23].Under physiological conditions adenosine acts as an inhibitory neuromodulator inthe brain [10, 42]. Neuropathological conditions like stroke, ischemia andseizures lead to the release of large amounts of adenosine in the extracellularspace. Under these conditions adenosine acts as a neuroprotective agent and anendogenous anticonvulsant [13, 15]. Adenosine promotes neuronal survival andmoreover counteracts seizure activity by presynaptically inhibiting the release ofthe excitatory neurotransmitter glutamate [33, 34]. Postsynaptically, adenosinecauses hyperpolarization of the cell membrane, mainly by increasing K+
conductance [19, 40]. These actions of adenosine are mainly mediated byadenosine A1 receptors. Accordingly, the neuroprotective effect of adenosine willbe increased by upregulation of adenosine A1 receptors [34].
Chapter 5
90
The neuromodulator adenosine is released in the brain in large amounts during neuropathologicalevents, like stroke or seizures. Under these circumstances adenosine induces neuroprotectiveeffects by activating adenosine A1 receptors in neurons. This makes the adenosine system one ofthe most important neuroprotective systems in the brain. Significant upregulation of adenosine A1receptor expression has been shown in mice after seizures, thereby increasing the neuroprotectivefunction of adenosine. This upregulation is a rather unusual feature, since G-protein coupledreceptors are normally down regulated in the presence of large amounts of ligand. We proposehere a mechanism that induces upregulation of adenosine A1 receptor expression in the presenceof high concentrations of extracellular adenosine.In vitro experiments have shown that stimulation of glia cells with the pro-inflammatory cytokine,interleukin-6 (IL-6) leads to an increase of adenosine A1 receptor expression in cultured braincells. In order to investigate the role of IL-6 in adenosine A1 receptor upregulation after seizureswe have performed experiments in wild type and IL-6 knock out mice. We report that wild typemice show increased expression of adenosine A1 receptors in hippocampus and cortex afterpentylenetetrazole-induced seizures, measured by autoradiography. In contrast, no upregulationwas observed in IL-6 knock out mice. These results suggest that IL-6 is involved in theupregulation of adenosine A1 receptor expression after seizures. We conclude that IL-6 plays animportant role in the complex mechanism of adenosine-induced neuroprotection.

Upregulation of adenosine A1 receptors in the brain has been observed in mouseand rat after induction of seizures and in post mortem tissue of humans sufferingfrom epilepsy [5, 30, 41]. It has moreover been assumed that in mice thisupregulation of adenosine A1 receptors protects neurons against subsequentseizures [4]. However, the observed upregulation of adenosine A1 receptors inthe presence of high concentrations of extracellular adenosine is a rather unusualfeature, since TM7 receptors are normally downregulated (desensitized) in thepresence of large amounts of ligand [21, 36]. Indeed, desensitization of adenosineA1 receptors in response to prolonged agonist treatment in brain tissue has beenreported [1, 2, 36]. It may thus be assumed that the expression of adenosine A1
receptors during seizures is regulated such that the desensitizing properties ofhigh adenosine concentrations are counteracted. We have recently suggested amechanism that involves the cytokine IL-6, which could account for such anupregulation [8]. IL-6 is released in brain under pathological conditions likeischemia and seizures [26, 28, 31] where it is known to have significantneuroprotective effects [3, 18, 25, 39]. Previous work from our group has shownthat adenosine induces the release of IL-6 in astrocytes [16, 38] and that IL-6treatment increases adenosine A1 receptor expression in nervous tissue [8]. A direct relationship between IL-6 expression in brain and upregulation ofadenosine A1 receptors in vivo has yet not been demonstrated. In order to addressthis question we have now investigated the potential role of IL-6 in regulatingadenosine A1 receptor expression in vivo, using IL-6 deficient mice. We reporthere that the increased expression of adenosine A1 receptors, observed in thehippocampus and cortex of wild type mice in response to pentylenetetrazole-induced seizures is absent in IL-6 deficient mice. This finding corroborates theassumption that IL-6 increases A1 receptor expression in brain.
Materials and Methods
Animals and pentylenetetrazole treatment
Male wild type C57Bl6 mice (Harlan) and male IL-6 knock out mice (Jackson), 3months old, weighing 23-29 g were used. The animals were housed in groups offive per cage in a room with controlled light/dark (12h/12h) cycle andtemperature (21ºC). Food and water was supplied ad libitum. The mice receivedonce daily, for two consecutive days one intraperitoneal (i.p.) injection with 55mg/kg pentylenetetrazole (PTZ, Sigma) dissolved in 0.9% saline solution.Control animals received saline only. Latency of seizure onset was timed andanimals were subsequently observed for one hour. Animals were assigned aseizure score according to the following criteria adopted from Kondziella andcolleagues [24]. 0= normal behavior, 1= myoclonic jerks, 2= minimal seizures
IL-6-induced upregulation of adenosine A1 receptor expression
91

without Straub-tail, 3= minimal seizures with Straub-tail, 4= generalized tonic-clonic seizures, 5= like 4 with loss of consciousness and postictal phase, 6= like5 with rotation on their axis, 7= like 6 and death. All pentylenetetrazole (PTZ)treated animals showed at least score 2 (minimal seizures) within 5 minutes.Mice were killed by decapitation 26 hours after the second injection. Allexperiments were in accordance with the regulations of the Committee for Use ofExperimental Animals of the University of Groningen (DEC No. 2976).The mice which received PTZ injections did not all show tonic-clonic seizures.Therefore, the groups of mice (both wild type and IL-6 knock out) that receivedPTZ injections, were subdivided in two groups according to the severity of theseizures. The “PTZ-high” groups consisted of animals which showed tonic-clonicseizures (score 4) after at least one of the two PTZ injections. All other animals,which received PTZ injections, were assigned to the “PTZ-low” groups.
Tissue preparation
The brains were removed immediately after decapitation. The two hemisphereswere separated and subsequently frozen and stored at –80ºC until use. The righthemispheres were prepared for RNA analysis and radioligand binding studieswhile the left hemispheres were used for autoradiography. The right hemispheres of animals in the same experimental group were pooledbefore RNA extraction and membrane preparation. Wild type and IL-6 knock outcontrol groups contained 15 animals, while the four PTZ treated groups had 6-8animals per group. From the right hemispheres 25 coronal brain sections of 30 µmwere cut and lysed in guanidinium isothiocyanate containing mercaptoethanol.Lysed samples were stored at -20ºC until RNA analysis. The remaining part of theright hemispheres was collected in 0.32 M sucrose solution (4ºC) and immediate-ly processed further into brain membranes according to Lohse and colleagues[29]. In brief, brain tissue was homogenized using a Potter-homogenizer in 10volumes of 0.32 M sucrose. The homogenate was centrifuged at 1000 g for 15 minand the supernatants were centrifuged at 48000 g for 40 min. The pellets wereresuspended in 50 mM Tris-HCl buffer, pH 7.4, and subsequently centrifuged at48000 g for 15 min. After repeating the last centri-fugation step membranes wereresuspended in Tris-HCl buffer and incubated at 37ºC for 30 min with 2 IU/mladenosine deaminase. Brain membranes were frozen and stored at –80ºC untilthe binding assay was performed. Left hemispheres were cut into coronal brain sections of 20 µm using a Leicamicrotome (at -14ºC). The tissue sections were thaw-mounted on gelatine-coatedmicroscope slides and dried overnight sealed in plastic with silica gel. Microscopeslides were stored at –80ºC until autoradiography.
Chapter 5
92

RNA isolation and reverse transcription-polymerase chain reaction (RT-PCR)
Tissue was lysed in guanidinium isothiocyanate/mercaptoethanol buffer and totalRNA was extracted with one phenol/chloroform step according to [9].Subsequently, RNA was treated with RNAse free DNAse (Sigma-Aldrich,Bornhem, Belgium) for 30 min and precipitated. Reverse transcription (RT): 1 µgof total RNA was transcribed into cDNA in a total volume of 25 µl containing 0.5µl of M-MLV reverse transcriptase, 1 µl of RNase inhibitor, 1 µl of randomhexamers (0.2 mM), 5 µl of 5x buffer, 1.25 µl of deoxynucleosidetriphosphates(dNTPs) (10 mM) and H2O adjusted to 25 µl. After 60 min at 42°C, the reactionwas stopped by heating at 95°C for 5 min. The quality of the cDNA wascontrolled using glyceraldehyde-3-phosphate dehydrogenase (GAPDH) primers(see Table 5.1). Potential contamination by genomic DNA was checked for byrunning the reactions without reverse transcriptase and using GAPDH primers insubsequent polymerase chain reaction (PCR) amplifications. Conventional PCR:2 µl of the RT reaction was used in subsequent PCR amplification as describedpreviously [7]. In brief, the following reagents were added: 5 µl 10x PCR-buffer,2.5 µl MgCl2 (50 mM), 0.5 µl dNTPs (10 mM), 1 µl of a mix containing bothprimers, 39 µl H2O and 0.1 µl Taq-polymerase.
Real-time semi-quantitative polymerase chain reaction (Q-PCR)
Adenosine A1 receptor mRNA expression in the brains of wild type and IL-6knock out mice was analyzed by real-time PCR using the iCycler (Bio-rad,Veenendaal, The Netherlands) and the iQ SYBR Green supermix (Bio-rad).Mouse ribosomal protein L32-3A (rpL32A) primers and hypoxanthine guaninephosphoribosyl transferase (HPRT1) primers were used for normalization tohousekeeping genes. These genes did not show variations in response to theexperimental treatment. Primers used for adenosine A1 receptor analysis and thehousekeeping gene primers, listed in table 5.1, have been designed by the use ofthe Primer Designer program (Scientific and Educational Software, Version 3.0).The comparative Ct method (amount of target amplicon X in sample S,
IL-6-induced upregulation of adenosine A1 receptor expression
93
Table 5.1. Primers used for conventional reverse transcriptase and real-time (*) PCR
Gene accession Forward primer (5’-3’) Backward primer (5’-3’) Productnumber size(bp)
GAPDH AF106860 CATCCTGCACCACCAACTGCTTAG GCCTGCTTCACCACCTTCTTGATG 346rpL32A* X06483 GCTGGAGGTGCTGCTGATGT ACTCTGATGGCCAGCTGTGC 114HPRT1* X62085 GACTTGCTCGAGATGTCA TGTAATCCAGCAGGTCAG 102Ado A1R* AJ555877 CCTCTCCGGTACAAGACAGT GGTGTCAGGCCTACCACAAG 92

normalized to a reference R and related to a control sample C, calculated by2-{(CtX,S-CtR,S)- (CtX,C-CtR,C)} was used to determine the relative expression levelsbetween animal groups treated with saline and PTZ [27]. Results are given asmean ± S.E.M. (n=4). Relative expression values of adenosine A1 receptormRNA in animal groups treated with PTZ were compared to control levels (salinetreatment) by the one-sample t test, using 1 as the test value. p < 0.05 wasconsidered significant.
Radioligand binding assay
For the determination of the maximum bound radioligand, membranes wereincubated for 1 hr at 25°C in 50 mM Tris/HCl (pH 7.4 at 25°C) in the presence of4 nM [3H]DPCPX (Ki = 1.4 nM). Experiments were performed three times intriplo. Aspecific binding was determined in the presence of 100 µM CPA. Totalvolume during incubation was 200 µl. The protein concentration was measured inthe suspension used in the different experiments with the bicinchonic acidmethod. Incubations were stopped by rapid dilution with 1 ml ice-cold buffer andbound radioligand was subsequently recovered by filtration through WhatmanGF/B filters using either Millipore system or Brandel Harvester under reducedpressure. Filters were then washed three times with 2 ml buffer. The retainedradioactivity was measured by liquid scintillation counting (LKB Wallac, 1219Rackbeta).
Autoradiography
Sections were preincubated in Tris-HCl buffer, 170 mM, pH 7.4 containing 0.5IU/ml adenosine deaminase (Sigma Co.) for 3 x 10 min at room temperature,washed out and dried in a stream of cold air. For the labeling of adenosine A1
receptors, sections were incubated with 3.0 nM [3H]- N6-cyclohexyladenosine([3H]-CHA, Moravek Biochemicals, Brea, CA, USA, spec. activity 20 Ci/mmol) in170 mM Tris-HCl buffer including 0.5 IU/ml adenosine deaminase at roomtemperature for 90 min. To determine non-specific binding sections wereincubated in the presence of 3 µM N6-cyclopentyladenosine (CPA, Sigma-RBI,Saint Louis, MA, USA). Following incubation, unbound radioactivity wasremoved by rinsing the sections three times for 2 min in ice-cold Tris-HCl buffer,50 mM, pH 7.4, and rinsing once for 5 s in ice-cold distilled water to removebuffer salts. The sections were then dried in a stream of cold air. Dried sections were exposed to a tritium-sensitive phosphor screen (Packard)along with tritium standards ([3H]-microscales, Amersham) and stored in an X-ray film cassette for 96 hours at room temperature. Screens were read using theCyclone Storage Phosphor system (Packard). Quantification analysis of theresulting autoradiographic images was performed by using an automatic imageanalysis system (Quantimet 500, Leica, Cambridge). Optical density was measured
Chapter 5
94

in hippocampus and cortex in six sections per mouse (see Figure 5.1 for brainareas measured). The values of the sections were averaged for each mouse.Optical density measurements from each brain area were converted into fmolligand bound per mg tissue equivalent, according to the calibration curveobtained from the tritium standards. Statistical comparisons of the four animalgroups were performed using one-way ANOVA followed by Tukey posthoc test.
Results
Seizure scores
Seizure scores consist of ordinal data, therefore an average seizure score pergroup is not informative on the actual seizure scores. We therefore compared theseizure severity after the first and second PTZ-injection per animal. Figure 5.2shows the percentage of animals with similar seizure scores after both PTZ-injections and animals that showed higher scores after the first injection or afterthe second injection. In wild type mice one animal showed lower seizure scoresafter the second injection. The majority of animals (7 out of 10 animals) showedequal seizure scores on both days and in two animals higher seizure scores afterthe second injection were found. This pattern was different in IL-6 deficient mice,where the majority of animals (7 out of 13 animals) showed higher scores after
IL-6-induced upregulation of adenosine A1 receptor expression
95
12
3
Figure 5.1. Overview of the brain areas measured with autoradiography. Autoradiography wasperformed as described in the methods part. Adenosine A1 receptor expression was measured inone area of the hippocampus, which showed high expression of adenosine A1 receptor (area 1) andin two areas of the cortex. Area 2 included the parietal/somatosensory cortex while area 3 includedthe piriform cortex. All sections in which adenosine A1 receptor expression was measured weretaken at approximately Bregma –1.75.

the second injection and the minority showed equal (5 out of 13) or lower seizurescores (1 out of 13) after the second PTZ-injection. In order to compare thescores after first and second injection statistically, the Wilcoxon signed ranks testwas used. IL-6 knock out mice showed significantly (p < 0.05) higher seizurescores after the second injection compared to the first, whereas no significantdifference in seizure scores between the two injection rounds was observed inwild type animals.
Adenosine A1 receptor mRNA expression
In order to investigate possible effects of seizure activity in IL-6 deficient mice andwild type animals on A1 receptor mRNA expression, Q-PCR analysis wasperformed. No differences were observed in mice with lower seizure scorescompared to control mice (data not shown). Similarly no seizure effect on A1
receptor expression was measured in wild type with high seizures scores comparedto untreated mice (Figure 5.3). In contrast a significant (p < 0.05) downregu-lation of adenosine A1 receptor mRNA expression was detected in IL-6 deficientmice with high seizure scores compared to untreated controls (Figure 5.3).
Adenosine A1 receptor expression in brain membranes (radioligand binding)
Radioligand binding experiments using [3H]DPCPX have been performed inorder to investigate the expression of adenosine A1 receptors in the brain. Theuse of [3H]DPCPX as a specific mouse adenosine A1 receptor binding molecule
Chapter 5
96
0
40
60
80
20
% o
f ani
mal
s
day 1>2
wild typeIL-6 KO
seizure scoresday 1=2 day 1<2
Figure 5.2. Comparison of seizure scores of wild type and IL-6 knock out after first and secondPTZ-injection. Day 1>2 indicates higher seizure scores on day 1; 1=2 indicates no differences inseizure scores between the two injections; 1<2 indicates that the observed seizure score on day 2was higher than on day 1. Statistical analysis using the wilcoxan signed ranks test showed that IL-6knock out mice showed significantly higher scores on day 2 compared to day 1 while wild typeanimals showed similar scores on both days.

has been evaluated in an earlier study [43]. Since a high concentration (3 times ofthe Ki) of the radioligand was used, one can assume that all adenosine A1
receptors present in the preparation will be occupied by the ligand. Therefore theamount of [3H]DPCPX binding is informative on the number of adenosine A1
receptors. The graph in figure 5.4 shows [3H]DPCPX binding in fmol per mgtissue in wild type animals and IL-6 deficient mice. In IL-6 knock out mice thereare no differences between control and PTZ-injected animals (both PTZ low scoreand PTZ high score). Similarly we did not observe a difference in [3H]DPCPXbinding between the PTZ low score group and control wild type animals. In wildtype mice that showed high seizure scores after PTZ injection more [3H]DPCPXbinding was observed. This difference, however, just failed to reach significance,which is most likely due to the fact that seizures do not upregulate adenosine A1
receptor expression in all brain regions [4, 30]. It was therefore further decided toinvestigate the seizure effects in autoradiographic experiments as described [4].Since animals which showed low seizure scores, did not show any sign ofincreased [3H]DPCPX binding only animals with high seizure scores were usedfor autoradiography experiments.
IL-6-induced upregulation of adenosine A1 receptor expression
97
0.9
0.7
1.0
1.1
0.8re
lativ
e ad
enos
ine
A1
mRN
Aex
pres
sion
leve
l (ra
tio P
TZ/c
ontr
ol)
wild type
*
IL-6 KO
Figure 5.3. Real-time PCR analysis of adenosine A1 receptor mRNA expression in brains of wildtype and IL-6 knock out mice (normalized to rpl32A and HPRT1). Graph shows the relativeexpression level of mRNA in PTZ-injected animals (“PTZ high score”) compared to controls. Theexpression level of controls is set to 1. Data represent mean ± S.E.M of 4 experiments. * =significant reduction of mRNA expression compared to controls (p < 0.05), measured with theone-sample T-test.

Adenosine A1 receptor expression in brain sections (autoradiographicanalysis)
Figure 5.5 shows the results of the autoradiography experiments. We investigatedadenosine A1 receptor expression in the hippocampus and cortex, brain regionsthat showed an upregulation of adenosine A1 receptors in response to PTZ-induced seizures [4, 30]. In hippocampus (Figure 5.5A) and cortex (parietal/somatosensory cortex) (Figure 5.5B) of wild type mice a significant upregulation(p < 0.05) of [3H]CHA binding was observed in response to seizures. Thesefindings indicate an upregulation of adenosine A1 receptor expression ofapproximately 20% resembling values found in an earlier study [30]. In strikingcontrast no upregulation of adenosine A1 receptor expression was observed in IL-6 knock out mice, thereby corroborating the results obtained in membranepreparations. In another area of the cortex (including the piriform cortex) nodifferences in [3H]CHA binding were observed in both animal groups, wild typemice and IL-6 knock out animals (Figure 5.5C).
Correlation between seizure scores and adenosine A1 receptor expression
By plotting the seizure scores against the [3H]CHA binding results of theautoradiography experiments of individual animals, a possible correlationbetween seizure score (after the first PTZ-injection) and adenosine A1 receptorexpression was investigated (Figure 5.6). Data of both control (which all hadseizure score 0) and PTZ treated animals (score 2-4) were plotted. Statisticalanalysis revealed that the Pearson’s correlation coefficient was significant; p =0.003 (hippocampus) and p = 0.0019 (cortex1), indicating a positive correlation
Chapter 5
98
0
200
400
800
1000
[3 H] D
PCPX
bin
ding
in fm
ol/m
g tis
sue
wild type
600
IL-6 KO
controlPTZ low scorePTZ high score
Figure 5.4. Specific radioligand [3H]DPCPX binding to adenosine A1 receptors in brainmembranes of wild type and IL-6 knock out mice. Data are represented as mean ± S.E.M. of 3experiments. No significant differences could be found using one-way ANOVA.

between seizure scores observed on day 1 and [3H]CHA binding in wild typeanimals (Figure 5.6A and C respectively). No significant correlations were foundin IL-6 knock out animals (Figure 5.6B and D). It was also clear that the two wildtype animals that showed lower seizure scores (2 and 3) after PTZ injection alsohad lower adenosine A1 receptor expression levels.
IL-6-induced upregulation of adenosine A1 receptor expression
99
B
A
65
70
80
[3 H] C
HA
bin
ding
in fm
ol/m
g tis
sue
wtcontrol
60
cortex 2
wtPTZ
Il-6 KOcontrol
Il-6 KOPTZ
C
75
90
110
80
cortex 1*
100
hippocampus*
100
110
120
130
150
140
Figure 5.5. Autoradiographic analysis of adenosine A1 receptor expression in brain areas of wildtype and IL-6 knock out mice. Graphs show [3H]CHA binding in hippocampus (A), cortex 1 (B)and cortex 2 (C). Data are represented as mean ± S.E.M. per group (6-8 animals per group). * =significant increase in adenosine A1 receptor expression in hippocampus (A) and cortex 1 (B);p<0.05 using one-way ANOVA followed by Tukey post-hoc test.

Discussion
The current experiments were aimed to investigate the potential regulatory effectof IL-6 on the expression of adenosine A1 receptors in brain tissue. Bothadenosine and IL-6 are released under neuropathological conditions like seizures[26, 31, 32, 34, 37] and both factors induce pronounced neuroprotection [14, 18,25, 28, 35]. However, in contrast to the well-understood neuroprotective effectsof adenosine the mechanism of IL-6-induced neuroprotection remains to beelucidated [18, 20].The main receptor involved in the neuroprotective effect of adenosine is theadenosine A1 receptor [14, 34] and it has long been known that an upregulationof its expression increases its neuroprotective capacities [34]. Accordingly, a
Chapter 5
100
BA
120
100
[3 H] C
HA
bin
ding
in c
orte
x 1
(in fm
ol/m
g tis
sue)
60
wild type
C
80
100
120
0seizure score on day 1
1 2 3 4
D
0seizure score on day 1
1 2 3 4
[3 H] C
HA
bin
ding
in h
ippo
cam
pus
(in fm
ol/m
g tis
sue)
140
160
IL-6 KO
Figure 5.6. Adenosine A1 receptor expression in wild type mice is correlated to seizure scoresafter the first PTZ-injection. Correlation analysis of adenosine A1 receptor expression and seizurescores were performed by plotting the expression data against the seizure scores. Each data pointin the graphs represents an individual animal. Regression lines were calculated using Sigma Plot8.0. Positive correlations were found between adenosine A1 receptor expression in hippocampusand cortex area 1 and seizure scores induced by the first PTZ-injection in wild type animals (A, C)but not in IL-6 knock out animals (B, D).

marked adenosine A1 receptor upregulation has been observed in mice and ratsby chemically induced seizures and in post mortem tissue of humans sufferingfrom epilepsy. In all cases this upregulation is assumed to protect neurons againstsubsequent seizures [4, 5, 30, 41]. The upregulation of adenosine A1 receptorsduring seizures presents a paradox, since adenosine A1 receptors have been foundto be downregulated by prolonged ligand treatment [1, 2, 21, 36]. We haverecently described an upregulating effect of IL-6 treatment on adenosine A1
receptor expression in vitro and proposed that the cytokine could explain theincreased expression of A1 receptors after seizures [8]. In order to evaluate thishypothesis we have induced seizures in IL-6 deficient mice and wild type miceand compared the expression of adenosine A1 receptors in brain.First, we found that IL-6 deficient mice showed higher seizures scores after thesecond injection compared to the first injection, whereas this was not the case inwild type animals. Moreover, two animals in the IL-6 deficient group died as aresult of PTZ induced seizures, whereas all wild type animals survived theprocedure. These findings suggest that IL-6 knock out mice are more susceptibleto PTZ induced seizures, which was also concluded in two very recent studies[11, 12]. Second, no seizure effect on A1 receptor mRNA expression was found in wild typemice, whereas PTZ treatment caused a significant down regulation of adenosineA1 receptor mRNA expression in brain of IL-6 deficient mice. These findings thusindicate a stimulatory effect of IL-6 on the adenosine A1 receptor mRNAexpression in brain, verifying data that were obtained in vitro previously [8].Third, using autoradiography we observed a significant increase in adenosine A1
receptor expression in the hippocampus and part of the cortex in wild type miceafter PTZ-induced seizures. In contrast to wild type mice we did not find anyeffect of PTZ treatment on the expression of adenosine A1 receptors in the brainof IL-6 deficient mice. These autoradiographic findings were supported byradioligand binding experiments in membrane preparations of the brain. Takentogether these findings strongly support our previous assumption that IL-6increases the expression of adenosine A1 receptors in nervous tissue [8]. In a recent paper it was shown that seizures could be induced in IL-6 knock outmice at much lower dosages of several convulsant substances when compared towild type littermates [12]. By analyzing the effect of these different convulsantsthe authors conclude that IL-6 knock out mice might have impaired GABAinhibitory inputs and enhanced neurotransmission through glutamate receptors.Since adenosine A1 receptors are regulating both transmitter systems [14], wesuggest that IL-6 knock out mice are more susceptible to seizures due to a lack ofadenosine A1 receptor upregulation. Stimulation of adenosine A1 receptor expression might not be the onlyconnection between IL-6 and the adenosinergic system. We and others have
IL-6-induced upregulation of adenosine A1 receptor expression
101

shown that stimulation of adenosine A2B receptors in astrocytes leads to thesynthesis and release of IL-6 [16, 38]. Since adenosine A2B receptors have a lowaffinity for adenosine, high concentrations of the ligand (which are found duringseizures) are required to activate this receptor. We therefore propose thefollowing sequence of events: high concentrations of adenosine (like in seizures)activate adenosine A2B receptors in astrocytes and lead to the release of IL-6,which in turn increases the expression of adenosine A1 receptors augmenting theneuroprotective properties of this receptor [8]. Interestingly, a significant correlation was found in this study between the seizurescores and the induction of adenosine A1 receptor expression in the hippocampusand cortex of wild type mice. Thus, increased expression of adenosine A1
receptors was only found in animals with high seizure scores. Since according tothe above mentioned assumptions high levels of extracellular adenosine arerequired to initiate the circuitry, it is tempting to speculate that only in animalswith high seizure scores a sufficient amount of adenosine was released tostimulate the synthesis of IL-6. However, in order to confirm this hypothesis,further experiments are needed that would investigate the correlation of seizureseverity and IL-6 release in the brain.The data presented here and the results of others strongly suggest a beneficialrole of the pro-inflammatory cytokine IL-6 in brain during seizures. Findingspointing in the same direction have been published earlier by Asanuma andcolleagues [6]. This group found that an inhibition of brain inflammation bycyclosporine A treatment, which blocks the expression of several inflammatorygenes [22], significantly aggravated the severity of PTZ-induced seizures.Interestingly, it was observed that cyclosporine A treatment only affected seizureactivity after the second PTZ injection very similar to the effect we observed in IL-6 deficient mice [6]. These authors concluded that the brain inflammationoccurring in response to the first seizures would initiate processes that protectthe brain from upcoming seizure events. We have now added data in support ofthis assumption, showing that inflammatory events (release of IL-6) upregulatethe expression of the adenosine A1 receptor. We and others found that seizures did not induce an increase in adenosine A1
receptor expression in all brain regions [30]. Due to our data it would thereforebe of particular interest to investigate the question whether brain regions withoutan effect on A1 receptor expression might have different IL-6 release capacities orwhether the expression of A1 receptors is differentially regulated. In conclusion, this study demonstrates a clear relationship between the expressionof IL-6 and the upregulation of adenosine A1 receptors in response to seizures.The current data not only verify findings that have been obtained in vitro, theymoreover provide an explanation for the well-known, but not well-understoodneuroprotective properties of IL-6.
Chapter 5
102

References
IL-6-induced upregulation of adenosine A1 receptor expression
103
1. Abbracchio, M. P., Cattabeni, F., Fredholm, B. B., and Williams, M. 1993. Purinoceptor Nomenclature: A status report. Drug development research 28: 207-213
2. Abbracchio, M. P., Fogliatto, G., Paoletti, A. M., Rovati, G. E., and Cattabeni, F. 1992. Prolonged in vitro exposure of rat brain slices to adenosine analogues: selective desensitization of adenosine A1
but not A2 receptors. Eur J Pharmacol. 227(3): 317-3243. Ali, C., Nicole, O., Docagne, F., Lesne, S., MacKenzie, E. T., Nouvelot, A., Buisson, A., and Vivien,
D. 2000. Ischemia-induced interleukin-6 as a potential endogenous neuroprotective cytokine againstNMDA receptor-mediated excitotoxicity in the brain. J Cereb.Blood Flow Metab 20(6): 956-966
4. Angelatou, F., Pagonopoulou, O., and Kostopoulos, G. 1991. Changes in seizure latency correlate with alterations in A1 adenosine receptor binding during daily repeated pentylentetrazol-induced convulsions in different mouse brain areas. Neurosci Lett 132(2): 203-206
5. Angelatou, F., Pagonopoulou, O., Maraziotis, T., Olivier, A., Villemeure, J. G., Avoli, M., and Kostopoulos, G. 1993. Upregulation of A1 adenosine receptors in human temporal lobe epilepsy: a quantitative autoradiographic study. Neurosci Lett 163(1): 11-14
6. Asanuma, M., Ogawa, N., Nishibayashi, S., Kondo, Y., and Mori, A. 1995. Effects of repeated injection of cyclosporin A on pentylenetetrazol-induced convulsion and cyclophilin mRNA levels in rat brain. Neurochem.Res. 20(1): 101-105
7. Biber, K., Klotz, K. N., Berger, M., Gebicke-Harter, P. J., and van Calker, D. 1997. Adenosine A1
receptor-mediated activation of phospholipase C in cultured astrocytes depends on the level of receptor expression. J Neurosci 17(13): 4956-4964
8. Biber, K., Lubrich, B., Fiebich, B. L., Boddeke, H. W., and van Calker, D. 2001. Interleukin-6 enhances expression of adenosine A(1) receptor mRNA and signaling in cultured rat cortical astrocytes and brain slices. Neuropsychopharmacology 24(1): 86-96
9. Chomczynski, P. and Sacchi, N. 1987. Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal.Biochem. 162(1): 156-159
10. Cunha, R. A. 2001. Adenosine as a neuromodulator and as a homeostatic regulator in the nervous system: different roles, different sources and different receptors. Neurochem Int 38(2): 107-125
11. De Luca, G., Di Giorgio, R. M., Macaione, S., Calpona, P. R., Costantino, S., Di Paola, E. D., De Sarro, A., Ciliberto, G., and De Sarro, G. 2004. Susceptibility to audiogenic seizure and neurotransmitter amino acid levels in different brain areas of IL-6-deficient mice. Pharmacol.Biochem.Behav. 78(1): 75-81
12. De Sarro, G., Russo, E., Ferreri, G., Giuseppe, B., Flocco, M. A., Di Paola, E. D., and De Sarro, A. 2004. Seizure susceptibility to various convulsant stimuli of knockout interleukin-6 mice. Pharmacol.Biochem.Behav. 77(4): 761-766
13. Dragunow, M. 1988. Purinergic mechanisms in epilepsy. Prog.Neurobiol. 31(2): 85-10814. Dunwiddie, T. V. and Masino, S. A. 2001. The role and regulation of adenosine in the central
nervous system. Annu Rev Neurosci 24: 31-5515. Dunwiddie, T. V. and Worth, T. 1982. Sedative and anticonvulsant effects of adenosine analogs in
mouse and rat. J.Pharmacol.Exp.Ther. 220(1): 70-7616. Fiebich, B. L., Biber, K., Gyufko, K., Berger, M., Bauer, J., and van Calker, D. 1996. Adenosine A2b
receptors mediate an increase in interleukin (IL)-6 mRNA and IL-6 protein synthesis in human astroglioma cells. J Neurochem 66(4): 1426-1431
17. Fredholm, B. B., IJzerman, A. P., Jacobson, K. A., Klotz, K. N., and Linden, J. 2001. International Union of Pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol Rev 53(4): 527-552
18. Gadient, R. A. and Otten, U. H. 1997. Interleukin-6 (IL-6)--a molecule with both beneficial and destructive potentials. Prog Neurobiol 52(5): 379-390
19. Gerber, U. and Gahwiler, B. H. 1994. GABAB and adenosine receptors mediate enhancement of the K+ current, IAHP, by reducing adenylyl cyclase activity in rat CA3 hippocampal neurons. J Neurophysiol 72(5): 2360-2367

Chapter 5
104
20. Gruol, D. L. and Nelson, T. E. 1997. Physiological and pathological roles of interleukin-6 in the central nervous system. Mol Neurobiol 15(3): 307-339
21. Hettinger, B. D., Leid, M., and Murray, T. F. 1998. Cyclopentyladenosine-induced homologous down-regulation of A1 adenosine receptors (A1AR) in intact neurons is accompanied by receptor sequestration but not a reduction in A1AR mRNA expression or G protein alpha-subunit content.J Neurochem. 71(1): 221-230
22. Ho, S., Clipstone, N., Timmermann, L., Northrop, J., Graef, I., Fiorentino, D., Nourse, J., and Crabtree, G. R. 1996. The mechanism of action of cyclosporin A and FK506. Clin.Immunol.Immunopathol. 80(3 Pt 2): S40-S45
23. Klotz, K. N., Hessling, J., Hegler, J., Owman, C., Kull, B., Fredholm, B. B., and Lohse, M. J. 1998. Comparative pharmacology of human adenosine receptor subtypes - characterization of stably transfected receptors in CHO cells. Naunyn Schmiedebergs Arch Pharmacol 357(1): 1-9
24. Kondziella, D., Bidar, A., Urfjell, B., Sletvold, O., and Sonnewald, U. 2002. The pentylenetetrazole-kindling model of epilepsy in SAMP8 mice: behavior and metabolism. Neurochem.Int. 40(5): 413-418
25. Kunioku, H., Inoue, K., and Tomida, M. 2001. Interleukin-6 protects rat PC12 cells from serum deprivation or chemotherapeutic agents through the phosphatidylinositol 3-kinase and STAT3 pathways. Neurosci.Lett. 309(1): 13-16
26. Lehtimaki, K. A., Peltola, J., Koskikallio, E., Keranen, T., and Honkaniemi, J. 2003. Expression of cytokines and cytokine receptors in the rat brain after kainic acid-induced seizures.Brain Res.Mol.Brain Res. 110(2): 253-260
27. Livak, K. J. and Schmittgen, T. D. 2001. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 25(4): 402-408
28. Loddick, S. A., Turnbull, A. V., and Rothwell, N. J. 1998. Cerebral interleukin-6 is neuroprotective during permanent focal cerebral ischemia in the rat. J Cereb Blood Flow Metab 18(2): 176-179
29. Lohse, M. J., Lenschow, V., and Schwabe, U. 1984. Interaction of barbiturates with adenosine receptors in rat brain. Naunyn Schmiedebergs Arch.Pharmacol. 326(1): 69-74
30. Pagonopoulou, O., Angelatou, F., and Kostopoulos, G. 1993. Effect of pentylentetrazol-induced seizures on A1 adenosine receptor regional density in the mouse brain: a quantitative autoradiographic study. Neuroscience 56(3): 711-6
31. Peltola, J., Hurme, M., Miettinen, A., and Keranen, T. 1998. Elevated levels of interleukin-6 may occur in cerebrospinal fluid from patients with recent epileptic seizures. Epilepsy Res. 31(2): 129-133
32. Peltola, J., Laaksonen, J., Haapala, A. M., Hurme, M., Rainesalo, S., and Keranen, T. 2002. Indicatorsof inflammation after recent tonic-clonic epileptic seizures correlate with plasma interleukin-6 levels. Seizure. 11(1): 44-46
33. Ribeiro, J. A. 1995. Purinergic inhibition of neurotransmitter release in the central nervous system. Pharmacol Toxicol 77(5): 299-305
34. Rudolphi, K. A. and Schubert, P. 1996. Purinergic interventions in traumatic and ischemic injury. InNovel Thearapies for CNS Injuries, Peterson, P. L. and Willis, P. W. 327-342. Boca Raton, New York, London, Tokyo: CRC Press Inc.
35. Rudolphi, K. A., Schubert, P., Parkinson, F. E., and Fredholm, B. B. 1992. Neuroprotective role of adenosine in cerebral ischaemia. Trends Pharmacol Sci 13(12): 439-445
36. Ruiz, A., Sanz, J. M., Gonzalez-Calero, G., Fernandez, M., Andres, A., Cubero, A., and Ros, M. 1996.Desensitization and internalization of adenosine A1 receptors in rat brain by in vivo treatment with R-PIA: involvement of coated vesicles. Biochim.Biophys.Acta 1310(1): 168-174
37. Samland, H., Huitron-Resendiz, S., Masliah, E., Criado, J., Henriksen, S. J., and Campbell, I. L. 2003. Profound increase in sensitivity to glutamatergic- but not cholinergic agonist-induced seizuresin transgenic mice with astrocyte production of IL-6. J Neurosci.Res. 73(2): 176-187
38. Schwaninger, M., Neher, M., Viegas, E., Schneider, A., and Spranger, M. 1997. Stimulation of interleukin-6 secretion and gene transcription in primary astrocytes by adenosine. J Neurochem69(3): 1145-1150
39. Swartz, K. R., Liu, F., Sewell, D., Schochet, T., Campbell, I., Sandor, M., and Fabry, Z. 2001. Interleukin-6 promotes post-traumatic healing in the central nervous system. Brain Res 896(1-2): 86-95

IL-6-induced upregulation of adenosine A1 receptor expression
105
40. Trussell, L. O. and Jackson, M. B. 1985. Adenosine-activated potassium conductance in cultured striatal neurons. Proc Natl Acad Sci U S A 82(14): 4857-4861
41. Vanore, G., Giraldez, L., Rodriguez de Lores, Arnaiz G., and Girardi, E. 2001. Seizure activity produces differential changes in adenosine A1 receptors within rat hippocampus. Neurochem.Res. 26(3): 225-230
42. Williams, M. 1989. Adenosine: the prototypic neuromodulator. Neurochemistry international 14(3): 249-264
43. Wittendorp, M. C., Von Frijtag Drabbe Kunzel J., IJzerman, A. P., Boddeke, H. W. G. M., and Biber, K. 2004. The mouse brain adenosine A(1) receptor: functional expression and pharmacology.Eur J Pharmacol. 487(1-3): 73-79

106

107
Summary and conclusions
Chapter 6

The adenosinergic system
The energy-rich molecule ATP is used by all living cells as a “currency”, providingenergy for most cellular activities. In most organisms the daily turnover of ATPthus equals their bodyweight [12]. Adenosine, a metabolite of ATP, plays animportant role as a regulator of metabolic processes, but probably duringevolution it has gained an additional role as a signaling molecule. In chapter 1general features and functions of the adenosinergic system have been discussed indetail. In the subsequent chapters of the thesis we have focussed on theneuroprotective activity of adenosine receptors.
Summary/scope of the thesis
During neuropathological conditions large amounts of adenosine are released inthe brain. Whereas increased levels of adenosine mediate neuroprotection, in thelong term down regulation of adenosine receptors will occur. We hypothesizedthat in order to maintain the neuroprotective effects of adenosine there must besome factor involved to uphold or even increase adenosine receptor expression.Previous research in vitro has indicated that IL-6 might be involved in such amechanism [3, 7]. In figure 1.6 we have postulated our hypothesis on the key ofevents occurring during pathological conditions in the brain.
“ IL-6 is involved in upregulation of adenosine A1 receptors in neuropathological conditionsin vivo”
In order to prove this hypothesis a number of projects has been performed, whichare described in chapters 3, 4 and 5.
The mouse adenosine A1 receptor resembles the receptor in other species
Whereas the adenosine A1 receptor of many animal species has been cloned andcharacterized, hardly any literature data on the mouse A1 receptor were availableconcerning mouse adenosine A1 receptor pharmacology. Since we intended to usea mouse knock out model as approach to prove our hypothesis, we had tocharacterize the binding properties of ligands towards the mouse adenosine A1
receptor. We therefore cloned the adenosine A1 receptor from mouse astrocytesand expressed it in CHO cells. In chapter 4 the cloning, functional expressionand pharmacology of the mouse brain adenosine A1 receptor has been presented.The results show that the protein sequence of the adenosine A1 receptorresembles the rat and human sequences. Moreover, we found that commonlyused ligands bind the mouse receptor with similar affinities as the rat and human
Chapter 6
108

receptor. Therefore, we can conclude that in our experiments we can use thesame ligands, which have been used successfully in rats. Furthermore, thepharmacological characterization of the adenosine A1 receptor will be of help forthe study of other adenosine-mediated physiological processes in mouse models.
Working out the hypothesis
Possible regulatory effects of IL-6Currently, no effective IL-6 receptor antagonists are available. Therefore, in orderto study the possible regulatory effect of IL-6 on adenosine A1 receptor express-ion in vivo we have decided to use a IL-6 knock out mouse model. The con-stitutive IL-6 knock out model has the disadvantage that mice without the genecould have adapted and modified their physiology to cope with the loss of thegene. We have taken this risk because in general knock out mouse models haveproved to be useful tools to study the role of one specific factor. No overt changesin brain morphology and also on clear physiological phenotype was observed inIL-6 knock out mice. Furthermore, as expected, the lack of IL-6, clearly preventedupregulation of the A1 adenosine receptors in the seizure model.
Main findings (chapter 5)● IL-6 knock out mice were more susceptible to PTZ-induced seizures. Thisfinding implicates that the increased seizure susceptibility in IL-6 knock out miceis possibly due to a lower adenosine A1 receptor expression.● A subsequent autoradiography study indeed showed a lack of adenosine A1
receptor upregulation in IL-6 mice in the seizure model. Further analysis of theresults showed a clear correlation between the severity of the seizures and therate of adenosine A1 receptor expression in wild type animals. Wild type animalswith higher adenosine A1 receptor expression levels had generally a lower seizuresusceptibility, while in IL-6 knock out mice this correlation was absent.● Radioligand binding results in total mouse brain homogenates failed to show adifference in A1 expression levels, both under control conditions and after PTZ-induced seizures. The most likely explanation for the different findings inradioligand binding and autoradiography is the fact that the A1 adenosinereceptor is upregulated in distinct brain areas. The detection of this upregulationis lost in whole brain.
From the seizure experiments and the subsequent autoradiography we concludethat IL-6 is involved in upregulation of adenosine A1 receptor expression afterseizures, thereby enhancing the neuroprotective effects of adenosine. In the current literature contradicting effects of IL-6 in the brain have beenreported. Thus both detrimental and neuroprotective effects of IL-6 in the brain
Summary and conclusions
109

have been described. These different effects are probably due the differences oflevels of IL-6 involved and differences in brain areas studied. Our findings suggesta pronounced neuroprotective role of IL-6 in the brain. Moreover, our results alsoprovide the mechanism involved in IL-6 induced neuroprotection.
Future implicationsWe have observed adenosine A1 receptor upregulation 24-48 hours after PTZ-induced seizures in wild type mice but not in IL-6 knock out mice, implicating thatIL-6 induced upregulation of A1 receptors takes place within this time. It is how-ever, not yet clear what triggers the release of IL-6 from astrocytes in vivo: previousexperiments in vitro suggest involvement of adenosine A2B receptors [7, 14]. Furthermore, questions remain on the timing of the sequence of eventsconcerning the production of IL-6 and its’ release from astrocytes. In order tostudy this in detail, IL-6 release in the brain should be measured and adenosineA1 receptor mRNA and protein samples should be taken at various time pointsafter seizures. Since this would require a large number of test animals it would bedesirable to investigate this issue in a brain slice culture model using RNAsilencing techniques to knock out the IL-6 gene. Due to its’ neuroprotective effects, several reports have suggested that IL-6 couldhave some therapeutic potential in neurodegenerative diseases [9, 11, 13]. Due tothe widespread actions of IL-6 leading to peripheral inflammatory responses, IL-6based drugs can not be administered systemically. Therefore it is of utmostimportance that IL-6 is delivered exactly at the site where its actions are needed.Possible techniques which have been proposed are mini-osmotic pumps, gene-therapy or carrier molecules [9]. However, it seems rather unlikely that IL-6based drugs will be used in clinical setting. IL-6 itself might not be a good candidate as a target to increase the neuro-protective effects of adenosine. But there might be other factors that couldinteract with the adenosinergic system, thereby enhancing the neuroprotectiveeffects of adenosine. All other members of the IL-6 cytokine family (IL-11, LIF,OSM, CNTF and CT-1), use the same signaling cascade and are known to induceneuronal survival [16]. Interestingly, no research has been performed to checkwhether these neuroprotective effects are due to an interaction with theadenosinergic system. More research is needed to study this complex system andits interactions with cytokines, neurotrophins and other substances.
Neuroprotective substances released by glia cells: IL-6 and more?
IL-6 is a neuroprotective cytokine and is released by astrocytes upon adenosinereceptor stimulation. As described in chapter 2, astrocytes and microglia areknown to release other neuroprotective factors upon stimulation of variousadenosine receptor types. Using RT-PCR analysis we have screened mRNA levels
Chapter 6
110

of factors considered to be involved in neuroprotection like VEGF and pentraxin 3(PTX3), to determine whether adenosine stimulates the release of these factorsfrom glia cells. We also screened mRNA levels of chemokines like CCL2 andRANTES. We thus found that cultured astrocytes showed increased expression ofCCL2 mRNA after adenosine receptor stimulation (data not shown). Furtheranalysis of protein levels using ELISA showed that CCL2 protein release fromastrocytes was mediated by the adenosine A3 receptor subtype (chapter 3).This is a remarkable finding, since the role of the A3 receptor in the brain is stillunclear. Although previous reports suggest that the adenosine A3 receptormediates neuroprotection, our findings are among the first to show an action ofadenosine A3 receptors [1, 6, 17]. Our results indicate A3 might also be involvedin adenosine-induced neuroprotection via the induction of the potentiallyneuroprotective chemokine CCL2. CCL2 has been shown to mediate neuroprotective effects [4, 5], although its’mechanism is unknown. CCL2 may also have beneficial effects on astrocytes.CCL2 has been shown to induce cytoskeleton rearrangement, which might bebeneficial for cell survival. Thus the effect of A3 receptor stimulation oncytoskeleton rearrangement reported previously [1] could be explained by theeffect of the adenosine A3 receptor on the release of CCL2.
Conclusions
Adenosine is an important endogenous neuroprotector in the brain. In this thesiswe have investigated mechanisms of adenosine-induced neuroprotection. Wehave shown that IL-6 is involved in upregulation of the adenosine A1 receptor invivo, resulting in increased neuroprotection. This finding explains at least part ofthe neuroprotective properties of IL-6. Although the focus in this thesis is mainlyon interactions between adenosine and IL-6 (Figure 6.1B), these pathways mostlikely constitute only a small part of the complex interactions between adenosineand other factors, which have been reviewed in chapter 2 (Figure 6.1A). Since allfour adenosine receptor subtypes are involved, it is likely that the release of specificneuroprotective factors from glia cells might be precisely regulated by adenosine. Adenosine and adenosine receptors are thus involved in the complex interplaybetween neurons and glia cells. In this regard adenosine constitutes an importantfactor in neuron-glia communication like cytokines, chemokines and neurotrans-mitters [2, 8, 10, 15]. An improved understanding of the complex pathways of adenosine-inducedneuroprotection may reveal novel possibilities to enhance endogenous neuro-protective processes. This could lead to possible therapeutic opportunities totreat neurodegenerative disorders.
Summary and conclusions
111

Chapter 6
112
ATP
IL-6
(1)
ATP
AdoAdo
(1)
Ca2+-influxNT release
K+ ←←←
A1AR
NGF
S100BCCL2
A2AAR
P2Y
(4,5)
A1AR
A2BAR
A3AR
P2X7
(3)
astrocyte
microglia
neuron
(2) (2)(3)
A
B
upregulation
Neuropathologicalevent
(4)
stimulation(3)
Adenosine
Adenosine A1receptors
stimulation (2)
Adenosine A2Breceptors
release? (3)
IL-6
Neuroprotection
variousmechanisms? (2)(5)
release (1)
Neuropathologicalevent
Figure 6.1. A) Overview of neuroprotective substances released by glia cells and its effects onneurons. B) Interactions between adenosine and IL-6. Cascade of events in A and B: (1) Neuro-pathological events lead to an increase of extracellular adenosine concentration. (2) Adenosine A1receptors in neurons are stimulated, resulting in membrane hyperpolarization and inhibition ofneurotransmitter release, which will protect neurons. (3) Stimulation of adenosine A2B receptorson glia cells leads to a release of IL-6. The question mark indicates that it is not known if thisoccurs in vivo. (4) Interleukin-6 is involved in the upregulation of adenosine A1 receptors in vivo,which likely increases adenosine’s neuroprotective effects. (5) Beside upregulation of adenosine A1receptors there might be additional mechanisms of IL-6-induced neuroprotection.

References
Summary and conclusions
113
1. Abbracchio, M. P., Rainaldi, G., Giammarioli, A. M., Ceruti, S., Brambilla, R., Cattabeni, F., Barbieri,D., Franceschi, C., Jacobson, K. A., and Malorni, W. 1997. The A3 adenosine receptor mediates cell spreading, reorganization of actin cytoskeleton, and distribution of Bcl-XL: studies in human astroglioma cells. Biochem.Biophys.Res.Commun. 241(2): 297-304
2. Bezzi, P. and Volterra, A. 2001. A neuron-glia signalling network in the active brain. Curr.Opin.Neurobiol. 11(3): 387-394
3. Biber, K., Lubrich, B., Fiebich, B. L., Boddeke, H. W., and van Calker, D. 2001. Interleukin-6 enhances expression of adenosine A(1) receptor mRNA and signaling in cultured rat cortical astrocytes and brain slices. Neuropsychopharmacology 24(1): 86-96
4. Bruno, V., Copani, A., Besong, G., Scoto, G., and Nicoletti, F. 2000. Neuroprotective activity of chemokines against N-methyl-D-aspartate or beta-amyloid-induced toxicity in culture. Eur J Pharmacol 399(2-3): 117-121
5. Eugenin, E. A., D'Aversa, T. G., Lopez, L., Calderon, T. M., and Berman, J. W. 2003. MCP-1 (CCL2) protects human neurons and astrocytes from NMDA or HIV-tat-induced apoptosis. J Neurochem. 85(5): 1299-1311
6. Fedorova, I. M., Jacobson, M. A., Basile, A., and Jacobson, K. A. 2003. Behavioral characterization ofmice lacking the A3 adenosine receptor: sensitivity to hypoxic neurodegeneration.Cell Mol.Neurobiol. 23(3): 431-447
7. Fiebich, B. L., Biber, K., Gyufko, K., Berger, M., Bauer, J., and van Calker, D. 1996. Adenosine A2b
receptors mediate an increase in interleukin (IL)-6 mRNA and IL-6 protein synthesis in human astroglioma cells. J Neurochem 66(4): 1426-1431
8. Fields, R. D. and Stevens-Graham, B. 2002. New insights into neuron-glia communication. Science298(5593): 556-562
9. Gadient, R. A. and Otten, U. H. 1997. Interleukin-6 (IL-6)--a molecule with both beneficial and destructive potentials. Prog Neurobiol 52(5): 379-390
10. Hanisch, U. K. 2002. Microglia as a source and target of cytokines. Glia 40(2): 140-15511. Kunioku, H., Inoue, K., and Tomida, M. 2001. Interleukin-6 protects rat PC12 cells from serum
deprivation or chemotherapeutic agents through the phosphatidylinositol 3-kinase and STAT3 pathways. Neurosci.Lett. 309(1): 13-16
12. Noji, H. 1998. The rotary enzyme of the cell: the rotation of F1-ATPase. Science 282(5395): 1844-1845
13. Otten, U., Marz, P., Heese, K., Hock, C., Kunz, D., and Rose-John, S. 2001. Signals regulating neurotrophin expression in glial cells. Prog.Brain Res. 132: 545-554
14. Schwaninger, M., Neher, M., Viegas, E., Schneider, A., and Spranger, M. 1997. Stimulation of interleukin-6 secretion and gene transcription in primary astrocytes by adenosine. J Neurochem69(3): 1145-1150
15. Streit, W. J., Walter, S. A., and Pennell, N. A. 1999. Reactive microgliosis. Prog.Neurobiol. 57(6): 563-581
16. Taga, T. and Kishimoto, T. 1997. Gp130 and the interleukin-6 family of cytokines. Annu.Rev.Immunol. 15: 797-819
17. von Lubitz, D. K., Lin, R. C., Popik, P., Carter, M. F., and Jacobson, K. A. 1994. Adenosine A3
receptor stimulation and cerebral ischemia. Eur.J Pharmacol. 263(1-2): 59-67

114

115
Nederlandse samenvatting
Betrokkenheid van glia cellen en cytokinesbij neuroprotectieve processen
geïnduceerd door adenosine

Inleiding
Adenosine tri-fosfaat (ATP) is een belangrijke energierijke verbinding, die dienstdoet als energiedonor voor allerlei cellulaire processen. In situaties waarbij deenergie consumptie verhoogd is, wordt meer ATP afgebroken dan gevormd. Deconcentratie van het afbraakproduct, adenosine stijgt dan snel en wordtonmiddellijk over de celmembraan de cel uit getransporteerd. Dit extracellulaireadenosine bindt aan adenosine receptoren op de celmembraan, waardoor proces-sen in werking worden gezet die het metabolisme van de cel afremmen. Zo wordthet evenwicht tussen ATP afbraak en productie weer hersteld. Dit is een zeerbasaal mechanisme dat in alle celtypes aanwezig is. In de hersenen is de concentratie adenosine verhoogd tijdens neuropathologischeaandoeningen zoals herseninfarcten en epilepsie. Onder deze omstandighedenheeft adenosine neuroprotectieve eigenschappen die gemediëerd worden dooradenosine A1 receptoren. Verhoging van het aantal receptoren resulteert in eensterker neuroprotectief effect van adenosine. Langdurige blootstelling van adeno-sine A1 receptoren aan hoge concentraties adenosine, zoals tijdens neuropatho-logie kan echter leiden tot een vermindering van het aantal receptoren. Om debeschermende werking van adenosine te handhaven, is het noodzakelijk dat hetaantal functionele receptoren in de membraan op peil blijft. Voorafgaandonderzoek in gekweekte hersencellen heeft aangetoond dat het pro-inflammatoirecytokine, interleukine-6 (IL-6) hier waarschijnlijk bij betrokken is. Stimulatie vanadenosine receptoren in glia cellen resulteerde in een verhoogde afgifte van IL-6.Bovendien werd een verhoging van het aantal adenosine A1 receptoren in gliacellen geconstateerd nadat deze cellen met IL-6 waren gestimuleerd.
In dit proefschrift werd de hypothese onderzocht of IL-6 ook in vivo betrokken isbij de regulatie van adenosine A1 receptoren tijdens hersenschade. Om deze hypothese te kunnen bewijzen, is een reeks van experimenten gedaandie staan beschreven in hoofdstukken 3, 4 en 5.
Eigenschappen van de muis adenosine A1 receptor
Hoewel de eigenschappen van de adenosine A1 receptor van veel diersoortenbekend zijn, zijn er nauwelijks gegevens in de literatuur over de farmacologie vande muis adenosine A1 receptor. Omdat we de muis als proefdier wilden gebruikenin onze experimenten, was het nodig om eerst de bindingseigenschappen vanliganden voor de muis receptor te onderzoeken. Hiertoe werd de adenosine A1
receptor gekloneerd uit muis astrocyten en in de CHO cellijn tot expressiegebracht. Dit proces, evenals de resultaten zijn beschreven in hoofdstuk 4. De
Nederlandse samenvatting
116

resultaten laten zien dat de muis receptor, qua structuur en eigenschappenovereenkomt met de adenosine A1 receptor van de rat en de mens. Dit betekentdat we in onze experimenten de liganden kunnen gebruiken die met succesgebruikt zijn bij ratten.
Betrokkenheid van IL-6 bij de regulatie van adenosine A1 receptoren
Om de rol van IL-6 te kunnen onderzoeken (hoofdstuk 5), werd gebruik gemaaktvan een IL-6 knock out model; bij de IL-6 knock out muizen is het IL-6 gen d.m.v.genetische manipulatie uitgeschakeld. In controle en IL-6 knock out muizen werden kunstmatig convulsies opgewektd.m.v. injecties met de convulsant pentylenetetrazole (PTZ), waarna de expressievan adenosine A1 receptoren werd onderzocht. IL-6 knock out muizen waren veel gevoeliger voor PTZ dan controle muizen. Ditverschil zou wellicht veroorzaakt kunnen zijn door een verschil in adenosine A1
receptor expressie.Uit autoradiografie experimenten bleek dat bij controle muizen waarin convulsieswaren opgewekt, de expressie van adenosine A1 receptoren in de hippocampus eneen deel van de cortex was toegenomen. Bij IL-6 knock out muizen was er echtergeen effect van convulsies op adenosine receptor expressie waarneembaar. Hieruitkan geconcludeerd worden dat IL-6 een rol speelt bij de regulatie van adenosineA1 receptor expressie tijdens convulsies. IL-6 stimuleert op deze manier deneuroprotectieve effecten van adenosine. In de huidige literatuur worden tegenstrijdige (schadelijke en beschermende)effecten van IL-6 beschreven. Onze resultaten suggereren dat IL-6 een belangrijkeneuroprotectieve rol speelt in de hersenen. Bovendien geven de resultaten inzichtin het mechanisme van de beschermende werking van IL-6.
Afgifte van neuroprotectieve stoffen door glia cellen
Verder is in dit proefschrift de rol van adenosine receptoren in glia cellenonderzocht. Het is algemeen bekend dat stimulatie van adenosine receptoren inglia cellen resulteert in de productie en afgifte van neuroprotectieve stoffen, zoalsnerve growth factor (NGF), S100B en IL-6. In dit proefschrift is gevonden datastrocyten (het meest voorkomende glia cel type) na stimulatie van de adenosineA3 receptor het chemokine CCL2 afgeven (hoofdstuk 3). Dit is een bijzonderevinding omdat niet bekend is welke rol de adenosine A3 receptor speelt in dehersenen. Onze resultaten laten zien dat de adenosine A3 receptor, door het sti-muleren van de afgifte van het mogelijk neuroprotectieve chemokine CCL2,
Nederlandse samenvatting
117

betrokken zou kunnen zijn bij neuroprotectieve processen geïnduceerd dooradenosine.
Conclusie
Samenvattend heeft dit proefschrift bijgedragen aan het inzicht in de complexeinteracties tussen adenosine, adenosine receptoren en cytokines. Adenosine is een belangrijke stof met intrinsieke beschermende eigenschappen inde hersenen. In dit proefschrift hebben we de mechanismen onderzocht van deneuroprotectieve processen geïnduceerd door adenosine. We hebben laten ziendat het cytokine IL-6 ook in vivo betrokken is bij regulatie van adenosine A1
receptoren en daardoor de neuroprotectieve eigenschappen van adenosine kanversterken. Hoewel de focus in dit proefschrift voornamelijk gericht is op deinteracties tussen adenosine en IL-6, maken deze slechts deel uit van een grotergeheel van complexe interacties tussen adenosine en andere factoren (samengevatin hoofdstuk 2). Omdat alle vier adenosine receptoren erbij betrokken zijn, is het waarschijnlijkdat de afgifte van specifieke neuroprotectieve stoffen door glia cellen preciesgereguleerd wordt door adenosine. Zo zijn adenosine and adenosine receptorenbetrokken bij de complexe wisselwerking tussen neuronen en glia cellen. Hieruitkan geconcludeerd worden dat adenosine, evenals cytokines, chemokines enneurotransmitters, een rol speelt bij de communicatie tussen neuronen en gliacellen. Verbeterde kennis van de ingewikkelde processen verantwoordelijk voor deneuroprotectieve werking van adenosine, zou nieuwe mogelijkheden kunnencreëren om de intrinsieke beschermende werking van adenosine te versterken.Dit zou uiteindelijk kansen kunnen scheppen om methoden te ontwikkelen voorde behandeling van neurodegeneratieve ziekten.
Nederlandse samenvatting
118

Hier rest voor mij nog de taak om personen te bedanken die op de één of anderemanier hebben bijgedragen aan de totstandkoming van dit proefschrift. Voordatik begin met het opnoemen van namen, wil ik eerst mijn excuses aanbieden aandiegenen die ik hier wellicht vergeten ben te noemen.Allereerst wil ik hier mijn promotor, Erik Boddeke en copromotor Knut Biberbedanken. Op allerlei vlakken stonden ze voor me klaar. Erik was er met zijn uitdagendevragen en advies. Met Knut kon ik van gedachten wisselen over allerleiwetenschappelijke zaken maar natuurlijk vooral over adenosine en adenosinereceptoren, ook het onderwerp van zijn eigen proefschrift. Verder wil ik een aantal mensen buiten de vakgroep bedanken voor hun bijdragenaan dit proefschrift. Ad IJzerman en Jacobien Künzel voor het uitvoeren van deradioligand bindingsstudies, Katalin Horvath, Jan Keijser en Viktor Roman voorhun hulp bij de autoradiografie experimenten. Auke Meinema en GerardusZuidema voor het beschikbaar stellen van muis neonaten voor de glia celkweek. Voor hun praktische ondersteuning wil ik een aantal collega’s speciaal bedanken: Nieske Brouwer, Marjolein Hensens, Britta Küst en Ietje Mantingh-Otter. Mijn (ex-) kamergenoten, Ineke Dijkstra (alvast gefeliciteerd met jouw promotieop 20 oktober!), Eiko de Jong en Mike Zuurman, wil ik bedanken voor degezelligheid en meer. Ook al was ik de enige niet-chemokine A.I.O. tussen jullie,ik heb zeker wat gehad aan onze wetenschappelijke gedachtewisselingen. Tenslotte wil ik alle andere collega’s bedanken, die de vakgroep medischefysiologie maakten tot een zeer plezierige werkplek: Rob Bakels, VeerakumarBalasubramaniyan, Valentine Bouet, Sjef Copray, Hiske van Duinen, AlbertGramsbergen, Josje IJkema-Paassen, Koen Jansen, Daniel Kernell, Jaap van derLeest, Joke van der Mark-van der Wouden, Wieb Patberg, Angelica Rappert,Manon van Riezen, Trix van der Sluis-Rozema, Koen Vaartjes, Evelyn Wesselingen Inge Zijdewind.Gerry Hoogenberg wil ik bedanken voor haar administratieve ondersteuning ende leuke gesprekken op haar houten bankje.
119
Dankwoord

Een groot deel van hoofdstuk 1 en 2 zijn geschreven in de tijdvrije ruimte van hetBiologisch centrum in Haren, waar ik als proefpersoon meedeed aan eenonderzoek van de vakgroep Chronobiologie. Daarom bedank ik hierbij MarijkeGordijn, Andrei Zavada en de studenten voor het aangename verblijf daar in dekelder.Kameraad Sylvia Brugman wil ik bedanken voor de steun in onze gezamenlijkestrijd voor politieke vrijheid van meningsuiting in het FMW- gebouw.Saskia van der Schaaf wil ik niet alleen bedanken voor het toepasselijke ontwerpvoor de omslag van dit proefschrift, maar ook voor onze jarenlange vriendschap. Mijn ouders wil ik bedanken voor alles wat ze voor mij hebben gedaan, zodat ikdit kon bereiken.
Tenslotte wil ik nog een heel bijzonder iemand bedanken. Kwang-Yul Jang,zonder jouw liefde, gezelschap en steun was dit proefschrift er niet geweest.
Dankwoord
120





























