Embed Size (px)
Citation preview

Neurobiology of Disease 43 (2011) 86–98
Contents lists available at ScienceDirect
Neurobiology of Disease
j ourna l homepage: www.e lsev ie r.com/ locate /ynbd i
Adult neural precursors isolated from post mortem brain yield mostly neurons:An erythropoietin-dependent process
Giovanni Marfia a, Laura Madaschi a, Francesca Marra a, Mauro Menarini b, Daniele Bottai a,Alessandro Formenti c, Carmelo Bellardita c, Anna Maria Di Giulio a, Stephana Carelli a,d, Alfredo Gorio a,d,⁎a Laboratory of Pharmacology, Department of Medicine, Surgery and Dentistry, University of Milan, Polo H. San Paolo, via A. di Rudinì 8, Milan, Italyb Ospedale Montecatone, via Montecatone 37, Imola, Bologna, Italyc Department of Human Physiology, School of Medicine, University of Milan, via Mangiagalli 32, Italyd Clinical Pharmacology, IRCCS Humanitas, via Manzoni 56, Rozzano, Milan, Italy
Abbreviations: PM-NPCs, Post-Mortem Neural PrecuEPO-R, Erythropoietin Receptor; NPCs, Neural PrecursorSVZ, Sub Ventricular Zone.⁎ Corresponding author at: Laboratory of Pharmaco
Surgery andDentistry, University ofMilan, PoloH. San PaoFax: +39 0250323033.
E-mail address: [email protected] (A. Gorio).Available online on ScienceDirect (www.scienced
0969-9961/$ – see front matter © 2011 Elsevier Inc. Aldoi:10.1016/j.nbd.2011.02.004
a b s t r a c t
a r t i c l e i n f oArticle history:Received 23 December 2010Revised 2 February 2011Accepted 7 February 2011Available online 13 February 2011
Keywords:Adult neural progenitorsErythropoietinGene regulationHypoxia- inducible factor-1 (HIF-1)mTORNeural plasticity
This studywas aimed at the isolation of neural precursor cells (NPCs) capable of resisting to a prolonged ischemicinsult as thismay occur at the site of traumatic and ischemic CNS injuries. Adultmicewere anesthetized and thenkilled by cervical dislocation. The cadavers were maintained at room temperature or at 4 °C for different timeperiods. Post mortem neural precursors (PM-NPCs) were isolated, grown in vitro and their differentiationcapability was investigated by evaluating the expression of different neuronal markers. PM-NPCs differentiatemostly in neurons, show activation of hypoxia-inducible factor-1 and MAPK, and express both erythropoietin(EPO) and its receptor (EPO-R). The exposureof PM-NPCs toneutralizing antibodies to EPOorEPO-Rdramaticallyreduced the extent of neuronal differentiation to about 11% of total PM-NPCs. The functionality of mTOR andMAPK is also required for the expression of the neuronal phenotype by PM-NPCs. These results suggest thatPM-NPCs can be isolated from animal cadaver even several hours after death and their self-renewable capabilityis comparable to normal neural precursors. Differently, their ability to achieve a neural phenotype is superior tothat of NPCs, and this ismediated by the activation of hypoxia-induced factor 1 and EPO signaling. PM-NPCsmayrepresent good candidates for transplantation studies in animal models of neurodegenerative diseases.
rsor Cells; EPO, Erythropoietin;Cells; SCI, Spinal Cord Injury;
logy, Department of Medicine,lo, via A di Rudinì 8,Milan, Italy.
irect.com).
l rights reserved.
© 2011 Elsevier Inc. All rights reserved.
Introduction
Traumatic lesions of the cord are characterized by a secondaryprocess of degeneration, that is a complex condition of ischemia-likesyndrome and neuroinflammation leading to a large expansion of theearly mechanical damage (Bottai et al., 2008, 2010; Moreno-Manzanoet al., 2009, 2010; Popovich et al., 2003; Ronaghi et al., 2010). Asuccessful transplantation of neural precursors requires their survival insuch an unfavorable environment. We have previously reported thatneural precursor cells (NPCs), accumulate at spinal cord injury (SCI) siteand improve the rate of hind limb functional recovery. Their viability inthe injured spinal cord, however, lasts 12–20 days, then NPCs arephagocytated by macrophages (Bottai et al., 2008). A similar outcomewas observed with embryonic stem cells (Bottai et al., 2010) In view ofsuch results we aimed at isolating adult neural stem cells from the
subventricular zone (SVZ) after a prolonged global ischemia. Theisolation of ischemia-resistant neural precursors may supply cells withproperties different from NPCs, and able to survive in the unfavorableenvironment of the traumatically injured central nervous system(Molcanyi et al., 2007; Popovich et al., 2003).
Oxygen is an important energy source for cell metabolism (Bruick,2003), and its utilization is tightly regulated in the CNS (Hoge and Pike,2001). Lower oxygen levels determine hypoxia, that affects cellularproliferation and differentiation, and may have salutary effects onneural precursors (Chen et al., 2007; Studer et al., 2000) and neurons(Moreno-Marzano et al., 2010; Zhu et al., 2005). For instance dopamineexpression is induced by hypoxia in bone marrow-derived mesenchy-mal stem cells and P19 cells (Semenza, 2000; Wu et al., 2008). Hypoxiamay also positively influence stem cell differentiation (Santilli et al.,2010). The cellular responses to hypoxia aremanifested by activation ofthe hypoxia-inducible factor-1 (HIF-1), a transcriptional factor consist-ing of two subunits, the O2-regulated HIF-1 α subunit and theO2-independent HIF-1 β subunit. Under hypoxic conditions, HIF-1 αandβdimerize andmigrate to thenucleus; this enhances the expressionof several key target genes such as erythropoietin (EPO), vascularendothelial growth factor, tyrosinehydroxylase andOCT4,which acts asregulators of cellular proliferation and differentiation (Adelman andSimon, 2002; Ramírez-Bergeron and Simon, 2001; Richard et al., 1999).
87G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
Hypoxia also activates ERK1/2, which phosphorylates HIF-1αdirectly, thereby enhancing HIF-1α transcriptional activity (Berraet al., 2000; Minet et al., 2000). In this study we report that we wereable to isolate from the SVZ adult neural stem cells resistant to theischemic condition present at several hours after mouse death (6 and16 h). These cells were called Post-Mortem Neural Precursor Cells(PM-NPCs). Their proliferation is similar to that of NPCs obtained atkilling time (T0) (Gritti et al., 1996), while their differentiation yieldsabout 33% β tubulin III- and 36% of MAP2-positive cells compared to10–12% of NPCs. Differentiated PM-NPCs show higher HIF-1α activa-tion, express both EPO and EPO-R, and active voltage-dependent Ca++
channels. Such a higher differentiation requires the functionality ofmTOR andMAPK systems and is prevented by exposure to anti-EPO andanti-EPO-R antibodies. This study shows the existence of novel typeEPO-dependent adult neural stem cells, that can be extracted from thebrain of cadavers, andmay represent a more successful approach to thetreatment of traumatic CNS injuries.
Materials and methods
Ethics statement
All animals were handled in strict accordance with good animalpractice as defined by the Italian Guidelines for Laboratory Animals
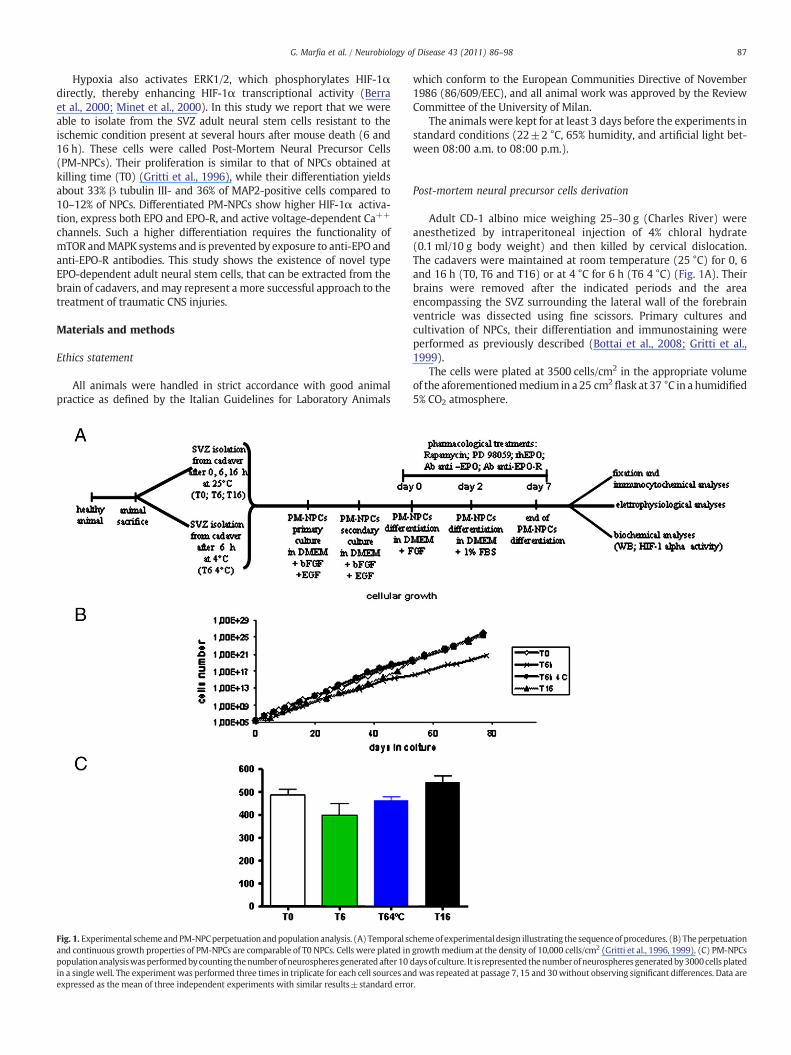
Fig. 1. Experimental schemeandPM-NPCperpetuation andpopulation analysis. (A) Temporal scand continuous growth properties of PM-NPCs are comparable of T0 NPCs. Cells were plated inpopulation analysiswasperformedbycounting thenumber of neurospheresgeneratedafter10in a single well. The experiment was performed three times in triplicate for each cell sources anexpressed as the mean of three independent experiments with similar results±standard erro
which conform to the European Communities Directive of November1986 (86/609/EEC), and all animal work was approved by the ReviewCommittee of the University of Milan.
The animals were kept for at least 3 days before the experiments instandard conditions (22±2 °C, 65% humidity, and artificial light bet-ween 08:00 a.m. to 08:00 p.m.).
Post-mortem neural precursor cells derivation
Adult CD-1 albino mice weighing 25–30 g (Charles River) wereanesthetized by intraperitoneal injection of 4% chloral hydrate(0.1 ml/10 g body weight) and then killed by cervical dislocation.The cadavers were maintained at room temperature (25 °C) for 0, 6and 16 h (T0, T6 and T16) or at 4 °C for 6 h (T6 4 °C) (Fig. 1A). Theirbrains were removed after the indicated periods and the areaencompassing the SVZ surrounding the lateral wall of the forebrainventricle was dissected using fine scissors. Primary cultures andcultivation of NPCs, their differentiation and immunostaining wereperformed as previously described (Bottai et al., 2008; Gritti et al.,1999).
The cells were plated at 3500 cells/cm2 in the appropriate volumeof the aforementionedmedium in a 25 cm2
flask at 37 °C in a humidified5% CO2 atmosphere.
hemeof experimentaldesign illustrating the sequenceof procedures. (B) Theperpetuationgrowth medium at the density of 10,000 cells/cm2 (Gritti et al., 1996, 1999). (C) PM-NPCsdaysof culture. It is represented thenumberofneurospheres generatedby3000cells platedd was repeated at passage 7, 15 and 30without observing significant differences. Data arer.
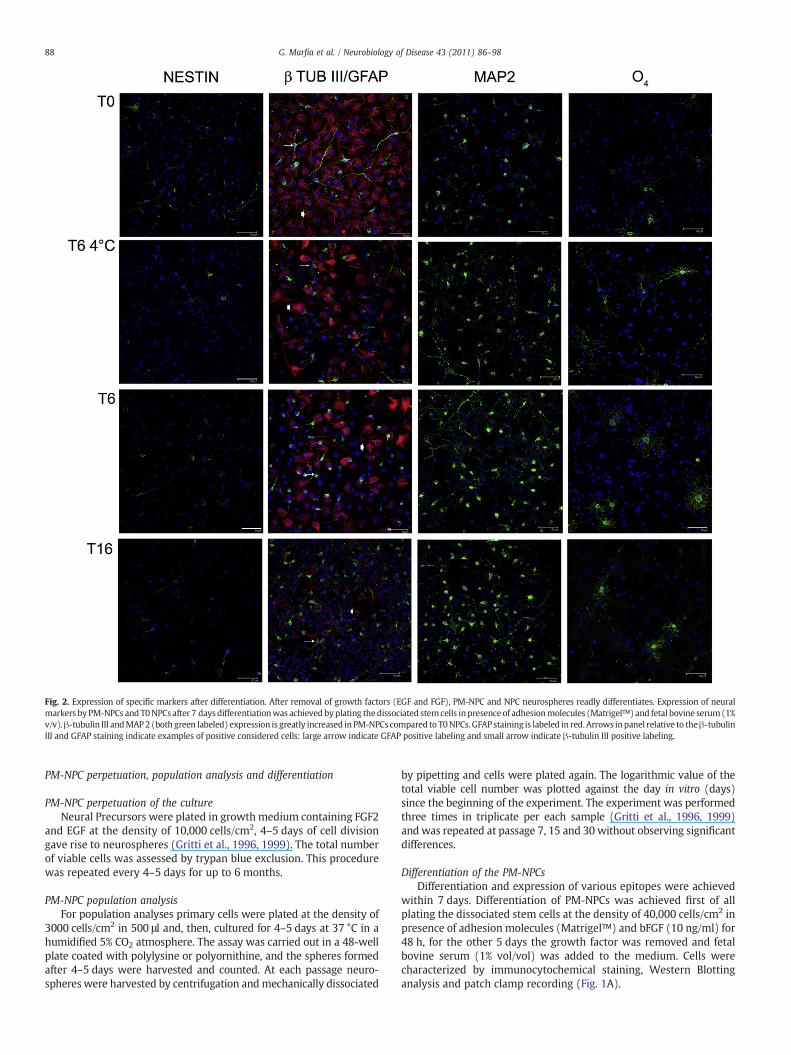
Fig. 2. Expression of specific markers after differentiation. After removal of growth factors (EGF and FGF), PM-NPC and NPC neurospheres readly differentiates. Expression of neuralmarkers by PM-NPCs and T0NPCs after 7 daysdifferentiationwas achieved byplating thedissociated stemcells in presence of adhesionmolecules (Matrigel™) and fetal bovine serum(1%v/v).β-tubulin III andMAP2 (both green labeled) expression is greatly increased in PM-NPCs compared to T0NPCs. GFAP staining is labeled in red.Arrows inpanel relative to theβ-tubulinIII and GFAP staining indicate examples of positive considered cells: large arrow indicate GFAP positive labeling and small arrow indicate β-tubulin III positive labeling.
88 G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
PM-NPC perpetuation, population analysis and differentiation
PM-NPC perpetuation of the cultureNeural Precursors were plated in growth medium containing FGF2
and EGF at the density of 10,000 cells/cm2, 4–5 days of cell divisiongave rise to neurospheres (Gritti et al., 1996, 1999). The total numberof viable cells was assessed by trypan blue exclusion. This procedurewas repeated every 4–5 days for up to 6 months.
PM-NPC population analysisFor population analyses primary cells were plated at the density of
3000 cells/cm2 in 500 μl and, then, cultured for 4–5 days at 37 °C in ahumidified 5% CO2 atmosphere. The assay was carried out in a 48-wellplate coated with polylysine or polyornithine, and the spheres formedafter 4–5 days were harvested and counted. At each passage neuro-spheres were harvested by centrifugation andmechanically dissociated
by pipetting and cells were plated again. The logarithmic value of thetotal viable cell number was plotted against the day in vitro (days)since the beginning of the experiment. The experiment was performedthree times in triplicate per each sample (Gritti et al., 1996, 1999)and was repeated at passage 7, 15 and 30 without observing significantdifferences.
Differentiation of the PM-NPCsDifferentiation and expression of various epitopes were achieved
within 7 days. Differentiation of PM-NPCs was achieved first of allplating the dissociated stem cells at the density of 40,000 cells/cm2 inpresence of adhesion molecules (Matrigel™) and bFGF (10 ng/ml) for48 h, for the other 5 days the growth factor was removed and fetalbovine serum (1% vol/vol) was added to the medium. Cells werecharacterized by immunocytochemical staining, Western Blottinganalysis and patch clamp recording (Fig. 1A).

89G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
The immunostaining was performed as follows, differentiatedPM-NPCs were fixed with 4% paraformaldehyde in PBS for 10 min atroom temperature, and, then, washed with PBS. The coverslips wereincubated for 90 min at 37 °C (or overnight at 4 °C) in PBS containing10% normal goat serum (NGS), 0.3% Triton X-100, and the appropriateprimary antibodies or antisera. After thorough washing with PBS and10% NGS, cells were reacted for 45 min (room temperature) withsecondary antibody Alexa Fluor 488 or 543 anti mouse or rabbit(Invitrogen, Carlsbad, California). The cells were then counterstainedwith the DNA-binding dye 4′-6′-diamidino-2-phenylindole (DAPI)(2 μg/ml in PBS) for 10 min at room temperature, twice washed inPBS followed. Coverslips containing the stained differentiated cellsweremounted onto glass slides and examined under a fluorescencemicroscope (Leica, Wetzlar, Germany). Acquisition of the stained cellswas performed using the image-analysis software (Leica, Wetzlar,Germany) or by confocal microscope Leica SP2 microscope with He/Krand Ar lasers (Leica, Wetzlar, Germany). Images were taken with a 40×objective lens and 1× zoom (Leica). In control experiments, primaryantibodies were either omitted or replaced with equivalent concentra-tions of unrelated Ig of the same subclass. Moreover, in double labelingexperiments, sections incubated with one primary antibody and twosecondary anti-sera revealed no appreciable cross-reaction. Neuraldifferentiation was assessed by immunocytochemistry with antibodiesagainst: Nestin (monocl.1:100; Pharmingen, San Jose, CA, USA),β-tubulin III (monocl. 1:150; Sigma), microtubule-associated protein 2(MAP2; monocl. 1:200; Sigma), Glial fibrillary acidic protein (GFAP;monocl. 1:400; Roche, Basel, CH), Coline Acetyl Transferase (ChAT;monocl. 1:1000; Chemicon, Temecula, CA), Tyrosine Hydroxylase(TH; monocl. 1:400; Chemicon), γ-aminobutyrric acid (GABA; monocl.1:500; Chemicon), Glutammate (GLU; monocl. 1:400; Chemicon),Serotonin (5HT; monocl. 1:50; Chemicon), Dopamine Transporter(DAT; monocl. 1:400; Chemicon), O4 (O4, sulfatide mAb clone:81 1:150;Millipore,Milano, Italy) PAX6 (monocl. 1:6; IowaUniversity, USA),3CB2 (monocl. 1:50; Iowa University, USA), and RC2 (monocl. 1:50;Iowa University, USA).
Pharmacological treatmentsIn order to investigate the role of erythropoietin pathway, NPCs
and PM-NPCs were differentiated as described above in presence ofdifferent amount of human recombinant erythropoietin (rh-EPO,Janssen & Cilag, Italy), or antibodies against both erythropoietin (anti-EPO; Santa Cruz, CA; 3 μg/ml) or erythropoietin receptor (anti-EPO-R;Santa Cruz CA; 3 μg/ml) during 7 days.
To investigate MAPK (ERK 1/2) andmTOR involvement in PM-NPCdifferentiation, NPCs and PM-NPCs were exposed to PD98059 (3 μMand 30 μM) or rapamycin (0.05 ng/ml) throughout the differentiationtest. In PD98059 and rapamycin treatments, stock solutions wereprepared in Dimethylsulfoxide (DMSO) and diluted in differentiationmedium to reach the final concentration (see above). Control culturesreceived vehicle alone (0.001% DMSO). Differentiation media with orwithout rapamycin or PD98059 were replenished every 48 h duringthe course of experiments. Each differentiation assay was performedthree times and repeated at passage 7, 15 and 30 without observingsignificant differences.
Cell count
The quantification of positive cells to neural markers (see above)was performed, by confocal microscopy, after specifical immunocy-tochemical staining following 7 days differentiation in culture. Briefly,cell counts were performed on a minimum of 9 independent fields(3 fields/3 coverslips/treatment) of photomicrographs captured with40× objective. Total counts of each neural markers immunoreactivecells were performed and the number of positive cells per culture wasexpressed as the percentage to the total cells. DAPI supplied the totalnumber of cells being a nuclear staining.
Protein extraction and Western Blotting
Cell protein extracts were obtained by means of TRIzol™ reagentfollowing manufacturer instructions (Invitrogen, Carlsbad, California).Equal amounts of solubilized protein (20 μg) were heated in Laemmlisample buffer (Biorad, Hercules, CA) containing 2-β mercaptoethanol(70 mM), separated by SDS-PAGE gel and electroblotted onto nitrocel-lulose membrane. The membrane was blocked in TBS with 0.05%Tween-20 (v/v), containing 5% slim milk (w/v), and probed withprimary antibodies overnight at 4 °C. The membrane was thenincubated with specific horseradish peroxidase-conjugated secondaryantibodies (1:10,000 dilution; Chemicon, Temecula, CA). The proteinswere visualized by means of an enhanced chemioluminescencedetection system (ECLTM, Amersham, Arlington Heights, IL). Afteracquisition by a GelDocTM image capture system (Kodak), theautoradiograms were quantified using Quantity OneTM software.Primary antibodies used were: erythropoietin (EPO; polyclonal 1:200;Santa Cruz, CA), erythropoietin receptor (EPO-R; polyclonal 1:200;SantaCruz) (Verdier et al., 2006) ERK (polyclonal 1: 1000. Cell SignallingTechnology, Denvers MA), phospho-ERK (polyclonal 1: 1000; CellSignalling Technology), and Hypoxia inducible factor 1 α (HIF-1 α,polyclonal 1:200; Vinci Biochem, Italy).
Evaluation of HIF-1 α activity detection
HIF-1 α activity detection was quantitatively evaluated by meansof TransAMTM HIF-1 assay Kit (Active Motif, Europe). Proteic nuclearphase was extracted according to the manufacturer instructions.Shortly, TransAMTM contains a 96-well plate on which has beenimmobilized an oligonucleotide containing a HIF responsive element(HRE) (5′-TACGTGCT-3′) from the EPO gene. HIF contained in nuclearextracts specifically binds to this oligonucleotide. The primaryantibody used in TransAMTM Kit recognizes an epitope on HIF-1 αprotein that is accessible upon DNA binding. Addition of a secondaryHRP-conjugated antibody provides a sensitive colorimetric readoutquantified by spectrophotometry. The nuclear extracts were preparedusing TransAMTM nuclear protein extraction kit starting from bothfloating T0 NPCs and PM-NPC neurospheres and after their differen-tiation. Following manufacturer instructions COS-7 cells (CoCl2treated cells) were used as positive control. Eight μg from eachsample of nuclear extract were employed to evaluate HIF activity.
Patch-clamp recordings
Whole-cell patch-clamp technique was utilized to assess thepresence of ion channels in the plasma membrane of both NPCs andPM-NPCs during their differentiation, as described previously (Cinoand Formenti, 2008), by means of pipettes of 3–4 MΩ resistance, withan Axopatch 200B integrating patch-clamp amplifier (Axon Instru-ments, USA) interfaced with a PC. Analog signals were low-passfiltered at 5 kHz and the data were digitized at sampling times of100 μs using a Digidata 1200 Series Interface (Axon Instruments).Stimulation, data acquisition and analysis were carried out usingpCLAMP programs by Axon Instruments. Data are expressed as meanvalue±SEM.
Statistical analysis
All of data are expressed as themean±Standard Error Mean (SEM).We performed an analysis of variance (ANOVA) test followed byBonferroni's post-test. Statistical analyses were performed usingGraphPad Prism 4.0 software; p values of b0.05 were consideredstatistically significant.
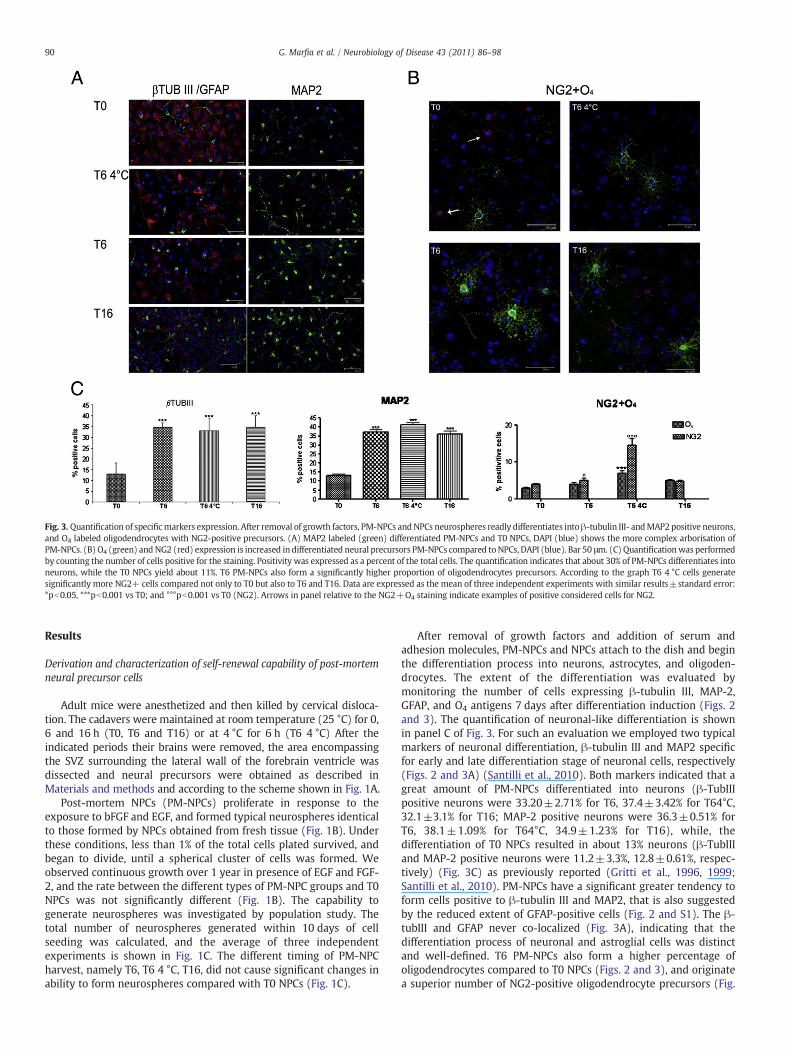
Fig. 3. Quantification of specificmarkers expression. After removal of growth factors, PM-NPCs andNPCs neurospheres readly differentiates intoβ-tubulin III- andMAP2 positive neurons,and O4 labeled oligodendrocytes with NG2-positive precursors. (A) MAP2 labeled (green) differentiated PM-NPCs and T0 NPCs, DAPI (blue) shows the more complex arborisation ofPM-NPCs. (B) O4 (green) and NG2 (red) expression is increased in differentiated neural precursors PM-NPCs compared toNPCs, DAPI (blue). Bar 50 μm. (C) Quantificationwas performedby counting the number of cells positive for the staining. Positivity was expressed as a percent of the total cells. The quantification indicates that about 30% of PM-NPCs differentiates intoneurons, while the T0 NPCs yield about 11%. T6 PM-NPCs also form a significantly higher proportion of oligodendrocytes precursors. According to the graph T6 4 °C cells generatesignificantly more NG2+ cells compared not only to T0 but also to T6 and T16. Data are expressed as the mean of three independent experiments with similar results±standard error:*pb0.05, ***pb0.001 vs T0; and °°°pb0.001 vs T0 (NG2). Arrows in panel relative to the ΝG2+O4 staining indicate examples of positive considered cells for NG2.
90 G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
Results
Derivation and characterization of self-renewal capability of post-mortemneural precursor cells
Adult mice were anesthetized and then killed by cervical disloca-tion. The cadavers were maintained at room temperature (25 °C) for 0,6 and 16 h (T0, T6 and T16) or at 4 °C for 6 h (T6 4 °C) After theindicated periods their brains were removed, the area encompassingthe SVZ surrounding the lateral wall of the forebrain ventricle wasdissected and neural precursors were obtained as described inMaterials and methods and according to the scheme shown in Fig. 1A.
Post-mortem NPCs (PM-NPCs) proliferate in response to theexposure to bFGF and EGF, and formed typical neurospheres identicalto those formed by NPCs obtained from fresh tissue (Fig. 1B). Underthese conditions, less than 1% of the total cells plated survived, andbegan to divide, until a spherical cluster of cells was formed. Weobserved continuous growth over 1 year in presence of EGF and FGF-2, and the rate between the different types of PM-NPC groups and T0NPCs was not significantly different (Fig. 1B). The capability togenerate neurospheres was investigated by population study. Thetotal number of neurospheres generated within 10 days of cellseeding was calculated, and the average of three independentexperiments is shown in Fig. 1C. The different timing of PM-NPCharvest, namely T6, T6 4 °C, T16, did not cause significant changes inability to form neurospheres compared with T0 NPCs (Fig. 1C).
After removal of growth factors and addition of serum andadhesion molecules, PM-NPCs and NPCs attach to the dish and beginthe differentiation process into neurons, astrocytes, and oligoden-drocytes. The extent of the differentiation was evaluated bymonitoring the number of cells expressing β-tubulin III, MAP-2,GFAP, and O4 antigens 7 days after differentiation induction (Figs. 2and 3). The quantification of neuronal-like differentiation is shownin panel C of Fig. 3. For such an evaluation we employed two typicalmarkers of neuronal differentiation, β-tubulin III and MAP2 specificfor early and late differentiation stage of neuronal cells, respectively(Figs. 2 and 3A) (Santilli et al., 2010). Both markers indicated that agreat amount of PM-NPCs differentiated into neurons (β-TubIIIpositive neurons were 33.20±2.71% for T6, 37.4±3.42% for T64°C,32.1±3.1% for T16; MAP-2 positive neurons were 36.3±0.51% forT6, 38.1±1.09% for T64°C, 34.9±1.23% for T16), while, thedifferentiation of T0 NPCs resulted in about 13% neurons (β-TubIIIand MAP-2 positive neurons were 11.2±3.3%, 12.8±0.61%, respec-tively) (Fig. 3C) as previously reported (Gritti et al., 1996, 1999;Santilli et al., 2010). PM-NPCs have a significant greater tendency toform cells positive to β-tubulin III and MAP2, that is also suggestedby the reduced extent of GFAP-positive cells (Fig. 2 and S1). The β-tubIII and GFAP never co-localized (Fig. 3A), indicating that thedifferentiation process of neuronal and astroglial cells was distinctand well-defined. T6 PM-NPCs also form a higher percentage ofoligodendrocytes compared to T0 NPCs (Figs. 2 and 3), and originatea superior number of NG2-positive oligodendrocyte precursors (Fig.
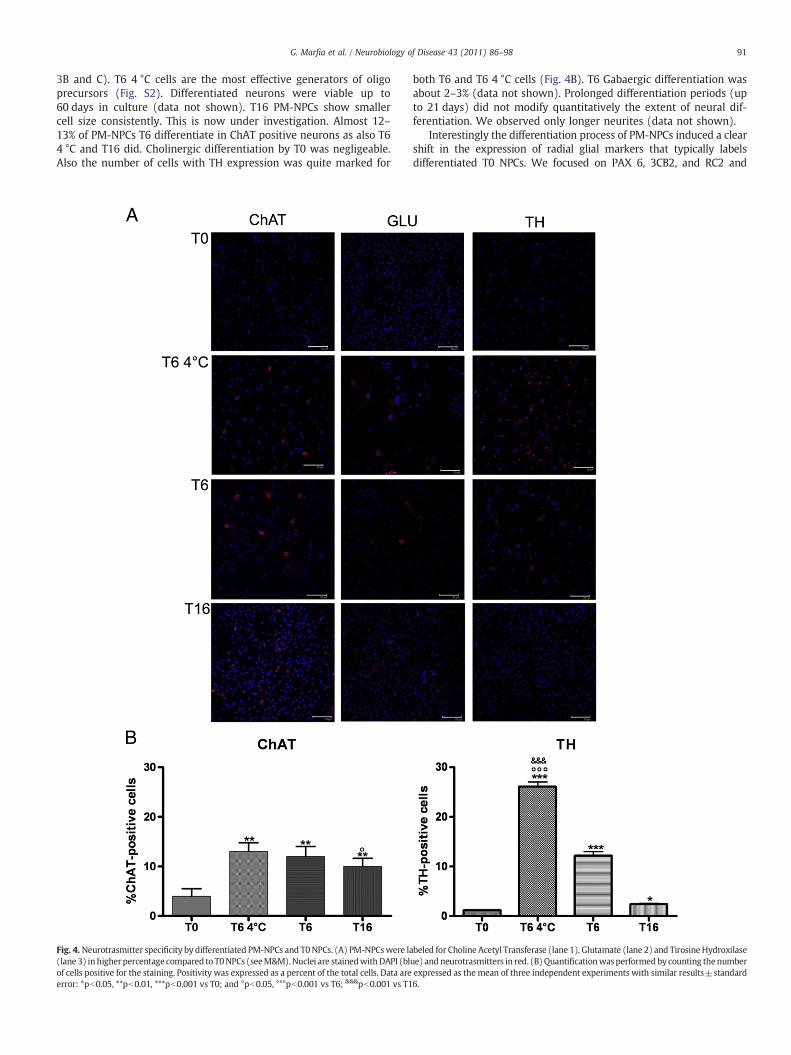
91G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
3B and C). T6 4 °C cells are the most effective generators of oligoprecursors (Fig. S2). Differentiated neurons were viable up to60 days in culture (data not shown). T16 PM-NPCs show smallercell size consistently. This is now under investigation. Almost 12–13% of PM-NPCs T6 differentiate in ChAT positive neurons as also T64 °C and T16 did. Cholinergic differentiation by T0 was negligeable.Also the number of cells with TH expression was quite marked for
Fig. 4. Neurotrasmitter specificity by differentiated PM-NPCs and T0NPCs. (A) PM-NPCswere la(lane 3) inhigherpercentage compared to T0NPCs (seeM&M).Nuclei are stainedwithDAPI (blof cells positive for the staining. Positivity was expressed as a percent of the total cells. Data areerror: *pb0.05, **pb0.01, ***pb0.001 vs T0; and °pb0.05, °°°pb0.001 vs T6; &&&pb0.001 vs T1
both T6 and T6 4 °C cells (Fig. 4B). T6 Gabaergic differentiation wasabout 2–3% (data not shown). Prolonged differentiation periods (upto 21 days) did not modify quantitatively the extent of neural dif-ferentiation. We observed only longer neurites (data not shown).
Interestingly the differentiation process of PM-NPCs induced a clearshift in the expression of radial glial markers that typically labelsdifferentiated T0 NPCs. We focused on PAX 6, 3CB2, and RC2 and
beled for Choline Acetyl Transferase (lane 1), Glutamate (lane 2) and Tirosine Hydroxilaseue) and neurotrasmitters in red. (B)Quantificationwas performedby counting thenumberexpressed as the mean of three independent experiments with similar results±standard6.
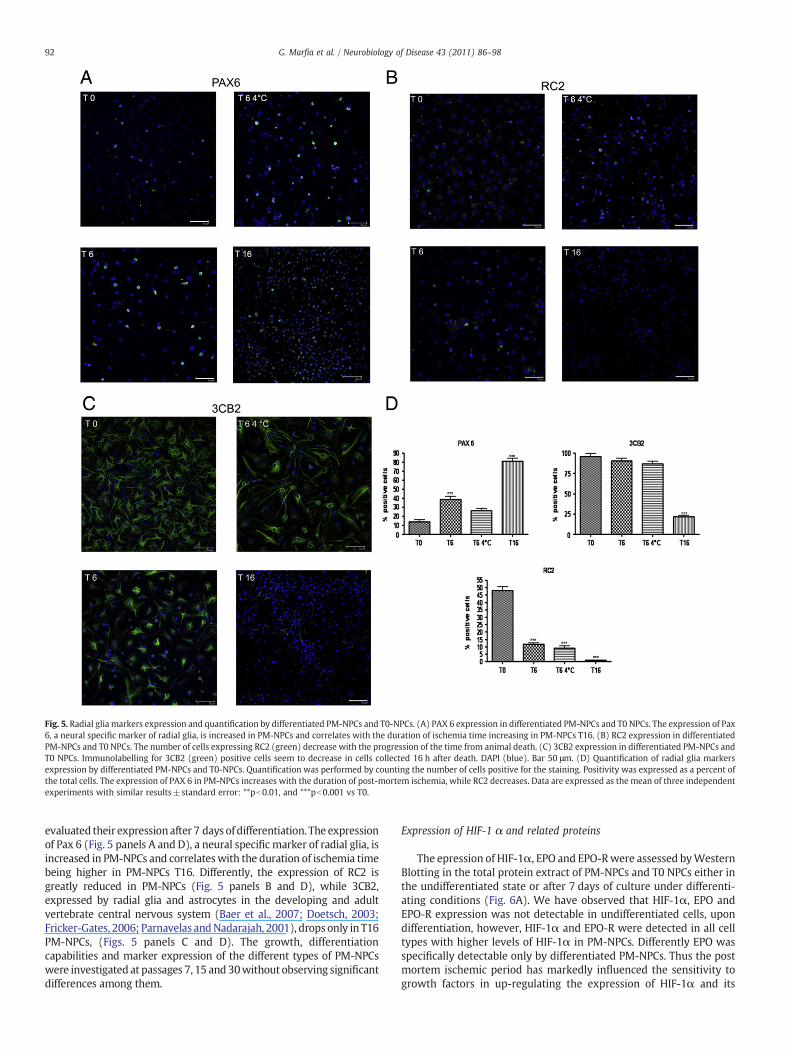
Fig. 5. Radial glia markers expression and quantification by differentiated PM-NPCs and T0-NPCs. (A) PAX 6 expression in differentiated PM-NPCs and T0 NPCs. The expression of Pax6, a neural specific marker of radial glia, is increased in PM-NPCs and correlates with the duration of ischemia time increasing in PM-NPCs T16. (B) RC2 expression in differentiatedPM-NPCs and T0 NPCs. The number of cells expressing RC2 (green) decrease with the progression of the time from animal death. (C) 3CB2 expression in differentiated PM-NPCs andT0 NPCs. Immunolabelling for 3CB2 (green) positive cells seem to decrease in cells collected 16 h after death. DAPI (blue). Bar 50 μm. (D) Quantification of radial glia markersexpression by differentiated PM-NPCs and T0-NPCs. Quantification was performed by counting the number of cells positive for the staining. Positivity was expressed as a percent ofthe total cells. The expression of PAX 6 in PM-NPCs increases with the duration of post-mortem ischemia, while RC2 decreases. Data are expressed as the mean of three independentexperiments with similar results±standard error: **pb0.01, and ***pb0.001 vs T0.
92 G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
evaluated their expression after 7 days of differentiation. The expressionof Pax 6 (Fig. 5 panels A and D), a neural specific marker of radial glia, isincreased in PM-NPCs and correlateswith the duration of ischemia timebeing higher in PM-NPCs T16. Differently, the expression of RC2 isgreatly reduced in PM-NPCs (Fig. 5 panels B and D), while 3CB2,expressed by radial glia and astrocytes in the developing and adultvertebrate central nervous system (Baer et al., 2007; Doetsch, 2003;Fricker-Gates, 2006; Parnavelas andNadarajah, 2001), drops only in T16PM-NPCs, (Figs. 5 panels C and D). The growth, differentiationcapabilities and marker expression of the different types of PM-NPCswere investigated at passages 7, 15 and 30without observing significantdifferences among them.
Expression of HIF-1 α and related proteins
The epression of HIF-1α, EPO and EPO-Rwere assessed byWesternBlotting in the total protein extract of PM-NPCs and T0 NPCs either inthe undifferentiated state or after 7 days of culture under differenti-ating conditions (Fig. 6A). We have observed that HIF-1α, EPO andEPO-R expression was not detectable in undifferentiated cells, upondifferentiation, however, HIF-1α and EPO-R were detected in all celltypes with higher levels of HIF-1α in PM-NPCs. Differently EPO wasspecifically detectable only by differentiated PM-NPCs. Thus the postmortem ischemic period has markedly influenced the sensitivity togrowth factors in up-regulating the expression of HIF-1α and its
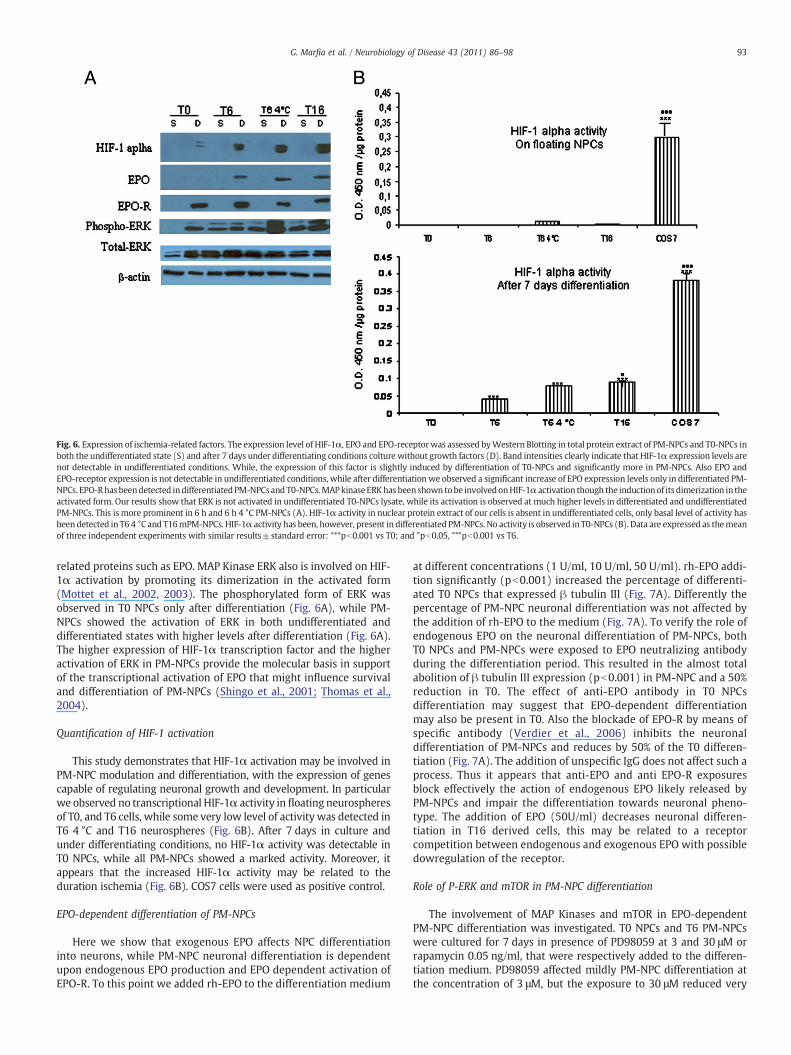
Fig. 6. Expression of ischemia-related factors. The expression level of HIF-1α, EPO and EPO-receptorwas assessed byWestern Blotting in total protein extract of PM-NPCs and T0-NPCs inboth the undifferentiated state (S) and after 7 days under differentiating conditions colture without growth factors (D). Band intensities clearly indicate that HIF-1α expression levels arenot detectable in undifferentiated conditions. While, the expression of this factor is slightly induced by differentiation of T0-NPCs and significantly more in PM-NPCs. Also EPO andEPO-receptor expression is not detectable in undifferentiated conditions, while after differentiation we observed a significant increase of EPO expression levels only in differentiated PM-NPCs. EPO-Rhas beendetected indifferentiatedPM-NPCsand T0-NPCs.MAPkinaseERKhas been shown tobe involvedonHIF-1α activation though the inductionof its dimerization in theactivated form. Our results show that ERK is not activated in undifferentiated T0-NPCs lysate, while its activation is observed at much higher levels in differentiated and undifferentiatedPM-NPCs. This is more prominent in 6 h and 6 h 4 °C PM-NPCs (A). HIF-1α activity in nuclear protein extract of our cells is absent in undifferentiated cells, only basal level of activity hasbeendetected in T6 4 °C and T16mPM-NPCs. HIF-1α activity has been, however, present in differentiated PM-NPCs. No activity is observed in T0-NPCs (B). Data are expressed as themeanof three independent experiments with similar results±standard error: ***pb0.001 vs T0; and °pb0.05, °°°pb0.001 vs T6.
93G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
related proteins such as EPO. MAP Kinase ERK also is involved on HIF-1α activation by promoting its dimerization in the activated form(Mottet et al., 2002, 2003). The phosphorylated form of ERK wasobserved in T0 NPCs only after differentiation (Fig. 6A), while PM-NPCs showed the activation of ERK in both undifferentiated anddifferentiated states with higher levels after differentiation (Fig. 6A).The higher expression of HIF-1α transcription factor and the higheractivation of ERK in PM-NPCs provide the molecular basis in supportof the transcriptional activation of EPO that might influence survivaland differentiation of PM-NPCs (Shingo et al., 2001; Thomas et al.,2004).
Quantification of HIF-1 activation
This study demonstrates that HIF-1α activation may be involved inPM-NPC modulation and differentiation, with the expression of genescapable of regulating neuronal growth and development. In particularwe observed no transcriptional HIF-1α activity in floating neurospheresof T0, and T6 cells, while some very low level of activity was detected inT6 4 °C and T16 neurospheres (Fig. 6B). After 7 days in culture andunder differentiating conditions, no HIF-1α activity was detectable inT0 NPCs, while all PM-NPCs showed a marked activity. Moreover, itappears that the increased HIF-1α activity may be related to theduration ischemia (Fig. 6B). COS7 cells were used as positive control.
EPO-dependent differentiation of PM-NPCs
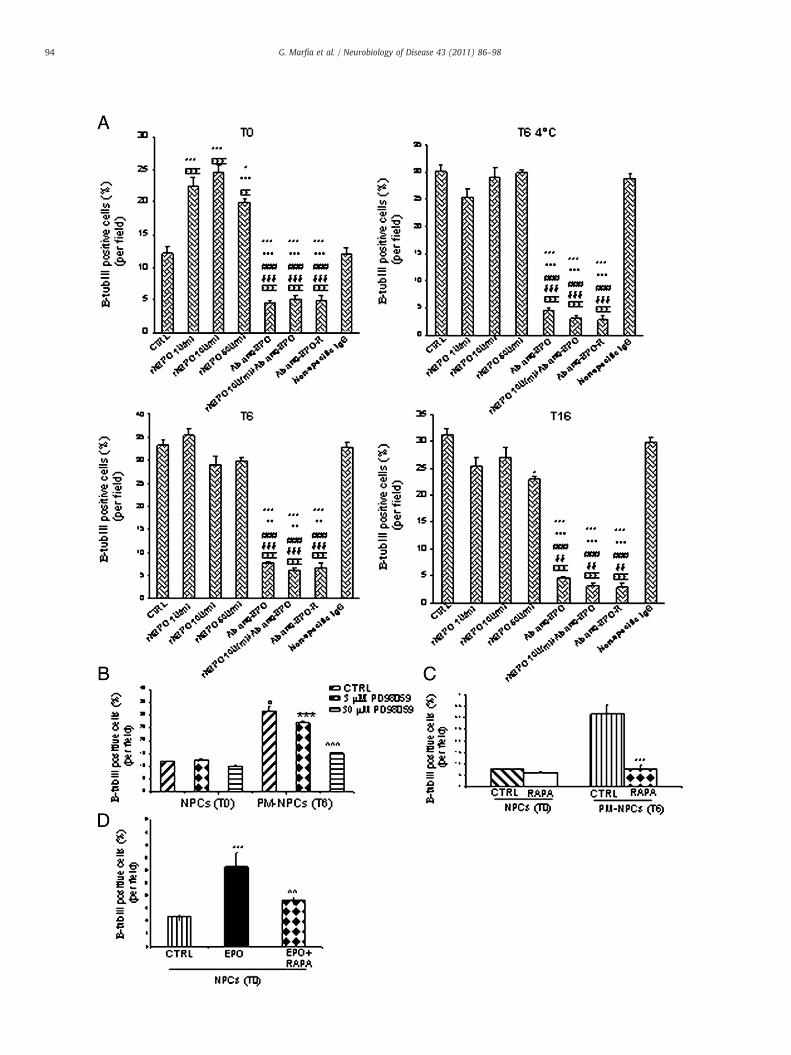
Here we show that exogenous EPO affects NPC differentiationinto neurons, while PM-NPC neuronal differentiation is dependentupon endogenous EPO production and EPO dependent activation ofEPO-R. To this point we added rh-EPO to the differentiation medium
at different concentrations (1 U/ml, 10 U/ml, 50 U/ml). rh-EPO addi-tion significantly (pb0.001) increased the percentage of differenti-ated T0 NPCs that expressed β tubulin III (Fig. 7A). Differently thepercentage of PM-NPC neuronal differentiation was not affected bythe addition of rh-EPO to the medium (Fig. 7A). To verify the role ofendogenous EPO on the neuronal differentiation of PM-NPCs, bothT0 NPCs and PM-NPCs were exposed to EPO neutralizing antibodyduring the differentiation period. This resulted in the almost totalabolition of β tubulin III expression (pb0.001) in PM-NPC and a 50%reduction in T0. The effect of anti-EPO antibody in T0 NPCsdifferentiation may suggest that EPO-dependent differentiationmay also be present in T0. Also the blockade of EPO-R by means ofspecific antibody (Verdier et al., 2006) inhibits the neuronaldifferentiation of PM-NPCs and reduces by 50% of the T0 differen-tiation (Fig. 7A). The addition of unspecific IgG does not affect such aprocess. Thus it appears that anti-EPO and anti EPO-R exposuresblock effectively the action of endogenous EPO likely released byPM-NPCs and impair the differentiation towards neuronal pheno-type. The addition of EPO (50U/ml) decreases neuronal differen-tiation in T16 derived cells, this may be related to a receptorcompetition between endogenous and exogenous EPO with possibledowregulation of the receptor.
Role of P-ERK and mTOR in PM-NPC differentiation
The involvement of MAP Kinases and mTOR in EPO-dependentPM-NPC differentiation was investigated. T0 NPCs and T6 PM-NPCswere cultured for 7 days in presence of PD98059 at 3 and 30 μM orrapamycin 0.05 ng/ml, that were respectively added to the differen-tiation medium. PD98059 affected mildly PM-NPC differentiation atthe concentration of 3 μM, but the exposure to 30 μM reduced very

Fig. 8. Calcium currents. Voltage-dependent currents through Ca-channels have been evoked by means of step potentials from a holding potential of−90 mV, scaled between −50to +50 mV (inset in panel A). Ba2+ (10 mM) was utilized in the extracellular side of the membrane as charge carrier because of its high permeation through high voltage activated(HVA) Ca-channels and its blocking action on many potassium channels. (A) Ca-channel currents were blocked by means of 100 μM Cd2+, the spared currents were subtracted fromthose recovered (R) after washing out the blocker and the results are shown in panel B. The current-to-voltage graph (panel C) shows a pick current at −20 mV. Ca-currents werealmost completely recovered removing the blockers from the external solution. Data are expressed as the mean of three independent experiments with similar results±standarderror: ***pb0.001 vs (control) ; °°°pb0.001 vs rhEPO 10 U/ml; ###pb0.001 vs rhEPO 1U/ml; $$pb0.01 rhEPO 50 U/ml.
95G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
significantly the number of β tubulin III-positive PM-NPCs. T0 wasunaffected (Fig. 7B). PM-NPC neuronal differentiation is, however,extremely dependent upon mTOR function, since rapamycin at doseas low as 0.05 ng/ml obliterated the higher neuronal-differentiationtypical of PM-NPCs (Fig. 7C). This low dose of rapamycin did not affectdifferentiation of T0 NPCs (Fig. 7C). Differently rapamycin reduced T0neuronal differentiation promoted by EPO addition (Fig. 7D), and thatwas totally abolished when rapamycin dosage was increased (0.5 ng/ml, data not shown). These data strongly suggest that EPO-mediatedneuronal differentiation of neural progenitor requires mTOR func-tionality. To verify whether the presence of rapamycin or PD98059could alter cell survival, during the same assays we quantified the
Fig. 7. EPO-dependent differentiation of SVZ-neural progenitors and PM-NPCs into neurons anpresence of different amount of rhEPO (1 U/ml;10 U/ml; 50 U/ml) that has been added to the di3 μg/ml or rhEPO 10 U/ml plus Ab anti-EPO 3 μg/ml or a non specific antibody anti-ED1 3 μg/mpercentage of positive cells. It is shown that the addition of EPO enhances the neuronal diffethe neuronal differentiation of T0-NPCs and all PM-NPCs.Mean±S.E.M.: A*pb0.05, ***Pb0.001$$$pb0.001 vs rhEPO 50U/ml; ££pb0.01, £££pb0.001 vs Ab anti-ED1. (B- D). (B) PM-NPCs and N7 days. Differentiation towards the neuronal phenotype was investigated by immunostaining fodifferentiation in a dose-dependent manner. (C) PM-NPCs (T6) and NPCs (T0) were differDifferentiationwas investigated as indicated for panel A. The addition of Rapamycin prevents thdifferentiated in absence (CTRL) or presence of rhEPO (EPO 10U/ml), with orwithout out RapamA and B. The exposure to rapamycin affects and reduces the EPO-mediated enhanced neuronasimilar results±standard error; B° pb0,05; ^^^pb0,001; ****pb0,001; C***pb0.001 T6 RAPA
total numbers of cells. After 7 days of differentiation we observed nosignificative change in the total number of cells in cultures treatedwith rapamycin (0.05 ng/ml and 0.5 ng/ml) or PD (3 and 30 μM) vscontrol (data not shown).
Calcium channel currents
The family of voltage-gated calcium channels serves as the keytransducers of cell-surface membrane-potential changes into localintracellular calcium transients that initiate many different physio-logical events. High-voltage-activated (HVA) calcium currents (ICa)have been found in NPCs (Arnhold et al., 2000; Catterall et al., 2005;
d MAP-kinase ERK and mTOR involvement. (A) PM-NPCs and NPCs were differentiated infferentiationmedium; in some caseswe added also the Ab anti-EPO 3 μg/ml or anti-EPO-Rl. Differentiation was investigated by the staining for β-tubulin III and was reported as arentiation of T0-NPCs only, while the anti EPO exposure suppresses, almost completelyvs CTRL; ###pb0.001 vs rhEPO1 U/ml; °°pb0.01, °°° pb0.001 vs rhEPO 10 U/ml; $$pb0.01,PCs were differentiated in absence (CTRL) or presence of PD98059 (3 μM and 30 μM) forr β-tubulin III andwas reported as a percentage of positive cells. PD98059 affects PM-NPCentiated in absence (CTRL) or presence of Rapamycin (RAPA; 0.05 ng/ml) for 7 days.e higher neuronal differentiation of PM-NPCs,while T0 are unaffected. (D) NPCs (T0)wereycin (RAPA; 0.05 ng/ml) for 7 days. Differentiationwas investigated as indicated for panell differentiation. Data are expressed as the mean of three independent experiments withvs T6 CTRL; and D ***pb0.001 EPO vs CTRL, ^^pb0.01 EPO+RAPA vs CTRL.

96 G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
D'Ascenzo et al., 2006). In the present study, ICa has been studied bymeans of whole-cell patch-clamp technique at different stages ofPM-NPC maturation. The recordings were carried out between 5 and45 days after induction of differentiation. ICa has been identified bymeans of biophysical and pharmacological criteria. Barium ions(10 mM) were utilized as charge carriers to take advantage of itsblocking effect on potassium channels, and for its grater permeationof HVA Ca-channels. HVA ICa was detected in all of the cells tested(n=64). The maximum conductance (Gmax) was 0.153±0.02nS∙pF-1. Fig. 8 from A to C shows Ba2+ currents through Ca-channelselicited by means of steps of increasing potential from −50 mV to+50 mV. Cadmium ions (100 μM) added to the external solutionblocked almost completely the Ba2+ currents (Fig. 8). Subtractingthe current traces in presence of Cd2+ from those obtained afterwashing out the blocker (R – Cd2+; Fig. 8) it was possible to isolatethe current component blocked by Cd2+ and panel C of the sameFigure shows the current to voltage relationship of this component.On the whole, the characteristics of these currents are compatiblewith HVA calcium channel currents as far as activation and deacti-vation kinetics, voltage to peak of the I–V relationship, and sensiti-vity to Cd2+ are concerned.
Interestingly enough, the addition of anti-EPO antibodies to the PM-NPC cultures immediately after plating and during the differentiationtime, completely abolished or prevented the appearance of Ca-currents(n=10 out of 10 cells tested). The same results were obtained utilizinganti EPO-R antibodies (n=5 out of 5 cells recorded; data not shown).
Discussion
Cell transplantation therapy based on stem cells is a majorcomponent of regenerative medicine. An issue limiting the clinicalapplication and effectiveness of stem cell therapies is the high extentof transplanted cell death. Oxygen tension is an important regulator ofcell metabolism, survival, and fate, although extreme fluctuations inoxygen availability are a major cause of cell death. In this presentinvestigation we show the isolation of neuronal precursors under nonphysiological conditions, at about 6 h post-mortem, that afterdifferentiation yield significantly higher proportion of both β-tubulinIII and MAP-2 labeling in comparison with neural precursors isolatedimmediately after the animal sacrifice. These post-mortem acquiredneural progenitors cells were, thus, named PM-NPCs.
Here we report, in accordance with other observations (Francisand Wei, 2010; Santilli et al., 2010), that the exposure to non phy-siological conditions, such as the ischemic stimulus, drasticallyenhances neuronal and oligodendroglial differentiation. Hypoxic orischemic preconditioning was initially discovered as an endogenousprotective mechanism in animals, and later confirmed in virtuallyall types of cultured cells and organs (Kirino, 2002; Pong, 2004).Hypoxic/ischemic preconditioning usually produces two phases ofcytoprotection: Phase I, an acute protection that appears early andlasts for a few hours after insult and Phase II, a delayed but strongerprotection which emerges many hours after hypoxia and can last fordays or even weeks (Semenza, 2001; Kirino, 2002; Pong, 2004). Thedelayed phase of hypoxic/ischemic preconditioning is mediated bysynthesis of mRNAs and proteins, including the transcription factorhypoxia-inducible factor 1-α (HIF1-α) and HIF target genes, such aserythropoietin (EPO). The findings here reported, show that aprolonged ischemic insult leads to neural precursors that can beextracted and differentiate mostly in neurons after in vitro differen-tiation. This feature is likely driven by higher levels of HIF-1αactivation and EPO expression. The finding that supplementation ofboth anti-EPO and EPO-R antibodies to the differentiating mediumleads to the suppression of typical expression of neuronal markerssuch as β-tubulin III and Ca2+ currents supports such an hypothesis.Similar features of hypoxia impact on cell fate decision have recentlybeen shown both by Prado-Lopez and colleagues (2010) trough the
investigation the effects of low O2 on gene expression in humanembryonic stem cells and by Francis and Wei (2010) through theinvestigation of the effects of low O2 on neuronal differentiation ofhuman embryonic stem cells. Studies have suggested a role for EPO inthe proliferation and differentiation of neuronal progenitors and stemcells, while others reported that EPOmay affect the extent of neuronalproduction through the enhancement of forebrain NPCs differen-tiation (Shingo et al., 2001; Park et al., 2006). For example, theadministration of EPO into the lateral ventricle increased the numberof newly generated cells migrating to the olfactory bulb and olfactorybulb neurons (Bernaudin et al., 2002; Ruscher et al., 2002), an effectthat could be mimicked by subjecting the animal to hypoxia (Kirino,2002; Pong, 2004). Furthermore, low levels of oxygen enhancedproliferation, survival and differentiation of CNS neural precursors(Wang et al., 2004), and post-ischemia treatment with EPO enhancedneurogenesis and improved neurological function (Moreno-Manzanoet al., 2009, 2010). The inductive effect of ischemia on theproliferation of neural progenitors has also been shown in agedadult human forebrain, where an elevated number of Ki-67-positivecells were formed in the SVZ ipsilateral to the ischemic zone (Martí-Fàbregas et al., 2010). Differently, in our study we observed that post-mortem ischemia triggered the generation of EPO-dependent neuralstem cells, where EPO enhances differentiation rather than prolifer-ation. Thus the above observations and the isolation of the EPOproducing PM-NPCs with a higher tendency to differentiate intoneuronal phenotypes support the potential role for endogenous EPOin the plasticity and repair of the central nervous system.
Moreover higher HIF-1α activation and ERK phosphorylation inundifferentiated PM-NPCs do not affect their in vitro proliferationability that is comparable to T0 NPCs. The differentiation process,however, clearlymarks the difference between these twokinds of SVZ-derived neural progenitors. The higher expression of neuronalphenotypes requires functionality of MAPK (Schlessinger, 2000) andmTOR complex (Park et al., 2008), since the exposure to eitherPD98059 or rapamycin, counteracts the induction of β tubulin IIIexpression and neurite formation. PM-NPCs T6 are particularlysensitive to mTOR activity, being the rapamycin dose 100 fold lowerthan the average dose normally used to inhibit mTOR in TSC2 cells(Carelli et al., 2007; Lesma et al., 2005, 2008). mTOR function is alsorequired for the differentiation into olygodendrocytes, although thereported dosage of rapamycinwas 200 times higher (Tyler et al., 2009;Park et al., 2008). The higher ERK activation observed in undifferen-tiated PM-NPCs may have a role in their differentiation process, sincethe exposure to the MAPK inhibitor PD98059 affects significantly theextent of neuronal differentiation. This study shows that the enhancedneuronal PM-NPC differentiation requires EPO and the functionality ofmTOR and ERK, but other mechanisms may also be involved. Inlowered O2 levels HIF-1α interacts with activated Notch to repressterminal differentiation in neural precursors, while increasing O2
levels prevents this direct HIF-1α/Notch interaction, thereby allowingprecursor cells to differentiate decreasing gliogenesis and stimulatingPAX6 gene expression (Corbin et al., 2008; Gustafsson et al., 2005).
Radial glia serves as a guide for neurons, and has, also, propertiesof neural progenitors during neurogenesis (Goetz, 1998; Misson et al.,1991). It is known that a subset of radial glia may give rise to sub-ventricular zone astrocytes that function as adult neural stem cell(Aguado et al., 2006; Merkle et al., 2004). We observed that theexpression of PAX6, by differentiated PM-NPCs, increases with theduration of donor ischemia, while RC2 decreases. PAX6 is one of the keyfactors inCNSpatterningandplays a role in thedifferentiation of corticalradial glia and in the specification of progenitor cells into neurons(Simpson and Price, 2002). Inhibition of PAX6 expression impairs theformation of neural progenitors (Tuoc et al., 2009). RC2 is expressedby asubpopulation of neural progenitors addressed in the astroglial lineageand evolving into GFAP expressing cells (Mottet et al., 2002, 2003). Thusthe differential expression of radial glia markers, with higher PAX 6 and

97G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
lower RC2, is in support of the high neurogenic ability of PM-NPCs and,also, suggests a close relationship to a radial glia lineage.
T6 PM-NPCs, derived from sacrificed mouse kept at 4 °C, differ fromnormal T6 for their ability to yield a significantly higher percentage ofoligodendrocytes and precursors during differentiation. Thus it wouldbe interesting to assess their reparatory ability in animal models ofdemyelinating disease.
PM-NPC differentiation yields a significantly higher percentage ofneurons which are mostly immunopositive for ChAT, TH, glutamate,and at much lower percentage for GABA. Moreover T6 4 °C show ahigher propension to differentiate in TH positive neurons. The clearindication of their neuronal differentiation comes from the classicalelectrophysiological properties. After induction of differentiation, PM-NPCs show the delayed-rectifier K+ and the fast inactivating Na+
currents (manuscript in preparation), and the Ca2+ currents. Theselatter currents show the biophysical and pharmacological character-istics of HVA currents. The peak of the I–V relationship at −20 mV(Fig. 8), considering the presence of 10 mM Ba2+ in the externalsolution that should exert a screening effect on the fixed negativecharges on the cell membrane shifting the voltage-dependentcharacteristics rightward on the voltage axis, seem to indicate thepresence of the low voltage activating components of these channels,i.e. Cav1.3–4 (Lipscombe et al., 2004). Interestingly enough, in addi-tion to the suppression of all neuronal markers, the inhibition of EPOpathway by means of anti-EPO and EPO-R antibodies completelyabolishes the Calcium currents. This could be explained both by theimmaturity of neurons present in the plate of recording and by thesignificant decrease of their number.
In summary, we have demonstrated the isolation of adult PM-NPCsand characterized their neuronal differentiation ability. Furthermore,we show that brief exposures of neural precursors to an ischemicstimuli can induce cell preconditioning and attenuate cell deathinduced by injurious events typical to ischemic stress. Preconditioninghad no detrimental effect on neuronal differentiation or proliferation.HIF stabilization and activation of several key neuroprotective genesmost likely contribute to the cytoprotection. It is conceivable thattransplanting preconditioned cells, such as PM-NPCs, may replace someof the lost neurons in lesioned CNS and improve recovery of function.This is supported by the outcome of their application to mice aftertraumatic injury to the cord, where PM-NPCs survive, form neurites andpromote recovery of function (manuscript in preparation).
Supplementarymaterials related to this article can be found onlineat doi:10.1016/j.nbd.2011.02.004.
Founding
This work was supported in part by a grant from FondazioneMontecatone and PRIN 2005 to Alfredo Gorio.
Acknowledgment
We acknowledge the continuous scientific support by DrsGuglielmo Brayda, Fabio Calbucci, Roberto Piperno, Maria Taricco,Giulio Cicognani of the scientific advisory board of the FondazioneMontecatone. We are grateful to Raffaella Adami, Davide Merli,Maria Vittoria Traversa, Ambra Vittoria Gualeni, Melissa Sorosina,Francesco Bolzoni and Francesco Naselli for the outstanding supportand suggestions.
References
Adelman, D.M., Simon, M.C., 2002. Hypoxic gene regulation in differentiating ES cells.Methods Mol. Biol. 185, 55–62.
Aguado, T., Palazuelos, J., Monory, K., Stella, N., Cravatt, B., et al., 2006. Theendocannabinoid system promotes astroglial differentiation by acting on neuralprogenitor cells. J. Neurosci. 26, 1551–1561.
Arnhold, S., Andressen, C., Angelov, D.N., Vajna, R., Volsen, S.G., et al., 2000. Embryonicstem-cell derived neurones express a maturation dependent pattern of voltage-gated calcium channels and calcium-binding proteins. Int. J. Dev. Neurosci. 18,201–212.
Baer, K., Eriksson, P.S., Faull, R.L., Rees, M.I., Curtis, M.A., 2007. Sox-2 is expressed by glialand progenitor cells and Pax-6 is expressed by neuroblasts in the humansubventricular zone. Exp. Neurol. 204, 828–831.
Bernaudin, M., Nedelec, A.S., Divoux, D., MacKenzie, E.T., Petit, E., et al., 2002.Normobaric hypoxia induces tolerance to focal permanent cerebral ischemia inassociation with an increased expression of hypoxia-inducible factor-1 and itstarget genes, erythropoietin and VEGF, in the adult mouse brain. J. Cereb. BloodFlow Metab. 22, 393–403.
Berra, E., Milanini, J., Richard, D.E., Le Gall, M., Vinals, F., et al., 2000. Signalingangiogenesis via p42/p44 MAP kinase and hypoxia. Biochem. Pharmacol. 60,1171–1178.
Bottai, D., Madaschi, L., Di Giulio, A.M., Gorio, A., 2008. Viability-dependent promotingaction of adult neural precursors in spinal cord injury. Mol. Med. 14, 634–644.
Bottai, D., Cigognini, D., Madaschi, L., Adami, R., Nicora, E., Menarini, M., Di Giulio, A.M., Gorio,A., 2010. Embryonic stem cells promote motor recovery and affect inflammatory cellinfiltration in spinal cord injured mice. Exp. Neurol. 223 (2), 452–463 2010 Jun.
Bruick, R.K., 2003. Oxygen sensing in the hypoxic response pathway: regulation of thehypoxia-inducible transcription factor. Genes Dev. 17, 2614–2623.
Carelli, S., Lesma, E., Paratore, S., Grande, V., Zadra, G., et al., 2007. Survivin expression intuberous sclerosis complex cells. Mol. Med. 13, 166–177.
Catterall, W.A., Perez-Reyes, E., Snutch, T.P., Striessnig, J., 2005. International Union ofPharmacology. XLVIII. Nomenclature and structure-function relationships ofvoltage-gated calcium channels. Pharmacol. Rev. 57, 411–418.
Chen, H.L., Pistollato, F., Hoeppner, D.J., Ni, H.T., McKay, R.D., et al., 2007. Oxygen tensionregulates survival and fate of mouse central nervous system precursors at multiplelevels. Stem Cells 25, 2291–2301.
Cino, I., Formenti, A., 2008. Spermine biphasically affects N-type calcium channel currentsin adult dorsal root ganglion neurons of the rat. Biochim. Biophys. Acta 1778,2437–2443.
Corbin, J.G., Gaiano, N., Juliano, S.L., Poluch, S., Stancik, E., et al., 2008. Regulation ofneural progenitor cell development in the nervous system. J. Neurochem. 106,2272–2287.
D'Ascenzo,M., Piacentini, R., Casalbore, P., Budoni, M., Pallini, R., et al., 2006. Role of L-typeCa2+ channels in neural stem/progenitor cell differentiation. Eur. J. Neurosci. 23,935–942.
Doetsch, F., 2003. The glial identity of neural stem cells. Nat. Neurosci. 6, 1127–1134.Francis, K.R., Wei, L., 2010. Human embryonic stem cell neural differentiation and
enhanced cell survival promoted by hypoxic preconditioning. Cell Death Dis. 1, e22.Fricker-Gates, R.A., 2006. Radial glia: a changing role in the central nervous system.
NeuroReport 17, 1081–1084.Goetz, M., 1998. How are neurons specified: master or positional control? Trends
Neurosci. 21, 135–136.Gritti, A., Parati, E.A., Cova, L., Frolichsthal, P., Galli, R., et al., 1996. Multipotential stem
cells from the adult mouse brain proliferate and self-renew in response to basicfibroblast growth factor. J. Neurosci. 16, 1091–1100.
Gritti, A., Frölichsthal-Schoeller, P., Galli, R., Parati, E.A., Cova, L., et al., 1999. Epidermaland fibroblast growth factors behave as mitogenic regulators for a singlemultipotent stem cell-like population from the subventricular region of the adultmouse forebrain. J. Neurosci. 19, 3287–3297.
Gustafsson, M.V., Zheng, X., Pereira, T., Gradin, K., Jin, S., et al., 2005. Hypoxia requiresnotch signaling to maintain the undifferentiated cell state. Dev. Cell 9, 617–628.
Hoge, R.D., Pike, G.B., 2001. Oxidative metabolism and the detection of neuronalactivation via imaging. Chem. Neuroanat. 22, 43–52.
Kirino, T., 2002. Ischemic tolerance. J. Cereb. Blood Flow Metab. 22, 1283–1296.Lesma, E., Grande, V., Carelli, S., Brancaccio, D., Canevini, M.P., et al., 2005. Isolation and
growth of smoothmuscle-like cells derived fromtuberous sclerosis complex-2humanrenal angiomyolipoma: epidermal growth factor is the required growth factor. Am. J.Pathol. 167, 1093–1103.
Lesma, E., Grande, V., Ancona, S., Carelli, S., Di Giulio, A.M., et al., 2008. Anti-EGFRantibody efficiently and specifically inhibits human TSC2−/− smooth muscle cellproliferation. Possible treatment options for TSC and LAM. PLoS ONE 3, e3558.
Lipscombe, D., Helton, T.D., Xu, W., 2004. L-type calcium channels: the low down. J.Neurophysiol. 92, 2633–2641.
Martí-Fàbregas, J., Romaguera-Ros, M., Gómez-Pinedo, U., Martínez-Ramírez, S., Jiménez-Xarrié, E., Marín, R., Martí-Vilalta, J.L., García-Verdugo, J.M., 2010. Proliferation in thehuman ipsilateral subventricular zone after ischemic stroke. Neurology 74 (5),357–365.
Merkle, F.T., Tramontin, A.D., Garcia-Verdugo, J.M., Alvarez-Buylla, A., 2004. Radial gliagive rise to adult neural stem cells in the subventricular zone. Proc. Natl Acad. Sci.USA 101, 17528–17532.
Minet, E., Arnould, T., Michel, G., Roland, I., Mottet, D., et al., 2000. ERK activation uponhypoxia: involvement in HIF-1 activation. FEBS Lett. 468, 53–58.
Misson, J.P., Austin, C.P., Takahashi, T., Cepko, C.L., Caviness Jr., V.S., 1991. The alignmentof migrating neural cells in relation to the murine neopallial radial glial fibersystem. Cereb. Cortex 1, 221–229.
Molcanyi, M., Riess, P., Bentz, K., Maegele, M., Hescheler, J., et al., 2007. Trauma-associated inflammatory response impairs embryonic stem cell survival andintegration after implantation into injured rat brain. J. Neurotrauma 24, 625–637.
Moreno-Manzano, V., Rodríguez-Jiménez, F.J., García-Roselló, M., Laínez, S., Erceg, S.,Calvo, M.T., Ronaghi, M., Lloret, M., Planells-Cases, R., Sánchez-Puelles, J.M.,Stojkovic, M., 2009. Activated spinal cord ependymal stem cells rescue neurologicalfunction. Stem Cells 27 (3), 733–743 Mar.

98 G. Marfia et al. / Neurobiology of Disease 43 (2011) 86–98
Moreno-Manzano, V., Rodríguez-Jiménez, F.J., Aceña-Bonilla, J.L., Fustero-Lardíes, S.,Erceg, S., Dopazo, J., Montaner, D., Stojkovic, M., Sánchez-Puelles, J.M., 2010.FM19G11, a new hypoxia-inducible factor (HIF) modulator, affects stem celldifferentiation status. J. Biol. Chem. 285 (2), 1333–1342 Jan 8.
Mottet, D., Michel, G., Renard, P., Ninane, N., Raes, M., et al., 2002. ERK and calcium inactivation of HIF-1. Ann. NY Acad. Sci. 973, 448–453.
Mottet, D., Michel, G., Renard, P., Ninane, N., Raes, M., et al., 2003. Role of ERK andcalcium in the hypoxia-induced activation of HIF-1. J. Cell. Physiol. 194, 30–44.
Park,M.H., Lee, S.M., Lee, J.W., Son, D.J., Moon, D.C., Yoon, D.Y., Hong, J.T., 2006. ERK-mediatedproduction of neurotrophic factors by astrocytes promotes neuronal stem celldifferentiation by erythropoietin. Biochem. Biophys. Res. Commun. 339, 1021–1028.
Park, K.K., Liu, K., Hu, Y., Smith, P.D., Wang, C., et al., 2008. Promoting axon regenerationin the adult CNS by modulation of the PTEN/mTOR pathway. Science 322, 963–966.
Parnavelas, J.G., Nadarajah, B., 2001. Radial glial cells: are they really glia? Neuron 31,881–884.
Pong, K., 2004. Ischaemic preconditioning: therapeutic implications for stroke? ExpertOpin. Ther. Targets 8, 125–139.
Popovich, P.G., van Rooijen, N., Hickey, W.F., Preidis, G., McGaughy, V., 2003.Hematogenous macrophages express CD8 and distribute to regions of lesioncavitation after spinal cord injury. Exp. Neurol. 182, 275–287.
Prado-Lopez, S., Conesa, A., Armiñán, A., Martínez-Losa, M., Escobedo-Lucea, C., Gandia,C., Tarazona, S., Melguizo, D., Blesa, D., Montaner, D., Sanz-González, S., Sepúlveda,P., Götz, S., O'Connor, J.E., Moreno, R., Dopazo, J., Burks, D.J., Stojkovic, M., 2010.Hypoxia promotes efficient differentiation of human embryonic stem cells tofunctional endothelium Mar 31 Stem Cells 28 (3), 407–418.
Ramírez-Bergeron, D.L., Simon, M.C., 2001. Hypoxia-inducible factor and the develop-ment of stem cells of the cardiovascular system. Stem Cells 19, 279–286.
Richard, D.E., Berra, E., Gothie, E., Roux, D., Pouyssegur, J., 1999. p42/p44 mitogen-activated protein kinases phosphorylates hypoxia inducible factor 1 α (HIF-1α)and enhance the transcriptional activity of HIF-1. J. Biol. Chem. 274, 32631–32637.
Ronaghi, M., Erceg, S., Moreno-Manzano, V., Stojkovic, M., 2010. Challenges of stem celltherapy for spinal cord injury: human embryonic stem cells, endogenous neuralstem cells, or induced pluripotent stem cells? Stem Cells 28 (1), 93–99.
Ruscher, K., Freyer, D., Karsch, M., Isaev, N., Megow, D., et al., 2002. Erythropoietin is aparacrine mediator of ischemic tolerance in the brain: evidence from an in vitromodel. J. Neurosci. 22, 10291–10301.
Santilli, G., Lamorte, G., Carlessi, L., Ferrari, D., Rota Nodari, L., Binda, E., Delia, D.,Vescovi, A.L., De Filippis, L., 2010. Mild hypoxia enhances proliferation andmultipotency of human neural stem cells. PLoS ONE 5 (1), e8575.
Schlessinger, J., 2000. Cell signaling by receptor tyrosine kinases. Cell 103, 211–225.Semenza, G.L., 2000. Surviving ischemia: adaptive responses mediated by hypoxia-
inducible factor 1. J. Clin. Invest. 106, 809–812.Semenza, G.L., 2001. HIF-1 and mechanisms of hypoxia sensing. Curr. Opin. Cell Biol. 13,
167–171.Shingo, T., Sorokan, S.T., Shimazaki, T.,Weiss, S., 2001. Erythropoietin regulates the in vitro
and in vivo production of neuronal progenitors by mammalian forebrain neural stemcells. J. Neurosci. 21, 9733–9743.
Simpson, T.I., Price, D.J., 2002. Pax-6: a pleiotropic player in development. Bioessays 24,1041–1051.
Studer, L., Csete, M., Lee, S.H., Kabbani, N., Walikonis, J., et al., 2000. Enhancedproliferation, survival, and dopaminergic differentiation of CNS precursors inlowered oxygen. J. Neurosci. 20, 7377–7383.
Thomas, D.D., Espey, M.G., Ridnour, L.A., Hofseth, L.J., Mancardi, D., et al., 2004. Hypoxicinducible factor 1α, extracellular signal-regulated kinase, and p53 are regulated bydistinct threshold concentrations of nitric oxide. Proc. Natl Acad. Sci. USA 101,8894–8899.
Tuoc, T.C., Radyushkin, K., Tonchev, A.B., Piñon, M.C., Ashery-Padan, R., et al., 2009.Selective cortical layering abnormalities and behavioural deficits in cortex-specificPax6 knock-out mice. J. Neurosci. 29, 8335–8349.
Tyler, W.A., Gangoli, N., Gokina, P., Kim, H.A., Covey, M., et al., 2009. Activation of themammalian target of rapamycin (mTOR) is essential for oligodendrocytedifferentiation. J. Neurosci. 29, 6367–6378.
Verdier, F., Gomez, S., Lacombe, C., Mayeux, P., 2006. Selected anti-Epo receptorantibodies predict Epo receptor expression. Blood 107, 1892–1895.
Wang, L., Zhang, Z., Wang, Y., Zhang, R., Chopp, M., 2004. Treatment of stroke witherythropoietin enhances neurogenesis and angiogenesis and improves neurolog-ical function in rats. Stroke 35, 1732–1737.
Wu, L.Y., Wang, Y., Jin, B., Zhao, T., Wu, H.T., et al., 2008. The role of hypoxia in thedifferentiation of P19 embryonal carcinoma cells into dopaminergic neurons.Neurochem. Res. 33, 2118–2125.
Zhu, L.L., Zhao, T., Li, H.S., Zhao, H.,Wu, L.Y., et al., 2005. Neurogenesis in the adult rat brainafter intermittent hypoxia. Brain Res. 1055, 1–6.