Embed Size (px)
Citation preview

Geobios xxx (2015) xxx–xxx
G Model
GEOBIO-701; No. of Pages 46
Original article
Ammonites of the subfamily Mayaitinae Spath, 1928 from theOxfordian of Kachchh, Western India§
Matthias Alberti a,*, Dhirendra K. Pandey b, Manja Hethke c, Franz T. Fursich c
a Institut fur Geowissenschaften, Christian-Albrechts-Universitat zu Kiel, Ludewig-Meyn-Straße 10, 24118 Kiel, Germanyb Department of Geology, University of Rajasthan, 302004 Jaipur, Indiac GeoZentrum Nordbayern, Fachgruppe PalaoUmwelt, Friedrich-Alexander-Universitat Erlangen-Nurnberg, Loewenichstraße 28, 91054 Erlangen, Germany
A R T I C L E I N F O
Article history:
Received 7 August 2014
Accepted 3 February 2015
Available online xxx
Keywords:
Ammonoids
Taxonomy
Biostratigaphy
Upper Jurassic
Kachchh Basin
India
A B S T R A C T
A collection of 100 ammonites of the subfamily Mayaitinae Spath, 1928 (Sphaeroceratidae,
Stephanocerataceae) from the Oxfordian (Upper Jurassic) of the Kachchh Basin, Western India,
represents 19 morphospecies. Their study confirms most of the species originally described from the
Kachchh Basin more than 80 years ago, but it strongly extends the knowledge on their diagnostic
features. Most species can be readily differentiated by the shape and ontogenetic development of their
whorl section as illustrated by a Fourier shape analysis. The Oxfordian subfamily Mayaitinae is largely
endemic to the Malagasy Gulf and adjacent regions at the southern margin of the Tethys. Finds of isolated
specimens from other areas of the northern Tethys or southern Pacific are suggested to be drifted shells or
misinterpretations of homeomorphic taxa. Although commonly found in the Kachchh Basin, the group
has received little attention through the last decades due to their seemingly limited use in
biostratigraphy. In the present study the biostratigraphic potential of a series of taxa is discussed;
e.g., Epimayaites subtumidus seems to be restricted to the Stenocycloides Subzone (Bifurcatus Zone) of the
Upper Oxfordian.
� 2015 Published by Elsevier Masson SAS.
Available online at
ScienceDirectwww.sciencedirect.com
1. Introduction
The Jurassic succession of the Kachchh Basin in Western Indiahas been studied for more than a century, beginning with surveysof Sykes (1834), Grant (1837), Blanford (1869), and Wynne(1872). Subsequently, scientists started to examine and describethe rich and well preserved fossil content of the rocks, especially ofthe Jurassic part of the succession. Taxonomic monographs werepublished on different fossil groups, but particularly the cepha-lopods were studied in detail due to their diversity, abundance, andmost importantly their significance for stratigraphic correlations.Particularly outstanding are the works of Waagen (1873-1875) andSpath (1927-1933), which are still used today as a basis for theidentification of cephalopods from the basin. With continuation ofgeological and palaeontological research in the Kachchh Basin thelithostratigraphic framework and ammonite biostratigraphy arenow much better understood (Fursich et al., 2013). Four recentfield surveys in the Kachchh Basin, however, which concentratedmainly on the Oxfordian part of the succession, yielded more than
§ Corresponding editor: Stephane Reboulet.
* Corresponding author.
E-mail address: [email protected] (M. Alberti).
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
http://dx.doi.org/10.1016/j.geobios.2015.02.005
0016-6995/� 2015 Published by Elsevier Masson SAS.
800 ammonoids and showed that the fossil record of this group isstill not fully known. In addition, a modern and comprehensivetaxonomic study of the Jurassic ammonite fauna based on materialcollected with a high stratigraphic resolution is not yet available.The present study is one in a series to enlarge our knowledge of theUpper Jurassic cephalopods of the Kachchh Basin (see Alberti et al.,2011; Pandey et al., 2012a, 2013a, 2013b, 2015).
2. Geological overview
The Kachchh Basin is situated in Western India, wedgedbetween the Arabian Sea and the border to Pakistan (Fig. 1). Thebasin formed during the early break-up of Gondwana with riftingbetween India and Africa from the Late Triassic onwards (Biswas,1982, 1991). After an initial phase of terrestrial sedimentation, thesea reached the area in the Jurassic. Ammonites of the genusLeptosphinctes Buckman, 1920 point to marine conditions since theBajocian (Singh et al., 1982), but the first transgression probablyoccurred already in the Early Jurassic (Pandey and Fursich, 2001;Fursich et al., 2004a; Rai and Jain, 2013). Since then, marineconditions dominated in the small, fault-bordered basin until mid-Cretaceous times (Fursich, 1998).
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 1. Geological sketch map of the Kachchh Basin showing the sample localities.
Modified after Patel et al. (2012) and Fursich et al. (2013).
M. Alberti et al. / Geobios xxx (2015) xxx–xxx2
G Model
GEOBIO-701; No. of Pages 46
Traditionally, localities with Jurassic outcrops in the KachchhBasin have been divided into three regions: the Kachchh Mainland,the Wagad Uplift, and the Island Belt (Fig. 1). The KachchhMainland occupies the central part of the basin and contains mostof the well-known sections. While the southern part of theKachchh Mainland is covered by the volcanics of the Deccan traps,the northern part adjacent to the Great Rann of Kachchh ischaracterized by an E-W-oriented chain of anticlinal or domalstructures offering outcrops of Jurassic rocks ranging from the?Bajocian to the Tithonian (Pandey and Fursich, 1998; Fursichet al., 2004a, 2004b; Krishna et al., 2009a). Furthermore, a chain ofoutcrops south of the district capital Bhuj displays Jurassic stratauplifted along the Katrol Hill Fault (i.e., the Charwar Hills; Biswas,1993). The Oxfordian succession of the Kachchh Mainland ischaracterized by strong condensation (commonly only a fewmetres in thickness) and sedimentary gaps. The examined intervalstarts within the top of the Upper Callovian Gypsiferous Shalemember of the Chari Formation (Fig. 2). This unit consists mainly ofbioturbated, argillaceous silt with several levels of small ferrugi-nous concretions and abundant secondary gypsum. The Gypsifer-ous Shale member is coarsening upwards into the fine-grainedsandstone beds of the Dhosa Sandstone member, which ischaracterized by occasional cross-bedding indicating an elevated
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
energy level due to a lower water depth. The boundary betweenboth members seems to be diachronous. Based on peltoceratids,the basal part of the Dhosa Sandstone member is Late Callovian inage in the east (e.g., at the village Lodai) and Early Oxfordian in thewest (e.g., at Jumara Dome; John H. Callomon, pers. comm. 2000;Pandey et al., 2015). The occurrence of allochthonous, ferruginousooids marks the boundary to the Dhosa Oolite member, whose top,the Dhosa Conglomerate Bed, serves as an important markerhorizon throughout the Kachchh Mainland. This strongly con-densed unit has received considerable attention due to an intricatemixture of different lithologies and its complex genetic history(Singh, 1989; Fursich et al., 1992; Alberti et al., 2013a). Althoughbeing mostly less than 1 m thick, the Dhosa Conglomerate Bedrepresents most parts of the Oxfordian. It contains largeconcretionary slabs with a Lower Oxfordian ammonite faunafloating in a fine-grained micritic matrix with reworked Lower andMiddle Oxfordian ammonite taxa (e.g., Alberti et al., 2011; Pandeyet al., 2012a). Additionally, at a few outcrops of the Habo Dome(e.g., 3 km east of the temple Rudra Mata) the conglomeraticmatrix fills crevices cutting down into thin layers of oldersandstone. After a long depositional gap encompassing parts ofthe Late Oxfordian and Early Kimmeridgian, sedimentationresumed in the late Early Kimmeridgian with the sandstones of
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 2. Lithostratigraphic framework of the Oxfordian of the Kachchh Basin. The dashed lines denote subunits of the various members.
Modified after Pandey et al. (2013b) and Fursich et al. (2013).
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 3
G Model
GEOBIO-701; No. of Pages 46
the Katrol Formation (Krishna and Pathak, 1993; Krishna et al.,1996, 2000; Pandey et al., 2013b).
In contrast to the condensed Oxfordian succession of theKachchh Mainland, the same interval is represented by severalhundred metres of sediments in the Wagad Uplift. The overallsiliciclastic sediments are alternatively dominated by thin-beddedsilty shales or more massive, cross-bedded sandstones (Fig. 2).Most parts of the succession are devoid of macrofossils andtherefore received less attention. Major concentrations of ammo-nites occur only in the Middle to lower Upper Oxfordian KanthkotAmmonite Beds, as described in a series of studies during the lastfew years (e.g., Krishna et al., 1996, 1998, 2009a, 2009b, 2009c; Royet al., 2012; Pandey et al., 2012a).
The Island Belt consists of a chain of several smaller upliftedareas, which are completely surrounded by the salt marshes ofthe Great Rann of Kachchh. These areas become true islands onlyduring monsoonal floods, but even then they are well connectedto the rest of Kachchh by roads built on dams elevated above themarshes. While most outcrops of the Island Belt display olderstrata, Oxfordian rocks are exposed on Khadir Island as well ason some smaller islets towards the southwest (i.e., Kakindia Betand Gangta Bet; Fig. 1). The predominantly siliciclastic succes-sion of the Khadir Formation at the southernmost tip of KhadirIsland contains a rich fossil record with ammonites beingespecially abundant in a number of conglomeratic beds at thetop of the succession. Several specimens from this interval havepreviously been described as reworked and pointing to Early andMiddle Oxfordian ages (Alberti et al., 2011; Pandey et al., 2012a).The small islet Gangta Bet is particularly interesting as itssedimentary succession contains an abundant and diverse LateOxfordian fauna (upper Bifurcatus Zone), unknown from other
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
parts of the Kachchh Basin (Pandey et al., 2013a; Alberti et al.,2013b).
3. Material and methods
During four recent field surveys (2008-2013) to the KachchhBasin, the authors collected more than 800 ammonoids fromsections through the Upper Callovian, Oxfordian, and Kimmer-idgian. These fossils formed the base for a series of studies eachconcentrating on a particular taxonomic group. The Oxfordianammonite fauna of the area is strongly dominated by perisphinc-tids (Pandey et al., 2012a, 2013a, 2013b), followed by peltoceratids(Alberti et al., 2011; Pandey et al., 2015) and mayaitins. Ammonitesof other groups are very rare, but include oppeliids (e.g.,Taramelliceras) and phylloceratids (Pandey et al., 2013a). Anoverview of the stratigraphic distribution of the recorded taxa isgiven below.
The present study concentrates on the mayaitins, which arerepresented by 100 specimens in the collection. These fossils aremarked with the prefix GZN and currently housed at theGeoZentrum Nordbayern of the Friedrich-Alexander-UniversitatErlangen-Nurnberg, Germany, but are planned to be storedultimately at the Department of Geology of the University ofRajasthan, Jaipur, India. Further ammonites examined had beencollected by the Late John H. Callomon during his travels toKachchh between the years 1990 and 1998. This material carriesthe identifier OUMNH and is kept in the collections of the OxfordUniversity Museum of Natural History, England.
During the research, all localities in the Kachchh Basin known toexhibit Oxfordian outcrops have been investigated and corre-sponding sections measured. The specimens were collected with
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Table 1Geographic coordinates of the sampled sections.
Locality Latitude Longitude
Jara Dome N 23843’26.7’’ E 068859’26.9’’
Jumara Dome N 23841’13.6’’ E 069802’51.0’’
Keera Dome N 23835’05.3’’ E 069814’44.0’’
Medisar N 23823’38.2’’ E 069830’18.5’’
Jhura Camp N 23824’54.2’’ E 069836’26.3’’
Rudra Mata N 23822’11.7’’ E 069844’55.0’’
Lodai N 23822’08.4’’ E 069853’30.3’’
Mandvi Road N 23811’32.4’’ E 069836’08.0’’
Ler N 23811’40.9’’ E 069845’41.4’’
S-Khadir N 23847’00.3’’ E 070822’12.9’’
Kantkote N 23828’46.0’’ E 070827’26.3’’
Tramau River N 23829’26.4’’ E 070829’39.9’’
M. Alberti et al. / Geobios xxx (2015) xxx–xxx4
G Model
GEOBIO-701; No. of Pages 46
the highest possible stratigraphic resolution; only few have beentaken as stray samples in riverbeds adjacent to the Oxfordianoutcrops (as indicated in the descriptions below). The localitiesfrom which mayaitins have been collected for this study aremarked in Fig. 1, while Table 1 contains the geographic coordinatesof the corresponding sections. Only the material from Gangta Bet,which includes three mayaitins, has been described separately(Pandey et al., 2013a).
For most modern taxonomic studies on ammonites it isdesirable to work on material combining internal whorls (ornuclei), juveniles, and full-grown adults of a species to compre-hensively characterize its ontogenetic stages as well as intraspe-cific variability. In this context, the fossil ammonite record of theKachchh Basin suffers from a series of limitations as most of theammonites are found only as internal whorls. This problem isalready apparent in the material of Waagen (1873-1875) andSpath (1927-1933), but is also present in more recent studies (e.g.,Pandey et al., 2012a). Nevertheless, the mayaitins are compara-tively well preserved with eight species being represented byspecimens showing at least parts of the body chamber. Further-more, many specimens have been cross-sectioned to study theirpotential allometric growth and ontogenetic changes in whorl-section shape. The cross-sections obtained this way were scanned
Fig. 3. Measured dimensions of the ammonoids and abbreviations used. D:
diameter; Wh: whorl height; Ww: whorl width; U: width of the umbiculus.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
and measured digitally. Dimensions at different diameters aregiven in millimeters (numbers in parentheses are proportionaldimensions as percentage of diameter) and stored as electronicSupplementary Data (Tables S1-S19 in Appendix A). Measure-ments and abbreviations used are explained in Fig. 3. In general,suture lines were not visible very well on the collected specimens.Only four suture lines could be drawn from the venter to theumbilical seam, representing Mayaites maya, Mayaites rotundus,Epimayaites transiens, and Epimayaites smeei (Fig. 4).
The whorl sections of the cut specimens were furtherexamined using a Fourier shape analysis to evaluate theirpotential in differentiating the individual morphospecies. Two-dimensional whorl sections exhibit only a limited number ofdiscrete morphological features, making outline analysis a suitablealternative to the otherwise advantageous landmark analysis(Hammer and Harper, 2006). One-hundred-and-three whorlsections of 37 specimens at shell diameters > 50 mm havebeen outlined and subsequently formatted to represent1000 xy-coordinates using the image capture software tpsDig2
Fig. 4. Suture lines of some of the collected Mayaitinae. A. Mayaites maya
(GZN2009II 230); note that the suture line was drawn from a partly abraded flank
and therefore appears simpler. B. Mayaites rotundus (GZN2010I 1040). C.Epimayaites transiens (GZN2009II 220). D. Epimayaites smeei (GZN2010I 1092).
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

M. Alberti et al. / Geobios xxx (2015) xxx–xxx 5
G Model
GEOBIO-701; No. of Pages 46
(http://life.bio.sunysb.edu/morph/). Then, the Fourier shape anal-ysis reduces these coordinates to a smaller number of parameters(harmonics), which are finally subjected to a principal componentsanalysis.
The Fourier shape analysis decomposes each whorl-sectionshape described by xy-coordinates into harmonically-related sineand cosine curves with two Fourier coefficients per harmonic. Theharmonic spectrum is computed by Fast Fourier Transform (FFT)with the help of the program HANGLE, which operates on the tangentangle as a function of arc-length (Haines and Crampton, 2000).Mathematical terms are listed in the appendix of Haines andCrampton (2000). The first 12 harmonics have been chosen tocapture the whorl-section shape. This method calculates coeffi-cients that are computationally independent of each other. Incontrast, harmonics yielded by elliptic Fourier analysis ([EFA];Kuhl and Giardina, 1982) are increasingly down-weighted relativeto the first (Haines and Crampton, 2000). Consequently the choiceof using Fourier shape analysis over EFA enhances the discrimina-tory power of higher-order harmonics.
Crampton and Haines (1996) published three programs to beused for Fourier shape analysis: HANGLE, HMATCH, and HCURVE.Prior tothe computation of the FFT, HANGLE smoothes outlines by reducingthe high frequency pixel noise from the automatic tracing ofoutlines. For this, a weighted moving average over threecoordinates has been taken. Afterwards, the whorl sectionsunderwent ten smoothing iterations (Haines and Crampton,2000). HMATCH normalizes for the starting position and orientationof the whorl sections by adjusting the output of HANGLE. Itminimizes the sums of the squares of all differences between the103 whorl sections (Haines and Crampton, 2000). In a final step,multivariate statistical analysis of the Fourier coefficients wascarried out using a principal component analysis on a variance-covariance matrix.
4. The subfamily Mayaitinae Spath, 1928
Spath (1928b: p. 222) originally introduced the familyMayaitidae for ‘‘the later Macrocephalites’’ and included fivegenera: Mayaites, Epimayaites, Dhosaites, Paryphoceras, andPrograyiceras. Since then some authors have downgraded thegroup to the subfamily Mayaitinae, integrating it into the familySphaeroceratidae Buckman, 1920 (see Donovan et al., 1981;Brookfield and Westermann, 1982; Howarth, 1998; Krishna et al.,2000; Westermann et al., 2002; Jana et al., 2005), while othersretained its original status as a family (e.g., Thierry, 1975; Stevens,1997; Enay and Cariou, 1997, 1999; Enay, 2009). In the following itis considered to be a subfamily. A detailed discussion of itsaffinities is beyond the scope of the present article.
At present, the Mayaitinae consist of the genera Mayaites,Epimayaites, Dhosaites, and Paryphoceras. Arkell et al. (1957) placedthe genus Grayiceras Spath, 1923 into the affinity of this subfamily,but the status of this taxon is still doubtful. Specimens previouslyassigned to Grayiceras have been found to be of Callovian age andbetter related to the Eucycloceratinae (Westermann and Wang,1988; Enay and Cariou, 1997). However, Jana et al. (2005) argue forthe inclusion of this genus within the Mayaitinae based onseemingly Oxfordian specimens from the Spiti Valley, Himalaya. Inany case, Grayiceras does not occur in the Kachchh Basin, andmaterial from this region therefore cannot help in solving thisproblem. Another genus previously accommodated within theMayaitinae is Prograyiceras Spath, 1928, but this was included intoParyphoceras by Howarth (1998), a view which is followed here.
The origin of the Mayaitinae is still debated. Arkell et al. (1957)proposed that they derived from the Callovian-Oxfordian Pachy-ceratidae, but other authors (Thierry, 1975, 1976; Westermann
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
and Wang, 1988; Jana et al., 2005) discussed that the Eucyclocer-atinae represent the link between the Callovian Macrocephalitinaeand the Oxfordian Mayaitinae based on similarities in the suturelines between these groups (Donovan et al., 1981). Jana et al.(2005) suggested that the group formed in the Himalaya regionfrom where it spread into neighbouring areas.
Most members of the subfamily Mayaitinae have beendescribed from the Kachchh Basin by Waagen (1873-1875) andSpath (1927-1933). However, subsequent studies on the ammonitefauna of the basin have rather neglected this group. Although manyauthors have collected mayaitins during their research andincluded them in their faunal lists, they have rarely been describedor illustrated. This is probably due to their limited use instratigraphic correlations, as mayaitins have a comparativelylimited geographic occurrence and many species seem to be longranging. Thierry (1976) was the first to present a palaeogeographicmap showing the distribution of mayaitins known at that time, butsince then additional material has been described.
Apart from the Kachchh Basin, members of the subfamilyMayaitinae have been recorded from other localities of the Indiansubcontinent, such as the Jaisalmer Basin (e.g., Kachhara andJodhawat, 1981; Krishna, 1987; Chatterjee, 1990; Prasad, 2006;Pandey et al., 2012b, 2014) and the Indian Himalaya (Brookfieldand Westermann, 1982; Pandey et al., 2013c). Neighbouring areaswhere mayaitins have been found include Pakistan (Noetling,1895; Arkell, 1956), Nepal (Gradstein et al., 1991; Enay and Cariou,1997, 1999), and Tibet (Xiaochi and Grant-Mackie, 1988; Wester-mann and Wang, 1988; Xiaoqiao et al., 2000). Within the MalagasyGulf, separating eastern and western Gondwana, members of thesubfamily have been recorded from the Arabian Peninsula(Howarth and Morris, 1998; Howarth, 1998), East Africa (Torn-quist, 1893; Futterer, 1894; Arkell, 1956; Enay, 2009), andMadagascar (Lemoine, 1911; Collignon, 1959a, 1959b; Thierry,1975; Hoffmann and Keupp, 2008). The geographically furthestreliable records of the Mayaitinae are from the Sula Islands/NewGuinea (Westermann et al., 1978).
Specimens originally described as mayaitins from New Zealandand Argentina have later on been placed in the genus Araucanites,which belongs to the separate Eurycephalitinae (Stipanicic et al.,1975; Westermann and Riccardi, 1985; Hudson et al., 1987; vonHillebrandt and Groschke, 1995; Stevens, 1997; Hudson, 1999;Westermann et al., 2002; Parent, 2006; Parent et al., 2006; Riccardi,2008). A single specimen measuring 30 mm in diameter fromAntarctica has been described by Thomson (1982; Riccardi, 1991)as Epimayaites aff. transiens, but this record is rather doubtful(Westermann et al., 2002) and, in the light of the presently reviseddescription (see below), does not fit within this species. Braun andJordan (1976) described one fragmentary, deformed and flattenedsteinkern (diameter 50 mm) from Western Thailand as Epimayaites
cf. falcoides. This would be the only specimen known from thenorthern margin of the Tethys (Sato and Westermann, 1991).However, due to the poor preservation and the singularity of thisrecord, it should be regarded with great caution.
It seems therefore much more reliable to assume that thesubfamily Mayaitinae is restricted to the area around the MalagasyGulf and the northern margin of eastern Gondwana, while thesouthern and western margins of the supercontinent are inhabitedby members of the Eurycephalitinae (Fig. 5). Shells of deadmayaitins may have been drifting across ocean basins forconsiderable distances (as seen in Recent nautiloid shells andassumed for the extinct ammonites; Reyment, 2008), alsoemphasizing caution with singular records.
Since ammonites being an extinct group of organisms, theirtaxonomy is largely restricted to the concept of morphospecies.This designation implies that the described taxa do not necessarilycorrespond to a particular biospecies. Especially since the degree of
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 5. Palaeogeographic reconstructions. A. The Bathonian-Callovian world with the approximate range of the Macrocephalitinae and Eurycephalitinae. B. The Oxfordian
world with the described records and approximate range of the Mayaitinae and the genus Araucanites.
Modified and extended after Enay and Cariou (1997, 1999); see text for further references on the occurrences.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx6
G Model
GEOBIO-701; No. of Pages 46
intraspecific, morphological variability, and of gradual morpho-logical changes within a species through time cannot always beresolved (e.g., due to the limited number of specimens), thisconcept still forms the basis of taxonomy in palaeontology(Gingerich, 1985; Smith, 1994). This means that differentiationof fossil material into species often follows artificial morphologicalboundaries not necessarily congruent with those between livingbiospecies. Studies such as that of Kowalewski et al. (1997) showedthat the reliance on morphogroups does not automatically lead totaxonomic under-resolution. Nevertheless, this unsatisfying situ-ation is easily illustrated by the hypothesis of sexual dimorphismin ammonites, which has been proposed more than a century ago,and received new attention with the studies of Makowski (1962)and Callomon (1963). Following this hypothesis, two morphos-pecies represent the sexes of a single biospecies. These macro- andmicroconchs are characterized by certain features, such as adifference in size, a particular development of the peristome, or amodification of the ornamentation on the body chamber.Consequently, if two taxa show a parallel evolution with repeatedand simultaneous appearances of new characters in the ornamen-tation, they might belong to a single biospecies (Callomon, 1963).Since the rediscovery of this phenomenon, many dimorphic pairshave been described and the hypothesis has become wellestablished in research on ammonites (e.g., Makowski, 1971;Brochwicz-Lewinski and Rozak, 1976; Datta et al., 1996; Scher-zinger et al., 2006). Commonly, entire genera or subgenera havebeen found to represent either macro- or microconchs (Pandeyet al., 2012a).
Sexual dimorphism is also present in the subfamily Mayai-tinae. Callomon (1963) described members of the genusMayaites as macroconchs, while representatives of Epimayaites,Dhosaites, and Paryphoceras were thought to be the correspond-ing microconchs. The present collection shows that thisstatement is an oversimplification. While all known membersof Mayaites show a very large size and therefore justify theiridentification as macroconchs, there are also gigantic forms ofEpimayaites which surely represent macroconchs (e.g., Epi-
mayaites polyphemus, E. transiens, E. lemoini, and E. subtumidus).Still, smaller species of Epimayaites might indeed be
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
microconchs. Due to their size, the present collection ofmembers of Dhosaites and Paryphoceras supports their descrip-tion as microconchs. Following their general appearance, speciesof Dhosaites might be the microconchs to Mayaites, whilemembers of Paryphoceras may correspond to some of themacroconchs in Epimayaites. The study of sexual dimorphism inthe Mayaitinae of the Kachchh Basin is, however, hampered bythe fact that no specimens are preserved with a complete bodychamber and therefore peristome. In general, the low number ofspecimens for a series of species (especially of microconchs)does not allow the pairing of dimorphs at the species level.Much more material is needed to reliably recognize dimorphicpairs or validate the status of some of the morphospecies. Thus,only one preliminary attempt for the identification of adimorphic pair might be justified: the common co-occurrenceand generally similar appearances of Epimayaites transiens andE. falcoides suggest that they belong to the same biospecies. Themacroconch E. transiens attains a very large adult size and ischaracterized by a strongly compressed shell ornamented byfine and dense ribs, which fade towards the body whorl. Themicroconch E. falcoides is medium-sized and also has a verycompressed shell with fine and dense ribs which, however,persist across the adult body chamber.
Other taxa described herein might indeed be only variants of amorphologically flexible species. Again, more material is necessaryto reliably evaluate the intraspecific variability. Examples includeMayaites aff. maya and Mayaites jumarensis as possible variants ofM. maya. These and other cases are discussed in the followingdescriptions of the taxa, which are restricted to morphospeciesbecause of the aforementioned reasons.
5. Systematic Palaeontology
The suprageneric classification used below follows Donovanet al. (1981).
Class CEPHALOPODA Cuvier, 1797Order AMMONOIDEA Zittel, 1884Suborder AMMONITINA Hyatt, 1889
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

M. Alberti et al. / Geobios xxx (2015) xxx–xxx 7
G Model
GEOBIO-701; No. of Pages 46
Superfamily STEPHANOCERATACEAE Neumayr, 1875Family SPHAEROCERATIDAE Buckman, 1920Subfamily MAYAITINAE Spath, 1928Genus Mayaites Spath, 1924Type species: Ammonites maya Sowerby, 1840.Remarks: The genus Mayaites contains ammonites of large to
gigantic size, which are commonly inflated. The shells are rather
Fig. 6. Mayaites maya (Sowerby, 1840). A, B. GZN2009II 230 from the matrix of the Dhosa
Dhosa Conglomerate Bed near Medisar, Jhura Dome. The cross denotes the approxima
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
coarsely ribbed, except for the outer whorl, which becomessmooth. The ornamentation is comparatively simple anddominated by straight ribs. Characteristically, the ribs passstraight across the venter (Spath, 1928b; Arkell et al., 1957).
Mayaites maya (Sowerby, 1840)Figs. 4(A), 6, 7(A), 8(A-C)
Conglomerate Bed of the Jumara Dome. C, D. GZN2010I 1104 from the matrix of the
te position of the final septum of the phragmocone. Scale bar: 50 mm.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 7. Ontogenetic development of the whorl section A. Mayaites maya (Sowerby, 1840). B. Mayaites aff. maya (Sowerby, 1840). C. Mayaites jumarensis Spath, 1928.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx8
G Model
GEOBIO-701; No. of Pages 46
1840. Ammonites maya nov. sp. – Sowerby, p. 719, pl. 61, fig. 8.pars 1875c. Stephanoceras polyphemus (non Waagen) – Waagen,
p. 116, pl. 29, figs. 1, 3.pars 1895. Macrocephalites polyphemus non Waagen – Noetling,
pl. 11.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
1928b. M. maya (Sowerby) – Spath, p. 225, pl. 28, fig. 5, pl. 29,fig. 5, pl. 30, fig. 2, pl. 31, fig. 1(a, b), pl. 32, fig. 6, pl. 37, fig. 11(a, b),12, pl. 40, fig. 2, pl. 49, fig. 6, pl. 46, fig. 8.
1931b. M. maya (Sowerby) – Spath, pl. 119, fig. 1.1951. M. maya Sowerby – Basse and Perrodon, p. 53, pl. 4, fig. 4(a, b).
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 8. Cross-sections of specimens of Mayaites Spath, 1924. A-C. Mayaites maya (Sowerby, 1840); cross-section of GZN2009II 230 at a diameter of 159 mm (A); cross-section of
GZN2010I 1104 at a diameter of 202 mm (B); cross-section of GZN2010I 1103 at a diameter of 213 mm (C). D. Mayaites aff. maya (Sowerby, 1840); cross-section of GZN2010I
1105 at a diameter of 224 mm. E. Mayaites jumarensis Spath, 1928; cross-section of GZN2010I 1094 at a diameter of 194 mm. F, G. Mayaites rotundus Spath, 1928; cross-
section of GZN2010I 1040 at a diameter of 140 mm (F); cross-section of GZN2010I 1038 at a diameter of 95 mm (G). H. Mayaites subkobyi Spath, 1928; cross-section of
GZN2009II 135 at a diameter of 84 mm. Note the reduced size of the drawings. Scale bar: 50 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 9
G Model
GEOBIO-701; No. of Pages 46
1959a. M. maya (Sowerby) – Collignon, pl. 40, fig. 208.Holotype: The specimen figured by Sowerby (1840: pl. 61,
fig. 8) and refigured by Spath (1928b: pl. 31, fig. 1, and pl. 32, fig. 6).Material: Three specimens. One specimen from the matrix of
the Dhosa Conglomerate Bed of the Jumara Dome (GZN2009II 230).Two specimens from the matrix of the Dhosa Conglomerate Bednear Medisar, Jhura Dome (GZN2010I 1103, 1104).
Earlier records in the Kachchh Basin: The locality of theholotype is doubtful, but Waagen (1875c) and Spath (1928b)mention M. maya from the Dhosa Oolite member of the Jhura Dome(Niruna, Badi), Habo Dome (Lodai), and south of Bhuj (Samatra,Barasar, Fakirwari, Ler).
Measurements: Table S1.Description: Very large (diameter > 240 mm), involute shell,
septate until a diameter of 160 to 180 mm. Shells depressed in theinner whorls, but becoming more and more compressed duringontogeny (Fig. 7(A)). Whorl section subrounded with flanksmerging smoothly into rounded venter without prominentventrolateral shoulder. Umbilical edge rounded but distinct,
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
umbilical wall slightly rounded, steeply inclined, umbilicus deep(Fig. 8(A-C)). Maximum inflation just above the umbilical edge. Theornamentation consists of weakly flexuous ribs on the innermostwhorls, which become straight on the middle whorls and turn intobroad weak folds on the body chamber. Primary ribs on innerwhorls branch slightly below the Middle of the flank into two tothree secondaries. Free intercalatory ribs are common. In general,for each primary rib there are around three to five secondaries,which pass almost straight across the venter. Secondary ribsdisappear on the body chamber.
Remarks: The present material matches the photographs anddescriptions of the larger specimens of M. maya provided by Spath(1928b, 1931b). The holotype of the species is a large shell(maximum diameter 195 mm) with probably not more than thebeginning of the body chamber preserved. Diagnostic for thespecies is an increasing whorl height to whorl width ratio with thewhorl section of the body chamber being almost as high as wide(Fig. 7(A)). Furthermore, the ribbing is comparatively straight onthe flanks and on the venter. In addition to the larger specimens,
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

M. Alberti et al. / Geobios xxx (2015) xxx–xxx10
G Model
GEOBIO-701; No. of Pages 46
Spath (1928b) included a series of smaller nuclei or Middle whorlsof phragmocones into this species, thereby demonstrating a largeintraspecific variability. Such a variability in the dimensions andshape of the inner whorls can also be seen in the cross-sections ofthe present material (Fig. 8(A-C)). Nevertheless, some of the nucleiillustrated by Spath (1928b) might actually be too small for areliable identification, but they have been left in the synonymysince the originals could not be examined during the present study.
The diagnosis of M. maya has created problems shortly after itsfirst description. Waagen (1875c) has described specimens asStephanoceras maya, but these were later on included partly inE. lemoini by Spath (1928b). Similar problems exist with the earlydescriptions by Tornquist (1893), Uhlig (1910), and Lemoine(1911) (Spath, 1928b: p. 227). Collignon (1959a) described aspecimen of M. maya from Madagascar which is incomplete andcomparatively compressed, but might still fit within the intraspe-cific variability of the species.
The closest comparable form is M. jumarensis, which occurs atthe same stratigraphic level. This species has been separated from
Fig. 9. A. Mayaites jumarensis Spath, 1928; GZN2010I 1094 from the Dhosa Oolite memb
the matrix of the Dhosa Conglomerate Bed near Medisar, Jhura Dome. The cross denotes t
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
M. maya by its stronger ribbing as well as a smaller umbilicus. Asdescribed below, the species also seems to be slightly moredepressed than the average M. maya. Another similar species isE. polyphemus which also occurs at the same localities. However,this species can be differentiated by being compressed throughoutontogeny, having a comparatively pointed venter, and by reachinga slightly larger size.
Mayaites aff. maya (Sowerby, 1840)Figs. 7(B), 8(D), 9(B), 10(F)aff. 1840. A. maya nov. sp. – Sowerby, p. 719, pl. 61, fig. 8.aff. 1928b. M. maya (Sowerby) – Spath, p. 225, pl. 31, fig. 1(a, b),
pl. 32, fig. 6.aff. 1931b. M. maya (Sowerby) – Spath, pl. 119, fig. 1.Material: One specimen from the matrix of the Dhosa
Conglomerate Bed near Medisar, Jhura Dome (GZN2010I 1105).Measurements: Table S2.Description: Very large (diameter > 260 mm), depressed,
involute shell, septate until a diameter of not more than
er of the Jumara Dome. B. Mayaites aff. maya (Sowerby, 1840); GZN2010I 1105 from
he approximate position of the final septum of the phragmocone. Scale bar: 50 mm.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 10. A-D. Mayaites sp. indet.; GZN2009II 157 from the matrix of the Dhosa Conglomerate Bed of the Keera Dome (A, B); GZN2009II 144 from the Dhosa Oolite member near
Jhura Camp, Jhura Dome (C, D). E. Mayaites jumarensis Spath, 1928; GZN2010I 1094 from the Dhosa Oolite member of the Jumara Dome. F. Mayaites aff. maya (Sowerby, 1840);
GZN2010I 1105 from the matrix of the Dhosa Conglomerate Bed near Medisar, Jhura Dome. The cross denotes the approximate position of the final septum of the
phragmocone. Scale bars: 10 mm (A-D), 30 mm (E, F).
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 11
G Model
GEOBIO-701; No. of Pages 46
150 mm (Fig. 7(B)). Whorl section subrounded with flanksgrading smoothly into broadly rounded venter without pro-nounced ventrolateral shoulder (Fig. 8(D)). Umbilical edgerounded, umbilical wall steeply inclined, umbilicus deep.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Maximum inflation just above the umbilical edge. Theornamentation consists of moderately thick primary ribs, whichturn into broad folds on the otherwise almost smooth bodychamber. Secondary ribs of inner whorls not observable,
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

M. Alberti et al. / Geobios xxx (2015) xxx–xxx12
G Model
GEOBIO-701; No. of Pages 46
folds on the outer whorl cross the venter straight withoutbranching.
Remarks: The closest comparable form to the present specimenis M. maya as described and illustrated by Spath (1928b,1931b). However, the specimen has a more broadly roundedventer and is depressed throughout its ontogeny (Fig. 7(B)). Inaddition, M. maya seems to be slightly larger with phragmoconesreaching sizes between 160 and 180 mm. As Spath (1928b)described a large intraspecific variability, it might be connected toM. maya through an array of intermediate forms, but it might alsorepresent a distinct morphospecies. Additional material is neces-sary to clarify this point.
M. jumarensis is also a closely allied form, but has a smallerumbilicus and is more coarsely ribbed (Spath, 1928b: p. 228).Mayaites subkobyi and Mayaites rotundus are generally moredepressed and the latter species also has a slightly widerumbilicus (Spath 1928b: pp. 227, 229). Some ontogenic stages ofMayaites radiatus Spath, 1928 may have similar dimensions asthe present specimen, but according to Spath (1928b: p. 231)the change from a less depressed to a more depressed form istypical for the species. Such a change is not present in thecurrent specimen. E. polyphemus shows some resemblance to thepresent specimen, especially the illustration of the lectotype byNoetling (1895: pl. 12). However, this species, as describedbelow, seems to be restricted to compressed forms (Noetling,1895; Spath, 1928b).
Mayaites jumarensis Spath, 1928Figs. 7(C), 8(E), 9(A), 10(E)1928b. M. jumarensis nov. sp. – Spath, p. 228, pl. 43, fig. 1(a, b).Holotype: The specimen figured by Spath (1928b: pl. 43, fig.
1(a, b)).Material: One specimen from the Dhosa Oolite member of the
Jumara Dome (GZN2010I 1094).Earlier records in the Kachchh Basin: M. jumarensis has been
originally described by Spath (1928b) from the Dhosa Oolitemember of the Jumara Dome as well as from Samatra andFakirwari, south of Bhuj.
Measurements: Table S3.Description: Part of a depressed, involute phragmocone
(septate until a diameter of at least 195 mm; Fig. 7(C)). Whorlsection subrounded with flanks merging smoothly into broadlyrounded venter (Fig. 8(E)). Umbilical edge and umbilical wallrounded. Umbilicus narrow. Maximum inflation just above theumbilical edge. The ornamentation consists of coarse, almoststraight, slightly prorsiradiate ribs originating at the umbilicaledge. Primary ribs branch into two to three finer secondaries atabout one-third of lateral height which cross the venter straight.Free intercalatory secondary ribs are present.
Remarks: The present specimen matches the description andillustration of M. jumarensis by Spath (1928b). The holotype has aslightly smaller umbilicus (Table S3), but this is considered to fallwithin the range of intraspecific variability.
M. maya is a closely related form but can be separated fromM. jumarensis by its more compressed whorl section of the outerwhorls, a weaker ribbing, and possibly a slightly larger umbilicus.Furthermore, the present specimen is slightly larger than thosedescribed above as M. maya. Although these differences can beobserved in the current collection, they may be the expression of avariable biospecies. This view is supported by the observation ofSpath (1928b) that there are specimens fitting morphologically inbetween M. jumarensis and M. maya. For now, however, thedistinction is retained, awaiting further material.
Mayaites rotundus Spath, 1928Figs. 4(B), 8(F, G), 11, 12(A)
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
1928b. Mayaites rotundus nov. sp. – Spath, p. 227, pl. 30, fig. 3(a-c), pl. 44, fig. 7(a, b).
Holotype: The specimen figured by Spath (1928b: pl. 30, fig.3(a-c)).
Material: Three specimens. One specimen from the DhosaOolite directly underneath the Dhosa Conglomerate Bed nearRudra Mata, Habo Dome (GZN2009II 168). Two specimens fromthe matrix of the Dhosa Conglomerate Bed near Ler, south of Bhuj(GZN2010I 1038, 1040).
Earlier records in the Kachchh Basin: The species has beenoriginally described from the Dhosa Oolite member near Fakirwari,south of Bhuj (Spath, 1928b).
Measurements: Table S4.Description: Parts of moderately large, strongly depressed,
involute phragmocones (septate until a diameter of at least140 mm). Whorl section subrounded with flanks grading smoothlyinto broadly rounded venter with indistinct ventrolateral shoulder(Fig. 8(F, G)). Umbilical edge pronounced, umbilical wall nearlyvertical. Umbilicus deep. Maximum inflation just above theumbilical edge. The ornamentation consists of moderately thickprimary ribs, which branch indistinctly into two to three secondaryribs at one-third of lateral height. Intercalatory secondary ribsoccur. Ribs are comparatively straight, originate at the umbilicalwall rursiradiately, but turn recti- to slightly prorsiradiately at theumbilical edge. Secondary ribs pass straight across the venter.
Remarks: These specimens match the illustration and descrip-tion of M. rotundus by Spath (1928b). The dimensions noted bySpath (1928b: p. 228) for the holotype of the species indicate aratio between whorl height to whorl width of 0.65 compared tovalues above 0.70 in the present specimens (Fig. 12(A)). However,measurements based on the illustrations of the holotype by Spath(1928b) indicate rather comparable ratios (pl. 30, fig. 3(c): 0.79; pl.30, fig. 3(a, b): 0.73; Table S4). In addition, the outermost part of theholotype is slightly fragmented, and therefore the width of thewhorl can only be estimated there.
The species can be differentiated from most other members ofthe genus by its strongly depressed whorl section and compara-tively straighter ribs. M. maya and M. jumarensis are related forms,but have a more compressed whorl section (Spath, 1928b: p. 225,228). Mayaites obesus Spath, 1928 shows a much smaller umbilicusand concave-forward-directed ribbing on the inner part of theflank and backward-directed secondary ribs on the outer part ofthe flank (Spath, 1928b: p. 228). Mayaites subkobyi is characterizedby considerably wider ribs (Spath, 1928b: p. 229).
Mayaites subkobyi Spath, 1928Figs. 8(H), 12(B), 131928b. Mayaites subkobyi nov. sp. – Spath, p. 229, pl. 37, fig. 13,
pl. 45, fig. 1(a, b).1931b. Mayaites aff. subkobyi Spath – Spath, pl. 103, fig. 5.Holotype: The specimen figured by Spath (1928b: pl. 45, fig.
1(a, b)).Material: One specimen from a thin layer of Dhosa Oolite on top
of the Dhosa Conglomerate Bed at Rudra Mata, Habo Dome(GZN2009II 135).
Earlier records in the Kachchh Basin: The species hasoriginally been described by Spath (1928b) from the Dhosa Oolitemember near Fakirwari, south of Bhuj.
Measurements: Table S5.Description: Part of a small, strongly depressed, involute
phragmocone (Fig. 12(B)). Whorl section subrounded with broadlyrounded venter (Fig. 8(H)). Umbilical edge distinct, umbilical wallslightly rounded. Umbilicus deep. Maximum inflation slightlyabove the umbilical edge. The ornamentation consists of coarse,comparatively sparse primary ribs originating at the umbilicaledge, which branch indistinctly into two to three weaker
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 11. A, B. Mayaites rotundus Spath, 1928; GZN2010I 1040 from the matrix of the Dhosa Conglomerate Bed near Ler, south of Bhuj. Scale bar: 20 mm.
Fig. 12. Ontogenetic development of the whorl section. A. Mayaites rotundus Spath, 1928. B. Mayaites subkobyi Spath, 1928.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 13
G Model
GEOBIO-701; No. of Pages 46
Please cite this article in press as: Alberti, M., et al., Ammonites of the subfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,Western India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2015.02.005

Fig. 13. Mayaites subkobyi Spath, 1928; GZN2009II 135 from a thin layer of Dhosa
Oolite on top of the Dhosa Conglomerate Bed at Rudra Mata, Habo Dome. Scale bar:
20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx14
G Model
GEOBIO-701; No. of Pages 46
secondary ribs at around one-third of lateral height. Characteristicis a thickening or broadening of the primary ribs on the inner flank.Secondary ribs cross the venter straight or with a very slightforward-directed sinuosity.
Remarks: The specimen matches the description of M. subkobyi
by Spath (1928b) and shows the characteristic thickening of theprimary ribs. The illustration of the holotype seems to have slightlysparser primary ribs, but this is regarded to be within theintraspecific variation. The holotype is the inner part of aphragmocone with a diameter of 85 mm (Table S5). Accordingto Spath (1928b) the species was still septate at diameters of190 mm. M. maya is a comparable taxon, but has a generally morecompressed whorl section. M. rotundus has similar dimensions, butits ribs are much denser.
Mayaites sp. indet.Fig. 10(A-D)Material: Fourteen specimens. One specimen from the matrix
of the Dhosa Conglomerate Bed of the Keera Dome (GZN2009II157). Five specimens from the Dhosa Oolite member near JhuraCamp, Jhura Dome (GZN2009II 144, 161, 188, 239, GZN2010I 014).One specimen from a thin layer of Dhosa Oolite on top of the DhosaConglomerate Bed at Rudra Mata, Habo Dome (GZN2009II 155).One specimen from the Dhosa Oolite member near Lodai, HaboDome (GZN2009II 147). One specimen from the matrix of theDhosa Conglomerate Bed at the Mandvi Road, south of Bhuj(GZN2009II 182). Five specimens from the Dhosa Oolite memberdirectly underneath the Dhosa Conglomerate Bed at Ler, south ofBhuj (GZN2009II 153, 154, 186, 187, 245).
Description: Small, depressed to slightly compressed, involutenuclei of phragmocones. Whorl section subelliptical to rounded.Umbilical edge rounded, umbilical wall steep, umbilicus narrow.The ornamentation consists of comparatively rectiradiate ribs,branching at different heights on the flank into finer secondaries,which pass straight across the venter.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Remarks: These are nuclei of phragmocones, which are toosmall for reliable specific identification. The rectiradiate ribbing, arather depressed whorl section, and the straight ribs on the ventersupport an assignment to the genus Mayaites. Differences inproportional dimensions and style of ribbing between the speci-mens show that they belong to several species.
Genus Epimayaites Spath, 1928Type species: Stephanoceras transiens Waagen, 1875.Remarks: The genus Epimayaites contains ammonites of
variable adult size, which are characterized by a sharper andmore elegant ribbing than Mayaites. Characteristically, the ribspass the venter with a forward-directed sinuosity. While somespecies are inflated and depressed, the group contains severalcompressed forms (Spath, 1928b; Arkell et al., 1957).
Epimayaites polyphemus (Waagen, 1875)Figs. 14, 15(A, B), 16(A), 17(A, B)pars 1875c. Stephanoceras polyphemus nov. sp. – Waagen, p.
116, pl. 29, fig. 2 (non figs. 1, 3).pars 1895. Macrocephalites polyphemus Waagen – Noetling, pl.
12 (non pl. 11).1928b. Epimayaites polyphemus (Waagen) – Spath, p. 233.Lectotype: The specimen figured by Waagen (1875c: suture
line of pl. 29, fig. 2) and Noetling (1895: pl. 12).Material: Three specimens. One specimen from the matrix of
the Dhosa Conglomerate Bed of the Jara Dome (GZN2009II 181).Two specimens from the matrix of the Dhosa Conglomerate Bednear Lodai, Habo Dome (GZN2010I 1101, 1102).
Earlier records in the Kachchh Basin: Waagen (1875c)recorded the lectotype of E. polyphemus from the Dhosa Oolitemember near Lodai, Habo Dome. Spath (1928b: p. 239) describedanother, very similar specimen transitional to E. lemoini orE. excentricus from Kantkote, Wagad Uplift.
Measurements: Table S6.Description: Very large (diameter > 260 mm), compressed,
involute shell, septate until a diameter of around 190 mm(Fig. 16(A)). Whorl section subrounded with flanks gradingsmoothly into rounded (body chamber) to slightly pointed venterwithout pronounced ventrolateral shoulder (Fig. 17(A, B)).Umbilical edge rounded, umbilical wall slightly rounded, steeplyinclined. Umbilicus narrow, but strongly widening at the bodychamber. Maximum inflation just above the umbilical edge. Theornamentation consists of primary ribs branching slightly belowthe Middle of the flank into finer, forward-directed secondary ribs.The number of secondaries per primary rib increases duringontogeny (generally ranging from two to five secondaries perprimary). Secondary ribs cross the venter with slight forward-directed sinuosity on inner and Middle whorls. Body chamberalmost smooth with only weak broad folds. Adult shell showsdistinct uncoiling of the body chamber (Fig. 14(B)).
Remarks: The specimens match previous descriptions ofE. polyphemus as well as the illustration of the lectotype byNoetling (1895). Characteristic of the species is its large size andthe compressed shell with slightly prorsiradiate secondary ribs(Spath, 1928b: p. 233). Specimens illustrated previously under thisname by Waagen (1875c: pl. 29, figs. 1, 3) have been assigned toM. maya by Spath (1928b). Additionally, Spath (1928b) mentionedanother specimen from Pakistan described by Noetling (1895: pl.11) as E. polyphemus to belong to M. maya. These problematicidentifications already suggest a great resemblance between bothspecies, especially if only the outermost smooth whorl isexamined. However, the inner and Middle whorls of M. maya
are depressed, becoming compressed only in the outer whorls. Incontrast, E. polyphemus is compressed with an almost constantratio of whorl height to whorl width throughout ontogeny(Fig. 16(A)).
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005
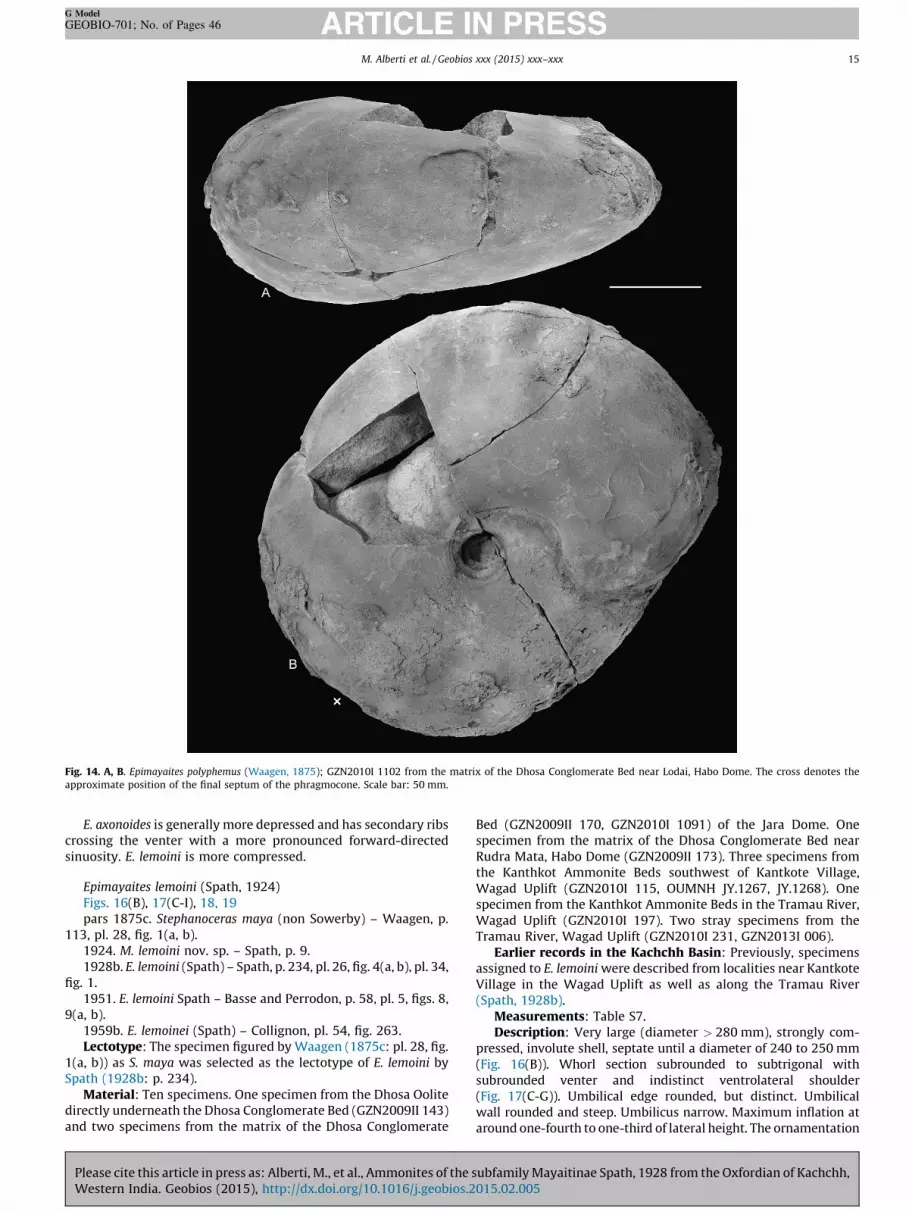
Fig. 14. A, B. Epimayaites polyphemus (Waagen, 1875); GZN2010I 1102 from the matrix of the Dhosa Conglomerate Bed near Lodai, Habo Dome. The cross denotes the
approximate position of the final septum of the phragmocone. Scale bar: 50 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 15
G Model
GEOBIO-701; No. of Pages 46
E. axonoides is generally more depressed and has secondary ribscrossing the venter with a more pronounced forward-directedsinuosity. E. lemoini is more compressed.
Epimayaites lemoini (Spath, 1924)Figs. 16(B), 17(C-I), 18, 19pars 1875c. Stephanoceras maya (non Sowerby) – Waagen, p.
113, pl. 28, fig. 1(a, b).1924. M. lemoini nov. sp. – Spath, p. 9.1928b. E. lemoini (Spath) – Spath, p. 234, pl. 26, fig. 4(a, b), pl. 34,
fig. 1.1951. E. lemoini Spath – Basse and Perrodon, p. 58, pl. 5, figs. 8,
9(a, b).1959b. E. lemoinei (Spath) – Collignon, pl. 54, fig. 263.Lectotype: The specimen figured by Waagen (1875c: pl. 28, fig.
1(a, b)) as S. maya was selected as the lectotype of E. lemoini bySpath (1928b: p. 234).
Material: Ten specimens. One specimen from the Dhosa Oolitedirectly underneath the Dhosa Conglomerate Bed (GZN2009II 143)and two specimens from the matrix of the Dhosa Conglomerate
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Bed (GZN2009II 170, GZN2010I 1091) of the Jara Dome. Onespecimen from the matrix of the Dhosa Conglomerate Bed nearRudra Mata, Habo Dome (GZN2009II 173). Three specimens fromthe Kanthkot Ammonite Beds southwest of Kantkote Village,Wagad Uplift (GZN2010I 115, OUMNH JY.1267, JY.1268). Onespecimen from the Kanthkot Ammonite Beds in the Tramau River,Wagad Uplift (GZN2010I 197). Two stray specimens from theTramau River, Wagad Uplift (GZN2010I 231, GZN2013I 006).
Earlier records in the Kachchh Basin: Previously, specimensassigned to E. lemoini were described from localities near KantkoteVillage in the Wagad Uplift as well as along the Tramau River(Spath, 1928b).
Measurements: Table S7.Description: Very large (diameter > 280 mm), strongly com-
pressed, involute shell, septate until a diameter of 240 to 250 mm(Fig. 16(B)). Whorl section subrounded to subtrigonal withsubrounded venter and indistinct ventrolateral shoulder(Fig. 17(C-G)). Umbilical edge rounded, but distinct. Umbilicalwall rounded and steep. Umbilicus narrow. Maximum inflation ataround one-fourth to one-third of lateral height. The ornamentation
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 15. A, B. Epimayaites polyphemus (Waagen, 1875); GZN2009II 181 from the matrix of the Dhosa Conglomerate Bed of the Jara Dome. C, D. Epimayaites axonoides Spath,
1928; GZN2010I 072 from the Kanthkot Ammonite Beds southwest of Kantkote Village, Wagad Uplift. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx16
G Model
GEOBIO-701; No. of Pages 46
consists of slightly flexuous ribs with primaries originating at theumbilical edge and branching mostly into two secondaries slightlyabove the Middle of the whorl height (Fig. 17(H, I)). Generally, two tothree free secondary ribs are intercalated. Density of primariesvaries between specimens. Number of secondaries per primary rib
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
increases with size. Secondaries cross the venter with pronouncedforward-directed sinuosity. Body chamber increasingly smooth.
Remarks: The specimens match the illustration and descriptionof E. lemoini by Spath (1928b). The species can be distinguishedfrom its relatives by its dimensions as well as by the style of
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 16. Ontogenetic development of the whorl section. A. Epimayaites polyphemus (Waagen, 1875). B. Epimayaites lemoini (Spath, 1924).
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 17
G Model
GEOBIO-701; No. of Pages 46
ribbing. E. transiens is more compressed, has a smaller umbilicus,and finer and more numerous ribs, which cross the venter withonly a slight forward-directed sinuosity. The specimen GZN2009II143 also shows a slightly denser ribbing and a weaker forward-directed sinuosity on the venter (Fig. 18(A, B)), but its dimensionsas well as the more flexuous ribbing allow its identification asE. lemoini. E. axonoides is less compressed and has coarser ribs(Spath, 1928b: p. 235).
Epimayaites axonoides Spath, 1928Figs. 15(C, D), 17(J-O), 20, 21(A)1928b. E. axonoides nov. sp. – Spath, p. 235, pl. 25, fig. 8(a, b), pl.
34, fig. 3(a, b), pl. 47, fig. 2.1931a. E. axonoides Spath – Spath, pl. 48, fig. 4, pl. 49, fig. 8.1959b. E. axonoides Spath – Collignon, pl. 53, figs. 261, 262.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Holotype: The specimen figured by Spath (1928b, 1931a: pl. 47,fig. 2, pl. 48, fig. 4, and pl. 49, fig. 8).
Material: Seven specimens. Two specimens from the KanthkotAmmonite Beds southwest of Kantkote Village, Wagad Uplift(GZN2010I 072, GZN2011I 349). Four specimens from theKanthkot Ammonite Beds in the Tramau River, Wagad Uplift(GZN2010I 136, 142, 159, 201). One stray specimen from theTramau River, Wagad Uplift (GZN2013I 004).
Earlier records in the Kachchh Basin: Spath (1928b) describedE. axonoides from localities near Kantkote Village in the WagadUplift.
Measurements: Table S8.Description: Fragments of involute phragmocones. Inner
whorls with a subrounded whorl section with slightly archedflanks merging into subrounded venter along a smooth curve.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 17. Cross-sections and ribbing patterns of Epimayaites Spath, 1928. A, B. Epimayaites polyphemus (Waagen, 1875); cross-section of GZN2010I 1102 at a diameter of
253 mm (A); cross-section of GZN2009II 181 at a diameter of 110 mm (B). C-I. Epimayaites lemoini (Spath, 1924);cross-section of GZN2013I 006 at a diameter of 233 mm (C); cross-
section of GZN2010I 1091 at a diameter of 182 mm (D); cross-section of GZN2010I 197 at a diameter of 135 mm (E); cross-section of GZN2009II 143 at a diameter of 84 mm (F);
cross-section of GZN2010I 231 at a diameter of 70 mm (G); ribbing pattern on left side of OUMNH JY.1267 at a diameter of 90 mm (H); ribbing pattern on left side of GZN2009II
143 at a diameter of 68 mm (I). J-O. Epimayaites axonoides Spath, 1928; cross-section of GZN2013I 004 at a diameter of 127 mm (J); cross-section of GZN2011I 349 at a diameter of
127 mm (K); cross-section of GZN2010I 072 at a diameter of 108 mm (L); cross-section of GZN2010I 201 at a diameter of 85 mm (M); ribbing pattern on left side of GZN2011I
349 at a diameter of 127 mm (N); ribbing pattern on right side of GZN2010I 142 at a diameter of 44 mm (O). Note the reduced size of the drawings. Scale bar: 50 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx18
G Model
GEOBIO-701; No. of Pages 46
Venter becomes slightly pointed on Middle and outer whorls.Whorl width almost equal to whorl height (Figs. 17(J-M), 21(A)).Umbilical edge rounded, but distinct. Umbilical wall roundedand steep. Umbilicus deep. Maximum inflation slightly aboveumbilical edge. The ornamentation consists of slightly flexuousribs originating on the outer umbilical wall (Fig. 17(N, O)).Primary ribs concave-forward on inner flank, branching slightlyabove mid-flank into two, more rarely three, secondary ribs. Thenumber of secondaries per primary rib increases with diameteras does the number of free intercalatory secondary ribs. Inaddition, the ribs become straighter during ontogeny.Secondaries cross the venter with prominent forward-directedsinuosity.
Remarks: The specimens match the description and illustrationof E. axonoides by Spath (1928b, 1931a). The holotype of the speciesis a large phragmocone with a diameter of 193 mm (Spath, 1928b:p. 234; Fig. 21(A); Table S8). The present material consists only ofMiddle and inner whorls of phragmocones showing considerable
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
variation in whorl shape. Nevertheless, the ratio of whorl height towhorl width is largely between 0.85 and 1.05. Thereby it is lowerthan in the closely related E. lemoini, which could be confusedregarding its very similar ornamentation. E. subtumidus, incontrast, is a much more inflated form occurring at the samestratigraphic level.
Epimayaites subtumidus (Waagen, 1875)Figs. 21(B), 22–24(A-C)1875c. Stephanoceras subtumidum nov. sp. – Waagen, p. 118, pl.
28, fig. 4(a, b).1928b. E. subtumidus (Waagen) – Spath, p. 237, pl. 25, fig. 5, pl.
29, fig. 6(a, b), pl. 31, fig. 7, pl. 32, fig. 9.Holotype: The specimen figured by Waagen (1875c: pl. 28, fig.
4(a, b)).Material: Five specimens from the Kanthkot Ammonite Beds
southwest of Kantkote Village, Wagad Uplift (GZN2010I 095, 113,114, GZN2011I 337, 341).
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 18. Epimayaites lemoini (Spath, 1924). A, B. GZN2009II 143 from the Dhosa Oolite directly underneath the Dhosa Conglomerate Bed of the Jara Dome. C, D. OUMNH
JY.1268 from the Kanthkot Ammonite Beds southwest of Kantkote Village, Wagad Uplift. E, F. OUMNH JY.1267 from the Kanthkot Ammonite Beds southwest of Kantkote
Village, Wagad Uplift. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 19
G Model
GEOBIO-701; No. of Pages 46
Earlier records in the Kachchh Basin: E. subtumidus isoriginally known from localities near Kantkote Village and inthe Tramau River in the Wagad Uplift (Waagen, 1875c; Spath,1928b).
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Measurements: Table S9.Description: Parts of large, strongly depressed, involute
phragmocones (septate until a diameter of at least 190 mm;Fig. 21(B)). Whorl section rounded with flanks merging into
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 19. A, B. Epimayaites lemoini (Spath, 1924); GZN2013I 006 from the Tramau River, Wagad Uplift. The cross denotes the approximate position of the final septum of the
phragmocone. Scale bar: 30 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx20
G Model
GEOBIO-701; No. of Pages 46
broadly rounded venter without prominent ventrolateral shoulder(Fig. 24(A-C)). Umbilical edge rounded, but distinct. Umbilical wallnearly vertical and steep. Umbilicus very deep and narrow.Maximum inflation just above the umbilical edge. The ornamen-tation consists of broad, comparatively straight primary ribsoriginating at the umbilical edge and branching into two or threesecondaries at the Middle of the flank. In the outermost whorls ofthe phragmocone, the ribbing is still dense, but becomes weakerand the branching points are indistinct. Secondary ribs cross theventer with prominent forward-directed sinuosity.
Remarks: The specimens match descriptions and illustrationsof E. subtumidus by Waagen (1875c) and Spath (1928b). Due to thestrongly depressed whorl section in combination with theforward-directed sinuosity of the secondary ribs on the venter,this species can be easily differentiated from other related forms.E. axonoides occurs at the same locality and stratigraphic level, butis distinctly more compressed. At some ontogenetic stagesM. obesus Spath, 1928 shows a similarly depressed whorl sectionand a narrow umbilicus, but at larger diameters it becomes more
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
compressed. Furthermore, M. obesus has distinctly curved ribs(Spath, 1928b: p. 228).
Epimayaites cf. evolutus Spath, 1928Figs. 24(D-G), 25(A-C), 26(A)cf. 1928b. E. evolutus nov. sp. – Spath, p. 238, pl. 36, figs. 13, 14,
pl. 37, fig. 5(a-c).Holotype: The specimen figured by Spath (1928b: pl. 37, fig.
5(a-c)).Material: Three specimens. Two specimens from the Kanthkot
Ammonite Beds southwest of Kantkote Village, Wagad Uplift(GZN2010I 068, 070). One specimen from the Kanthkot AmmoniteBeds in the Tramau River, Wagad Uplift (GZN2010I 207).
Earlier records in the Kachchh Basin: Spath (1928b) recordedE. evolutus from localities around Kantkote Village in the WagadUplift.
Measurements: Table S10.Description: Parts of compressed, involute phragmocones
(septate until a diameter of at least 155 mm; Fig. 26(A)). Whorl
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 20. Epimayaites axonoides Spath, 1928. A, B. GZN2011I 349 from the Kanthkot Ammonite Beds southwest of Kantkote Village, Wagad Uplift. C. GZN2010I 136 from the
Kanthkot Ammonite Beds in the Tramau River, Wagad Uplift. D, E. GZN2010I 142 from the Kanthkot Ammonite Beds in the Tramau River, Wagad Uplift. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 21
G Model
GEOBIO-701; No. of Pages 46
section elliptical with slightly flattened flanks and rounded venter(Fig. 24(D, E)). Umbilical edge rounded, umbilical wall steep.Umbilicus narrow. Maximum inflation at about one-third of lateralheight. The ornamentation consists of coarse, sharp ribs originat-ing on the umbilical wall. Primary ribs pass umbilical edge withprominent forward-directed concavity and branch at the Middle ofthe flank into mostly two secondary ribs of similar strength(Fig. 24(F, G)). Commonly, a free secondary rib is intercalated. Theribs are slightly biconcave and slightly prorsiradiate. Secondariescross the venter with pronounced forward-directed sinuosity.
Remarks: The specimens match E. evolutus as described andillustrated by Spath (1928b). Due to their small size not allcharacteristics of the species could be seen. The present specimens
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
show that the innermost whorls of the phragmocone are depressedand with ontogeny the whorl section gets increasingly morecompressed. Due to their small size, however, their whorl section isstill not as compressed as that of the wholly septate holotype witha diameter of 153 mm (Fig. 26(A); Table S10). Unfortunately, Spath(1928b) did not list dimensions of smaller specimens. Further-more, Spath (1928b) recorded that the primary ribs become moredistant on the outer part of the phragmocone, which could also notbe observed in the present specimens at smaller diameters. Due tothese reasons, they have only tentatively been placed in thisspecies.
Another similar form is E. lemoini, but this species has adistinctly smaller umbilicus. Furthermore, the ribs of E. evolutus
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 21. Ontogenetic development of the whorl section. A. Epimayaites axonoides Spath, 1928. B. Epimayaites subtumidus (Waagen, 1875).
M. Alberti et al. / Geobios xxx (2015) xxx–xxx22
G Model
GEOBIO-701; No. of Pages 46
seem to be coarser. Other taxa, such as E. transiens or E. falcoides,are much more compressed and have much denser and finerribbing.
Epimayaites transiens (Waagen, 1875)Figs. 4(C), 24(P-T), 26(B), 27, 281875c. Stephanoceras transiens nov. sp. – Waagen, p. 111, pl. 32,
figs. 2(a, b), 3(a, b).pars 1875c. Stephanoceras opis (Sowerby) – Waagen, p. 140, pl.
36, fig. 2.1895. Macrocephalites transiens Waagen – Noetling, p. 13, pl. 10,
figs. 1, 2.1928b. E. transiens (Waagen) – Spath, p. 240, pl. 26, fig. 2, pl. 28,
fig. 3, pl. 36, fig. 11.aff. 1951. Epimayaites aff. transiens Waagen – Basse and
Perrodon, p. 60, pl. 4, fig. 5(a, b).aff. 1959b. E. transiens (Waagen) – Collignon, pl. 81, fig. 325.Lectotype: The specimen figured by Waagen (1875c: pl. 32, fig.
2(a, b)).
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Material: Seven specimens. One specimen from the DhosaOolite (GZN2010I 1093) and three specimens from the matrix ofthe Dhosa Conglomerate Bed (GZN2009II 167, 171, 220) of theJumara Dome. Two specimens from the Kanthkot Ammonite Bedssouthwest of Kantkote Village, Wagad Uplift (GZN2010I 078, 098).One stray specimen from the Tramau River, Wagad Uplift(GZN2013I 005).
Earlier records in the Kachchh Basin: E. transiens hasoriginally been described by Waagen (1875c) from the DhosaOolite member near Lodai (Habo Dome) and northwest of Soorka.The specimen described as Stephanoceras opis (Sowerby, 1840) byWaagen (1875c) and later on assigned to E. transiens by Spath(1928b) has been collected from the Dhosa Oolite member of theJhura Dome. Spath (1928b) described additional specimens fromlocalities around Kantkote Village in the Wagad Uplift.
Measurements: Table S11.Description: Very large (diameter > 260 mm), strongly com-
pressed, involute shell, septate until a diameter of 200 to 220 mm(Fig. 26(B)). Whorl section subrounded to subtrigonal with slightly
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 22. Epimayaites subtumidus (Waagen, 1875); GZN2011I 341 from the Kanthkot Ammonite Beds southwest of Kantkote Village, Wagad Uplift. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 23
G Model
GEOBIO-701; No. of Pages 46
acute venter merging into flanks along an indistinct, roundedventrolateral shoulder (Fig. 24(Q-T)). Umbilical edge prominent,rounded, umbilical wall steep. Umbilicus very narrow. Maximuminflation at about one-fourth of lateral height. The ornamentationconsists of numerous, fine, nearly straight ribs (Fig. 24(P)). Primaryribs originate at the umbilical edge and branch around the Middleof the flank into several secondaries. Intercalatory secondaries arecommon. Secondary ribs cross the venter with slight forward-directed sinuosity. Ribbing fades towards the adult body chamberwhere the shell is almost smooth.
Remarks: The specimens match the descriptions and illustra-tions of the lectotype of Epimayaites transiens by Waagen (1875c)and Spath (1928b). The species can be distinguished from relatedtaxa by its strongly compressed form, narrow umbilicus, anddense, fine ribbing. As the specimens measured by Waagen (1875c)and Spath (1928b) were slightly damaged, the dimensions given bythese authors can only be seen as a rough indication (Table S11).E. lemoini is more inflated and E. patella Spath, 1928 has a widerumbilicus. E. falcoides seems to have very similar inner whorls, but
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
the ribbing is, in general, more flexuous. Epimayaites lautus is morecoarsely and sparsely ribbed (Spath, 1928b: p. 241).
Basse and Perrodon (1951) illustrated a specimen described asEpimayaites aff. transiens from Madagascar which might well fallwithin the range of variation of the present species. In contrast,Collignon (1959b) assigned an ammonite from Madagascar toE. transiens, but this specimen is less compressed and has a widerumbilicus. In addition, its illustration points to a slightly coarserribbing.
Epimayaites lautus Spath, 1928Figs. 24(J), 29, 30(A)1928b. Epimayaites lautus nov. sp. – Spath, p. 242, pl. 45, fig. 3(a,
b).1951. Epimayaites lautus Spath – Basse and Perrodon, p. 61, pl. 4,
fig. 6.1959b. Epimayaites lautus Spath – Collignon, pl. 84, fig. 336.Holotype: The specimen figured by Spath (1928b: pl. 45, fig.
3(a, b)).
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 23. Epimayaites subtumidus (Waagen, 1875); GZN2011I 341 from the Kanthkot Ammonite Beds southwest of Kantkote Village, Wagad Uplift. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx24
G Model
GEOBIO-701; No. of Pages 46
Material: One specimen from the matrix of the DhosaConglomerate Bed of the Jara Dome (GZN2009II 166).
Earlier records in the Kachchh Basin: Epimayaites lautus hasbeen originally described from the Dhosa Oolite member of theJumara Dome (Spath, 1928b).
Measurements: Table S12.Description: Part of a small, strongly compressed, involute
phragmocone (Fig. 30(A)). Whorl section subrounded to ellipticalwith rounded venter and indistinct ventrolateral shoulders(Fig. 24(J)). Umbilical edge rounded, umbilical wall steep androunded. Umbilicus narrow. Maximum inflation at about one-thirdof lateral height. The ornamentation consists of dense, moderatelystrong, comparatively straight ribs originating at the umbilicaledge. Primary ribs slightly concave-forward on the inner part of theflank before they bifurcate at the Middle of the flank. Commonly afree secondary rib is intercalated between two pairs of secondaries.Secondary ribs cross the venter with a slight forward-directedsinuosity.
Remarks: The specimen matches the illustration and descrip-tion of E. lautus by Spath (1928b) based on the wholly septate
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
holotype with a diameter of 88 mm. The closest relative isE. transiens, but this form is compressed throughout ontogeny,whereas the innermost whorls of E. lautus are depressed (Table S12).Furthermore, E. lautus shows coarser ribbing as already pointed outby Spath (1928b: p. 242). E. falcoides has more flexuous ribs.
Epimayaites falcoides Spath, 1928Figs. 24(K-O), 30(B), 311928b. E. falcoides nov. sp. – Spath, p. 242, pl. 38, fig. 6, pl. 40, fig. 6.1931a. E. falcoides Spath – Spath, pl. 67, fig. 7(a, b).1951. E. falcoides Spath – Basse and Perrodon, p. 62, pl. 1, fig. 9.1959b. E. falcoides Spath – Collignon, pl. 49, figs. 244, 246, pl. 56,
fig. 269.Holotype: The specimen figured by Spath (1928b: pl. 40, fig. 6).Material: Sixteen specimens. One specimen from the matrix of
the Dhosa Conglomerate Bed of the Jara Dome (GZN2009II 139).One specimen from Kakindia Bet, south of Khadir Island (OUMNHJY.1162). One specimen from the Kanthkot Ammonite Bedssouthwest of Kankote Village, Wagad Uplift (GZN2010I 089).Thirteen specimens from the Kanthkot Ammonite Beds in the
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 24. Cross-sections and ribbing patterns of Epimayaites Spath, 1928. A-C. Epimayaites subtumidus (Waagen, 1875); cross-section of GZN2011I 341 at a diameter of 189 mm
(A); cross-section of GZN2010I 095 at a diameter of 97 mm (B); whorl sections of GZN2010I 114 at unknown diameter (C). D-G. Epimayaites cf. evolutus Spath, 1928; cross-
section of GZN2010I 207 at a diameter of 86 mm (D); cross-section of GZN2010I 068 at a diameter of 76 mm (E); ribbing pattern on left side of GZN2010I 068 at a diameter of
70 mm (F); ribbing pattern on right side of GZN2010I 207 at a diameter of 84 mm (G). H, I. Epimayaites smeei (Spath, 1928); ribbing pattern on right side of GZN2010I 1092 at a
diameter of 144 mm (H); cross-section of GZN2010I 1092 at a diameter of 142 mm (I). J. Epimayaites lautus Spath, 1928; cross-section of GZN2009II 166 at a diameter of
63 mm. K-O. Epimayaites falcoides Spath, 1928; cross-section of GZN2010I 160 at a diameter of 95 mm (K); cross-section of GZN2010I 166 at a diameter of 84 mm (L); cross-
section of GZN2010I 187 at a diameter of 66 mm (M); cross-section of GZN2010I 246 at a diameter of 45 mm (N); ribbing pattern on left side of GZN2010I 247 at a diameter of
49 mm (O). P-T. Epimayaites transiens (Waagen, 1875); ribbing pattern on right side of GZN2010I 1093 at a diameter of 53 mm (P); cross-section of GZN2013I 005 at a
diameter of 245 mm (Q); cross-section of GZN2009II 220 at a diameter of 173 mm (R); cross-section of GZN2009II 167 at a diameter of 73 mm (S); cross-section of GZN2010I
1093 at a diameter of 65 mm (T). U, V. Epimayaites sp. A; cross-section of GZN2010I 036 at a diameter of 57 mm (U); cross-section of GZN2009II 223 at a diameter of 92 mm
(V). Note the reduced size of the drawings. Scale bars: 50 mm (A-N, Q-V), 10 mm (O, P).
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 25
G Model
GEOBIO-701; No. of Pages 46
Tramau River, Wagad Uplift (GZN2010I 153, 160, 166, 180, 187,192, 194, 196, 221, 225, 226, 246, 247).
Earlier records in the Kachchh Basin: E. falcoides has originallybeen described from localites near Kantkote Village, Wagad Uplift(Spath, 1928b).
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Measurements: Table S13.Description: Moderately large (diameter > 120 mm), strongly
compressed, involute shell, septate until a diameter of around90 mm (Fig. 30(B)). Whorl section elliptical with flanks mergingsmoothly into rounded to slightly acute venter with indistinct
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 25. A-C. Epimayaites cf. evolutus Spath, 1928; GZN2010I 068 from the Kanthkot Ammonite Beds southwest of Kantkote Village, Wagad Uplift (A, B); GZN2010I 207 from
the Kanthkot Ammonite Beds in the Tramau River, Wagad Uplift (C). D, E. Epimayaites sp. A; GZN2009II 223 from a concretionary slab in the Dhosa Conglomerate Bed of the
Jara Dome. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx26
G Model
GEOBIO-701; No. of Pages 46
ventrolateral shoulder (Fig. 24(K-N)). Umbilical edge rounded,umbilical wall steeply inclined, umbilicus comparatively shallow.Maximum inflation slightly above the umbilical edge. Theornamentation consists of dense, fine, flexuous ribs (Fig. 24(O)).Primary ribs originate at umbilical edge, are concave-forward onthe inner part of the flank, and branch into two finer secondariesaround mid-flank. Intercalatory secondary ribs common. Thesecondaries cross the venter with a prominent forward-directedsinuosity.
Remarks: The specimens match illustrations and descriptionsof E. falcoides by Spath (1928b, 1931a). The present collection
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
includes mostly phragmocones, but there seem to be twospecimens with parts of their body chambers preserved (GZN2010I160, 166; Fig. 31(I, K)). The shell of specimen GZN2010I 187 isabnormal in being reminiscent of an ‘‘opening’’ of the shell duringthe final stage of ontogeny (Fig. 31(C)); however, this bent in thewhorl is restricted to the right side of the individual and mightreflect a repaired injury during the life time of the ammonite.
E. falcoides can be distinguished from other members of thesubfamily by its dense, flexuous ribs, the strongly compressedshell, and the prominent forward-directed sinuosity of thesecondary ribs on the venter. E. transiens is similar, but has a
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 26. Ontogenetic development of the whorl section. A. Epimayaites cf. evolutus Spath, 1928. B. Epimayaites transiens (Waagen, 1875).
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 27
G Model
GEOBIO-701; No. of Pages 46
smaller umbilicus and its secondary ribs cross the venter with onlya slight forward-directed sinuosity. E. patella Spath, 1928 describedfrom the same locality and stratigraphic unit is closely related, butdescribed as having less flexuous ribs (Spath, 1928b: p. 243). Thephotograph of the holotype and its dimensions (Table S13),however, suggest that it might fall within the range of theintraspecific variability of E. falcoides.
Epimayaites smeei (Spath, 1928) nov. comb.Figs. 4(D), 24(H, I), 30(C), 32(A, B)1928b. Mayaites smeei nov. sp. – Spath, p. 230, pl. 37, fig. 6, pl.
45, fig. 7(a, b).
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Holotype: The specimen figured by Spath (1928b: pl. 45, fig.7(a, b)).
Material: One specimen from a concretionary slab in the DhosaConglomerate Bed of the Jara Dome (GZN2010I 1092).
Earlier records in the Kachchh Basin: The species hasoriginally been described by Spath (1928b) from the Dhosa Oolitemember of Ler or Fakirwari, south of Bhuj, as well as from East ofBadi, Jhura Dome.
Measurements: Table S14.Description: Part of a large, compressed, involute phragmo-
cone (septate until a diameter of at least 170 mm; Fig. 30(C)).Whorl section subrounded, with flanks grading smoothly into
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 27. A, B. Epimayaites transiens (Waagen, 1875); GZN2009II 220 from the matrix of the Dhosa Conglomerate Bed of the Jumara Dome. Scale bar: 30 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx28
G Model
GEOBIO-701; No. of Pages 46
rounded venter (Fig. 24(I)). Umbilical edge and umbilical wallrounded, and umbilicus comparatively wide. Maximum inflationat about one-third of lateral height. The ornamentation consists ofsharp primary ribs originating around the umbilical edge with aslight forward-concave bent. Ribs cross the flank slightlyprorsiradiate and comparatively straight to slightly concave-forward (Fig. 24(H)). Primary ribs branch into two to three slightlyfiner secondaries at around the Middle of the flank. Free,intercalatory secondaries occur. Secondary ribs cross the venterwith prominent forward-directed sinuosity.
Remarks: The specimen matches the description and illustra-tion of Mayaites smeei by Spath (1928b). Nevertheless, this taxonhas to be placed within the genus Epimayaites. Members of thespecies have a compressed whorl section and a fine, elegant ribbingwith secondary ribs crossing the venter with a prominent forward-directed sinuosity. These characteristics, already mentioned bySpath (1928b), strongly support the assignment of the species tothe genus Epimayaites. In the present specimen the verycompressed inner whorls tend to become more inflated towards
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
the Middle whorls (Fig. 30(C), Table S14). Whether this is the truecharacter of the species or just a preservational bias has to beclarified with the help of further specimens, so far not available.
The closest relative of E. smeei seems to be E. lemoini, which hasa very similar ribbing pattern, but a sharper umbilical edge and asmaller umbilicus. Moreover, it is slightly more compressed.
Epimayaites sp. AFigs. 24(U, V), 25(D, E), 30(D)Material: Two specimens. One specimen from a concretionary
slab in the Dhosa Conglomerate Bed of the Jara Dome (GZN2009II223). One specimen from the southernmost tip of Khadir Island(GZN2010I 036).
Measurements: Table S15.Description: Parts of strongly depressed, involute phragmo-
cones (septate until a diameter of at least 100 mm; Fig. 30(D)).Whorl section rounded, with flanks merging smoothly into broadlyrounded venter (Fig. 24(U, V)). Umbilical edge rounded, umbilicalwall steep, umbilicus deep and narrow. Maximum inflation at
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 28. Epimayaites transiens (Waagen, 1875). A, B. GZN2010I 1093 from the Dhosa Oolite member of the Jumara Dome. C. GZN2013I 005 from the Tramau River, Wagad
Uplift. The cross denotes the approximate position of the final septum of the phragmocone. Scale bars: 20 mm (A, B), 30 mm (C).
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 29
G Model
GEOBIO-701; No. of Pages 46
about one-third of lateral height. The ornamentation consists ofcoarse, dense, almost rectiradiate ribs branching very low (at one-fourth of lateral height) into two secondary ribs of equal strength.Commonly, one free secondary is intercalated. Secondary ribs crossthe broad venter with prominent forward-directed sinuosity.
Remarks: The present specimens can be differentiated frommost of the members of the subfamily Mayaitinae by their low-branching ribs. M. rotundus has similar dimensions, but itssecondary ribs pass almost straight across the venter. M. obesus
Spath, 1928 has a smaller umbilicus. E. axonoides is morecompressed, whereas E. subtumidus is more depressed.E. pseudindicus Spath, 1928 has a wider umbilicus. The specimensdo not fit any previously described species, but due to theircomparatively poor preservation, no new taxon is defined here.
Epimayaites sp. indet.Fig. 32(C-E)Material: Twelve specimens. Two specimens from the matrix of
the Dhosa Conglomerate Bed of the Jara Dome (GZN2009II 140,
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
151). Three specimens from the Dhosa Oolite directly underneaththe Dhosa Conglomerate Bed near Jhura Camp, Jhura Dome(GZN2009II 137, 138, 185). Four specimens from the KanthkotAmmonite Beds in the Tramau River, Wagad Uplift (GZN2010I 150,164, 179, 205). Three stray specimens from the Tramau River,Wagad Uplift (GZN2010I 228, 229, 241).
Description: Small, compressed, involute nuclei of phragmo-cones. Whorl section elliptical. Umbilical edge distinct, umbilicalwall steep, umbilicus narrow. The ornamentation consists offlexuous ribs branching around the Middle of the flank intoslightly finer secondaries. Free intercalatory secondaries occur.Secondary ribs cross the venter with prominent forward-directedsinuosity.
Remarks: The present specimens represent nuclei of phrag-mocones, which are too small to be reliably identified on thespecies level. Due to their flexuous ribbing, compressed whorlsection, and forward-directed sinuosity of the ribs on the venter,they can be assigned to the genus Epimayaites, but might very wellbelong to several species.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 29. A, B. Epimayaites lautus Spath, 1928; GZN2009II 166 from the matrix of the
Dhosa Conglomerate Bed of the Jara Dome. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx30
G Model
GEOBIO-701; No. of Pages 46
Genus Dhosaites Spath, 1924Type species: Dhosaites elephantoides Spath, 1924.Remarks: The genus Dhosaites contains small to moderately-
sized ammonites characterized by a coarse, sharp, rectiradiateribbing throughout their ontogeny. Ribs pass mostly straightacross the venter. Compared to Mayaites and Epimayaites, speciesof Dhosaites have a comparatively wide umbilicus (Spath, 1924,1928b; Arkell et al., 1957).
In addition to the material used for the taxonomic descriptions,there are three more ammonites of the genus Dhosaites in thecollection, which are too poorly preserved to justify a detaileddescription. They come from the Dhosa Oolite member near JhuraCamp, Jhura Dome (GZN2010I 013), from the matrix of the DhosaConglomerate Bed near Ler, south of Bhuj (GZN2010I 1002), andfrom the Kanthkot Ammonite Beds in the Tramau River, WagadUplift (GZN2010I 170).
Dhosaites otoitoides Spath, 1928Figs. 33(A, B), 34(A), 35(A)1928b. D. otoitoides nov. sp. – Spath, p. 245, pl. 44, fig. 1(a, b).1951. D. otoitoides Spath – Basse and Perrodon, p. 65, pl. 7, figs.
7(a, b), 8, 9.1959a. D. otoitoides Spath – Collignon, pl. 39, figs. 206, 207.2013a. D. otoitoides Spath – Pandey et al., p. 142, pl. 2, fig. 1, Fig.
5(A, B), Table 2.Holotype: The specimen figured by Spath (1928b: pl. 44, fig.
1(a, b)).Material: One specimen from the southernmost tip of Khadir
Island (GZN2010I 035).Earlier records in the Kachchh Basin: Dhosaites otoitoides has
originally been described by Spath (1928b) from Samatra andFakirwari, south of Bhuj. Pandey et al. (2013a) described thespecies from the Gangta Bet.
Measurements: Table S16.Description: Fragment of a depressed, involute phragmocone
(Fig. 34(A)). Whorl section rounded, with flanks merging smoothlyinto broadly rounded venter (Fig. 35(A)). Umbilical edge roundedand indistinct, umbilical wall rounded. Umbilicus comparativelywide. Maximum inflation at about one-third of lateral height. Theornamentation consists of coarse, relatively sparse, sharp, almoststraight ribs originating at the indistinct umbilical edge. Primary
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
ribs bifurcate more or less regularly at or slightly below the Middleof the flank. Secondary ribs are of equal strength and pass straightacross the venter.
Remarks: The specimen is comparatively poorly preserved, butshows all characteristics of Dhosaites otoitoides as illustrated anddescribed by Spath (1928b). Typical for the species is thedepressed, broadly rounded whorl section, which gets more andmore compressed throughout the ontogeny (Fig. 34(A)). The innerwhorls of Dhosaites primus are generally more compressed andmore densely ribbed. D. elephantoides is more depressed and has awider umbilicus.
Dhosaites aff. primus Spath, 1928Figs. 33(C, D), 34(B), 35(B, C)aff. 1928b. D. primus nov. sp. – Spath, p. 245, pl. 32, fig. 5, pl. 36,
fig. 10(a, b), pl. 37, fig. 1(a, b), pl. 39, figs. 3, 4, pl. 40, fig. 3, pl. 44, fig.5(a, b), pl. 45, fig. 2(a, b).
aff. 1931a. D. primus Spath – Spath, pl. 49, figs. 3(a, b), 4, 13.aff. 1951. D. primus Spath – Basse and Perrodon, p. 66, pl. 1, fig. 11.aff. 2013a. D. primus Spath – Pandey et al., p. 144, pl. 2, fig. 2,
Fig. 5(C, D), Table 3.Holotype: The specimen figured by Spath (1928b: pl. 37, fig.
1(a, b)).Material: Two specimens. One specimen from the matrix of the
Dhosa Conglomerate Bed of the Jumara Dome (GZN2009II 248).One specimen from the matrix of the Dhosa Conglomerate Bed atLer, south of Bhuj (GZN2010I 1015).
Earlier records in the Kachchh Basin: Spath (1928b) describedD. primus from the Dhosa Oolite member of Fakirwari, Samatra, andWanda on the Kachchh Mainland. Pandey et al. (2013a) describedthe species from the Gangta Bet.
Measurements: Table S17.Description: Fragments of a depressed, involute phragmocone
(Fig. 34(B)). Whorl section rounded, with flanks merging smoothlyinto broadly rounded venter (Fig. 35(B, C)). Umbilical edgerounded, umbilical wall rounded. Umbilicus comparatively wide.Maximum inflation at about one-third of lateral height. Theornamentation consists of coarse, sharp, almost straight, slightlyprorsiradiate ribs originating on the umbilical wall. Primary ribsbifurcate more or less regularly at about one-third of lateral height.Ribs slightly heightened at the branching points. Secondary ribs ofequal strength, crossing the venter with very slight forward-directed sinuosity.
Remarks: The specimens match the characteristics of the genusDhosaites. The closest related form seems to be D. primus, whichhas a wider umbilicus and is more compressed (Table S17).Collignon (1959a: pl. 39, fig. 204; 1959b: pl. 83, fig. 331) illustratedfurther specimens assigned to D. primus from Madagascar, but oneof these forms is strongly compressed and might thereforerepresent another taxon. The holotype of D. primus illustratedand described by Spath (1928b) is septate only until a diameter ofaround 55 mm, pointing to a smaller adult size than that ofspecimen GZN2009II 248 which is septate until a diameter of atleast 90 mm (Fig. 34(B)).
For these reasons, the present specimens most likely representa new species of Dhosaites, but the poor state of preservationwithout a body chamber prevents a comprehensive description.Already Spath (1928b: p. 246) proposed the existence of furtherspecies of Dhosaites in the Oxfordian of the Kachchh Basin, basedon a series of small nuclei, which are highly variable in dimensionsand style of ribbing.
Genus Paryphoceras Spath, 1928Type species: Paryphoceras badiense Spath, 1928.Remarks: The genus Paryphoceras contains moderately-
sized ammonites which are strongly ribbed throughout ontogeny
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 30. Ontogenetic development of the whorl section. A. Epimayaites lautus Spath, 1928. B. Epimayaites falcoides Spath, 1928. C. Epimayaites smeei (Spath, 1928). D.Epimayaites sp. A.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 31
G Model
GEOBIO-701; No. of Pages 46
and have a compressed whorl section. The ribbing is slightlymore elegant and flexuous than in Dhosaites, and the umbilicusis wider than in Mayaites and Epimayaites. In general, ribs passthe venter with a forward-directed sinuosity (Spath, 1928b;Arkell et al., 1957; Howarth, 1998). Howarth (1998) hasincluded the taxon Prograyiceras into the synonymy of thisgenus.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Paryphoceras badiense Spath, 1928Figs. 35(D, E), 36(A, B), 371928b. P. badiense nov. sp. – Spath, p. 248, pl. 35, fig. 9, pl. 40, fig.
7(a, b).1951. P. badiense Spath – Basse and Perrodon, p. 67, pl. 6, fig. 9.1959b. P. badiense Spath – Collignon, pl. 76, fig. 315.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 31. Epimayaites falcoides Spath, 1928. A. OUMNH JY.1162 from Kakindia Bet, south of Khadir Island. B-K. Specimens from the Kanthkot Ammonite Beds in the Tramau
River, Wagad Uplift; GZN2010I 247 (B), GZN2010I 187 (C, J), GZN2010I 246 (D-F), GZN2010I 153 (G, H), GZN2010I 160 (I, K). The cross denotes the approximate position of the
final septum of the phragmocone. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx32
G Model
GEOBIO-701; No. of Pages 46
Holotype: The specimen figured by Spath (1928b: pl. 40, fig. 7).Material: One specimen from the Dhosa Oolite directly
underneath the Dhosa Conglomerate Bed of the Jara Dome(GZN2009II 141).
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Earlier records in the Kachchh Basin: Paryphoceras badiense
was originally described by Spath (1928b) from the Dhosa Oolitemember of Badi (Jhura Dome) and Fakirwari (south of Bhuj).
Measurements: Table S18.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 32. A, B. Epimayaites smeei (Spath, 1928); GZN2010I 1092 from a concretionary slab in the Dhosa Conglomerate Bed of the Jara Dome. C-E. Epimayaites sp. indet.;
GZN2009II 140 from the matrix of the Dhosa Conglomerate Bed of the Jara Dome (C); GZN2009II 137 from the Dhosa Oolite directly underneath the Dhosa Conglomerate Bed
near Jhura Camp, Jhura Dome (D, E). Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 33
G Model
GEOBIO-701; No. of Pages 46
Description: Fragment of a strongly compressed, involutephragmocone (Fig. 37). Whorl section elliptical, with flanksmerging smoothly into rounded venter (Fig. 35(D)). Umbilicaledge rounded, but distinct. Umbilical wall nearly vertical.Umbilicus narrow. Maximum inflation almost at mid-flank. Theornamentation consists of comparatively dense, coarse, sharp,biconcave ribs (Fig. 35(E)). Primary ribs originate at the umbilicalwall and cross the umbilical edge with a prominent forward-directed concavity. Ribs branch mostly into two secondary ribs ofequal strength at around mid-flank. Occasionally, a free secondary
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
rib is intercalated. Secondaries cross the venter with a pronouncedforward-directed sinuosity.
Remarks: The specimen matches the illustration and descrip-tion of Paryphoceras badiense by Spath (1928b). Spath (1928b)based the species on two illustrated specimens. The paratype isstill septate at a diameter of 70 mm, while the holotype with adiameter of 90 mm is described by Spath (1928b) as most likelyshowing the beginning of the body chamber (compare withFig. 37). Furthermore, it seems that the whorls of the holotypebegin to uncoil as is visible from Spath’s (1928b: pl. 40, fig. 7)
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 33. A, B. Dhosaites otoitoides Spath, 1928; GZN2010I 035 from the southernmost tip of Khadir Island. C, D. Dhosaites aff. primus Spath, 1928; GZN2010I 1015 from the
matrix of the Dhosa Conglomerate Bed at Ler, south of Bhuj (C); GZN2009II 248 from the matrix of the Dhosa Conglomerate Bed of the Jumara Dome (D). Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx34
G Model
GEOBIO-701; No. of Pages 46
illustration. Another specimen described by Collignon (1959b)from Madagascar has a diameter of 110 mm with no suture linesvisible on its photograph.
Paryphoceras badiense can be differentiated from other mem-bers of the genus by its dimensions, strongly biconcave ribbing onthe flank, and secondary ribs crossing the venter with apronounced forward-directed sinuosity. The latter, for example,is much weaker to absent in Paryphoceras rugosum Spath, 1928.
Fig. 34. Ontogenetic development of the whorl section. A. Dhosait
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Paryphoceras stephanoides Spath, 1928 has a less compressed whorlsection (Collignon, 1959b: pl. 85, fig. 340).
Paryphoceras tramauense (Spath, 1928)Fig. 36(E, F)1928b. Prograyiceras tramauense nov. sp. – Spath, p. 251, pl. 28,
fig. 7.1931a. Prograyiceras tramauense Spath – Spath, pl. 50, fig. 5.
es otoitoides Spath, 1928. B. Dhosaites aff. primus Spath, 1928.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 35. Cross-sections and ribbing patterns of Dhosaites Spath, 1924 and Paryphoceras Spath, 1928. A. Dhosaites otoitoides Spath, 1928; cross-section of GZN2010I 035 at a
diameter of 48 mm. B, C. Dhosaites aff. primus Spath, 1928; cross-section of GZN2009II 248 at a diameter of 90 mm (B); cross-section of GZN2010I 1015 at a diameter of 42 mm
(C). D, E. Paryphoceras badiense Spath, 1928; cross-section of GZN2009II 141 at a diameter of 68 mm (D); ribbing pattern on left side of GZN2009II 141 at a diameter of 67 mm
(E). Scale bar: 10 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 35
G Model
GEOBIO-701; No. of Pages 46
1959b. Prograyiceras sp. aff. tramauense Spath – Collignon, pl.58, fig. 274.
Holotype: The specimen figured by Spath (1928b, 1931a: pl. 28,fig. 7, and pl. 50, fig. 5).
Material: One specimen from the Kanthkot Ammonite Beds ofthe Tramau River, Wagad Uplift (GZN2010I 202).
Earlier records in the Kachchh Basin: Spath (1928b) describedthe species from the Tramau River of the Wagad Uplift.
Measurements: Table S19.Description: Fragment of a compressed, involute shell. Whorl
section elliptical, with flanks merging smoothly into acutelyrounded venter. Umbilical edge rounded, indistinct. Umbilical wallrounded, umbilicus comparatively wide. Maximum inflation ataround one-third of lateral height. The ornamentation consists ofsharp, sparse ribs originating around the umbilical edge, passingthe flank with a slight forward-directed concavity. Ribs bifurcateregularly at around mid-flank. Secondary ribs of same strength onthe flank, but thickening on the venter, which they cross withpronounced forward-directed sinuosity.
Remarks: The specimen matches the description and illustra-tion of the species by Spath (1928b, 1931a), except for a slightlyless inflated whorl section which may, however, lie within theintraspecific variability. Suture lines are barely visible, traces arerestricted to the earlier part of the fragmented whorl, whichmight therefore represent part of the body chamber. Followingthis assumption, the phragmocone would have a diameter of70 to 80 mm. This is comparable to the holotype of Spath(1928b, 1931a) which is also described as being only part of thebody chamber with a diameter of less than 120 mm. Parypho-
ceras grayi (Spath, 1924) is less compressed and more denselyribbed.
Paryphoceras sp. indet.Fig. 36(C, D)
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Material: Two specimens from the matrix of the DhosaConglomerate Bed of the Jumara Dome (GZN2009II 136, 159).
Description: Fragments of strongly compressed, involutephragmocones. Whorl section elliptical, with flanks mergingsmoothly into rounded venter. Umbilical edge sharp, umbilicalwall nearly vertical. Umbilicus narrow. Maximum inflation ataround one-third of lateral height. The ornamentation consists ofcomparatively straight, dense, and sharp ribs. Primary ribs originateat the umbilical edge with prominent forward-directed concavity.Primaries branch mostly into two secondary ribs of equal strength ataround mid-flank. Occasionally, a free secondary rib is intercalated.Secondaries cross the venter with forward-directed sinuosity.
Remarks: The specimens share the general characteristics ofthe genus Paryphoceras, but do not match any of the previouslydescribed species. The closest relative known from the area andstratigraphic unit is Paryphoceras badiense. That species hasdistinctly biconcave ribs, whereas in the present specimens theribs are comparatively straight. In addition, the ribbing inParyphoceras badiense is sparser and the forward-directed sinuosityof the ribs on the venter seems more pronounced. In contrast,Paryphoceras rugosum Spath, 1928 has secondary ribs which passnearly straight across the venter. Paryphoceras stephanoides Spath,1928 has a less compressed whorl section (Collignon, 1959b: pl. 85,fig. 340).
Members of the genus Paryphoceras Spath, 1928 are rare in thesedimentary rocks of the Kachchh Basin; all known taxa arerepresented only by very few specimens. Although the presentspecimens most likely belong to a new species, their preservationdoes not justify the description of a new taxon here.
6. Fourier shape analysis
As documented above, several species within the subfamilyMayaitinae share an overall similar morphology and/or have a
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 36. A, B. Paryphoceras badiense Spath, 1928; GZN2009II 141 from the Dhosa Oolite member directly underneath the Dhosa Conglomerate Bed of the Jara Dome. C, D.Paryphoceras sp. indet.; GZN2009II 136 from the matrix of the Dhosa Conglomerate Bed of the Jumara Dome. E, F. Paryphoceras tramauense (Spath, 1928); GZN2010I 202 from
the Kanthkot Ammonite Beds of the Tramau River, Wagad Uplift. The cross denotes the last visible septum of the phragmocone. Scale bar: 20 mm.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx36
G Model
GEOBIO-701; No. of Pages 46
Please cite this article in press as: Alberti, M., et al., Ammonites of the subfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,Western India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2015.02.005

Fig. 37. Ontogenetic development of the whorl section in Paryphoceras badiense
Spath, 1928.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 37
G Model
GEOBIO-701; No. of Pages 46
highly variable ornamentation (e.g., the ribbing pattern ofE. lemoini). The present study therefore emphasizes the importanceof the shape and dimensions of the whorl section in theidentifications as these are often characteristic of a certain species,in particular regarding its ontogenetic changes. This fact can beeasily illustrated by the species M. maya and E. polyphemus, which
Fig. 38. Comparison of ontogenetic trajectories in the shape of the whorl sections of t
particular diameters, but their ontogenetic changes are characteristic.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
share a strikingly similar whorl shape in the smooth body chamberof adult specimens but differ in the whorl section of inner whorls(Fig. 38). It is therefore mandatory for the identification of taxa toevaluate the ontogenetic trajectories of the species since thisinformation is mostly lacking in the original descriptions byWaagen (1873-1875) and Spath (1927-1933). Although theseauthors illustrated many small individuals, the identification ofsuch nuclei is complicated and not very reliable. Instead, it isnecessary to cut large individuals and examine the ontogeneticchanges within single specimens.
The Fourier shape analysis of a multispecies sample shows thatthe shape of the whorl section can indeed be used to differentiatebetween species within the subfamily Mayaitinae. Fig. 39 illus-trates the resulting plot based on 103 whorl sections at shelldiameters of more than 50 mm for all well-preserved specimens inthe collection. The first two axes spanning the morphospace,together explaining 86.1% of the overall morphometrical variationin the dataset, principally follow the ratio of whorl height to whorlwidth (PC 1) and the ratio between the height of the whorl sectionand the apertural height (PC 2), with a high apertural heighttowards negative scores on PC2. In general, the species occupywell-defined areas within the morphospace, with the depressedforms occupying the left side and the compressed forms the rightside of the diagram.
Furthermore, the ontogenetic trends illustrated in Fig. 39indicate that the growth of many species was allometric. Whorlsections at smaller diameters (juveniles) are marked by a largeapertural height compared to the whorl height. The aperturalheight shows a relative decrease along the ontogenetic trajectoriestowards the outer whorls of the ammonites. In other words, theindividuals become more and more involute during ontogeny. Thearrows indicating the ontogenetic trajectories in Fig. 39 aretherefore mostly directed upwards, with larger (older) individualsassuming progressively more positive scores on PC2. The arrowsare, however, not merely vertical, but also follow characteristicchanges in compression for many species. Those taxa with a verycompressed whorl section tend to become less compressed duringontogeny (e.g., E. transiens, E. lemoini, and E. smeei). In these casesthe ontogenetic trajectories point towards the upper left corner of
hree mayaitins. The whorl sections of different species can appear very similar at
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 39. Principal component analysis of Fourier coefficients describing the whorl shapes of different ontogenetic stages in 17 species in the subfamily Mayaitinae Spath, 1928,
based on 103 whorl sections at diameters above 50 mm. Most taxa occupy distinct fields within the morphospace and show characteristic ontogenetic trajectories (grey
arrows). Species with similar whorl sections and consequently overlapping fields can be differentiated by their size and/or ribbing pattern.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx38
G Model
GEOBIO-701; No. of Pages 46
the morphospace spanned by PC1 and PC2. In contrast, depressedforms become either more compressed (M. maya; arrow pointstowards the upper right corner) or change only very little(E. subtumidus, M. rotundus; arrow much steeper). The ratio ofwhorl height to whorl width (PC1) and the ratio between theheight of the whorl section and the apertural height (PC2)therefore show an interdependence possibly explaining theslightly ‘‘arch’’-shaped distribution of the points in Fig. 39.
Other factors overprint these two main characters of whorlshape, such as the roundness of the venter with outer whorls beingcommonly more acute than inner ones and thereby occasionallyturning almost triangular (e.g., E. transiens in Fig. 39). In any case,the interdependence of different parameters in the ammoniteshells is striking and also includes the ornamentation. Buckman’sfirst law of covariation states that the coarseness of ribs and otherornamentation in ammonite shells correlates negatively with thedegree of compression and involution (Westermann, 1966; Guexet al., 2003; Hammer and Bucher, 2005; Moulton et al., 2015). Thistrend can also be seen in the present mayaitins, in which theindividuals become more and more involute during ontogenywhile the ribbing fades and the distance between ribs increasesuntil the outermost whorls are almost smooth (e.g., E. lemoini,Fig. 19(B); E. transiens, Fig. 28(C)). Most recently, Moulton et al.(2015) proposed a morpho-mechanical model for the development
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
of ammonite shells, which explains the principles of Buckman’slaw and might also account for the covariation found in mayaitins.
Some of the morphospecies illustrated in Fig. 39 show rathersimilar whorl sections leading to overlapping distributions in thefirst principal plane of the morphospace. The characteristics usedin discriminating these species are briefly reiterated in thefollowing. Both M. rotundus and Epimayaites sp. A have depressedwhorl sections and a smoothly rounded venter, but can bedifferentiated by their ribbing which is very low-branching inEpimayaites sp. A. Furthermore, the secondary ribs pass the venterstraight in M. rotundus, whereas in Epimayaites sp. A they do thiswith a slight forward-directed sinuosity. The strongest overlapsexist in the field of M. maya (Fig. 39), which shows similarities withMayaites aff. maya, M. jumarensis, and E. axonoides. Still, Mayaites
aff. maya can be well differentiated by its more rounded venter andmore depressed whorl section, while M. jumarensis shows astronger ribbing and a smaller umbilicus. E. axonoides has a moreflexuous ribbing with secondary ribs crossing the venter with aprominent forward-inclined sinuosity absent in the species ofMayaites. In addition, the ontogenetic trajectories of M. maya andE. polyphemus illustrate very well the trends already describedabove and shown in Fig. 38. While the inner whorls show stronglydifferent whorl sections, the outer whorls are very similar, asindicated by converging ontogenetic trajectories (Fig. 39).
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 40. Summary of the main diagnostic features of the ten most abundant Mayaitinae Spath, 1928 in the Kachchh Basin.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 39
G Model
GEOBIO-701; No. of Pages 46
Epimayaites cf. evolutus differs from E. polyphemus by its largerumbilicus and slightly more compressed whorl section. E. falcoides
can be differentiated from E. transiens by its much more flexuousribbing as well as its smaller size. E. lautus is more coarsely ribbedand has more depressed innermost whorls than E. transiens. Themain characteristic features of the most abundant species of thesubfamily Mayaitinae present in the Kachchh Basin have beensummarized in Fig. 40.
7. Biostratigraphy
The current study comprises 100 specimens representing19 morphospecies of the subfamily Mayaitinae from localitiesthroughout the Kachchh Basin (Figs. 41–43; for detailed sections of
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
most of the localities, see Pandey et al., 2012a; Fursich et al., 2013;Alberti et al., 2013a). The oldest members of the subfamily presentin the collection are specimens of M. jumarensis and E. transiens
from the Dhosa Oolite member, more than 20 cm below the DhosaConglomerate Bed (interval A of Fig. 44). The occurrence ofPeltoceratoides (Parawedekindia) arduennensis in the same horizonsindicates an Early Oxfordian age (Cordatum Zone) for these rocks(Alberti et al., 2011). Additionally, a series of perisphinctids andtwo species of Euaspidoceras have been recorded. The next unitwith now abundant mayaitins is the Dhosa Oolite member directlybelow the Dhosa Conglomerate Bed containing P. badiense,M. rotundus, and E. lemoini (interval B of Fig. 44). Again, thesespecies occur together with members of Peltoceratoides (e.g., P. (P.)constantii, P. (Parawedekindia) arduennensis), indicating an Early
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 41. Lithologs from localities in the western Kachchh Mainland yielding specimens of the subfamily Mayaitinae Spath, 1928, with the stratigraphic range of the taxa.
Numbers below stratigraphic ranges indicate the number of specimens collected from the interval. See Fig. 43 for legend.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx40
G Model
GEOBIO-701; No. of Pages 46
Oxfordian age (Cordatum Zone; Alberti et al., 2011). Additional co-occurring taxa in these horizons are perisphinctids and four speciesof Euaspidoceras.
The Dhosa Conglomerate Bed itself is a highly complex unitconsisting of concretionary slabs floating within a ferruginous,micritic matrix. Its complicated genetic history has been inter-preted by several authors (Singh, 1989; Fursich et al., 1992; Albertiet al., 2013a). The ammonite fauna collected from the concre-tionary slabs (interval C of Fig. 44) strongly resembles that of theunderlying Dhosa Oolite and can also be assigned to the CordatumZone (Lower Oxfordian) due to the presence of Peltoceratoides
(Parawedekindia) cf. arduennensis (Alberti et al., 2011). Mayaitinsfrom these slabs include M. subkobyi, E. smeei, and Epimayaites sp.A. The ammonites within the matrix of the Dhosa ConglomerateBed (interval D of Fig. 44) are mostly reworked and belong toseveral ammonite zones (Lower to Middle Oxfordian; Cordatum toTransversarium zones). Characteristic taxa occurring in this unitare Peltoceratoides (Peltoceratoides) constantii, P. (Parawedekindia)cf. arduennensis, Perisphinctes sp. cf. P. (Dichotomosphinctes)antecedens, P. (D.) subhelenae, and P. (D.) cf. tizianiformis
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
(Alberti et al., 2011; Pandey et al., 2012a). This level also yieldeda large number of mayaitins belonging to M. maya, M. aff. maya,M. rotundus, E. polyphemus, E. lemoini, E. transiens, E. lautus,E. falcoides, and Dhosaites aff. primus. A special situation exists inthe outcrops in the vicinity of Rudra Mata in the western part of theHabo Dome, where a thin layer of seemingly laterally continuousoolitic sandstone covers the Dhosa Conglomerate Bed; itsammonite fauna, however, points to an Early Oxfordian age(Alberti et al., 2011), meaning, that the sandstone is indeed olderthan the conglomeratic matrix below. Actually, the matrix fills onlycrevices below the sandstone, which therefore can be correlated inage with the concretionary slabs otherwise floating in the DhosaConglomerate Bed.
Reworked ammonites of the Lower to Middle Oxfordian(Cordatum to Transversarium zones) have also been found at asection at the southernmost tip of Khadir Island (interval E of Fig. 44).These rocks continue south under the cover of the recent sedimentsof the Rann of Kachchh, but crop out once more on the small isletKakindia Bet, where the Late John H. Callomon collected specimenOUMNH JY.1162 described herein as E. falcoides. Although the
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 42. Lithologs from localities in the eastern Kachchh Mainland and southern Khadir Island yielding specimens of the subfamily Mayaitinae Spath, 1928, with the
stratigraphic range of the taxa. Numbers below stratigraphic ranges indicate the number of specimens collected from the interval. See Fig. 43 for legend.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 41
G Model
GEOBIO-701; No. of Pages 46
present authors did not measure a section on Kakindia Bet, theapproximate stratigraphic level is comparable to that at thesouthernmost tip of Khadir Island (Fig. 44). Apart from mayaitins,these horizons contain Peltoceratoides (Peltoceratoides) eugeniforme
with an Early Oxfordian age and several perisphinctids known fromthe Dhosa Conglomerate Bed of the Kachchh Mainland or from theMiddle Oxfordian of the Wagad Uplift (e.g., Perisphinctes (Peri-
Perisphinctes) indogermanus, P. (Kranaosphinctes) nerunaensis, P.(Dichotomosphinctes) cf. obliqueplicatus; Alberti et al., 2011; Pandeyet al., 2012a).
The Kanthkot Ammonite Beds of the Wagad Uplift in easternKachchh have been divided into three intervals (F-H of Fig. 44)based on perisphinctids (Pandey et al., 2012a). The lowermostinterval F has been assigned to the basal Plicatilis Zone, butcontains only few perisphinctids and no mayaitins (Pandey et al.,2012a). The presence of Perisphinctes (Perisphinctes) martelli, P.(Arisphinctes) cf. cotovuiformis, P. (Liosphinctes) plicatilis, P. (Dichot-
Dichotomosphinctes) tizianiformis, and P. (D.) subhelenae ininterval G indicates a Middle Oxfordian age (Plicatilis toTransversarium zones; Pandey et al., 2012a). The uppermost
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
levels of the Kanthkot Ammonite Beds (interval G) contain manyperisphinctids, some of which typical of the Upper Oxfordian(Stenocycloides Subzone, Bifurcatus Zone), including Perisphinctes
(Dichotomosphinctes) elisabethae, P. (Dichotomoceras) cf. bifurcatus,P. (D.) gr. crassus, P. (D.) cf. dichotomus, and most notably P. (D.)stenocycloides. Following on top of the Kanthkot Ammonite Beds isthe Patasar Shale member (interval I of Fig. 44) which so far yieldedonly very few perisphinctids, including Perisphinctes (Dichotomo-
ceras) cf. besairiei.Two of the intervals in the Kanthkot Ammonite Beds contain
numerous mayaitins (Fig. 44), but members of the genusMayaites have been found in situ only in the Lower OxfordianDhosa Oolite member of the Kachchh Mainland. The Middle andUpper Oxfordian might therefore lack Mayaites and insteadcontain only species of Epimayaites (most notably E. transiens,E. lemoini, E. falcoides, E. axonoides, and E. subtumidus) andParyphoceras. A single, poorly preserved specimen of Dhosaites
from the Kanthkot Ammonite Beds in the Tramau River, WagadUplift, extends the range of this genus into the MiddleOxfordian.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 43. Lithologs from localities in the Wagad Uplift yielding specimens of the subfamily Mayaitinae Spath, 1928, with the stratigraphic range of the taxa. Numbers below
stratigraphic ranges indicate the number of specimens collected from the interval.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx42
G Model
GEOBIO-701; No. of Pages 46
The ammonite fauna of the small isolated islet Gangta Bet,which consists mainly of Upper Oxfordian rocks, contains threemayaitins: Dhosaites otoitoides, D. primus, and Paryphoceras cf.crassum (Pandey et al., 2013a). The two Dhosaites species comefrom beds relatively low in the succession of the islet (interval J ofFig. 44). Except for a poorly preserved specimen of Euaspidoceras
sp., no further identifiable ammonites have been collected fromthese horizons up to now, but Patel et al. (2012) have described afew (albeit quite doubtful; Kanjilal, 2014) specimens of Mayaites
and Peltoceratoides from the Gangta Bet, the latter being indicativeof the Lower Oxfordian (Alberti et al., 2011). However, it cannot beexcluded that interval J also encompasses the Middle Oxfordian(compare Pandey et al., 2013a). Paryphoceras cf. crassum occurstogether with a series of perisphinctids indicative of the UpperOxfordian (Grossouvrei Subzone, Bifurcatus Zone; interval L ofFig. 44), including Perisphinctes (Dichotomoceras) bifurcatoides, P.(D.) cf. dichotomus, and most notably P. (D.) grossouvrei (Pandeyet al., 2013a).
In most studies dealing with the biostratigraphy of the KachchhBasin, species of the Mayaitinae have been rather neglected due totheir presumed long-ranging nature. The present collection
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
corroborates the long range for several taxa occurring from theLower to Upper Oxfordian (E. transiens, E. lemoini), but it alsoidentifies a few taxa, which might indeed be indicative of a certainammonite zone. As already mentioned above, species of Mayaites
seem to be restricted to the Lower Oxfordian (Cordatum Zone), eventhough they occur reworked together with taxa from the Plicatilisand Transversarium zones in the Dhosa Conglomerate Bed. Somepresently described species have been collected only as singlespecimens which do not allow an in-depth statement on theirstratigraphic distribution at the moment, but the common occur-rence of E. subtumidus in the Upper Oxfordian (Bifurcatus Zone;Stenocycloides Subzone) might indicate its usefulness as an indexfossil for the Kachchh Basin and neighbouring areas. E. axonoides
might be at least indicative of the Middle to Upper Oxfordian,occurring at several levels within the Kanthkot Ammonite Beds.
The uppermost Oxfordian is missing on the Kachchh Mainlandand is largely unfossiliferous in the Wagad Uplift. Only on GangtaBet the upper Bifurcatus Zone (Grossouvrei Subzone) is repre-sented by a very rich ammonite fauna (Pandey et al., 2013a).However, in a collection of almost 150 specimens from these units,only one member of the Mayaitinae has been found to belong to
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

Fig. 44. Comparison of the stratigraphic occurrences of mayaitins, perisphinctids, and peltoceratids collected during four recent field surveys (2008-2013) of the Oxfordian
succession of the Kachchh Basin (data on perisphinctids and peltoceratids from Alberti et al., 2011; Pandey et al., 2012a, 2013a, 2015). Data are presented separately for the
Kachchh Mainland, S-Khadir & Kakindia Bet, the Wagad Uplift, and the Gangta Bet. Levels with predominantly reworked ammonites (D, E) are shaded in grey. Oxf.: Oxfordian;
Pli.: Plicatilis Zone; Transv.: Transversarium Zone.
M. Alberti et al. / Geobios xxx (2015) xxx–xxx 43
G Model
GEOBIO-701; No. of Pages 46
Please cite this article in press as: Alberti, M., et al., Ammonites of the subfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,Western India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2015.02.005

M. Alberti et al. / Geobios xxx (2015) xxx–xxx44
G Model
GEOBIO-701; No. of Pages 46
Paryphoceras cf. crassum. This might mark the gradual disappear-ance of mayaitins in the Kachchh Basin before the end of theOxfordian.
8. Conclusions
Although Jurassic ammonites of the Kachchh Basin, WesternIndia, have been studied for more than a century, the presentanalysis of members of the subfamily Mayaitinae Spath, 1928,shows that there is still room to further deepen our understandingof this fossil cephalopod fauna. The Mayaitinae show a geographicoccurrence restricted to the southern Tethys and are nowadaysfound mainly in East Africa, Madagascar, India, and New Guinea.Isolated records further afield are probably incorrect identifica-tions or drifted shells. Although the group and its genera hadoriginally been defined based on material from the Kachchh Basin,the subfamily was largely neglected over the following decades.This is mainly due to their limited use in biostratigraphy caused bytheir restricted palaeogeographic distribution as well as by thelong ranges of its members. Nevertheless, the current collectionindicates that a few taxa can be used for intra- and inter-basinalcorrelations as described above. The present study revises theoriginal species descriptions by Waagen (1873-1875) and Spath(1927-1933) by identifying maximum phragmocone diameters formost of the taxa as well as recognizing the potential of the shape ofthe whorl section and its ontogenetic changes as diagnosticfeatures for most species using a Fourier shape analysis.
Acknowledgements
The Late John H. Callomon shared his knowledge on Jurassicammonites with the authors. Birgit Leipner-Mata prepared thefossils, and Marie-Luise Neufert and Lisa Eggert took thephotographs. Christian Schulbert scanned some of the ammonitetransects. During the field surveys in the Kachchh Basin thefollowing persons offered their assistance in various ways: Mr. P.H.Bhatti and family, Narendra Swami, Mr. M.G. Thakkar, KeyurPandya, Kaustubh Sane, Ahesan Sheliya, and Valsamma Fursich.Financial support has been granted by the German ResearchSociety (FU 131/34-1). D.K. Pandey gratefully acknowledgesfinancial support by the Alexander von Humboldt Foundation.The authors thank the editors Gilles Escarguel and StephaneReboulet for their support. The manuscript benefitted fromreviews by Kevin Page, Subhendu Bardhan, and Jacques Thierry.
Appendix A. Supplementary data
Supplementary data (Tables S1-S19) associated with this articlecan be found, in the online version, at http://dx.doi.org/10.1016/j.geobios.2015.02.005.
References
Alberti, M., Fursich, F.T., Pandey, D.K., 2013a. Deciphering condensed sequences: acase study from the Oxfordian (Upper Jurassic) Dhosa Oolite member of theKachchh Basin, western India. Sedimentology 60, 574–598.
Alberti, M., Pandey, D.K., Fursich, F.T., 2011. Ammonites of the genus PeltoceratoidesSpath, 1924 from the Oxfordian of Kachchh, western India. Neues Jahrbuch furGeologie und Palaontologie, Abhandlungen 262, 1–18.
Alberti, M., Nutzel, A., Fursich, F.T., Pandey, D.K., 2013b. Oxfordian (Late Jurassic)gastropods from the Kachchh Basin, western India. Neues Jahrbuch fur Geologieund Palaontologie, Abhandlungen 270, 275–300.
Arkell, W.J., 1956. Jurassic geology of the world. Oliver & Boyd, Edinburgh.Arkell, W.J., Kummel, B., Wright, C.W., 1957. Mesozoic Ammonoidea. In: Moore, R.C.
(Ed.), Treatise on Invertebrate Paleontology. Part L, Mollusca 4, Cephalopoda,Ammonoidea. The Geological Society of America and the University of KansasPress, New York and Lawrence, pp. 80–437.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Basse, E., Perrodon, M., 1951. Macrocephalitides du sud-ouest de Madagascar:Macrocephalitidae, Eucycloceratidae, Mayaitidae. Recoltes de la mission E.Basse 1930-31, t. 3065, Memoires de la Societe Geologique de France, pp. 1–100.
Biswas, S.K., 1982. Rift basins in the western margin of India and their hydrocarbonprospects with special reference to Kutch basin. American Association ofPetroleum Geologists, Bulletin 66, 1497–1513.
Biswas, S.K., 1991. Stratigraphy and sedimentary evolution of the Mesozoic basin ofKutch, western India. In: Tandon, S.K., Pant, C.C., Casshyap, S.M. (Eds.), Stratig-raphy and sedimentary evolution of Western India. Gyanodaya Prakashan,Nainital, pp. 74–103.
Biswas, S.K., 1993. Geology of Kutch, Volume1. K.D. Malaviya Institute of PetroleumExploration, Dehradun.
Blanford, W.T., 1869. On the geology of a portion of Cutch. Memoirs of theGeological Survey of India 6, 17–39.
Braun, E., Jordan, R., 1976. The stratigraphy and paleontology of the Mesozoicsequence in the Mae Sot area in western Thailand. Geologisches Jahrbuch B 21,5–51.
Brochwicz-Lewinski, W., Rozak, Z., 1976. Some difficulties in recognition of sexualdimorphism in Jurassic perisphinctids (Ammonoidea). Acta PalaeontologicaPolonica 21, 115–124.
Brookfield, M.E., Westermann, G.E.G., 1982. Mesozoic ammonites from the SpongValley, Zanskar, N.W. India. Journal of the Geological Society of India 23, 263–266.
Callomon, J.H., 1963. Sexual dimorphism in Jurassic ammonites. Transactions of theLeicester Literary and Philosophical Society 57, 21–56.
Chatterjee, T.K., 1990. The systematics of the ammonoid fauna from the Callovian-Tithonian sequence of Jaisalmer, Rajasthan, and their significance in biostra-tigraphy (Ph.D. thesis). Indian School of Mines, Dhanbad (unpublished).
Collignon, M., 1959a. Atlas des fossils caracteristiques de Madagascar. Fascicule III(Oxfordien), Pls. 34-46. Service geologique, Tananarive.
Collignon, M., 1959b. Atlas des fossils caracteristiques de Madagascar. Fascicule IV(Argovien-Rauracien), Pls. 47-95. Service geologique, Tananarive.
Crampton, J.S., Haines, A.J., 1996. Users’ manual for programs HANGLE, HMATCH, andHCURVEfor the Fourier shape analysis of two-dimensional outlines. Institute ofGeological & Nuclear Sciences, Science Report 96/37, 1–28.
Datta, K., Bhaumik, D., Jana, S.K., Bardhan, S., 1996. Age, ontogeny and dimorphismof Macrocephalites triangularis Spath – the oldest macrocephalitid ammonitefrom Kutch, India. Journal of the Geological Society of India 47, 447–458.
Donovan, D.T., Callomon, J.H., Howarth, M.K., 1981. Classification of the JurassicAmmonitina. In: House, M.R., Senior, J.R. (Eds.), The Ammonoidea - Theevolution, classification, mode of life and geological usefulness of a major fossilgroup, Special Volume 18, The Systematics Association, pp. 101–155.
Enay, R., 2009. Les faunes d’ammonites de l’Oxfordien au Tithonien et la biostrati-graphie des Spiti-Shales (Callovien superieur-Tithonien) de Thakkhola, NepalCentral. Documents des Laboratoires de Geologie Lyon 166, 1–351.
Enay, R., Cariou, E., 1997. Ammonite faunas and palaeobiogeography of the Hima-layan belt during the Jurassic: initiation of a Late Jurassic austral ammonitefauna. Palaeogeography, Palaeoclimatology, Palaeoecology 134, 1–38.
Enay, R., Cariou, E., 1999. Jurassic ammonite faunas from Nepal and their bearing onthe palaeobiogeography of the Himalayan belt. Journal of Asian Earth Sciences17, 829–848.
Fursich, F.T., 1998. Environmental distribution of trace fossils in the Jurassic ofKachchh (western India). Facies 39, 243–272.
Fursich, F.T., Alberti, M., Pandey, D.K., 2013. Stratigraphy and palaeoenvironmentsof the Jurassic rocks of Kachchh – Field Guide. Beringeria (7), 1–174.
Fursich, F.T., Callomon, J.H., Pandey, D.K., Jaitly, A.K., 2004a. Environments andfaunal patterns in the Kachchh rift basin, western India, during the Jurassic.Rivista Italiana di Paleontologia e Stratigrafia 110, 181–190.
Fursich, F.T., Oschmann, W., Singh, I.B., Jaitly, A.K., 1992. Hardgrounds, reworkedconcretion levels and condensed horizons in the Jurassic of western India: theirsignificance for basin analysis. Journal of the Geological Society 149, 313–331.
Fursich, F.T., Oschmann, W., Pandey, D.K., Jaitly, A.K., Singh, I.B., Liu, C., 2004b.Palaeoecology of middle to lower Upper Jurassic macrofaunas of the KachchhBasin, western India: an overview. Journal of the Palaeontological Society ofIndia 49, 1–26.
Futterer, K., 1894. Beitrage zur Kenntniss des Jura in Ost-Afrika. Zeitschrift derDeutschen geologischen Gesellschaft 46, 1–48.
Gingerich, P.D., 1985. Species in the fossil record: concepts, trends, and transitions.Paleobiology 11, 27–41.
Gradstein, F.M., Gibling, M.R., Sarti, M., von Rad, U., Thurow, J.W., Ogg, J.G., Jansa, L.F.,Kaminski, M.A., Westermann, G.E.G., 1991. Mesozoic Tethyan strata of Thak-khola, Nepal: evidence for the drift and breakup of Gondwana. Palaeogeogra-phy, Palaeoclimatology, Palaeoecology 88, 193–218.
Grant, C.W., 1837. Memoir to illustrate a Geological Map of Cutch. Transactions ofthe Geological Society of London second series 5, 289–329.
Guex, J., Koch, A., O’Dogherty, L., Bucher, H., 2003. A morphogenetic explanation ofBuckman’s law of covariation. Bulletin de la Societe Geologique de France 174,603–606.
Haines, A.J., Crampton, J.S., 2000. Improvements to the method of Fourier shapeanalysis as applied in morphometric studies. Palaeontology 43, 765–783.
Hammer, Ø., Bucher, H., 2005. Buckman’s first law of covariation – a case ofproportionality. Lethaia 38, 67–72.
Hammer, Ø., Harper, D.A.T., 2006. Paleontological data analysis. Blackwell Publish-ing, Malden.
Hoffmann, R., Keupp, H., 2008. A giant member of the genus Lobolytoceras Buckman,1923 (Ammonitina, Lytoceratinae) from the Oxfordian of SW Madagascar.Neues Jahrbuch fur Geologie und Palaontologie, Abhandlungen 250, 53–64.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

M. Alberti et al. / Geobios xxx (2015) xxx–xxx 45
G Model
GEOBIO-701; No. of Pages 46
Howarth, M.K., 1998. Ammonites and nautiloids from the Jurassic and LowerCretaceous of Wadi Hajar, southern Yemen. Bulletin of the Natural HistoryMuseum, Geology Series 54, 33–107.
Howarth, M.K., Morris, N.J., 1998. The Jurassic and Lower Cretaceous of Wadi Hajar,southern Yemen. Bulletin of the Natural History Museum, Geology Series 54, 1–32.
Hudson, N., 1999. The middle Jurassic of New Zealand – A study of the Lithostrati-graphy and Biostratigraphy of the Ururoan, Temaikan and Lower HeterianStages (?Pliensbachian to?Kimmeridgian). (Ph.D. thesis). University of Auck-land (unpublished).
Hudson, N., Grant-Mackie, J.A., Helby, R., 1987. Closure of the New Zealand ‘‘MiddleJurassic Hiatus’’? Search 18, 146–148.
Jana, S.K., Bardhan, S., Halder, K., 2005. Eucycloceratin ammonites from the Callo-vian Chari Formation, Kutch, India. Palaeontology 48, 883–924.
Kachhara, R.P., Jodhawat, R.L., 1981. On the age of Jaisalmer Formation, Rajasthan,India. In: Proceedings of IX Indian Colloquium on Micropalaeontology andStratigraphy, Udaipur, pp. 235–247.
Kanjilal, S., 2014. Revisiting the proposed ammonite zonation of Jurassic rocks ofGangta Bet, Wagad, Eastern Kachchh. Journal of the Palaeontological Society ofIndia 59, 103–107.
Kowalewski, M., Dyreson, E., Marcot, J.D., Vargas, J.A., Flessa, K.W., Hallman, D.P.,1997. Phenetic discrimination of biometric simpletons: paleobiological impli-cations of morphospecies in the lingulide brachiopod Glottidia. Paleobiology 23,444–469.
Krishna, J., 1987. An overview of the Mesozoic stratigraphy of Kachchh andJaisalmer basins. Journal of the Palaeontological Society of India 32, 136–149.
Krishna, J., Pathak, D.B., 1993. Late Lower Kimmeridgian–Lower Tithonian virgato-sphinctins of India: Evolutionary succession and biogeographic implications.Geobios 15, 227–238.
Krishna, J., Pandey, B., Ojha, J.R., 2009a. Gregoryceras in the Oxfordian of Kachchh(India): Diverse eventful implications. Geobios 42, 197–208.
Krishna, J., Pandey, B., Pathak, D.B., 2009b. Characterization of Dichotomoceras in theOxfordian of Kachchh. Journal of the Geological Society of India 74, 469–479.
Krishna, J., Pathak, D.B., Pandey, B., 1996. Quantum refinement in the Kimmeridgianammonoid chronology in Kachchh (India). GeoResearch Forum 1-2, 195–204.
Krishna, J., Pathak, D.B., Pandey, B., 1998. Development of Oxfordian (Early UpperJurassic) in the most proximally exposed part of the Kachchh Basin at Wagadoutside the Kachchh Mainland. Journal of the Geological Society of India 52,513–522.
Krishna, J., Pathak, D.B., Pandey, B., Ojha, J.R., 2000. Transgressive sediment intervalsin the Late Jurassic of Kachchh, India. GeoResearch Forum 6, 321–332.
Krishna, J., Pandey, B., Ojha, J.R., Pathak, D.B., 2009c. Reappraisal of the ageframework, correlation, environment and nomenclature of Kachchh Mesozoiclithostratigraphic units in Wagad. Journal of Scientific Research, Banares HinduUniversity, Varanasi 53, 1–20.
Kuhl, F.P., Giardina, C.R., 1982. Elliptic Fourier features of a closed contour. Com-puter Graphics and Image Processing 18, 236–258.
Lemoine, P., 1911. Ammonites du Jurassique de superieur du cercle d’Analalava(Madagascar). Annales de Paleontologie 6, 45–64.
Makowski, H., 1962. Problem of sexual dimorphism in ammonites. PalaeontologiaPolonica 12, 1–92.
Makowski, H., 1971. Some remarks on the ontogenetic development and sexualdimorphism in the Ammonoidea. Acta Geologica Polonica 21, 321–340.
Moulton, D.E., Goriely, A., Chirat, R., 2015. The morpho-mechanical basis of ammo-nite form. Journal of Theoretical Biology 364, 220–230.
Noetling, F., 1895. The fauna of the Kellaways of Mazar Drik. Memoirs of theGeological Survey of India, Palaeontologia Indica, series 16 1, 1–22.
Pandey, B., Pathak, D.B., Krishna, J., 2013c. Preliminary remarks on new ammonoidcollection from freshly exposed succession of the Spiti Formation betweenLidang and Giumal, Spiti Valley, Himachal Himalaya, India. Himalayan Geology34, 124–134.
Pandey, D.K., Fursich, F.T., 1998. Distribution and succession of Jurassic rocks inGora Dongar, Pachchham ‘‘Island’’, Kachchh, India. Journal of the GeologicalSociety of India 51, 331–344.
Pandey, D.K., Fursich, F.T., 2001. Environmental distribution of scleractinian coralsin the Jurassic of Kachchh, western India. Journal of the Geological Society ofIndia 57, 479–495.
Pandey, D.K., Alberti, M., Fursich, F.T., 2012a. Ammonites of the genus PerisphinctesWaagen, 1869 from the Oxfordian of Kachchh, western India. Revue de Paleo-biologie 31, 483–587.
Pandey, D.K., Alberti, M., Fursich, F.T., 2013a. Ammonites from the Oxfordian(Bifurcatus Zone) strata of Gangta Bet, Kachchh, western India. Journal of thePalaeontological Society of India 58, 139–174.
Pandey, D.K., Alberti, M., Fursich, F.T., 2015. Ammonites of the genera PeltocerasWaagen, 1871, Metapeltoceras Spath, 1931, and Euaspidoceras Spath, 1931 fromthe Upper Callovian and Oxfordian of Kachchh, western India, and theirbiostratigraphic potential. Journal of the Palaeontological Society of India.
Pandey, D.K., Fursich, F.T., Alberti, M., 2014. Stratigraphy and palaeoenvironmentsof the Jurassic rocks of the Jaisalmer Basin – Field Guide. Beringeria (9), 1–111.
Pandey, D.K., Alberti, M., Fursich, F.T., Głowniak, E., Oloriz, F., 2013b. Ammonitesfrom the Oxfordian-Kimmeridgian boundary and the Lower-Upper Kimmer-idgian of Kachchh, western India. Volumina Jurassica 11, 97–146.
Pandey, D.K., Choudhary, S., Bahadur, T., Swami, N., Poonia, D., Sha, J., 2012b. Areview of the Lower-lowermost Upper Jurassic facies and stratigraphy of theJaisalmer Basin, western Rajasthan, India. Volumina Jurassica 10, 61–82.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Parent, H., 2006. Oxfordian and Late Callovian ammonite faunas and biostratig-raphy of the Neuquen-Mendoza and Tarapaca basins (Jurassic, Ammonoidea,western South-America). Boletın del Instituto de Fisiografıa y Geologıa 76,1–70.
Parent, H., Schweigert, G., Melendez, G., 2006. Oxfordian perisphinctid ammonitesfrom Chacay Melehue, Argentina. Palaontologische Zeitschrift 80, 307–324.
Patel, S.J., Joshi, P.N., Joseph, J.K., 2012. Ammonite zonation of the Jurassic rocks ofthe Gangta Bet area, Wagad region, eastern Kachchh, India. Journal of thePalaeontological Society of India 57, 129–133.
Prasad, S., 2006. Ammonite biostratigraphy of Middle to Late Jurassic rocks ofJaisalmer Basin, Rajasthan, India. Memoirs of the Geological Survey of India 52,1–146.
Rai, J., Jain, S., 2013. Pliensbachian nannofossils from Kachchh: Implications on theearliest Jurassic transgressive event on the western Indian margin. Zitteliana 53,105–120.
Reyment, R.A., 2008. A review of the post-mortem dispersal of cephalopod shells.Palaeontologia Electronica 11 (12A.), 1–13.
Riccardi, A.C., 1991. Jurassic and Cretaceous marine connections between theSoutheast Pacific and Tethys. Palaeogeography, Palaeoclimatology, Palaeoecol-ogy 87, 155–189.
Riccardi, A.C., 2008. The marine Jurassic of Argentina: a biostratigraphic framework.Episodes 31, 326–335.
Roy, A., Bardhan, S., Das, S., Mondal, S., Mallick, S., 2012. Systematic revision andpalaeobiogeography of Perisphinctes Waagen (Ammonoidea) from the Oxfor-dian of Kutch, India: Stratigraphic and evolutionary implications. Palaeoworld21, 167–192.
Sato, T., Westermann, G.E.G., 1991. 4. Japan and South-East Asia. In: Westermann,G.E.G., Riccardi, R. (Eds.), Jurassic taxa ranges and correlation charts for thecircum Pacific, 24, Newsletters on Stratigraphy, pp. 81–108.
Scherzinger, A., Schweigert, G., Parent, H., 2006. New considerations on dimorphismand aptychus in Gravesia Salfeld (Ammonoidea; Late Jurassic). Neues Jahrbuchfur Geologie und Palaontologie, Abhandlungen 241, 269–286.
Singh, I.B., 1989. Dhosa Oolite – A transgressive condensation horizon of Oxfordianage in Kachchh, Western India. Journal of the Geological Society of India 34,152–160.
Singh, C.S.P., Jaitly, A.K., Pandey, D.K., 1982. First report of some Bajocian-Bathonian(Middle Jurassic) ammonoids and the age of the oldest sediments fromKachchh, W. India. Newsletters on Stratigraphy 11, 37–40.
Smith, A.B., 1994. Systematics and the fossil record: documenting evolutionarypatterns. Blackwell Science, London.
Sowerby, J. de C., 1840. Description of fossils from the Upper Secondary Formationof Cutch collected by C.W. Grant. Transactions of the Geological Society LondonSeries 2, 5, explanations of Plates 21-23 and figs. 8-17.
Spath, L.F., 1924. On the Blake collection of ammonites from Kachh, India. Memoirsof the Geological Survey of India, Palaeontologia Indica, New Series 9 1, 1–29.
Spath, L.F., 1927-1933. Revision of the Jurassic cephalopod fauna of Kachh(Cutch). Parts I-VI. Memoirs of the Geological Survey of India, PalaeontologiaIndica, new series 9 2, part I, 1-71, pls. 1-7 [1927]; part II, 72-161, pls. 8-19[1928a]; part III, 162-278, pls. 20-47 [1928b]; part IV, 279-550, pls. 48-102[1931a]; part V, 551-658, pls. 103-124 [1931b]; part VI, 659-945, pls. 125-130 [1933].
Stevens, G.R., 1997. The Late Jurassic ammonite fauna of New Zealand. Institute ofGeological & Nuclear Sciences monograph 18 (New Zealand Geological SurveyPaleontological Bulletin 74) 1–217.
Stipanicic, P.N., Westermann, G.E.G., Riccardi, A.C., 1975. The Indo-Pacific ammo-nite Mayaites in the Oxfordian of the southern Andes. Ameghiniana 12, 281–305.
Sykes, W.H., 1834. A notice respecting some fossils collected in Cutch, by Capt.Walter Smee, of the Bombay Army. Transactions of the Geological Society ofLondon, second series 5, 715–719.
Thierry, J., 1975. Ontogenese de la ligne de suture et origine des Mayaitidae Spath1928 (Ammonitina, Stephanocerataceae) de l’Oxfordien moyen de la provinceindo-malgache. Comptes rendus hebdomadaires des seances de l’Academie dessciences, serie D : sciences naturelles 280, 1543–1546.
Thierry, J., 1976. Paleobiogeographie de quelques Stephanocerataceae (Ammoni-tina) du Jurassique moyen et superieur ; une confrontation avec la theoriemobiliste. Geobios 9, 291–331.
Thomson, M.R.A., 1982. Late Jurassic fossils from Low Island, South Shetland Islands.British Antarctica Survey Bulletin 56, 25–35.
Tornquist, A., 1893. Fragmente einer Oxfordfauna von Mtaru in Deutsch-Ostafrika,nach dem von Dr. Stahlmann gesammelten Material. Mitteilungen aus demNaturhistorischen Museum in Hamburg 10, 265–288.
Uhlig, V., 1910. The fauna of the Spiti shales. Fasc. 2. Memoirs of the GeologicalSurvey of India, Palaeontologia India, series 15. Himalayan fossils 4, 133–306.
von Hillebrandt, A., Groschke, M., 1995. Ammoniten aus dem Callovium/Oxfor-dium-Grenzbereich von Nordchile. Berliner Geowissenschaftliche Abhandlun-gen, Reihe A 169, 1–40.
Waagen, W., 1873-1875. Jurassic fauna of Kutch. The Cephalopoda. Memoirs of theGeological Survey of India, Palaeontologia Indica, series 9 1, part I, 1-22, pls. 1-4[1873]; part II, 23-76, pls. 5-14 [1875a]; part III, 77-106, pls. 15-24 [1875b]; partIV, 107-247; pls. 25-60 [1875c]
Westermann, G.E.G., 1966. Covariation and taxonomy of the Jurassic ammoniteSonninia adicra (Waagen). Neues Jahrbuch fur Geologie und Palaontologie,Abhandlungen 124, 289–312.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005

M. Alberti et al. / Geobios xxx (2015) xxx–xxx46
G Model
GEOBIO-701; No. of Pages 46
Westermann, G.E.G., Riccardi, A.C., 1985. Middle Jurassic ammonite evolution in theAndean Province and emigration to Tethys. Sedimentary and EvolutionaryCycles, Lecture Notes in Earth Sciences 1, 6–34.
Westermann, G.E.G., Wang, Y.G., 1988. Middle Jurassic ammonites of Tibet and theage of the Lower Spiti Shales. Palaeontology 31, 295–339.
Westermann, G.E.G., Hudson, N., Grant-Mackie, J., 2002. New Jurassic Ammonitinafrom New Zealand: Bathonian-Callovian Eurycephalitinae. New Zealand Journalof Geology & Geophysics 45, 499–525.
Westermann, G.E.G., Sato, T., Skwarko, S.K., 1978. Brief report on the Jurassicbiostratigraphy of the Sula Islands, Indonesia. Newsletters on Stratigraphy 7,96–101.
Please cite this article in press as: Alberti, M., et al., Ammonites of the sWestern India. Geobios (2015), http://dx.doi.org/10.1016/j.geobios.2
Wynne, A.B., 1872. Memoir on the Geology of Kutch, to accompany the mapcompiled by A. B. Wynne and F. Fedden, during the seasons of 1867-68 and1868-69. Memoirs of the Geological Survey of India 9, 1–293.
Xiaochi, L., Grant-Mackie, J.A., 1988. Upper Jurassic and Lower Cretaceous Buchia(Bivalvia) from southern Tibet, and some wider considerations. Alcheringa 12,249–268.
Xiaoqiao, W., Jansa, L.F., Sarti, M., Zhao, W., 2000. New results of the Jurassicstratigraphic study in the Nyalam area of southern Tibet. GeoResearch Forum6, 263–268.
ubfamily Mayaitinae Spath, 1928 from the Oxfordian of Kachchh,015.02.005





























