Embed Size (px)
Citation preview

Amniotic Fluid can Act as anAppetitive UnconditionedStimulus in Preweanling Rats
Carlos AriasDepartment of Psychology, Binghamton
University—SUNY, P.O. Box 6000Binghamton, New York 13902-6000
E-mail: [email protected];[email protected]
M.G. ChotroFacultad de Psicologıa
Universidad del Paıs Vasco UPV/EHUSpain ABSTRACT: Studies in humans and animals indicate that exposure to flavors in
the amniotic fluid during the later gestational period may induce preferences forthose flavors. Considering that during the last prenatal period the amniotic fluidcontains substances that activate the opioid system, and that this system plays acritical role in the acquisition of olfactory preferences early in life, it has beenhypothesized that the amniotic fluid may acquire appetitive unconditionedproperties during this period. This has been tested in an experiment in whichpreweanling rats were exposed to alcohol odor (CS) paired or unpaired with theintraoral infusion of amniotic fluid (US) collected on gestational day 20. Thepairing of these two stimuli induced an enhanced palatability of alcohol’s flavor aswell an increased intake of the drug. These results support the idea that amnioticfluid acquires appetitive unconditioned properties during the last days of gestationand suggest that associative mechanisms involving the amniotic fluidcould be underlying odor and taste preferences acquired through fetal exposure.� 2007 Wiley Periodicals, Inc. Dev Psychobiol 49: 139–149, 2007.
Keywords: amniotic fluid; rat; ethanol; classical conditioning
INTRODUCTION
In humans, as in other mammals, innate preferences for or
aversions to certain flavors have been observed. In
general, during early ontogeny there is a preference for
sweet flavors, which are generally related to a high caloric
value, and an aversion towards bitter flavors that often
indicate toxicity (Hudson & Distel, 1999). However,
beyond these innate tendencies, there also seems to be a
sensitive period during early development in which
experiences with flavors produce enduring preferences
that ensure the acquisition of important information
about which foods are safe, appropriate, and nutritious
(Mennella & Beauchamp, 1998).
Several studies conducted with both animals
(Altbacker, Hudson, & Bilko, 1995; Arias & Chotro,
2005b; Bilko, Altbacker, & Hudson, 1994; Capretta &
Rawls, 1974; Dominguez, Lopez, & Molina, 1998;
Hepper & Waldman, 1992; Mennella & Beauchamp,
1998; Smotherman, 1982; Sneddon, Hadden, & Hepper,
1998) and humans (Mennella, Jagnow, & Beauchamp,
2001; Schaal, Marlier, & Soussignan, 2000) have found
that exposure to different tastes and odors during early
development can generate subsequent preferences for
these stimuli. In these studies, however, it is not clear
whether this preference is just the result of mere exposure,
or if certain associative processes are also involved. Odor
preferences can be acquired readily in rats without explicit
reinforcement (Caza & Spear, 1984; Hudson, 1993),
which probably helps the pup to become familiarized with
its environmental odors (Hudson & Distel, 1999). In other
cases, different sources of stimulation related to maternal
care strengthen the acquisition of olfactory preferences,
for example, thermo-tactile stimulation (Alberts & May,
1984). Milk and amniotic fluid are maternal-related fluids
that may also participate as unconditioned stimuli in
the acquisition of these preferences. Both fluids seem
able to induce appetitive unconditioned responses such as
mouthing movements and inhibition of fetal aversive
responses to sensory stimuli, like facial wiping. In
Received 11 April 2006; Accepted 10 November 2006Correspondence to: C. AriasPublished online in Wiley InterScience
(www.interscience.wiley.com). DOI 10.1002/dev.20205
� 2007 Wiley Periodicals, Inc.

addition, milk is able to induce an increase in rear limb
activity as part of a general reorganization in motor
behavior, an expression of the fetal stretch response
(Koffman, Petrov, Varlinskaya, & Smotherman, 1998;
Korthank & Robinson, 1998; Robinson & Smotherman,
1994). Furthermore, these fluids serve as vehicles through
which the fetus and the newborn are exposed to a variety of
chemosensorial stimulation (Mennella & Beauchamp,
1996; Mennella et al., 2001). Therefore, it is possible
that the fetus or the infant rat associates the different
odors or flavors presented in the amniotic fluid or milk,
respectively, with the reinforcing properties of such fluids.
To a certain extent, this association may play a role in
the development of preferences for the olfactory and
gustatory stimuli included in the mother’s diet during
gestation or lactation found in several studies (Galef &
Henderson, 1972; Galef & Sherry, 1973). Evidence
supporting this hypothesis is that milk can act as an
appetitive US in different procedures of classical
conditioning (Cheslock, Varlinskaya, Petrov, & Spear,
2000; Nizhnikov, Petrov, Varlinskaya, & Spear, 2002;
Robinson, Arnold, Spear, & Smotherman, 1993).
To our knowledge, the possibility that amniotic
fluid can act as an appetitive US has not been clearly
demonstrated. Nevertheless, there is evidence from
studies with humans and animals that indirectly supports
this hypothesis. First, during its intrauterine life the fetus
swallows and inhales large amounts of amniotic fluid
(Printchard, 1965; Ross & Nijland, 1997, 1998). The
intake of this fluid increases in the presence of some
flavors (De Snoo, 1937, cited by Ross & Nijland), which
may facilitate the processing of those stimuli. It is known
that a variety of substances included in the mother’s diet
reach the amniotic fluid (Abate, Pepino, Dominguez,
Spear, & Molina, 2000; Dominguez, Lopez, Chotro, &
Molina, 1996; Mennella, Johnson, & Beauchamp, 1995;
Nolte, Provenza, Callan, & Panter, 1992); and also that
prenatal experience with several chemosensory stimuli
increases their acceptance after birth (Abate, Spear, &
Molina, 2001; Arias & Chotro, 2005b; Bilko et al., 1994;
Capretta, Petersik, & Stewart, 1975; Dominguez et al.,
1998; Mennella et al., 2001; Smotherman, 1982). There is
some evidence to indicate that the amniotic fluid itself
induces unconditioned responses of appetence and
orientation. Human newborns show an orienting response
towards amniotic fluid (Marlier, Schaal, & Soussignan,
1997, 1998; Schaal, Marlier, & Soussignan, 1995, 1998)
and a preference for nipples that have been spread with
this fluid (Varendi, Porter, & Winberg, 1996). It has also
been found that the odor of amniotic fluid can calm crying
induced by maternal separation (Varendi, Christensson,
Porter, & Winberg, 1998).
In studies with rats it has also been found that amniotic
fluid guides the newborn towards the location of the nipple
during the first episode of breast-feeding (Mennella &
Beauchamp, 2002). Furthermore, pups show a preference
for nipples spread with an odor (citral) when this has been
added to the amniotic fluid before birth (Pedersen & Blass,
1982). It is interesting to indicate here that several studies
have shown that amniotic fluid contains substances
capable of activating the opioid system (Korthank &
Robinson, 1998; Kristal, Tarapacki, & Barton, 1990;
Kristal, Thompson, & Abbott, 1986). The amniotic fluid
collected during the last 2 days of gestation of the rat (GD
20–21) has been found to reduce facial wiping responses
induced by lemon (Korthank & Robinson), a behavior
expressed by the fetus in response to novel or aversive
stimuli which is known to be controlled by the opioid
system (Smotherman & Robinson, 1987). This same
effect has been observed when administering other
substances that activate the opioid system, such as milk
or morphine (Smotherman & Robinson, 1992a,b). In
general, there is evidence to indicate that the activation of
this neurochemical system plays an important role in the
acquisition of conditioned preferences both during the last
days of gestation (Arias & Chotro, 2005a; Arnold,
Robinson, Spear, & Smotherman, 1993; Chotro & Arias,
2003), and during the first days of postnatal life (Kehoe,
1988; Roth & Sullivan, 2001, 2003).
Taking into account all these antecedents, one may
expect the amniotic fluid to act as an appetitive US not
only during the intrauterine life, but also after birth. The
objective of the present experiment is to study this
possibility with a paradigm of classical conditioning in
preweanling rats. Ethanol odor was selected as the CS, in
light of previous studies demonstrating that preweanling
rats readily transfer conditioned learned responses from
the odor to the taste of ethanol (Molina, Hoffmann, &
Spear, 1986; Molina, Serwatka, Spear, & Spear, 1985;
Molina, Serwatka, & Spear, 1984, 1986; Serwatka,
Molina, & Spear, 1986). The hypothesis that guides this
study is that the association between the odor of ethanol
and the intraoral infusion of amniotic fluid will enhance
the palatability of ethanol’s flavor and will also increase
consumption of the drug.
METHOD
Subjects
Subjects for this experiment were 39 preweanling Wistar rats
(19 males and 20 females) derived from 7 litters. Pups were born
and reared in a temperature-controlled vivarium at the University
of the Basque Country (Spain). The colony room was maintained
on a 12-hr light/12-hr dark illumination cycle, with light onset at
8 a.m. Female rats were time-mated to provide subjects for this
study and were housed individually in standard maternity cages
with continuous access to food (Panlab, Barcelona, Spain,
Developmental Psychobiology. DOI 10.1002/dev140 Arias and Chotro

maternity formula) and water. In all experiments the European
regulations for care and treatment of experimental animals
were followed, and procedures were controlled and approved
by the Diputacion Foral de Gipuzkoa (Spain), in compliance
with the European Communities Council Directive (86/609/
EEC).
Procedures
Amniotic Fluid Collection. The US used in this experiment
was amniotic fluid collected on GD 20. To obtain this fluid,
pregnant rats were put down in a CO2 gas chamber.
Subsequently, a ventral laparotomy was performed and the fluid
was extracted from the amniotic sacs, placed in sterilized tubes
and frozen at �20�C until conditioning day. On conditioning
day, the fluid was defrosted to room temperature and placed into
the syringes used for its intraoral administration with an infusion
pump.
Conditioning Trials. On PDs 12 and 13 pups were separated
from their mothers, and distributed randomly into three groups,
Paired (n¼ 14), Unpaired (n¼ 13), and Nonexposed (n¼ 12).
Each group was placed separately in a holding chamber
(15� 8� 15 cm) maintained at 28–30�C with heating pads.
After 15 min, pups from the Unpaired group were exposed to the
ambient odor of ethanol, the CS. To carry out this exposure, a
cotton piece (4� 2 cm) was saturated with 5 mL of ethanol (96%
v/v) and attached to the internal surface of the cover of the testing
chamber. Fifteen minutes later, all pups were intraorally
cannulated using a procedure described in previous studies
(Hall & Rosenblatt, 1977). Cannulae are made from 5-cm
sections of polyethylene tubing (PE 10, Clay Adams) and
one end of the section is heated to form a small flange. A thin wire
attached to the nonflanged end of the cannula is placed on the
internal surface of the pup’s cheek and the wire is then pushed
through the oral mucosa until the flanged end of the cannula is
positioned over the internal surface of the cheek while the
remainder of the cannula exits from the oral cavity. The entire
procedure takes less than 5 s per pup and induces minimal stress.
These cannulae were later used to infuse the amniotic fluid
during the conditioning trials.
Two hours later, the bladders of pups from the Paired and
Unpaired conditions were voided by gentle brushing of the
anogenital area and their body weights were registered.
Immediately, these pups were placed in individual clear
Plexiglas chambers (15� 8� 15 cm) maintained at 28–30�C
with heating pads. Then, pups received intraorally the infusion of
amniotic fluid (US) delivered through an infusion pump (KD
Scientific, New Hope, PA) connected to the oral cannula of each
pup by a polyethylene catheter (PE 50, Clay Adams). The
volume administered to each subject’s mouth during this trial
was equivalent to 4% of their body weight and was infused with a
continuous flow during 15 min, with pups being able to either
consume or reject the infused solution. During these trials, pups
from group Paired were exposed simultaneously to the odor of
ethanol using a procedure similar to the one described for group
Unpaired. At the end of the session, postinfusion weights were
registered. Immediately after the conditioning trial, all pups
were returned to the home cage.
The intake of amniotic fluid was expressed as a percentage of
body weight gain after the administration of the ethanol solution,
using the following formula:
½ðpostinfusion weight � preinfusion weightÞ=preinfusion weight� � 100
Taste Reactivity Test
On PD 14 all pups were evaluated using an ethanol taste
reactivity test. Pups were separated from their mothers, placed in
heated holding chambers and intraorally cannulated, following
the procedure described previously. Three hours later, pups’
reaction to the taste of a 6% ethanol solution (v/v) was tested
following a procedure already described in previous studies
(Arias & Chotro, 2005a,b). The evaluation was conducted in a
trapezoid-shaped chamber with a front wall (29 cm wide)
made of clear glass and the remaining walls (back 18 cm, sides
11.5 cm) and floor made of mirror, so as to allow observation of
the pups’ orofacial expression and body movements in any
position. The chamber was 12.5 cm high and was divided into
two equal sections. Two pups were evaluated at a time, one per
section of the chamber. Pups remained in the test chamber for
2 min before the start of the intraoral infusions of ethanol; this
was considered the baseline period. Intraoral infusions were
performed using procedures similar to those described for
the conditioning trials, with two exceptions: the total volume
administered to each subject was reduced to 2.5% of their body
weight and it was infused in five 15-s pulses, with the onset of
each pulse separated by 2 min. The duration of the infusion test
was 10 min, preceded by a 2 min baseline period. During this
period pups were videotaped for subsequent analysis of their
behavior. Once the entire litter had been tested, pups were
returned to the maternal cage.
Based on previous studies using taste reactivity tests with
infant rats (Arias & Chotro, 2005a,b, 2006; Chotro, Kraebel,
McKinzie, Molina, & Spear, 1996; Hall & Bryan, 1981; Vigorito
& Sclafani, 1988) and adult rats (Grill & Norgren, 1978; Parker,
1988; Parker, 1995), as well as on our own observation of the
pups’ reaction to the taste of ethanol, the following behavioral
measures were selected as dependent variables under analysis:
general activity and wall climbing, considered here as aversive
reactions, and mouthing and paw licking, considered here as
ingestive/appetitive responses.
The behavioral measures were analyzed from the videotapes
and were scored every 30 s. General activity was rated in the
seven categories described by Hall & Bryan (1981): 0¼ no
movement, except for occasional twitches; 1¼ slight movement,
usually of the head or paw, sustained for 5 s; 2¼ substantial
movement of the head and paws, including grooming, but no
locomotion; 3¼ locomotion involving forelimbs and often
including rooting and probing but with hind limbs motionless
and usually serving as a pivot; 4¼ clear and sustained
locomotion about the test container; 5¼ vigorous locomotion,
often including rolling, kicking, and wall climbing; 6¼ an
extreme, but occasionally observed, form of rating five in which
the pup tumbled about its container for most of the 30 s interval,
locomoting, rolling, probing, wall climbing and jumping.
Developmental Psychobiology. DOI 10.1002/dev Amniotic Fluid as an Unconditioned Stimulus 141

A pup was considered to be wall climbing when standing on
its rear limbs with its forepaws against the wall of the testing
chamber. Time in seconds for wall climbing was registered for
each pup every 30 s during the baseline and test intervals.
Mouthing was defined as any obvious movement of the mouth
and jaws, and the total time (seconds) was also registered during
the testing trials. Mouthing and paw licking were independent
measures, that is, mouthing movements displayed during
paw licking were not measured as mouthing. Since mouthing
and paw licking are behaviors directly related to the intraoral
infusion of the liquid, no data were obtained during baseline
periods for these variables, with baseline data therefore
only being analyzed for general activity and wall climbing. As
with general activity, all these behaviors were scored every 30 s.
So, for general activity and wall climbing there were data for four
30-s baseline bins followed by twenty 30-sec infusion bins,
whereas for mouthing and paw licking there were data for only
twenty 30-sec testing bins. All behavioral observations were
performed by a trained researcher blind to the experimental
conditions.
Ethanol Intake Test
On the 2 days following the taste reactivity test (PDs 15 and 16)
all pups were tested in terms of ethanol intake. In the morning of
the corresponding day, pups were separated from their mothers,
placed in a holding chamber (15� 8� 15 cm) and intraorally
cannulated, as described above. The procedures used for this
ethanol intake test were similar to those described for the
intraoral infusion of amniotic fluid, except for the solution
infused (ethanol 6% v/v) and the volume administered to each
subject (5.5% of their body weight). The dependent variable
analyzed was ethanol intake expressed as a percentage of body
weight gained during the test.
Data Analysis
The effect of conditioning (Paired, Unpaired, and Nonexposed)
on body weights on PD 14 was analyzed with a one-way analysis
of variance (ANOVA).
Consumption data from the conditioning trials were analyzed
with a 2 (Conditioning: Paired and Unpaired)� 2 (Day) mixed
ANOVA, while data from the ethanol intake test were analyzed
with a 3 (Conditioning: Paired, Unpaired, and Nonexposed)�2 (Day) mixed ANOVA. Wall climbing data during baseline
trials were analyzed separately from the remaining infusion
trials with a 3 (Conditioning: Paired, Unpaired and Non-
exposed)� 4 (Bin) mixed ANOVA. Mouthing, paw licking, and
wall climbing data collected during the 20 test bins were
analyzed with a 3 (Conditioning: Paired, Unpaired, and
Nonexposed)� 20 (Bin) mixed ANOVA. Significant effects
were further explored using Tukey HSD post hoc tests and
repeated measures were compared using t-test for dependent
samples. Since general activity scores are ordinal data,
nonparametric tests (Kruskal–Wallis ANOVA and Mann–
Whitney U-test) were used. A rejection criterion of p< .05
was adopted for all the analyses presented in this study.
RESULTS
Body Weights
No differences between groups were detected when
comparing body weights on PD 14 (mean 29.18 g,
standard deviation¼ 1.54).
Amniotic Fluid Intake
The consumption of amniotic fluid during both condition-
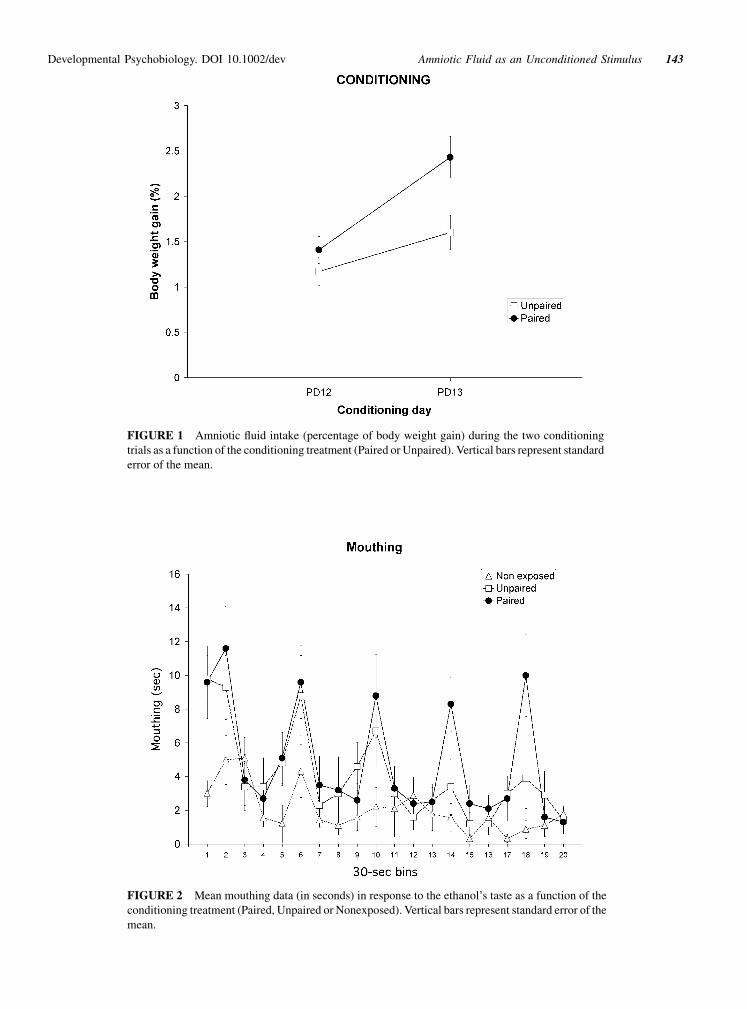
ing trials is presented in Figure 1. As can be observed, the
intake of amniotic fluid increased on the second day of
conditioning. However, this effect was observed mainly in
group Paired, which showed a higher intake of amniotic
fluid when compared to group Unpaired. The ANOVA
showed a significant effect of the main factors Condition-
ing [F(1,25)¼ 6.15, p< .05] and day [F(1,25)¼ 25.53,
p< .001], indicating that pups ingested more amniotic
fluid on the second conditioning trial, and that pups from
group Paired consumed more than those from group
Unpaired. The interaction between both factors was
nearly significant [F(1,25)¼ 4.22, p¼ .051]. Planned
comparisons indicated that during the second trial, group
Paired consumed significantly more amniotic fluid than
group Unpaired [F(1,25)¼ 7.61, p< .05].
Taste Reactivity Test
Figure 2 presents the mean duration of mouthing response
to ethanol during the twenty 30-sec scoring intervals as
a function of the conditioning treatment. As can be
observed in this figure, groups Paired and Unpaired
displayed more mouthing than group Nonexposed during
all infusion bins. During the last ones, however, group
Paired showed more mouthing than the other two groups.
It is important to note that there was a peak in mouthing
response during bins 2, 6, 10, 14, and 18. These bins
correspond to the 30 s immediately following the offset of
the intraoral infusions of ethanol. This peak in mouthing
response was observed mainly in groups Paired and
Unpaired, and seems to be habitual across the session for
groups Unpaired and Nonexposed, although not for group
Paired. The ANOVA confirmed these observations with a
significant effect of Conditioning [F(2,26)¼ 5,77,
p< .01], Bin [F(19,494)¼ 9.13, p< .001], as well as the
interaction between both factors [F(38,494)¼ 1.70,
p< .01]. Post hoc analyses of the main factor Condition-
ing revealed that pups from group Paired showed
significantly more mouthing than those from group
Nonexposed, while group Unpaired did not differ from
the other groups. Post hoc analysis of the interaction
indicated that during bin 1, groups Paired and Unpaired
showed more mouthing than group Nonexposed; during
Developmental Psychobiology. DOI 10.1002/dev142 Arias and Chotro
Developmental Psychobiology. DOI 10.1002/dev
FIGURE 1 Amniotic fluid intake (percentage of body weight gain) during the two conditioning
trials as a function of the conditioning treatment (Paired or Unpaired). Vertical bars represent standard
error of the mean.
FIGURE 2 Mean mouthing data (in seconds) in response to the ethanol’s taste as a function of the
conditioning treatment (Paired, Unpaired or Nonexposed). Vertical bars represent standard error of the
mean.
Amniotic Fluid as an Unconditioned Stimulus 143
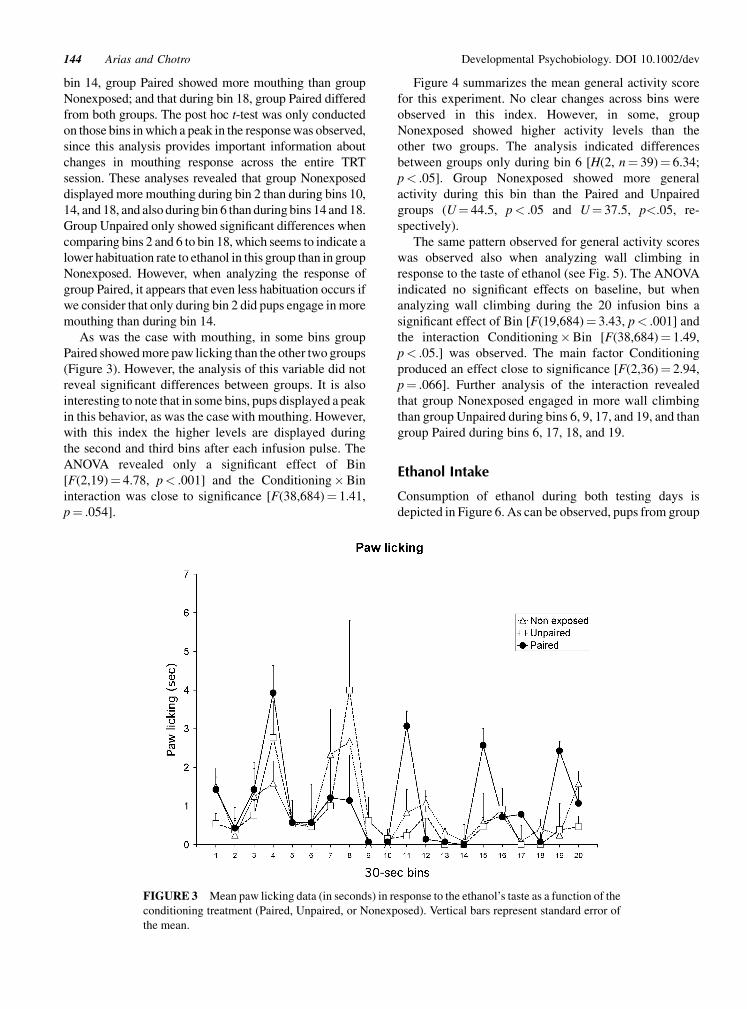
bin 14, group Paired showed more mouthing than group
Nonexposed; and that during bin 18, group Paired differed
from both groups. The post hoc t-test was only conducted
on those bins in which a peak in the response was observed,
since this analysis provides important information about
changes in mouthing response across the entire TRT
session. These analyses revealed that group Nonexposed
displayed more mouthing during bin 2 than during bins 10,
14, and 18, and also during bin 6 than during bins 14 and 18.
Group Unpaired only showed significant differences when
comparing bins 2 and 6 to bin 18, which seems to indicate a
lower habituation rate to ethanol in this group than in group
Nonexposed. However, when analyzing the response of
group Paired, it appears that even less habituation occurs if
we consider that only during bin 2 did pups engage in more
mouthing than during bin 14.
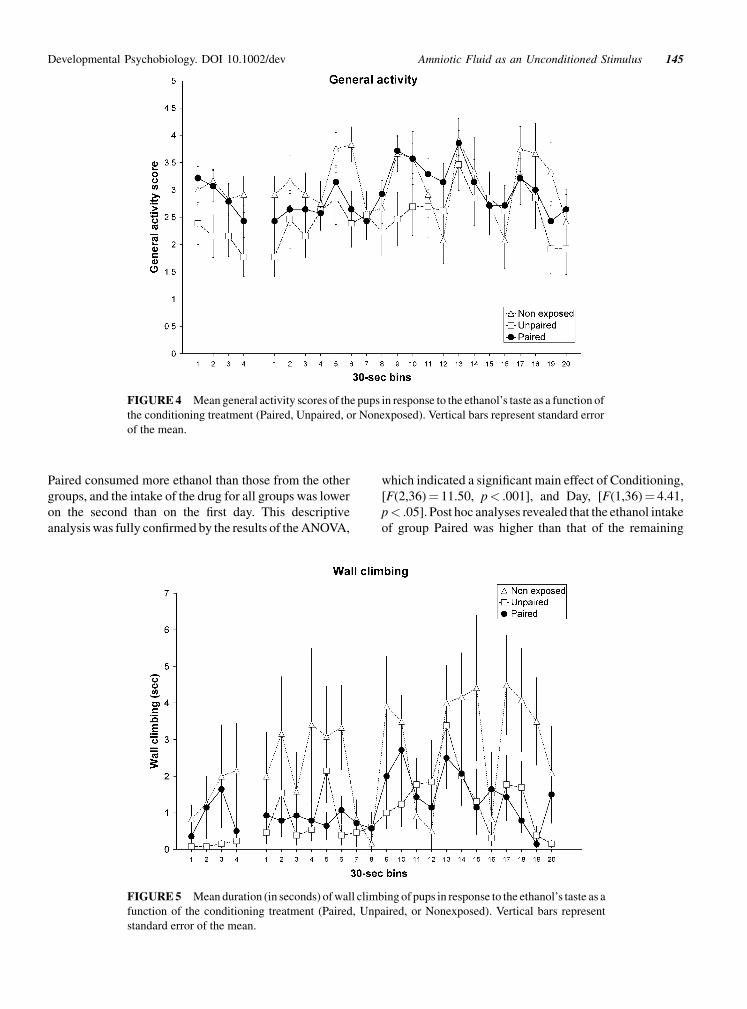
As was the case with mouthing, in some bins group
Paired showed more paw licking than the other two groups
(Figure 3). However, the analysis of this variable did not
reveal significant differences between groups. It is also
interesting to note that in some bins, pups displayed a peak
in this behavior, as was the case with mouthing. However,
with this index the higher levels are displayed during
the second and third bins after each infusion pulse. The
ANOVA revealed only a significant effect of Bin
[F(2,19)¼ 4.78, p< .001] and the Conditioning�Bin
interaction was close to significance [F(38,684)¼ 1.41,
p¼ .054].
Figure 4 summarizes the mean general activity score
for this experiment. No clear changes across bins were
observed in this index. However, in some, group
Nonexposed showed higher activity levels than the
other two groups. The analysis indicated differences
between groups only during bin 6 [H(2, n¼ 39)¼ 6.34;
p< .05]. Group Nonexposed showed more general
activity during this bin than the Paired and Unpaired
groups (U¼ 44.5, p< .05 and U¼ 37.5, p<.05, re-
spectively).
The same pattern observed for general activity scores
was observed also when analyzing wall climbing in
response to the taste of ethanol (see Fig. 5). The ANOVA
indicated no significant effects on baseline, but when
analyzing wall climbing during the 20 infusion bins a
significant effect of Bin [F(19,684)¼ 3.43, p< .001] and
the interaction Conditioning�Bin [F(38,684)¼ 1.49,
p< .05.] was observed. The main factor Conditioning
produced an effect close to significance [F(2,36)¼ 2.94,
p¼ .066]. Further analysis of the interaction revealed
that group Nonexposed engaged in more wall climbing
than group Unpaired during bins 6, 9, 17, and 19, and than
group Paired during bins 6, 17, 18, and 19.
Ethanol Intake
Consumption of ethanol during both testing days is
depicted in Figure 6. As can be observed, pups from group
Developmental Psychobiology. DOI 10.1002/dev
FIGURE 3 Mean paw licking data (in seconds) in response to the ethanol’s taste as a function of the
conditioning treatment (Paired, Unpaired, or Nonexposed). Vertical bars represent standard error of
the mean.
144 Arias and Chotro
Paired consumed more ethanol than those from the other
groups, and the intake of the drug for all groups was lower
on the second than on the first day. This descriptive
analysis was fully confirmed by the results of the ANOVA,
which indicated a significant main effect of Conditioning,
[F(2,36)¼ 11.50, p< .001], and Day, [F(1,36)¼ 4.41,
p< .05]. Post hoc analyses revealed that the ethanol intake
of group Paired was higher than that of the remaining
Developmental Psychobiology. DOI 10.1002/dev
FIGURE4 Mean general activity scores of the pups in response to the ethanol’s taste as a function of
the conditioning treatment (Paired, Unpaired, or Nonexposed). Vertical bars represent standard error
of the mean.
FIGURE5 Mean duration (in seconds) of wall climbing of pups in response to the ethanol’s taste as a
function of the conditioning treatment (Paired, Unpaired, or Nonexposed). Vertical bars represent
standard error of the mean.
Amniotic Fluid as an Unconditioned Stimulus 145
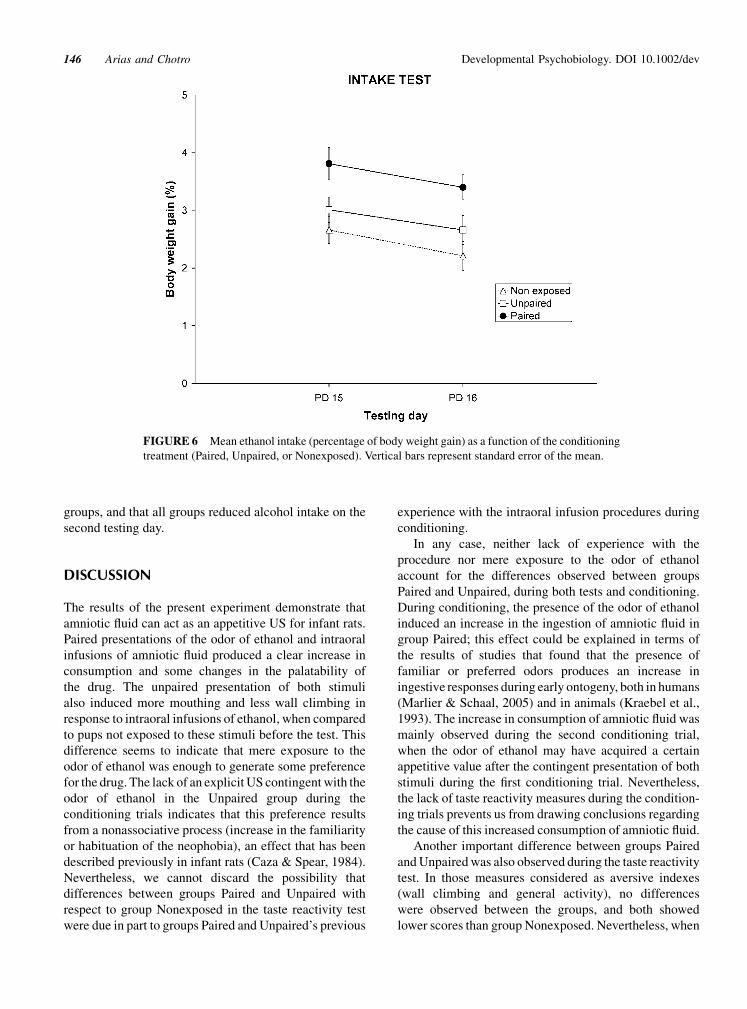
groups, and that all groups reduced alcohol intake on the
second testing day.
DISCUSSION
The results of the present experiment demonstrate that
amniotic fluid can act as an appetitive US for infant rats.
Paired presentations of the odor of ethanol and intraoral
infusions of amniotic fluid produced a clear increase in
consumption and some changes in the palatability of
the drug. The unpaired presentation of both stimuli
also induced more mouthing and less wall climbing in
response to intraoral infusions of ethanol, when compared
to pups not exposed to these stimuli before the test. This
difference seems to indicate that mere exposure to the
odor of ethanol was enough to generate some preference
for the drug. The lack of an explicit US contingent with the
odor of ethanol in the Unpaired group during the
conditioning trials indicates that this preference results
from a nonassociative process (increase in the familiarity
or habituation of the neophobia), an effect that has been
described previously in infant rats (Caza & Spear, 1984).
Nevertheless, we cannot discard the possibility that
differences between groups Paired and Unpaired with
respect to group Nonexposed in the taste reactivity test
were due in part to groups Paired and Unpaired’s previous
experience with the intraoral infusion procedures during
conditioning.
In any case, neither lack of experience with the
procedure nor mere exposure to the odor of ethanol
account for the differences observed between groups
Paired and Unpaired, during both tests and conditioning.
During conditioning, the presence of the odor of ethanol
induced an increase in the ingestion of amniotic fluid in
group Paired; this effect could be explained in terms of
the results of studies that found that the presence of
familiar or preferred odors produces an increase in
ingestive responses during early ontogeny, both in humans
(Marlier & Schaal, 2005) and in animals (Kraebel et al.,
1993). The increase in consumption of amniotic fluid was
mainly observed during the second conditioning trial,
when the odor of ethanol may have acquired a certain
appetitive value after the contingent presentation of both
stimuli during the first conditioning trial. Nevertheless,
the lack of taste reactivity measures during the condition-
ing trials prevents us from drawing conclusions regarding
the cause of this increased consumption of amniotic fluid.
Another important difference between groups Paired
and Unpaired was also observed during the taste reactivity
test. In those measures considered as aversive indexes
(wall climbing and general activity), no differences
were observed between the groups, and both showed
lower scores than group Nonexposed. Nevertheless, when
Developmental Psychobiology. DOI 10.1002/dev
FIGURE 6 Mean ethanol intake (percentage of body weight gain) as a function of the conditioning
treatment (Paired, Unpaired, or Nonexposed). Vertical bars represent standard error of the mean.
146 Arias and Chotro

analyzing the appetitive measures, group Unpaired,
although not group Paired, showed a decrease in mouthing
by the end of the test. This decreased mouthing, indicating
a habituation process, has been previously observed in
response to intraoral infusions of ethanol and other
solutions using this same procedure (Arias & Chotro,
2005b). The absence of this habituation effect in group
Paired allowed us to observe differences at the end of the
test between this group and group Unpaired. Furthermore,
as indicated above, group Paired showed not only
more mouthing in response to ethanol, but also a higher
intake of the drug. These differences in consumption and
ingestive responses can only be explained by the specific
treatment of group Paired, that is, the paired presentations
of the odor of ethanol and the intraoral infusion of
amniotic fluid, and the consequent appetitive conditioning
induced by this treatment. Considering all these results,
we can conclude that the amniotic fluid collected on GD
20 may act as an appetitive US in preweanling rats.
In this study, it is not possible to know if the amniotic
fluid acted as a primary or secondary reinforcer. As
mentioned before, amniotic fluid contains substances that
can activate the opioid system, and this neurochemical
system may be involved in the rewarding properties of
amniotic fluid (Korthank & Robinson, 1998). However,
we cannot be sure whether these substances act during
PDs 11 and 12 in the same way as during the fetal period,
considering the changes that occur in the opioid system
around PD 10 (Roth & Sullivan, 2003). It is possible also
that these opioid-activating substances may be degraded
after the procedure we used to collect and preserve this
fluid. If this were the case, in our experiment amniotic
fluid would most probably be acting as a secondary
reinforcer; in other words, its flavor may have acquired
rewarding properties in utero. Further research needs to be
done in order to investigate these possibilities. With the
present data we can conclude that amniotic fluid may
acquire appetitive unconditioned properties during the
last days of gestation.
It could be hypothesized that the preference for several
prenatally experienced substances described in several
studies (Abate et al., 2001; Arias & Chotro, 2005b; Bilko
et al., 1994; Dominguez et al., 1998; Mennella et al., 2001;
Molina, Chotro, & Dominguez, 1995; Schaal et al., 2000;
Smotherman, 1982), may be mediated by an associative
process. The presence of certain flavors in the amniotic
fluid seems to increase its ingestion (De Snoo, 1937, cited
by Ross & Nijland, 1997), which in turn may increase the
perception of the chemosensory aspects of these flavors.
During the last days of gestation it seems that the
composition of the physical characteristics of amniotic
fluid undergoes several physical and chemical
changes, and that during this period, this fluid may
activate the opioid system (Korthank & Robinson, 1998).
Considerable evidence indicates that this neurochemical
system plays an important role in the acquisition of
appetitive conditioned responses during early ontogeny
(Kehoe, 1988; Roth & Sullivan, 2001, 2003) as well as
during the last gestational days (Arias & Chotro, 2005a;
Arnold et al., 1993; Chotro & Arias, 2003). Therefore,
during this period of gestation, the fetus may associate the
flavors perceived in utero with the rewarding effects
produced by the ingestion of amniotic fluid, the latter
being mediated by the activation of the opioid system.
This hypothesis has not been supported in this study, and
remains open to future research.
The results of this experiment may also have implica-
tions for the analysis of the results found in other studies in
which the prenatal administration of a moderate ethanol
dose increased the preference for the chemosensory
properties of the drug (Arias & Chotro, 2005a,b; Chotro
& Arias, 2003; Dominguez et al., 1996, 1998; Molina
et al., 1995). This preference seems to be a conditioned
response mediated by the activation of the opioid system
(Arias & Chotro, 2005a; Chotro & Arias, 2003). In other
words, the fetus perceives the chemosensory properties of
ethanol in utero and, apparently, associates them with its
rewarding effects, mediated by the activation of the opioid
system. Nevertheless, it is not yet clear whether the
activation of this neurochemical system is induced by
ethanol, by amniotic fluid or by both.
NOTES
Carlos Arias was supported by the doctoral fellowship from
Gobierno Vasco (Programa de Formacion de Investigadores).
We thank Ana Medina and Oscar Vegas for their technical
assistance.
REFERENCES
Abate, P., Pepino, M. Y., Dominguez, H. D., Spear, N. E., &
Molina, J. C. (2000). Fetal associative learning mediated
through maternal alcohol intoxication. Alcoholism, Clinical
and Experimental Research, 24(1), 39–47.
Abate, P., Spear, N. E., & Molina, J. C. (2001). Fetal and
infantile alcohol-mediated associative learning in the rat.
Alcoholism, Clinical and Experimental Research, 25(7),
989–998.
Alberts, J. R., & May, B. (1984). Nonnutritive, thermotactile
induction of filial huddling in rat pups. Developmental
Psychobiology, 17(2), 161–181.
Altbacker, V., Hudson, R., & Bilko, A. (1995). Rabbit-mothers’
diet influences pups’ later food choice. Ethology, 99(2),
107–116.
Arias, C., & Chotro, M. G. (2005a). Increased palatability of
ethanol after prenatal ethanol exposure is mediated by the
Developmental Psychobiology. DOI 10.1002/dev Amniotic Fluid as an Unconditioned Stimulus 147

opioid system. Pharmacology, Biochemistry, and Behavior,
82(3), 434–442.
Arias, C., & Chotro, M. G. (2005b). Increased preference for
ethanol in the infant rat after prenatal ethanol exposure,
expressed on intake and taste reactivity tests. Alcoholism,
Clinical and Experimental Research, 29(3), 337–346.
Arias, C., & Chotro, M. G. (2006). Ethanol-induced preferences
or aversions as a function of age in preweanling rats.
Behavioural Neuroscience, 120(3), 710–718.
Arnold, H. M., Robinson, S. R., Spear, N. E., & Smotherman,
W. P. (1993). Conditioned opioid activity in the rat fetus.
Behavioural Neuroscience, 107(6), 963–969.
Bilko, A., Altbacker, V., & Hudson, R. (1994). Transmission of
food preference in the rabbit: The means of information
transfer. Physiology & Behavior, 56(5), 907–912.
Capretta, P. J., Petersik, J. T., & Stewart, D. J. (1975).
Acceptance of novel flavours is increased after early
experience of diverse tastes. Nature, 254(5502), 689–
691.
Capretta, P. J., & Rawls, L. H. III. (1974). Establishment of a
flavor preference in rats: Importance of nursing and weaning
experience. Journal of Comparative and Physiological
Psychology, 86(4), 670–673.
Caza, P. A., & Spear, N. E. (1984). Short-term exposure to an
odor increases its subsequent preference in preweanling rats:
A descriptive profile of the phenomenon. Developmental
Psychobiology, 17(4), 407–422.
Cheslock, S. J., Varlinskaya, E. I., Petrov, E. S., & Spear, N. E.
(2000). Rapid and robust olfactory conditioning with milk
before suckling experience: Promotion of nipple attachment
in the newborn rat. Behavioural Neuroscience, 114(3), 484–
495.
Chotro, M. G., & Arias, C. (2003). Prenatal exposure to ethanol
increases ethanol consumption: A conditioned response?
Alcohol, 30(1), 19–28.
Chotro, M. G., Kraebel, K. S., McKinzie, D. L., Molina, J. C., &
Spear, N. (1996). Prenatal and postnatal ethanol exposure
influences preweanling rats’ behavioral and autonomic
responding to ethanol odor. Alcohol, 13(4), 377–385.
De Snoo, K. (1937). Das trikende kind in uterus. Monatschr
f Geburtsh u Gynaek, 105, 88–97.
Dominguez, H. D., Lopez, M. F., Chotro, M. G., & Molina, J. C.
(1996). Perinatal responsiveness to alcohol’s chemosensory
cues as a function of prenatal alcohol administration during
gestational days 17–20 in the rat. Neurobiology of Learning
and Memory, 65(2), 103–112.
Dominguez, H. D., Lopez, M. F., & Molina, J. C. (1998).
Neonatal responsiveness to alcohol odor and infant alcohol
intake as a function of alcohol experience during late
gestation. Alcohol, 16(2), 109–117.
Galef, B. G. Jr., & Henderson, P. W. (1972). Mother’s milk: A
determinant of the feeding preferences of weaning rat pups.
Journal of Comparative and Physiological Psychology, 78(2),
213–219.
Galef, B. G. Jr., & Sherry, D. F. (1973). Mother’s milk: A
medium for transmission of cues reflecting the flavor of
mother’s diet. Journal of Comparative and Physiological
Psychology, 83(3), 374–378.
Grill, H. J., & Norgren, R. (1978). The taste reactivity test. I.
Mimetic responses to gustatory stimuli in neurologically
normal rats. Brain Research, 143(2), 263–279.
Hall, W. G., & Bryan, T. E. (1981). The ontogeny of feeding in
rats: IV. Taste development as measured by intake and
behavioral responses to oral infusions of sucrose and quinine.
Journal of Comparative and Physiological Psychology, 95(2),
240–251.
Hall, W. G., & Rosenblatt, G. (1977). Suckling behavior and
intake control in the developing rat pup. Journal of
Comparative and Physiological Psychology, 91, 1232–1247.
Hepper, P. G., & Waldman, B. (1992). Embryonic olfactory
learning in frogs. The Quarterly Journal of Experimental
Psychology. B, Comparative and Physiological Psychology,
44(3–4), 179–197.
Hudson, R. (1993). Olfactory imprinting. Current Opinion in
Neurobiology, 3(4), 548–552.
Hudson, R., & Distel, H. (1999). The flavor of life: Perinatal
development of odor and taste preferences. Schweizerische
Medizinische Wochenschrift, 129(5), 176–181.
Kehoe, P. (1988). Opioids, behavior, and learning in mamma-
lian development. In E. M. Blass (Ed.), Developmental
psychology and behavioral ecology (pp. 309–340). New
York and London: Plenum Press.
Koffman, D. J., Petrov, E. S., Varlinskaya, E. I., & Smotherman,
W. P. (1998). Thermal, olfactory, and tactile stimuli increase
oral grasping of an artificial nipple by the newborn rat.
Developmental Psychobiology, 33(4), 317–326.
Korthank, A. J., & Robinson, S. R. (1998). Effects of amniotic
fluid on opioid activity and fetal responses to chemosensory
stimuli. Developmental Psychobiology, 33(3), 235–248.
Kraebel, K. S., McKinzie, D. L., Chotro, M. G., Wurtzel, N. L.,
Molina, J. C., & Spear, N. E. (1993, November 4–7). Chronic
prenatal exposure to ethanol influences later responsivity to
the chemosensory cues of ethanol in infant rat pups.
Paper presented at the Annual meeting of the International
Society for Developmental Psychobiology, Alexandria,
Virginia.
Kristal, M. B., Tarapacki, J. A., & Barton, D. (1990). Amniotic
fluid ingestion enhances opioid-mediated but not nonopioid-
mediated analgesia. Physiology & Behavior, 47(1), 79–81.
Kristal, M. B., Thompson, A. C., & Abbott, P. (1986). Ingestion
of amniotic fluid enhances opiate analgesia in rats.
Physiology & Behavior, 38(6), 809–815.
Marlier, L., & Schaal, B. (2005). Human newborns prefer
human milk: Conspecific milk odor is attractive without
postnatal exposure. Child Development, 76(1), 155–168.
Marlier, L., Schaal, B., & Soussignan, R. (1997). Orientation
responses to biological odours in the human newborn. Initial
pattern and postnatal plasticity. Comptes Rendus de
L’Academie des Sciences Serie III-Sciences de la Vie-Life
Sciences, 320 (12), 999–1005.
Marlier, L., Schaal, B., & Soussignan, R. (1998). Neonatal
responsiveness to the odor of amniotic and lacteal fluids: A
test of perinatal chemosensory continuity. Child Develop-
ment, 69(3), 611–623.
Mennella, J. A., & Beauchamp, G. K. (1996). The early
development of human flavor preferences. In E. D. Capaldi
Developmental Psychobiology. DOI 10.1002/dev148 Arias and Chotro

(Ed.), Why we eat what we eat (pp. 83–112). Washington:
American Psychological Association.
Mennella, J. A., & Beauchamp, G. K. (1998). Early flavor
experiences: Research update. Nutrition Reviews, 56(7),
205–211.
Mennella, J. A., & Beauchamp, G. K. (2002). Flavor
experiences during formula feeding are related to preferences
during childhood. Early Human Development, 68(2), 71–82.
Mennella, J. A., Jagnow, C. P., & Beauchamp, G. K. (2001).
Prenatal and postnatal flavor learning by human infants.
Pediatrics, 107(6), E88.
Mennella, J. A., Johnson, A., & Beauchamp, G. K. (1995).
Garlic ingestion by pregnant women alters the odor of
amniotic fluid. Chemical Senses, 20(2), 207–209.
Molina, J. C., Chotro, M. G., & Dominguez, H. D. (1995). Fetal
alcohol learning derived from ethanol contamination of the
prenatal environment. In J. P. Lecanuet, W. P. Fifer, N.
Krasnegor, & W. P. Smotherman (Eds.), Fetal development:
A psychobiological perspective (pp. 419–438). Hillsdale,
NJ: Lawrence Erlbaum Associates.
Molina, J. C., Hoffmann, H., & Spear, N. E. (1986).
Conditioning of aversion to alcohol orosensory cues in 5-
and 10-day rats: Subsequent reduction in alcohol ingestion.
Developmental Psychobiology, 19(3), 175–183.
Molina, J. C., Serwatka, J., Spear, L. P., & Spear, N. E. (1985).
Differential ethanol olfactory experiences affect ethanol
ingestion in preweanlings but not in older rats. Behavioral
and Neural Biology, 44(1), 90–100.
Molina, J. C., Serwatka, J., & Spear, N. E. (1984). Changes in
alcohol intake resulting from prior experiences with alcohol
odor in young rats. Pharmacology, Biochemistry, and
Behavior, 21(3), 387–391.
Molina, J. C., Serwatka, J., & Spear, N. E. (1986). Alcohol
drinking patterns of young adult rats as a function of infantile
aversive experiences with alcohol odor. Behavioral and
Neural Biology, 46(3), 257–271.
Nizhnikov, M. E., Petrov, E. S., Varlinskaya, E. I., & Spear,
N. E. (2002). Newborn rats’ first suckling experience: Taste
differentiation and suckling plasticity. Physiology & Beha-
vior, 76(2), 181–198.
Nolte, D. L., Provenza, F. D., Callan, R., & Panter, K. E. (1992).
Garlic in the ovine fetal environment. Physiology &
Behavior, 52(6), 1091–1093.
Parker, L. (1988). Positively reinforcing drugs may produce a
different kind of CTA than drugs which are not positively
reinforcing. Learning and Motivation, 19, 207–220.
Parker, L. A. (1995). Rewarding drugs produce taste avoidance,
but not taste aversion. Neuroscience & Biobehavioral
Reviews, 19(1), 143–157.
Pedersen, P. E., & Blass, E. M. (1982). Prenatal and postnatal
determinants of the 1st suckling episode in albino rats.
Developmental Psychobiology, 15(4), 349–355.
Printchard, J. A. (1965). Deglution of normal and anencephalic
fetuses. Obstetrics and Gynecology, 25, 289–297.
Robinson, S. R., Arnold, H. M., Spear, N. E., & Smotherman,
W. P. (1993). Experience with milk and an artificial nipple
promotes conditioned opioid activity in the rat fetus.
Developmental Psychobiology, 26(7), 375–387.
Robinson, S. R., & Smotherman, W. P. (1994). Behavioral
effects of milk in the rat fetus. Behavioural Neuroscience,
108(6), 1139–1149.
Ross, M. G., & Nijland, M. J. (1997). Fetal swallowing:
Relation to amniotic fluid regulation. Clinical Obstetrics and
Gynecology, 40(2), 352–365.
Ross, M. G., & Nijland, M. J. (1998). Development of ingestive
behavior. American Journal of Physiology, 274(4 Pt 2),
R879–R893.
Roth, T. L., & Sullivan, R. M. (2001). Endogenous opioids and
their role in odor preference acquisition and consolidation
following odor-shock conditioning in infant rats. Develop-
mental Psychobiology, 39(3), 188–198.
Roth, T. L., & Sullivan, R. M. (2003). Consolidation and
expression of a shock-induced odor preference in rat pups is
facilitated by opioids. Physiology & Behavior, 78(1), 135–
142.
Schaal, B., Marlier, L., & Soussignan, R. (1995). Responsive-
ness to the odour of amniotic fluid in the human neonate.
Biology of the Neonate, 67(6), 397–406.
Schaal, B., Marlier, L., & Soussignan, R. (1998). Olfactory
function in the human fetus: Evidence from selective
neonatal responsiveness to the odor of amniotic fluid.
Behavioural Neuroscience, 112(6), 1438–1449.
Schaal, B., Marlier, L., & Soussignan, R. (2000). Human
foetuses learn odours from their pregnant mother’s diet.
Chemical Senses, 25(6), 729–737.
Serwatka, J., Molina, J. C., & Spear, N. E. (1986). Weanlings’
transfer of conditioned ethanol aversion from olfaction to
ingestion depends on the unconditioned stimulus. Behavioral
and Neural Biology, 45(1), 57–70.
Smotherman, W. P. (1982). In utero chemosensory experience
alters taste preferences and corticosterone responsiveness.
Behavioral and Neural Biology, 36, 61–68.
Smotherman, W. P., & Robinson, S. R. (1987). Prenatal
expression of species-typical action patterns in the rat fetus
(Rattus norvegicus). Journal of Comparative Psychology,
101(2), 190–196.
Smotherman, W. P., & Robinson, S. R. (1992a). Kappa opioid
mediation of fetal responses to milk. Behavioural Neu-
roscience, 106(2), 396–407.
Smotherman, W. P., & Robinson, S. R. (1992b). Prenatal
experience with milk: Fetal behavior and endogenous opioid
systems. Neuroscience & Biobehavioral Reviews, 16(3),
351–364.
Sneddon, H., Hadden, R., & Hepper, P. G. (1998). Chemosen-
sory learning in the chicken embryo. Physiology & Behavior,
64(2), 133–139.
Varendi, H., Christensson, K., Porter, R. H., & Winberg, J.
(1998). Soothing effect of amniotic fluid smell in
newborn infants. Early Human Development, 51(1), 47–
55.
Varendi, H., Porter, R. H., & Winberg, J. (1996). Attractiveness
of amniotic fluid odor: Evidence of prenatal olfactory
learning? Acta Paediatrica, 85(10), 1223–1227.
Vigorito, M., & Sclafani, A. (1988). Ontogeny of polycose and
sucrose appetite in neonatal rats. Developmental Psycho-
biology, 21(5), 457–465.
Developmental Psychobiology. DOI 10.1002/dev Amniotic Fluid as an Unconditioned Stimulus 149

























